bases quimicas de la estructura y plegado de …...bases quimicas de la estructura y plegado de...
TRANSCRIPT

BIOQUIMICA DE PROTEINAS BASES QUIMICAS DE LA ESTRUCTURA Y
PLEGADO DE PROTEINAS PROF. LUCIA CHEMES ([email protected])
J.C. Kendrew, modelos de Mioglobina

BIBLIOGRAFIA
FERSHT, A. STRUCTURE AND MECHANISM IN PROTEIN SCIENCE (1999) FREEMAN PRESS, CAP 1, 11, 17 18 Y 19
BRANDEN AND TOOZE. INTRODUCTION TO PROTEIN STRUCTURE (1999) GARLAND PUBLISHING
BERG TYMOCZKO AND STRYER. BIOCHEMISTRY. EDITORIAL FREEMAN AND COMPANY

PROTEINAS
MIOGLOBINA

PROTEINAS
MIOGLOBINA
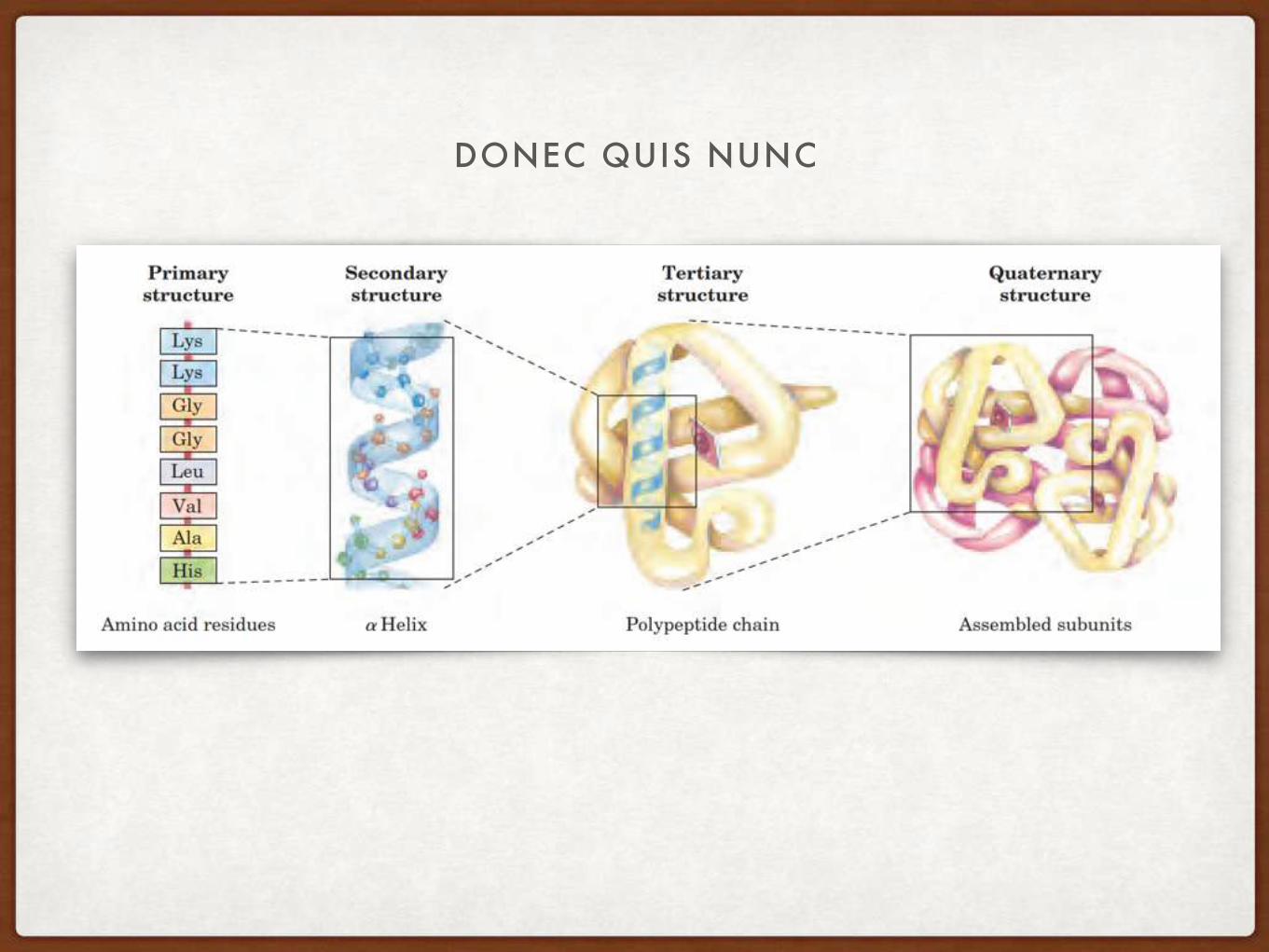
DONEC QUIS NUNC

LAS PROTEÍNAS NATURALES SON POLÍMEROS DE L -AMINOÁCIDOS
PROTEINAS
ISÓMERO L ISÓMERO D
Propiedades de aminoácidos: • Son moléculas quirales • Existen 20 variantes naturales de grupo R (cadena lateral)
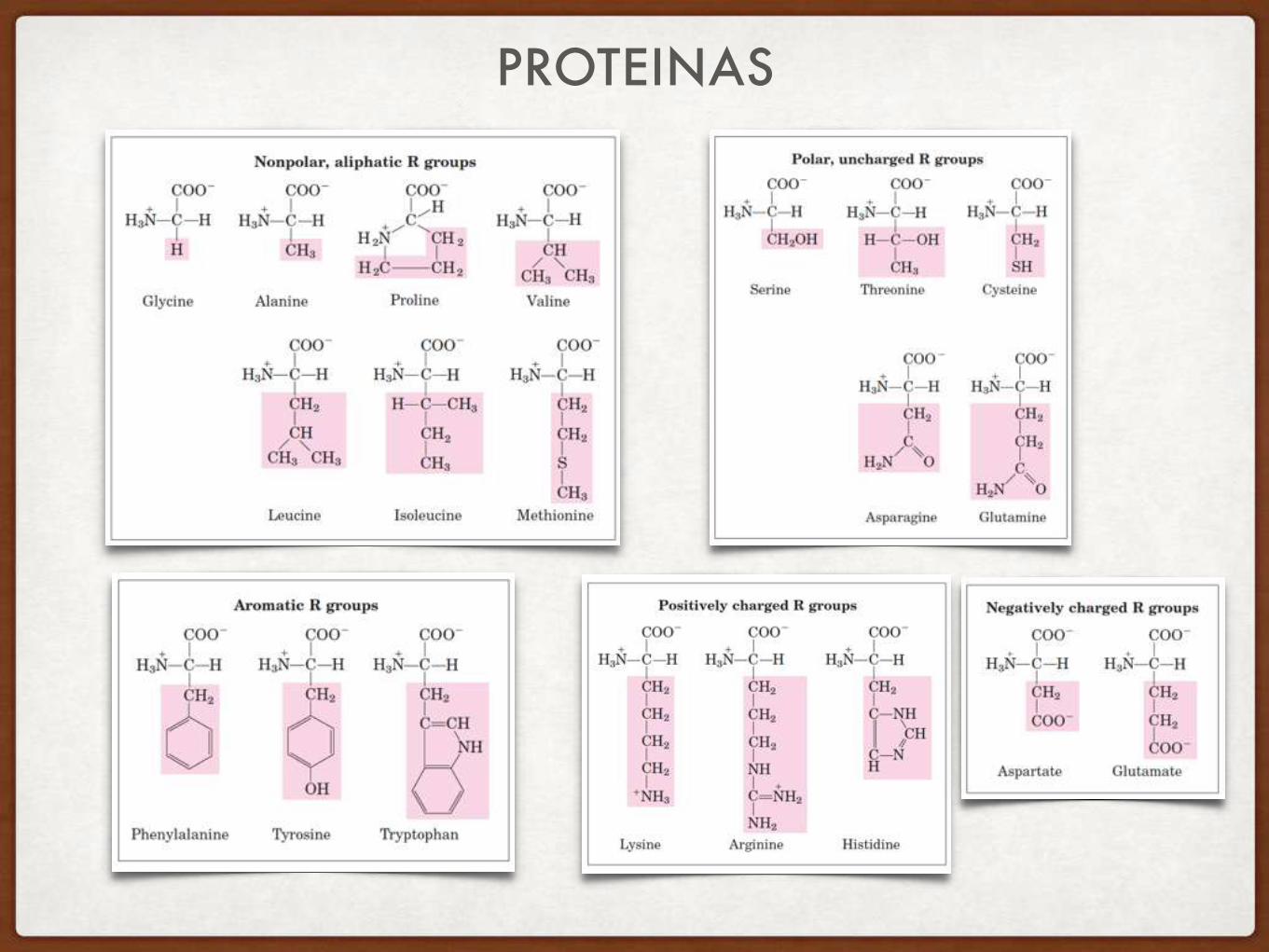
PROTEINAS
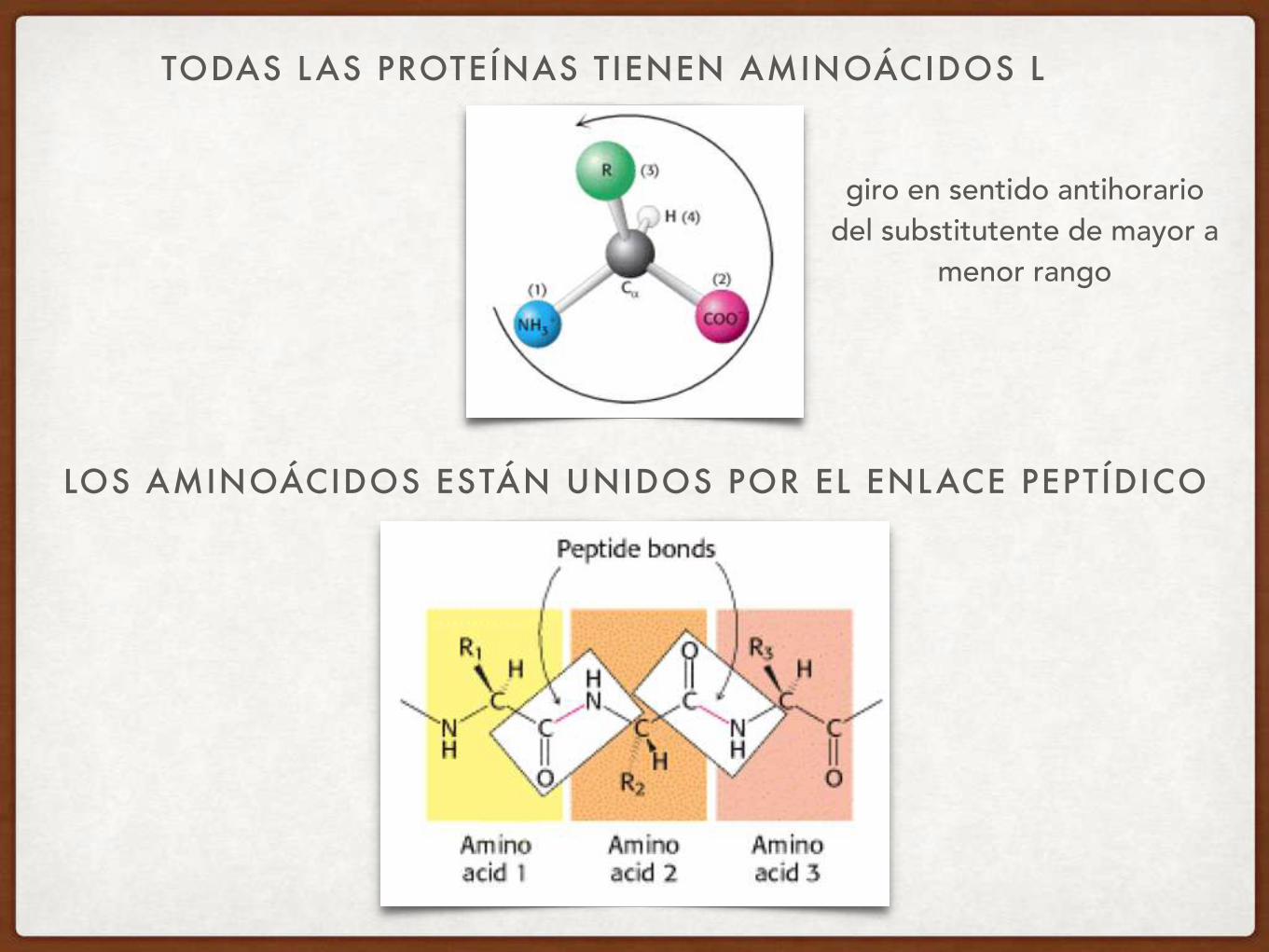
TODAS LAS PROTEÍNAS TIENEN AMINOÁCIDOS L
LOS AMINOÁCIDOS ESTÁN UNIDOS POR EL ENLACE PEPTÍDICO
giro en sentido antihorario del substitutente de mayor a
menor rango

EL ENLACE PEPTÍDICO EN DETALLE QUÍMICO
• Planar • Configuración “Trans” • Carácter de doble enlace entre C-N (enlace acortado)
Simple 1.49 Aº Doble 1.27 Aº
C-N Enlace peptídico 1.33 Aº 18-21 kcal/mol
Enlace C-N

UNA CADENA POLIPEPTIDICA
PROTEINAS

DEFINICION DE ANGULOS TORSIONALES
R1
R2
R1 R2
180 TRANS
0 CIS
cadena lateral C N H O
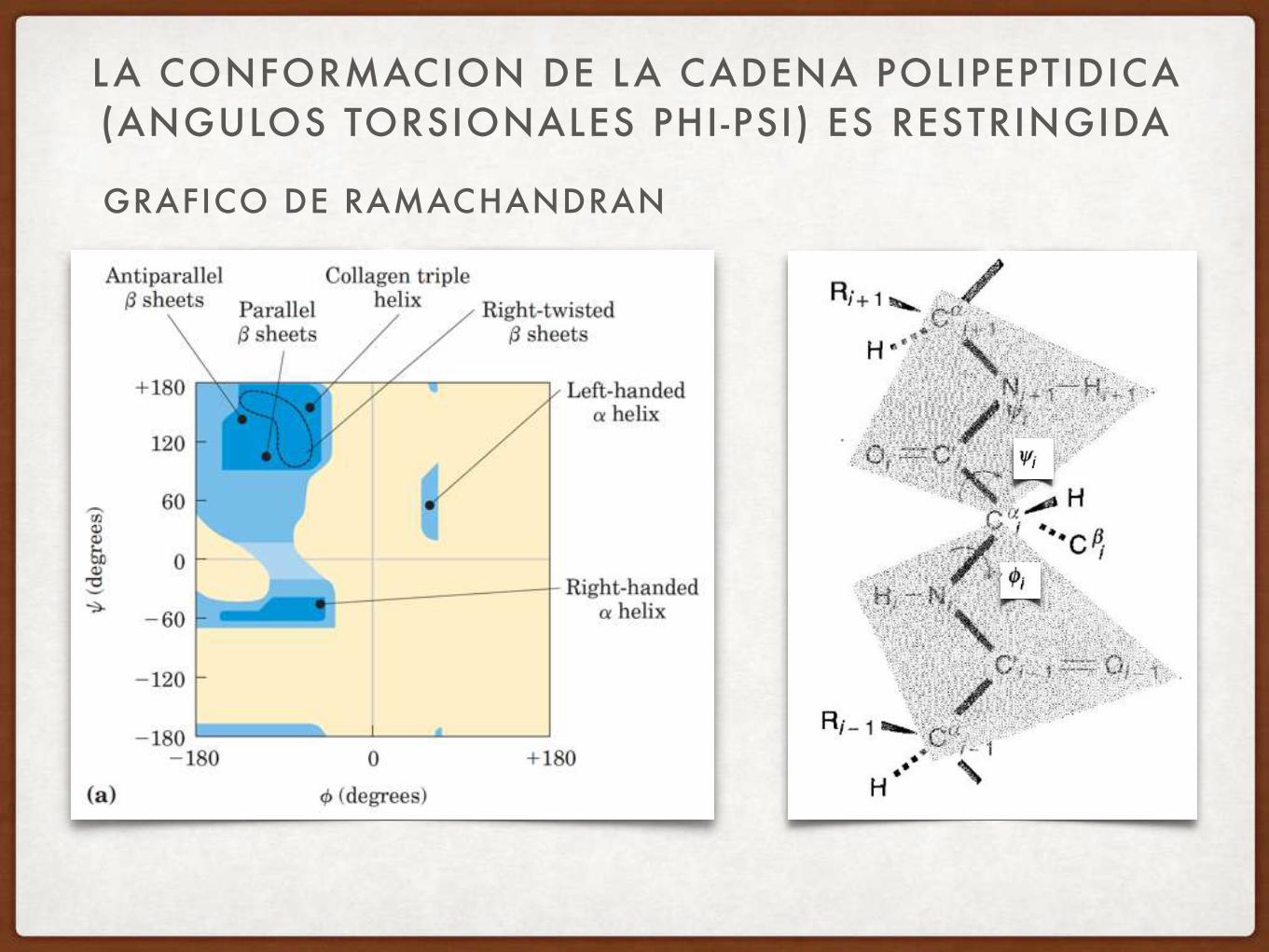
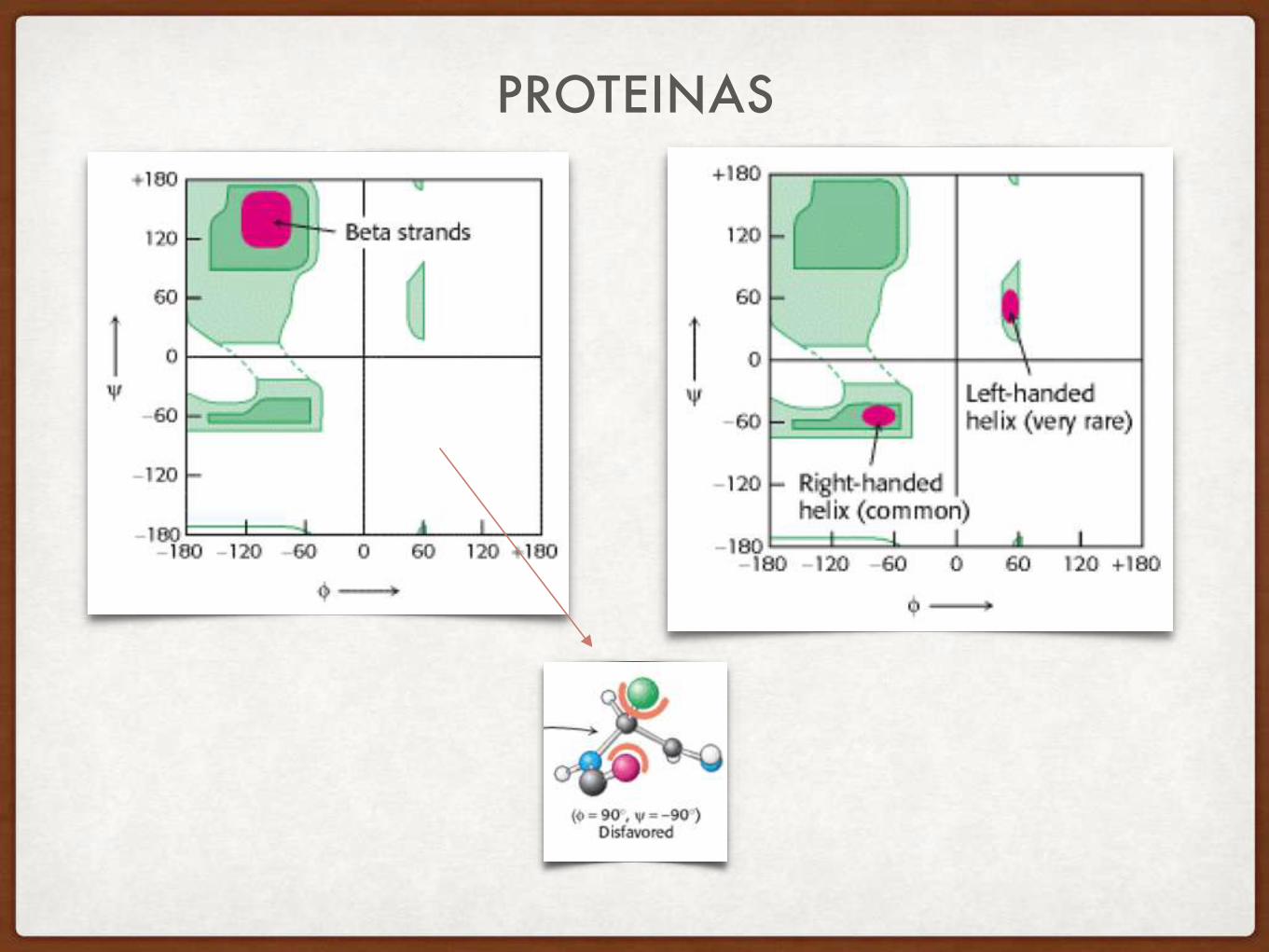
GRAFICO DE RAMACHANDRAN
LA CONFORMACION DE LA CADENA POLIPEPTIDICA (ANGULOS TORSIONALES PHI-PSI) ES RESTRINGIDA
PROTEINAS

DISTRIBUCIÓN DE ÁNGULOS PHI PSI EN PROTEÍNAS
La gran mayoría de los ángulos Phi-Psi (un par define a cada aminoácido) caen dentro de dos grandes zonas del espacio

AMINOACIDOS CON CONFORMACIÓN ATÍPICA
GLICINA (GLY, G)
• No es quiral • Alta flexibilidad puebla zonas
“prohibidas” del Ramachandran
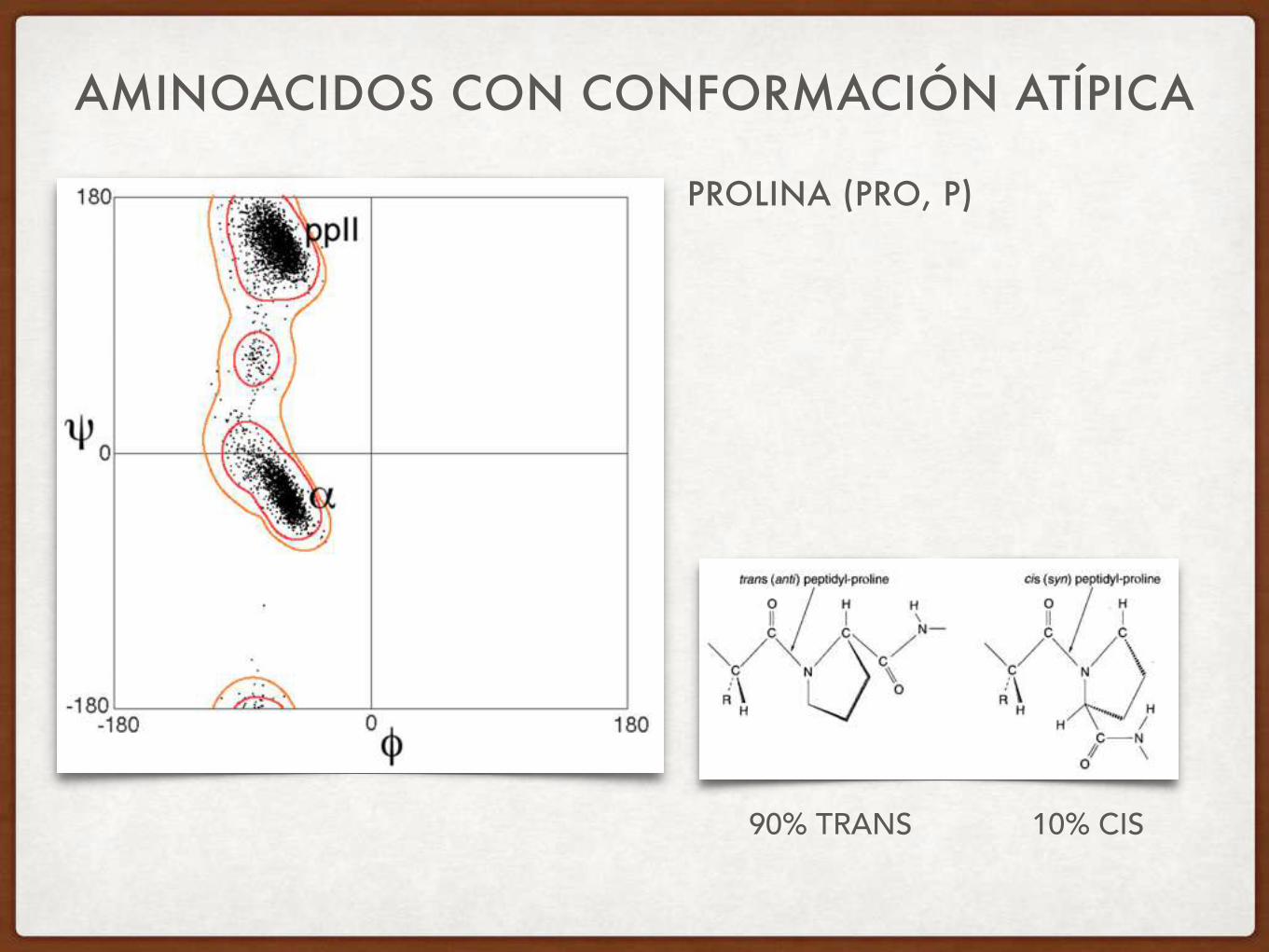
PROLINA (PRO, P)
AMINOACIDOS CON CONFORMACIÓN ATÍPICA
90% TRANS 10% CIS

PROTEINASALFA HELICE: 30% DE LOS RESIDUOS EN PROTEÍNAS
ALFA HELICE L. PAULING & R. COREY, 1951
ON THE NATURE OF THE CHEMICAL BOND CORNELL UNIVERSITY PRESS 1960
Experimentos de difracción de queratina
• Puentes H entre grupos carbonilo de residuo i y el grupo amida de i+4
• Estructura compacta de la cadena polipeptídica
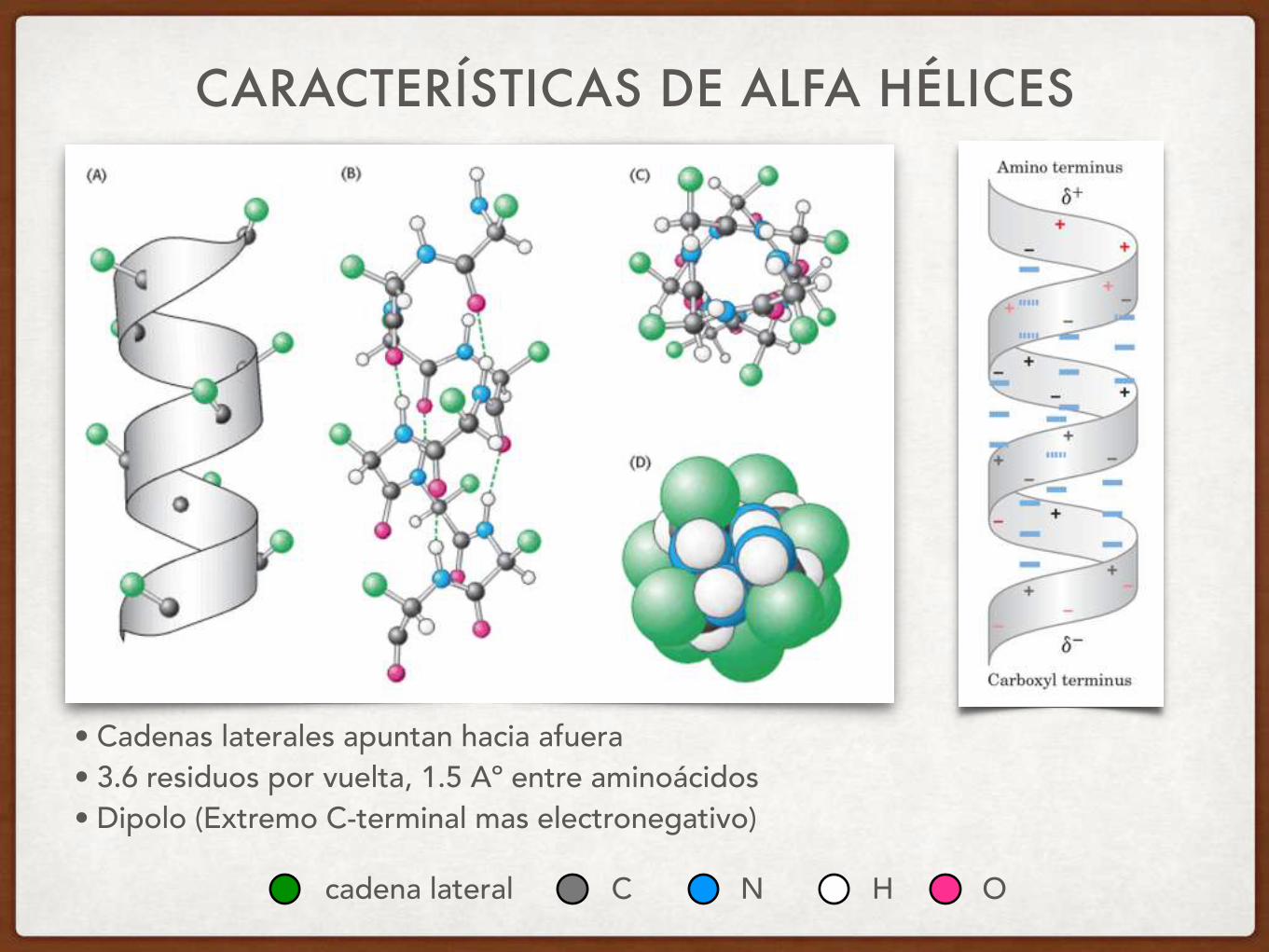
CARACTERÍSTICAS DE ALFA HÉLICES
• Cadenas laterales apuntan hacia afuera • 3.6 residuos por vuelta, 1.5 Aº entre aminoácidos • Dipolo (Extremo C-terminal mas electronegativo)
cadena lateral C N H O

EL DIPOLO DE LA HÉLICE PROVIENE DE LA DIRECCIÓN DE LOS PUENTES HIDROGENO
C
O
N
H OXÍGENO: ACEPTOR DE HNITRÓGENO: DONOR DE H
PUENTE H

ALFA HELICES
HELICE ANFIPÁTICA
CONECTIVIDAD
HELICE HIDROFÓBICA
Ri R (i+4)
aminoácidos hidrofílicos
aminoácidos hidrofóbicos aminoácidos hidrofóbicos
aminoácidos hidrofóbicos
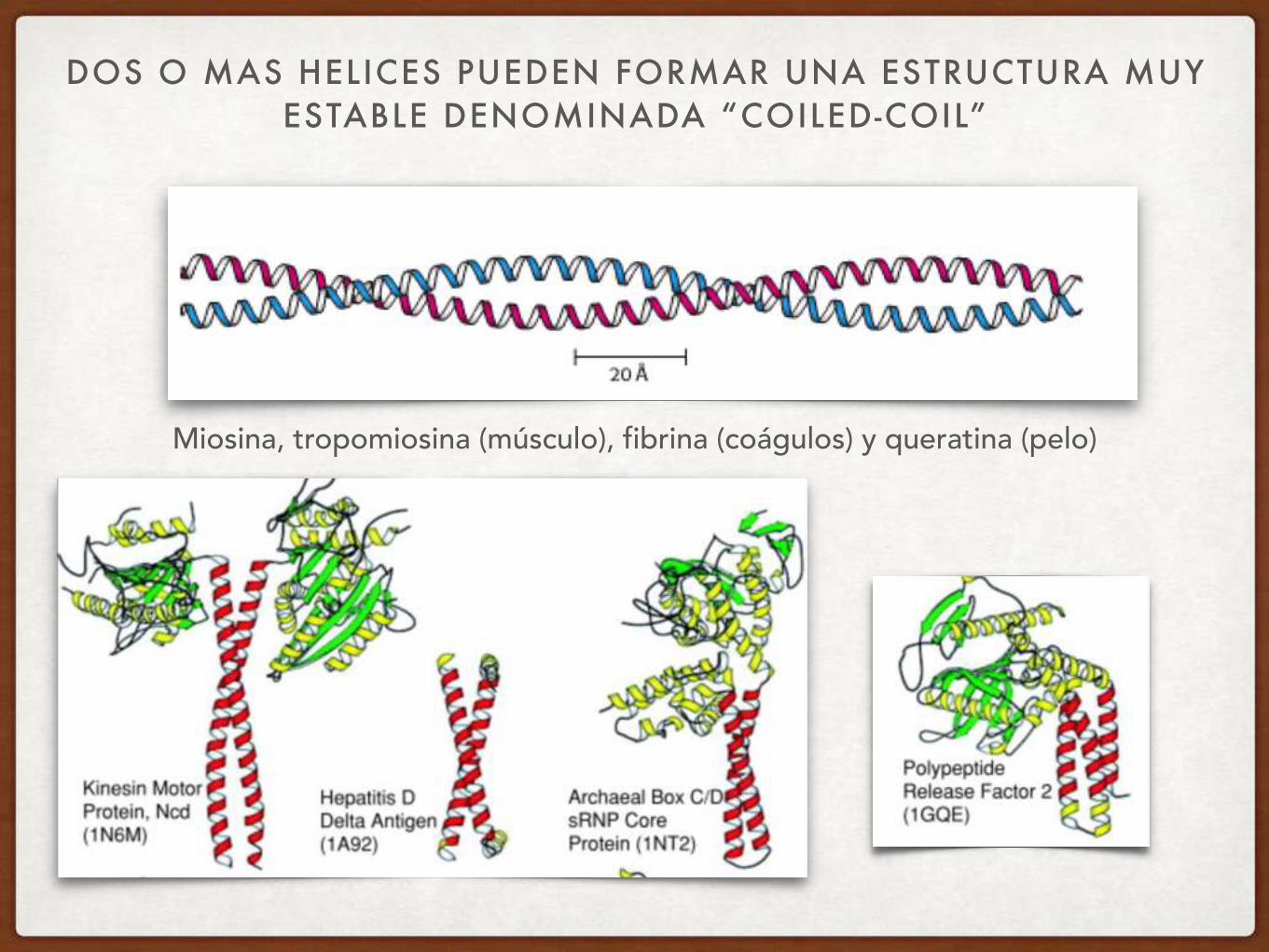
DOS O MAS HELICES PUEDEN FORMAR UNA ESTRUCTURA MUY ESTABLE DENOMINADA “COILED-COIL”
Miosina, tropomiosina (músculo), fibrina (coágulos) y queratina (pelo)

QUÉ FUERZAS ESTABILIZAN ESTA ESTRUCTURA?
COILED COILS
1- INTERACCIONES HIDROFÓBICAS ENTRE LOS AMINOÁCIDOS “A” Y “D” (D ES SIEMPRE LEUCINA, “A” FRECUENTEMENTE HIDROFÓBICO
2- PUENTES SALINOS ENTRE LAS POSICIONES “G” Y “E”
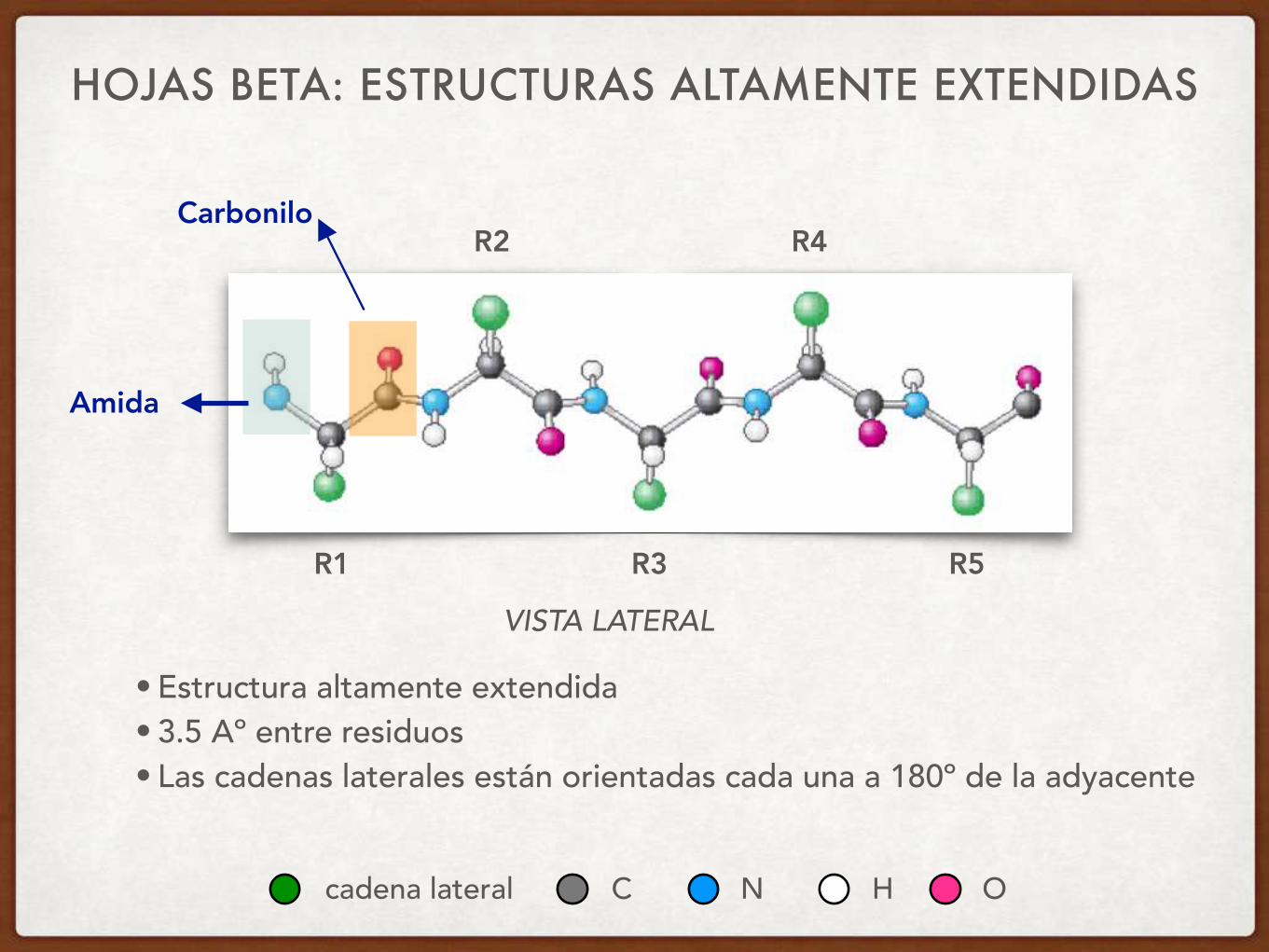
HOJAS BETA: ESTRUCTURAS ALTAMENTE EXTENDIDAS
• Estructura altamente extendida • 3.5 Aº entre residuos • Las cadenas laterales están orientadas cada una a 180º de la adyacente
R1
R2
R3
R4
R5
Carbonilo
Amida
VISTA LATERAL
cadena lateral C N H O

HOJA BETA ANTIPARALELA
cadena lateral C N H O
Carbonilo (aceptor)
Gr. Amido (donor)
VISTA SUPERIOR
NtCt
CtNt
LAS CADENAS R (VERDES) ENTRAN Y SALEN DEL PIZARRON
cadena lateral C N H O
CtNt
CtNt
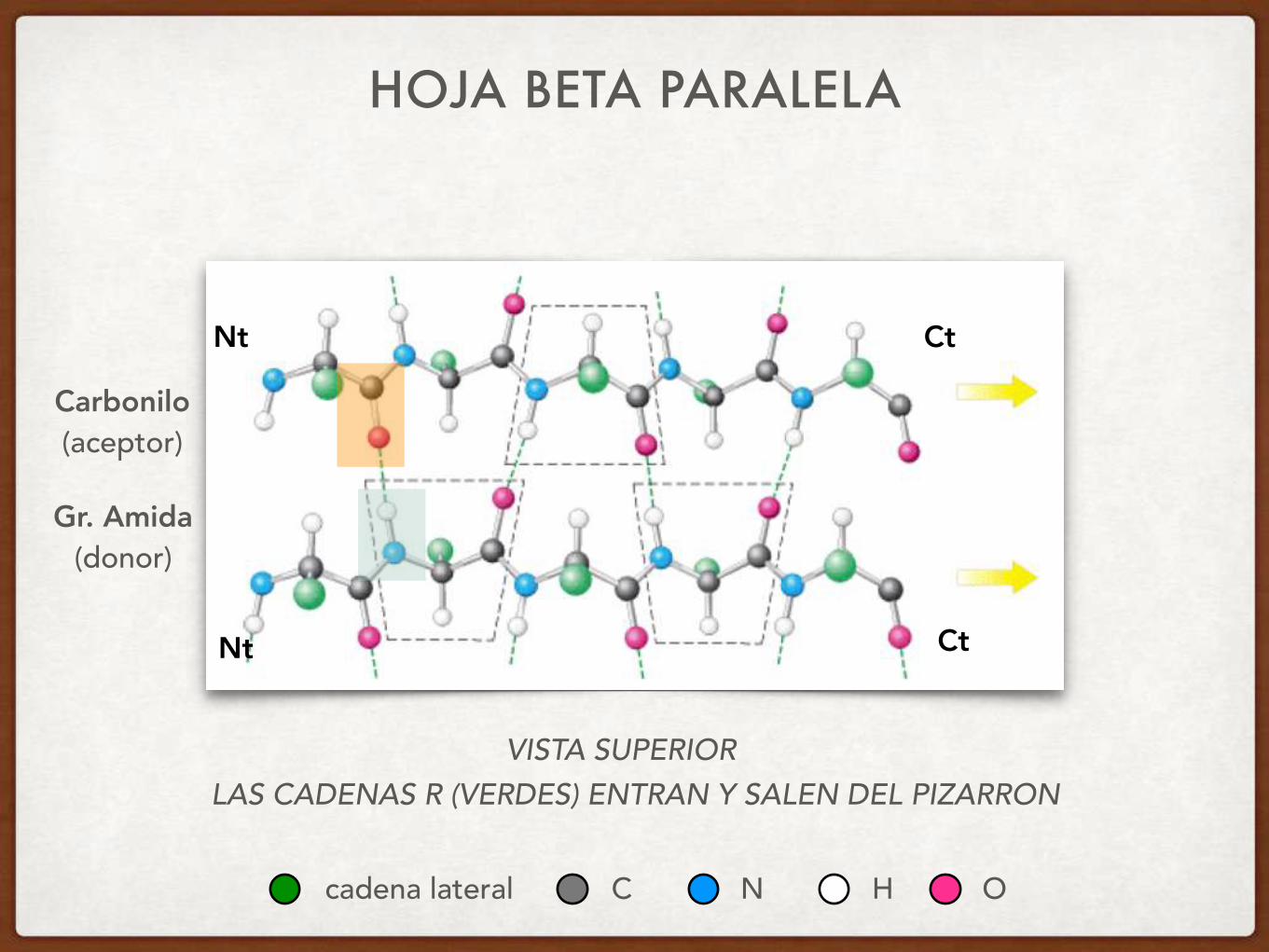
HOJA BETA PARALELA
VISTA SUPERIOR
Carbonilo (aceptor)
Gr. Amida (donor)
LAS CADENAS R (VERDES) ENTRAN Y SALEN DEL PIZARRON

“GIROS” ESTRUCTURAS DE VARIOS TIPOS DE GIROS
• Carbonilo de residuo i (iZQ) unido a grupo amida de residuo i+3 (DER)
• En el Tipo II, X siempre es Gly (G)

ENLACE COVALENTE QUE ESTABILIZA LA ESTRUCTURA TERCIARIA DE LAS PROTEÍNAS
PUENTE DISULFURO
Enlace covalente que se forma mediante una reacción de
oxidación-reducción (REDOX)
Más común en proteínas extracelulares Puede formarse dentro de una cadena (intracatenario) o unir dos subunidades (intercatenario)

Frecuencia relativa con que se encuentran los aminoácidos en diferentes estructuras secundarias de las proteínas
Val, Ile, Thr: Aminoácidos con ramificaciones en el C beta
Desfavorecidos en alfa hélices por choques estéricos con la cadena. Se acomodan bien en hojas beta donde los R se alejan de la estructura de la hoja

Frecuencia relativa con que se encuentran los aminoácidos en diferentes estructuras secundarias de las proteínas
Ser, Asp, Asn: Aminoácidos con cadenas polares que pueden formar puentes H Desfavorecidos en alfa hélices por competir con los ptes H de cadena ppal. Son más abundantes en Giros
Pro: Tiene valor Phi fijo de ~60º: estructura restringida y no tiene NH donor de ptes H: tiende a desestabilizar tanto hélice como hoja beta Es abundante en Giros
Gly: Muy flexible, más abundante en Giros

ESTRUCTURA TERCIARIA DE PROTEÍNAS
CÓMO DEFINIMOS UN “DOMINIO” PROTEICO?
Estructura plegada de la mioglobina

ESTRUCTURAL
LO PODEMOS DEFINIR A VARIOS NIVELES
EVOLUTIVO
FUNCIONAL
UNA UNIDAD INDEPENDIENTE DE PLEGAMIENTO
UNA UNIDAD EVOLUTIVA
UN MODULO O UNIDAD FUNCIONAL
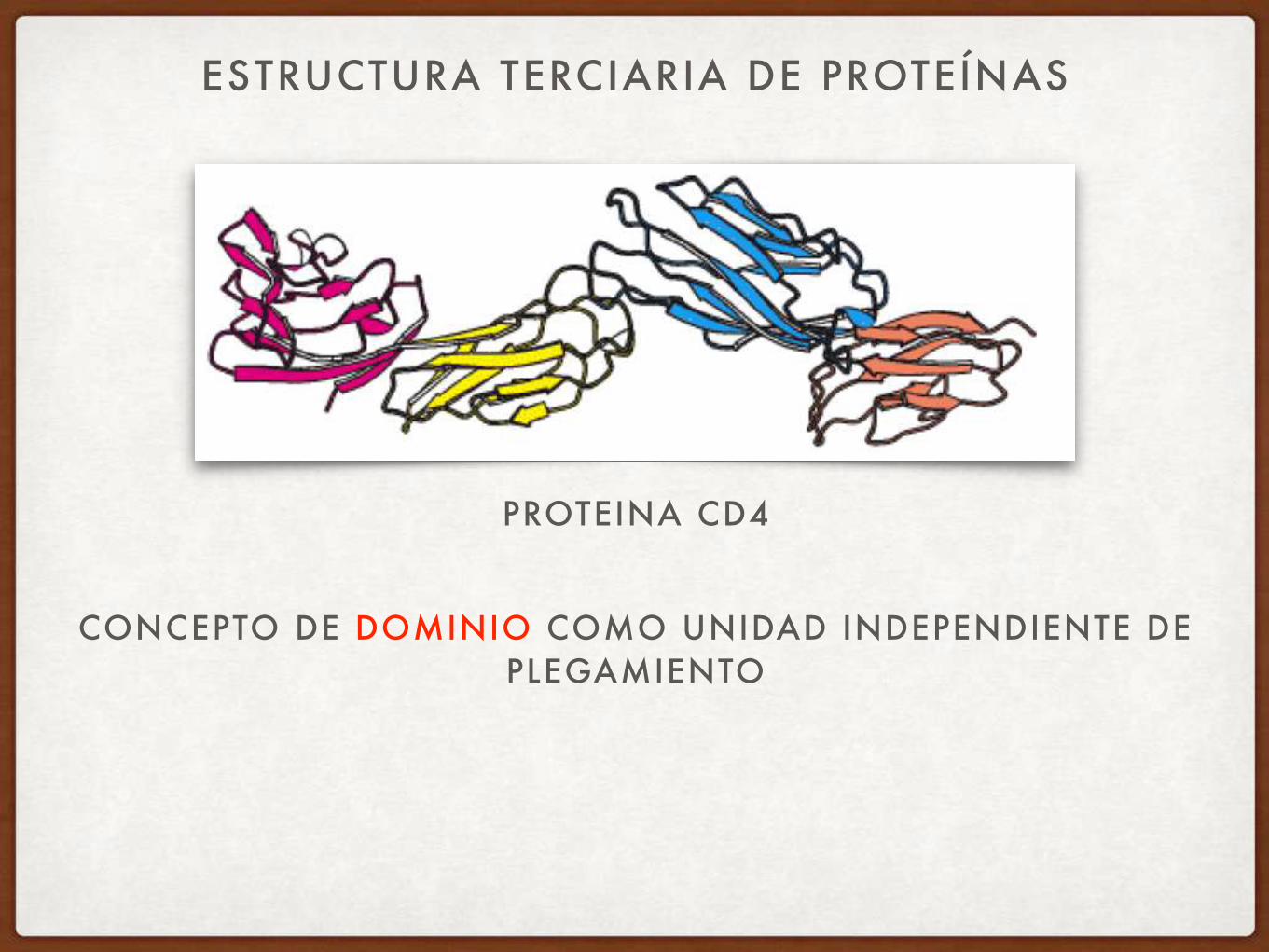
ESTRUCTURA TERCIARIA DE PROTEÍNAS
PROTEINA CD4
CONCEPTO DE DOMINIO COMO UNIDAD INDEPENDIENTE DE PLEGAMIENTO

FUERZAS MOLECULARES EN ESTRUCTURA DE PROTEÍNAS(EN SISTEMAS ACUOSOS!!)
• FUERZAS ELECTROSTÁTICAS
• FUERZAS DE VAN DER WAALS
• PUENTES DE HIDRÓGENO
• EL ENLACE O “EFECTO” HIDROFÓBICO
INTERACCIONES ENTRE ÁTOMOS NO ENLAZADOS (INTERACCIONES NO COVALENTES)

QUÉ NOS DICEN LAS LEYES DE LA TERMODINÁMICA?
Segunda ley de la termodinámica: Para un proceso espontáneo, la entropía total del sistema siempre aumenta
UN PROCESO ES ESPONTANEO SI EL CAMBIO EN ENERGÍA LIBRE DE GIBBS ES MENOR A CERO
ENERGÍA LIBRE DE GIBBS
prestar atención a las MAGNITUDES de las energías de unión

FUERZAS ELECTROSTÁTICAS
FUERZA DE “LARGO” ALCANCE PORQUE LA ENERGÍA DECAE LENTAMENTE CON R (COMO 1/R PARA GRUPOS CARGADOS)
FUERZA QUE SE ESTABLECE ENTRE GRUPOS CARGADOS O DIPOLARES, TAMBIÉN LLAMADA PUENTE SALINO O UNIÓN IÓNICA
Q1
+
Q2
_
E =q1*q2D*R
R = distancia
D = constante dieléctricaD (agua) = 80
D (proteína) = 2

ENERGIAS DE UNIÓN ELECTROSTÁTICA MEDIDAS EN PROTEÍNAS
Cada unión individual se encuentra en el orden de 0.2-0.5 kcal/mol

FUERZAS DE VAN DER WAALS
También llamadas fuerzas no polares o fuerzas de dispersión de LondonINTERACCIONES ESTABLECIDAS ENTRE DOS NUCLEOS ATOMICOS
Repulsión entre las nubes electrónicas de ambos
átomos a distancias cortasAtracción generada por los dipolos de ambos átomos,
que decae rápidamente con la distancia (como 1/R6)
EL PUNTO DE MÍNIMA ENERGÍA CORRESPONDE AL RADIO DE VAN DER WAALS
CORTO ALCANCE

FUERZAS DE VAN DER WAALS
Cada unión individual se encuentra en el orden de 0.1-0.3 kcal/mol pero la sumatoria de interacciones de todos los átomos puede volverse grande

PUENTES DE HIDRÓGENO
ÁTOMO ACEPTOR
ÁTOMO DONOR
3.13.04 2.932.72.63 2.88Distancia (Amstrong)
UN TIPO DE ENLACE MUY IMPORTANTE EN SISTEMAS BIOLÓGICOS
Enlace covalente
menor a Van der Waals
DISTANCIA
NO covalente entre dos átomos electronegativos
• Un hidrógeno es parcialmente donado a otro átomo más electronegativo • El ángulo preferido es 180º • Las energías típicas de cada enlace son de 3-9 kcal/mol
Alfa hélice y Hoja Beta

INTERACCIONES MOLECULARES EN UN AMBIENTE ACUOSO
Las energías medidas en solución son aquellas determinadas por las interacciones entre moléculas en competencia con el agua
Los reactivos se unen entre sí, desplazando al agua asociada a ellos. Esto está acompañado por cambios importantes en la entropía del solvente (AGUA)

Grupo en N-Octanol
ENERGIAS DE TRANSFERENCIA DE GRUPOS QUIMICOS DE UN AMBIENTE APOLAR (N-OCTANOL) A UN AMBIENTE POLAR (AGUA)
Grupos alifáticos prefieren estar en un solvente no polar
Grupo en Agua
△G > 0 desfavorable
△G < 0 favorable
Grupos polares prefieren estar en un solvente polar

Bases moleculares del “Efecto Hidrofóbico”Interacción ligando-enzima Coalescencia de lípidos
La interacción entre moléculas no polares “libera” moléculas de agua al solvente, aumentando la entropía total del sistema
Agua en el solventeLos puentes Hidrógeno entre moléculas se rompen y reforman en forma dinámica
Entropía de agua
Agua en torno a las moléculas hidrofóbicas
s e e n c u e n t r a n ordenadas en torno a la molécula apolar
Entropía de agua
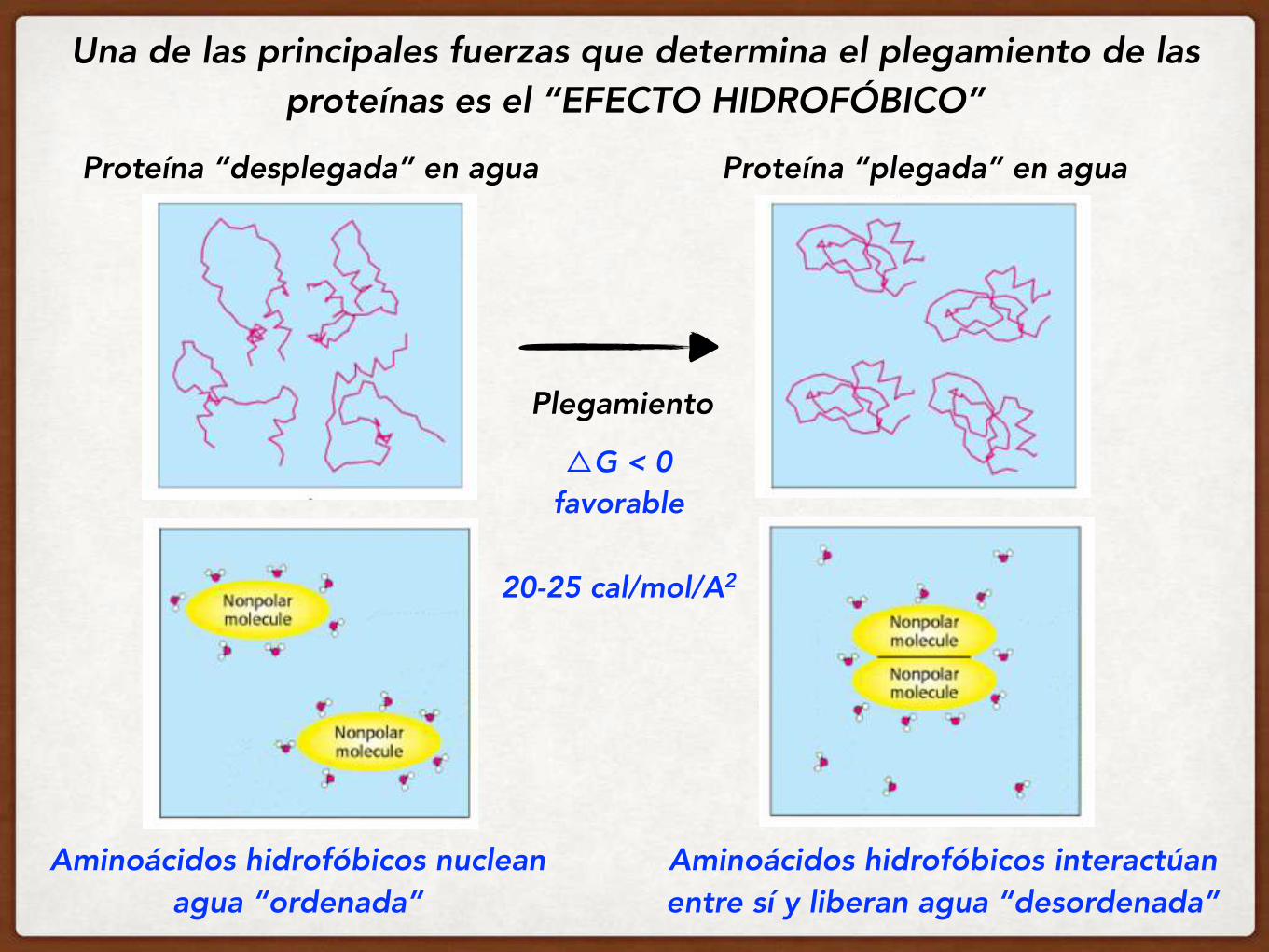
Una de las principales fuerzas que determina el plegamiento de las proteínas es el “EFECTO HIDROFÓBICO”
Aminoácidos hidrofóbicos nuclean agua “ordenada”
Proteína “desplegada” en agua
Plegamiento
Proteína “plegada” en agua
△G < 0 favorable
Aminoácidos hidrofóbicos interactúan entre sí y liberan agua “desordenada”
20-25 cal/mol/A2

LAS PROTEÍNAS PLEGADAS SE CARACTERIZAN
VISTA SUPERFICIAL CORTE TRANSVERSAL
MIOGLOBINA
POR UN CENTRO RICO EN AMINOÁCIDOS HIDROFÓBICOS
AA cargados AA hidrofóbicos AA polares

PROTEINAS
ESTRUCTURA CUATERNARIA DE PROTEÍNAS
COMBINACIONES DE SUBUNIDADES: UN DÍMERO PROTEINA CRO DE BACTERIOFAGO LAMBDA

PROTEINASESTRUCTURA CUATERNARIA DE PROTEÍNAS
COMBINACIONES DE SUBUNIDADES: LO MAS GRANDE?EL RIBOSOMA, EL PORO NUCLEAR
EL TETRÁMERO ALFA2/BETA2 DE HEMOGLOBINA

“AGENTES DESNATURALIZANTES”
CLORURO DE GUANIDINIO [Gdm:Cl] UREA
Son buenos solventes de las cadenas laterales y por ende estabilizan el estado desplegado de las proteínas
LA ESTRUCTURA NATIVA PUEDE SER DESNATURALIZADA POR AGENTES QUÍMICOS
Plegamiento (En agua)
Desplegamiento (En Gdm:Cl o Urea)
PROTEINA PLEGADAPROTEINA DESPLEGADA

DÓNDE ESTÁ CODIFICADA LA INFORMACIÓN PARA PLEGAR UNA PROTEÍNA?
EL EXPERIMENTO CENTRAL DE ANFINSEN, 1962
RNASA
Nativa, catalíticamente activa degrada RNA
4 Puentes Disulfuro
UREA MERCAPTOETANOL
RNASA
Desplegada, inactiva
Cisteínas reducidas

TODA LA INFORMACIÓN NECESARIA PARA PLEGAR ESTA PROTEÍNA ESTABA CODIFICADA
GENÉTICAMENTE EN SU SECUENCIA
RNASA
Desplegada, inactiva
Cisteínas reducidas
DIALISIS REMOVIO LENTAMENTE UREA y MERCAPTOETANOL
RNASA
Replegada Nativa Catalíticamente activa Puentes S-S correctos

ESTRUCTURA PLEGADA DE LA RIBONUCLEASA

SIN EMBARGO….LA RELACIÓN SECUENCIA -ESTRUCTURA, ES UNÍVOCA?
…VDLLKN…
🧐 NO !! La conformación que adquiere una secuencia depende a su vez del contexto estructural

EVOLUCIÓN MODULAR DE PROTEÍNAS
EL UNIVERSO DE DOMINIOS ES LIMITADO
CONCEPTO DE DOMINIO COMO UNIDAD EVOLUTIVA

EXISTE UNA CANTIDAD LIMITADA DE MOTIVOS ESTRUCTURALES
“BETA HAIRPIN”
FORMADOS POR ELEMENTOS PROXIMOS EN LA SECUENCIA PRIMARIA
“GREEK KEY”
VENENO DE SERPIENTE ERABUTOXIN
STAPHYLOCOCUS NUCLEASE

“HELIX -LOOP-HELIX”
EXISTE UNA CANTIDAD LIMITADA DE MOTIVOS ESTRUCTURALESMOTIVOS ESTRUCTURALES
PROTEINAS DE UNION A ADN FACTORES DE TRANSCRIPCION
PROTEINAS DE UNION A CA2+
CALMODULINA

LOS DOMINIOS SE FORMAN POR COMBINACION DE MOTIVOS
MOTIVO BETA -ALFA -BETA -ALFA EN LA TRIOSA FOSFATO ISOMERASA

LAS PROTEÍNAS SON MODULARESY SE FORMAN POR COMBINACIONES DE DOMINIOS
DOMINIO “EGF” 53 AA
DOMINIO “SERIN PROTEASA” 245 AA
DOMINIO “KRINGLE” 85 AA
DOMINIO “UNION A CA2+”

COMO SE GENERO LA DIVERSIDAD DE DOMINIOS?
SE ESTIMA QUE EXISTEN ~1000 TIPOS DE SUPERFAMILIAS DE DOMINIOS
LAS DIFERENTES PROTEINAS MODULARES PUDIERON HABERSE GENERADO POR EVENTOS DE DUPLICACION Y FUSION GENICA
EN EUCARIOTAS, LA APARICION DE INTRONES PUDO HABER AMPLIFICADO LAS COMBINATORIAS DE DOMINIOS

EVOLUCIÓN DE DOMINIOS DE UNIÓN A OXIGENO
Secuencias de Hemoglobina y Mioglobina humanas
Alineamiento23% IDENTIDAD
NARANJA: posiciones idénticas AMARILLO: sustituciones conservativas
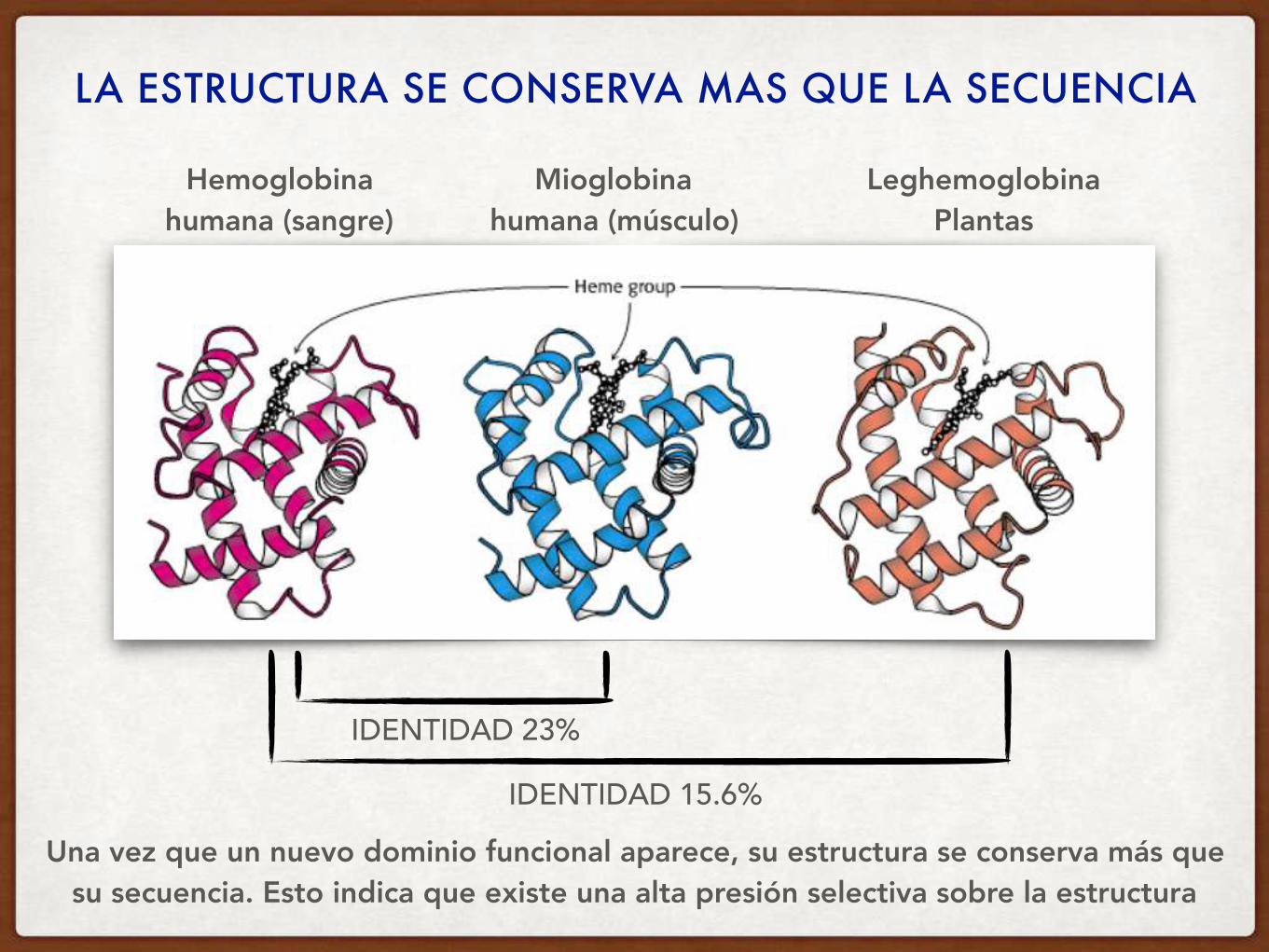
LA ESTRUCTURA SE CONSERVA MAS QUE LA SECUENCIA
IDENTIDAD 15.6%
IDENTIDAD 23%
Una vez que un nuevo dominio funcional aparece, su estructura se conserva más que su secuencia. Esto indica que existe una alta presión selectiva sobre la estructura
Hemoglobina humana (sangre)
Mioglobina humana (músculo)
Leghemoglobina Plantas
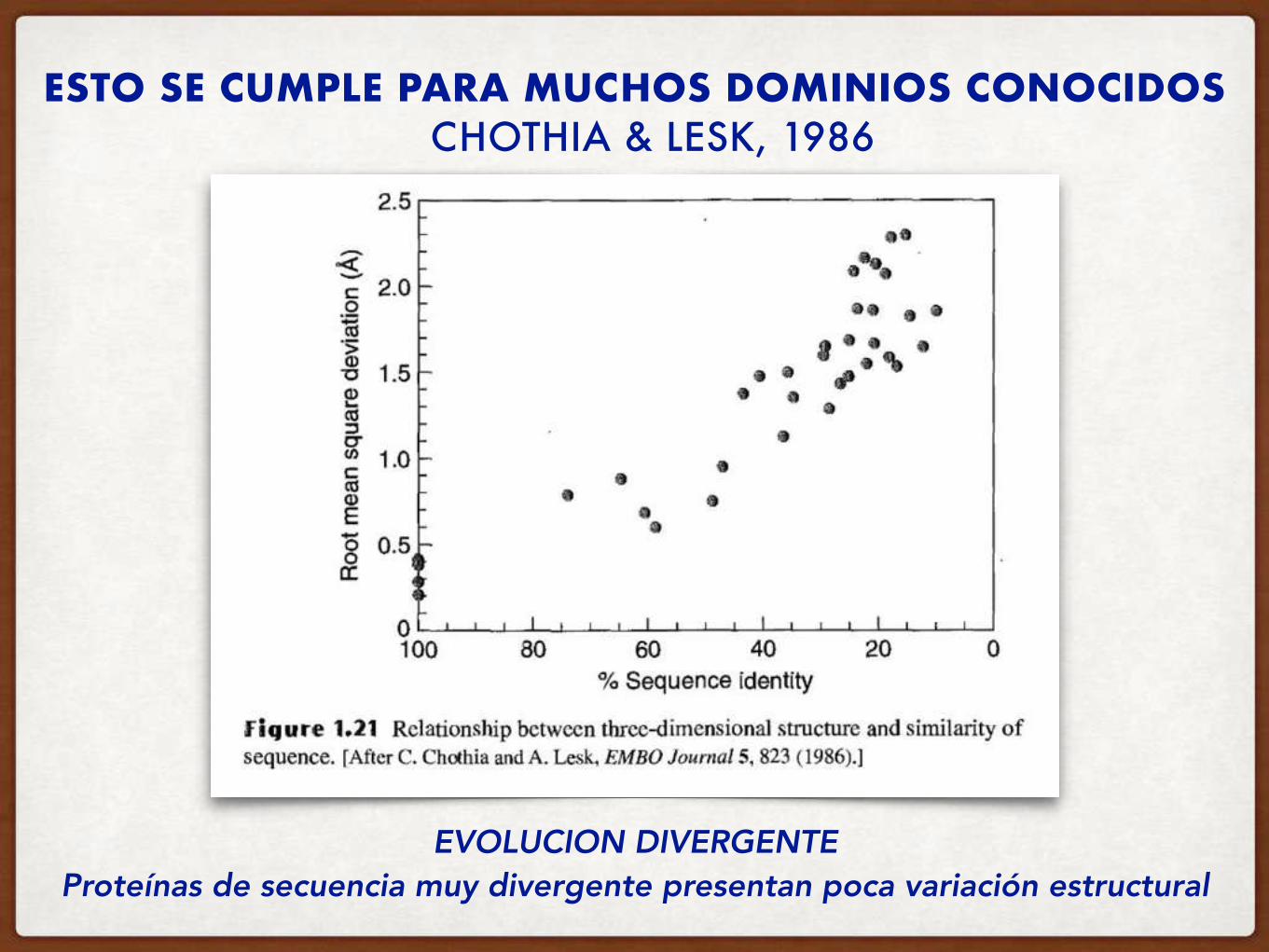
ESTO SE CUMPLE PARA MUCHOS DOMINIOS CONOCIDOS
Proteínas de secuencia muy divergente presentan poca variación estructural
CHOTHIA & LESK, 1986
EVOLUCION DIVERGENTE
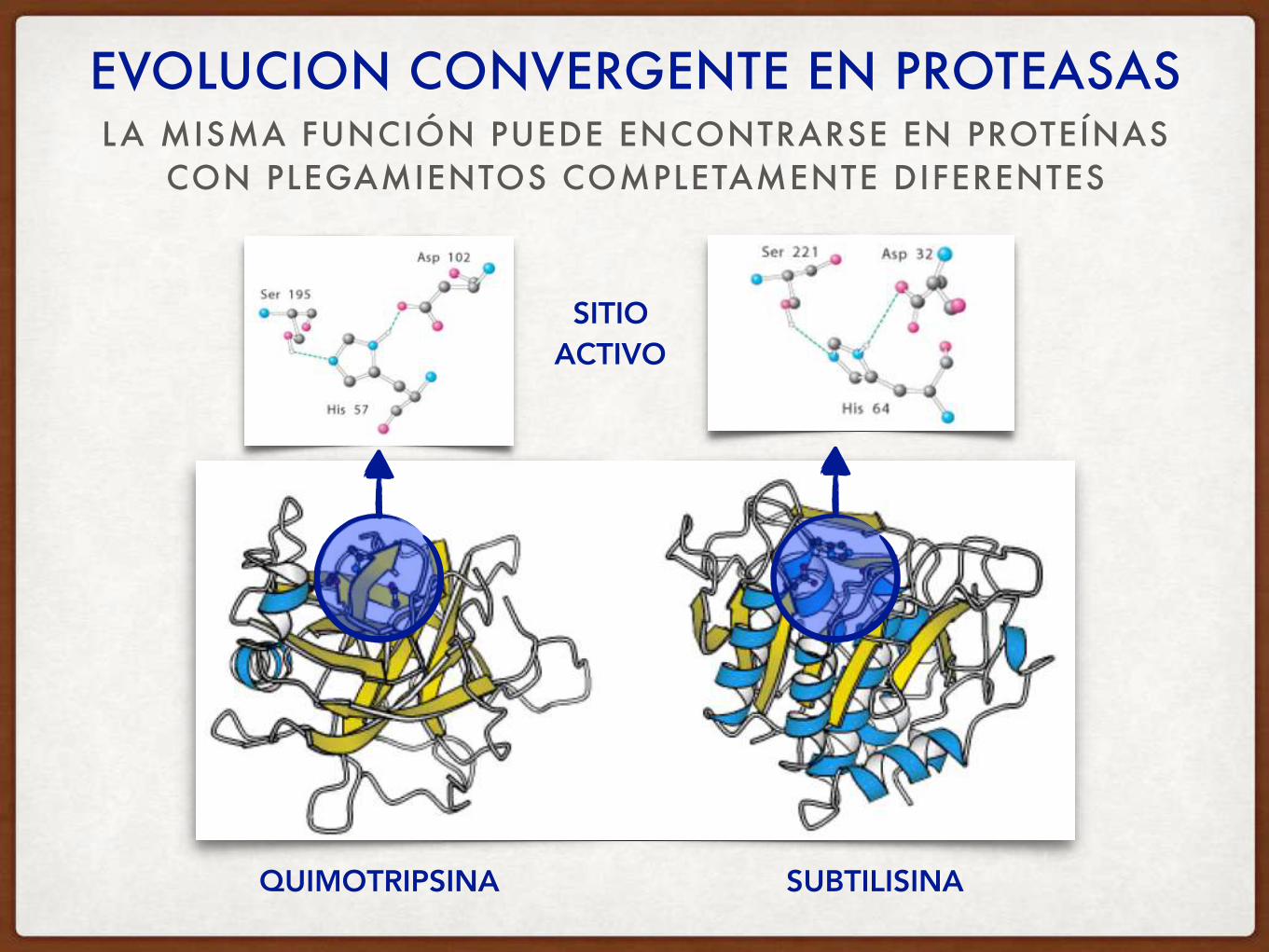
LA MISMA FUNCIÓN PUEDE ENCONTRARSE EN PROTEÍNAS CON PLEGAMIENTOS COMPLETAMENTE DIFERENTES
EVOLUCION CONVERGENTE EN PROTEASAS
SITIO ACTIVO
QUIMOTRIPSINA SUBTILISINA

TODAS LAS REGIONES DE LAS PROTEÍNAS SE PLIEGAN PARA FORMAR DOMINIOS?
NO!!
PROTEINAS “INTRINSECAMENTE DESORDENADAS” (IDPS) O “NATIVAMENTE DESPLEGADAS”
p53 tumor suppressor
Wells et al. PNAS 2008; 105: 5762
TAD DBD TD RD
disordered ordered disordered
NO ADQUIEREN ESTRUCTURA TERCIARIA ESTABLE
~30% DEL PROTEOMA EN EUCARIOTAS
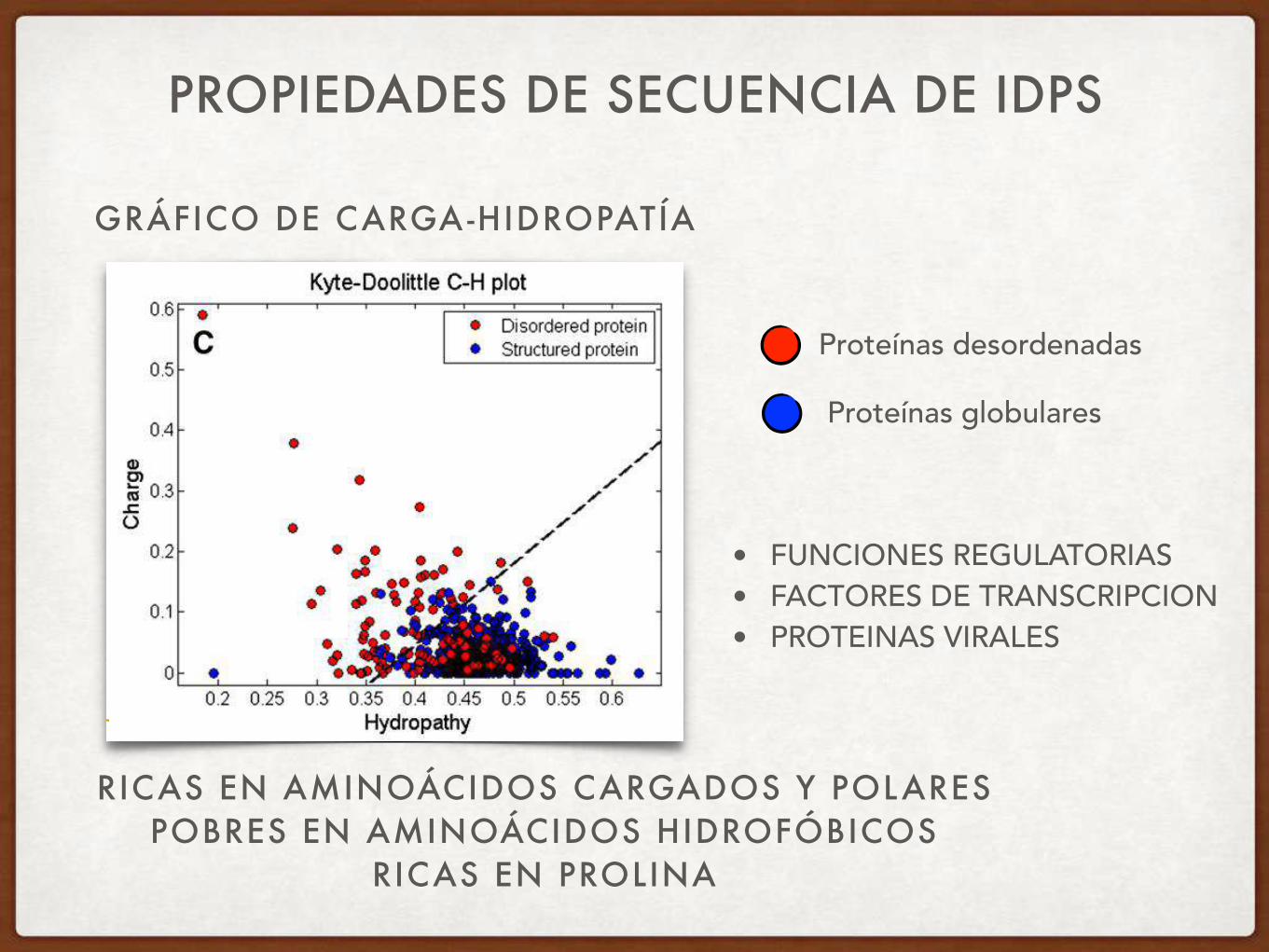
PROPIEDADES DE SECUENCIA DE IDPS
RICAS EN AMINOÁCIDOS CARGADOS Y POLARES POBRES EN AMINOÁCIDOS HIDROFÓBICOS
RICAS EN PROLINA
Charge-hydropathy plot GRÁFICO DE CARGA-HIDROPATÍA
Proteínas desordenadas
Proteínas globulares
• FUNCIONES REGULATORIAS • FACTORES DE TRANSCRIPCION • PROTEINAS VIRALES
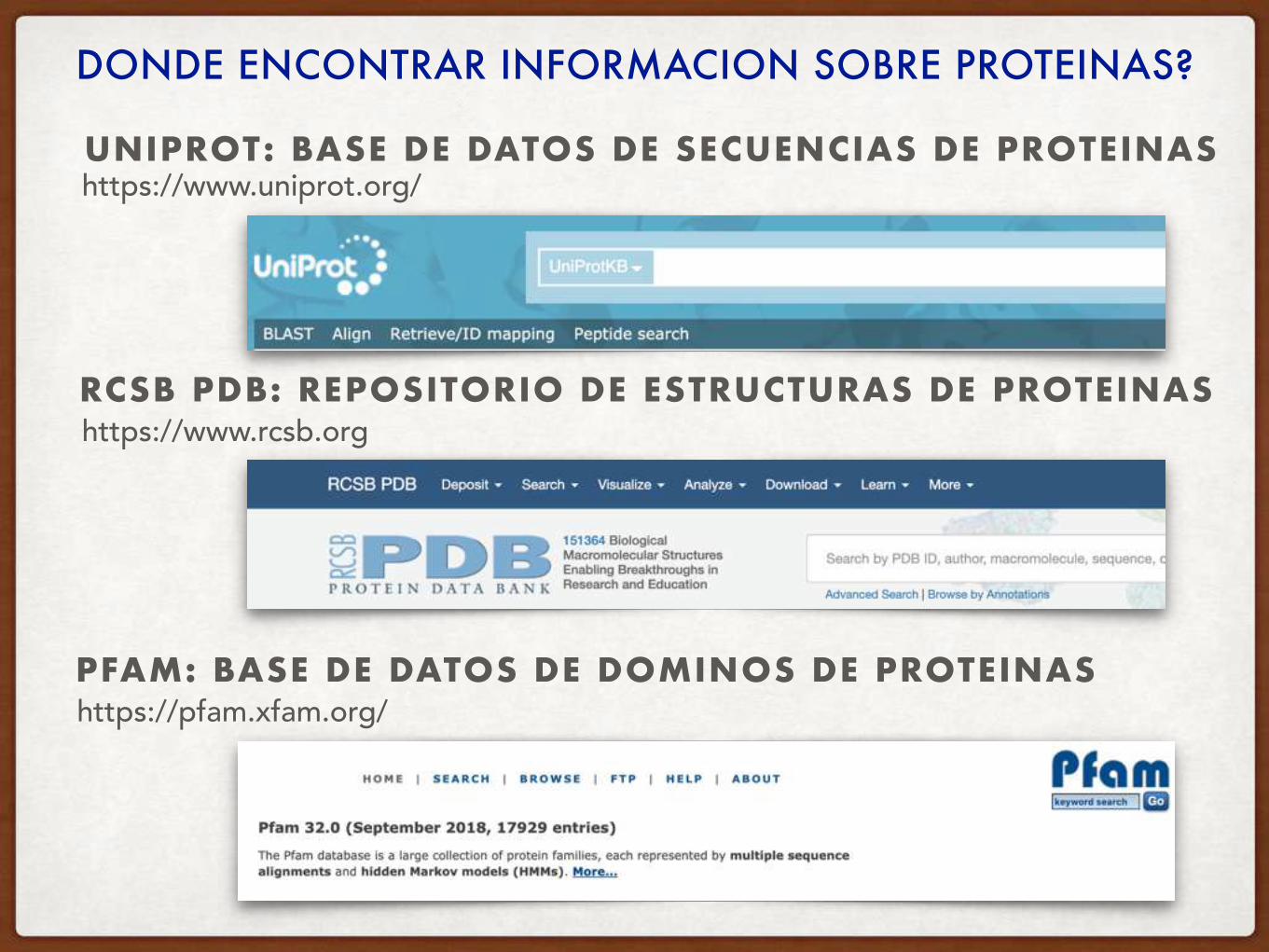
DONDE ENCONTRAR INFORMACION SOBRE PROTEINAS?
RCSB PDB: REPOSITORIO DE ESTRUCTURAS DE PROTEINAS
UNIPROT: BASE DE DATOS DE SECUENCIAS DE PROTEINAS
PFAM: BASE DE DATOS DE DOMINOS DE PROTEINAS
https://www.uniprot.org/
https://www.rcsb.org
https://pfam.xfam.org/

PLEGAMIENTO DE LAS PROTEINASLA PARADOJA DE LEVINTHAL
CUANTAS COMBINATORIAS DEBE EXPLORAR UNA SECUENCIA PARA PODER PLEGARSE? CUANTO TIEMPO LLEVARÍA?
Proteína “desplegada”
Plegamiento
Proteína “plegada”

PLEGAMIENTO DE LAS PROTEINASLA PARADOJA DE LEVINTHAL
Para una cadena de 100 aminoácidos…. Si hay sólo 3 conformaciones para cada aminoácidoCombinaciones azarosas que deben ser exploradas 3100
Si cada transición lleva 10-13 seg (muy poco!)
5 x 1047~
5 x 1034seg 1.6 x 1027 años!
CUANTAS COMBINATORIAS DEBE EXPLORAR UNA SECUENCIA PARA PODER PLEGARSE?
CUANTO TIEMPO LLEVA HACERLO?
EL PLEGAMIENTO DE LAS PROTEÍNAS NO OCURRE POR UN PROCESO DE EXPLORACIÓN AZAROSA DEL ESPACIO DE CONFORMACIONES !!!!!!
🤔
#ni corriendo se llega…no es compatible con la vida!
ENTONCES: CÓMO OCURRE?

LA ANALOGIA DEL MONO DE SHAKESPEARE
Hacen falta 1040 combinaciones!!!! para que un mono tipee el siguiente texto de Shakespeare al azar
Peeero….si se retienen las posiciones correctas
LA ESENCIA DEL PLEGAMIENTO DE PROTEINAS ES LA RETENCION DE INTERMEDIARIOS CORRECTOS DE PLEGADO
Sólo hacen falta ~3000 tipeosal azar = 2326 COMBINACIONES
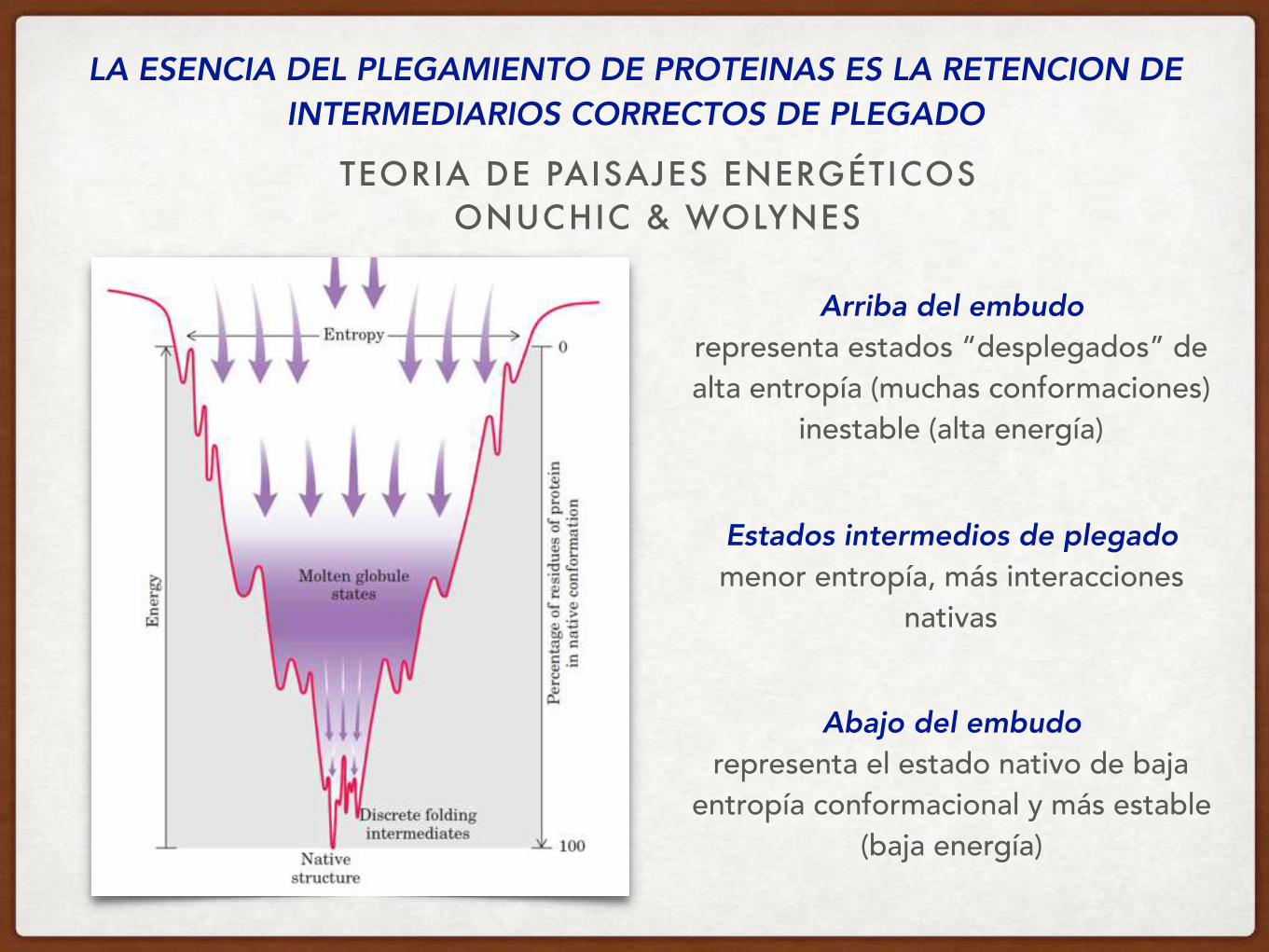
TEORIA DE PAISAJES ENERGÉTICOS ONUCHIC & WOLYNES
LA ESENCIA DEL PLEGAMIENTO DE PROTEINAS ES LA RETENCION DE INTERMEDIARIOS CORRECTOS DE PLEGADO
Arriba del embudo representa estados “desplegados” de alta entropía (muchas conformaciones)
inestable (alta energía)
Abajo del embudo representa el estado nativo de baja
entropía conformacional y más estable (baja energía)
Estados intermedios de plegado menor entropía, más interacciones
nativas