angiotensin ii signaling and hb-egf shedding via metalloproteinase in glomerular mesangial cells
TRANSCRIPT

Kidney International, Vol. 60 (2001), pp. 2153–2163
Angiotensin II signaling and HB-EGF shedding viametalloproteinase in glomerular mesangial cells
YOKO UCHIYAMA-TANAKA, HIROAKI MATSUBARA, YOSHIHISA NOZAWA, SATOSHI MURASAWA,YASUKIYO MORI, ATSUSHI KOSAKI, KATSUYA MARUYAMA, HIROYA MASAKI,YASUNOBU SHIBASAKI, SOICHIRO FUJIYAMA, ATSUKO NOSE, OSAMU IBA,TAKAMASA HASAGAWA, ERIKO TATEISHI, SHIGEKI HIGASHIYAMA, and TOSHIJI IWASAKA
Department of Medicine II, Kansai Medical University, Osaka; Pharmacological Laboratory, Taiho Pharmaceutical Co. Ltd.,Tokushima; and Department of Biochemistry, School of Allied Health Science, Osaka University Faculty of Medicine,Osaka, Japan
Angiotensin II signaling and HB-EGF shedding via metallo- Angiotensin II (Ang II) has been shown to have rolesproteinase in glomerular mesangial cells. in the development of glomerulosclerosis by induction
Background. Angiotensin II (Ang II) has been implicated of hemodynamic changes and synthesis of extracellularin the development of glomerulosclerosis by stimulating fibro-matrix proteins. Glomerular mesangial cells have abun-nectin (FN) synthesis. The processing and release of heparindant high-affinity Ang II receptors, which are classifiedbinding–endothelin growth factor (HB-EGF) are activated by
protein kinase C (PKC) and Ca2� signaling. We studied the pharmacologically as belonging to the Ang II type-1 re-roles of HB-EGF and endothelial growth factor (EGF) receptor ceptor (AT1-R) subtype [1–3]. The AT1-R signal is in-(EGFR) in Ang II-induced FN expression using mesangial cells. volved in glomerulosclerosis by stimulation of cell pro-Methods. Mesangial cells were prepared from mouse kidneys
liferation and synthesis of extracellular matrix proteinsby the explant method and cells were used at passages 4 and 5.[4, 5]. AT1-R stimulation initiates early biochemicalResults. Ang II stimulated FN mRNA levels dose-depen-
dently with a maximal increase (3.4-fold) after 12 hours of events, including rapid production of diacylglycerol andincubation. This action was completely inhibited by PKC inhib- inositol 1,4,5-trisphosphate by phospholipase C (PLC)-itors and slightly blocked by Ca2� chelating agents. FN mRNA mediated hydrolysis of inositol phospholipids and activa-accumulation by Ang II was abolished by tyrosine kinase inhib-
tion of protein kinase C (PKC) [6–9]. Ang II also inducesitors, a specific inhibitor for EGFR (AG1478) and extracellularsignal-regulated kinase (ERK) inactivation. Addition of neu- a rapid increase in expression of growth-associated nu-tralizing anti–HB-EGF antibody, as well as pretreatment with clear proto-oncogenes as observed in cellular events me-heparin or the metalloproteinase inhibitor batimastat abol- diated by peptide growth factors, and stimulates tyrosineished induction of FN expression by Ang II. In mesangial cells
phosphorylation of multiple substrates, including extra-stably transfected with a chimeric construct containing HB-cellular signal-regulated kinases (ERK) [10–15].EGF and alkaline phosphatase (ALP) genes, ALP activity in
incubation medium was rapidly increased by Ang II (1.7-fold Signaling through ERK in response to various G pro-at 0.5 min) and reached a 4.1-fold increase at two minutes. tein-coupled receptor agonists has been shown to relyAng II phosphorylated EGFR (maximal at 2 min) and ERK on transactivation of the EGFR [reviewed in 16, 17].(maximal at 8 min) in a PKC- and metalloproteinase-depen-
Daub et al originally found the role of EGFR-mediateddent manner. Ang II stimulated the expression and release oftransforming growth factor-� (TGF-�) via EGFR-mediated signaling in Ras/ERK activation by G-protein-coupledsignaling, and the released TGF-� also contributed to Ang II- receptors (GPCRs) [18]. Ang II also was found to trans-mediated FN expression via EGFR transactivation. activate EGFR in cardiac fibroblasts or vascular smooth
Conclusions. Ang II-mediated FN expression was regulatedmuscle cells (VSMC) [19–23]. EGFR transactivation wasby autocrine effects of HB-EGF and TGF-�, suggesting a novelinitially suggested to be independent of EGFR ligands,paradigm for cross-talk between Ang II and growth factor
receptor signaling pathways. based on the rapid onset of EGFR tyrosine phosphoryla-tion and the failure to detect EGFR ligands, in particularEGF, in conditioned medium of stimulated cells [18, 19,Key words: angiotensin II, HB-EGF, TGF-�, fibronectin, mesangial
cell, EGF receptor, cell signaling. 21]. Thereafter, several groups reported the involvementof HB-EGF in GPCR-mediated EGFR transactivationReceived for publication February 6, 2001[24–26]. Filardo et al demonstrated that estrogen alsoand in revised form July 23, 2001
Accepted for publication July 25, 2001 activated ERK via HB-EGF-mediated EGFR activation[27]. HB-EGF is a heparin-binding member of the EGF 2001 by the International Society of Nephrology
2153

Uchiyama-Tanaka et al: Angiotensin II and HB-EGF2154
family and is synthesized as a transmembrane precursor glyceraldehyde-3-phosphate dehydrogenase (GAPDH)that is then proteolytically processed into the mature, as an internal control. Signals were measured by scanningsoluble growth factor. This proteolytic processing was densitometry, and FN mRNA levels were normalizedreported to be regulated by PKC and Ca2� signaling [28]. relative to the GAPDH mRNA levels. Ca2� levels wereAlthough accumulating evidence suggested the involve- measured using fura-2/AM as reported [20].ment of HB-EGF in EGFR transactivation, the actual We constructed a pRC expression vector containingrelease of HB-EGF into incubation medium has not been the human alkaline phosphatase (ALP) cDNA subcloneddemonstrated, and it remains undefined whether such a between human HB-EGF signal sequence and the cod-signaling mechanism is operating in mesangial cells. ing region for mature human HB-EGF protein (pALP-
Fibronectin (FN) is an extracellular matrix and plasma HB-EGF) [33]. This pAlp-HB-EGF (30 �g) was trans-protein with various functions that plays important roles fected into mesangial cells using LIPOFECTAMINEin mesangial cells, cell adhesion and migration [29]. This PLUS� (Gibco BRL) and selected with G418 [19, 20].study focused on the signaling mechanism for Ang II- Three stable cell lines with high ALP activity in theinduced FN expression in mesangial cells, and examined incubation medium were selected and analyzed in thiswhether Ang II actually stimulates the processing and experiment. For determination of ALP activity, the incu-release of HB-EGF, which in turn transactivates the bation medium was heated at 70�C for ten minutes andEGFR-mediated ERK pathway to regulate FN gene ex- aliquots of 100 �L were further incubated at 37�C forpression in mesangial cells. five minutes in assay buffer (100 �L) containing dietha-
nolamine (2 mol/L, pH 9.8), MgCl2 (2 mmol/L), homo-arginine (0.4 mol/L) and disodium p-nitrophenyl phos-METHODSphate (12.5 mmol/L). The sample volume was expandedMaterialsto 1 mL and absorbance at 405 nm was read. The values
Heparin binding-EGF, genistein, AG1478, AG1295, were normalized with cellular protein using bovine serumherbimycin, BAPTA-AM, PD98059 and TMB-8 were albumin (BSA) as a standard (Pierce Chemical, Rockford,purchased from Calbiochem (La Jolla, CA, USA). Stau- IL, USA) and background absorbance in the medium,rosporin, calphostin-C, phorbol 12-myristate 13-acetate and are shown relative to the time 0 control value.(PMA) and heparin (porcine intestinal mucosa) werefrom Sigma (St. Louis, MO, USA). Angiotensin II and Measurement of ERK and EGFR phosphorylationEGF were from Peptide Institute Inc. (Osaka, Japan)
Tyrosine-phosphorylation of EGFR was analyzed inand Toyobo (Osaka, Japan), respectively. Antibodies werecells exposed to Ang II for two minutes as previouslyfrom the following vendors: Telios Pharm (FN), Santadescribed [19, 20]. After brief sonication, samples wereCruz Biotechnology (Santa Cruz, CA, USA; EGFR, HB-incubated for 30 minutes at 4�C, centrifuged for fiveEGF, normal rabbit IgG), New England Bio-Labs (Bev-minutes at 15,000 � g and the supernatant was incubatederly, MA, USA; phospho-specific ERK), Upstate Bio-with anti-rabbit EGFR antibody for 16 hours at 4�C.technology (Lake Placid, NY, USA; phospho-tyrosineImmune complexes were affinity-purified by anti-rabbit4G10), R&D Systems (Wiesbaden, Germany; pan-selec-IgG agarose beads (Sigma), transferred on nitrocellulosetive TGF-�). The metalloproteinase inhibitor batimastatmembranes, blotted with anti-phosphotyrosine antibodywas from British Biotech (Cowley, Oxford, UK). Theand detected with the enhanced chemiluminescence kitAT1-R antagonist RNH6270 (active form of CS866) was(Amersham Japan, Tokyo, Japan). Threonine/tyrosine-provided by Sankyo Co. Ltd (Tokyo, Japan).phosphorylation of ERK was determined in cells ex-posed to Ang II for eight minutes using phospho-ERKCell cultureantibody [19, 20].Mesangial cells were prepared from kidneys of 8-week-
old male C57BL/6 mice by the explant method [3], andDetection of FN released into mediumcultured with Dulbecco’s modified Eagle’s medium
Quantification of FN secreted into medium was per-(DMEM) containing 10% fetal calf serum (FCS). Sub-formed as previously reported [22]. Following 12 hoursconfluent cells (passages 4 and 5) were serum-starved forof incubation of cells with or without Ang II (100 nmol/L),24 hours and then used for the experiments.100 �Ci/ml of [35S]-methionine was added for further 12
Northern blotting, stable transfection of DNA, hours. Aliquots of the medium were incubated with rab-Ca2� analyses and measurement of ALP activity bit anti-FN antibody for 16 hours at 4�C. Immune com-
plexes were affinity-purified using protein A-sepharoseTotal RNA was extracted using TRIZOL Reagentbeads and analyzed by sodium dodecyl sulfate-polyacryl-(Gibco BRL, Paisley, Scotland, UK), fractionated on 1%amide gel electrophoresis (SDS-PAGE). The intensityagarose/formaldehyde gels, and transferred onto nylonof signals of 220 kD was determined by densitometry andmembranes [30–32]. Blots were then hybridized with
random-primed 32P-labeled cDNA probes for FN and the value in the control was arbitrarily expressed as 1 unit.

Uchiyama-Tanaka et al: Angiotensin II and HB-EGF 2155
Binding assay and measurement of TGF-� tained a similar level until 24 hours (data not shown).Mesangial cells expressed only AT1-R (kD � 0.61 �Membrane preparation and binding assay were per-0.02 nmol/L, Bmax � 1.55 � 0.14 pmol/mg protein), andformed as previously described [34–36]. Briefly, cellsAT2-R was not detectable in these cells (N � 4; Fig. 1B).were homogenized in ice-cold PBS containing protease in-Subsequent experiments were performed with 100 nmol/Lhibitors [1 �g/mL phenylmethylsulfonyl fluoride (PMSF),Ang II stimulation for 12 hours.1 �g/mL leupeptin, 0.2% (wt/vol) aprotinin, 1 �g/mL
antipain] and centrifuged at 1000 g for ten minutes. The Phorbol ester-sensitive PKC rather than Ca2� issupernatants were further centrifuged at 48000 g for 30 involved in Ang II-induced FN mRNA accumulationminutes. The resulting pellet was resuspended in assay To address the role of PKC in AT1-R signaling,buffer containing 50 mmol/L Tris (pH 7.6), 100 mmol/L cells were treated with the PKC inhibitors staurosporinNaCl, 10 mmol/L MgCl2, 1 mmol/L egtazic acid (EGTA), (40 nmol/L) or calphostin-C (50 nmol/L). These inhibi-0.25 mg/mL BSA and protease inhibitors. Protein con- tors interacted with the adenosine 5�-triphosphate (ATP)tent was determined using a BCA Protein Assay Kit binding site and regulatory subunit of PKC, respectively(Pierce) with BSA as a standard. Membrane fractions [37, 38], and were found to completely inhibit AT1-R-(50 �g of protein) were incubated with 125I-[Sar1, Ile8] mediated FN mRNA accumulation (Fig. 2A). PhorbolAng II for the saturation experiment (0.1 to 5 nmol/L) ester (PMA, 100 nmol/L) also markedly up-regulatedin a total assay volume of 300 �L for 90 minutes at 21�C. FN mRNA levels. Ang II-mediated FN mRNA accumu-
lation was partially inhibited by pretreatment withSpecific 125I-[Sar1, Ile8] Ang II binding was determinedBAPTA-AM (12.5 �mol/L) and TMB8 (100 �mol/L),from the difference between counts in the absence andcommonly used as intracellular Ca2� chelators, (�68%presence of 3 �mol/L unlabeled Ang II. Total Ang IIand �71%, respectively, vs. FN mRNA level in the pres-receptor numbers (Bmax) and kD values were determinedence of Ang II), whereas EGTA had no effect on theby Scatchard analysis, and AT1-R and AT2-R were sepa-induction (Fig. 2A). These concentrations of Ca2� chela-rately calculated from total Ang II receptor numbers ontors were sufficient to abolish Ang II-induced intracellu-the basis of distribution ratio determined by nonlinearlar Ca2� mobilization (Fig. 2B). These results suggestedleast-squares regression analysis by inhibition with losar-that PKC-mediated pathways rather than Ca2� signalingtan or CGP42112A using the GraphPad InPlot computerplay a dominant role in Ang II-induced FN mRNA accu-program (Graph Pad, San Diego, CA, USA) [34].mulation in mesangial cells.To measure the amounts of TGF�-1 protein in the
conditioned culture medium, we used an enzyme-linked Induction of FN mRNA by Ang II is regulated byimmunosorbent assay (ELISA) kit (R&D Systems) ac- EGFR-mediated ERK signalingcording to the manufacturer’s instructions. Cells were To examine whether tyrosine kinases and EGFR trans-incubated in serum-free medium for 12 hours and then activation are involved in Ang II-induced FN expression,stimulated with Ang II (100 nmol/L) for 12 hours. Ali- cells were pretreated with genistein 200 �mol/L or herbi-quots of medium were obtained and the assay was re- mycin 1 �mol/L with a strong preference for tyrosine-peated three times. specific kinases [39]. These inhibitors markedly inhibited
Ang II-induced FN mRNA accumulation (Fig. 3A) withStatistical analysis no effects on basal FN mRNA levels (data not shown). A
specific inhibitor of EGFR (AG1478, 1 �mol/L) [19] andThe results are expressed as means � SE. StatisticalERK inactivation by the MEK inhibitor PD98059 abol-analysis was performed by one-way ANOVA followedished Ang II-induced FN mRNA accumulation, althoughby pair-wise contrasts using the Dunnett multiple com-the platelet-derived growth factor receptor (PDGFR)parison test. Data were considered statistically significantinhibitor AG1295 had no effect (Fig. 3A). The specificitywhen P was 0.05.of AG1478 as an inhibitor for EGFR phosphorylationwas well established in our previous studies by testing
RESULTS the effects of a dominant negative mutant of EGFR [19].EGF stimulated FN mRNA levels, and this increase wasAng II-induced FN mRNA level is mediated byblocked by AG1478 and PD98059 (Fig. 3B). The phorbolAT1-R stimulationester also up-regulated FN expression, which was abol-
Angiotensin II dose-dependently induced FN mRNA ished by AG1478 and PD98059.accumulation with an EC50 of �45 nmol/L and a maximal
HB-EGF is processed and released by Ang IIpeak at 100 nmol/L, which were completely blocked bysignaling to up-regulate FN expressionthe AT1-R antagonist RNH6270 (1 �mol/L; Fig. 1A).
Ang II (100 nmol/L) significantly stimulated FN mRNA We next tested whether HB-EGF is actually releasedaccumulation after six hours of incubation, reaching a via proteolytic processing by metalloproteinase after the
addition of Ang II to regulate FN expression. Figure 4Amaximal increase after 12 hours and thereafter main-

Uchiyama-Tanaka et al: Angiotensin II and HB-EGF2156
Fig. 1. Dose-dependent effects of angiotensin II (Ang II) on fibronectin (FN) mRNA accumulation and characterization of Ang II receptors inmesangial cells. (A) Mesangial cells were stimulated by Ang II at the indicated concentrations for 12 hours. Total RNA (15 �g) was separatedon agarose formaldehyde gels and transferred onto nylon membranes for hybridization with 32P-labeled FN or GAPDH cDNA probes. Densitiesof FN mRNA signals were measured by densitometry and normalized relative to those of GAPDH mRNA signals. The results are arbitrarilyindicated as values relative to FN mRNA levels in the unstimulated cells (control). Results shown are means � SE of four separate experimentsand representative data are shown. *P 0.01 vs. the control. RNH is AT1-R antagonist (1 �mol/L). (B) Saturation curve and Scatchard plots formembrane fractions from mesangial cells. Membrane preparation and binding assay were performed as described in the Methods section. B/Fmeans the molar ratio of bound I125–Ang II to free I125-Ang II. Symbols are: (�) total; (�) non-specific; (�) specific.
shows that addition of neutralizing anti-HB-EGF anti- ated by measuring ALP activities in the incubation medium(Fig. 4B). ALP activity was rapidly increased after addi-body, heparin (which competes with cell surface-associ-
ated heparan sulfate proteoglycans as coreceptors for tion of Ang II (1.7-fold at 0.5 min) and reached a 4.1-foldincrease at two minutes. Pretreatment with calphostin-CHB-EGF binding to EGFR) [40], and the metalloprotei-
nase inhibitor batimastat completely inhibited Ang II- or batimastat completely abolished the Ang II-mediatedincrease in ALP activity. Pretreatment with BAPTA-mediated FN mRNA accumulation. Batimastat has a
collagen-mimicking hydroxamate’s structure, which fa- AM tended to reduce the induction of ALP activity byAng II, while AG1478 had no effect. Similar results werecilitates chelation of the zinc ion in the active site of
membrane-bound zinc-endopeptidases, metalloprotei- obtained in three different clones, indicating that Ang IIrapidly induces the proteolytic processing of pro-HB-nases (MMPs), including the interstitial collagenases,
stromelysins 1-3, matrilysin, 92 kD gelatinase B (MMP-9), EGF via metalloproteinases in a PKC-dependent man-ner to stimulate the maturation and release of HB-EGF.and the 72 kD gelatinase A (MMP-2) [41]. HB-EGF also
stimulated FN expression, which was blocked by heparinAng II transphosphorylates EGFR to activate ERKand neutralizing anti-HB-EGF antibody. FN induction
by PMA was abolished by pretreatment with HB-EGF Next, we tested whether Ang II actually tyrosine-phos-phorylates EGFR and activates ERK. Tyrosine-phosphor-and batimastat, indicating that PKC is located upstream
of proHB-EGF processing in the Ang II signaling pathway. ylation of EGFR was maximally activated two minutesafter addition of Ang II, and was completely abolished byTo define the mechanism of HB-EGF processing by
Ang II further, we established mesangial cells stably trans- addition of calphostin-C, heparin, batimastat, or AG1478(Fig. 5A). ERK activation was examined using an anti-fected with an expression vector containing ALP cDNA
between the HB-EGF signal sequence and HB-EGF phospho-ERK antibody. ERK activation was maximallyactivated eight minutes after addition of Ang II. In accor-cDNA. The processing and release of HB-EGF were evalu-

Uchiyama-Tanaka et al: Angiotensin II and HB-EGF 2157
Fig. 2. Effects of protein kinase C (PKC) and Ca2� signals on Ang II-induced FN mRNA levels. (A) Mesangial cells were pretreated with orwithout the PKC inhibitor staurosporin (40 nmol/L), calphostin-C (50 nmol/L), intracellular Ca2� chelator BAPTA-AM (12.5 �mol/L), TMB-8(100 �mol/L) or extracellular Ca2� chelator EGTA (5 mmol/L) for 30 minutes, and then stimulated with Ang II (100 nmol/L) or PMA (100 nmol/L)for 12 hours. Quantification of mRNA levels was performed as described in Fig. 1. Results shown are means � SE of four separate experimentsand representative data are shown. *P 0.01 vs. the control. (B) Mesangial cells were incubated in the presence of fura 2-AM and stimulatedwith Ang II (100 nmol/L). Traces are typical of those from three separate experiments. Intracellular Ca2� levels were expressed in nmol/L.
dance with the results regarding EGFR phosphorylation, with AG1478 or PD98059 completely abolished Ang II-induced FN secretion (Fig. 5C).Ang II-mediated ERK activation was abolished by pre-
treatment with PKC inhibitors, heparin, batimastat orAng II stimulates TGF-� mRNA accumulation andAG1478 (Fig. 5B).secretion leading to induction of FN
Ang II-induced FN mRNA expression results in an Angiotensin II has been shown to induce TGF-� ex-increase in FN protein release pression in mesangial cells [4]. However, it remains to
be determined whether HB-EGF-mediated EGFR acti-To examine whether FN mRNA accumulation resultsin an increase in FN protein level, FN protein secreted vation is involved in TGF-� expression. Figure 6A shows
that Ang II up-regulated TGF-� mRNA expression byinto the medium was analyzed by immunoprecipitationusing anti-FN antibody. Immunoprecipitated FN secreted 1.8-fold, which was abolished by PKC inhibitors, heparin,
batimastat, AG1478 and PD98059, indicating the involve-into medium was increased by 2- and 2.8-fold over thecontrol by Ang II and EGF, respectively. Pretreatment ment of the HB-EGF-EGFR-ERK pathway in Ang II-

Uchiyama-Tanaka et al: Angiotensin II and HB-EGF2158
Fig. 3. Effects of tyrosine kinase inhibitorsand epidermal growth factor receptor (EGFR)antagonist on Ang II-induced FN mRNA accu-mulation. Mesangial cells were pretreated withor without the tyrosine kinase inhibitors gen-istein (200 �mol/L), herbimysin (1 �mol/L), spe-cific inhibitors of EGFR (AG1478, 1 �mol/L)or PDGF (AG1295, 1 �mol/L), MEK inhibi-tor PD98059 (10 �mol/L) for 30 minutes andthen challenged with Ang II (100 nmol/L),EGF (10 ng/mL) or PMA (100 nmol/L) for12 horus. Quantification of mRNA levels wasperformed as described in Fig. 1. Resultsshown are means � SE of four separate exper-iments and representative data are shown.*P 0.01 vs. the control.
mediated TGF-� mRNA expression. The TGF-�1 con- To determine the contribution of released TGF-� toAng II-mediated FN expression, we found that the addi-centrations in the incubation medium were measured.
As shown in the right panel of Figure 6A, the mean tion of neutralizing anti-TGF-� antibody partially inhib-ited Ang II-mediated FN expression (�61% vs. FNamount of active TGF-�1 present in six control cell cul-
tures was 0.13 ng/mL. When stimulated with 100 nmol/L mRNA level in the presence of Ang II), while this con-centration of anti-TGF-� antibody completely blockedAng II, a 3.2-fold increase to 0.42 ng/mL was observed.

Uchiyama-Tanaka et al: Angiotensin II and HB-EGF 2159
Fig. 4. Metalloproteinase cleavage of pro-HB-EGF is involved in Ang II-induced FNmRNA accumulation. (A) Cells were pre-treated for 30 minutes with neutralizing anti-HB-EGF antibody (HB-EGF, 1 �g/mL),heparin (100 �g/mL) and the metallopro-teinase inhibitor batimastat (10 �mol/L), andthen stimulated with Ang II (100 nmol/L),HB-EGF (20 ng/mL) or PMA (100 nmol/L)for 12 hours. Quantification of mRNA levelswas performed as described in Fig. 1. (B) Cellsstably transfected with pALP-HB-EGF werestimulated for two minutes with Ang II (100nmol/L) with or without calphostin-C (50nmol/L), batimastat (10 �mol/L), AG1478 (1�mol/L) or BAPTA-AM (10 �mol/L). ALPactivity in the incubation medium was deter-mined as described in the Methods section.The ALP activities (N � 4 in each experiment)are shown relative to the values of 0 timecontrol arbitrarily given a value of 1. Resultsshown are means � SE of four separate exper-iments and representative data are shown.*P 0.01 vs. the control.
TGF-� (1 ng/mL)-mediated FN expression (Fig. 6B). manner rather than PKC signals in cardiac fibroblasts[19, 20] or VSMC [21]. Daub et al originally found theWe also studied whether the released TGF-� can phos-role of EGFR-mediated signaling in Ras/ERK activationphorylate EGFR to activate ERK. As shown in Figureby G-protein-coupled receptors (GPCRs) [18] and there-6C, TGF-� (1 ng/mL) has the ability to activate EGFRafter several groups reported the involvement of HB-and ERK, and pretreatment with AG1478 abolished thisEGF in GPCR-mediated EGFR transactivation [24–26].action, suggesting that TGF-� released by Ang II signal-Filardo et al demonstrated that estrogen also activateding also contributes to the induction of FN protein byERK via this HB-EGF-mediated EGFR activation [27].activating the EGFR-ERK pathway.However, the actual release of HB-EGF into the incuba-tion medium was not demonstrated in these previous
DISCUSSION studies, and it remains to be determined whether suchFibronectin plays an important role in extracellular a signaling mechanism is operating in the mesangial cells.
matrix accumulation secondary to glomerulosclerosis [29]. The major new findings of the present study are thatAng II induced the synthesis and secretion of FN and (1) the processing and release of HB-EGF are actuallycollagen types I and III from mesangial cells [2, 4]. Recent caused by AT1-R-mediated PKC signaling in a metallo-evidence demonstrated that Ang II-induced ERK activa- proteinase-dependent manner; (2) the released HB-EGFtion was mainly mediated via downstream signaling of binds to EGFR, which transactivates EGFR-mediated
ERK pathway to up-regulate FN expression; (3) EGFREGFR transactivated by AT1-R in a Ca2�-dependent
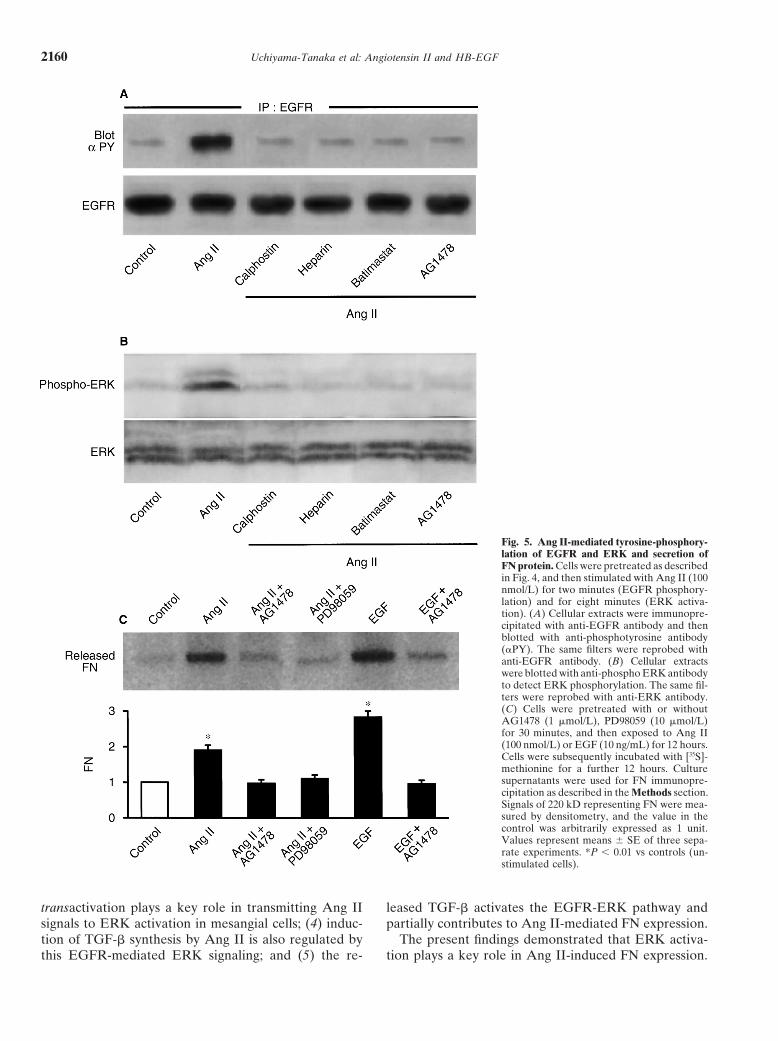
Uchiyama-Tanaka et al: Angiotensin II and HB-EGF2160
Fig. 5. Ang II-mediated tyrosine-phosphory-lation of EGFR and ERK and secretion ofFN protein. Cells were pretreated as describedin Fig. 4, and then stimulated with Ang II (100nmol/L) for two minutes (EGFR phosphory-lation) and for eight minutes (ERK activa-tion). (A) Cellular extracts were immunopre-cipitated with anti-EGFR antibody and thenblotted with anti-phosphotyrosine antibody(PY). The same filters were reprobed withanti-EGFR antibody. (B) Cellular extractswere blotted with anti-phospho ERK antibodyto detect ERK phosphorylation. The same fil-ters were reprobed with anti-ERK antibody.(C) Cells were pretreated with or withoutAG1478 (1 �mol/L), PD98059 (10 �mol/L)for 30 minutes, and then exposed to Ang II(100 nmol/L) or EGF (10 ng/mL) for 12 hours.Cells were subsequently incubated with [35S]-methionine for a further 12 hours. Culturesupernatants were used for FN immunopre-cipitation as described in the Methods section.Signals of 220 kD representing FN were mea-sured by densitometry, and the value in thecontrol was arbitrarily expressed as 1 unit.Values represent means � SE of three sepa-rate experiments. *P 0.01 vs controls (un-stimulated cells).
transactivation plays a key role in transmitting Ang II leased TGF-� activates the EGFR-ERK pathway andpartially contributes to Ang II-mediated FN expression.signals to ERK activation in mesangial cells; (4) induc-
tion of TGF-� synthesis by Ang II is also regulated by The present findings demonstrated that ERK activa-tion plays a key role in Ang II-induced FN expression.this EGFR-mediated ERK signaling; and (5) the re-
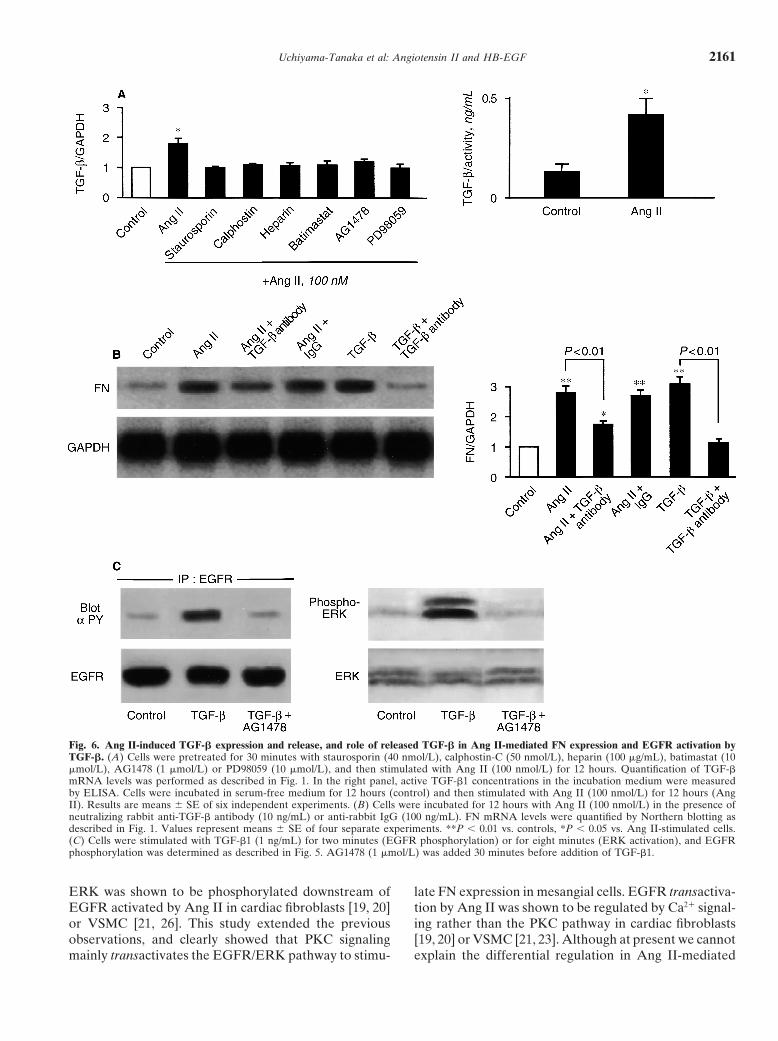
Uchiyama-Tanaka et al: Angiotensin II and HB-EGF 2161
Fig. 6. Ang II-induced TGF-� expression and release, and role of released TGF-� in Ang II-mediated FN expression and EGFR activation byTGF-�. (A) Cells were pretreated for 30 minutes with staurosporin (40 nmol/L), calphostin-C (50 nmol/L), heparin (100 �g/mL), batimastat (10�mol/L), AG1478 (1 �mol/L) or PD98059 (10 �mol/L), and then stimulated with Ang II (100 nmol/L) for 12 hours. Quantification of TGF-�mRNA levels was performed as described in Fig. 1. In the right panel, active TGF-�1 concentrations in the incubation medium were measuredby ELISA. Cells were incubated in serum-free medium for 12 hours (control) and then stimulated with Ang II (100 nmol/L) for 12 hours (AngII). Results are means � SE of six independent experiments. (B) Cells were incubated for 12 hours with Ang II (100 nmol/L) in the presence ofneutralizing rabbit anti-TGF-� antibody (10 ng/mL) or anti-rabbit IgG (100 ng/mL). FN mRNA levels were quantified by Northern blotting asdescribed in Fig. 1. Values represent means � SE of four separate experiments. **P 0.01 vs. controls, *P 0.05 vs. Ang II-stimulated cells.(C) Cells were stimulated with TGF-�1 (1 ng/mL) for two minutes (EGFR phosphorylation) or for eight minutes (ERK activation), and EGFRphosphorylation was determined as described in Fig. 5. AG1478 (1 �mol/L) was added 30 minutes before addition of TGF-�1.
ERK was shown to be phosphorylated downstream of late FN expression in mesangial cells. EGFR transactiva-tion by Ang II was shown to be regulated by Ca2� signal-EGFR activated by Ang II in cardiac fibroblasts [19, 20]
or VSMC [21, 26]. This study extended the previous ing rather than the PKC pathway in cardiac fibroblasts[19, 20] or VSMC [21, 23]. Although at present we cannotobservations, and clearly showed that PKC signaling
mainly transactivates the EGFR/ERK pathway to stimu- explain the differential regulation in Ang II-mediated

Uchiyama-Tanaka et al: Angiotensin II and HB-EGF2162
EGFR activation between mesangial cells, fibroblasts TGF-� as well as whether HB-EGF plays a role in TGF-�and VSMC, it may be partially due to cell-type depen- signaling are necessary.dent differences in expression of PKC isoforms or MMP In summary, we demonstrate that PKC signals acti-isoforms. Further studies are needed to clarify this cell- vated via AT1-R played a critical role in the processingtype specific regulation in Ang II-mediated EGFR acti- and release of HB-EGF in mesangial cells, and that thevation signal. released HB-EGF was an intervening molecule to trans-
We found that processing of HB-EGF by Ang II was mit AT1-R signals to EGFR transactivation. Furthermore,rapidly induced (0.5 min) before maximal activation of we showed that Ang II stimulated the expression andEGFR (2 min), and that the transactivation mechanism release of TGF-� via EGFR-mediated ERK signaling,of EGFR was mostly mediated by HB-EGF processed and that the released TGF-� activates the EGFR-ERKand released via MMPs. HB-EGF is a heparin-binding pathway and partially contributes to Ang II-mediated FNmember of the EGF family and is synthesized as a trans- expression. Thus, Ang II-mediated FN expression is reg-membrane precursor that is then proteolytically pro- ulated by autocrine effects of HB-EGF and TGF-�, sug-cessed into the mature, soluble growth factor, and its gesting a novel paradigm for cross-talk between Ang IIproteolytic processing was reported to be regulated by and growth factor receptor signaling pathways.MMPs in a PKC- and Ca2� -dependent manner [28],consistent with the present observations. Estrogen-medi- ACKNOWLEDGMENTSated activation of the HB-EGF/EGFR pathway was
This study was supported in part by research grants from the Minis-shown to require Src-related kinase activities via Gi pro- try of Education, Science and Culture, Japan, Study Group of Molecu-tein-coupled receptors [27]. These findings suggest that lar Cardiology, Naito Foundation, Clinical Pharmacology Foundation
and Japan Medical Association, and Japan Smoking Foundation, JapanMMPs can be activated by tyrosine kinases associatedHeart Foundation.with PKC or Ca2� signaling. The MMPs, which include
the interstitial collagenases, stromelysins 1-3, matrilysin, Reprint requests to Hiroaki Matsubara, M.D., Department of Medi-cine II, Kansai Medical University, Moriguchi, Osaka 570-8507, Japan.92 kD gelatinase B (MMP-9), and the 72 kD gelatinaseE-mail: [email protected] (MMP-2), constitute a family of matrix-degrading en-
zymes with distinctive extracellular matrix substratespecificities [41]. MMP-2 and MMP-9 share a similar APPENDIXsubstrate specificity, encompassing type IV and V colla-
Abbreviations used in this article are: Ang II, angiotensin II; AT1-R,gen, FN and laminin [42]. MMP-2 acts not only as a angiotensin type 1 receptor; EGFR, epidermal growth factor receptor;matrix-degrading enzyme, but also directly stimulates ERK, extracellular signal-regulated kinase; FN, fibronectin; GPCRs,
G-protein–coupled receptors; HB-EGF, heparin binding-epidermalgrowth by glomerular mesangial cells or enhances syn-growth factor; MEK, ERK kinase; MMP, matrix metalloprotein;thesis of prosclerotic interstitial collagens, and proteo-PDGF, platelet-derived growth factor receptor; PKC, protein kinase C;
lytically-derived fibronectin cleavage fragments were PLC, phospholipase C; PMA, phorbol 12-myristate 13-acetate; TGF-�,transforming growth factor-�; VSMC, vascular smooth muscle cells.shown to be responsible for these growth-promoting and
prosclerotic actions [43]. MMP-2 expression by mesan-gial cells can be stimulated by interleukin-1�, tumor ne- REFERENCEScrosis factor-, and TGF-� [44]. The development of 1. Mendelsohn FA, Millan M, Quirion R, et al: Localization ofsclerotic lesions in chronic glomerulosclerosis is tempo- angiotensin II receptors in rat and monkey kidney by in vitro
autoradiography. Kidney Int 31(Suppl 20):S40–S44, 1987rally correlated with high levels of MMP-2 expression2. Matsubara H, Sugaya T, Murasawa S, et al: Tissue-specific ex-[45]. The present study extended these previous reports pression of human angiotensin II AT1 and AT2 receptors and
suggesting the involvement of MMPs in the pathophysi- cellular localization of subtype using in situ hybridization. Nephron80:25–34, 1998ology of glomerulosclerosis and added new evidence that
3. Goto M, Mukoyama M, Suga S, et al: Growth-dependent inductionthe glomerular MMPs are also involved in Ang II-medi-of angiotensin II type 2 receptor in rat mesangial cells. Hyperten-
ated FN synthesis via processing and release of HB-EGF. sion 30:358–362, 19974. Kagami S, Border W, Miller D, Noble N: Angiotensin II stimu-An important function of Ang II in the kidney includes
lates extracellular matrix protein synthesis through induction ofthe ability of mesangial cells to release growth factors.transforming growth factor-� expression in rat glomerular mesan-
Treatment of mesangial cells with Ang II secreted sig- gial cells. J Clin Invest 93:2431–2437, 1994nificant amounts of TGF-� to cause the synthesis of FN 5. Weiss R, Ramierz A: TGF-� and angiotensin-II-induced mesan-
gial matrix protein secretion is mediated by protein kinase C.and collagen proteins [4, 46]. Our study demonstrates,Nephrol Dial Transplant 13:2804–2813, 1998to our knowledge for the first time, that Ang II stimulates 6. Matsubara H, Inada M: Molecular insights into angiotensin II
the expression and release of TGF-� via EGFR-medi- type 1 and type 2 receptors: Expression, signaling and physiologicalfunction and clinical application of its antagonists. Endocrinolated ERK signaling, and that the released TGF-� alsoJ 45:137–150, 1998activates the EGFR-ERK pathway and contributes to
7. Inagami T, Kitami Y: Angiotensin II receptor: Molecular cloning,Ang II-mediated FN expression. Further studies to de- functions and regulation. Hypertens Res 17:87–97, 1994
8. Berk BC, Corson MA: Angiotensin II signal transduction in vascu-fine the molecules involved in EGFR activation by

Uchiyama-Tanaka et al: Angiotensin II and HB-EGF 2163
lar smooth muscle, role of tyrosine kinases. Circ Res 80:607–616, protease-dependent EGF receptor activation is required for activa-tion of ERK and p38 MAP kinase, but not for JNK. J Biol Chem1997
9. Booz GW, Dostal DE, Singer HA, Baker KM: Involvement of 276:7957–7962, 200127. Filardo E, Quinn JA, Kirby I, et al: Estrogen-induced activation ofprotein kinase C and Ca2� in angiotensin II-induced mitogenesis
of cardiac fibroblasts. Am J Physiol 267: C1308–C1318, 1994 Erk-1 and Erk-2 requires the G protein-coupled receptor homolog,GPR30, and occurs via trans-activation of the epidermal growth10. Schorb W, Peerler TC, Madigan NN, et al: Angiotensin II induced
protein tyrosine phosphorylation in neonatal rat cardiac fibroblasts. factor receptor through release of HB-EGF. Mol Endocrinol 14:1649–1660, 2000J Biol Chem 269:19626–19632, 1994
11. Bernstein KE, Marrero MB: The importance of tyrosine phos- 28. Raab G, Klagsbrum M: Heparin-binding EGF-like growth factor.Biochim Biophys Acta 1333:F179–F199, 1997phorylation in angiotensin II signaling. Trend Cardiovasc Med 6:
179–187, 1996 29. Hynes R: Molecular biology of fibronectin: Annu Rev Cell Biol1:67–90, 198512. Zou Y, Komuro I, Yamazaki T, et al: Protein kinase C, but not
tyrosine kinases or Ras, plays a critical role in angiotensin II- 30. Suzuki J, Matsubara H, Inada M: Rat angiotensin II (type 1A)receptor mRNA regulation and subtype expression in myocardialinduced activation of Raf-1 kinase and extracellular signal-regu-
lated protein kinases in cardiac myocytes. J Biol Chem 27:33592– growth and hypertrophy. Circ Res 73:439–447, 199331. Nio Y, Matsubara H, Murasawa S, et al: Regulation of gene33597, 1996
13. Sadoshima J, Izumo S: The heterotrimetric Gq protein-coupled transcription of angiotensin II receptor subtypes in myocardialinfarction. J Clin Invest 95:46–54, 1995angiotensin II receptor activates p21ras via the tyrosine kinase-
Shc-Grb2-Sos pathway in cardiac myocytes. EMBO J 15:775–787, 32. Murasawa S, Matsubara H, Kijima K, et al: Down-regulation bycAMP of angiotensin II type 2 receptor gene expression in PC121996
14. Sadoshima J, Qiu Z, Morgan JP, Izumo S: Angiotensin II and cells. Hypertens Res 19:271–279, 199633. Tokumura S, Higashiyama S, Endo T, et al: Ectodomain sheddingother hypertrophic stimuli mediated by G protein-coupled recep-
tors activate tyrosine kinase, mitogen-activated protein kinases, of epidermal growth factor receptor ligands is required for kera-tinocyte migration in cutaneous wound healing. J Cell Biol 151:209–and 90-kD S6 kinase in cardiac myocytes. The critical role of Ca2�-
dependent signaling. Circ Res 76:1–15, 1995 220, 200034. Nozawa Y, Haruno A, Oda N, et al: Angiotensin II receptor15. Eguchi S, Matsumoto T, Motley ED, et al: Identification of an
essential signaling cascade for mitogen-activated protein kinase subtypes in bovine and human ventricular myocardium. J Pharma-col Exp Ther 270:566–571, 1994activation by angiotensin II in cultured rat vascular smooth muscle
cells. J Biol Chem 271:14169–14175, 1996 35. Ohkubo N, Matsubara H, Nozawa Y, et al: Angiotensin type2 receptors are re-expressed by cardiac fibroblasts from failing16. Zwick E, Hackel PO, Prenzel N, Ullrich A: The EGF receptor
as central transducer of heterologous signaling systems. Trends myopathic hamster hearts and inhibit cell growth and fibrillar colla-gen metabolism. Circulation 96:3954–3962, 1997Pharmacol Sci 20:408–412, 1999
17. Harris RC: EGF receptor activation by G-protein coupled recep- 36. Tsutusmi Y, Matsubara H, Masaki H, et al: Angiotensin II type2 receptor overexpression activates the vascular kinin system andtors. Kidney Int 8:898–899, 2000
18. Daub H, Weiss FU, Wallasch C, Ullrich A: Role of transactiva- causes vasodilation. J Clin Invest 104:925–935, 199937. Kobayashi E, Nakano H, Morimoto M, Tamaoki T: Calphostintion of the EGF receptor in signalling by G-protein-coupled recep-
tors. Nature 379:557–560, 1996 C (UCN-1028C), a novel microbial compound, is a highly potentand specific inhibitor of protein kinase C. Biochem Biophys Res19. Murasawa S, Mori Y, Nozawa Y, et al: Angiotensin II AT1
receptor-induced extracellular signal-regulated protein kinase acti- Commun 159:548–553, 198938. Herbert JM, Seban E, Maffrand JP: Characterization of specificvation is mediated by Ca2�/calmodulin-dependent transactivation
of epidermal growth factor receptor. Circ Res 82:1338–1348, 1998 binding sites for [3H]-staurosporine on various protein kinases.Biochem Biophys Res Commun 171:189–195, 199020. Murasawa S, Mori Y, Nozawa Y, et al: Role of calcium-sensitive
tyrosine kinase Pyk2/CAKbeta/RAFTK in angiotensin II induced 39. Levitzki A, Gazit A: Tyrosine kinase inhibition: An approach todrug development. Science 267:1782–1787, 1995Ras/ERK signaling. Hypertension 32:668–675, 1998
21. Eguchi S, Numaguchi K, Iwasaki H, et al: Calcium-dependent 40. Higashiyama S, Abraham JA, Klagsbrun M: Heparin-bindingEGF-like growth factor stimulation of smooth muscle cell migra-epidermal growth factor receptor transactivation mediates the an-
giotensin II-induced mitogen-activated protein kinase activation tion: Dependence on interactions with cell surface heparan sulfate.J Cell Biol 122:933–940, 1993in vascular smooth muscle cells. J Biol Chem 273:8890–8896, 1998
22. Moriguchi Y, Matsubara H, Mori Y, et al: Angiotensin II-induced 41. Wojtowicz-Praga SM, Dickson RB, Hawkins MJ: Matrix metal-loproteinase inhibitors. Invest New Drugs 15:61–75, 1997transactivation of epidermal growth factor receptor regulates fi-
bronectin and transforming growth factor-� synthesis via transcrip- 42. Collier IE, Wilhelm SM, Eisen AZ, et al: H-ras oncogene-trans-formed human bronchial epithelial cells (TBE-1) secrete a singletional and posttranscriptional mechanism. Circ Res 84:1073–1074,
1994 metalloprotease capable of degrading basement membrane colla-gen. J Biol Chem 263:6579–6587, 199823. Bokemeyer D: Schmitz U, Kramer HJ: Angiotensin II-induced
growth of vascular smooth muscle cells requires a Src-dependent 43. Turck J, Pollock A, Lovett DH: Gelatinase A is a glomerularmesangial cell growth and differentiation factor. Kidney Int 51:activation of the epidermal growth factor receptor. Kidney Int
58:549–558, 2000 1397–1400, 197744. Marti HP, Lee L, Kashgarian M, Lovett DH: Transforming24. Pranzel N, Zwick E, Daub H, et al: EGF receptor transactivation
by G-protein-coupled receptors requires metalloproteinase cleav- growth factor-beta 1 stimulates glomerular mesangial cell synthesisof the 72-kd type IV collagenase. Am J Pathol 144:82–94, 1994age of proHB-EGF. Nature 402:23–30, 1999
25. Kalmes A, Vesti BR, Daum G, et al: Heparin blockade of throm- 45. Carome MA, Striker LJ, Peten EP, et al: Assessment of 72-kilodal-ton gelatinase and TIMP-1 gene expression in normal and scleroticbin-induced smooth muscle cell migration involves inhibition of
epidermal growth factor (EGF) receptor transactivation by hepa- murine glomeruli. J Am Soc Nephrol 5:1391–1396, 199446. Sun Y, Zhang JQ, Ramires F: Local angiotensin II and trans-rin-binding EGF-like growth factor. Circ Res 87:92–98, 2000
26. Eguchi S, Dempsey PJ, Frank GD, et al: Activation of MAP forming growth factor-�1 in renal fibrosis of rats. Hypertention35:1078–1084, 2000kinases by angiotensin II in vascular smooth muscle cells: Metallo-