b2 bradykinin receptor immunoreactivity in rat brain
TRANSCRIPT

B2 Bradykinin ReceptorImmunoreactivity in Rat Brain
ER-YUN CHEN,1 DWAINE F. EMERICH,2 RAYMOND T. BARTUS,2AND
JEFFREY H. KORDOWER1*1Research Center for Brain Repair and Department of Neurological Sciences, Rush
Presbyterian Medical Center, Chicago, Illinois 606122Alkermes Inc., Cambridge, Massachusetts 02139
ABSTRACTBradykinin has long been known to exist in the central nervous system and has been
hypothesized to mediate specific functions. Despite an increasing understanding of thefunctions of bradykinin, little is known about the cell types expressing the bradykininreceptor within the brain. The present investigation employed a monoclonal antibody di-rected against the 15-amino-acid portion of the C-terminal of the human bradykinin B2receptor to establish the cellular distribution of bradykinin B2 receptor immunoreactivity inthe rat brain. Bradykinin B2 receptor immunoreactivity was ubiquitously and selectivelyobserved in neurons, including those within the olfactory bulb, cerebral cortex, hippocampus,basal forebrain, basal ganglia, thalamus, hypothalamus, cerebellum, and brainstem nuclei.Bradykinin B2 receptor immunoreactivity was also present in the circumventricular organsincluding choroid plexus, subfornical organ, median eminence, and area postrema. Double-labeling experiments colocalizing the bradykinin B2 receptor with the neuronal markerNeuN or the astrocytic marker glial fibrillary acidic protein revealed that virtually 100% ofthe bradykinin B2 receptor-immunoreactive positive cells were neurons. The widespreaddistribution of bradykinin B2 receptor immunoreactivity in neuronal compartments suggestsa greater than previously appreciated role for this peptide in neuronal function. J. Comp.Neurol. 427:1–18, 2000. © 2000 Wiley-Liss, Inc.
Indexing terms: bradykinin; kininogen; immunohistochemistry
Bradykinin is a nonapeptide formed from kininogen bythe serine protease kallikrein and plays important roles inthe pathogenesis of inflammation, pain, asthma, hyper-tension, and other diseases (Marceau et al., 1980, 1983;Regoli and Barabe, 1980; Bhoola et al., 1992; Hall, 1992;Dray and Perkins, 1993; Walker et al., 1995; Kaplan et al.,1998). In addition to its well-documented effects in theperiphery, bradykinin may also act as a modulator or/andneurotransmitter in the central nervous system (CNS;Snyder 1980; Yaksh and Hammond 1982; Yano et al.,1984; Yau et al., 1986). The biological effects of bradykininare mediated by its interactions with two distinct brady-kinin receptor subtypes, the bradykinin B1 and B2 recep-tors. The bradykinin B1 receptor is expressed at very lowlevels and has a limited distribution under normal condi-tions but can be induced and up-regulated under certainpathological conditions, such as tissue damage or inflam-mation (Marceau et al., 1983, 1997; Bathon and Proud,1991; Marceau, 1995). The bradykinin B2 receptor is con-stitutively and widely expressed in various mammaliantissues, where it is believed to mediate the majority of the
biological actions of bradykinin (Kaplan et al., 1998). Ex-pression and cloning of human bradykinin B2 receptorcDNA that is highly homologous to rat bradykinin B2receptor cDNA (81%) reveals that this receptor is com-posed of 364 amino acids, with a molecular mass of 41,442Da, and belongs to the G-protein-coupled superfamily ofreceptors. The receptor is characterized by seven trans-membrane regions interconnected by three extracellularand three intracellular loops (Burch and Axelrod, 1987;McEachern et al., 1991; Hess et al., 1992; 1994a,b; Powellet al., 1993; Menke et al., 1994; Bachvarov et al., 1995).
The various components of the kallikrein-kinin systemhave been identified in the mammalian nervous system.
*Correspondence to: Dr. Jeffrey H. Kordower, Department of Neurolog-ical Sciences, Rush Presbyterian Medical Center, 2242 West HarrisonStreet, Chicago IL 60612. E-mail: [email protected]
Received 1 February 2000; Revised 28 March 2000; Accepted 17 April2000
THE JOURNAL OF COMPARATIVE NEUROLOGY 427:1–18 (2000)
© 2000 WILEY-LISS, INC.

Correa and coworkers (1979) first reported thatbradykinin-like immunoreactive neuronal cells were local-ized to the hypothalamus and that bradykinin-like immu-noreactive fibers were distributed within the periaqueduc-tal gray matter, hypothalamus, perirhinal and cingulatecortices, ventral portion of caudate-putamen, and lateralseptal area of the rat brain. These initial observationswere confirmed and extended by many demonstrating thewidespread binding, protein immunoreactivity, andmRNA of the kallikrein-kinin system in nervous tissues,including the cerebral cortex, thalamus, hypothalamus,cerebellum, brainstem, pituitary, pineal gland, and spinalcord (Kariya et al., 1981, 1985; Perry and Snyder, 1984;Chao et al., 1987; Kizuki et al., 1994; Lopes et al., 1995;Raidoo et al., 1996a).
The majority of studies examining bradykinin receptorswithin the mammalian CNS employed receptor bindingtechniques to reveal a widespread distribution of bindingsites (Steranka et al., 1988; Fujiwara et al., 1988, 1989;Privitera et al., 1991; Lopes et al., 1995; Murone et al.,1996, 1997). Steranka and coorkers (1988) localized[3H]bradykinin binding sites to the substantia gelatinosa,dorsal root, and a subset of small cells in both the dorsalroot and trigeminal ganglia of the guinea pig. Specifichigh-affinity bradykinin binding has also been seen in thepons, medulla oblongata, cerebral cortex, hippocampus,and spinal cord of the guinea pig brain (Fujiwara et al.,1989). [125I-Tyr8]bradykinin-specific binding sites havebeen localized in the medulla oblongata to the nucleus ofthe solitary tract, the area postrema, the dorsal motornucleus of the vagus nerve, the caudal subnucleus of thespinal trigeminal nucleus (Privitera et al., 1991), and thesuperficial laminae of the dorsal horn (Lopes et al., 1995).Most recently, using a radiolabelled and specific bradyki-nin B2 receptor antagonist analogue, [125I]HPP-HOE140,Murone and coworkers (1996, 1997) found that the brady-kinin B2 receptor was widely distributed in both motorand sensory areas of sheep brain.
Together these data demonstrate a wide distribution ofbradykinin B2 receptors in the CNS, implicating a func-tionally diverse endogenous system. However, these stud-ies have not extensively characterized the cellular distri-bution of the bradykinin B2 receptor. Using specificpolycolonal antibodies, Raidoo and coworkers (Raidoo etal., 1996b; Raidoo and Bhoola, 1997) localized bradykininB2 receptors to the neurons of brainstem, basal nuclei,cerebral cortex, and hypothalamus in the human brain.However, a precise mapping of bradykinin B2 receptordistribution has yet to be described. The present studyemployed a monoclonal antibody directed to the 15-amino-acid portion of the C-terminal of the human B2 bradykininreceptor to investigate the cellular distribution of brady-kinin B2 receptor immunoreactivity throughout the ratbrain.
MATERIALS AND METHODS
Subjects
Adult male Fischer 344 rats (Harlan; 200–250 g; n 5 6)were used for this study. They were housed in pairs in atemperature- and humidity-controlled colony room thatwas maintained on a 12:12 hours light-dark cycle. Foodand water were available ad libitum throughout the ex-periment. All experiments were conducted in accordancewith National Institutes of Health guidelines.
Preparation of tissues
Rats were administered an overdose of sodium pento-barbital (120 mg/kg) and then perfused transcardiallywith 0.9% saline, followed by fixation with a 4% parafor-maldehyde in 0.1 M sodium phosphate buffer. Brains wereremoved from the calvaria, postfixed for 1 hour in thesame fixative, and then cryoprotected overnight in 30%sucrose in 0.1 M sodium phosphate buffer (pH 7.2) at 4°C.The brains were sectioned frozen at a thickness of 40 mmon a sliding knife microtome and stored at 220°C in cryo-protectant.
Immunocytochemistry
Three primary antibodies were used in this study, in-cluding mouse antibradykinin B2 receptor monoclonal an-tibody (Transduction Laboratories, Lexington, KY), mouseantineuronal nuclear antigen (NeuN) monoclonal anti-body (Chemicon, Temecula, CA), and rabbit antiglialfibrillary acidic protein (GFAP) polycolonal antibody(Dako, Glostrup, Denmark). Two adjacent series of sec-tions from each brain were processed for immunohisto-chemical visualization of both bradykinin B2 receptor andNeuN, via the biotin-labeled antibody procedure (Hsu etal., 1981). Briefly, following several washes in the Tris-buffered saline (pH 7.2, TBS) solution containing 0.05%Triton X-100 (TBS-Tx), sections were incubated for 20minutes in a TBS solution containing 0.1 M sodium perio-date to eliminate endogenous peroxidase activity. Afterthree washes in TBS-Tx, background staining was theninhibited by a 1 hour incubation in 5% normal horseserum and 2% bovine serum albumin, after which the twoprimary antibodies at the appropriate dilution (1:1,000)were applied for 20 hours at room temperature in a 0.1 Msodium phosphate-buffered saline (pH 7.2) solution con-taining 0.4% Triton X-100 and 3% normal horse serum.After six washes in TBS-Tx, sections were then sequen-tially incubated in the biotinylated horse antimouse IgG(Vector Laboratories, Burlingame, CA; 1:200) for 1 hourand the Elite avidin-biotin complex (ABC Kits; VectorLaboratories; 1:500) for 75 minutes, separated by threewashes in TBS-Tx. The chromogen solution that com-pleted the reaction consisted of 0.05% 39,3-diaminobenzidine (DAB), 0.005% H2O2, and 2.5% nickel IIsulfate. Sections were mounted on gelatin-coated slides,dehydrated through graded alcohol (50%, 70%, 95%, 99%),cleared in xylene, and coverslipped with Cytoseal.
Double-label procedures
To assess whether B2 receptor immunoreactivity waslocalized to neurons or astrocytes, we employed double-label immunofluorescence and double-label immunoperox-idase protocols colocalizing bradykinin B2 receptor-containing cells with the neuronal marker NeuN or theastrocyte marker GFAP. For the double-immuno-fluorescences protocol, a dual-channel confocal laser scan-ning microscope with an argon/krypton laser was used forimaging the double-labeled fluorescent tissues. Two differ-ent fluorescent probes were assessed simultaneously orseparately allowing the two different antigens to be de-tected in the same section. Because the first primary an-tibody was a mouse antibradykinin B2 receptor monoclo-nal antibody, and the second primary antibody was arabbit anti-GFAP polyclonal antibody, an indirect immun-ofluoresence double-label procedure was used to double-
2 E.-Y. CHEN ET AL.
label the bradykinin B2 receptor with GFAP. Followingseveral washes in a 0.1 M sodium phosphate-bufferedsaline solution (PBS) containing 0.1% Triton X-100 (PBS-Tx), pH 7.4, one series of sections was incubated for 1 hourin the PBS-Tx containing 5% normal goat serum and 2%bovine serum albumin to inhibit background staining andthen incubated in the PBS-Tx containing 3% normal goatserum and the first primary antibody to bradykinin B2receptor (1:100) overnight at room temperature. Sectionswere then washed in PBS-Tx and incubated sequentiallyin PBS-Tx containing 3% normal goat serum and the goatanti-mouse antibody coupled to Cy2 (1:200; Jackson Im-munoresearch Laboratories, Inc., West Grove, PA) for 1hour. Following several washes in PBS-Tx, sections wereincubated for 1 hour in the PBS-Tx containing 5% normalgoat serum and 2% bovine serum albumin to inhibit back-ground staining again. Sections were then incubated inthe PBS-Tx containing 3% normal goat serum and the
second primary antibody to GFAP (1:1,000) overnight atroom temperature. Sections were then washed in PBS-Txand incubated sequentially in PBS-Tx containing 3% nor-mal goat serum and the goat anti-rabbit antibody coupledto Cy3 (1:400; Jackson Immunoresearch Laboratories,Inc.) for 1 hour. Following several washes in PBS, sectionswere mounted on gelatin-coated slides, dehydratedthrough graded alcohol (50%, 70%, 95%, 99%), cleared inxylene, and coverslipped with DPX.
Because the first primary antibody (bradykinin B2 re-ceptor) and the second primary antibody (NeuN) are bothmonoclonal antibodies that were raised in the same spe-cies (mouse), it is difficult to use a simple indirect immun-ofluoresence double-label procedure to double label thesetwo antigens. With this procedure, the second primaryantibody could bind to the first secondary antibody, andthe second secondary antibody could also bind to the firstprimary antibody. To avoid any potential artificial stain-
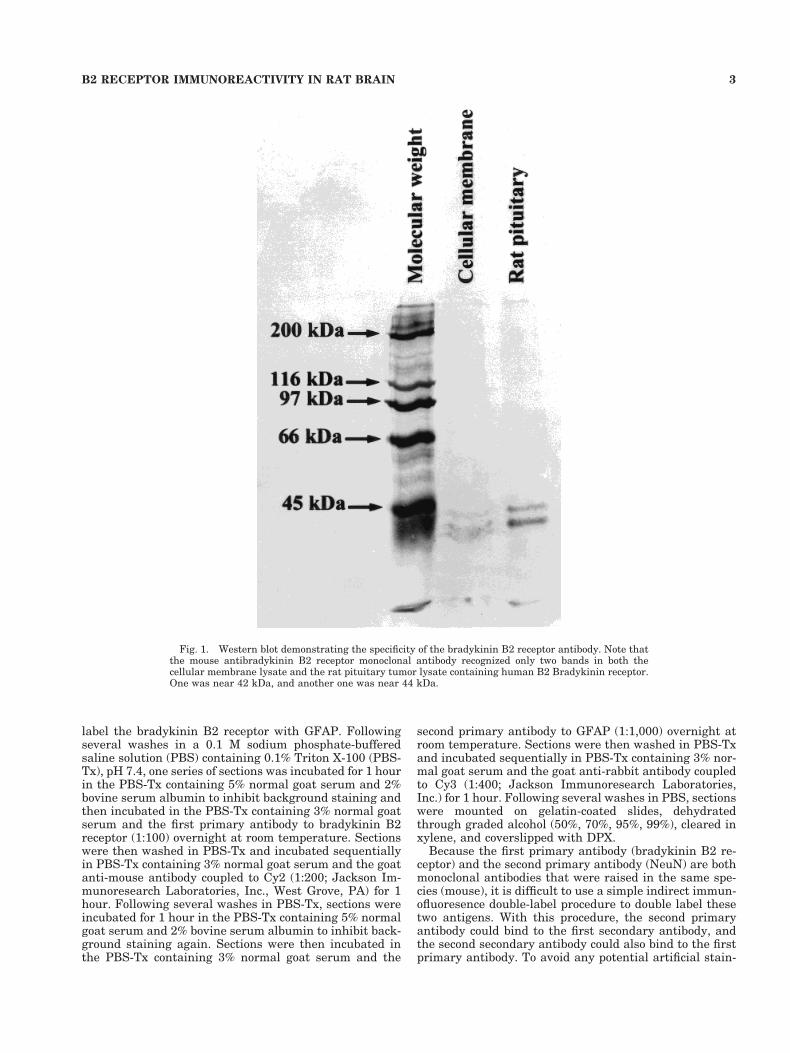
Fig. 1. Western blot demonstrating the specificity of the bradykinin B2 receptor antibody. Note thatthe mouse antibradykinin B2 receptor monoclonal antibody recognized only two bands in both thecellular membrane lysate and the rat pituitary tumor lysate containing human B2 Bradykinin receptor.One was near 42 kDa, and another one was near 44 kDa.
3B2 RECEPTOR IMMUNOREACTIVITY IN RAT BRAIN
ing, a modification of the indirect immunofluoresencedouble-label procedure (Wessel and McClay, 1986; Fran-zusoff et al., 1991; Lewis Carl et al., 1993; Negoescu et al.,1994) was employed. In this procedure, the most impor-tant step is to use monovalent Fab fragments of the sec-ondary antibody to cover the surface of the first primaryand secondary antibodies sterically after the first antigenlabeling is complete. This step prevents the capture of thesecond primary antibody by residual free binding sites ofthe first secondary antibody and the linkage of the secondsecondary antibody to the first primary antibody. In thisprocedure, sections were first processed for the immuno-fluoresence labeling of bradykinin B2 receptor in a man-ner identical to the immunofluoresence double-label pro-cedure of bradykinin B2 receptor and GFAP. Then, thesections were incubated with normal mouse serum con-taining normal mouse IgG for 1 hour. This step is tosaturate any open antigen binding sites on the first sec-ondary antibody (goat anti-mouse antibody coupled toCy2), preventing it from binding to the second primaryantibody (NeuN). After several washes in PBS-Tx, sec-tions were incubated sequentially in PBS-Tx containingan excess unconjugated Fab fragments of goat anti-mouseIgG (20 mg/ml; Jackson Immunoresearch Laboratories,
Inc.) for 1 hour. This step is to saturate any open antigenbinding sites of the first primary antibody (bradykinin B2receptor) as well as the normal mouse IgG preventing itfrom binding to the second secondary antibody (goat anti-mouse antibody coupled to Cy3). After several washes inPBS-Tx, sections were incubated for 1 hour in the PBS-Txcontaining 5% normal goat serum and 2% bovine serumalbumin to inhibited the background staining again andthen incubated in the PBS-Tx containing 3% normal goatserum and the second primary antibody (mouse anti-NeuN monoclonal antibody) at the appropriate dilution(1:200) overnight at room temperature. Sections werethen washed in PBS-Tx and incubated sequentially inPBS-Tx containing 3% normal goat serum and the goatanti-mouse antibody coupled to Cy3 (1:400; Jackson Im-munoresearch Laboratories, Inc.) for 1 hour. Followingseveral washes in PBS, sections were mounted on gelatin-coated slides, dehydrated through graded alcohol (50%,70%, 95%, 99%), cleared in xylene, and coverslipped withDPX.
Immunofluoresence double-labeled sections were exam-ined by a Fluoview laser confocal system equipped with anOlympus microscope and an argon/krypton lasers. Thedifferent magnification images were obtained by a dual-
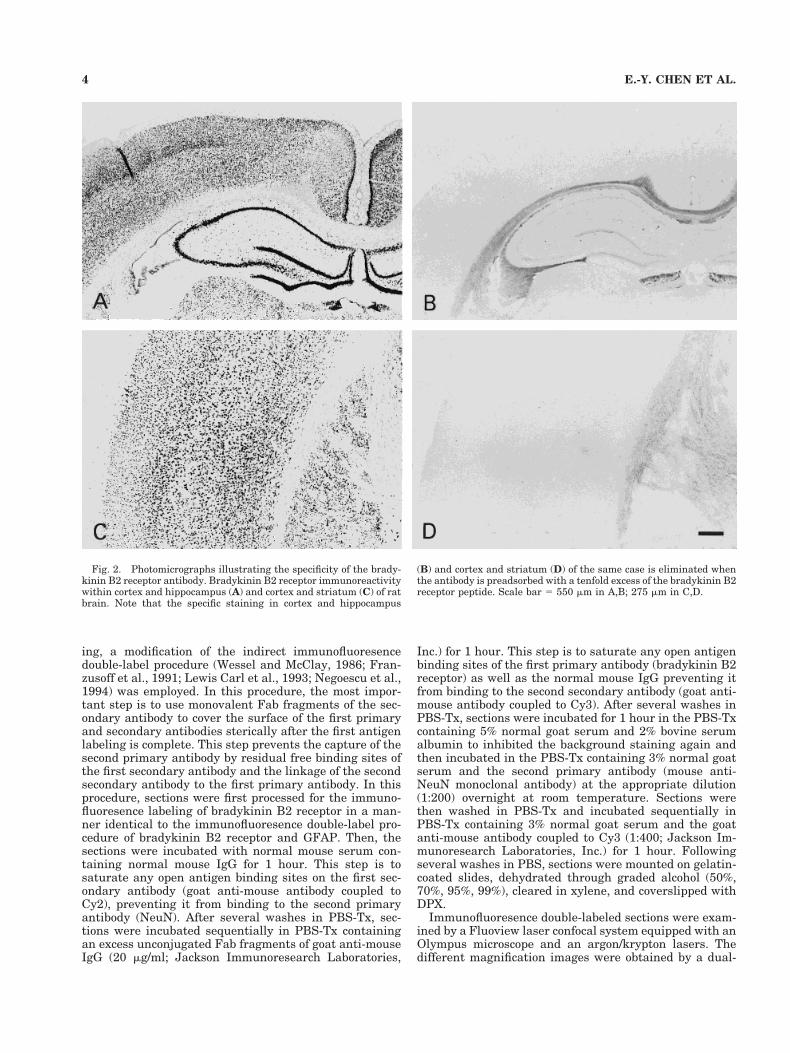
Fig. 2. Photomicrographs illustrating the specificity of the brady-kinin B2 receptor antibody. Bradykinin B2 receptor immunoreactivitywithin cortex and hippocampus (A) and cortex and striatum (C) of ratbrain. Note that the specific staining in cortex and hippocampus
(B) and cortex and striatum (D) of the same case is eliminated whenthe antibody is preadsorbed with a tenfold excess of the bradykinin B2receptor peptide. Scale bar 5 550 mm in A,B; 275 mm in C,D.
4 E.-Y. CHEN ET AL.
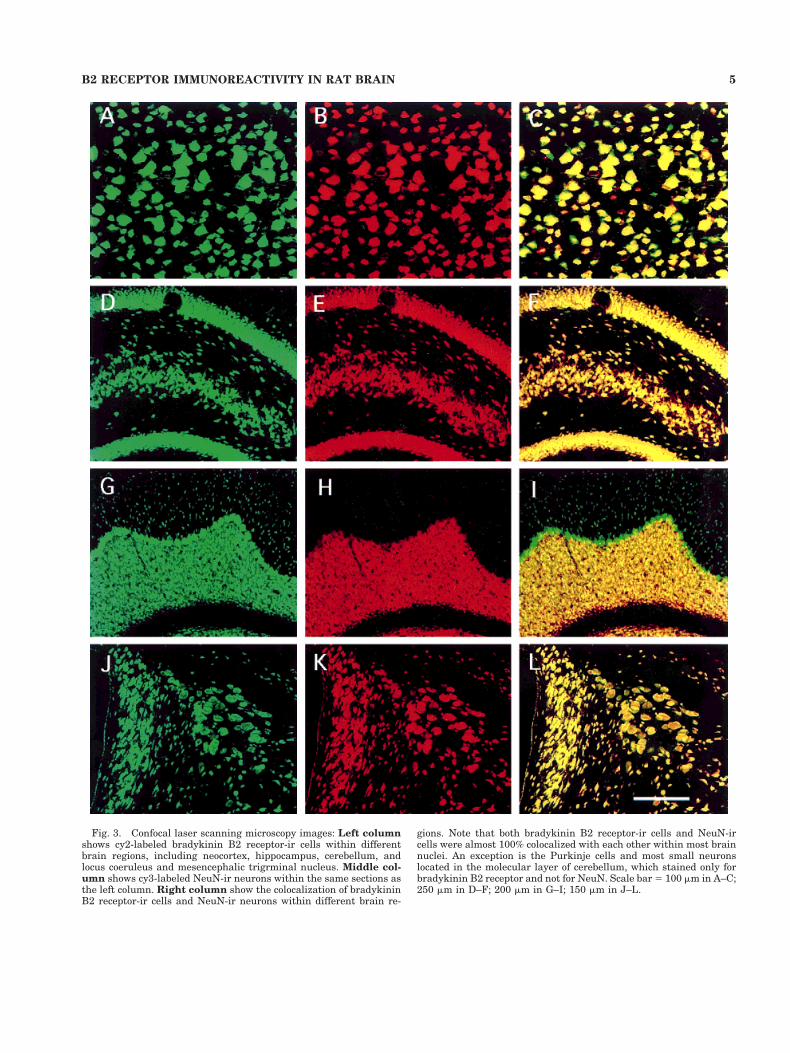
Fig. 3. Confocal laser scanning microscopy images: Left columnshows cy2-labeled bradykinin B2 receptor-ir cells within differentbrain regions, including neocortex, hippocampus, cerebellum, andlocus coeruleus and mesencephalic trigrminal nucleus. Middle col-umn shows cy3-labeled NeuN-ir neurons within the same sections asthe left column. Right column show the colocalization of bradykininB2 receptor-ir cells and NeuN-ir neurons within different brain re-
gions. Note that both bradykinin B2 receptor-ir cells and NeuN-ircells were almost 100% colocalized with each other within most brainnuclei. An exception is the Purkinje cells and most small neuronslocated in the molecular layer of cerebellum, which stained only forbradykinin B2 receptor and not for NeuN. Scale bar 5 100 mm in A–C;250 mm in D–F; 200 mm in G–I; 150 mm in J–L.
5B2 RECEPTOR IMMUNOREACTIVITY IN RAT BRAIN
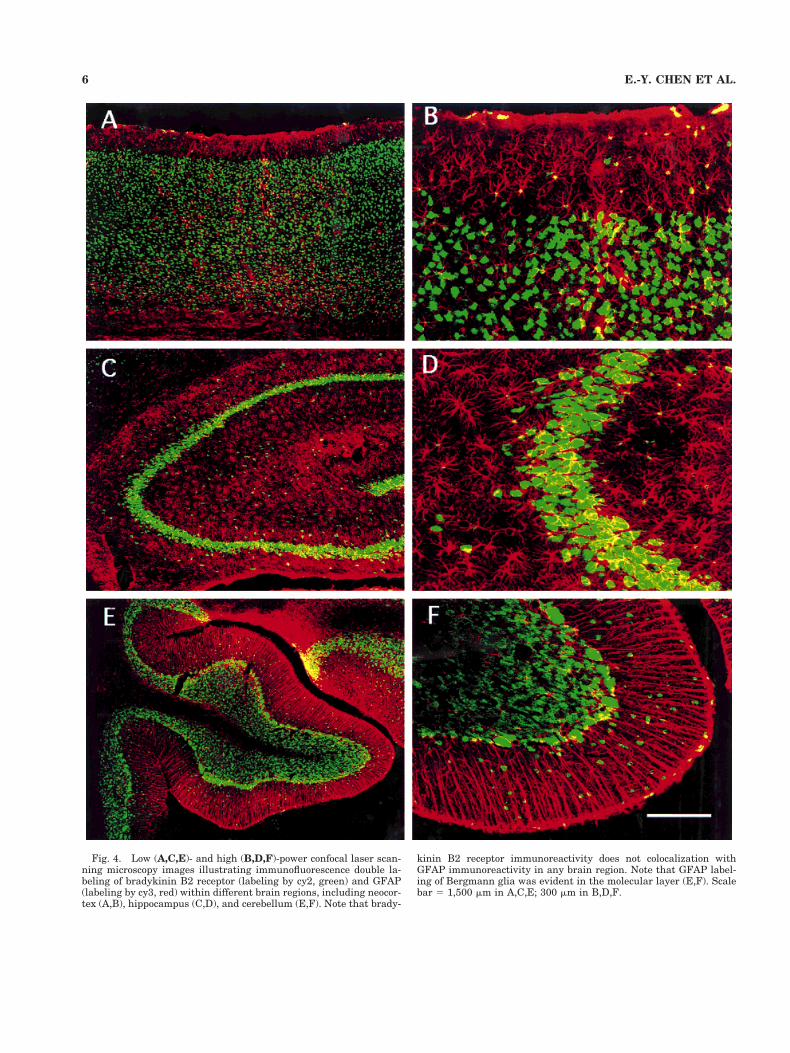
Fig. 4. Low (A,C,E)- and high (B,D,F)-power confocal laser scan-ning microscopy images illustrating immunofluorescence double la-beling of bradykinin B2 receptor (labeling by cy2, green) and GFAP(labeling by cy3, red) within different brain regions, including neocor-tex (A,B), hippocampus (C,D), and cerebellum (E,F). Note that brady-
kinin B2 receptor immunoreactivity does not colocalization withGFAP immunoreactivity in any brain region. Note that GFAP label-ing of Bergmann glia was evident in the molecular layer (E,F). Scalebar 5 1,500 mm in A,C,E; 300 mm in B,D,F.
6 E.-Y. CHEN ET AL.
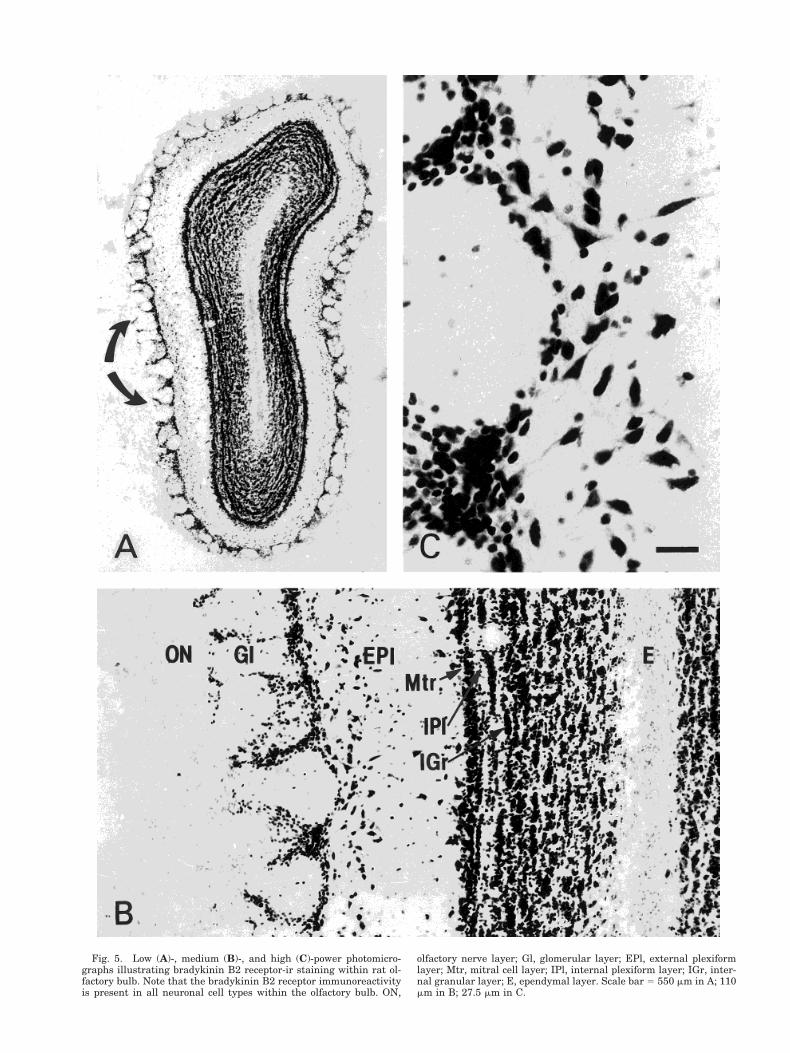
Fig. 5. Low (A)-, medium (B)-, and high (C)-power photomicro-graphs illustrating bradykinin B2 receptor-ir staining within rat ol-factory bulb. Note that the bradykinin B2 receptor immunoreactivityis present in all neuronal cell types within the olfactory bulb. ON,
olfactory nerve layer; Gl, glomerular layer; EPl, external plexiformlayer; Mtr, mitral cell layer; IPl, internal plexiform layer; IGr, inter-nal granular layer; E, ependymal layer. Scale bar 5 550 mm in A; 110mm in B; 27.5 mm in C.
channel confocal laser scanning the sections, and theseimages were collected using appropriate emission filters:for Cy3, excitation at 568 nm and a BA585IF filter; forCy2, excitation at 488 nm and two emission filters,BA510IF and BA550RIF.
To confirm the findings seen with the double-immunofluorescence protocol, a double-label immunoper-oxidase procedure was employed using nickel-DAB-H2O2and DAB-H2O2 as chromogens for the two different anti-gens (Hsu and Soban, 1982; Kordower et al., 1988; Mufsonet al., 1989, 1993, 1998). Nickel-DAB-H2O2 produces ablack reaction product that masks the antibodies previ-ously added to the sections and allows a second antigen tobe detected with a subsequently added antibody. DAB-H2O2 as the second chromogen produces a brown reactionproduct allowing an easily identifiable two-color immuno-histochemical profile. Briefly, sections were first processedfor bradykinin B2 receptor immunoreactivity (1:1,000) asdescribed above. Nickel-DAB-H2O2 was used as the chro-mogen, yielding a black reaction product within the cellnucleus and cytoplasm. Following completion of the firstreaction, sections were incubated in 0.5% H2O2 for 30minutes to eliminate excess peroxidase activity. Then, thesections were processed for NeuN (1:1,000) or GFAP(1:2,000) immunoreactivity using DAB-H2O2 as the chro-
mogen to yield a brown reaction product. Sections weremounted on gelatin-coated slides, dehydrated throughgraded alcohol (50%, 70%, 95%, 99%), cleared in xylene,and coverslipped with Cytoseal.
Control experiments
Multiple control experiments including primary dele-tions, substitutions, preadsorbtions, and Western blottingwere performed. First, control sections were processed forbradykinin B2 receptor, NeuN, and GFAP immunoreac-tivity in a manner identical to that described above exceptfor the deletion of the primary antibody or the substitutionof an irrelevant IgG matched for protein concentration. Itis important to note that, even though staining was elim-inated in sections with the primary antibody deleted, anirrelevant IgG substituted or, with the antibody pread-sorbed against bradykinin B2 receptor peptide, the poten-tial for antiserum to react with structurally related pro-teins cannot be excluded. Thus, a degree of caution ininterpreting data from immunohistochemical procedure isalways warranted. In this regard, the term immunoreac-tivity in this study refers to -like immunoreactivity. Addi-tional sections were immunohistochemically processed asdescribed above except that the bradykinin B2 receptorantibody was preadsorbed with tenfold or 20-fold excess of
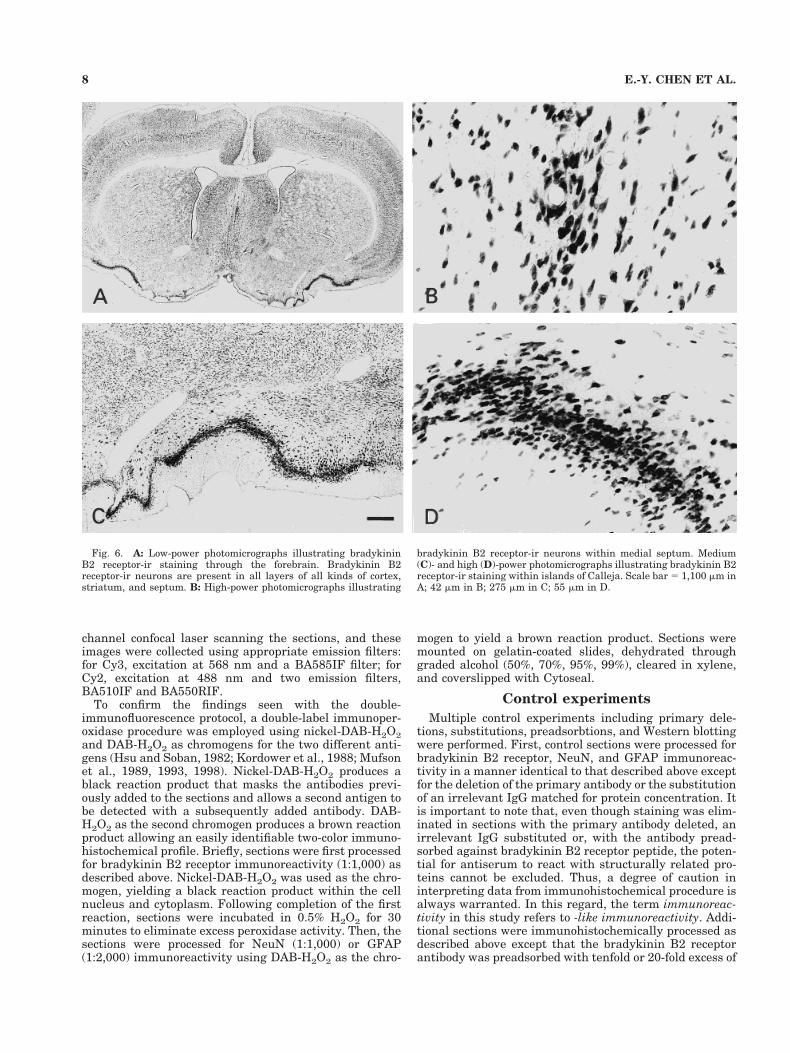
Fig. 6. A: Low-power photomicrographs illustrating bradykininB2 receptor-ir staining through the forebrain. Bradykinin B2receptor-ir neurons are present in all layers of all kinds of cortex,striatum, and septum. B: High-power photomicrographs illustrating
bradykinin B2 receptor-ir neurons within medial septum. Medium(C)- and high (D)-power photomicrographs illustrating bradykinin B2receptor-ir staining within islands of Calleja. Scale bar 5 1,100 mm inA; 42 mm in B; 275 mm in C; 55 mm in D.
8 E.-Y. CHEN ET AL.

bradykinin B2 receptor peptide (Transduction Laborato-ries) for 48 hours prior to incubation.
To establish further the specificity of the bradykinin B2receptor antibody, 4 ml of cellular membrane lysate con-taining human B2 bradykinin receptor (Receptor Biology,Inc., Beltsville, MD) or 6 ml rat pituitary tumor lysate(Transduction Laboratories) were heated for 2 minutes at100°C in the sample buffer containing 0.02% SDS, 0.01%glycerol, 10% 1 M Tris (pH 6.8), 0.01% bromophenol blue,and 5% 2-ME. Samples were cooled on ice and spun for 2minutes in an Eppendorf microcentrifuge (model 5415C).Forty microliters of each sample and diluted biotinylatedhigh-molecular-weight protein standards (Bio-Rad, Rich-mond, CA) were loaded into a 10% SDS gel. The gel wasrun in the Bio-Rad minigel tank at 40 V initially to let allprotein go through the stacking gel. Once the blue frontline reached the running gel, voltage was increased to 80V until the blue front line reached the limit of running gel.Proteins in the gel were blotted overnight onto a sheet ofnitrocellulose membrane in the minigel tank in Towbinsolution (25 mM Tris, 192 mM glycine, 20% methanol, pH8.3). The membrane was washed three times in TBS (20mM Tris-HCl, 500 mM NaCl, 0.05% Tween-20, pH7.5) andblocked for 2 hours in TBS containing 3% nonfat dry milk.The membrane was incubated in mouse antibradykininB2 receptor monoclonal antibody (1:1,000) overnight atroom temperature in TBS. After six washes in TBS, mem-brane was then sequentially incubated in the biotinylatedhorse anti-mouse IgG (1:200; Vector Laboratories) for 1hour and the Elite avidin-biotin complex (ABC Kits; 1:500;Vector Laboratories) for 75 minutes separated by threewashes in TBS. After thorough washing in TBS, the colorwas developed in DAB solution consisting of 0.05% DABand 0.5% H2O2. Color development was stopped by thor-ough wash with TBS and stored in TBS buffer prior tophotographic documentation of the data.
RESULTS
Specificity of immunostaining
Western blot control experiments confirmed that themouse antibradykinin B2 receptor monoclonal antibodyonly recognized two bands in both the cellular membranelysate and the rat pituitary tumor lysate containing hu-man B2 bradykinin receptor (Fig. 1). One was near 42kDa(rat bradykinin B2 receptor is a 366-amino-acid proteinwith a molecular mass of 41,696 Da), and the other wasnear 44kDa, which likely reflects the phosphorylated formof the receptor. The specificity of this antibody has beenconfirmed; specific bradykinin B2 receptor immunoreac-tivity was observed with a widespread distribution of ratbrain using the mouse anti-bradykinin B2 receptor mono-clonal antibody. Specific staining was eliminated in sec-tions in which the primary antibody solvent or irrelevantIgG was substituted for the primary bradykinin B2 recep-tor antibody. Moreover, specific staining was completelyeliminated when the bradykinin B2 receptor antibody waspreadsorbed with tenfold bradykinin B2 receptor peptide(Fig. 2). Both double-label procedures using Nickel-DAB-H2O2 and DAB-H2O2 as the chromogen and double-immunofluoresence-stained sections in which one primaryantibody was omitted revealed staining for only the ap-propriate antigens.
General characteristics
Immunostaining with mouse antibradykinin B2 re-ceptor monoclonal antibodies revealed that bradykininB2 receptor immunoreactivity was present throughoutrat brain nuclei, including those within the olfactorybulb, cerebral cortex, hippocampus, basal forebrain,basal ganglia, thalamus, hypothalamus, cerebellum,and brainstem nuclei (Figs. 2A,C, 3A,D,G,J, 4 –14). Bra-dykinin B2 receptor immunoreactivity was also ob-served in the circumventricular organs such as the sub-fornical organ, median eminence, and area postrema aswell as the choroid plexus (Fig. 14). Immunoreactivityfor the bradykinin B2 receptor was primarily localizedto cell bodies but was occasionally observed within bothcell bodies and proximal dendrites in select neuronalpopulations such as pyramidal cells in layer V of neo-cortex (Fig. 7B), neurons of all cranial nerve motornuclei (Figs. 10B, 11D, 12F, 13B), and in magnocellularneurons of the red nucleus (Fig. 10D). Although immu-noreactivity for the bradykinin B2 receptor was de-tected in both the cytoplasm and the nucleus, the nu-cleus was generally more strongly labeled. This wasparticularly true in the large pyramidal cells in layer Vof neocortex (Fig. 7B), Purkinje cells (Fig. 12C), and
Fig. 7. Low (A)- and high (B)-power photomicrographs illustratingbradykinin B2 receptor-ir staining within cortex. Bradykinin B2 re-ceptor immunoreactivity is present in all layers of neocortex andcingulate cortex. Note bradykinin B2 receptor-ir staining in the nu-cleus is markedly stronger than in the cytoplasm in pyramidal cells inlayer V of neocortex (B). Scale bar 5 345 mm in A; 55 mm in B.
9B2 RECEPTOR IMMUNOREACTIVITY IN RAT BRAIN
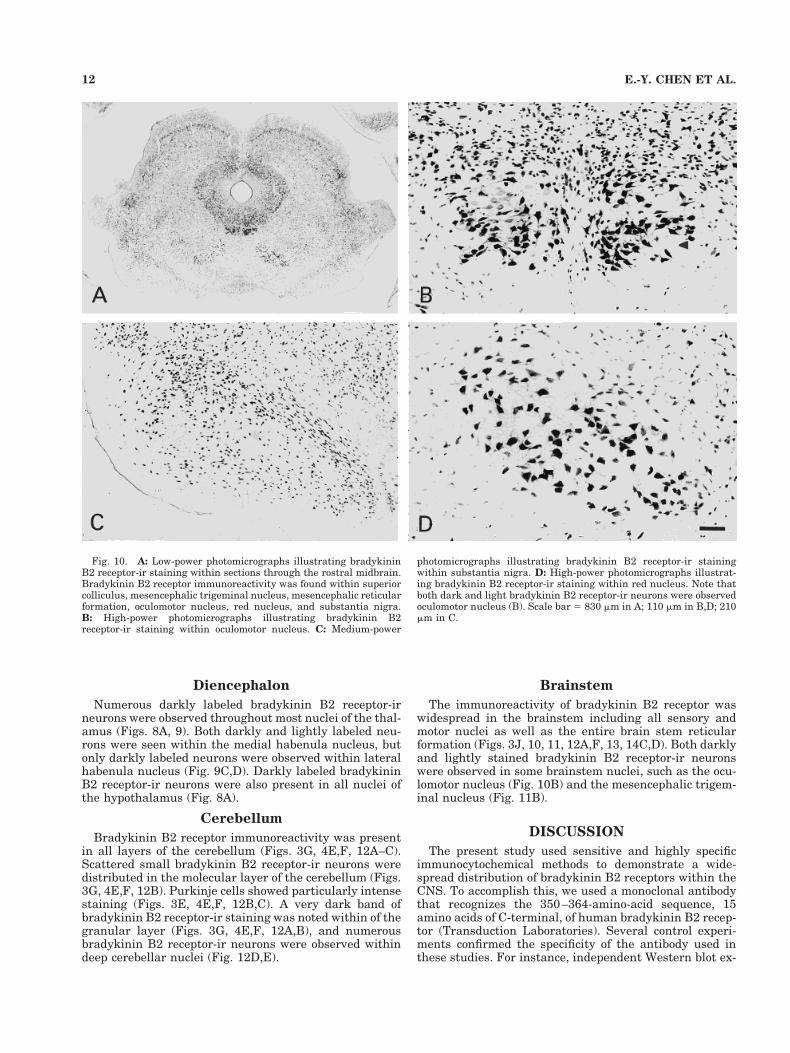
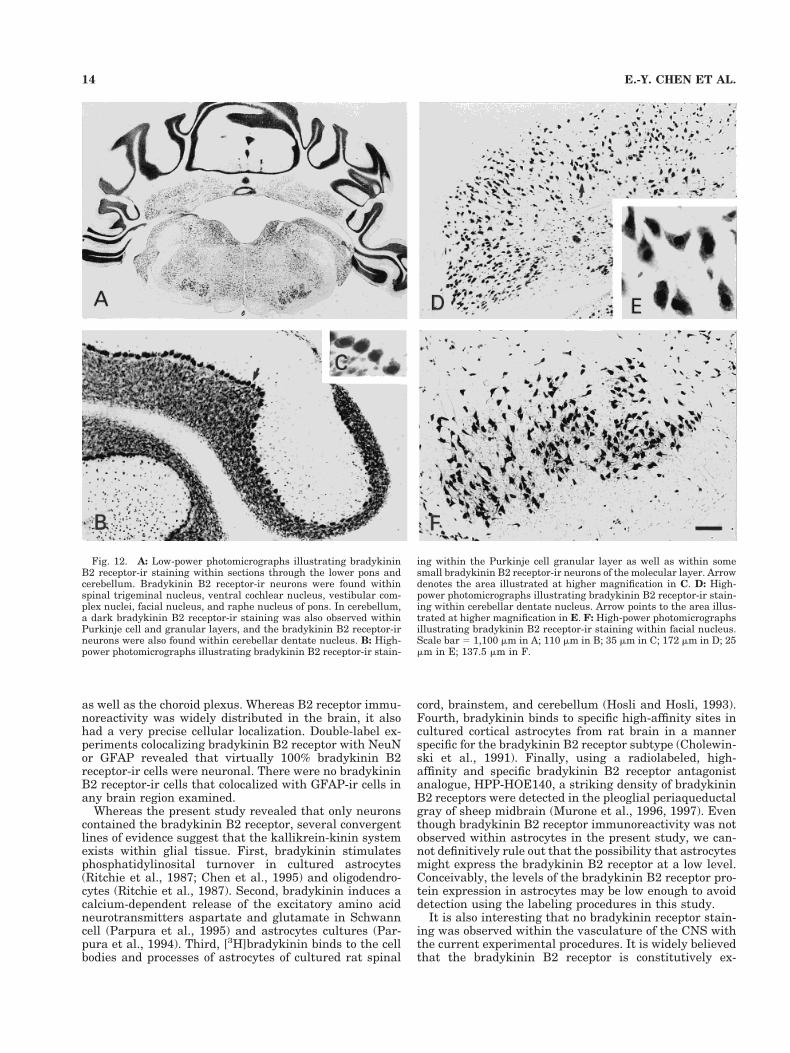
neurons in dentate nucleus of cerebellum (Fig. 12E).Both dark- and light-staining neurons were seen withinsome nuclei such as medial habenula (Fig. 9C,D), hilusof dentate gyrus (Fig. 8C,E), oculomotor nucleus (Fig.10B), and mesencephalic trigeminal nucleus (Fig. 11B).
Double-labeling experiments colocalizing the bradyki-nin B2 receptor and NeuN revealed that virtually 100% ofbradykinin B2 receptor-immunoreactive (ir) cells werealso NeuN immunoreactive (Fig. 3). The exception for thiswas a few neuronal populations that were only labeled bymouse antibradykinin B2 receptor antibody. Such popula-tions included Purkinje cells, most of small neurons lo-cated in the molecular layer of the cerebellum (Fig. 3G–I),mitral cells, and large tufted cells of the olfactory bulb(data not shown). Double-labeling of bradykinin B2 recep-tor and GFAP revealed that virtually no bradykinin B2receptor immunoreactivity was seen in astrocytes in anybrain region examined. Because bradykinin B2 receptorimmunoreactivity was observed only within neurons (Fig.4A–F), the present paper uses the phrase bradykinin B2receptor-ir neurons to denote bradykinin B2 receptor-ircells.
Olfactory bulb
Bradykinin B2 receptor-ir staining was observedwithin all neuronal types of the olfactory bulb (Fig. 5).No staining was observed within the olfactory nervelayer. Numerous small bradykinin B2 receptor-ir neu-rons were seen surrounding glomeruli in the periglo-merular region. An occasional bradykinin B2 receptor-irneuron was found within the glomeruli itself (Fig.5B,C). Within the external plexiform layer, most brady-kinin B2 receptor-ir neurons were medium-sized tolarge tufted cells (Fig. 5B,C). One or two rows of intensebradykinin B2 receptor-ir mitral cells were also ob-served within the mitral cell layer (Fig. 5A,B). Numer-ous bradykinin B2 receptor-ir neuron clusters were ob-served within the internal granular layer (Fig. 5A,B).Together these results indicate that bradykinin B2 re-ceptor immunoreactivity is present in all neuronal celltypes of all layers within rat olfactory bulb.
Cerebral cortex
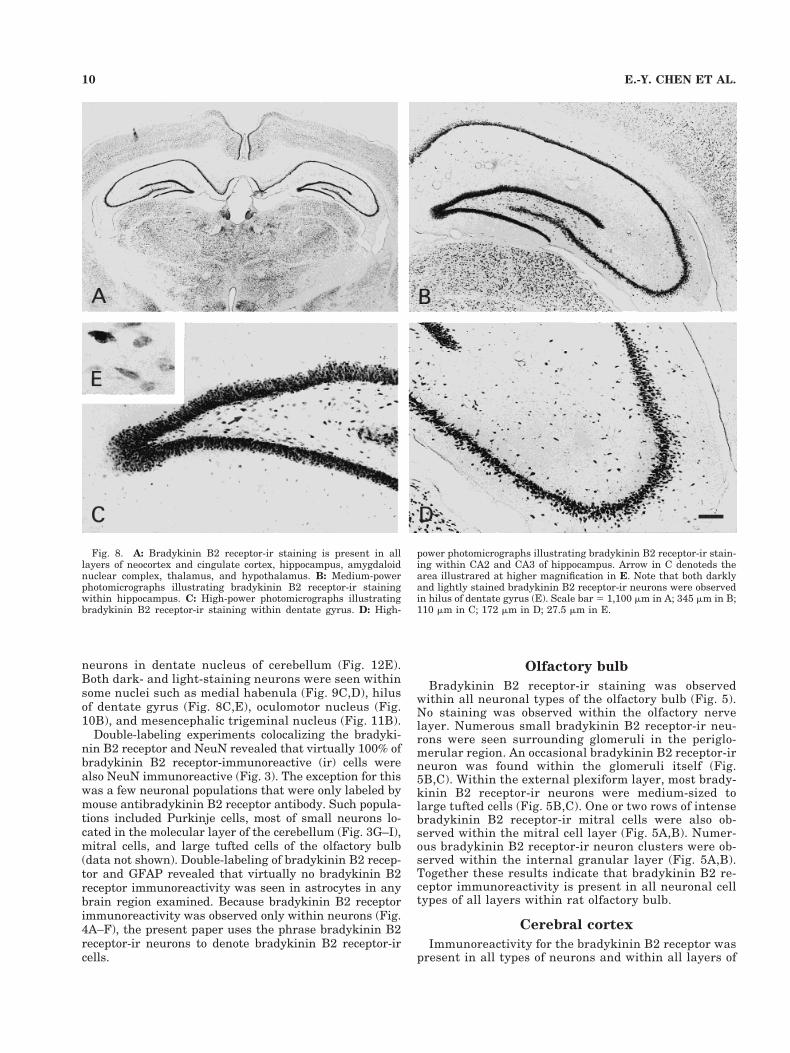
Immunoreactivity for the bradykinin B2 receptor waspresent in all types of neurons and within all layers of
Fig. 8. A: Bradykinin B2 receptor-ir staining is present in alllayers of neocortex and cingulate cortex, hippocampus, amygdaloidnuclear complex, thalamus, and hypothalamus. B: Medium-powerphotomicrographs illustrating bradykinin B2 receptor-ir stainingwithin hippocampus. C: High-power photomicrographs illustratingbradykinin B2 receptor-ir staining within dentate gyrus. D: High-
power photomicrographs illustrating bradykinin B2 receptor-ir stain-ing within CA2 and CA3 of hippocampus. Arrow in C denoteds thearea illustrared at higher magnification in E. Note that both darklyand lightly stained bradykinin B2 receptor-ir neurons were observedin hilus of dentate gyrus (E). Scale bar 5 1,100 mm in A; 345 mm in B;110 mm in C; 172 mm in D; 27.5 mm in E.
10 E.-Y. CHEN ET AL.
the cortex (Figs. 2A,C, 3A, 4A,B, 6A,C, 7A,B, 8A). Theintensity of immunoreactivity of bradykinin B2 receptorvaried over the different cortical layers and across cor-tical regions. In neocortex, dark immunoreactivity wasfound within the pyramidal cell somata of layer V; amoderate immunoreactivity was observed in layersI–III and layer VI; and light immunoreactivity was ob-served in layer IV (Figs. 6A, 7A,B, 8A). In contrast,darkly immunoreactivity was found in all layers of thecingulate, insular, piriform, and entorhinal cortices(Figs. 2A, 6A,C,D, Fig 7A, 8A).
Amygdaloid nuclear complex andhippocampus
Immunoreactivity for the bradykinin B2 receptor wasfound throughout the amygdaloid nuclear complex. Immu-noreactivity was also present in virtually all neurons of alllayers of both the hippocampal formation and the dentategyrus. A collection of darkly stained neurons was observedwithin the pyramidal layer of the hippocampal formationand the granule cell layer of the dentate gyrus (Figs. 2A,3D, 4C,D, 8). Scattered bradykinin B2 receptor-ir neuronswere seen within both polymorphic layer and molecularlayer of the hippocampal proper and molecular layer ofdentate gyrus (Figs. 3D, 4C,D, 8B–D). Both darkly and
lightly stained neurons were observed within hilus of den-tate gyrus (Fig. 8B,C,E).
Basal forebrain
Bradykinin B2 receptor-ir was observed within all thesubfields of the basal forebrain, including the lateral sep-tum, medial septal/diagonal band complex, and nucleusbasalis of Meynert. In the lateral septal nucleus, a groupof large and medium-sized multipolar cells with eithertriangular or round shapes were darkly stained (Fig.6A,B). Bradykinin B2 receptor-ir cells were present inboth magnocellular neurons and nonmagnocellular neu-rons within medial septal/diagonal band complex (Fig.6A,B) and nucleus basalis magnocellularis.
Basal ganglia
Numerous lightly labeled bradykinin B2 receptor-irneurons were scattered throughout the caudate putamen(Figs. 2C, 6A). Darkly labeled bradykinin B2 receptor-irneurons were also observed scattered throughout globuspallidus and entopeduncular nucleus. These bradykininB2 receptor-ir neurons were multipolar in morphology,with only short processes emanating from the soma.Darkly labeled bradykinin B2 receptor-ir neurons wereobserved throughout subthalamic nucleus.
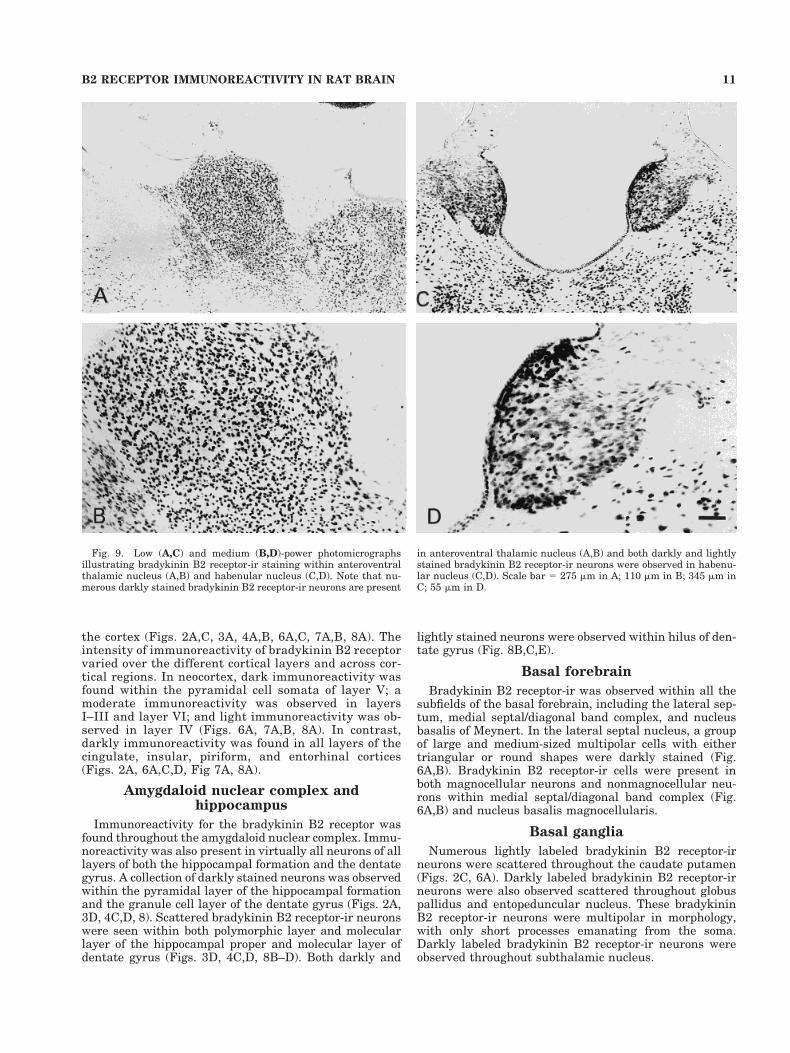
Fig. 9. Low (A,C) and medium (B,D)-power photomicrographsillustrating bradykinin B2 receptor-ir staining within anteroventralthalamic nucleus (A,B) and habenular nucleus (C,D). Note that nu-merous darkly stained bradykinin B2 receptor-ir neurons are present
in anteroventral thalamic nucleus (A,B) and both darkly and lightlystained bradykinin B2 receptor-ir neurons were observed in habenu-lar nucleus (C,D). Scale bar 5 275 mm in A; 110 mm in B; 345 mm inC; 55 mm in D.
11B2 RECEPTOR IMMUNOREACTIVITY IN RAT BRAIN
Diencephalon
Numerous darkly labeled bradykinin B2 receptor-irneurons were observed throughout most nuclei of the thal-amus (Figs. 8A, 9). Both darkly and lightly labeled neu-rons were seen within the medial habenula nucleus, butonly darkly labeled neurons were observed within lateralhabenula nucleus (Fig. 9C,D). Darkly labeled bradykininB2 receptor-ir neurons were also present in all nuclei ofthe hypothalamus (Fig. 8A).
Cerebellum
Bradykinin B2 receptor immunoreactivity was presentin all layers of the cerebellum (Figs. 3G, 4E,F, 12A–C).Scattered small bradykinin B2 receptor-ir neurons weredistributed in the molecular layer of the cerebellum (Figs.3G, 4E,F, 12B). Purkinje cells showed particularly intensestaining (Figs. 3E, 4E,F, 12B,C). A very dark band ofbradykinin B2 receptor-ir staining was noted within of thegranular layer (Figs. 3G, 4E,F, 12A,B), and numerousbradykinin B2 receptor-ir neurons were observed withindeep cerebellar nuclei (Fig. 12D,E).
Brainstem
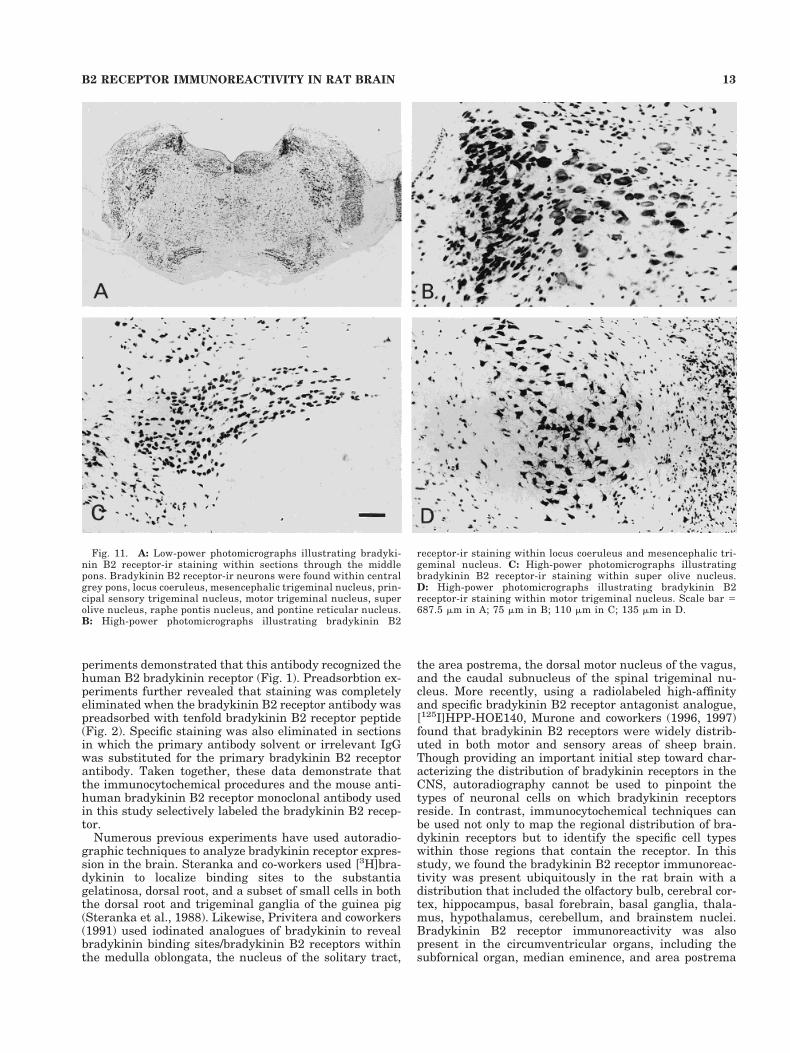
The immunoreactivity of bradykinin B2 receptor waswidespread in the brainstem including all sensory andmotor nuclei as well as the entire brain stem reticularformation (Figs. 3J, 10, 11, 12A,F, 13, 14C,D). Both darklyand lightly stained bradykinin B2 receptor-ir neuronswere observed in some brainstem nuclei, such as the ocu-lomotor nucleus (Fig. 10B) and the mesencephalic trigem-inal nucleus (Fig. 11B).
DISCUSSION
The present study used sensitive and highly specificimmunocytochemical methods to demonstrate a wide-spread distribution of bradykinin B2 receptors within theCNS. To accomplish this, we used a monoclonal antibodythat recognizes the 350–364-amino-acid sequence, 15amino acids of C-terminal, of human bradykinin B2 recep-tor (Transduction Laboratories). Several control experi-ments confirmed the specificity of the antibody used inthese studies. For instance, independent Western blot ex-
Fig. 10. A: Low-power photomicrographs illustrating bradykininB2 receptor-ir staining within sections through the rostral midbrain.Bradykinin B2 receptor immunoreactivity was found within superiorcolliculus, mesencephalic trigeminal nucleus, mesencephalic reticularformation, oculomotor nucleus, red nucleus, and substantia nigra.B: High-power photomicrographs illustrating bradykinin B2receptor-ir staining within oculomotor nucleus. C: Medium-power
photomicrographs illustrating bradykinin B2 receptor-ir stainingwithin substantia nigra. D: High-power photomicrographs illustrat-ing bradykinin B2 receptor-ir staining within red nucleus. Note thatboth dark and light bradykinin B2 receptor-ir neurons were observedoculomotor nucleus (B). Scale bar 5 830 mm in A; 110 mm in B,D; 210mm in C.
12 E.-Y. CHEN ET AL.
periments demonstrated that this antibody recognized thehuman B2 bradykinin receptor (Fig. 1). Preadsorbtion ex-periments further revealed that staining was completelyeliminated when the bradykinin B2 receptor antibody waspreadsorbed with tenfold bradykinin B2 receptor peptide(Fig. 2). Specific staining was also eliminated in sectionsin which the primary antibody solvent or irrelevant IgGwas substituted for the primary bradykinin B2 receptorantibody. Taken together, these data demonstrate thatthe immunocytochemical procedures and the mouse anti-human bradykinin B2 receptor monoclonal antibody usedin this study selectively labeled the bradykinin B2 recep-tor.
Numerous previous experiments have used autoradio-graphic techniques to analyze bradykinin receptor expres-sion in the brain. Steranka and co-workers used [3H]bra-dykinin to localize binding sites to the substantiagelatinosa, dorsal root, and a subset of small cells in boththe dorsal root and trigeminal ganglia of the guinea pig(Steranka et al., 1988). Likewise, Privitera and coworkers(1991) used iodinated analogues of bradykinin to revealbradykinin binding sites/bradykinin B2 receptors withinthe medulla oblongata, the nucleus of the solitary tract,
the area postrema, the dorsal motor nucleus of the vagus,and the caudal subnucleus of the spinal trigeminal nu-cleus. More recently, using a radiolabeled high-affinityand specific bradykinin B2 receptor antagonist analogue,[125I]HPP-HOE140, Murone and coworkers (1996, 1997)found that bradykinin B2 receptors were widely distrib-uted in both motor and sensory areas of sheep brain.Though providing an important initial step toward char-acterizing the distribution of bradykinin receptors in theCNS, autoradiography cannot be used to pinpoint thetypes of neuronal cells on which bradykinin receptorsreside. In contrast, immunocytochemical techniques canbe used not only to map the regional distribution of bra-dykinin receptors but to identify the specific cell typeswithin those regions that contain the receptor. In thisstudy, we found the bradykinin B2 receptor immunoreac-tivity was present ubiquitously in the rat brain with adistribution that included the olfactory bulb, cerebral cor-tex, hippocampus, basal forebrain, basal ganglia, thala-mus, hypothalamus, cerebellum, and brainstem nuclei.Bradykinin B2 receptor immunoreactivity was alsopresent in the circumventricular organs, including thesubfornical organ, median eminence, and area postrema
Fig. 11. A: Low-power photomicrographs illustrating bradyki-nin B2 receptor-ir staining within sections through the middlepons. Bradykinin B2 receptor-ir neurons were found within centralgrey pons, locus coeruleus, mesencephalic trigeminal nucleus, prin-cipal sensory trigeminal nucleus, motor trigeminal nucleus, superolive nucleus, raphe pontis nucleus, and pontine reticular nucleus.B: High-power photomicrographs illustrating bradykinin B2
receptor-ir staining within locus coeruleus and mesencephalic tri-geminal nucleus. C: High-power photomicrographs illustratingbradykinin B2 receptor-ir staining within super olive nucleus.D: High-power photomicrographs illustrating bradykinin B2receptor-ir staining within motor trigeminal nucleus. Scale bar 5687.5 mm in A; 75 mm in B; 110 mm in C; 135 mm in D.
13B2 RECEPTOR IMMUNOREACTIVITY IN RAT BRAIN
as well as the choroid plexus. Whereas B2 receptor immu-noreactivity was widely distributed in the brain, it alsohad a very precise cellular localization. Double-label ex-periments colocalizing bradykinin B2 receptor with NeuNor GFAP revealed that virtually 100% bradykinin B2receptor-ir cells were neuronal. There were no bradykininB2 receptor-ir cells that colocalized with GFAP-ir cells inany brain region examined.
Whereas the present study revealed that only neuronscontained the bradykinin B2 receptor, several convergentlines of evidence suggest that the kallikrein-kinin systemexists within glial tissue. First, bradykinin stimulatesphosphatidylinosital turnover in cultured astrocytes(Ritchie et al., 1987; Chen et al., 1995) and oligodendro-cytes (Ritchie et al., 1987). Second, bradykinin induces acalcium-dependent release of the excitatory amino acidneurotransmitters aspartate and glutamate in Schwanncell (Parpura et al., 1995) and astrocytes cultures (Par-pura et al., 1994). Third, [3H]bradykinin binds to the cellbodies and processes of astrocytes of cultured rat spinal
cord, brainstem, and cerebellum (Hosli and Hosli, 1993).Fourth, bradykinin binds to specific high-affinity sites incultured cortical astrocytes from rat brain in a mannerspecific for the bradykinin B2 receptor subtype (Cholewin-ski et al., 1991). Finally, using a radiolabeled, high-affinity and specific bradykinin B2 receptor antagonistanalogue, HPP-HOE140, a striking density of bradykininB2 receptors were detected in the pleoglial periaqueductalgray of sheep midbrain (Murone et al., 1996, 1997). Eventhough bradykinin B2 receptor immunoreactivity was notobserved within astrocytes in the present study, we can-not definitively rule out that the possibility that astrocytesmight express the bradykinin B2 receptor at a low level.Conceivably, the levels of the bradykinin B2 receptor pro-tein expression in astrocytes may be low enough to avoiddetection using the labeling procedures in this study.
It is also interesting that no bradykinin receptor stain-ing was observed within the vasculature of the CNS withthe current experimental procedures. It is widely believedthat the bradykinin B2 receptor is constitutively ex-
Fig. 12. A: Low-power photomicrographs illustrating bradykininB2 receptor-ir staining within sections through the lower pons andcerebellum. Bradykinin B2 receptor-ir neurons were found withinspinal trigeminal nucleus, ventral cochlear nucleus, vestibular com-plex nuclei, facial nucleus, and raphe nucleus of pons. In cerebellum,a dark bradykinin B2 receptor-ir staining was also observed withinPurkinje cell and granular layers, and the bradykinin B2 receptor-irneurons were also found within cerebellar dentate nucleus. B: High-power photomicrographs illustrating bradykinin B2 receptor-ir stain-
ing within the Purkinje cell granular layer as well as within somesmall bradykinin B2 receptor-ir neurons of the molecular layer. Arrowdenotes the area illustrated at higher magnification in C. D: High-power photomicrographs illustrating bradykinin B2 receptor-ir stain-ing within cerebellar dentate nucleus. Arrow points to the area illus-trated at higher magnification in E. F: High-power photomicrographsillustrating bradykinin B2 receptor-ir staining within facial nucleus.Scale bar 5 1,100 mm in A; 110 mm in B; 35 mm in C; 172 mm in D; 25mm in E; 137.5 mm in F.
14 E.-Y. CHEN ET AL.

pressed on the endothelial cells of the brain’s capillaries,where its activation of capillary receptors induces a potentvasogenic response (Wahl et al., 1983; Unterberg et al.,1984; Yong et al., 1992). Murone and coworkers (1996,1997) have also described a high density of bradykinin B2receptors within the circumventricular organs and themiddle cerebral arteries. Moreover, kallikrein mRNA hasbeen identified in endothelial cells of rat arteries andveins (Saed et al., 1990), and B2 receptor immunoreactiv-ity is present in endothelial cells and smooth muscle ofhuman arteries and veins (Raidoo et al., 1997). Giventhese considerations, it is surprising that no bradykininB2 receptor staining was observed on blood vessels in thisstudy. Again, it is possible that the level of bradykinin B2receptor protein expression in endothelial cells was insuf-ficient for detection using current labeling procedures.However, it is notable that none of the other studies thathave employed antibodies directed against the bradykininB2 receptor have reported staining on CNS blood vessels(e.g., Raidoo et al., 1996b; Raidoo and Raidoo, 1997).
One of the better recognized roles of bradykinin is tomodulate cardiovascular homeostasis via central mecha-nisms. Intraventricular injections of bradykinin produce adose-dependent increase in blood pressure and heart rate
that is mediated by bradykinin B2 receptors and activa-tion of the kallikrein system (Lindsey et al., 1989; Yang etal., 1989; Yong et al., 1992; Madeddu et al., 1994, 1996).Local injections of bradykinin into brain parenchyma haveidentified several regions involved in the cardiovasculareffects of bradykinin. For instance, Diz and Jacobwitz(1984) demonstrated that bradykinin injections into therostral hypothalamus produce increases in blood pressureand heart rate, whereas injections into the medial ante-rior hypothalamus decrease heart rate but have no effecton blood pressure. Lindsey and coworkers (1997) foundthat microinjection of bradykinin directly over the paratri-geminal nucleus of unanaesthetized rats produced a sig-nificant increase in arterial pressure and a moderate in-crease in heart rate. Autoradiographic studies haveidentified bradykinin receptors in several other brain re-gions associated with cardiovascular control, including thenucleus of the solitary tract, dorsal nucleus of the vagus,and area postrema (Murone et al., 1997). Although thesedata alone are not sufficient to determine whether brady-kinin plays a role in normal cardiovascular homeostasis,the present demonstration of B2 receptor immunostainingin multiple brain areas that could modulate vascular toneprovides further circumstantial support for such a role.
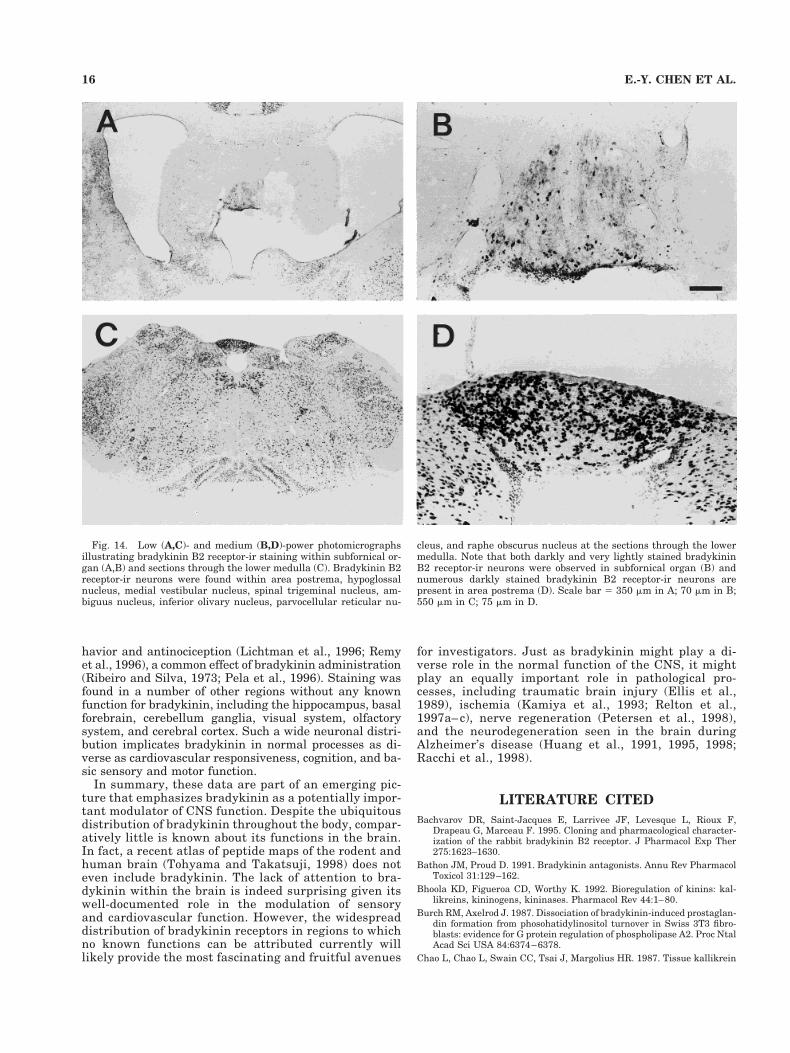
Consistent with previous reports, bradykinin B2-ir neu-rons were found at high density within the circumven-tricular organs, including the subfornical organ, medianeminence, and area postrema. An important feature thatdistinguishes the circumventricular organs from the restof the CNS is the lack of a blood–brain barrier that allowsrelatively free exchange of compounds between the brainand periphery (McKinley et al., 1990). The presence ofbradykinin B2 receptors may indicate that bradykininparticipates in a system that functions as a gate for hu-moral factors to penetrate the brain and exert effects onregions accessible by local subsequent diffusion. Intraven-tricular administration of kinins has been reported toproduce a wide variety of effects, including hypertension,bradycardia, antinociception, and antidiuresis. Each ofthese effects has been suggested to be mediated, at least inpart, by a local bradykinin system within circumventricu-lar organs. For instance, lesions of the median eminence(Hoffman and Schmid, 1978) abolish the antidiuretic ef-fects of bradykinin. Lesions of the area postrema (Wilkin-son and Scroop, 1985) overcome the increased blood pres-sure and heart rate that occurs with intracarotid infusionsof bradykinin. Moreover, the presence of kininogenaseactivity and kallikrein-like immunoreactivity in humancerebrospinal fluid suggests that bradykinin’s pressor ef-fects might be locally regulated within the circumven-tricular organs (Scicli et al., 1984).
Widespread bradykinin B2 receptor staining was ob-served within brainstem nuclei, including the red nucleus,oculomotor nucleus, and mesencephalic trigeminal nu-cleus. Although no role of bradykinin is known in theseregions, it is tempting to speculate on its potential involve-ment in brain function. The observation of staining withinthe red nucleus is intriguing because of the role of thisstructure and its efferent pathways in eliciting sustainedpostural tremor and other extrapyramidal symptoms. Theinvolvement of the red nucleus in locomotor function inthe rat suggests that bradykinin might be involved in theregulation and control of motor function (Ruigrok et al.,1996, Wishaw et al., 1996). B2 staining was also observedin the raphe nucleus, suggesting a role in anxiolytic be-
Fig. 13. A: Low-power photomicrographs illustrating bradykininB2 receptor-ir staining within sections through the lower medulla.Bradykinin B2 receptor-ir neurons were found within hypoglossalnucleus, medial vestibular nucleus, spinal trigeminal nucleus, am-biguus nucleus, inferior olivary nucleus, parvocellular reticular nu-cleus, and raphe obscurus nucleus. B: High-power photomicrographsillustrating bradykinin B2 receptor-ir staining within hypoglossalnucleus. Scale bar 5 687.5 mm in A; 110 mm in B.
15B2 RECEPTOR IMMUNOREACTIVITY IN RAT BRAIN
havior and antinociception (Lichtman et al., 1996; Remyet al., 1996), a common effect of bradykinin administration(Ribeiro and Silva, 1973; Pela et al., 1996). Staining wasfound in a number of other regions without any knownfunction for bradykinin, including the hippocampus, basalforebrain, cerebellum ganglia, visual system, olfactorysystem, and cerebral cortex. Such a wide neuronal distri-bution implicates bradykinin in normal processes as di-verse as cardiovascular responsiveness, cognition, and ba-sic sensory and motor function.
In summary, these data are part of an emerging pic-ture that emphasizes bradykinin as a potentially impor-tant modulator of CNS function. Despite the ubiquitousdistribution of bradykinin throughout the body, compar-atively little is known about its functions in the brain.In fact, a recent atlas of peptide maps of the rodent andhuman brain (Tohyama and Takatsuji, 1998) does noteven include bradykinin. The lack of attention to bra-dykinin within the brain is indeed surprising given itswell-documented role in the modulation of sensoryand cardiovascular function. However, the widespreaddistribution of bradykinin receptors in regions to whichno known functions can be attributed currently willlikely provide the most fascinating and fruitful avenues
for investigators. Just as bradykinin might play a di-verse role in the normal function of the CNS, it mightplay an equally important role in pathological pro-cesses, including traumatic brain injury (Ellis et al.,1989), ischemia (Kamiya et al., 1993; Relton et al.,1997a– c), nerve regeneration (Petersen et al., 1998),and the neurodegeneration seen in the brain duringAlzheimer’s disease (Huang et al., 1991, 1995, 1998;Racchi et al., 1998).
LITERATURE CITED
Bachvarov DR, Saint-Jacques E, Larrivee JF, Levesque L, Rioux F,Drapeau G, Marceau F. 1995. Cloning and pharmacological character-ization of the rabbit bradykinin B2 receptor. J Pharmacol Exp Ther275:1623–1630.
Bathon JM, Proud D. 1991. Bradykinin antagonists. Annu Rev PharmacolToxicol 31:129–162.
Bhoola KD, Figueroa CD, Worthy K. 1992. Bioregulation of kinins: kal-likreins, kininogens, kininases. Pharmacol Rev 44:1–80.
Burch RM, Axelrod J. 1987. Dissociation of bradykinin-induced prostaglan-din formation from phosohatidylinositol turnover in Swiss 3T3 fibro-blasts: evidence for G protein regulation of phospholipase A2. Proc NtalAcad Sci USA 84:6374–6378.
Chao L, Chao L, Swain CC, Tsai J, Margolius HR. 1987. Tissue kallikrein
Fig. 14. Low (A,C)- and medium (B,D)-power photomicrographsillustrating bradykinin B2 receptor-ir staining within subfornical or-gan (A,B) and sections through the lower medulla (C). Bradykinin B2receptor-ir neurons were found within area postrema, hypoglossalnucleus, medial vestibular nucleus, spinal trigeminal nucleus, am-biguus nucleus, inferior olivary nucleus, parvocellular reticular nu-
cleus, and raphe obscurus nucleus at the sections through the lowermedulla. Note that both darkly and very lightly stained bradykininB2 receptor-ir neurons were observed in subfornical organ (B) andnumerous darkly stained bradykinin B2 receptor-ir neurons arepresent in area postrema (D). Scale bar 5 350 mm in A; 70 mm in B;550 mm in C; 75 mm in D.
16 E.-Y. CHEN ET AL.

in rat brain pituitary: regional distribution and estrogen induction inthe anterior pituitary. Endocrinology 120:475–482.
Chen C-C, Chang J, Chen W-C. 1995. Role of protein kinase C subtypes aand d in the regulation of bradykinin-stimulated phosphoinositidebreakdown in astrocytes. Mol Pharmacol 39:39–47.
Cholewinski AJ, Stevens G, McDermott AM, Wilkin GP. 1991. Identifica-tion of B2 bradykinin binding sites on cultured cortical astrocytes.J Neurochem 57:1456–1458.
Correa FMA, Innis RB, Uhl GR, Snyder SH. 1979. Bradykinin-like immu-noreactive neuronal system localized histochemically in rat brain. ProcNatl Acad Sci USA 76:1489–1493.
Diz DI, Jacobowitz DM. 1984. Cardiovascular effects of discrete intrahy-pothalamic and preoptic injections of bradykinin. Brain Res Bull 12:409–417.
Dray A, Perkins M. 1993. Bradykinin and inflammatory pain. TrendsNeurosci 16:99–104.
Ellis EF, Chao J, Heizer ML. 1989. Brain kininogen following experimentalbrain injury: evidence for a secondary event. J Neurosurg 71:437–442.
Franzusoff A, Redding K, Crosby J, Fuller RS, Schekman R. 1991. Local-ization of components involved in protein transport and processingthrough the yeast Golgi apparatus. J Cell Biol 112:27–37.
Fujiwara Y, Mantione CR, Yamamura HI. 1988. Identification of B2 bra-dykinin binding sites in guinea-pig brain. Eur J Pharmacol 147:487–488.
Fujiwara Y, Mantione CR, Vavrek RJ, Stewart JM, Yamamura HI. 1989.Characterization of [3H]bradykinin binding sites in guinea-pig centralnervous system: possible existence of B2 subtypes. Life Sci 44:1645–1653.
Hall JM. 1992. Bradykinin receptors: pharmacological properties and bio-logical roles. Pharmacol Ther 56:131–190.
Hess JF, Borkowski JA, Young GS, Strader CD, Ransom RW. 1992. Clon-ing and pharmacological characterization of a human bradykinin(BK-2) receptor. Biochem Biophys Res Commun 184:260–268.
Hess JF, Borkowski JA, Macneil T, Stonesifer GY, Fraher J, Strader CD,Ransom RW. 1994a. Differential pharmacology of cloned human andmouse B2 bradykinin receptors. Mol Pharmacol 45:1–8.
Hess JF, Borkowski JA, Stonesifer GY, MacNeil T, Strader CD, RansomRW. 1994b. Cloning and pharmacological characterization of bradyki-nin receptors. Brazil J Med Biol Res 27:1725–1723.
Hoffman WE, Schmid PG. 1978. Separation of pressor and antidiureticeffects of intraventricular bradykinin. Neuropharmacology 17:999–1002.
Hosli E, Hosli L. 1993. Autoradiographic localization of binding sites forneuropeptide Y and bradykinin on astrocytes. Neuroreport 4:159–162.
Hsu SM, Soban E. 1982. Color modification of diaminobenzidine (DAB)precipitation by metallic ions and its application for double immuno-histochemistry. J Histochem Cytochem 30:1079–1082.
Hsu SM, Raine L, Fanger H. 1981. Use of avidin-biotin peroxidase complex(ABC) in immunoperoxidase techniques. A comparison between ABCand unlabeled antibody (PAP) procedures. J Histochem Cytochem 29:577–580.
Huang H-M, Toral-Barza L, Thaler H, Tofel-Grehl B, Gibson GE. 1991.Inositol phosphates and inteacellular calcium after bradyinin stimula-tion in fibroblasts from young, normal aged and Alzheimer donors.Neurobiol Aging 12:469–473.
Huang H-M, Lin T-A, Sun GY, Gibson GE. 1995. Increased inositol 1,4,5-trisphoisphate accumulation correlates with an up-regulation of bra-dykinin receptors in Alzheimer’s disease. J Neurochem 64:761–766.
Huang H-M, Ou H-C, Hsueh S-J. 1998. Amyloid b peptide enhancedbradykinin-mediated inositol (1,4,5)trisphosphate formation and cyto-solic free calcium. Life Sci 63:195–203.
Kamiya T, Katayama Y, Kashiwagi F, Terashi A. 1993. The role of brady-kinin in mediating ischemic brain edema in rats. Stroke 24:571–576.
Kaplan AP, Joseph K, Shibayama Y, Nakazawa Y, Ghebrehiwet B, Reddi-gari S, Silverberg M. 1998. Bradykinin formation. Plasma and tissuepathways and cellular interactions. Clin Rev Allerg Immunol 16:403–429.
Kariya K, Kawauchi R, Okamoto H. 1981. Regional distribution of kininasein rat brain. J Neurochem 36:2086–2088.
Kariya K, Yamaucki A, Sasaki T. 1985. Regional distribution and charac-terization of kinin in the CNS of the rat. J Neurochem 44:1892–1897.
Kizuki K, Suzuki T, Kudo M, Noguchi T. 1994. Immunohistochemicaldemonstration of tissue kallikrein in the neurons of rat brain. BrainRes 634:305–309.
Kordower JH, Sladek JR, Bing G, Fiandaca MS, Gash DM. 1988. Tyrosinehudroxylase immunoreactive neurons in the primate subfornical or-gan: species specificity. Brain Res 461:221–229.
Lewis Carl SA, Gillete-Ferguson I, Ferguson DG. 1993. An indirect immu-nofluorescence procedure for staining the same cryosection with twomouse monoclonal primary antibodies. J Histochem Cytochem 41:1273–1278.
Lichtman AH, Cook SA, Martin BR. 1996. Investigation of brain sitesmediating cannabinoid-induced antinociception in rats: evidence sup-porting periaqueductal gray involvement. J Pharmacol Exper Ther276:585–593.
Lindsey CJ, Nakaie CR, Martins DTO. 1989. Central nervous system kininreceptors and the hypertensive response mediated by bradykinin. Br JPharmacol 97:763–768.
Lindsey CJ, Buck HS, Fior-Chadi DR, Lapa RC. 1997. Pressor effectmediated by bradykinin in the paratrigeminal nucleus of the rat.J Physiol 502:119–129.
Lopes P, Kar S, Chretien L, Regoll D, Quirion R, Couture R. 1995. Quan-titative autoradiographic localization of [125I-Tyr]bradykinin receptorbinding sites in the rat spinal cord: effects of neuronal capsaicin,noradrenergic deafferentation, dorsal rhizotomy and peripheral axo-tomy. Neuroscience 68:867–881.
Madeddu P, Glorioso N, Varoni MV, Demontis MP, Fattaccio MC, AnaniaV. 1994. Cardiovascular effects ofbrain kinin receptor blockade in spon-taneously hypertensive rats. Hypertension 23:189–192.
Madeddu P, Parpaglia PP, Glorioso N, Chao L, Chao J. 1996. Antisenseinhibition of the brain kallikrein-kinin system. Hypertension 28:980–987.
Marceau F. 1995. Kinin B1 receptors: a review. Immunopharmacology30:1–26.
Marceau F, Barabe J, St.-Pierre S, Regoli D. 1980. Kinin receptors inexperimental inflammation. Can J Physiol Pharmacol 58:536–542.
Marceau F, Lussier A, Regoli D, Giroud JP. 1983. Pharmacology of kinins:their relevance to tissue injury and inflammation. Gen Pharmacol14:209–29.
Marceau F, Larrivee JF, Saint-Jacques E, Bachvarov DR. 1997. The kininB1 receptor: an inducible G protein coupled receptor. Can J PhysiolPharmacol 75:725–730.
McEachern AE, Shelton ER, Bhakta S, Obernolte R, Bach C, Zuppan P,Fujisaki J, Aldrich RW, Jarnagin K. 1991. Expression cloning of a ratbradykinin receptor. Proc Natl Acad Sci USA 88:7724–7728.
McKinley MJ, McAllen RM, Mendelsohn FAO, Allen AM, Chai SY, OldfieldBJ. 1990. Circumventricular organs: neuroendocrine interfaces be-tween the brain and the hemal milieu. Front Neuroendocrinol 11:91–127.
Menke JG, Borkowski JA, Bierilo KK, MacNeil T, Derrick AW, SchneckKA, Ransom RW, Strader CD, Linemeyer DL, Hess JF. 1994. Expres-sion cloning of a human B1 bradykinin receptor. J Biol Chem 270:21583–21586.
Mufson EJ, Bothwell M, Hersh LB, Kordower JH. 1989. Nerve growthfactor receptor immunoreactive profiles in the normal aged humanbasal forebrain: colocalization with cholinergic neurons. J Comp Neurol285:196–217.
Mufson EJ, Cochran E, Benzing WC, Kordower JH. 1993. Galaninergicinnervation of the cholinergic vertical limb of the diagonal band (Ch2)and bed nucleus of the stria terminalis in aging, Alzheimer’s diseaseand Down’s syndrome. Dementia 4: 237–250.
Mufson EJ, Jaffar S, Levey AI. 1998. m2 Muscarinic acetylcholine receptorimmunoreactive neurons are not reduced within the nucleus basalis inAlzheimer’s disease: Relationship with cholinergic and galaninergicperikarya. J Comp Neurol 392:313–329.
Murone C, Perich RB, Schlawe I, Chai SY, Casley D, MacGregor DP,Muller-Esterl W, Mendelsohn FAO. 1996. Characterization and local-ization of bradykinin B2 receptors in the guinea-pig using a radioiodi-nated HOE140 analogue. Eur J Pharmacol 306:237–247.
Murone C, Paxinos G, McKinley MJ, Oldfield BJ, Muller-Esterl W, Men-delsohn FAO, Chai SY. 1997. Distribution of Bradykinin B2 receptorsin sheep brain and spinal cord visualized by in vitro autoradiography.J Comp Neurol 381:203–218.
Negoescu A, Labat-Moleur F, Lorimier P, Lamarcq L, Guillermet C. 1994.F(ab) secondary antibodies: a general method for double immunolabel-ing with primary antisera from the same species. Efficiency control bychemiluminescence. J Histochem Cytochem 42:433–437.
Parpura V, Basarsky TA, Liu F, Jeftinija K, Jeftinija S, Haydon PG. 1994.
17B2 RECEPTOR IMMUNOREACTIVITY IN RAT BRAIN

Glutamate-mediated astrocyte-neuron signalling. Nature 369:744–747.
Parpura V, Liu F, Jeftinija KV, Jeftinija S, Haydon PG, Jeftinija SD. 1995.Neuroligand-evoked calcium-dependent release of excitatory amino ac-ids from Schwann cells. J Neurosci 15:5831–5839.
Pela IR, Rosa AL, Silva CAA, Huidobro-Toro JP. 1996. Central B2 receptorinvolvement in the antinociceptive effect of bradykinin in rats. Br JPharmacol 118:1488–1492.
Perry DC, Snyder SH. 1984. Identification of bradykinin in mammalianbrain. J Neurochem 43:1072–1080.
Petersen M, Eckert AS, Vonbanchet GS, Heppelmann B, Klusch A, KniffkiK-D. 1998b. Plasticity in the expression of bradykinin binding sites insensory neurons after mechanical nerve injury. Neuroscience 83:949–959.
Powell SJ, Slynn G, Thomas C, Hopkins B, Briggs I, Graham A. 1993.Human bradykinin B2 receptor: nucleotide sequence analysis and as-signment to chromosome 14. Genomics 15:435–438.
Privitera PJ, Daum PR, Hill DR, Hiley CR. 1991. Autoradiographic visu-alization and characteristics of [125I]bradykinin binding sites in guineapig brain. Brain Res 577:73–79.
Racchi M, Ianna P, Binetti G, Trabucchi M, Govoni S. 1998. Bradykinin-induced amyloid precursor protein secretion: a protein kinaseC-independent mechanism that is not altered un fibroblasts from pa-tients with sporadic Alzheimer’s disease. Biochem J 330:1271–1275.
Raidoo DM, Bhoola KD. 1997. Kinin receptors on human neurons. J Neu-roimmunol 77:39–44.
Raidoo DM, Ramchurren N, Naidoo Y, Naidoo S, Bhoola KD. 1996a. Re-gional distribution of tissue kallikrein in the human brain. Immuno-pharmacology 32:39–47.
Raidoo DM, Ramchurren N, Naidoo Y, Naidoo S, Muller-Esterl W, BhoolaKD. 1996b. Visualisation of bradykinin B2 receptors on human brainneurons. Immunopharmacology 33:104–107.
Raidoo DM, Ramsaroop R, Naidoo S, Muller-Esterl W, Bhoola KD. 1997.Kinin receptors in human vascular tissue: their role in atheromatousdisease. Immunopharmacology 36:153–160.
Regoli D, Barabe J. 1980. Pharmacology of bradykinin and related kinins.Pharmacol Rev 32:1–46.
Relton JK, Beckey VE, Rothwell NJ, Whalley ET. 1997a. Bradykinin B2receptor antagonist CP-0597 reduces infarct volume after focal cerebralischamia in the rat: comparison with HOE 140, NPC 17731 and MK801[abstract]. Br J Pharmacol 120:75P.
Relton JK, Rothwell NJ, Whalley ET. 1997b. Blockade bradykinin B1receptor reverses the neuroprotective effect of B2 receptor antagonismafter focal cerebral ischamia in the rat [abstract]. Br J Pharmacol120:76P.
Relton JK, Beckey VE, Hanson WL, Whalley TE. 1997c. CP-0597, a selectivebradykinin B2 receptor antagonist, inhibits brain injury in a rat model ofreversible middle cerebral artery occlusion. Stroke 28:1430–1436.
Remy SM, Schreiber R, Dalmus M, De-Vry J. 1996. Somatodendritic5-HT1A receptors are critically involved in the anxiolytic effects of8-OH-DPAT. Psychopharmacology 125:89–91.
Ribeiro SA, Silva M. 1973. Antinociceptive action of bradylinin and relatedkinins of large molecular weights by the intraventricular route. Br JPharmacol 47:517–528.
Ritchie T, Cole R, Kim H-S, De Vellis J, Noble EP. 1987. Inositol pkosphol-opid hydrolysis in cultured astrocytes and oligodendrocytes. Life Sci41:31–39.
Ruigrok TJ, Vanderburg H, Sabel-Goedknegt E. 1996. Locomotion coin-cides with c-Fos expression in related areas of inferior olive and cere-bellar nuclei in the rat. Neurosci Lett 214:119–122.
Saed GM, Carretero OA, MacDonald RJ, Scicli AG. 1990. Kallikrein mes-senger RNA in rat arteries and veins. Circ Res 67:510–516.
Scicli AG, Forbes G, Nolly HL, Dujovny M, Carretero OA. 1984. Kallifrein-kinins in the central nervous system. Clin Exp Theor Pract A6:1731–1738.
Snyder SH. 1980. Brain peptides as neurotransmitters. Science 209:976–983.
Steranka LR, Manning DC, DeHaas CJ, Ferkany JW, Borosky SA, ConnorJR, Vavrek RJ, Stewart JM. 1988. Bradykinin as a pain mediator:receptors are localized to sensory neurons, and antagonists have anal-gesic actions. Proc Natl Acad Sci USA 85:3245–3249.
Tohyama M, Takatsuji K. 1998. Atlas of neuroactive substances and theirreceptors in the rat. New York: Oxford University Press. p 1–337.
Unterberg A, Wahl M, Baethmann A. 1984. Effects of bradykinin onpermeability and diameter of pial vessels in vivo. J Cereb Blood FlowMetab 4:574–585.
Wahl M, Young AR, Edvinsson L, Wagner F. 1983. Effects of bradykinin onpial arteries and arterioles in vitro and in situ. J Cereb Blood FlowMetab 3:231–237.
Walker K, Perkins M, Dray A. 1995. Kinins and kinin receptors in thenervous sytem. Neurochem Int 26:1–16.
Wessel GM, McClay DR. 1986. Two embryonic, tissue-specific moleculesidentified by a double-label immunofluorescence technique for mono-clonal antibodies. J Histochem Cytochem 34:703–706.
Wilkinson DL, Scroop GC. 1985. Role of prostaglandins and the areaspostrema in the central pressor action of bradykinin. Eur J Pharmacol113:287–290.
Wishaw IQ, Gorny B. 1996. Does the red nucleus provide the tonic supportagainst which fractionated movements occur? A study on forepawmovements used in skilled reaching by the rat. Behav Brain Res74:79–90.
Yaksh TL, Hammond DL. 1982. Peripheral and central substrates in-volved in the rostrad transmission of nociceptive information. Pain13:1– 85.
Yang XP, Carretero OA, Akahoshi M, Scicli AG. 1989. Effects of bloodpressure of intracerebroventricular administration of a kinin antago-nist. Adv Exp Med Biol 247:439–445.
Yano K, Higashida H, Inoue R, Nozawa Y. 1984. Bradykinin-induced rapidbreakdown of phosphatidylinositol 4,5-bisphosphate in neuroblastomaX glioma hybrid NG108-15 cells. J Biol Chem 259:10201–10207.
Yau WM, Dorsett JA, Youther ML. 1986. Bradykinin releases acetylcholinefrom myenteric plexus by a prostaglandin-mediated mechanism. Pep-tides 7:289–292.
Yong T, Gao XP, Koizumi S, Conlon JM, Rennard SI, Mayhan WG, Rubin-stein I. 1992. Role of peptidases in bradykinin-induced increase invascular permeability in vivo. Circ Res 70:952–959.
18 E.-Y. CHEN ET AL.