human bradykinin b2 receptors isolated by receptor-specific
TRANSCRIPT

Proc. Natl. Acad. Sci. USAVol. 90, pp. 10994-10998, December 1993Biochemistry
Human bradykinin B2 receptors isolated by receptor-specificmonoclonal antibodies are tyrosine phosphorylatedY.-J. I. JONG, L. R. DALEMAR, B. WILHELM, AND N. L. BAENZIGER*Department of Anatomy and Neurobiology, Washington University School of Medicine, St. Louis, MO 63110
Communicated by P. Roy Vagelos, July 23, 1993
ABSTRACT We report the immunoaffinity isolation ofbradykinin B2 receptors in a tyrosine-phosphorylated statefrom WI-38 human lung fibroblasts. We generated six mono-clonal antibodies directed against B2 bradykinin receptor bi-ologic activity mediating prostaglandin E2 production in WI-38. These cells express a repertoire of bradykinin receptoraffinity forms with closely correlated biologic activity andt3H]bradykinin binding. Some of the monoclonal antibodiesselectively recognize intermediate-affinity (Kd = 5.6 nM) orlow-affinity (Kd = 42 nM) receptor forms, whereas othersrecognize epitopes common to both. The monoclonal antibodiesblock bradykinin binding and biologic activity. Immunoaffm-ity chromatography on an immobilized monoclonal antibody ofintermediate- plus low-affinity specificity yields WI-38 B2receptors with intact [3H]bradykinin binding activity and amolecular mass of78 kDa. The same band is immunoblotted byall the monoclonal antibodies, indicating a similar molecularmass for receptor forms of different affinity. Anti-phosphoty-rosine antibodies demonstrate that the receptors are tyrosinephosphorylated, with implications for receptor function andregulation. Genistein completely inhibits bradykinin-mediatedprostaglandin E2 production with an IC50 of 8 FM, indicatingthat tyrosine kinase activity is critical for the signal transduc-tion leading to arachidonic acid release.
Bradykinin (BK), Arg-Pro-Pro-Gly-Phe-Ser-Pro-Phe-Arg, isa multifunctional mediator that aids in maintaining tissuehomeostasis and plays a key role in events accompanyinginflammation. BK is proteolytically generated from kinino-gen precursors in circulation and interstitial tissue fluids; itslocal concentration is rapidly increased upon tissue injury.BK receptors belonging to the guanine nucleotide-bindingprotein (G-protein)-coupled family trigger neuronal transmis-sion of pain or irritant messages, enhancement of vascularpermeability, and altered vasomotor tone that are prominentfeatures of the inflammatory process. BK action on thesesame receptors also regulates mitogenesis and protein pro-duction in fibroblasts, initiating repair steps that restorenormal tissue form and function following injury (1-4).We (1) previously demonstrated that IMR-90 human lung
fibroblasts sequentially mobilize BK B2 subtype receptors ofKd = 2.5-5 nM and 44 nM that mediate prostaglandin E2(PGE2) production, upon progression through their finite lifespan in vitro. Further information is needed at the cellular andmolecular level to elucidate the nature of BK B2 receptorsand the means by which cells can express diverse receptorforms. Although recent cloning of two BK receptors hasdisclosed their primary sequences (5-7), the molecular struc-ture of the protein as expressed in intact cells remainsuncharacterized. We have generated monoclonal antibodies(mAbs) that distinguish the BK receptors of Kd = 5 nM and44 nM in human lung fibroblasts and have used them to isolatethe receptor protein and to demonstrate that it contains
The publication costs of this article were defrayed in part by page chargepayment. This article must therefore be hereby marked "advertisement"in accordance with 18 U.S.C. §1734 solely to indicate this fact.
phosphorylated tyrosine, which may serve a regulatory func-tion. We also show that tyrosine kinase activity is essentialfor BK-mediated PGE2 production. These mAbs will allow usto explore the role of BK receptors in the growth anddevelopment offibroblasts and in these cells' participation inonset and/or propagation of the injury/inflammation re-sponse.
MATERIALS AND METHODSReceptor Biologic Activity and [3H]BK Binding. Culture of
WI-38 fibroblasts (ATCC CCL 75), ELISA for BK-mediatedPGE2 production, and equilibrium binding of 25 pM-250 nM[3H]BK (DuPont; 70-107 Ci/mmol; 1 Ci = 37 GBq) to intactcells for 60 min at 4°C and B2 subtype assignment were asdescribed (1). Cells were permeabilized with streptolysin 0(8, 9), and equilibrium binding of 0.075-175 nM [3H]BK wasmeasured in the absence or presence of 100 ,uM guanosine5'-[y-thio]triphosphate (GTP[y-S]) for 15 min at 37°C.[3H]BK bound at input BK concentrations of 0.75-75 nM at4°C was covalently crosslinked to receptors with 0.4 mMdisuccinyl suberimidate (Pierce) at 4°C for 20 min. Afterquenching with 150mM Tris (pH 8), cells were solubilized 2.5min in 400 ,ul of 1% Triton X-100 or 12 mM 3-[(3-cholami-dopropyl)dimethylammonio-1-propane sulfonate (CHAPS)/0.5% digitonin (CD) in 0.15 M NaCl/15 mM Tris Cl, pH 7.4,containing protease inhibitors (100 ,uM phenylmethanesulfo-nyl fluoride, 16.8 uM leupeptin, chymostatin at 4 ,ug/ml, 5.8,tM pepstatin, 6.6 ;LM antipain, 0.08 trypsin inhibitor unit ofaprotinin per ml, 10 mM benzamidine, soybean trypsininhibitor at 0.25 mg/ml, and 5 mM e-amino caproic acid) per35-mm well. Crosslinked radiolabel was analyzed on 5-12%acrylamide gradient gels omitting the SDS (10), run 7 hr at 35mA, and counted as 2-mm gel lane slices in 3a70 scintillant.For membrane preparation, confluent 75-cm2 flasks (8-12
x 106 cells) at 4°C were washed three times in Dulbecco'sphosphate-buffered saline (PBS), washed once in 1 mM Tris(pH 7.5), and swelled 10 min in 3 ml of 1 mM Tris (pH 7.5).Buffer H (10 mM Hepes, pH 7.2/10 mM NaCl/2 mMEGTA/2 mM MgCl2 plus protease inhibitors) was added (9ml); cells were immediately scraped and lysed with 20 strokeswith a B Dounce homogenizer. The homogenate was centri-fuged 5 min at 50 x g and the resulting supernatant wascentrifuged 30 min at 47,000 x g; membranes were resus-pended in buffer H. Membrane aliquots (150-200 pg ofprotein per 50 ,ul) were added to 0.125-90 nM [3H]BK in 100
Abbreviations: BK, bradykinin; mAb, monoclonal antibody; CD, 12mM CHAPS/0.5% digitonin; G protein guanine nucleotide-bindingprotein; GTP[y.S], guanosine 5'-[Lythio]triphosphate; PGE2, pros-taglandin E2; HAT, hypoxanthine/aminopterin/thymidine; CHAPS,3-[(3-cholamidopropyl)dimethylammonio-1-propanesulfonate; H,high; I, intermediate; L, low.*To whom reprint requests should be addressed at: Department ofAnatomy and Neurobiology, Washington University School ofMedicine, Box 8108, 660 South Euclid Avenue, St. Louis, MO63110.
10994

Proc. Natl. Acad. Sci. USA 90 (1993) 10995
,ul of binding buffer (1) and incubated in the absence or
presence of 100 ,uM GTP[y-S] (Boehringer Mannheim) for 15min at 37°C. Reactions were stopped by dilution with 3 ml ofice-cold 50 mM Tris (pH 7.1), filtered through glass fiberfilters (Schleicher & Schuell no. 31) pretreated with 0.3%aqueous polyethylenimine, and washed with 3 ml of the samebuffer. Nonspecific binding in the presence of 30 ,M unla-beled BK was .25% of total binding.
Isolation of mAbs Blocking BK Receptor Biologic Activity.Seven 6- to 8-week-old female BALB/c mice (Harlan-Sprague-Dawley) were immunized subcutaneously at 0, 2, 5,and 8 weeks with 100 ,g ofWI-38 membranes. A seropositivemouse rested 6 weeks was boosted i.p. and fused 4 days laterwith P3X63 Ag8.653 myeloma cells (Washington UniversityHybridoma Center). Selection by standard hypoxanthine/aminopterin/thymidine (HAT) medium techniques yielded200 clones at 13 days; they were screened for mAb-specificinhibition of B2 receptor-mediated PGE2 production inHSDM1C1 cells (4) at 25 or 200 nM BK. Ascites preparationsor purified mAbs were tested, with anti-fibronectin ascitesmAb from HFN 7.1 hybridoma (American Type CultureCollection) as control.
Immunoaffinit Isolation ofBK Receptors. Affinity-purifiedmAb C3-8 (intermediate- plus low-affinity specificity, 0.7 mgof protein per 2 ml) was coupled to 0.6 ml of CNBr-activatedSepharose 4B. WI-38 membranes (4 mg of protein from 20 x106 cells; two 75-cm2 flasks) were incubated for 10 min at 4°Cwith buffer H containing CD, and unsolubilized residue was
sedimented at 50,000 x g for 30 min. Solubilized material wasapplied to the mAb C3-8 affinity column equilibrated withbuffer H containing 0.5% Triton X-100 or CD, and theunadsorbed fraction was cycled back through the column fivetimes. The column was washed with the same buffer, thenwashed successively with 0.05 M Tris (pH 8.2) containingprotease inhibitors, CD or Triton X-100, and 0.1, 0.25, and0.5 M NaCl (5 ml each). Eluates were exchanged into bufferH containing 0.5% Triton X-100 or CD by repeated dilutionand microconcentration.
Receptor Binding and Immunoblotting. CD extracts ofmembranes or column eluates were incubated with 15 nM[3H]BK in the absence or presence of 30 ,uM unlabeled BKfor 1 hr at 4°C in 200 Al of SR buffer (0.2% bovine serumalbumin/2 mM bacitracin/20 ,uM phosphoramidon/20 AMSQ20881/CD in buffer H). Gel filtration on Sephadex G-50(bed volume of 1.8 ml in SR buffer) resolved bound ligand,excluded from the G-50, from free [3H]BK. Binding incuba-tions were washed onto the G-50 column with 200 ,ul of SRbuffer, and two-drop (=100 ,ul) fractions were collected.Membranes or mAb C3-8 column eluates were solubilized 30min at 60°C in sample buffer (10) containing 1% SDS/8 Murea without 2-mercaptoethanol and electrophoresed on an
0.1% SDS/8.5% polyacrylamide/8 M urea gel, followed byWestern blotting (11) with anti-BK receptor mAbs or anti-phosphotyrosine mAb Py2O (ICN) with chemiluminescentdetection.
RESULTS
Expression of a BK B2 Receptor Repertoire in WI-38 Fibro-blasts. We previously demonstrated that early passageIMR-90 fibroblasts expressed a single BK B2 receptor site ofKd = 2.5 nM that mediated PGE2 production; later an
additional B2 site of Kd = 44 nM was concurrently expressed(1). We detected three sites in WI-38 fibroblasts, two corre-sponding to those of IMR-90 and one with higher affinity. Ineach case the respective Kd for [3H]BK binding closelycorrelated with the ED50 (half-maximal stimulatory concen-
tration) for BK-mediated PGE2 production. We termed thesesites high-affinity (H; Kd = 440 pM, ED50 = 790 pM),
intermediate-affinity (I; Kd = 5.6 nM, ED50 7.9 nM), andlow-affinity (L; Kd = 42 nM, ED50 75 nM) forms.These affinity forms could be expressed individually or in
combination as the cells progressed through their life spanfrom passage 13 to passage 28 (n = 100 for PGE2 production;n = 38 for [3H]BK binding). Matched sets of cells from thesame plating yielded matching profiles ofbiologic activity andradioligand binding (n = 10). The most frequently observedreceptor profiles were predominant L site (40%) and concur-rent I and L sites (21%); I site or H site alone or othercombinations accounted for the remaining 39%. Single-sitecultures yielded linear Scatchard plots (Hill coefficients of0.97-1.03), whereas combinatorial expression generated cur-vilinear Scatchard plots (Hill coefficient of 0.51-0.75) withvarying proportions of the represented sites. In either case,the total number of receptors detected per cell was 200,000-250,000. WI-38 cells were able to produce the same range ofmaximum amounts ofPGE2 upon BK stimulation, which was5- to 20-fold, yielding 20-160 ng/106 cells, regardless of theprofile of receptor affinity sites expressed.Development of mAbs Against BK Receptor Affinity Forms.
Many BK B2 receptors expressed in other tissue systems andimmortalized cell lines appear to correspond to one of thethree affinity forms identified in WI-38 cells (5-7, 12-15). Tounderstand the basis of this diversity, mAbs specific fordifferent affinity forms of BK receptors were obtained byimmunizing with membranes from WI-38 cells concurrentlyexpressing I- and L-affinity receptors and screening for mAbinhibition of B2 BK-mediated PGE2 production in theHSDMLCj tumor cell line. The properties of six mAbsobtained by secondary expansion and subcloning are pre-sented in Table 1. The selected mAbs inhibited only PGE2production occurring at 25 nM BK, which saturated theI-affinity receptor (mAb All-5), or at 200 nM, which satu-rated the L site (mAbs G12-3 and C3-3); these mAbs did notinhibit basal or HAT-stimulated PGE2 production. Selectiveinhibition of BK receptor activation at 25 and 200 nM BKsuggested that mAbs All-5, C3-3, and G12-3 distinguishepitopes that contribute to differences in receptor affinity.mAbs C3-8, D9-7, and D9-11 blocked the stimulation ofPGE2production at both BK concentrations, suggesting that theyrecognized epitopes present in both I- and L-affinity forms.mAbs Selectively Recognize Human Lung Fibroblast BK
Receptor Affinity Forms. Fig. 1 demonstrates the ability ofmAbs All-5 and G12-3 alone and in combination to inhibitbinding of 15 nM [3H]BK to WI-38 membranes; this concen-
Table 1. Screening of mAbs directed against human fibroblastBK receptors
Maximal PGE2 output, ng
Hybridoma per 106 cells
medium 25 nM BK 200 nM BK
HAT alone 219 294All-5 94 294C3-3 219 185G12-3 239 156C3-8 145 162D9-7 145 100D9-11 94 144
A representative screen of final subclones is shown (see Materialsand Methods). HAT medium alone evoked 52 ng of PGE2 per 106cells, and buffer alone evoked basal PGE2 production of 2-10 ng per106 cells; BK stimulation shown includes this background. Specificinhibition was scored when a hybridoma supernatant (i) decreasedthe increment of BK-mediated PGE2 production by 50%o or more,(ii) did not suppress HAT medium background below its 52-ng level,and (iii) did not suppress buffer background, in three successivescreens each for initial 200 HAT-selected clones, their 22 positives,and 15 subclones.
Biochemistry: Jong et al.
Proc. Natl. Acad. Sci. USA 90 (1993)
tration is halfway between the Kd values of the I and L sitesexpressed by these cells and thus detects both sites. mAbA11-5 (I form specific) reduced binding of 15 nM [3H]BK by55%. Likewise mAb G12-3 (L form specific) reduced WI-38[3H]BK binding by 70%. No further inhibition was observedwith higher levels of either mAb alone. However, the twoantibodies in combination completely blocked specific[3H]BK binding. In contrast, an equivalent amount of acontrol mAb directed against fibronectin, which is expressedby WI-38 cells, did not inhibit [3H]BK binding. mAb C3-3,which is specific for the L-affinity form, inhibited [3H]BKbinding comparably to mAb G12-3 (n = 3). Thus these mAbsclearly were able to block both [3H]BK binding and BK-mediated PGE2 production.
In WI-38 cells concurrently expressing I- and L-affinity BKreceptor forms, mAb All-5 (2.5-5 ul of ascites fluid per150-,ul incubation) specifically blocked PGE2 production onlyat BK concentrations in the range of the I-affinity receptor.The component of the dose-response curves that reflectedPGE2 production at BK concentrations -25 nM was selec-tively abolished by mAb A1l-5, leaving residual dose-response curves with ED50 = 70-100 nM BK characteristicof a strictly L-form response (n = 6). A similar property hasbeen demonstrated by a mAb against one affinity form of theepidermal growth factor receptor in A431 cells (16). Affinity-purified mAb C3-3 against the L-form receptor proved to bea BK receptor agonist that stimulated PGE2 synthesis inWI-38 cells (Fig. 2). Its ED50 for PGE2 production was 124nM, <2-fold above the characteristic ED50 forBK itself at thehuman L-form receptor. Similar behavior has previouslybeen noted for anti-insulin receptor mAbs (17). mAb C3-3was originally identified by its ability to block PGE2 produc-tion at 200 nM but not 25 nM BK in rodent HSDMLCj cells.Its ability to mimic rather than inhibit BK biologic activity atthe human B2 receptor of the L form may reflect speciesdifferences in the interaction between this mAb and rodentvs. human B2 receptors.Immunoaffinty Isolation ofFibroblast BK B2 Receptors. We
enriched [3H]BK binding activity 1.6-fold in isolated WI-38membranes relative to intact or permeabilized cells. Theobserved BmS, increased from 510 fmol/mg of protein inwhole cells to 820 in membranes (n = 9); in both cells andmembranes, L-form receptor affinity was decreased 3-fold byGTP[y-S] treatment from Kd = 51-61 nM to Kd = 181-197nM, which is characteristic of G-protein-coupled receptors(18). Specific binding measured at 15 nM [3H]BK, which
100 -CTcn
* 80Fn
60I
x
00
40
20
No rrnAb All - G12-3 A1 1-5I-form L-form plus
G12-3
co0
c)oco0
w0-0)a
0 50 100 150[C3-3] nM
200 250 300
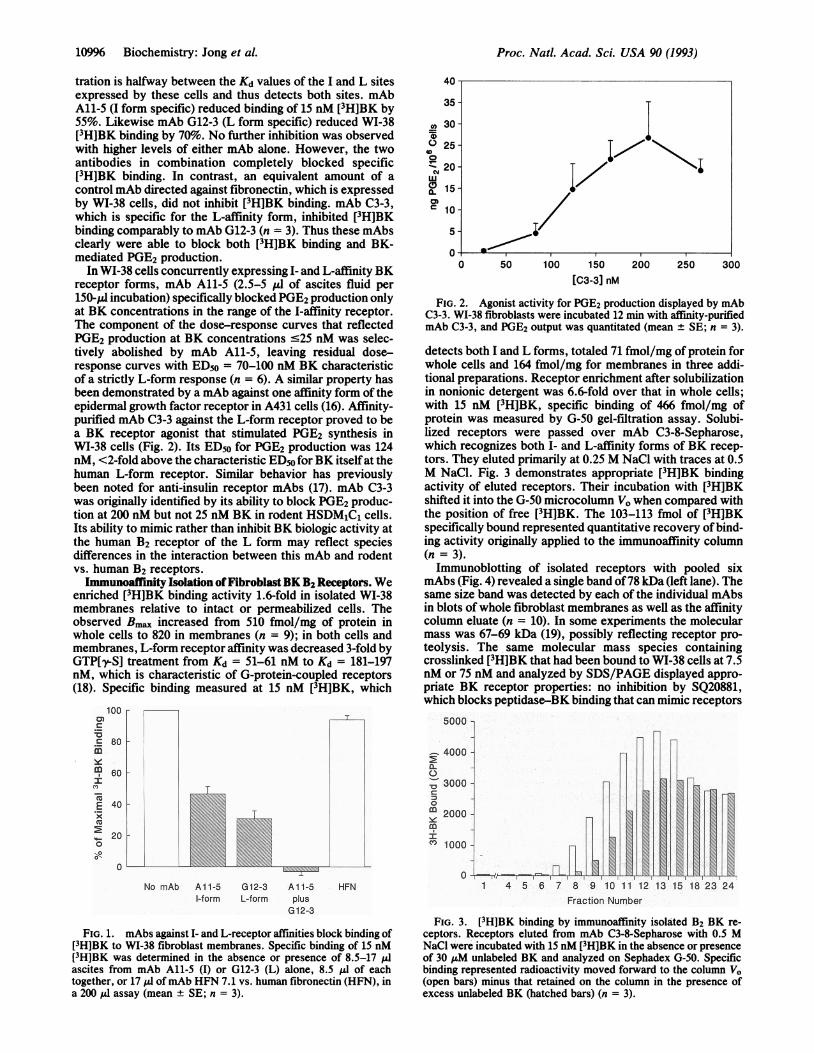
FIG. 2. Agonist activity for PGE2 production displayed by mAbC3-3. WI-38 fibroblasts were incubated 12 min with affinity-purifiedmAb C3-3, and PGE2 output was quantitated (mean ± SE; n = 3).
detects both I and L forms, totaled 71 fmol/mg of protein forwhole cells and 164 fmol/mg for membranes in three addi-tional preparations. Receptor enrichment after solubilizationin nonionic detergent was 6.6-fold over that in whole cells;with 15 nM [3H]BK, specific binding of 466 fmol/mg ofprotein was measured by G-50 gel-filtration assay. Solubi-lized receptors were passed over mAb C3-8-Sepharose,which recognizes both I- and L-affinity forms of BK recep-tors. They eluted primarily at 0.25 M NaCl with traces at 0.5M NaCl. Fig. 3 demonstrates appropriate [3H]BK bindingactivity of eluted receptors. Their incubation with [3H]BKshifted it into the G-50 microcolumn VO when compared withthe position of free [3H]BK. The 103-113 fmol of [3H]BKspecifically bound represented quantitative recovery of bind-ing activity originally applied to the immunoaffinity column(n = 3).Immunoblotting of isolated receptors with pooled six
mAbs (Fig. 4) revealed a single band of78 kDa (left lane). Thesame size band was detected by each of the individual mAbsin blots of whole fibroblast membranes as well as the affinitycolumn eluate (n = 10). In some experiments the molecularmass was 67-69 kDa (19), possibly reflecting receptor pro-teolysis. The same molecular mass species containingcrosslinked [3H]BK that had been bound to WI-38 cells at 7.5nM or 75 nM and analyzed by SDS/PAGE displayed appro-priate BK receptor properties: no inhibition by SQ20881,which blocks peptidase-BK binding that can mimic receptors
5000
4000
', 3000
C10
CO 1000
0HFN
FIG. 1. mAbs against I- and L-receptor affinities block binding of[3H]BK to WI-38 fibroblast membranes. Specific binding of 15 nM[3H]BK was determined in the absence or presence of 8.5-17 ILIascites from mAb All-5 (I) or G12-3 (L) alone, 8.5 ,ul of eachtogether, or 17 ,ul ofmAb HFN 7.1 vs. human fibronectin (HFN), ina 200 /, assay (mean ± SE; n = 3).
1 4 5 6 7 8 9 10 11 12 13 15 18 23 24
Fraction Number
FIG. 3. [3H]BK binding by immunoaffinity isolated B2 BK re-ceptors. Receptors eluted from mAb C3-8-Sepharose with 0.5 MNaCl were incubated with 15 nM [3H]BK in the absence or presenceof 30 ,uM unlabeled BK and analyzed on Sephadex G-50. Specificbinding represented radioactivity moved forward to the column VO(open bars) minus that retained on the column in the presence ofexcess unlabeled BK (hatched bars) (n = 3).
10996 Biochemistry: Jong et al.
-j

Proc. Natl. Acad. Sci. USA 90 (1993) 10997
(1), and 289%o inhibition by B2 antagonists but not a B1antagonist (n = 6). The [3H]BK-crosslinked receptor could besolubilized from whole cells in nonionic detergent and sep-arated by nondenaturing acrylamide gel electrophoresis. Pre-vious studies by others had indicated a molecular mass of250kDa for BK receptors thus solubilized into detergent micellecomplexes (20, 21). We found that cross-linked radiolabelfrom 7.5 nM [3H]BK binding (primarily I form) migrated asa 412-kDa band, whereas that from 75 nM [3H]BK binding(both I and L forms) had a peak at 321 kDa and a shoulder inthe range of412 kDa (n = 2); this implied a possible structuraldifference in I- vs. L-form receptors altering their apparentrelative size.BK B2 Receptors Are Tyrosine Phosphorylated and Trans-
duce Signals through Tyrosine Phosphorylation. The B2 BKreceptors affinity isolated from WI-38 fibroblasts containedphosphotyrosine residues, detected by immunoblotting withanti-phosphotyrosine mAb Py2O (Fig. 4, right lane). BK B2receptors in WI-38 fibroblasts thus represent a constitutivesubstrate for one or more tyrosine kinase activities. Todetermine if tyrosine phosphorylation is essential for theexpression of BK receptor activity, we treated WI-38 cellswith the tyrosine kinase inhibitor genistein and examined itsimpact on BK-mediated PGE2 production (Fig. 5). Genisteinspecifically inhibited BK-mediated PGE2 production with anIC50 of 8 ,uM and complete inhibition at 50 ,uM, withoutaffecting basal PGE2 synthesis or [3H]BK binding. Thustyrosine phosphorylation is critical in accomplishing BKreceptor signal transduction, which links the latter to PGE2synthesis. The tyrosine kinase activity mediating this stepmay or may not be the same kinase that acts upon the BKreceptor itself.
DISCUSSIONOur exploration of BK receptor expression in WI-38 humanlung fibroblasts seeks to identify properties that relate thestructure and function ofthis receptor system to its role in thebiological activities of fibroblasts. Structural and regulatorydetails,regarding BK receptors have not been available at theprotein level due to receptor instability to traditional meansof purification. Moreover, expression cloning ofB2 receptorsfrom different species and cell types has not yet provided anunderstanding of mechanisms that might contribute to BKreceptor diversity (5-7). We therefore raised mAbs directedagainst the biologic activity of B2 BK receptors. By screeningfor differential inhibition ofPGE2 production at two differentBK concentrations, we selected for mAbs that differentiallyinteract with the I- or L-affinity forms, as well as mAbsrecognizing epitopes that are common to both forms. Usingthese mAbs, we have now isolated and characterized BK B2
97 _ 78 8-
69-46-
FIG. 4. Immunoaffrnity-isolated BK receptor protein is tyrosinephosphorylated. Solubilized WI-38 B2 BK receptors were adsorbedto mAb C3-8-Sepharose, eluted with 0.25 M NaCl, and analyzed bySDS/PAGE. Left lane, immunoblot with six pooled mAbs, indicat-ing receptor from 2 x 106 cells at 78 kDa (arrow) (n = 10). Blottingof this band was highly specific for anti-BK receptor mAbs; it did notimmunoblot with mAb HFN 7.1, which blotted fibronectin at 200-220 kDa in WI-38 membranes. Right lane, immunoblot with mAbPy2O (ICN) against phosphotyrosine residues. Py20 immunoblottingwas completely blocked by 2 mM phosphotyrosine; n = 6 where Py20and anti-receptor mAbs blotted the same molecular mass band.
1400(0
0 1200.0ir 100
-c 80'aE0 60
o 40-Wc02 200a.
0 _0.0 0.5 1.0
log [Genistein] s1M1.5 2.0
FIG. 5. Genistein specifically inhibits BK-mediated PGE2 pro-duction. Cells were incubated with genistein for 32 min; controlbuffer or 25 nM BK was added, and PGE2 elaborated in 12 min wasmeasured. Basal PGE2 output was subtracted (mean ± SE; n = 5).
receptors as constitutively expressed in a single cell type.The mAb immunoaffinity isolated receptor species displaysappropriate [3H]BK binding capability. Its molecular mass of78 kDa is similar to Vl vasopressin, bombesin, and otherG-protein-coupled receptors (22-25) and larger than the mo-lecular mass of 41-42 kDa predicted from BK B2 receptorcloned sequences (5-7), which omits the contribution ofN-linked glycosylation (19). I- and L-receptor affinity formsidentified by each form-specific mAb represent proteins ofthe same or a highly similar molecular mass.We have further demonstrated tyrosine phosphorylation of
these receptors, a posttranslational modification that hasimplications for their function and regulation. Sequenceinformation on a cloned human B2 BK receptor has disclosedfive tyrosine residues that would be candidates for thismodification, by virtue of their location in cytoplasmic do-mains or just at the border of putative transmembranesegments. The C-terminal tail, often a region important inreceptor interactions with signal transduction pathways,bears such a tyrosine residue (5, 7). Tyrosine phosphoryla-tion of BK B2 receptors represents, to our knowledge, thefirst example of this modification in a neuropeptide G-pro-tein-coupled receptor. /32-Adrenergic receptor tyrosine phos-phorylation (26) has been attributed to insulin receptor tyro-sine kinase activity. G-protein-coupled receptors lack anendogenous tyrosine kinase activity, in contrast to classicalgrowth factor or insulin receptors (26-29). Growth factors inthe culture medium may stimulate growth factor receptortyrosine kinase activities in WI-38 prior to receptor isolation.Activation ofinsulin or fibroblast growth factor receptors (27)augments BK receptor signal transduction, postulated tooccur by a tyrosine phosphorylation mechanism; by analogywith the A32-adrenergic receptor, BK receptors may representsubstrates for these and/or other tyrosine kinases (30-32).
Finding receptor tyrosine phosphorylation prompted us toidentify a requirement for tyrosine phosphorylation in signaltransduction leading from BK receptor activation to PGE2production. Thus the widespread role of tyrosine phosphor-ylation in transducing the signal from growth factor receptorsextends to G-protein-coupled receptors for BK and for bom-besin (32), an intriguing parallel given the mitogenic activitydistinctive for these two neuropeptides. The tyrosine phos-phorylation-dependent link is an absolute requisite betweenBK receptor activation and arachidonate metabolism, sincegenistein inhibits WI-38 BK-mediated PGE2 synthesis com-pletely but inhibits G-protein-coupled stimulation of inositoltrisphosphate formation and Ca2+ signaling by only 35-50%o(33, 34). The arachidonate and inositol trisphosphate meta-
Biochemistry: Jong et al.

Proc. Natl. Acad. Sci. USA 90 (1993)
bolic pathways may differ in their phosphorylation cascadedownstream from receptor activation. One such cascademember has recently been identified as focal adhesion kinase(FAK), itself a tyrosine kinase, which becomes tyrosinephosphorylated in response to bombesin (32).
Diverse affinity forms of BK receptors may reflect molec-ular differences in receptor species, as indicated by theselective recognition properties of mAbs A11-5, G12-3, andC3-3 toward the repertoire of BK B2 receptor affinitiesmediating a common biologic activity. These mAbs are not"conformation specific" in the traditional sense. They ap-pear to target receptor configurational differences in wholecells and membrane preparations, yet sufficient epitope rec-ognition features for mAb recognition are preserved upon thedenaturing conditions of SDS/PAGE. BK receptor affinityforms may arise from different primary sequences, posttrans-lational modification of a single polypeptide, or associationwith a molecular companion.
Diversity in primary sequences within various G-protein-coupled receptor subfamilies can arise from different geneproducts or alternative splicing (35); it encompasses regionsof homology that would be consistent with mAb recognitionof BK receptor affinity form-specific and common epitopes.Posttranslational modification of a common polypeptidewould agree with the relatively similar BK B2 receptornumber expressed by WI-38 cells whatever the specificaffinity forms present. Either ofthese two mechanisms wouldbe consistent with a similar molecular mass of I and L formsby SDS/PAGE and yet an apparent difference of '-90 kDaunder nondenaturing electrophoretic conditions, which aremore sensitive to charge differences affecting receptor con-formation. Association in a complex with a molecular com-panion could also explain the latter receptor behavior. Re-ceptor complexes with G-protein subunits and coupled vs.uncoupled states have been invoked as a basis for affinitydiversity (18, 36, 37), although the WI-38 L form is clearlyG-protein-coupled itself, not simply an uncoupled I form (18).Tyrosine phosphorylation of BK receptors suggests theymight utilize this residue to dock with another protein via anSH2 domain. Signaling pathways mediated by classicalgrowth factor receptors employ this mechanism, physicallyassociating with a variety of cytoplasmic macromolecules(reviewed in ref. 31). Some candidate phosphorylatable ty-rosine residues in the cloned B2 receptor sequence areadjacent to known SH2 domain recognition motifs (5, 7, 38).Thus this cellular strategy for assembling receptor-companion complexes may be applicable to G-protein-coupled receptors as well.The three BK receptor affinity forms expressed in WI-38
human lung fibroblasts have respective Kd values corre-sponding closely to in vivo BK levels in humans (reviewed inref. 1) under normal baseline conditions (H form) and allergyor inflammation (I and L forms). Fibroblasts' ability tomobilize successive receptor affinity forms in tissue injurysituations with escalating BK generation may govern thebiologic response potential of tissues. We have identifieddifferent cellular signaling/second messenger pathways thatdrive expression of specific profiles of H, I, and L BKreceptor affinity forms (N.L.B., L.R.D., and Y.-J.I.J., un-published results). Thus targeted receptor mobilization maylink internal cellular metabolic processes to extracellularevents that prompt cells to adjust their complement of BKreceptors in a given environment.
We thank Drs. Steven J. Hamrs and Donna M. Crecelius forcontributing to this work. This work was supported by Grant HL36409 from the National Institutes of Health, a grant from theAmerican Federation for Aging Research (AFAR), and an AmericanHeart Association Established Investigatorship to N.L.B.
1. Baenziger, N. L., Jong, Y.-J. I., Yocum, S. A., Dalemar, L. R.,Wilhelm, B., Vavrek, R. & Stewart, J. M. (1992) Eur. J. Cell Biol.58, 71-80.
2. Steranka, L. R., Manning, D. C., DeHaas, C. J., Ferkany, J. W.,Borosky, S. A., Connor, J. R., Vavrek, R. J., Stewart, J. M. &Snyder, S. H. (1988) Proc. Natl. Acad. Sci. USA 85, 3245-3249.
3. Goldstein, R. H. & Wall, M. (1984) J. Biol. Chem. 259, 9263-9268.4. Becherer, P. R., Mertz, L. F. & Baenziger, N. L. (1982) Cell 30,
243-251.5. Hess, J. F., Borkowski, J. A., Young, G. S., Strader, C. S. &
Ransom, R. W. (1992) Biochem. Biophys. Res. Commun. 184,260-268.
6. McEachern, A. E., Shelton, E. R., Bhakta, S., Obernolte, R.,Bach, C., Zuppan, P., Fujisaki, J., Aldrich, R. W. & Jarnagin, K.(1991) Proc. Natl. Acad. Sci. USA 88, 7724-7728.
7. Eggerickx, D., Raspe, E., Bertrand, D., Vassart, G. & Parmentier,M. (1992) Biochem. Biophys. Res. Commun. 187, 1306-1313.
8. Ahnert-Hilger, G., Bader, M.-F., Bhakdi, S. & Gratzl, M. (1989) J.Neurochem. 52, 1751-1758.
9. Bhakdi, S., Roth, M., Sziegoleit, A. & Truanum-Jensen, M. (1984)Infect. Immun. 46, 394-400.
10. Laemmli, U. K. (1970) Nature (London) 227, 680-685.11. Towbin, H., Staehelin, T. & Gordon, J. (1979) Proc. Natl. Acad.
Sci. USA 76, 4350-4354.12. Manning, D. C., Vavrek, R., Stewart, J. M. & Snyder, S. H. (1986)
J. Pharmacol. Exp. Ther. 37, 504-512.13. Snider, R. M. & Richelson, E. (1984) J. Neurochem. 43, 1749-1754.14. Roberts, R. A. & Gullick, W. J. (1990) Biochemistry 29, 1975-1979.15. Fredrick, M. J. & Odya, C. E. (1987) Eur. J. Pharmacol. 134,
45-52.16. Kawamoto, T., Sato, J. D., Le, A., Polikoff, J., Sato, G. H. &
Mendelsohn, J. (1983) Proc. Natl. Acad. Sci. USA 80, 1337-1341.17. Steele-Perkins, G. & Roth, R. A. (1990) J. Biol. Chem. 265, 9458-
9463.18. Dickey, B. F., Fishman, J. B., Fine, R. E. & Navarro, J. C. (1987)
J. Biol. Chem. 263, 2577-2580.19. AMa, S. A., Buschko, J., Quitterer, U., Maidhof, A., Haasemann,
M., Breipohl, G., Knolle, J. & Muller-Esterl, W. (1993) J. Biol.Chem. 268, 17277-17285.
20. Faussner, A., Heinz-Erian, P., Klier, C. & Roscher, A. A. (1991) J.Biol. Chem. 266, 9442-9446.
21. Snell, P. H. & Snell, C. R. (1989)Biochem. Soc. Trans. 17,569-570.22. Fishman, J. B., Dickey, B. F. & Fine, R. E. (1987) J. Biol. Chem.
262, 14049-14055.23. Millar, J. B. A. & Rozengurt, E. (1990) J. Biol. Chem. 265, 12052-
12058.24. Benovic, J. L., Regan, J. W., Matsui, H., Mayor, F., Cotecchia, S.,
Leeb-Lundberg, L. M., Caron, M. G. & Lefkowitz, R. J. (1987) J.Biol. Chem. 262, 17251-17253.
25. Leeb-Lundberg, L. M., Cotecchia, S., DeBlasi, A., Caron, M. G.& Lefkowitz, R. J. (1987) J. Biol. Chem. 262, 3098-3105.
26. Hadcock, J. R., Port, J. D., Gelman, M. S. & Malbon, C. C. (1992)J. Biol. Chem. 267, 26017-26022.
27. Pandiella, A. & Meldolesi, J. (1989) J. Biol. Chem. 264, 3122-3130.28. Zhang, B., Tavares, J. M., Ellis, L. & Roth, R. A. (1991) J. Biol.
Chem. 266,990-996.29. Ek., B. & Heldin, C.-H. (1982) J. Biol. Chem. 257, 10486-10491.30. Morrison, D. K., Kaplan, D. R., Escobedo, J. A., Rapp, U. R.,
Roberts, T. M. & Williams, L. T. (1989) Cell 58, 649-657.31. Twamley, G. M., Kypta, R. M., Hall, B. & Courtneidge, S. (1992)
Oncogene 7, 1893-1901.32. Zachary, I., Sinnet-Smith, J. & Rozengurt, E. (1992) J. Biol. Chem.
267, 19031-19034.33. Leeb-Lundberg, L. M. & Song, X.-H. (1991) J. Biol. Chem. 266,
7746-7749.34. Sargeant, P., Farndale, R. W. & Sage, S. 0. (1993) FEBS Lett. 315,
242-246.35. Todd, R. D., Khurana, T. S., Sajovic, P., Stone, K. R. & O'Malley,
K. L. (1989) Proc. Natl. Acad. Sci. USA 86, 10134-10138.36. Mathis, S. A. & Leeb-Lundberg, L. M. F. (1991) Biochem. J. 276,
141-147.37. Werling, L., Puttfarcken, P. & Cox, B. (1988) Mol. Pharmacol. 33,
423-431.38. Songyang, Z., Shoelson, S. E., Chaudhuri, M., Gish, G., Pawson,
T., Haser, W. G., King, F., Roberts, T., Ratnofsky, S., Lechleider,R. J., Neel, B.-G., Birge, R.-B., Fajardo, J. E., Chou, M. M.,Hanafusa, H., Schaffhausen, B. & Cantley, L. C. (1993) Cell 72,767-778.
10998 Biochemistry: Jong et al.