biology, relationships, and control of aedes albopictus … pajrio library cataloguing in...
TRANSCRIPT

165143165143



Bcf’gNSR PEST MGMT INFOi-^AL’i’Sia CTR, AFPMB, FORESTfeliyi SKeTION, WRAMC,WASH,, B.& 20307-5001
BIOLOGY, DISEASE RELATIONSHIPS,AND CONTROL OF
Aedes albopictus
Jose G. Estrada-Franco, Ph.D.Department of EntomologyUniversity of Maryland
College Park,’Maryland, U.S.A.
and
George B. Craig, Jr., Ph.D.Vector Biology Laboratory
Department of Biological Sciences
University of Notre DameNotre Dame, Indiana, U.S.A.
Technical Paper No. 42
PAN AMERICAN HEALTH ORGANIZATIONPan American Sanitary Bureau, Regional Office of the
WORLD HEALTH ORGANIZATION525 Twenty-third Street, N.W.
Washington, D.C. 20037

PAJrIO Library Cataloguing in Publication Data
Estrada-Franco, Jos6 G.Biology, disease relationships, and control olAcdes albopictus Jose G. Estrada-Franco, Ph.D., and
George B. Craig, Jr., Ph.D. Washington, D.C. PAITO, (C)1995v, 49 p. (Technical Paper 42)
ISBN 92 75 13042
I. Title IT. (Series) Til. Craig, George B.1. AEDES 2. ARBOVIRUS INFECTIONS3. MOSQUITO CONTROL-methodsNI-M 0X525
ISBN 92 75 13042
The Pan American Health Organization welcomes requests for permission reproduce translate itspublications, in part in full. Applicalions and inquiries should be addressed the Publications Program,Pan American Health Organization, Washington, D.C., which will be glad provide the latest information
any changes made to the text, plans for editions, and reprints and translations already available.
(C) Pan American Health Organization 1995
Publications of the Pan American Health Organisation enjoy copyright protection in accordance withthe provisions of Protocol of the Universal Copyright Convention. All rights reserved.The designations employed and the presentation of the material in this publication do impiv (he
expression of any opinion whatsoever the pan of the Secretariat of the Pan American Health Organ-ization concerning the legal of any country, territory, city, of its authorities, concerningthe delimitation of its frontiers boundaries.The mention of spccilic companies of certain manufacturers’ products does imply that they
endorsed recommended by the Pan American Health Organization in preference others of similarthat mentioned. Errors and omissions exccpicd, the of proprietary products
distinguished by initial capital tellers.

CONTENTS
PagePREFACE
INTRODUCTION
PIIYLOGENETIC RELATIONSHIPS AND GEOGRAPHICALDISTRIBUTION
BIOLOGY 2
Eggs............................................................................................................ 5Egg Diapausc, Photoperiod, and Adaptations 6Egg Cold Hardiness 7Egg Morphology 8
Larvae 8Larval Seasonal Prevalence 10Larval Morphology 10Larval Habitat.......................................................................................... 10Larval Pathogens and Predators 16
Pupae 18
Adults 18
Longevity................................................................................................. 18Mating Characteristics 19Oviposition 20Host Preference 22Biting Behavior........................................................................................ 22Plight Range 23
Resting Behavior...................................................................................... 23Adult Morphology 23
Competition Studies 23
DISEASE RELATIONSHIPS 27
General Information Arboviral Diseases 28
Arbovirus Field Isolations in the Americas 28Eastern Equine Encephalitis (EEE) 28Potosi (POT)............................................................................................ 30Tcnsaw (TEN) and Keystone (KEY)......................................................... 30
Laboratory Vector Competence Studies of Some ImportantArboviruses from the Americas 31Dengue (DEN)......................................................................................... 31

Yellow Fever (YF) 31Venezuelan Equine Encephalitis (VEE)..................................................... 32La Crosse (LAC) 32Mayaro (MAY) and Oropouche (ORO)..................................................... 33
Arboviruses Common in Areas Outside the Amcricas 34Japanese Encephalitis (JE)........................................................................ 34Sindbis (SIN)........................................................................................... 34
Chikungunya (CHIK) 34Ross River (RR) 34
Transovarial Transmission 35
Other Arboviral Studies 35
Protozoan and Filarial Diseases 35Filarial Worms 35Avian Malarias 35
DISSEMINATION AND CONTROL 35
Mode of Dissemination and Origin............................................................... 35
Control 37Chemical Control 37Biological Control.................................................................................... 38Environmental Control 39Health Education...................................................................................... 39Legal Measures 39Integrated Control 40
CONCLUSIONS 40
ACKNOWLEDGMENTS 41
REFERENCES................................................................................................ 41

PREFACE
The first version of this document, titled "Ecology, Biology, and Control ofAedes albopictus (Skuse)," was prepared in January 1987 to provide scientific andtechnical background for the "Plan of Action onArdes albopictus in the Ameri-
cas." It discussed at the 99th Meeting of the Executive Committee of the PanAmerican Health Organization (PAHO), held in Washington, D.C., in June 1987
(PAHO Document CE99/15, 1987). The document produced by the Communic-
able Diseases Program under the coordination of Dr. F. J. Lopez-Antunano and
joint effort of Dr. Jose G. Estrada-Franco, PAHO Temporary Adviser; Dr.George B. Craig, Biology Department, University of Notre Dame; and Drs. Fran-cisco Pinhciro, Bruce Knudsen, and Michael Nelson of PAHO/WHO. Since the
1987 version produced, three excellent reviews the biology, genetics, and
disease relationships of Aedes albopictus have appeared, in addition to plethora of
scientific articles the species (Hawley, 1988; Rai, 1991; Mitchell, 1991).This second version is presented in order to include important information about
this mosquito species that has been brought to light during the past five years byseries of studies and research projects conducted primarily in the United States ofAmerica. The findings of these studies provide insight into what could be-
of the important arthropod vectors of human diseases and in theAmerican Region. This document is published in the hope that it will be of value to
health agencies in the Americas that concerned with vector surveillance and
control.

INTRODUCTION
Until the early 1980s Aedes alhopicius assumed to
solely inhabit certain islands of the Indian Ocean, variouscountries in the oriental region of Asia, and the HawaiianIslands in the Pacific Ocean (Huang, 1972). In the mid-
1980s, however, this picture underwent sudden and
alarming shift.
In August 1985 Ae. alhopictus mosquitoes discov-ered breeding in lire dumps in Harris County, in and around
Houston, Texas. The Houston Ae. (ilbopictus populationrepresented the first established infestation reported in theWestern Hemisphere (Centers for Disease Control and Pre-vention [CDC], 1986a; Monath, 1986; Sprengcr and
Wuithiranyagool, 1986). To date, Ac. albopictus has been
reported from 22 other states in the United States of Americait has moved northward, the northernmost point thus far
being Chicago, located at about 42 latitude (D. Gubler, pcrs.comm., 1993). The southward movement of Ae. alhopictusin the United States has reached the city of Brownsville,Texas, located at 2554.3’ latitude (Womack, 1993).
Elsewhere in the Americas, Ac. albopictu’, first
ported in Brazil in June 1986 in the states of Espirito Santo.Minas Gerais, Rio de Janeiro, and Sao Paulo (Forattini,1986; Parigot, 1986). By April 1994 the species had been
reported in three stales-Maranhao (San Luis de Mar-
anhao) and Bahia and Parana (several locations)-for to-
tal of 673 infested municipalities (J. C. Mangabeira daSuva, pcrs. comm., 1994). The extent of Brazil’s Ae. al-
bopiclns infestation prompted panel of experts, meetingat PAHO Headquarters in 1986, to hypothesize that the spe-cies first introduced into the country years ago(PAHO, 1987; CDC, )986b).
Furthermore, the third and fourth countries with estab-
lished infestations have recently been identified: the Do-minican Republic, where the mosquitoes have been found
mainly in parkland of the capital city of SantoDomingo (C. Pena, National Malaria Program, pers. comm.,June 1993), and Mexico, with infestations in towns along theU.S.-Mexico border in the slates of Coahuila and Tamauli-
pas (C. Castancda-Barron, Tamaulipas State Health Services,pers. comm., 1992; C. Moore, pers. comm., 1993). The spe-cies has also been reported in continental Africa and Europe(Savage et al., 1992; Sabatani et al., 1990).The potential threat posed by the presence in the Ajner-
icas of this exotic and efficient vector species of dengueand possibly yellow fever and other arboviruscs has gen-erated great among the countries (PAHO, 1987).
The intent of this document is to bring together the in-
formation available to date the biology, disease relation-
ships, and control ofAc. albopicliis. The Asian information,summarized in 1987 document, is supplemented by thefindings of studies conducted mainly in the United States,with emphasis data generated in the 1990s.
PHYLOGENETIC RELATIONSHIPS ANDGEOGRAPHICAL DISTRIBUTION
Aedes albopicliis first described by Skuse in 1894from specimens collected in the city of Calcutta the In-dian subcontinent (Barraud, 1928). Phylogcnetically, the
species has been placed in the order Diptcra, suborder Ne-matocera, family Culicidae, genus Ac’dc’s, subgenus Stego-myia. group sculcllaris, and subgroup albopictus (Iluang,1972; Rai ct al., 1982).
It is believed that Ac. ulboplctiis originated in the tropicalforest of Southeast Asia. where many closely related spe-cies known to coexist (Smith, 1956). This is also
considered to be the place where classical dengue had its
origin. Another medically important Stegomyia, Aedesgypli, is thought to have been introduced into Asia fromAfrica (Mattingly, 1957). These two species the most
important vectors of dengue in Southeast Asia (Smith.1956; Pant et al., 1973; Chan, 1985). In Asia, Ac. ulbo-
piclus far north Bcijing, China, at 40 latitude
(World Health Organization fWHOJ, 1980). It is also dis-tributed in the Korean peninsula and in Japan to northern
Honshu, Japan’s main island, reaching the city of Sendai
at 36 N (Kamimura, 1968; Hong ct al., 1971). Beijing and
Chicago, Illinois (at about 42 north latitude in the UnitedStates), the northernmost reported distribution points ofAc. albopictus in the Palcarctic and Nearclic regions,respectively.
In Southeast Asia the species has extended its range to
the coastal cities of Irian Jaya, the Solomon and Santa CruzIslands, and Papua New Guinea; it has also been found in
Brisbane and Darwin, Australia, where the infestations
promptly eradicated (Elliot, 1980; D. Gubler, pers.comm., 1993), and recently in New Zealand (Kay ct al.,1990; Craig, 1993). It has been reported in the Mariana,Caroline, and Hawaiian Islands, Palau, the Asian continent,and westward to several islands in the Indian Ocean such
Madagascar, Mauritius, Reunion, the Seychelles, and the
Chagos Archipelago (Surtees, 1966; I-ambrccht and VanSomcren, 1971; Ho el a)., 1972). In addition, the species

has recently been found in Europe (Albania and Italy) andin mainland Africa (coastal South Africa and various vil-
lages in Nigeria) (Sabatani et al., 1990; Savage et al., 1992;Craig, 1993; references in Hanson and Craig, 1994).Also. noted above, Aedes albopicliis has recently colo-nized portions of four countries of the Americas. It has beenfound in midwestern and southern states in the United Stales.three towns in northern Mexico along the border with Texas,
states in Braxil, and parts of the city of Santo Domingoin the Dominican Republic (Moore et al., 1988; Mangabeirada SUva, pers. comm., 1994; Moore, pers. comm., 1994).The species is known to all continents exceptAustralia and Antarctica. Map summarizes the worldwidedistribution of Ac. albopicliis. Maps 2 and illustrate its
distribution in three countries of the Americas.
In terms of altitude, Aedes alhopictus has been found upto 1,800 meters in the mountains of Thailand (Scankm and
Esah. 1965). Pant and co-workers (1973) reported the pres-of Ac. alhopiclus in Thai village located at 1,700
meters. Both studies noted finding the species at every al-
titude surveyed, from level to the highest. Ho el al.
(1972) reported maximum altitudes forAc’, albopictus of 24to ISO meters in West Pakistan, Sri Lanka, Taiwan, and
Malaya, but this study appears to underestimate thequito’s vertical range. In the United States the highest al-titude reported for the species is 305 meters at New Alsace,Indiana (Focks ct al., 1994).One of the most important differences between the North
American populations of Ae. albopiciiis and Ac. aegypti is
their latitudinal distribution. While Ae. aegypti populationslimited to the southern portions of the United States due
mainly to their inability to tolerate very low temperatures,Ae. alhopictus populations have evolved photoperiod-induccd egg diapause, which allows them to colonize tem-
perate and northern latitudes. In addition, temperate strainsof Ae. albopiclus, such the North American ones, show
egg cold hardiness, which enables the species to survivethe suboptimal winter temperatures found in the northernlatitudes (Hawley, 1988).While the distribution of Aedes albopictus and that of
Aedes aegypti overlap in the urban environment
(Harrison ct al., 1972; Azzizar, 1980; Chan, 1985), Ac. al-
hopictus is commonly found in suburban and ruralwhere open, vegetated spaces prevalent (Ho et al.,
1972; Pant et al., 1973; Chan, 1985). It is generallycepted that Ae. albopictus originally sylvan species,breeding and feeding in forest fringes, which began to adaptto the domestic environment throughout several parts of its
geographical range (Bonnet and Worchester, 1946; LaCasse and Yamaguti, 1950; Surtccs, 1970; Miyagi et a!.,1983). In the United States, studies based ovitrap col-lections carried out in New Orleans, Louisiana, in 1987showed that Ae. albopictus the most abundant speciesin suburban environments. These habitats characterized
by open with plentiful vegetation, surrounded by
buildings. By contrast, in urbanized sections of the
city, Ae. aegypti appears to remain the dominant species(New Orleans Mosquito Control Board [NOMCBj/ 1987).The tendency ofAc. albopiclus to inhabit natural containers
(tree holes and bromcliads) in places where vegetation is
abundant has also been observed in several other ofthe United States and Brazil (O’Mcura et al., 1992; Gomeset al., 1992). As in Asia, American Ae. albopiclus have
shown much greater propensity for using natural contain-than have Ae, aegypli.
BIOLOGY
In the United States, the duration of the larva-to-adult
phase of the life cycle of Ae. albopiclus in tires esti-
mated by comparing the population density of early instar
larvae with that of emerging aduits. Studies conducted dur-
ing the spring of 1989 in New Orleans (average ambient
temperature of about 26 C) with field populations of Ae.albopictus established that the average time period between
peak abundance of first instar larvae and peak abun-
dance of emerging adults about 10 days (NOMCB,1989). Under natural conditions in Viet Nam, eggs ot’A<?.albopiclus have been reported to reach adulthood in 7-20days in spring and 24 days in winter (Surtees, 1966; Ho et
al., 1972). Under laboratory conditions, studies thatsessed the effect of environmental parameters such tem-
perature and other variables produced similar results,ported by different authors.Matsuzawa and Kitahara (1966) found that for Ae. al-
bopictus the interval from the moment of oviposition to
emergence of the adult stage lasted 12, 13.7, and 24.3 daysat water temperatures of 30 C, 25 C, and 20 C, respec-tively. Hien (1975a) reported development times from ovi-
position to emergence of 11 days at 30 C, 14 days al
25 C, and 23 days at 20 C. Galliard and Golvan (1957),working with laboratory-reared strain of Ae. albopictus,found that development from egg to adult took 25 days and
10 days at 18 C and 25 C, respectively. Livingstonc and
Krishnamoorthy (1982) determined that at tempera-ture (26 C to 30 C) development from egg to adult offemale Ae, albopictus ranged from 11 to 12.4 days, whereasmales reached the adult stage in 10 to 11.4 days. The op-timal water temperature for Ac’, alhopictus development hasbeen reported to be 25 C. At this temperature the egg in-cubation period is two days (Sheng and Wu, 1953; Ho et
al., 1972).In addition to temperature, food supply also affects the
size and development time of the Ae. albopiclus larvae.Larvae placed in contaminated water with high organictent found to develop rapidly, while starvation and
’Address: 6601 Lakcshore Drive, New Orleans, LA7012A.

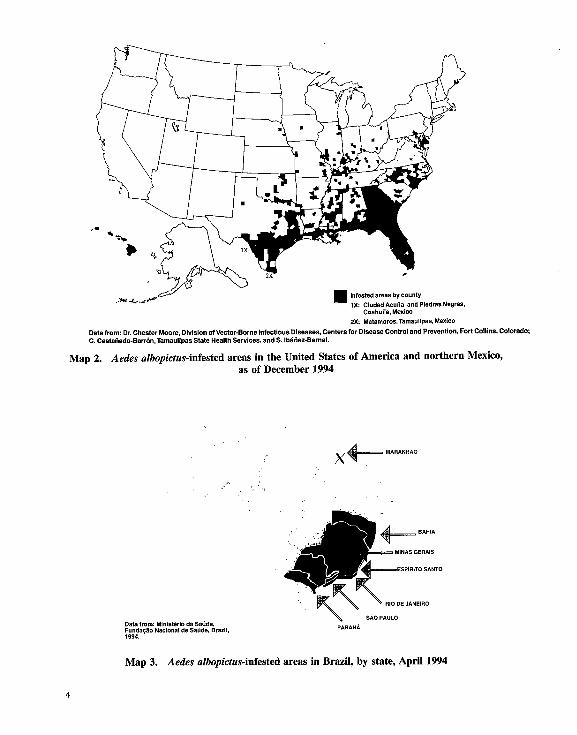
<;?-’ ^H by county
andPledrasNegras,Coatiuila,
Matamoros.Tamaulipas,
Data from: Dr. Chester Moore, Division ofVector-Borne Infectious Diseases, Centers for Disease Control and Prevention, Fort Collins, Colorado;
C. Castanedo-Barron, Tamaulipas State Health Services, and S. Ibanez-Bernal.
Map 2. Aedes alhopicfus-infestcd areas in the United States of America and northern Mexico,as of December 1994
Saude,Saude, Brazil,
-ESPl’RITO
Map 3. Aedes albopictus-’mfested areas in Brazil, by state, April 1994

overcrowding retarded development and increased mortal-ity under laboratory and field conditions (Mod, 1979). Ingeneral, the duration of the larval stage extended bydecreasing the food supply and shortened by increasing it.
At 0.035 mg tbod/larva/ml, the development timefrom egg hatching to adult 6.1 days. while at 0.004rog/larva/ml, the duration 32.4 days (Chan, 1971).
Variables known to affect the life cycle of any mosquitospecies include food, photoperiod, relative humidity, andtemperature. Significant information how these factorsaffect each developmental stage is summarized below.
Eggs
Mosquito eggs (including those of Aedes spp.) not
fertilized until the moment before they actually laid.
Sperm stored in the spermatheca enters each fully developedegg (oocyte) it passes the opening of the spermathecalduct its way out. Once the eggs have been laid, kary-ogamy and development of the embryos proceeds.Embryonic development may be defined the develop-mental changes taking place in the egg between fertilizationand hatching (Christophers, 1960). The length of the periodof embryonic development depends mainly the temper-ature and relative humidity to which the eggs exposed.
Laboratory observations have shown that Asian strainsof Ae. albopiclus eggs embryonated in periods of two to
four days at tempenitures between 24 C and 27 C (ref-in Hawlcy, 1988). Ideal conditions for embryogen-
esis of Ac. albopictus eggs from the Americas appear to beexposure under natural and laboratory conditions to tem-
perature of 21 C, relative humidity of 70%-80%, andphotoperiod of 16L:8D hours for six to days(NOMCB, 1989; Edgerly el al., 1993; Hanson and Craig,1994). Once embryogenesis has been completed, the eggs
stay unsubmerged and almost completely dry for longperiods without loss of viability.Egg survival may depend combination of density-
dependent and density-independent selective pressures.Events that may promote egg loss include desiccation, pre-dation, and freezing. Egg survival also may be affected bycertain female parental factors such unembryonated eggsthat collapse eggs that fall off because they not prop-erly attached to substrate. We will focus such factors
desiccation, predation, and freezing, since to the best ofknowledge studies have dealt with the female pa-
rental effects Ae. albopiclus egg survival.Ac. albopictus eggs survive desiccation in wide variety
of indoor and outdoor artificial containers (flower pots,earthenware jars, tin cans, coconut shells, tree holes, bam-
boo stumps, etc.) and hatch the containers fillup with water (Fcng, 1937; Ho et al., 1972). The numberof Ae. albopictus eggs that survive low humidity appearsto depend upon the development stage of the embryos be-
fore they exposed to dry conditions. Gubler (i970a)found that Ac. allwpictus eggs highly resistant to dryconditions if they kept in humid conditions for fourdays before being exposed to drought. The studycorded maximum egg longevity of 243 days at 25 C and
relative humidity of 70%-75%. In another laboratoryperiment Hien (1975b) showed that eggs kept wet cottonwool for four days and then exposed to 25-26C air at 60%-70% relative humidity highly resistant to dryness. Aftertwo months under these conditions of exposure, 94.7% ofthe eggs with conditioned embryos produced larvae. Maxi-
mal resistance to desiccation has been observed when eggsexposed to humidity for at least 24 hours, whereas
partial total mortality has been recorded when eggs lessthan 16 hours old dried (references in Hawley. 1988).A density-dependent factor that appears to play im-
portant role in the survival of Ae. alhopictus eggs is pre-dation. Gubler (1971a) found that the egg mortality ofnatural population of Ae. alhopictus from Calcutta, India,
greater than 50% during the first 24 hours, primarilyresult of predation by ants. A study conducted in New
Orleans, Louisiana, in 1989 examined the survivorship ofAe. albopictus eggs in tires (NOMCB. 1989). Eggs forthese experiments obtained from field sources, keptunder insectary conditions for five to days to favorembryonation and viability, and then returned to the
study Ovislrips placed in "dry" conditions
(tires with holes to prevent flooding) and "wet" conditions
(tires filled up with water). The study found that survivor-
ship of the field-collected eggs under both sets of conditionsbetween 10% and 30% after 24 hours. Thus, the rate
of egg loss between 70% and 90% and quite similarin both "wet" and ^dry" conditions. It presumed thategg loss caused mainly by predation by other arthro-pods, such arachnids, psocids, orlhopterans, isopods, andothers. The initial loss of eggs occurred when thenumber of intact eggs (to differentiate from collapseddamaged eggs) per ovistrip approximately 50 and the
eggs in clumps. The rate of egg loss became negli-gible when the number of intact eggs diminished to
about per ovistrip. Thus, the data indicated that egg lossis related to egg density. Egg survival in Ac. alhopk’lusappears to depend small numbers of eggs that avoid
predation by being widely spaced.Like all Aedes of the suhgenus Sffgomyiu. Ae. albopictus
females lay their eggs singly and spread them around the
container at varying distances from the water surface. Eachegg is attached at the water’s edge in the meniscus inthe wet just above the surface. Amerasinghe and AI-agoda (1984) observed that female Ae. alhopictus followset pattern when they oviposit bamboo traps. About 75%of their eggs deposited within 16 of the waterface, vertically and obliquely, with wide spacing left be-tween singly laid eggs- Oviposition extends to 53above the water level.

Eggs of Ae. albopicius hatch when they exposed to
specific stimulus. After embryonic development has been
completed, hatching within minutes of flooding,and larval growth and development proceed immedi-
ately. As batch of Ae. albopictus eggs begins to hatch,the time between the emergence of the first and the lastlarvae range from few minutes to several dozen daysdepending mainly water temperature and food avail-
ability. A widespread phenomenon within the genus Aedes,including Ae. alhopictus, is that eggs may hatch read-
ily in response to the stimulus of inundation, while othersmay remain dormant for varying periods despite being sub-merged (Edgerly et al., 1993). This phenomenon is known
installment hatching (Gillel, 1971), and is in Ae.alhopiclus when batch of eggs undergoes hatchingfollowed by several additional small hatchings (Gublcr,1971b; Hien, l975c; Hanson and Craig, 1994).Another parameter that is important for the hatching of
Ae. albopicius eggs is the amount of oxygen dissolved in
the water. Low oxygen levels, usually associated with highlevels of microbial activity and nutrients in the water, in-
hatching (Hien, l975c; Imai and Maeda, 1976; Ed-gerly et al., 1993). After egg flooding, microorganisms col-onize the egg surfaces, resulting in decrease in dissolved
oxygen owing to increase in metabolic microbial activ-
ity; this, in turn, stimulates egg hatching (Edgerly ct al.,1993). Thus, larval feeding microorganisms to bebasic mechanism that may inhibit egg hatching. As
sult of this observation, studies have focused larval graz-ing activity possible explanation of interspecific and
intraspecific egg hatch inhibition among At.’, albopicius and
congeneric species (Edgerly et al., 1993). This research is
discussed further in the section competitive interactions.The effects of egg freezing in Ae. albopictus, together
with diapause, discussed in the following section.
Egg Diupuuse, Photoperiod, and Adaptations
In temperate regions of the world, particularly the Aedes
aUMpictus-infested of North America and the north-ernmost points of its distribution in Asia (Japan and China),the species overwinters in the egg state (Ishii ct al., 1954;Wang, 1962; Wang. 1966; Imai and Maeda, 1976; Mori and
Wada, 1978; Mori et al., 1981). Survival of the northern
Ae. tilhopictus depends all most of the eggs enteringstate of hatching-suppression sufficient to outlast winter.
This is genetically determined property known diapause.The diapause slate is characterized by reduced morphogen-esis, increased resistance to environmental extremes, and al-tered reduced behavioral activity. All known of eggdiapause in the Culicidae correspond to suppression of
sponse to hatching stimuli by fully formed first instar larvaewithin the egg; state of suspended embryonic developmenthas been demonstrated.
The phenomenon of diapause is considered to be
hormonally mediated, resulting in state of low metabolic
activity imposed by specific stimulus. For Ae. alhopictusstrains from temperate areas, the stimulus appears to berelated mainly to photoperiod and temperature, and it is
adaptive in nature (Hawley, 1988; Focks et al., 1994). Inthis the word "adaptive" genetically deter-mined characteristic that enhances the ability of organ-ism to cope with its environment; adaptation is processwhereby population is altered in such way to be bettersuited to its environment (Ricklefs> 1979; Futuyma, 1986).Overall, shorter days (<13.5 hours) trigger egg diapause,with low temperatures enhancing the photopcriodicsponse. I-onger days tend to favor continuous nondiapausegrowth and development.
Other environmental variables such latitude (geo-graphical origin of the strains) and elevation also playrole in the induction of diapause in Ae. albopicius. Geo-graphical variation in the "critical photoperiod" (Cpp-thephotoperiod at which 50% of the individuals in the
quito population produce diapause eggs) has been reportedamong strains ofAe. albopictus from the United States and
Japan and is known to vary clinally with latitude (Pumpuni,1989; Pumpuni et al., 1992). Some modeling studies also
indicate that higher elevations associated with in-
creased Cpp in Ae. albopictus (Focks et al., 1994). StrainsofAc. albopictus from tlie tropics and subtropics do not dem-
onstrate sensitivity to photoperiod. Also, Ac. ulhopictus eggsfrom tropical have not demonstrated overwinteringpabilities. For tropical Ae. alhopictus, survival to de-
pend erratic rather than delayed hatching. Ae. albopictusis considered to display facultative diapause.2 Mori et al.
(1981) demonstrated that pupal and adult Ae. albopictusthe stages that sensitive to photoperiod.The photoperiodic response ofAe. albopicius from tem-
perate appears to have favored the rapid northerlyspread of the species in the central and eastern United
States, where it has colonized vast territory in just few
years. Ae. albopicius moved northward from latitude of
30 N in Houston. Texas, to 42 N in Chicago, Illinois,within three years. This observation also reinforced earlyconclusions that Ae. albopictus from North America have
temperate origin in Asia (Hawley et al., 1987). The north-distribution of Ae. alhopictus in the United States is
clearly limited by cold tolerance. Nawrocki and Hawley(1987) have concluded that the -5 C January isotherm is
Diapause in this is defined period during which growthdevelopment of Ihc insect is spontaneously suspended. In species with
obligatory diapau;>e, only single generation per year is found (univol-tine species). Some Audi’s species present this phenomenon
and southern latitudes. species with facultative diap.iu.se, such
albopiclu’i, multiple generations possible during ihc (mul-li\olline species). This type of diapause usually midnorlhcrn and
midsouthcrn latitudes (approximately 35-65).

good predictor of the northernmost distribution of thespecies in the United States.By contrast, the southward movement of Ae. albopictus
has been relatively slow, leading scientists to hypothesizethat the temperate origin and associated photopcriodicsponse of the U.S. populations would prevent their colo-
nization of the Neotropics. For instance, for Ae. albopictuspopulations to remain active in southern Texas and Florida
during significant portions of the year, they will requirelowering of the day length threshold total elimination ofthe photoperiodic response, because in these regions daylengths for most of the year less than 13.5 hours. How-
rapid selection for both of these features has been
noticed in Ae. alhopictus populations along the southernedge of its Florida distribution, suggesting rapid adaptationto shorter photoperiods (O’Meara et al., 1993). A theoret-ical explanation for this phenomenon could resemble thefollowing scenario: A temperate-zone population trans-
ported to location (southern of TexasFlorida) would experience combination of short daylengths and favorable temperatures for egg and larval de-
velopment for part of the year. During these periods,photoperiodic Ae. ulbopiclu’, genotypes would multiply,while photopcriodic genotypes would remain diapausingeggs. Thus, the transformation of photoperiodic popula-tion into nonphotoperiodic requires mutation, butmerely change in gene frequency.
In the laboratory, selection of nonphotoperiodic Ae. al-
bopiclus from originally photoperiodic population has
been accomplished within generations. In naturalpopulations of Ae. alhopictus from southern Texas andFlorida, the ability to diapausc has been lost three-
year period (Craig, 1993). This indicates that the selectionfor loss of photosensitivity is phenomenon of southernlatitudes, Ae. alhopictus’ invasion of the southern UnitedStates (i.e., peninsular Florida) began slowly in comparisonto its northern and eastern expansion. The genes had to
change via natural selection. Now the species has lost itsability to diapause, and the southern populations of Texasand Florida analogous to their tropical ancestors anddifferent from their temperate antecedents of the 1980s. Itis clear that the species is already in position to invade
any of Neotropical America, and the process may al-ready have begun (i.e., northern Mexico and the DominicanRepublic).
Finally, also in regard to the photoperiodic response,strain from Cariacica City, Brazil, showed sensitivity to
any photoperiod. Based this observation, Hawley andco-workers (1987) hypothesized that all strains from theUnited States might have origin in northern Asia
(probably Japan), while those introduced into Brazil haddifferent origin (likely tropical one). However, recent
studies based allozyme analysis indicated that the U.S.and Brazilian populations appear to share origin,with the evidence pointing to northern Japan (Kambhampati
et al., 1991). A recent publication by Hawley rein-forced the assumption that the Brazilian populations mayshare temperate ancestor with the U.S. popula-tions (Hawley, 1991).
Egg Cold Hardiness
The ability of insect to survive suboptimal tempera-tures is known "cold hardiness" (Hanson, 1991). Coldhardiness plays important role in the life cycle of severalinsects inasmuch it affects factors such adaptation to
seasonal changes, long-term population fluctuations, and
the geographical distribution and colonization trends of in-
vading species,It is well accepted that there (wo different types of
cold-hardy insects: freeze avoiding and freeze tolerant.Freeze-avoiding insects produce antifreezes such low-
molecular-wcight carbohydrates, polyols, and thermalhysteresis proteins, and may also discard ice-nucleatingsubstances to decrease their supercooling points (SCP). TheSCP of insect is the temperature at which it freezes.Freezc-tolcrant insects survive by producing ice-nucleatingagents that increase their SCP by inducing ice formation.Low-molecular-weight sugars and polyols to
both freezc-avoiding and freeze-tolerant insects. Glyccrolto be the most frequently encountered substance of
this type (Somme, 1982). It appears that the most
stimulus for cold hardiness is low temperature (0-5 C)(Baust and Lee, 1981; Horwalh and Duman, 1983; Kukalel al., 1989). Other environmental factors associated withthis phenomenon photoperiodic diapause (Lee and Den-linger, 1985), desiccation (Patterson and Duman, 1978),diet (Knight and Bale, 1986), thcrmoperiod (Horwath andDuman, 1986), and anoxia hypoxia (Somme, 1966).
Cold-hardiness studies have focused eggs of several
orders of the class Insecta, particularly those of the orderDiptera. All insect eggs studied to dale freeze avoiding.Egg cold hardiness studies have been carried out in severalspecies of the genus Aedes, including Ac. albopictus(Hatchett, 1946; Hawley et al., 1987; Copeland and Craig,1990; Hanson, 1991).
Hawley et al. (1987) found that eggs of Ae. ulbopiclusfrom northern Asia and the United States coldtolerant than eggs from tropical sites. In this study eggs
subjected to -10 C for 24 hours, treatment that
killed all Aedes aegypti and tropical Ae. albopictus, U.S.strains, however, showed hatch rate of 80%-90%. Eggsplaced outdoors in South Bend, Indiana, in mid-October1986 showed survival rate of 100% in mid-February1987. These studies clearly demonstrated that the speciesoverwinters in the northern United States and that coldclimation and diapause increase the cold hardiness of tem-
perate Ae. albopictus strains (Hawley et al.. 1989).

Laboratory studies based genetic between
temperate and tropical strains of Ac. albopiclus showed that
hybrid eggs with temperate female parent 36%cold hardy than those with tropical female parent (Han-son, 1991). The male parent docs influence the trait but
appears to play lesser role. The results suggest that themode of inheritance of cold hardiness "is under maternalinfluence by either cytoplasmic sex-linked factors’’
(Hanson, 1991).A set of experiments carried out in East St. Louis, Illi-
nois. which designed to insert genes to reduce the
overwintering ability of Ae. albopictus field strains, pro-duced decrease in cold hardiness and diapause incidence
in the populations (Hanson, 1991). In the studies, malesfrom tropical strain of Ae. alhopictus with distinctive
isozyme marker released. Subsequent egg samplesfrom the release site showed high (24%) frequency of the
genetic marker accompanied by reduced cold hardiness ofthe field populations. These experiments may open the wayfor genetic control of Ae. albopictus in the colder parts ofits range.
All in all, cold acclimation and diapause enhance the coldhardiness of only temperate strains of Ac’, alhopictus.
Egg Morphology
The eggs of Ae. albopiclus cigar-shaped, like those
of Ac’, aegypli, and devoid of floats. Egg morphology, based
scanning electron microscopy observations, is fol-
lows: Eggs average of 609.8 |j,m (+/- 5.9) in
length and 192.9 (im (+/- 2.4) in width (Linley, 1989). Theyhave "large, central outer chorionic tubercles smoothlyrounded, with surrounding cell fields almost invariably de-
void of small tubercles, leaving distinct clear aroundeach large tubercle," shown in Figure (Linley, 1989).For purposes of comparing the structure of the eggs of thetwo species of subgenus Stegomyia in the Amcricas,electron micrograph of Ae. aegypti egg shows the outer
chorionic tubercles surrounded by fairly narrow, feature-less ring (Figure 2). Ae. albopictus eggs blunt at theanterior end and tapering in the posterior region. The
ventral margin is crescendo than the dorsal marginand has conspicuous micropylar collar that is moderatelyprominent, compared with the very prominent foundin Ae. aegypti (Linley, 1989). This characteristic visibly dif-
ferentiates the two species.
Larvae
Under natural conditions Ae. albopictus larvae de-
velop in water with low turbidity and at pl-1 ranging from
5.2 to 7.6, with optimal pH between 6.8 and 7-6 in Asia
(Ho et al., 1972). Water containing amino acids, ammonia,and, in general, high organic nitrogen content appears to
Figure 1. Electron micrograph of entire egg of
Aedes albopictus. Scale: bar 100 (JLM. (Photo byDr. John Linley, Florida Medical Entomology
Laboratory, University of Florida at Vcro Beach.)

Figure 2. Electron micrograph of entire egg ofAedes aegypti. Scale: bar 100 [Jim. (Photo byDr. John Linley, Florida Medical Entomology
Laboratory, University of Florida at Vcro Beach.)
be the ideal habitat lor Ae. albopictus (Laird, 1959). Tothe effects of pH fourth instarAf. albopiclus lar-
vae, laboratory studies conducted with colonizedstrains in New Orleans. Larvae tested to determinetheir tolerance to various pH levels. The results showed thatpH equal to acid than 2.0, equal to
alkaline than 12.0, is fatal to mature larvae after four-hour
exposure (NOMCB, 1987), The studies also reconfirmedthe observation that the species is able to survive in water
containing relatively high concentrations of strong electro-lytes. Measurements in the natural larval habitats of thestrains in the New Orleans study showed pH ranges of6.33-8.35 for tires and 6.43-8.23 for tree holes.
Larvae have been known to survive ranges of dissolvedoxygen of 3-6 ppm in variety of habitats (Ho et al., 1972).Much lower survival ranges have been observed in the tree
holes of New Orleans (sugarberry and red maples), whichhad range of 0.6-2.7 ppm. Also, the larvae have
been found alive at values low 1.3 ppm of dissolvedoxygen in tire yards in New Orleans (NOMCB, 1987).
Adaptability to varied habitats enables the species to
breed successfully in wide variety of water-retainingtainers. It also appears that Ae. ulhopicfus larvae
tolerant of water with high organic content thanAe. aegypli larvae, although the latter species has beenreported from highly organic water conditions suchfound in tree holes, septic tanks, soak away pits, and thelike.
lAirval size and the duration of larval developmentinfluenced by temperature, food supply, crowding, and
Under laboratory conditions, Hien (1975a) demonstrated that
temperature affects the duration of larval development. At30 C development look 6 days, while at 25 C and 20 C,it took 9 and 13 days, respectively. Laboratory studies of the
Brazilian strains have shown larval developmental periodranging from 4 to 9 days at temperature of 25 C (Xavicret <U, 1991). Hien (1975a) ;ilso demonstrated that the fourthlarval instar period is the longest and that the effect of tem-
perature is clearly defined for that stage.Regarding the effect of food supply, starvation extends
the larval development period to average of 42 days,with 80% mortality rate. One of the findings of Hien
(1975a) thai Ae. alhopictus successfully fast for
long periods of time. Thirty-five percent of 100 specimensemerged from larvae that had been starved during the first
three developmental stages.Under natural and artificial conditions, high larval den-
sity appears to produce high mortality and increase inlarval development time. In comparison to Ae. aegypli,however, Ae. albopiclu:, resistant to crowding(Hien, 1975a; Mori and Wada, 1978).
Livingstone and Krishnamoorthy (1982), conducting lab-
oratory experiments with Ae. ulbopictus larvae in India,demonstrated that the larval period of females (119-149hours) greater than that of males (115-141 hours).

Larval Seasonal Prevalence
Larval seasonal prevalence otAe. albopictus appeared lo
be closely related to rainfall. Studies in Singapore (Ho cl
al., 1971), Bangladesh (Azzizar, 1980). Japan (Toma ct al.,
1982; Sota ct al., 1992), and the United States (NOMCB,1987; NOMCB, 1988) have shown thai the species is abun-
dant during the rainy It is important to point outthat since Ae. albopictus1 range extends from the tropics to
the temperate zone, seasonal peaks will differ with
In representative of the tropics such the city ofSingapore, Ac. alhopiclus presents three well-defined larval
population peaks: December, April-May, and August-September. The population peaks of the larvae precede thoseof the adults by almost exactly two months (Ho et al., 1971).In temperate regions, such northern Japan, larvae of Ae.albopiclus present from late March through mid-Septem-ber. High larval population peaks have been observed in
July-August and October, shortly after the rainfall peaks(Toma et al., 1982; Sola et al., 1992). High larval mortalityhas been observed to in the winter (Makiya, 1973;Mori and Wada, 197S; Mod ct al., 1981).
In Okinawajima Island in (lie Japanese subtropics, largepeak in the larval population observed from June to
mid-July, and the population became very low during No-vember through March. Larvae collected every monthof the year (Toma et al., 1982).
Studies in the United States conducted in New Orleansin tire yards and tree hole habitats during the of1987 indicated that the abundance of Ae. albopictus larvae
peaked in June (almost simultaneously with rainfall peaks)and decreased gradually through August (NOMCB, 1987).Conversely, maximum larval production in Ircc holes in
of the Paraiba Valley, Sao Paulo, Brazil, docs not
appear to coincide with rainfall peaks (Gomes ef a]., 1992).The highest larval production observed in the monthsof March and April, which correspond to the fall inthe Southern Hemisphere.
Larval Morphology
Microscopic examination is necessary to identify Ae. al-
bopictus larvae and differentiate them from other Aedcs
species, especially Andes aegypii larvae. The most signifi-cant differences between Ae. uegypli and Ae. albopiclus,which sometimes share the habitat, the following:On the mesolhorax and mctathorax.Ac. albopictus has pleu-ral hair groups that lack the long spines found in Ae. aegypli.In Ae. albopictus there two branches in seta 7-C the
head, while Ae. aegypti has only The ventral brush ofAe. albopiclus has four pairs of hairs, whereas Ae. aegypii’sbrush has five pairs. Ae. alhopictus comb scales have bare
apical spine and of small spinules basally each
side, while Ae. uegypti’s comb scales present several shorter,
stout, subapical spines (Huang, 1972) (see Figure 3).
Larval Habitat
Ae. albopictus is container breeder. It breeds in bothnatural and artificial receptacles. Ae. albopictus has beenfound breeding in such natural habitats tree holes, bam-
boo holes and stumps, coconut shells, plant axils (bromc-liads), ground pools, and rock pools. Its artificial habitats
include rubber tires, tin cans, drums, earthenware contain-
ers, bottles, flower pots, cisterns, and buckets.In the continental United States, infestations of Ae. al-
bopiclus have been found in used tire stockpiles, salvageyards, illegal dumps, the premises of tire dealers workingwith trucks and large equipment, and tire retreaders. Sur-veillance conducted in residential of the United Stateshas also reported infestations in tin cans, bird baths,clogged rain gutters, water bowls for pets, buckets, plasticand metal containers holding rainwater, and cemeteryflower pots (Moore et al., 198S; O’Meara and Gettman,1991). Natural containers such tree holes and tank bro-mcliads appear to be among the preferred habitats of Ae.albopictus larvae in suburban settings of the United Statesand Brazil (NOMCB, 1987; O’Meara et al.. 1993; Gomesct al., 1992). Recent reports from the Dominican Republichave determined that tree holes, rock holes, and fast-foodcontainers the main larval breeding places. Tree holesof Celtis luevigala, species of sugarbcrry, of themain breeding habitats of the species in New Orleans, Lou-isiana (NOMCB, 1987). Examples of larval breeding placesfor Ae. albopiclus from the United States presented in
Figures 4 to 14.
Preferred natural larval habitats in various countries ofAsia bamboo stumps, coconut shells, rock holes, andground pools (La Cassc and Yamaguti, 1950; Hong et al.,1971; Harrison et al., 1972; Ho el al., 1972; Huang, 1972;Rozeboom and Bridges, 1972; Miyagi et al., 1983). Incounlries of Southeast Asia other major breeding habitatsinclude earthenware, tin cans, ant traps/ rubber tires, bowls,and drums. Ant traps (he most indoor habitat
in places such Singapore, Dacca, and urban of Ma-laysia (Ho ct al., 1972; Azzizar, 1980; Chan, 1985).
Landscape lank bromeliads have been studied breed-
ing places for Ae. albopictus in number of locations incentral and southern Florida in the United States (O’Mearact al., 1993). In the northern latitudes of Florida, immatureAe. albopictus have been recovered from Aeclimea disti-chantha, cold-hardy tank bromeliad, which grows intral Florida in the city of Gainesvillc (2940’ N latitude).Before Ae. albopiclus became established in the area, the
^inall ceramic jars fillixl with pjaced under the leys ol satesin houses without refrigerator;,.
10

Figure 3. Drawing illustrating the most important differences
between Aedes albopictus and Aedes aegypti larval stages. (Drawingprovided by Dr. George O’Meara, Medical Entomology
Laboratory, University of Florida at Vero Beach.)
bromeliad the dominant microhabital for Wyeomyiaspp. It appears that Ae. alhopiclus is the main speciesin Aechmea distichantha, replacing other culicids. The spe-cies has been found in than 70% of all bromeliads
sampled in the Gainesville studies. This observation ap-pears to have been influenced by the fact that many other
species of tank bromeliads killed severely damagedby hard freeze. Two months after the freeze and forperiod of than 15 months, Ae. albopictus the onlyspecies recovered from the cold-hardy tank bromeliad.
Thus, because of the cold winter, and in the absence of
’Wyeomyia, Ac. ulbopictus is the dominant species (O’Mearaet al.. 1993).
Farther south of Gainesville in Vero Beach, Florida
(2740’ latitude), Ae. albopictus larvae have been found inother bromeliads such Neorcgalia spectabilis, BUlbergia
pyramidalis, and Aechmea fasciatus. In these microhabitatsAe. albopictus collected from fewer than 10% of theplants sampled, while \^/yeomyia spp. found in nearly90% of the bromeliads sampled. Thus, the main culicids atthis site Wyeomyia spp. (O’Meara et al., 1993).
In countries of tropical Asia, the breeding preferences ofAe. albopiclus in plant axils include leaves of Pundanusand banana trees and leaves of Colocusis esculenta (taro)(Kurihara, 1984).A study conducted by Laird (1959) in several of
the city of Singapore described of the characteristicsof the larval microhabitat ofAe. albopictus where the larvaehad optimum development. In relation to the aquatic floraand fauna, the water of the larval habitat classified
p mesosaprobic. p mesosaprobic waters supportedrich and diverse flora and fauna consisting of several spe-
ll

Figure 4. Aedes albopictus-mfested tires, Houston, Texas, U.S.A., March1986. (Photo by Dr. C. G. Moore, Division of Vector-Borne Infectious
Diseases [DVBID], CDC.)
Figure 5. Placement of a CDC ovitrap for Ac. albopictus surveillance
(Photo by Dr. C. G. Moore, DVBID, CDC.)

Figure 6. Tree hole containing water and Ae. alhopictus larvae, NewOrleans, Louisiana, U.S.A. (Photo by Dr. J. E. Freier, DVB1D, CDC.)
Figure 7. Bottle with water and Ac. albopictus larvae. (Photo byDr. C. G. Moore, DVBID, CDC.)

Figure 8. Discarded refrigerator door, with water and leaf litter infestedwith Ae. albopictus larvae. New Orleans, Louisiana, U.S.A. (Photo by
Dr. J. E. Freier, DVBID, CDC.)
Figure 9. Checking for adult Ac. atbopictus at used tireretread facility, Atlanta, Georgia, U.S.A. (Photo by Dr. C. G. Moore,
DVBID, CDC).

Figure 10. Aedes albopictus larvae in discarded toilet bowl tilled with
water. (Photo by Dr. C. G. Moore, DVBID, CDC.)
Figure 11. Aedes albopictus larvae in plastic sheet with water and leaf
litter. Note adjacent tire casing also containing water. New Orleans,Louisiana, U.S.A. (Photo by Dr. J. G. Freier, DVBID, CDC.)

Figure 12. Miscellaneous container habitats formosquitoes, infested with Ae. alboplctus larvae.(Photo by Dr. C. G. Moore, DVBID, CDC.)
cics of diatoms, desmids, microcmstaceans, phytoflagellates(Eiiglenaccac), ciliates. and many different rotifers, cyclo-poids, ostracods, and dccapods. All of the above providedideal conditions for development of Ac. albopictus becauseof the abundance of food and the relatively low concentra-tion of deleterious substances. The mesosaprobic habitat
less well suited for Ac. alhopiclus development;theless, substantial densities of Ae. alhopiclus larvaefound. That water contained free-living green algae, blue-grecn algae (Oscillatorio), flagellates, phytoflagellatcs (Eu-gle-na), Paramecium, vorticellas. and rotifers. Thetacean fauna abundant in cyclopoids and cladoccrans.
Larval Pathogens and Predators
Several pathogens and predators oiAe. albopiclus larvaehave been described in the literature.
The fungus Coelomomyces slegomyiae foundseveral Ae. albopiclus larvae in Singapore. The second in-star larvae showed the heaviest infections, concentrated inthe head capsule, thorax, and anal papillae (Laird, 1959).High infection rates in experimental trials with Coelomo-myces slegomyiae chapmani have demonstrated the po-tential of the fungus to biological control agentfor Ae. alhopicfu’i well other Culieidae (Lien and Lin,1990). The infection process of Ae. alhopiclus larvae byanother fungus, Tolypocliiumm cylyndrosporum, has alsobeen described. The pathogcnesis of the fungal invasion ofthe exoskeleton and the digestive apparatus of the larvaehas been studied to extent, but the potential ofthis fungus biological control agent for the species isnot yet clear (Ravallcc et aL, 1989).A gregarine protozoan parasite, Lankvsteriu culicis
(= Ascogregarina iaiwanensis Hen & Levine, 1980) isknown to develop trophozoites in the cells lining the larval
midgut ofAeiley albopiclus (Huang, 1972). Recent studiesconducted in Illinois (Munstermann and Wesson, 1990)have demonstrated the presence of Asc. taiwanensis instrains ofAc. albopictus from North America. Field-collectedspecimens showed infection rates of 67% to 95% at toursampling sites; subsequent isolations of the parasite fromfield-collected Ae. albopictus hosts to show path-ogenicity in the larvae and pupae of laboratory strains of Ac.alhopictus. In general, although the gregarine protozoan haslittle effect its natural host, it does appear to havemarked deleterious effect indigenous mosquitoes of NorthAmerica such Ae. aegypti and Ae. alropalpus (see thediscussion parasitism in the section competition stud-
ies). Also, the study reports strain variation in suscep-tibility to infection among the different North American Ae.alhopictus populations. This study is the first to find thisgregarine protozoan in culicid larvae in North America.Some workers have reported experimental infections of
Ae. albopictus larvae with the nematode Komanomermisculicivot-ax (Petersen and Chapman, 1979; Blackmore,1992). So far, parasitism of Ae: albopiclus by mermithidncmatodes has not been reported in any natural populationsof the species. The laboratory studies mentioned abovehave shown intermediate infection rates in North Americanstrains of Ae. albopiclus. Mermithids known to infectlarval mosquitoes by penetrating the cuticle and subse-quently maturing in the hcmocoel of the host.
Laird (1959) also found variety of other pathogens,such the ciliates Tetraliymena pyriformh and EpislylislacuslrLs, the alga Oscillaiona hrevis, and the bacteriumSphae-roiilus dichotomus.
In regard to predators, Marten (1984) observed cope-pod predator, Mesocyclops leuckarii pilosa, feeding uponfirst instar larvae of Ae. albopictus. The copcpodpable of eliminating them entirely. A drawback to theof this predator is that it appears to be in Ae. albo-piclus habitats. Further research conducted by the in-
16

Figure 13. Discarded tires potential Ae. albopictus breeding site.
(Photo by Dr. C. G. Moore, DVBID, CDC.)
Figure 14. Inspection of tire casings from Asia for Ae. albopictus larvae.
(Photo by R. E. Schoeppncr, San Mateo Mosquito Abatement District,Burlingame, California, U.S.A.)

vcstigator has demonstrated Ihc successful of other spe-cies of larvivorous copcpods in controlling Ac. albopicluslarvae in tire piles. In period of S-10 weeks the crusta-
Macrocyclops albidus (Jurine), species with broaddistribution in the Amcricas, able to completely elim-inate larvae in tire piles found in and around New Orleans(Marten, 1989a; 1989b; 1990a; 1990b). Marten also dis-
the of mixtures of several species of cyclopoidcopcpods to control species otSlegomyia and other culicidsof medical importance and these copepods’ potential effect
larval aquatic habitats. The mixtures of Macrocyclopsatbidus and Mesocyclops rultneri, both collected in naturalhabitats in New Orleans, proved to he effectiveMacrocyclops by itself. An advantage of this approachthe increase in survival rates of both species in the mixture,compared to the survival rate when single speciesintroduced (Marten, 1990b).
Finally, various Toxorynchiles mosquito speciesknown to prey Ae. albopiclus larvae. One of these pred-ators that has been found breeding successfully with Ae.alhopiclus larvae in Asia is Toxorynchiles splendens (Siir-tccs, 1966). II appears that Toxorynchites spp. may beeffective in controlling Ae. albopiclus larvae. Two speciesof To-wrynchites, introduced (amboinensis) and theother indigenous (rulilus septenirionalis}, have beensively produced in insectaries, and control trials beingcarried out in of New Orleans to test their efficacy inthe field (Freier, pcrs. comm., 1993).
Pupae
Under ideal conditions, Ae. albopiclus remains in the pu-pa! stage for about two days (Sheng and Wu, 1951; Surtces,1966; Hicn, 1975a), and. in other Aedes species, Ae.albopiclus males emerge before females. Livingslonc andKrishnamoorthy (1982) showed that the pupal developmentperiod 32-36 hours tor males, while females reachedthe adult stage in 49-52 hours. The experiment sug-gested that pupal development of females requires mini-
dark period of 12 hours.Hicn (1975a) studied the ctfcci of water temperature
pupal development. The results showed that pupal devel-opment lasted two days at 30 C, three days at 25 C, andfive days at 20 C. Ae. albopiclus pupae survive desiccationfor up to two days at 26 C and relative humidity of 87%.Pupal mortality about 1% under field conditions(Ho et al., 1972).
Adults
Longevity
Studies of longevity (survival) in different populations ofvector mosquitoes (particularly females) often provide im-
portant epidcmiologic information, because greater longev-ity increases the probability of transmitting diseases. Oneof the most important questions concerning Ac. tillmpictusis whether adult females of this species capable of livinglong enough to become infected and subsequently transmitinfectious agents. Studies of male survival may also be im-portant, especially in the application of certain genetictrol In addition, survival data may be useful inthe assessment and administration of vector controloperations.The bulk of the data longevity ofAe. albopiclus adults
(survival studies) from laboratory research (reportedin Hawley. 1988). Few studies have attempted to estimate
longevity otAe. albopiclus adults in the field. Thus far, twostudies have been carried out with North American strains.Some of these laboratory and field studies described below.
Environmental factors stimulate responses thai greatlymodify the behavior and biology of the adult mosquito.Temperature and relative humidity among the factorsthat play vital role in adult survival. Gao et al. (1984)reported that during eight years of observations of cagedAe. albopictus (kept at temperature conditions of 25 +/-
C and relative humidity of 80 +/- 5%). the maximumlongevity of the adult females 30-40 days. Thestudy demonstrated that the upper thermal survival limit
40 C (LT;(, 0.9 hour) and the lower limit-5 C (LTso 0.85 hour).Hylton (1969) showed that longevity of the adult Ae.
albopiclus female reached about 84 days at temperatureof 22.2 C and relative humidity of 80%. Ae. albopiclus
tolerant of low temperatures (15.5 C) and higli andlow humidities, whereas high temperatures (32.2 C) de-creased survival, independent of the relative humidity. Itappears that the ability ofAc. albopiclus to survive varioustemperatures and humidities may have favored its distri-bution in climatic conditions that range from temperate totropical. It has been suggested thatAc. alhopictusbody fluids at low humidities, thereby preventing (issue
desiccation, through spiracular control another moistureregulating mechanism (Hylton, 1967).
longevity in the laboratory is also affected by feeding.Gublcr and Bhaltacharya (1971) maintained mosquitoes at
temperature of 26 C and relative humidity of 50%-60%, and demonstrated that blood-nourished Ac. albopictusfemales lived from 38 to 112 days. A maximum of 117days found by Hien (1976a).By contrast, mosquitoes that fed with water and
kept at temperatures of 25-26 C and 6()%-70% relativehumidity began to die after three days. The majority ofstarving Ac. albopiclus did not live longer than five to
days (Hicn, 1976a). Under natural conditions femalewould normally find suitable host before this time intervalelapsed.
There two general approaches for measuring the rateof survival in insects: indirect methods based physio-
18

logical age within population and direct estimates bymark-rclease-rccapture experiments (Service, 1976).
Indirect methods to female survival based
chiefly tracheation of the ovaries and ovariolar dilata-tions (Polovodova, 1949; Detinova, 1962). Analysis of thetrachcaiion of the ovariolcs yields the parity rate, i.e.. the
proportion of the population that has oviposited. Therefore,the mosquito population be separated into two groups:
older group with oviposition cycle (parous)and younger group with oviposition cycle (nulHpa-rous). The greater the proportion of parous females, theolder the population. Parity rates determined by dis-
secting the ovaries of unfed recently fed females and
examining them for the presence absence of clumpedcoiled iracheoles. It’ the tracheoles coiled, the female is
nulliparous; if they not coiled, the female is parous.Parity rates important for comparing the age of popu-lations of different species in different after in-secticide treatment. Most important, if the oviposition cycle
the gonadotrophic cycle is known, both the daily survivalrate (proportion surviving through day) and the averageage of the population be estimated mathematically. Thedaily survival rate /; is estimated by extracting from the
proportion of parous females the root equal to the numberof day?, in the oviposition cycle. The formula is follows:
p ==-\/proportion ol’ parons females, where the ovipo-sition cycle in days. The ovariolar dilatation method deter-mines the number of times that female mosquito has
oviposited by examining dilatations in the basal part of thefollicular tube (pedicel), small duct that connects eachovariole in the ovary to the lateral oviduct (calix). Each
oviposition produces dilatation; thus, the number of di-
latations found in the follicular tubes of ovary indicates
the number of times the mosquito has oviposited. Theber of ovipositions indicates the physiological age of the
mosquito, which, when multiplied by the estimated number
of days in the oviposition cycle, yields estimate of the
chronological age.Estimates of adult longevity of Ae. ulbopictus based
ovariolar dilatations and tracheation of the ovaries have
been carried out in several laboratory studies. Hawley(1988) discusses their shortcomings, pointing out that agemay be underestimated when the estimate is basedriolar dilatations. For instance, studies have shownthat Ae. albopicfus females do indeed form recognizabledilatations, but that their number does not equal the known
number of gonotrophic cycles completed- Most of thetimates of daily survival based the assessment of parityrates of Ae. ulbopictus have been conducted with Asianstrains. The results displayed/? values ranging from 0.71 to
0.88, which correspond to average female longevity of2.9 to 7.8 days (Hawley, 1988). The average longevity of
the population is calculated by taking the reciprocal of the
negative, natural logarithm of p, that life expectancyl/-ln p.
Mark-release-recapture methods used to directly esti-mate survival usually involve marking known number ofindividuals with various dyes, releasing them into the wild,and then trying to recapture them by different Aregression line is then constructed by plotting the values ofthe logm(recaptures + 1) against the number of days post-release. Next. the slope of the regression is calculated in
simple way to obtain the probability of daily survival, p
(Walker et al., 1987). The antilog of the regression coeffi-cient yields the daily survival rate. Aside from being ofchief importance in survival studies, direct methods, such
mark-recapture analysis, also used for the followingpurposes: to estimate adult densities; to dispersal;to determine the timing of reproductive events; to evaluatethe extent to which individual vector returns to the
species for its blood source; and to identity resting places,especially outdoors and during the of minimum
density.In Singapore the oldest females collected in houses andhuman baits had undergone three gonotrophic cycles,
suggesting period greater than 11 days, while in Cal-
cutta dissections of wild females showed only two
ovipositions, indicating shorter life-span than in
Singapore (Ho ct ai- 1972; Gubler and Bhattacharya,1971). Some other studies have reported collection ofmarked Ae. ulbopictus females at periods greater than 20
days postrelease (Bonnet and Worchestcr. 1946; Rosen et
al., 1976; Niebylski, 1992).Mark-reieasc-recapture studies conducted with North
American Ae. alhopiclus strains in the eastern part of the
city of New Orleans during the rainy of 1987showed overall average probability of daily survival of0.77 for both males and females (NOMCB, 1987). Thus,the life expectancy value for the population about four
days. More recent research, also from North America,carried out in tire yard in Potosi, Missouri, during 1989-1991 and showed higher survival rates for Ae. albopictusfemales than those reported in other studies. In this researchthe p value for females 0.89, which corresponds to 8.6
(or about 9) days life expectancy in this population (Nie-bylski, 1992). In this particular study, single femaleobserved to survive 24 days. The survival rate p of the adultmales 0.77, which is comparable to the values reportedby other researchers. An average 8-day survival ofA<?. al-
bopicius females combined with average gonotrophiccycle of four days that Ae. ulbopictus potentially will
take two blood meals in lifetime, increasing the vectorial
potential of the species.
Mating Characteristics
As slated before, cross-specitic mating between Ac. al-
bopiclus and other species has been observed. The phenom-
19

enon, form of competition for mates, is known satyr-ism, and it has been noticed in laboratory experiments and
in the field, particularly with maleAe. aihopictus. Further-
more, it is known that injections of matrone, the Ae. ai-hopictus male accessory gland pheromonc. renders the fe-male Ae. aegypti refractory to subsequent inseminations(Mi and Rozeboom, 1973; Nasci et al., 1989; Craig, 1993).On the other hand, matrone from male Ae. aegypti haseffect female Ae. albopictus. It is believed that the pro-ficiency of the Ac. albopictus males in mating with other
species could be involved in the dramatic displacementtrends of Ae. aegypli by the Ae. albopicliif populations in
the southern United States (Nasci et al., 1989).Natural mating behavior of Ae. albopicluf reported
in Calcutta study (Gubler and Bhattacharya, 1972). Itfound that male Ae. alhopiclus swarmed around the feetand ankles of human observer. Female Ae. albopictusrived later and mated they approached the host to
feed. Mating took place in flight and usually occurredwithin to three feet of the ground. Most of the matingslasted from 5 to 15 seconds. A majority of the copulatingfemales that captured nulliparous, and mostinseminated in the vicinity of the host. The study also
reported swarming of males at the bases of trees andmetal baskets, leading to copulation and inseminationof females. It has been reported that copulation is suf-ficient to fertilize batch of eggs and that femalesnot inseminated until they 30 to 36 hours old (Ho ct
al., 1972).
Oviposition
The speed of egg production, like that of other physio-logical processes of insects, is critically dependentvironmental factors such temperature.The underlying mechanism that triggers oviposilion in
Ae. albopictus has been studied to extent in strainsfrom India and from Lake Charles, Louisiana, in the UnitedStates (Lcahy and Craig- 1965; Klowden and Chambers,1992; Klowden, 1993). Experimental studies have indicatedthat male accessory gland components of Ac. alhopiclusplay role in inducing egg deposition in the females (Leahyand Craig, 1965; Klowden, 1993). It appears that the ovi-
position stimulant for females is not related to the mechan-ics of mating but rather to stimulant substances producedby the male accessory gland. Likewise, abdominal disten-
tion resulting from blood meal to provide secondstimulus that initiates ovarian development (Klowden,1993).
Although it is generally accepted that Ae. alhopiclus fe-males begin to feed two days after emergence (Del Rosario,1963; Gubler and Bhattacharya, 1971; Hawley, 1988),Ae. alhopiclus have been observed to take their first bloodmeal during their first day postemergence (Hien. 1976b).
Hien (1976b) observed that Ae. albopictus femalesready to lake blood meals several times during singlegonolrophic cycle, that is, the second and third daysafter the first meal. Gubler and Bhattacharya (1971) alsoobserved that Ae. albopictus females took blood forond time in single gonotrophic cycle. Multiple feedingduring gonotrophic cycle could be importantponent in the epidemiology of vector-borne diseases (Bur-kot et al., 1988). This behavior could increase the abilityofAf. alhopiclus to acquire and deliver infections from and
to different hosts, thereby spreading pathogens to largersusceptible human population. Information regarding Ae.albopiclus feeding patterns is sketchy; however, it is knownthat the species has wide host range compared with other
Slegomyia species. Thus, there is real possibility that Ae.alhopiclus may pick up pathogens in ?.oonotic cycle and
carry them to humans in the domestic environment.
Knowledge of the interval between successive oviposi-tions (the duration of the gonotrophic cycle) allowsversion of the physiological age into longevityfor the population and also the study of population dynam-ics. In general, Ac. aihopictus laboratory experimentsto agree with field studies, pointing to average durationof five days for the first and second gonotrophic cycles. Forinstance, Mori and Wada (1977) demonstrated that under nat-
ural conditions, with average field temperature of 25 C,the period from emergence to the first blood meal of Ac.albopictus about two days and the duration of gono-lrophic cycle five days. These results in accordwith laboratory experiments. Using Detinova’s method, the
studies showed that Ae. albopiclus females readyto feed again within 24 hours after oviposition.Under laboratory conditions, Del Rosario (1963) found
that the first blood meal taken two days after
gence, and the gonotrophic cycle found to last three to
four days before oviposition at temperature of 24--29 Cand 80%-93% relative humidity. Gubler and Bhattacharya(1971) also observed that the first blood meal of Ae. al-
bopictus females took place two days after emergence, with
gonotrophic cycle lasting three to five days after bloodmeal at temperature of 25-26 C and relative humidity of50%-60%. Hien (1976b) showed that the gonotrophic cycleafter blood meal lasted three to three-and-a-half daysat temperature of 25-26 C and relative humidity of60%-70%.The number of eggs laid by Ac. albopictus depends
the physiological age of the mosquito, the body weight after
emergence, and, particularly, the size of the blood meal
(Hien, 1976a). In general, it appears that there is linearcorrelation between blood meal size and number of eggsoviposited. Hien (1976a) observed that, average, Ae. al’bopictus females lay eggs after blood meat of al least 0.7mg. For females taking from 0.8 mg to 2.5 mg of blood.the average number of eggs produced in the first gono-trophic cycle 72 per female (Hien, 1976a). The study
20

also demonstrated that the number of eggs produced in-creased with the size of the blood meal taken by the female(85 more eggs at 2.0 mg of blood).A study conducted by Klowden and Chambers (1992)
showed that Ae. albopictus females possess the ability to
utilize extremely small blood meals to develop their eggs,in contrast to other species ofSfegomyia (i.e., Ae. aegyp{i).Values low 0.1 |xl of blood enough to triggeregg development in Ac’, albopiclus strains from Lake
Charles, Louisiana. The average number of eggs oviposited18 per female. In contrast, the other North American
Slegomyici, Ae. uegypli, unable to develop any eggsunless the blood meal volume than (xl. Gublcrand Bhatlacharya (1971) found that the number ofeggs in the first gonotrophic cycle 63.4 per female.Similar results obtained in two other studies: Udaka(1959) reported 62 eggs per female feeding sugar and
blood, and Soekiman et al. (1984) observed average of57.5 eggs per female after single blood feeding.
Based measurements of parity rates for Ac. albopicius,Hawlcy (1988) determined that the number of egg batcheslaid bv female wild mosquito in its life-span is in the
range of 0.2 to 2.1. The average number of ovipositionsinferable from these data is approximatelyThe number of eggs laid also depends the physiolog-
ical age of the female, and the number progressively de-
age increases (Gublcr and Bhattacharya. 1971;Hien, .1976a). Generally speaking, the first gonotrophic cy-cle produces the highest number of eggs, with gradualdecrease in subsequent cycles. On average, Ae. albopiclusfemales the laboratory found to produce 283-344eggs per mosquito during their life cycle (Gubler and Bhat-tacharya, 1971; Hien. 1976a).Autogcny has been observed in laboratory strains of Ac.
albopictus (Bat-Miriam and Craig, 1966; Cui, 1982; Klow-den and Chambers, 1992). Autogeny in this is defined
egg maturation without exogenous of protein.namely, the blood meal. Conversely, species requiring in-
gestion of proteinaceous foodstuffs for egg development is
known anautogenous species (Roubaud, 1929). It hasbeen suggested that autogenous mosquitoes accumulateconsiderably during their larval life than do
anaulogenous This situation may be related to the
somewhat longer larval developmental periods in autoge-strains. Autogeny in mosquitoes is not controlled
solely by larval nutritional conditions but also by geneticcomponents (Twohy and Rozcboom, 1957). Autogenytherefore has ecological significance, since it allows the
mosquito to lay eggs in the absence of host for
feeding, thus ensuring the continuity of the species.In Ae. allwpictus strains from Lake Charles. Louisiana,
autogenous behavior expressed in adult females main-
tained sugar diet (Klowden and Chambers, 1992). Insample of 100 individuals, 5% displayed autogeny. No
data the number of eggs oviposited per female pro-
vided by the researchers. Based data from this research,it hypothesized that food accumulated inextended larval life cycle, compared to that of Ae.gypli, may favor stores of lipids and proteins that enhancethe expression of autogeny in Ae. albopiclus. These obser-vations part of studies assessing the reproductive andmetabolic differences between North American strains ofAe. albopiclus and Ae. aegypll.
With strains from Mauritius and Madagascar, Bat-Miriam
and Craig (J966) demonstrated that adults that fedonly sugar and raisins produced few eggs. The strainfrom Mauritius able to present autogeny for 12 gen-erations. Cui (1982) observed that larvae collected in
Guangzhou City and reared in the laboratory displayedtogeny. The aulogeny index reported for the Guangzhoustrain varied from 1.96 to 2.96. while in the Mauritius strainit 2.7 eggs per female.
Oviposition activity and horizontal and vertical ecologi-cal stratification have been studied by Gubler (197 Ib). Hedetermined from series of 18 ovitrap collections that most
of the eggs laid al ground level, but de-
posited at height of 15 meters, in the tops of trees in
Calcutta, India. In Paradeniya, Sri Lanka, Amerasinghe and
Alagoda (1984) made similar observations; Ae. alhopictusfemales laid about 52% of their eggs at ground level, andthere decrease in oviposilion with increasing heightabove ground. Only 33% of the eggs laid at 3.5 meters
and just 16% of the eggs laid at 7 meters. The results
suggest that Ae. albopictus females, although preferringhorizontal range stratification for oviposition, able to
migrate and oviposit in the treclops.Oviposition sites affected by habitat type, light, tem-
perature, and humidity, well by such subtle influencescharacteristics of the water (biolic and abiotic) and
the oviposition surface (Bently et al.. 1976; Amerasingheand Alagoda, 1984). Ho ct al. (1972) reported that, under
laboratory conditions, female Ae. albopictus preferred to
oviposit in habitats with rough gray surface and lowflectivity rather than smooth black surface with highreflectivity. Yap (1975) observed that ovitraps coaled withred and black preferred ovitraps of other colors.
In their natural habitat, Ae. alhopiclus usually lay eggs inwater reservoirs containing decaying vegetable matter
(Hien, 1976a).The quality of larval food appears to affect the adult’s
oviposition capacity. This effect observed in Ac. al-hopiclus females reared with banana powder in the larvalstage: egg laying particularly weak except al constant
temperature (Ho ct al., 1972).In nature, it observed that Ae. alhopictus females
seldom laid all of their mature eggs in single oviposition.Instead, they appeared to from place to place, at each
site leaving behind few eggs. The female probably laysall mature eggs during the of several ovipositions,periodically interrupting her egg laying to fly to another
21

container. This strategy is considered species survivalmechanism (Rozcboom el al., 1973).
Laboratory observations assessing the influence of bloodin oviposition have been carried out in New Or-
leans, Louisiana. The results showed that mosquitoes feed-ing rodent species laid significantly eggs thanthose feeding avian, rabbit, human hosts (NOMCB,1989).
findings of other researchers (Tempclis ct al., 1970) andadds to the potential of the species to become involved inthe cnzootic/endcmic transmission cycles of indigenousboviroses and other vector-borne diseases. Ac. albopictusbreeds in surrounding the human environment and hasdemonstrated highly anthropophilic behavior; thus, the spe-cies participate in the sylvan zoonotic cycle and carrydiseases to
Host Preference
Aedes alhopictus females feed wide range ofmals well birds. Host-feeding patterns for wild Ac.albopictus populations in the New World showed that thespecies is opportunistic feeder that appears to be pri-marily attracted to mammals rather than other types of hosts(Savage ct a]., 1993). Savage and co-workers conductedstudies in the of 1989 and 1990 in large tiredump located in Potosi, Missouri. Their sample consistedof 172 Ac’, alhopictus females, which tested byof the precipitin test and direct ELISA. The results showedthat 64% of the mosquitoes had fed mammals and 16.9%
birds. This study identified nine mammalian taxahosts: rabbits, deer, and dogs (the three most common),followed in decreasing order by humans, squirrels, opos-sums, myomorphs (except Raltus), bovincs, andFour avian taxa also identified asAe. albopiclus bloodhosts: passcritbrms, columbiforms, ciconiit’orms, and quail.Various blood meals from unidentified mammalianand avian hosts.
In extension of the Missouri studies to fourstates (Florida, Illinois, Louisiana, and Indiana), Niebylski(1992) and Niebylski et al. (I994a) found twomalian laxa (rats and cats), well turtles, preferredhosts, in addition to the nine mammalian taxa named above.The variation in blood hosts between sites and within sitescould be governed by the relative abundance of specificvertebrates at the time of sampling.
In Thailand and the Hawaiian Islands, Ae. alhopictus fe-males collected in sylvan environment fed humans,horses, pigs, buffalo, bovine species, dogs, chickens, andboobies (Tempclis et al., 1970; Sullivan cl al., 1971). In theHawaiian Islands, 93% of the females fed mammals and7% birds.Under laboratory conditions, Ae. alhopictus feeds hu-
mans, rabbits, mice, chickens, rats, and guinea pigs (DelRosario, 1963; Ho cl al., 1972). Ae. albopiclus females inbreeding cages have also been reported to teed snakesand frogs (Miyagi, 1972).
According to Ho et al. (1972), Ae. albopictus prefers tofeed humans, but host availability appears to play fun-damental role in the average behavior of the mosquito pop-ulations. The confirmation in the U.S. studies that Ae. ai-hopictus is opportunistic blood feeder supports the
Biting Behavior
On average, Ae. albopictif, females take their first bloodmeal two days after emergence (Del Rosario. 1963; Hien,1976a). Adult females vigorous daytime biters (al-though they sometimes bite at night) and typically bite out-doors (Feng, 1938; Sheng and Wu, 1951; Surtccs, 1966;Ho et al., 1972; Pant ct al., 1973; J. Frcicr, unpublishedinformation). The diurnal biting cycle ofAc. albopiclus ap-pears to be generally bimodal, with period in earlymorning and the other in evening (Gubler, I971b; Ho el
al., 1973; Basic and Santos-Basio, 1974).Did studies conducted during the of 1987
Ae. albopictus strains from New Orleans, Louisiana, showedthis bimodal cycle of biting activity. The first biting cycleoccurred between 6:00 and 10:00 and the second be-tween 4:00 and 10:00 p.m. Statistical analysis suggestedpositive correlation between biting activity and relative hu-midity (r 0.33, p 0.005). The study observed bitingactivity throughout the 24-hour period, decreasing but notdisappearing during the night-time hours (NOMCB, 1987).
In Calcutta, Gubler (I971b) observed thai the diurnaltivity of Ae. albopiclus females presented primary peakbetween 6:00 and 8:00 and secondary peak between4:00 and 6:00 p.m. In Singapore, well-defined biting activ-ity of female Ae. alhopictus found, with morningpeak just after sunrise at 7:30 and evening peakaround sunset from 5:30 to 6:30 p.m. The prominentning peak may be important factor contributing to dis-
transmission, since in certain tropical human out-door activity increases in late afternoon (Ho ct al., 1973).
In the Philippines, Basio and Santos-Basio (1974) ob-served minor biting peak between 9:00 and 10:00and major peak between 4:00 and 5:00 p.m. The bitingcycle similar in four different geographical
Although biting activity of females occurred predomi-nantly at ground level, it observed high 9 meters
above the ground during the peak biting time in Calcutta(Gubler, I971b). Population studies of Ae. albopiclusconducted by Mogi and Yamanura (1981) indicate tlialAe.albopiclus attraction to humans depends host factorssuch sex, age, race, and clothing, well mosquitoresponsiveness influenced by circadian rhythm, micro-climatic conditions, and undetermined factors related to theindividual host. Thus, females attack humans under the
22

guidance of carbon dioxide, moisture, organic chemicals,;ind visual factors including movement. The studyfound that the range of attraction of Ae. iilbopiclus to
human is circle with radius of about 4-5 meters (anof 50-80 m2). The host-seeking pattern of Ae. albo-
piclus appears to follow two phases: initial random flightand directional flight after encountering stimuli from
host. This assertion is supported by the observation that Ac.alhopklus distribute vertically within the range where stim-uli from human being the ground detectable.
Seasonal abundance of adults varies according to the
geographical Therefore, seasonal biting activity is in-fluenced mainly by temperature and rainfall.
In Ncvf Orleans, adult mosquito abundance peaksdays after the beginning of rainfall. Larval mosquito pop-ulations reach their peak densities in tree holes and tires
during the month of June. Adults found between themonths of May and August. Most adult activity disappearsin late September following the onset of diapause(NOMCB, 1987; NOMCB, 1988).
In the tropics, tor example, in the city of Singapore, therethree adult population peaks: in March, in June-July,
and in November-December. Thus, female feeding activityis highest during those three periods. The population peaksclosely follow those of rainfall (Ho ct al., 1971).On the subtropical island of Okinawa, Toma et al. (1982)
found females feeding throughout the year, with peaks infeeding activity occurring from April to June and from Au-gust to November. In temperate of Japan, adult peaks
from late March to mid-September. La Cassc andYamaguti (1950), in Honshu, Japan, observed that bitingrates of Ae. alhopiclus peaked during the last two weeks ofAugust.
Flight Range
Bonnet and Worchester (1946) determined during mark-rclease-recapture studies with Ac. albopictu.f thai the
imum recapture distance 134 meters. Thus, it is ap-parent that Ac. albopictui has short flight range, rarelyreaching 500 meters (Stojanovich and Scott, 1965). Thisobservation has been confirmed by recent dispersal studiesconducted with North American Ac. albopictus strains fromPotosi. Missouri (Nicbylski, 1992; Nicbylski el al.,1994b). Using mark-release-rccaplure trials, the maximumdispersal distance estimated for females 525 meters,whereas the maximum dispersal distance for males225 meters. It also noted that than 90% of the
Ac. albopictus specimens from the study dispersedless than 100 meters.
Resting Behavior
Aedfs alhopiclw, adults have been found resting outdoorsin clearings and rubber plantations in Malaysia. In China,
the adults appeared in mosquito nets, kitchens, drawingrooms, pig sties, among weeds in the field (Surtees,1966; Ho et al., 1972).
In the United States resting Ac. iilbopiclus have beenfound in forest fringes where both canopy and understory
present (Nicbylski. 1992).
Adult Morphology
Adults of the genus Aedcs be differentiated fromother Culicidac because their abdomen is pointed. The main
characteristic that distinguishes Ac*, tilbopicliis from Ac.gypti is the conspicuous lyre-shaped silver pattern thescutum of Ae. acgypli, compared with the distinct longitu-dinal sil\ line of Ae. aibopictus. If the scales from thethorax inadvertently rubbed off during collection ifother handling problems occur, there is another character
that distinguish these species. Tlic clypcus, structurelocated the head between the palpi, is covered withwhite scales in the Ae, aegypti female and black scales in
the Ac. albopictus female. A pictorial key provided by Dr.G. O’Meara and another trom the Supcrintcndcncia de
Campanhas de Saudc Publica (SUCAM, 1989) in Brazil
(Figures 15 and 16) show differences in greater detail forthe adult stages of both species. In addition, Walter Reed
Biosystematics Unit plate illustrates the external morphol-ogy of adult Ae. albopictus (Figure 17).
Competition Studies
Arclcs albopictus utilizes the kind of artificialter containers that the principal of Ac. acgypti,and several studies report that Ac. albopictus and Ae.aegypii may share the habitat (Gilotra el al., 1967;Chan, K. L., ct al., 1971; Ho et al., 1972; Azzizar, 1980;Sprenger and Wuithiranyagool, 1986; Nasci el a)., 1989;O’Meara el al., 1993). Because of this association, it has
been hypothesized that in parts of Southeast AsiaAe. aegypti has completely replaced the indigenous Ac’, al-Iwpiclus in urban (Pant et al., 1973). Conversely, ob-servations the spread of Ae. albopictus in the southern
coastal slates of the United States indicate that the expan-sion appears to be occurring al the expense of Ac. aefypti.The introduction of Ac. albopictus has been accompaniedby drastic and rapid decline of Ae. aegypti populations(Nasci et a)., 1989; O’Meara et al., 1993).
Historically, laboratory studies of larval competitionducted with different Asian strains of Ac. ulbopictiis andAc. aegypti showed Acdes ac^ypti better able to competethan Ae. albopictus (Macdonald, 1956; Gilotra et al., 1967;Moore and Fisher, 1969; Sucharit et al., 1978). Informationobtained by these researchers suggested that Ac. albopicluswould not become established in locales inhabited by Ae.
23

Aedesalbopictus
^ I \ 7T W / I ^\_i_ Clypeus __/ fr-
Figure 15. Differences in the clypcus ofAt’, albopictus andAe. aegypti. The female Ae. albopictus clypeus is covered only
with dark scales, while Ae. aegypti shows white scales.(Drawings provided by Dr. George O’Meara, Medical
Entomology Laboratory, University of Florida at Vero Beach.)
aegypli because of competitive displacement competitiveexclusion.’1The phenomenon of competitive displacement involving
Ac. ulhopiclus and other species has also been suggestedby other scientists. For instance, in cage experiments Gub-ler (l970b) demonstrated induced sterility when males ofAe. alhopkliis cross-inseminated females of Ae. polyne-sieiisis. Gublcr (1970c) suggested competitive displace-ment principle by showing experimentally that at ratio of10 Ae. albopictus males to Ae. polynesiensis male, cagecolony of Ar. polynesiensis completely eradicated.
Likewise, Ac. albopiclus is reported to have displaced Ae.guiimensis from of Guam and the Mariana Islands.Ae. guumeiisis is indigenous to Guam. bul Ae. albopiclus isknown to have been introduced in 1944. One study hypoth-esized that (in part due to competition) the population densityof Ae. guamensis decreased by much 95% in artificial
competitive exclusion principle, (iiiusc’s hypothesis, [h Hie prin-ciple Unit species 1ming identical ecological requirementsexist indefinitely.
containers and by 30% in natural habitats, while thai ofAe.albopiclus increased (Rozcboom and Bridges, 1972).
In North America, Ae. albopiclus has been foundciated with Ae. Iriserialus in tires and tree holes (NOMCB,1987; Livdahl and Willey, 1991; O’Meara et al.. 1993).Laboratory larval competition studies between the two spe-cies using tree hole and tire fluid suggest that they couldcoexist in tree holes, but that Ae. Iriserialus would becomelocally extinct in lire habitats (Livdahl and Willey, 1991).The mechanism proposed by the researchers lo explain thisresult is that "resources exist within tires that be used
effectively by Ac. albopiclus than Ae. Iriseriams, andthat tree hole fluid contains that each speciesploits differentially." In general, these laboratory-basedsumptions have been supported by field studies conductedin Florida which showed "progressive increase in thefrequency of artificial containers with Ac. albopiclus andconcomitant decrease in those with Ae. Iriserialus"(O’Meara el al., 1993).The apparent of Ae. albopictus competitor
to be corroborated by observations the relativeabundance of the other New World Stegomyia, Ae. aegypti,
24


Figure 17. Adult morphology ofAc. albopictus. A, dorsal aspect of the
male; B, lateral aspect of the male abdomen; C, lateral aspect of the female
head; D, lateral aspect of the male thorax; E, anterior surface of the malelegs. (Drawings provided by Walter Reed Biosystematics Unit,
Smithsonian Institution, Washington, D.C.)

in various southern cities of the United Stales in the 1980s,and the subsequent dramatic decrease in Ac. aegypti pop-ulation;; following the introduction of Ae. albopictus. Fieldobservations of artificial habitats, including tires, in Texas,Alabama, and Florida appear to reconfirm the phenomenon(Sprengcr and Wuithiranyagool, 1986; Hobbes et al., 1991;O’Meara et al., 1993).
Another hypothetical mechanism suggested for compet-itive replacement otAe. aegypti by Ae. albopictus in NorthAmerica is parasitism. For instance, the intestinal gregarineprotozoan Ascogregarina taiwanen’iis, which parasitizes Ac.albopictus larvae, appears to have to the New World
along with Ac. albopictus (Munstermann and Wesson,1990). After spores ingested by the larvae, the sporo-zoites penetrate and grow in the midgut cells, then emergeand go to the Malphigian tubes where their life cycle is
completed by sexual fusion of the gametes, cyst production,and spore excretion by the emerged adults. Ae. alhopictustolerates its parasite well; very heavy infections producemaximum larval mortality rate of 10%-20%. Experimentscarried out by G. Craig at the University of Notre Damehave demonstrated cross-infections of Asc. taiwanensis in
Ae. aegypti. However, in that species the infection settlesin the midgut and does not complete its sexual cycle. Moreimportant, heavy infections of Asc. taiwanensis have beenshown to produce high larval mortality in Ae. aegypti.Thus, it has been suggested that At’, albopiclus’ parasitegives it major competitive edge in the field, which in turn
may help to explain the decrease in Ae. aegypti populations.Edgcrly ct al. (1993) studied the inhibitory effects of lar-
the hatch rate of conspecific and congeneric eggspotential population regulatory mechanism. Their labo-
ratory experiments assessed the egg hatch rates of Ae. al-
hopictus in response to increases in the larval density of
two possible competitors, Ac. aegypti and Ae. friserialus.
The rationale behind the research approach is that after eggflooding, "microorganisms colonize the egg surfaces andthe subsequent decline in dissolved oxygen due to micro-
bial respiration triggers hatch." Therefore, larval feedingmicroorganisms could be simple mechanism for hatch
inhibition. "Foraging larvae of other Aedes species mayimpose interspecific, well intraspecific, hatch inhibi-
tion." Rggs of each species exposed to varyingbinations of larval species and density for 24 hours and
their respective hatch rates then assessed. Of the three
species, Aedes albopictus eggs showed the lowest level ofinhibition when exposed to high larval densities. In addi-
tion, al the lowest larval density, Ae. albopictus imposedthe most intense interspecific hatch inhibition. It hy-pothesized that the observed differential hatch responsesthe result of differences in larval feeding rates, respirationrates, and oxygen thresholds required for hatch, well
differential changes in larval grazing behavior with in-
creased density. The conclusions of the study suggest thatthe relative of "Ae. albopictus in inhibiting egg
hatch of Ae. triserialus and Ae. aegypti, combined with
their propensity to hatch into occupied habitats, may pro-vide the species with significant advantage theycounter resident populations." Edgerly and co-workers
(1993) concluded that the "competitive ability of Ae. al-
bopictus larvae may help to explain why Ae. albopictus hasachieved rapid dominance in domestic container habitats
previously occupied by Ae. triseriatus and Ae. aegypti inthe southern United States.’’
Further studies focusing adult mating behavior haveexamined mating interference (interspecific matings) to tryto explain the observed differences between Ae. ullwpicfusand Ae. aegypti. Interspecific matings between Ae. alho-
pictus males and Ae. aegypti females have been demon-strated in both field and laboratory experiments (Nasci et
al., 1989). Ae. aegypli females inseminated by Ae. albo-pictus males contained clumps of few dead sperm in their
spermatheca, which implies refractoriness to further matingby the female. In turn, the observations suggest mating in-terference at high Ae. albupiclus densities, which may be
important factor in determining the distribution of the
two Stegomyia in North America. The problem with this
likely explanation is that the experiments have not been
replicated either in cages in the field by other researchers.Additional approaches have been pursued to try to
plain the decline ofAe. aegypti and the role Ae. allmpiclushas played in that decline in North America, hut conclu-
sive general explanation has not yet emerged. More field
studies needed to the true impact of Ae. albo-
pictus Ae. aegypti populations- In general, researchers
agree that not enough evidence is available to provewhether the decline ofAe. aegypli and Ae. triseriatus hasbeen caused by Ae. alhopictus. In all likelihood, combi-
nation of several factors (including larval competition) maybe contributing to the decline of Ae. aegypti and otherAedes in North America.
Regarding the Asian situation, in which Ae. aegypti ap-pears to have displaced Ae. albopictus, lidgerly et ai.
(1993), in agreement with Hawley (1988), consider that"habitat changes have dramatic effects the outcomeof biological invasions; rampant urbanization in SoutheastAsia apparently stimulated increase in abundance in Ae.aegypti introduced from Africa, at the expense of nativeAe. ailwpictus.’’
Despite several likely explanations, both phenomena-the Asian and North American competitive Aedes replace-ments-still constitute ecological enigma.
DISEASE RELATIONSHIPS
The potential public health importance cifAe. alhopictusis indicated by its ability to transmit various arboviruses,filarial worms, and protozoa. Both field and laboratory stud-
27

ies indicate that this species is susceptible to infection with
and able to transmit pathogens of medical im-
portance. However, it is important to note that thus far Ae.
aibopiclus has not been implicated in the transmission of
diseases of public health importance in the Amcricas. Gen-eral information arboviral diseases and overview of
of the important of these diseases, well
their potential relationship to Ac. alboplctus, presentedbelow.
General Information on Arboviral Diseases
Arthropod-borne viruses (arboviruses) consist of groupof animal viruses that able to reproduce in arlhropodand be transmitted to vertebrate host. According to the
World Health Organization (1985), "arboviruses viruses
that maintained in nature principally, to importantextent, through biological transmission bchveen susceptiblevertebrate hosts by hematophagous arthropods throughtransovarial and possibly venereal transmission in arthro-
pods; the viruses multiply and produce viraemia in the
tebrates, multiply in the tissues of arthropods, and passedto vertebrates by the bites of arthropods after period
of extrinsic incubation." Vertical (transovarial) and venereal
transmission in arthropods may basic long-termmaintenance mechanisms for arboviruses.
DeFoliart et al. (1987) consider vertical transmission to
be survival mechanism developed by the virus to resist
adverse climatic conditions that may affect its arthropodhost. Vertical transmission involves "the direct transfer ofinfection from parent organism to his. her, its off-
spring." In contrast, horizontal biological transmission in-
volves ingcstion of the virus in the blood of vircmic
tebrate, multiplication of the virus in the midgut of Ihc
mosquito, dissemination of the virus to the hemocoel, in-
fection of the salivary glands, and transmission of the virus
to susceptible vertebrate host by bite (Turell, 1988). The
virus also be transmitted cither horizontally by arthro-
pods infected vertically, mechanically by arthropods that
obtain partial blood meal from infected host and then
continue their feeding susceptible (i.e., arthropodscontaminated flying syringes).
Sliroycr (1986) and Mitchell (1991) reviewed the vector
competence of Ae. albopictus, including North and South
American strains, to arbovirus infections. The term "vectorcompetence’’ used herein corresponds to the definition
cited by Mitchcll (1983): "the combined effect of all of the
physiological and ecological factors of vector, host, patho-gen, and environment that determine the vector status of
given arthropod population." laboratory studies have dem-onstrated that Ar. alhopiclus is competent vector of most
of the American encephalitis viruses. These include
bers of the family Togaviridac, genus Alphavirus (easternequine encephalitis [EEE], western equine encephalitis
[WEEJ, Venezuelan equine encephalitis [VEE], and Mayaro[MAY] viruses); the family Flaviviridae, genus Fliivivirus
(St. Louis encephalitis [SLE], yellow fever, and dengue vi-
ruses); and the family Bunyaviridac, genus Biinyavinif (LaCrossc encephalitis [LAC], trivittatus \TVr\. and James-town Canyon []C] viruses). Table summarizes the keyvector competence information available for Ae. albopiclitsin studies carried out primarily with strains from the Amer-icas. It also includes relevant information from stud-
ies conducted with strains from Hawaii Asia that pre-dated the discovery of Ae. albopiclus in the New World.
Aedes albopiclus has been shown to be susceptible to in-
fection with at least 26 different arboviruses and able to
transmit at least 24 of them. Information natural and
experimental infections of Ae. albopiclus and its potentialto transmit these viruses to humans and other vertebrates is
discussed below.
Arbovirus Field Isolations in the Americas
Four arboviruses have been isolated from Ae. albopicluscollected at various locations in the continental United
States. These Potosi (POT) virus from mosquitoes col-
lected in central Missouri in 1989, Tensaw (TEN) virus
from Texas in 1991, Keystone (KEY) virus from central
Florida in 1991, and EEE virus from central Florida in
1991. The role ofAe. ulbopicius in the natural transmission
cycles of these four viruses remains obscure. Of the four
isolations, probably the most disturbing is that of EEEvirus (Mitchell et a]., 1992).
Eastern Equine Encephalitis (EEE)
This RNA virus, belonging to the genus Alphavirus ofthe family Togaviridac, forms part of complex of enccph-alitidcs transmitted by mosquito species (Acha and Szyfres,
1987). In North America the virus is maintained in fresh-
water swamp habitats in cnzoolic cycle principally in-
volving Culiseta melanuru and variety of bird species
(Scott and Weaver, 1989). Various species of mosquitoesbelieved to be bridge vectors between this maintenance
cycle and susceptible mammalian hosts. In North America,
these vectors include Coquillettidia pcrlurhans and Aede’i
sollicitan’i. in the tropical countries of the Amcricas, the
vectors appear to be Culcx nigripalpiis, Culcx laeniopus,and Aedes taemorhynchus (Calisher et al., 1981).EEE is but disease of humans in
the Americas with overall case-fatality rate among clin-
ical of between 50% and 80% (Shopc, 1985a). The
disease in humans has sudden onset and is manifested byhigh fever, cephalalgia, conjunctivitis, nausea, vomiting,and lethargy, progressing rapidly to delirium and
Neurologic signs include neck stiffness, convulsions,
spasms of the muscles of the extremities, and altered
28

Hexes. A biphasic is in children, with fever,vomiting, and headaches for two days, followed first
by apparent recovery and then by rapidly fatal
cephalitis. Sequelae and in the
young. About 60% of the survivors show mental retardation
(Shope, 1985a; Acha and Szyfrcs. 1987). Sporadic humanof REE have been reported from Brazil, the Domin-
ican Republic, Jamaica, and Trinidad and Tobago. (WHO.1985). Elsewhere in the Americas, the virus has been iso-
lated in Argentina, Canada, Colombia, Cuba, Guatemala,Guyana, Haiti, Mexico, Panama, Peru, the United States,and Venezuela (Acha and Szyfres, 1987).
Although FEE disease in humans, it
primarily affects domestic mammals and exotic birds. Nu-epizootics have been reported among horses, with
high mortality rates. Such outbreaks may may not be
accompanied by clinical in the human population. Ex-otic birds that be affected by EEE include pheasants,
Table 1. Susceptibility ofAc. iilbopictus to oral infection with arboviruses, ability to transmit themby bite, and status in nature.
Virus
Chikungunya’"1’Dengue 1,2,3,4""
Oralinfeel.
Oraltrans.
Infectedin
NoYes
References
Tcsh al., 1976; Turcll ill., 19921)
Roscn ill., 1985; Milchcll al..
EEF.-
Jamestown Canyon’’JE-Keystone1’La Crosse1’Mayaro1’Nodamnra’’Oropouctn^Oningo’’
Polosi"Ril’l Valley lever"Ross River’’’
Sinilbis"
San Angelo’’SLE"
Tensaw*Triviiiatus1’West Nile"WEE"
VEE IAB"Yellow fcvel""
Tcsh al., 1976; Turoll al., 19921)
Roscn al., 1985; Milchcll al..
1987Yes Mitchcll al., 1992; Turcll al.,
1994No Grimslad al., 1989Yes Roscn al., 1978Yes Milchcll al.. 1992No Sircit and Grimslad, 1990No Smith and Prancy, 1991No Tesh. 1980No Smith and Franci, 1991No Kay al., 1982; Tomori and Aitken,
cited in Shroyer, 1986Yes Francy ct al., 1990aNo Turcll al.. 1988No Mitchcll and Guhlcr, 1987; Milchcll
al., 1987No Dohm and Turcll, unpublished data,
1993No Tcsh. 1980No Milarrmra al., 1940, cited in
Shroyer, 1986; Hardy al.. 1980Yes Mitchcll al., 1992No Grimslad al., )989
No Akiiler al., 1982No Kramcr al., unpublished
information, cilcd in Milchcll.1991; Simmons al.. died in
Shroyer, 1986No Tnrcll al.. 1992aNo Mitchcll al., 1987; Miller and
Ballinger. 1988
Studies conducted with Ac. uibopictus strains Irom Hawaii and/or Asia.
Studies conducted with {ilbopH’tna strains from United Slates and/or Brazil.
EEE equine encephalitis virus
JK Japanese encephalitis virus
SI.E Sl. I-ouis encephalitis \irus
WEE Western equine encephalitis virus,
VEP. Venezuelan equine encephalitisSwim’: taken from Shroyer (1986). Milchell (1991). and updaled information published o! Juh 1993.
Special emphasis is given studies carried with strains from Americas.
29

quails, peking ducks, emus, and partridges. In out-
breaks in the United States attack rates of up to 50% havebeen recorded, producing serious economic losses (WHO,1985).
Fourteen strains of EEE virus isolated from Ac’.albopictus collected during the of 1991 in tire
dump in Polk County, Florida (Mitchell et al., 1992). This
the first reported isolation of arbovirus of known
public health importance from wild-caught Ae. albopictusin the United States. In June 1992, after active participationby the local Mosquito Abatement District and destructionand removal of the tire dump, several research agenciesfailed to EEE virus fromA<?. albopictus pools fromthe
Several studies have examined the potential for Ae. al-hopiclu\’ to transmit EEE virus. A strain ofAe, albopictusfrom Houston, Texas, shown to be highly susceptibleto infection after feeding viremic chick; 100% (n10) and 40% (n 20) transmitted virus by bite (Scott el
al., 1990). Turcll et al. (1994) examined mosquitoes fromthe tire pile in Polk County, Florida, for their susceptibilityto strain of EEE virus recovered from Ae. alhopictus.Again, these mosquitoes highly susceptible to oral in-fection [98% (n 360)], and 40% (n 216) transmittedvirus by bite. These Ae. albopiclus significantlysusceptible to infection with EEE virus than Ae. sol-licitans Ae. iaeniorhynchus, known bridge vectors ofEEE virus (Turell, unpublished data). Although neitherScott et al. (1990) Turell et al. (1994) observed anyevidence of vertical transmission (2,952 progenyined), transovarial transmission may still play role in the
long-term maintenance of this virus in nature. To date, the
public health significance of the EEE field isolations fromAe. albopicius remains unclear. However, because of the
high sensitivity of this species to EEE viral infection, its
opportunistic feeding behavior, and its continued expansionwithin the Americas, the natural transmission cycles ofEEEvirus may be modified by the inclusion ofAe. albopictus.
Potosi (POT)
Ten strains of POT, newly recognized virus,covered from Ac. albopicius collected in the of1989 in the United States (Francy et al., 1990a). The iso-
lates determined to belong to the family Bunyaviridae,genus Bunyavirns, and the Bunyamwera serogroup. Theseisolations made from pools ofA<?. ulbopiclus capturedin Potosi, Missouri; the minimal field infection rate in Sep-tember 1989 8.9 per 1,000 mosquitoes. Six additionalisolations from the made during the
collecting period (Mitchell et al., 1990). Attempts to isolate
additional strains during the of 1990 and 1991unsuccessful.
Studies of vertebrate susceptibility to POT virus
carried out with suckling mice, hamsters, rabbits, and pi-geons. This virus is pathogenic for suckling mice. 85%(n 59) became symptomatic after intracercbral inoculation
of POT virus (Niebylski, 1992). The range of symptomsdetected in mice included loss of coordination, tremor andprostration, and limb paralysis- Overall mortality 62%,with average survival time of 8.8 days. Hamsters andrabbits developed low levels of viremia (2.0 login median
tissue culture infective doses fTCID^]/ml to 4.3 logicTCID,(i/ml of whole blood) with sign of illness. Neither
viremia signs of illness detected in pigeons In-oculated with POT virus (Niebylski, 1992). Transmission
studies indicate that Ac. albopicius transmit POT virus
horizontally (Heard ct al., 1991; Mitchell ct al., 1990) but
not vertically (Francy et al., 1990a). Preliminary testing ofpeople working at the sampling sites demonstrated the pres-
of neutralizing antibody to the virus In in-
dividual. However, the public health threat posed by POTvirus has not been fully resolved, and additional informa-
tion human pathology is needed.
Tensaw (TEN) and Keystone (KEY)
TEN virus isolated from pool of 38 Ae. albopictuscollected 25 July 1991 in Montgomery County, Texas(Mitchell et al., 1992). This virus belongs to the familyBunyaviridae, genus Bunyavirus, and isolated for thefirst time in 1960, from Anopheles crucians collected in
Baldwin County, Alabama (Coleman, 1969). The virus pro-duces clinical disease and death in suckling and adult micebut not in rabbits, guinea pigs, hamsters. Antibodies havebeen detected in dogs, raccoons, cattle, and humans (Cham-berlain et al.. 1969). The importance of TEN virus
public health threat remains unknown (Coleman, 1969).KEY virus recovered from pool ofA<:’. albopicius
collected during the Florida studies that recoveredEEE virus from this species. The virus belongs to the fam-
ily Bunyaviridae, genus Runyavirus, and has been isolated
from cotton rats (Sigmodon hispidus) in Georgia and Flor-ida (Sudia et al., 197 la). Aedes allunlicus and/or tormentor
appears to be the principal vector, although KEY virus hasbeen isolated from nine other mosquito species. Antibodieshave been detected in rice rats, cottontail rabbits, graysquirrels, raccoons, white-tailed deer, sika deer, horses, and
humans (Sudia et al., 1971a; Watts et al., 1982).Although the impact of TEN and KEY viruses human
health remains unknown, the isolation of these viruses fromAe. albopicius and its ability to transmit them raises the
important question of how the presence of vector
may affect natural transmission cycles. As stated by Francyet al. (1990a), "the possibility remains that other viruses,which heretofore have not infected humans to any appre-ciable degree because of nonhuman feeding habits of their
30

arthropods vectors, may be transmitted to humans by Ac.albopictus result of the aggressive feeding o( this spe-cies humans well other mammalian hosts.’’
Laboratory Vector Competence Studies of SomeImportant Arboviruses from the Americas
Dengue (DEN)
DEN viruses consist of antigenic subgroup of four
closely related, but antigcnically distinct, viruses designatedDEN-1, -2, -3, and -4. which belong to the genus Flavivi-
rus, family Flaviviridac (Westaway et al., 1985).The classical form of dengue is characterized by acute
febrile illness with headache and joint and muscle pains(WHO, 1986a). Epidemics of dengue annually affect mil-lions oF people in tropical of the world. Dengue hem-
orrhagic fever/dengue shock syndrome (DHF/DSS) is
form primarily in children, although it affect
adults well. It is caused by the DEN virus clas-sical dengue. However, DHF/DSS may have immuno-
pathologic basis, and the case-fatality rate is 5% to 10%(WHO. 1986a). DHF is major of morbidity and
mortality among children in many countries of Southeast
Asia, and recently its importance has increased in the
Americas (Halstead, 1980; Monath, 1986; Kouri ct al.,1989; I’AHO. 1992).
In the Americas, dengue worsened public health
problem during the 1980s. Between 1980 and 1990than million dengue reported to PAHO. The
majority of these caused by serotypes 1, 2, and
4, will few imported related to DEN-3. Duringthose years, increase in of DHF/DSS observedin various countries of the continent. Thus far, the most
important outbreak of DHF/DSS in Cuba in 1981,when than 300,000 dengue cases, 10,312 DHF/DSScases, and 158 deaths recorded (Kouri et al,, 1989).A second major outbreak occurred in Venezuela from Oc-tober 19S9 to January 1990, with 5,990 and 70 deaths
(PAHO, 1992). Other countries in the Americas that have
reported DHF Aruba, Brazil, Colombia, the Dominican
Republic, El Salvador, French Guiana, Honduras, Mexico,
Nicaragua, Puerto Rico, Saint I.ucia, Surinamc, and the Vir-
gin Islands (Institut Pasteur dc Guyane, 1991; PAHO,1992).The ability of Ae. albopictus to transmit DEN virus
first shown in studies involving human volunteers early192fi (Siler et al., 1926). More recently, Ae. ulbopiftusshown to be able to transmit all four DEN serotypes
both horizontally (Gubler and Roscn, 1976) and vertically(Roscn et al., 1983). The species also found to be
susceptible to oral infection with all four DEN serotypesthan Ae. aegypli (Jumali et ai., 1979; Rosen ct al.,
1985). Naturally infected Ac. albopicliis collected inAsia by Rudnick and Chan (1965).
Various studies have found Ae. albopictus to be associ-ated with epidemics of dengue fever (Chan. Y. C., et al.,1971; Jumali et al., 1979; Fan et al.. 1989). However, be-
Ae. albopictus often overlaps in distribution with Ae.aegypti, another known vector of DEN viruses, it is ol’len
difficult to determine the relative contribution of the two
species to disease transmission. During massive dengue ep-idemics in the Seychelles Islands and in Guandong, China,where Ae. aegypti absent, Ae. albopictus incrimi-nated the principal vector (Mctsclaar et al., 1980; Oiu el
al., 1981). Thus, experimental and natural association of the
four DEN viruses with Ac. albopiclus has been documentedin several instances.
In the Americas, geographical strains of Ac. albopiclusfrom Brazil (Cariacica City) and the United States (Hous-ton) competent laboratory vectors of all four DBNrotypes (Mitchcll. 1991). In addition, both North and South
American strains of Ae. albopictus transmit all fourDEN serotypes vertically (Mitchell and Miller, 1990; Bosio
ct al., 1992). Thus, Ae. albopiclus may impor-tant maintenance vector of DEN viruses in endemic areas,and endemic foci may be initiated by importation of
vertically infected eggs (Gubler, 1987; Shroyer, 1990).
Yellow Fever (YF)
YF virus is member of the genus Flavivirus, family Fla-
viviridae- This viral disease is endemic in tropical regions ofboth the Americas and Africa. In the Americas, the endemic
for Yl’’ include the Amazon and Orinoco-Magdalenabasins in Bolivia, Brazil, Colombia, Ecuador, French Guiana,Peru, and Venezuela (Acha and Szyfrcs, 1987). The disease
may range from subclinical to fatal. Depending the
verity, the signs and symptoms of yellow fever include fe-
ver, headache, nausea, epistaxis, albummuria, and bodypain. In cases, the disease is characterized by oli-
guria, jaundice, and hcmorrhagic manitcslations that in-
clude epistaxis, black vomitus, mclena. and uterinehemorrhage (WHO, 1985).
Dingcr et al, (1929) reported that Ae. albopictus femalestransmitted YF virus to monkeys in laboratory experiments.However, infection and transmission rates not
determined.Monath (1986) suggested that, because of the ecological
adaptability of Ac. albopictus, this species could bridge the
gap between the jungle and urban YF viral transmission
cycles. This scenario would favor the spread of YF in vil-
lages and towns, particularly in endemic of South
America. Ae. alhopiclus could fill ecological niche anal-
ogous to that of Ae. simpsoni, major bridging and epi-demic vector of YF in East Africa. These assumptions have
been reinforced by experimental studies indicating that both
31

the U.S. (Houston) and Brazilian (Cariacica, Espfrito Santo
State) strains of Ac. ulbopictus susceptible lo oral in-
fection with YF virus (Mitchcll el al., 1987; Miller and
Ballingcr, 1988; Miller el al., 1989). However, the North
American strains significantly efficient vectors
than their southern counterparts.
Venezuelan Equine Encephalitis (VEE)
VEE virus is member of the genus Alplwvirus, family
Togaviridae. Based the kinetics of the hcmagglutination-inhibition test, Young and Johnson (1969) first established
classification for VEE virus consisting of complexgroup divided into four subtypes (I-IV). Subsequently, six
antigenically related virus subtypes numbered through VIdefined (Walton and Grayson, 1989; Calishcr, 1994).
Also recognized the varieties IAB, 1C, ID, III, and IFwithin subtype I, well the varieties II1A, IIIB, and
IIIC. for total of eight different variants (Calisher, 1994).Among the five antigenic variants of subtype I, two (IABand 1C) virulent to horses and responsible for
epidemics/epizootics, while the other members of the VEE
complex generally considered cnzootic viruses (Cal-ishcr, 1994). The VEE complex is native to the Americas
and is endemic in Central America and northern South
America well Mexico, Trinidad and Tobago, and the
state of Florida in the United States (WHO, 1985).VEE in humans is usually respiratory illness with fe-
ver, headache, and muscle pain similar to self-limiting
influenza-like febrile illness. However, infections
develop into serious encephalitis (Shope, 1985a; Acha
and Szyfrcs, 1987). In most cases, the onset of the disease
is accompanied by malaise, chills, ccphalalgia, and, fre-
quently, vomiting, and diarrhea. The illness may
resolve after three to five days unless encephalitisPeripheral neurological symptoms, such flaccid spastic
paralysis and alterations in reflexes similar to the neurolog-
ical symptoms of other arbovirus encephalitides, typicalof VEE encephalitis (Acha and Szyfres, 1987). Encephalitisis frequent in children than in adults. Mortality rates
low but may reach 3.6% in with deficient medical
services (Shope, )985a). Sequelae, including behavioral
changes and learning defects, also Horses may de-
velop rapid fulminating illness with sudden death
encephalitic syndrome. During the period 1967-1971.
100,000 equine deaths due to VEE, well hundreds of
thousands of human infections, reported from Central
America and the United States (WHO, 1985).Experimental studies indicate that both North and South
American strains of Ae. alhopiclus competent vectors
of VEE virus (Bcaman and Turell, 1991; Turcll and Bca-
man, 1992). However, susceptibility to infection and rates
of transmission varied significantly among the five strains
tested. Transmission rates higher for South American
strains (24%) than for U.S. strains from Indiana Texas
(5%) (Beaman and Turcll, 1991); however, strain from
New Orleans (Gcntilly), Louisiana, transmitted VEE virus
most efficiently (45%) (Turcll and Bcaman, 1992). The
transmission rate observed for the New Orleans strain
comparable to higher than those reported for Pso-
rophora confimis (33%) and Aedes soUicilam (38%), two
of the natural vectors of epizootic VEE (Sudia el al., 1971b;
Turcll et al., 1992a). Turcll and co-workers (1992a) spec-ulate that the observed differences in strain susceptibilitymay be the result of either founder effect separateintroductions in the of origin of the strains. The ef-
ficient transmission of VEE virus by of the As. al-
hopiclus strains presently found in the Americas suggests
that the presence of this species has increased the risk of
transmission of VEE.
La Crosse (LAC)
LAC virus is member of the genus Runyavirus of the
family Bunyaviridae and belongs to the California
group. Isolation of LAC virus has been reported only from
North America, and LAC is endemic throughout large por-tions of the midwestcrn United Stales. Infection with LAC
virus encephalitis, particularly in children and
adolescents under 15 years of age. The endemic in-
cludes Minnesota, Wisconsin, Illinois. Indiana, Ohio, West
Virginia, and New York. Although fewer than 100
usually reported each year, it is likely that many infections
with LAC virus incorrectly diagnosed of mild,
undift’crcntiated fever of unknown origin. Infection with
LAC virus may result in illness with high fever, with
without generalized convulsions. Vomiting, focal seizures,
paralysis, abnormal reflexes, and mental manifestations
(Shope, 1985b), and neurological sequelaepersist for years after the acute illness. The case-fatality rate
is less than 0.5%.Various species of mammals, especially rodents,
ceptible to infections by LAC virus, but it is believed that
these infections generally asymptomatic (Acha and
Szytres, 1987). The tree hole mosquito Aedes Iriserialiis is
considered to be the principal vector of LAC virus to both
amplifying rodent hosts and humans. The main vertebrate
hosts of LAC appear to be the eastern chipmunk (Tamiusstnalus), gray squirrel {Sciurus caroUnensis), and red fox
(Viilpes ftilva) (Calishcr, 1994).The introduction ofAe. alhopictus into cnzootic for
LAC has led to various studies of the vector potential of
this species for LAC virus (Tcsh and Gubler, 1975; Grim-
stad et al., 1989; Strcit and Grimstad, 1990; Cully et al.,1991; Cully ct al., 1992). Ac. alhopicliis is highly suscep-tible to oral infection with LAC virus, nearly all (1,633
>99%) of 1,637 Ae. alhopicltis contained LAC viruswhen tested 14 days after ingcstion of blood-LAC virus
32

suspension containing virus tilers simulating doses thatwould be encountered in nature (3.5-4.5 logs). These stud-ies included 27 strains of Ac. albopiclun, 14 from locationswithin the continental United States, 5 from Brazil. and therest from temperate Asia, tropical Asia, and Hawaii (Streitand Grimstad, 1990). Moreover, 97% of the 1,637head-positive for the virus, suggesting dissemination ofLAC virus beyond midgut barriers and throughout the he-mocoel. All 43 Ac’. Iriscriatus fed the blood-virussuspension also had disseminated LAC virus infections(Streit and Grimstad, 1990). Other experiments conductedwith LAC-viremic eastern chipmunks (T. siriatus) suggestthat the infection threshold otAe. albopictus is lower thanthat of Ae. triserialus (Streit, pers. 1993). Morethan 30% of fully engorged Ae. alhopicluf, became infected,compa-’ed to less than 109& of similarly exposed Ac’, iriser-iatus. after feeding chipmunk with LAC virus titerof 3,600 viral plaque-forming units (PFU)/ml blood (84hour postinfection). In transmission experiments conductedafter 14 days of extrinsic incubation, 43% ofAc’, albopictustransmitted virus to suckling mice (Streit and Grimstad,1990). Ae. albopictus infected either orally transovariallytransmitted LAC virus to susceptible eastern chipmunks(Cully cl al., 1992). As with other arboviruses, there
significant variation in transmission rates (18%-75%)among the various Ae. albopictus strains tested. Brazilianand tropical Asian strains tended to transmit LAC virus
efficiently than did North American and temperateAsian strains (Streit and Grimstad, 1990).
Ae. albopictus also transmit LAC virus transovarially(Streit and Grimsfad, 1990; Cully ct al., 1992). Minimumfilial i’.ifeclion rates range between 1% (Teresa Brazilian
strain) and 4.5% (Oahu strain). In recent study, filial in-fection rates ranged from 3%-22% (average 10%) for prog-eny of individual LAC virus-infected females (Streit andGrimslad, 1990). Streit (1994) found that transovarial trans-mission of LAC increased rapidly with selection. Afterthree generations, he established LAC-disseminated line
which continued 90%-r positive for 10 generations. Inother line he established, transovarial transmissiontinued for 10 generations among mosquitoes thatceived blood (i.e., completely autogenous). Thisability opens remarkable possibilities for the maintenanceof reservoir. In economically depressed in EastSt. Louis, Illinois, 50% of wild-caught Ac. albopiclus hadfed urban rats. Antibodies to LAC found in rats
and opossums. Streit (1994) suggests the possibility ofu’ban cycle for LAC.
The fact that Ae. albopictus becomes infected and trans-mits LAC virus at rates similar to those of the natural vector
(Ae. triseriatus), along with its establishment in whereLAC virus is endemic and its general feeding behavior, in-dicates that Ac. allwpicius may become involved in the nat-
ural LAC virus cycle and secondary vector in
where LAC virus is active.
Mayaro (MAY) and Oropouche (ORO)
MAY virus is member of the genus Alphaviru^, familyTogaviridae. Its activity has been documented only in theAmcricas, where it has been isolated in Bolivia, Brazil,Panama, Suriname, Trinidad, and the United States (Pin-heiro and LcDuc, 1989). In endemic areas, such the Am-
regions of Bolivia and Brazil, 10%-15% of residentshave antibodies to MAY virus (Pinheiro and LcDuc, 1989).The onset of the illness is abrupt, with fever, headache,epigastric pain, backache, arthralgia, photophobia, dizzi-ness, and chills (Pinheiro et al.. 1981; Pinheiro and LcDuc.1989). Humans appear to be accidental hosts that becomeinfected in forest where MAY virus circulates amongwild vertebrates and mosquitoes. MAY virus has been
isolated from several species of mosquitoes, especiallyHaemagogiis spp., although it has also been recovered fromCulex, Sahethes, and Aedes spp.A strain of Ac’, albopictus from Sao Paulo, Brazil,
competent laboratory vector for MAY virus, with infectionrates of 9%-85% and transmission rates of 45%-50%(Smith and Francy, 1991).
It concluded that "with large Ae. albopictus popu-lations and extensive MAY virus amplification, this speciescould secondary vector during outbreaks andperhaps bridging vector for MAY virus between theforest habitats and settled areas" (Smith and Francy.1991). The danger that Ae. alhopiclus will become in-
volved in the transmission cycle of this virus is increased
by the recently reported infestations of Ac. albopictus inthe state of Maranhao, which is contiguous with the stateof Para, where past outbreaks of MAY have been reported(Pinheiro et al., 1981).OHO is member of the Simbu serogroup of the genus
Runyavirus, family Bunyaviridae (Karabatsos, 1985). In-fection with ORO virus has been recognized major
of human febrile illness in the Amazonian region ofBrazil. Between the years 1962 and 1980, several outbreaksoccurred in Para State which affected at least 165,000 per-
in various urban centers (Pinheiro et al., 1982). Al-
though deaths have been attributed to infection with
ORO virus, many patients became severely ill and requiredprolonged convalescence (Shope, 1985b). The disease is
manifested by sudden onset of fever, chills, and malaiseand is marked by myalgia, dizziness, headache, photopho-bia, and sometimes mild epigastric pain, nausea, andvomiting.
Studies carried out with Ae. ulbopictus strain fromSao Paulo, Brazil, suggest that this species may not be
competent vector of ORO virus (Smith and Francy, 1991).Infection rates 2%-12%, and mosquito of thisstrain transmitted the virus by bite. This is not surprising,since Culicoides paraensis is considered the natural vector
(Pinheiro et al., 1982). However, although the risk thatAe.albopicfus will become involved in ORO virus transmission
33

appears low based the results of Smith and Francy(1991), other strains of Ac’, albopictus might be effi-
cient vectors of the virus.
Arboviruses Common in Areas Outside theAmericas
Ae. albopicius is competent vector of several viruses
not endemic to the Americas. These include Japanesecephalitis (JE), chikungunya (CHIK), Ross River (RR),Sindbis (SIN), and West Nile (WN) viruses. In addition,Ac. albopictus transmit JE virus (ransovarially, raisingthe possibility that JE virus could be introduced into the
Americas via infected eggs. Information JE, SIN, CHIK.and RR viruses is summarized below.
Japanese Encephalitis (JE)
JE virus is member of the West Nile antigenic complexof the genus Flavivirus, family Flaviviridae. Monath (1985)considers the disease it to be the most important ofthe arboviral encephalitides in terms of morbidity and
tality. Epidemics of JE seasonally in temperateof Asia and in the northern part of tropical Southeast Asia.
Illness may be manifested febrile headache syndrome,aseptic meningitis, encephalitis. When neurological dis-
orders appear, they usually accompanied by fever,chills, anorexia, nausea, vomiting, dizziness, and drowsi-
Case-fatality rates during epidemics range from 20%to 50%, but these high rates reflect recognition of only themost
In the laboratory, Ae. albopictus is susceptible to infec-
tion with JE virus and is able to transmit virus vertically to
its F, progeny after cither intrathoracic inoculation in-
gestion of virus-sucrosc-crythrocytc mixture (Rosen et al.,1978). As adults, at least five of the infected F, female Ac’.albopictus transmitted JE virus to chicks which they fed.
Moreover, Huang (1982) reported that strains of the
JE virus isolated from field-collected Ac’, albopictuslarvae from Fukien, China.
Sindbis (SIN)
S]N virus is member of the western equine encephalitiscomplex of the genus Alphavirus, family Togaviridae, and
dengue-like disease in humans. Clinical symptomsinclude fever, rash. headache, general malaise, and arthral-
gia (Niklasson, 1989). Although fatal have been
reported, arthralgia may be enough to require hos-
pitalization and may persist for up to three years. SINin Europe, Africa, Asia, and Australia- Various Culex
spp. have been implicated potential vectors, and the nat-
ural transmission cycle is believed to be between Culex
mosquitoes and birds, with various Acdes spp- actingbridge vectors that infect humans.
Laboratory studies indicate thai Ae. ulbopicliis is at least
efficient vector of SIN virus is Cx. pipiens, known
vector (D. Dohm and M. Turell, unpublished data). SINvirus has not yet been recovered from wild specimens ofAe. albopicius.
Chikungunya (CHIK.)
CHIK virus is also member of the genus Alphavirus.family Togaviridae. and dengue-like disease in hu-
This disease has abrupt onset, with high fever,myalgia, and sudden intense pain in joints(Shope, 1985a). Outbreaks of chikungunya fever have been
recognized in Angola, Congo, Nigeria, Senegal, and Zim-babwe in Africa and in Cambodia, India, Malaysia, the
Philippines, and Thailand in Asia (Karabatsos, 1985); theseoutbreaks affected tens of thousands of people.
Ae. albopicius adults highly susceptible to infection
with CHIK virus in laboratory experiments and also
capable of transmitting the virus by bite (Mangiafico, 1971;Yamanishi et al., 1983; Turell et al., 1992a). High infectionand transmission rates for Ae. albupictus (100% and 69%,respectively) readily testify to the importance of this
quito potential vector of CHIK virus (Mangiafico,1971). Furthermore, Turell and colleagues (1992b) dem-onstrated high infection rates (range, 72%-97%) in eightAe. albopictus strains from different of Asia, Brazil,and the United States after the mosquitoes fed vircmic
rhesus monkey. Disscminalion rates (36%-80%) observedin the experiments higher than those found in Ae.aegypti, the natural vector of CHIK virus in Southeast Asia.
Transmission rates not calculated. The virus has not
yet been recovered from wild specimens ofAe. allwpicius.
Ross River (RR)
RR virus is member of the genus Alphavirus and the
etiologic agent of epidemic polyarthritis (EPA) (Shope,1985a; MitcheH and Gublcr, 1987). Individuals affected bythe disease complain of pain, swelling, and limitation of
movement in the joints of the hands and ankles. Patients
may develop maculopapular rash the trunk andtremities. Other symptoms include throat, enlargedlymph nodes, and tender palms and soles (Shope, 1985a).Until the late 1970s, reports of epidemics of EPA in-
itially received only from Australia, the Solomon Islands,and New Guinea. In 1979 major outbreak of EPAcurred in the Western Pacific region, affecting than
50,000 individuals in Fiji, the Samoan Islands, the Cook
Islands, and parts of Melanesia (see references in Mitchell
and Gubler, 1987). Antibodies for the virus have been
found in domestic animals such dogs, sheep, pigs,
34

horses, and cattle. Antibodies also have been found in wildmammals such kangaroos, wallabies, and rodents(Shope, 1985a). The most important vectors of RR virus
appear to be Culex annulirostris, Aedes vigilax, and Aedespolynesiensis (Kay et al., 1982).
Studies the vector competence of several geographicalstrains of Ae. alhopicius from Houston, Hawaii, and Asia
showed the species to be efficient experimental vector
for RR virus (Mitchell and Gubler, 1987; Mitchell et al.,1987). Variation in vector competence among the geograph-ical strains also detected in the trials. Infection rates
approached 100% following ingestion of high-titer bloodmeals from golden Syrian hamsters (6.3 log;,, PFU/ml-7.1logio PFU/ml). Strains from Sri Lanka and Singapore dem-onstrated less susceptibility to infection (53%-80%). Trans-mission rates also high in the Houston and Hawaiistrains (52% and 77%, respectively) by day 14. Both strains
able to transmit RR virus to suckling mice by day 7postinfcction. When larvae from the second oviposition of
infected female parent tested, they failed to showproof of vertical transmission. Ross River virus has not yetbeen recovered from field-collected Ae. albopiclu.’s.
Transovarial Transmission
In addition to high susceptibility to oral infection, Ae.albopictus is able to transmit viruses to its prog-eny tran?.ovarially. Strains of Ae. albopictus from Hawaii
Asia known to be able to transmit vertically 15boviruse?;: DEN-1, DEN-2, DEN-3, DEN-4, Banzi, Bus-suquara, Ilheus, Kokobera, Kuirjin, JE, SLE, and Uganda Sviruses in the family Flaviviridae; and KEY, LAC, and SanAngelo viruses in the family Bunyaviridae (Tesh, 1980;Shroyer, 1986). Stabilized infections of San Angelo virus
in Ae. albopicius, and very efficient vertical trans-
mission takes place for up to 38 consecutive generations(Tesh and Shroycr, 1980; Shroyer, 1986). In the laboratory,Ae. albopicius strains from the Americas have verticallytransmitled DEN-1, DEN-4, YF, POT, and LAC viruses
(Mitchell, 1991; Streit and Grimstad, 1990).
Other Arboviral Studies
Shroyer (1986) lists arbovimses that able toreplicate in Ae. albopicius specimens following intrathora-cic inoculation- However, there is informationwhether Ae. albopicius be orally infected with thesearbovin’ses and subsequently transmit them by bite. Theseviruses include the phleboviruses Arumowat, Bujaru, Chi-
libre, Tcoaraci, Itaporanga, Karimabad (Tesh, 1975), and
Rift Valley virus (Turell et al., 1988), and the rhabdoviruses
Chandipura, Gray Lodge, Joinjakaka, Piry, Sigma, and
vesicuh.r stomatitis (Rosen, 1980).
Protozoan and Filarial Diseases
Filarial Worms
Field-collected specimens ot’A<?. albopicius from Sangla-buri, Thailand, found to be infected with Dirofilariaspp. (Harinasuta et al., 1970). Ae. albopictus is considered
important vector of Dirofilaria immitis, the causative
agent of dog heartworm in Japan and the United States(Mori and Wada, 1977; Scoles and Craig, 1993). Ae. al-
hopictus populations from the United States have marked
genetic variability in terms of susceptibility to Dirofilariaimmiiis. Work carried out in North Carolina found that Ae.albopictus entirely refractory to infection with D. im-
mitis (Apperson et al., 1989). However, recent workconducted at the University of Notre Dame showed that
40% of Ae. albopictus population from New Orleans
developed infectious L3 larvae: the results for other U.S.populations intermediate (Scoles and Craig, 1993).Several laboratory experiments also have demonstrated that
Ae. albopicius be infected with D. immitis (Galliard andNgu, 1938; Stojanovich and Scott, 1965; Konishi. 1989).
However, Ae. albopictus is not vector of human fila-
riasis caused by Hrugia Wuchereria spp. (Sasa, 1976).The larval tail to develop in the thoracic muscles.Roscn et al. (1976) justified their release of Ac. albopictus
small Pacific atoll attempt to replace vector offilariasis (Ae. poiynesiensis) with nonvector. Unfortu-nately, Ac. albopicius failed to persist the atoll.
Avion Malarias
Avian malarias have been experimentally transmitted byAe. aihopictus. The species became infected with and trans-
mitted Plasmodium lophurae beginning 12 days after feed-ing infected duck (Laird, 1941). Likewise, Ae. al-
Iwpictus is susceptible to infection with Pla^modiumgallinaceum; female mosquitoes developed sporozoites in
the salivary glands and gut 15 days after infectious bloodmeal (Russell and Menon, 1942). These mosquitoes alsotransmitted P. gallinaceum to normal fowls. The ability of
Ae. albopictus to harbor P. gallmaceum has made this
quito useful model for malaria studies (Hu ct al- 1989).
DISSEMINATION AND CONTROL
Mode of Dissemination and Origin
Since the discovery of Ac. alhopicfus in the Ainericas in1985 and its rapid spread to than 300 counties in 23states of the continental United States and 673 municipal-ities in 7 states in Brazil (C. Moore, pcrs. comm., Septem-ber 1994; J. Mangabeira da Silva, pers. comm., April 1994),
35

questions have arisen to how the mosquito intro-duced in the first place and how it is spread to
The spread of Ac. albopicfus is widely believed to beassociated with the massive movement of used tires aroundthe world (Rai, 1991). There is compelling evidence thatused tires imported in containers from Asia and holdinglarvae and of Ac. albopictus responsible for its in-troduction into the United States (Monath, 1986; Craven el
al., 1988; Francy et al., 1990b). After the Houston experi-ence, in which tires found to be the main breeding
of Ac. albopictus (Sprenger and Wuithiranyagool.1986), surveys carried out in other states have focusedtire yards.The recent infestations of Ac. aihopictus reported in
towns along the northern border of Mexico in the states of
Coahuila and Tamaulipas may also be due in part to themovement of used (ires and other artificial containers. It is
important to note that the officially reported figures thenumber of used tires exported from the United States to
Mexico have been increasing steadily since the 1980s. Fur-thermore, Mexico is the principal customer for U.S. used
tires in the Western Hemisphere, having bought 173,708units in 1991 alone (U.S. Department of Commerce, Bureauof the Census, National Trade Data Bank, CN TRADEXA4012200000-Used pneumatic tires of rubber). In addi-
tion, it is well known that many trucks transport scrap tiresfrom infested of the United States throughout Mexico,which clearly favors the spread of the infestation. Thus. it
is uncertain whether the Mexican infestations restrictedto those bordering the state of Texas whether the
species has already become well established in the country.In the four countries in the Americas known to harbor
the species-Brazil, the Dominican Republic, Mexico, andthe United States-Ac. albopictus has been reported breed-ing in tires, well in artificial and natural water-filledcontainers. Regarding the issue of used tire exports.O’Meara and co-workers (1993) pointed out that "there is
major threat of exporting Ac. allwplctus from the UnitedStates to countries that currently free of this mosquito.Used tires being shipped from Florida and other GulfCoast states, without adequate inspections for mosquitoes.to destinations in the Caribbean region and Central andSouth America." Introduction of Ae. albopictus to addi-tional countries in the Americas would appear to be im-
minent may have already occurred owing to such factorsthe rapid expansion of and traffic by land, air,
and Moreover, since adequate entomological surveil-
lance and vector control activities lacking in most ofthe countries of the Americas, the spread is likely tocontinue.
In the state of Florida, three other specific factors besidesthe movement of containers appear to be contributing to the
spread of the species. These factors the widespreadavailability of suitable aquatic habitats, the decline of Ac.aegypli populations, and the rapid adaptation of Ac. albo-
pictus to shorter photoperiods (O’Meara et al., 1993).Moreover, since 1986, in many in Louisiana Ae. al-
bopictus has shown that it spread from tire piles into
tree holes (NOMCB, 1987).The appearance of the species in the Dominican Republic
raises several iniiinswered questions. The infestation maybe strain of temperate origin (from either Asia conti-
nental America) that originally had photoperiodicsponse but progressively lost it result of genetic selec-
tion for nonphoroperiodic trait. Another possible scenariois that this infestation is independent of others in the Amer-icas. having been brought inadvertently in shipping cargosthat originated in tropical Asia. Current research will
doubtedly these questions in the future.Brazil’s infestation appears to be linked to the interna-
tional seaport of Victoria in the state of Espfrito Santo(PAHO, 1987). Since used tires from the United Statesregularly unloaded in this area, it is possible that secondarydispersion of Ac. albopiclus may have been responsible forits introduction into Brazil. It thus appears that all the infes-tations reported from the Americas classic examples of
the role played by increased travel and betweencountries in the proliferation of non-native species.
Studies North American and Brazilian strains of Ac.albopictus have shown that populations from both
cicctrophoretically similar to Ae. albopictus populationsfrom temperate of northern Asia (Kambhampati et al.,1991).
In the United States, studies carried out to determine the
degree of differentiation among mosquitoes collected at
several sites in the city and among populations fromdifferent cities showed levels of genetic variation in theHouston and New Orleans populations. These results sug-gested large and independent introductions oiAe.dcs to bothcities and support the idea that Houston and New Orleans(Black et al., 1987) the first sites of introduction in
the United States.Overall, enzyme electrophoretic observations carried out
by Drs. W. Hawley and L. Munstcrmann with SoutheastAsian populations of At’, albopiclus show extreme geneticdifferentiation, in marked contrast to the relative homoge-neity of North American populations. "This appears to in-dicate that gene flow in the introduced populations of theAmericas is much higher than that found among indigenouspopulations’’ (Hawley, pcrs. comm., 1991). Other molec-ular studies appear to reinforce the notion of high levels of
gene flow in the species. For instance, variation in mtDNAanalyzed in 17 populations of Ac. allwpictus using 18,
4, 5, and 6 bp cutting restriction enzymes (Kambhampatiand Rai, 1991), The results revealed low level of restric-tion fragment length polymorphism in the Ac. alhopictuspopulations, compared with other insect species. Only three
populations found to carry novel mtDNA haplotypcs.An analysis of individual mosquitoes revealed that thesepopulations included individuals carrying both the ancestral
36

and the novel mtDNA haplotypcs, suggesting that the di-
vergence of the populations is incomplete. The studycluded that the recent range expansion of the species and
gene flow facilitated by human activities could explain the
low level of polymorphism. Rai (1986 and 1991) reviewedstudies the genetics of Ac. allwpictus.
In general, it is believed that populations that highlyvariable, i.e., genetically plastic, better able to avoid
control and to develop resistance. Partial resis-
tance lo organophosphates, specifically malathion, hasarisen inAe. alhopictus populations from Houston and NewOrleans (NOMCB, 1987; Khoo et al, 1988; Robert and
Olson, 1989). Various levels of resistance to organophos-phates have been observed in several populations in theUnited States, while Brazilian strains from Kspirito SantoSlate (Cariacica City) have been found to be highlyceptible to the insecticides studied (Wesson, 1990).
Control
It is difficult to control eradicate Ae. alhopictusthan Ae. aegypti because Ac. alhopiclus is found fartherfrom human habitation and in wider range of habitats.
Population suppression may be economical only inhuman communities. Clearly, the best approach for
controlling Ae. athopiclus and other Stegomyia species is
to limit the availability of larval habitats. Massive educa-tional programs encouraging citizens to take actions to pre-vent mosquito breeding around and in their homes playcential role in control efforts.Prior to the U.S. infestation, very few studies evalu-
ations dealt with the control ofAe. allwpictus. In fact, this
species rarely, if ever, the primary target of controlprogram. More often, it considered secondary in im-
portance to Ae. aegypti. To date. the most extensive effortto control Ae. aegypti and Ae. alhopictus has been carriedout in Singapore. This integrated program, begun in 1968,includes environmental management, health education, le-
gal measures, and community participation; chemical
trol is reserved solely for outbreaks of dengue hemorrhagicfever (WHO, 1986b).
Chemical Control
House-to-house control campaigns, directed specificallyat Ar. aegypti, appear to be less effective against Ae. al-
bopicius, whose control requires coverage of larger areas,including the forest. For instance, island the Gulfof Thailand, temephos applications and malathion foggingeffectively suppressed the Ae. aegypti population but did
not significantly reduce the Ae. alhopiclus population(Gould et al., 1970).Dowling (1955) reported control ot’Ae. allwpictus using
15% dieldrin applied at 88 ml per ha with SwingFog(R)
portable thermal tog generator. With treatment of
entire 11.5-km’1 island Singapore, good controlachieved for 10 days; however, the population recovered
rapidly during the third week. With two treatments spacedweek apart, prolonged control achieved (a 92%
reduction for up to eight weeks).As already stated, in Singapore, chemical control is used
only during outbreaks of dengue hemorrhagic lever. Duringsuch epidemic in 1973, special attention given to
400 construction sites where vector breeding places and
harborages profuse. To kill the adult mosquitoes, all
the construction sites fogged with hioresmethrinmixed with equal parts of piperonyl butoxide synergist;the containers treated with 1% temephos sand gran-ules 1% malathion emulsion to kill the immature stages.The DHF continued, however, continuous dailytreatment implemented at these sites. This approachresulted in significant drop in DHF incidence (WHO,1986b).
In Louisiana, ultra-low-volume (ULV) application of the
synthetic pyrethroid bioresmethrin reduced the adult Ae.ulbopictus population by 60%, but the reduction lasted for
only about three days (Anderson, New Orleans MosquitoControl District, pers. comm., 1986). In New Orleans, for-mulations of permethrin sandcorc granules containing 1.5%active ingredient tested to control larvae in experi-mental plots containing tire piles. The pesticide appliedusing manually operated sprayers. The approximateweight per treated tire 8 g. A single application pro-duced larval control of nearly 100% during period of121 days (NOMCB, 1987) and greater than 80% for nearly300 days (NOMCB, 1988). A similar experiment, alsoried out in tire piles New Orleans, conducted to
the effects of combination of corncob fenoxicarb-
impregnated material (\% active ingredient) and granularformulations of Bti (Bacillus thuringicnsis Israelensix)(200 ITUs). The weight of granules received in eachtire 5.8 g. Mortality of larvae in the treated tires
ceeded 80% al 60 days, but tell to 9% at 81 days (NOMCB,1987).During the of 1989 aerial ULV malathion treat-
ments conducted in two urban of New Orleans.
The twin-engine aircraft equipped with two self-contained Micronair spray pod systems utilizing rotary at-
omisers. Aerial treatment rates 3 oz/acre, at treat-
ment altitude of 120-200 feet and flight speed of 150
mph, resulting in swath width of 500 feet; this treatment
succeeded in suppressing the population of Stegomyiamosquitoes for days (NOMCB, 1989),
Jardina (1990) reported the eradication of Ac. albopictusfrom Indianapolis in Marion County, Indiana, throughcombination of chemical treatment and reduction
(removal of tires and trash from yards). The classical treat-
ment of malathion and granular Abate replaced with
syncrgized rcsmethrin and granular Bti. In September 1987.
37

adulticiding performed around infested tire retread-ing company and surrounding roads using ULV foggingunit mounted truck. Synergized resmethrin usedat rate of 130 ml/min during five consecutive days. Theoperation conducted twice day at the optimal spray-ing times of mid-morning and early evening. In addition,
portable ULV unit used in inaccessible Simul-taneously, larvicidc operations with Bti carried out in
3,500 tires six times during period of 45 days. Surveil-lance operations conducted in 1988 and 1989 failed to de-tect any Ae. allwpictus in the
Other studies ofAf. ulbopictus populations from aroundHouston, Texas, demonstrated the adulticidal effects ofmethrin, which produced 96% mortality at concentrationsof 1.5 ng/feroale (48 hours posttreatmcnt at 21 C) (Khooet al., 1988). Another pyrethroid showing moderately toxic
effects Ae. ulhopiclus strains from Malaysia is cyfluthrin(Responsar) (Vythilingam et al., 1992). Both wettable andemulsifiable concentrates of cyfluthrin sprayed plywood(10 nig [aij/rar) produced about 60% mortality after 20 daysof exposure.
Studies carried out with U.S. strains of Ae. albopictushave shown considerable variation in the response ofious geographical populations to organophosphate insecti-cides (Khoo ct al., 1988; Robert and Olson, 1989; Wesson,1990; Sweeney, 1993). According to Wesson (1990), there
two possible for the variation in insecticide
ceptibility among the U.S. strains: the U.S. populations mayhave originated from number of different Asian withdistinct insecticide exposure conditions, selection for in-
secticide resistance may have taken place in the UnitedStates sinceAc’, alhopictus became established. Wesson the-orizes that the observed differences may be the result ofboth factors. Some selection for insecticide tolerance to
malathion is believed to have taken place in populationsfrom Texas and Louisiana.Brown (1986) and Neng et al. (1993) report that Ae. al-
bopictus is resistant the organochlorincs DDT and HCHin China, India, Japan, Malaysia, Southeast Asia, and thePhilippines and resistant to the organophosphate malathionin Singapore and Viel Nam, fenthion in Malaysia, and fen-itrothion in Madagascar.
Biological Control
With the exception of the trials carried out with cyclo-poid copepods (Marten, I990a, 1990b), there have been fewfield trials of predators, parasites, pathogens of Ae. al-bopictus. Reports have been limited mostly to findings offield infections and species associations.The predatory mosquito larva Toxorhynchites splendens
found to be frequently associated with Ae. ulbopictusin Singapore, and Chan (l968a) concluded that "... T.splendens probably be most effectively utilized for the
control QiAedes albopictus in rural areas....’’ On the otherhand, Ncwkirk (1947) believed that T. splendens wouldcupy minor role in the elimination of mosquitoes: "Itslong life and few offspring, when compared to Aedes al-
hopicJus..., and its low survival rate tend to nullify the
importance of (T.) splendens in biological control." An at-
tempt to reduce the breeding of Ae. albopictus by intro-
ducing small numbers of T. inornaius in Hawaii in the1920s failed when this predator died out (Gerberg, 1985).The technology developed for the rearing and field
trials of T. brevipalpis and T. rutilus for the control of Ac*.aegypli in the Amcricas (Gerberg, 1985) may be utilizedwith better results against Ae. albupictus, since both
the latter species and these predators breed readily in nat-
ural containers away from homes, while the breeding sitesof Ae. uegypli domestic.
In laboratory tests with the parasitic mermithid nematodeRomanomermis culicivorax, Ae. albopictus rated "3"
susceptibility scale of to 5, indicating that the hostshowed moderate physiological resistance (Finney-Crawlcy,1985).Roberts et al. (1983) report two observations of labora-
tory infection ofAe. ulbopictus with viruses: densonucleo-sis virus, which only retarded growth at low temperatures(16 C), and Nodamura virus, small RNA virus that in-fects vertebrates. This virus caused death in Ae. albopictuswhen inoculated into the thorax but little mortality when
ingested by adults immersed larvae.In the laboratory, susceptibility of Ae. albopictus to the
bacterium Bacillus ihuringlensis israe-lensis foundto be lower than that of Ae. aegypti from Enugu, Nigeria,and Djakarta, Indonesia, but higher than that ofAc. aegyptifrom Bora Bora (de Barjac and Coz, 1979).
Infection with three species of grcgarine protozoans ofthe genus Ascogregarina observed in Ae. alhopiclus in
Taiwan (Lien and Levinc, 1980). Beicr and Craig (1985)state, "Although many workers have suggested the poten-tial of gregarines control agents, these parasites appearto be relatively harmless to their natural mosquito hosts."
Various species of the pathogenic fungus Coelomomycesfound parasitizing A*?, ulbopictus larvae from bamboo
stumps and tree holes in Taiwan between 1955 and 1967
(Laird, 1967). Two species of this fungus (predominantlyC. stegomyiue) that parasitized Ae. albopictus in Singapore
collected and introduced into three atolls of the To-kelau Islands, New Zealand, in 1958. After two years the
fungi had significantly reduced the population of Ae.pofynesiensis, and by 1963 there fourfold increase in
parasitized larvae (Laird, 1967).Finally, the of copepods (Mesocyclops), discussed
earlier larval predators, has been demonstrated in field
trials in New Orleans (Marten el al., 1989a; Marten, I990a,1990b). Several species of cyclopoid copepods thatnaturally in New Orleans, such Mucrocyclops alhidus,Mesocyclops longisetus, and Mesocyclops aspericornis,
38

have been shown to be highly effective for controlling firstinstar larvae of Ac’, albopictus in discarded tires. The most
effective of the three appears to be Macrocyclops albidus,which, at density of 72 per tire, killed average of 45larvae in 24-hour period. Treatment of tires with this
pepod lias provided control for period of two years (Mar-ten, 1990b). Survival of this species is excellent when it is
introduced into tires around woodedAnother species that has shown promise in controlling
Ac. alhopiclus first instar larvae is Mesocyclops longisetus.This species appears to be highly effective in tires situatedin the open and exposed to hot conditions. Nine-teen specimens eliminate 40 larvae per day. The dis-
advantage of Mesocycfops longiselus is tliat tlie speciesdoes not survive the winter season, and it is sensitiveto freezing temperatures than Macrocyclops (Marten,1990b).Another approach that has been recently introduced in-
volves treating tire piles with mixtures of copcpods and Bti.Such treatment has been found to provide control for about
year. Bti at concentrations of 1,200 ITU/ml mixed with
Macroi-yclopv alhidiis had detectable toxic effect the
cyclopoid, but killed both late instars and instars ef-
fectively (Marten et al., 1993).
Environmental Control
Environmental management involves the eliminationreduction of vector breeding Commonly known
reduction, it appears to be the single most effectivemethod ^ Aedes vector control thus far available. The
derlying principle of reduction consists of the elim-
ination of breeding in order to disrupt the immaturelife cycle of the mosquito. This objective be imple-mented by such simple burying destroyingunwanted water-bearing receptacles. Particular attentionshould be given to reducing standing water in and aroundseaports, bus terminals, train stations, and airports, well
in containers such discarded tires, which favorite
breeding sites of the Acdcs species. Additional actions that
be taken include upending large containers such jars,drums, and water tanks that kept in the open and
ering water-holding containers with tight-fitting lids. Peoplebe encouraged to change the water weekly in containers
around their homes, such water jars, flower vases, andornamental jars, and to scrub and thoroughly rinse thesecontainers lo dislodge Aedes eggs before refilling withter. Achieving permanent mosquito control in naturaltainers such tank bromeliads is difficult, but homeownersmay consider limiting the numbers of these plants that theyplace in their yards. In such Florida it ismended that tank bromeliads be flushed out twice weekwith garden hose equipped with appropriate nozzle
(O’Meara and Gettman, 1991).
In Singapore, where Ae. albopictu’i populationsfound to have three peaks during the year, fluctuating in
step with rainfall, the best control aimed at
preventing the population peaks by destroying the majorbreeding during the period immediately prior tothose peaks (WHO, 1986b).The most effective form of environmental control is to
modify completely and permanently the wherequito production Slums, which the
sponsible for most Aedes breeding in Singapore, have beenvirtually eliminated by government housing program. By1984, than 75% of the total population housedin government-constructed apartment buildings. However,this effort did not solve the problem of infestations in largehouses with compounds, because although Ae. aegypfi in-
dices in such houses low, Ae. cilbopiclus house infes-
tation almost 100%. This the because thesepremises usually had suitable Ae. albopictus breeding hab-itats, such tin cans, tree holes, and leaf axils of plants(WHO, 1986b).
Health Education
Public health education is aimed particularly at the neigh-borhoods where Aedes indices highest. To motivate peo-ple to eliminate and prevent mosquito-borne diseases,number of public campaigns be imple-mented. These may involve the holding of seminars, work-shops, and exhibitions; the distribution of pamphlets and
posters; the preparation of feature articles for newspapersand magazines; the presentation of talk shows and theof slogans the radio, television, and in schools; the
of film strips; and the personal message of the vector
control officer working door-to-door. In Singapore, it
concluded that "there is doubt that continuous routinehealth education contributed immensely to the
prevention and control of Aedes breeding in premises"(WHO, 1986b).
Legal Measures
The legal component of Aedes control has two aspects:laws that permit the health inspector to enter each buildingand examine all potential breeding sources, and legislationthat makes it illegal to have breeding places the prem-ises. In Singapore, the Destruction of Disease-Bearing In-sects Act permits the control officer to orders and
offenders who maintain conditions condu-cive to breeding harboring mosquitoes. For privatehomcowner, order is served when breeding has beenconfirmed.To lessen the likelihood that Ae. ulbopiclus will
from the United States to other countries of the Americas,the Centers for Disease Control and Prevention has pro-
39

posed discussion of the following four options: (a) de-velop and implement regulations to require disinsection ofall used tires exported from the United States; (b) formulateguidelines for by PAHO countries to require fumigationof used tires shipped from the United States; (c) maintainconstant surveillance in port and at destinations ofimported used tires in each receiving country; and (d) pro-vide assistance by CDC through PAHO to countries wish-ing to design surveillance and control plans (CDC, 1993).
Integrated Control
No single control method is sufficient to lower
quito levels for long period of time. Instead, it issary to combine the various methods of the vector controlarsenal in the most effective, most economical, and safestpossible way. In Singapore, system of routine vectorveillance, continuous year-round reduction, healtheducation, and fogging of premises in where
than 5% of the houses infested implemented in1974. This system effective in achieving year-roundcontrol of the vectors Ae. aegypli and Ac. albopictus, and
also successful in preventing two epidemics that sweptthrough other parts of the region (WHO, 1986b).
CONCLUSIONS
Some health officials and members of the scientific
munity have suggested that Ae. albopictus is less impor-tant vector of dengue virus than Ae. aegypti. Experience inAsia the past 30 years supports this contention in that,with only few exceptions, the major epidemics of dengueand dengue hcmorrhagic fever have been transmitted by Ac.aegypti. Nevertheless, the threat of Ae. albopictus in theAmericas should not be underestimated. Craig (1993)points out the differences that exist between the two Sfe-gomyia species in the Ajnericas and describes themfollows:
Ac. albopictus has far greater geographical range thanAe. aegypli because of its cold tolerance.Ae. albopiclus has inaccessible habitat because itbreeds in plants, and therefore it is far harder to control.The species have different habitats, with Ae. aegyptifound in the inner city and Ac. albopiclus in suburbanand rural
Ae. albopiclus has already achieved far greater popula-tions in the United States than Ae. aegypti, which has
been major biting pest in that country; forample, in New Orleans, 80% of the mosquito complaints
currently about Ae. alhopictus.Ae. albopictus harbor the range of arboviruscsand is efficient vector for of them; Ae.
gypli has been known to enter into the cycle ofEEE.Ae. albopictus has the potential to enter into the LaCrosse virus cycle, while Ae. aegypti has chance of
doing because it cannot survive the winter cold.Ac. albopiclus feed wide variety of hosts, whileAe. aegypti outside Africa is highly domestic and is
likely to in contact with feral
The information about Ae. alhopictus presented in thisdocument portrays very complex species. Its ability to
harbor indigenous arboviruses has been demonstrated in the
United States, and wealth of experimental data haveinforced the evidence of its potential role in vectoring hu-
and animal arboviral diseases. Complicating the pic-ture is the catholic feeding behavior of the species, whichhas been reported feeding not only small and largemals, including humans, but also birds, snakes, and tur-
tles. Ae. albopiclus enter into sylvan zoonotic cyclesand bring such viruses to man; that is why it is danger-
Moreover, unlike swamp and marsh mosquitoes, Ae.albopiclus breeds close to human habitations.A very disturbing development is isolation of the eastern
equine encephalitis (EEE) virus from Ae. albopictusquitoes collected at the Polk County Tire Dump in Florida.EEE and often fatal disease in humans. It isalso important disease of domestic animals and has
killed significant number of animals belonging to endan-gered species, such the whooping
Houston (Texas) and Brazilian strains of Ae. albopictushave shown susceptibility to experimental oral infection withthe four dengue serotypes (Mitchell and Miller, 1990). IfAe.albopictus spreads into Central America and the Caribbean,it could add complexity to the scenario of dengue inthe Americas. Ae. albopictus breeds in urban, rural, and for-ested in water retained by artificial containers andnatural vegetation. It is exophilic and prefers to breed out-side human dwellings, which makes it potentially danger-
maintenance vector.
Seven years after its discovery in the Americas. Ae. al-
bopiclus continues to expand its range into geograph-ical In 1986, 11 states in the United Slates
ported to be infested; by 1991, 11 additional statesknown to have significant populations of Ae. albopiclus.Similarly, in Brazil infestation reported in 89 munici-
palities in 1986; by April 1994, 676 municipalities andthree states-Maranhao, Parana, and Bahia-had beenadded to the list of infested areas, indicating well-estab-lished breeding population in the country. In addition, the
expansion is not restricted to the Americas. In the last few
years, the species has spread rapidly to many communitiesin northern Italy.
There still many unanswered questions about Ac. al-
bopictus and its relationship to human diseases. For in-
stance, association of Ac. albopictus with arboviral dis-
40

outbreaks has been reported in the Americas. TheMinistry of Health of Brazil did not find any evidence
sociating/le. albopictus with the dengue outbreaks reportedduring the past five years, but detailed studies need
to be carried out during such events, especially sinceeral of the recent dengue epidemics took place in Brazilianstates wliereAc. ulbopictus is known to be present. Clearly,such studies help to resolve many of the enigmas whichsurround this species in natural settings.
Ae. alhopictus is highly adaptable species. Its evolu-
tionary plasticity is demonstrated by its ability to developinsecticide resistance after only lew generations of labo-
ratory selection. Insecticide selection studies of Ae. albo-piclus strains from the United States indicated unusuallyhigh tolerance to malathion, resulting in resistance ratioof 21 after just six generations (Wesson, 1990). Controlefforts in infested may be compromised if Ac. alho-
pictus develops resistance trait.
Enzyme electrophoretic studies appear to indicate that
gene flow in the introduced populations of the Americas is
higher than that found among indigenous populations ofSoutheast Asia. These observations render indirect evidenceof the importance of the tire trade in disseminating intro-duced populations, and show that interdiction of that tradecould prevent the colonization of other of the
hemisphere.The potential for the species to spread to other ecosys-
tems in the Americas is real, demonstrated by the recent
reports from northern Mexico and the Dominican Republic.If the trend continues, enlargement of the foci in Brazil, the
Dominican Republic, Mexico, and the United States be
expected, along with expansion of the infestation to othercounlriys of the Americas. involvement of Ac. albopictusin the dengue cycle, and involvement of the species intransmission of zoonotic arboviroses. Moreover, if Ae. al-
bopiciuh spreads into yellow fever endemic of South
America, it could act bridge vector and bring yellow-fever to South American urban centers.
The continuing spread of Ac. albopictus to
require’; the urgent attention of health agencies and the sci-
entific community throughout the Americas.
ACKNOWLEDGMENTS
We grateful to Dr. Duane Gubler, Director of theDivision of Vector-Borne Infectious Diseases, Center forInfectious Diseases, Centers for Disease Control and Pre-vention, Fort Collins, Colorado; Dr. George O’Meara, Flor-ida Medical Entomology Laboratory, Institute of Food and
Agricultural Sciences, University of Florida at Vero Beach,Florida; Dr. Michael J. Turell, Applied Research Division,U.S. Army Medical Research Institute of Infectious Dis-
eases, Kort Dctrick, Frederick, Maryland; and Drs. MichaelNelson and Francisco Pinheiro, PATIO/WHO, all of whom
read the drafts, made valuable comments, and suggestedchanges to improve the document. For providing data.drawings, and photographs indebted to Dr. ChesterMoore, Division of Vector-Borne Infectious Diseases, Cen-ter for Infectious Diseases, Centers for Disease Control andPrevention, Fort Collins. Colorado; Dr. John R. Linley,Florida Medical Entomology Laboratory, Institute of Foodand Agricultural Sciences, University of Florida at VeroBeach; Dr. Ralph Harbach, Walter Reed BiosystematicsUnit, Smithsonian Institution, Washington, D.C.; and Mr.Arthur Botelho dc Barros, Superintendencia de Campanhasde Saude Publica (SUCAM), Brazil. Part of the researchreported bv G. B, C. supported by NIH Research GrantNo. 02753.
REFERENCES
ACHA, P., and B. Sy-ytres. 1987. Zooiww", and CommunicableDiseases Common Man and Animals. Pan American
TIcallh Organization, Washington, D.C. Scientific Publication503.
AKHTER. R.. C. G. Haycs, S. Bagar. and W. Reisen. 1982. WcslNile virus in Pakistan. III. Compaialivc capability ofCulcx tritaeniorhyttchus and eight other species of mos(.]ui-
Trans R Sor Tiop Mcd Hyg. 76:449-453.ALI, S., and I- Rozeboom. 1973. Comparative laboratory obser-
vations selective mating oiAedcv (Sicgomyia) atbopiciusand Aedcs (Su’gvmyia) polynesicnsis Marks. Mosq News, 33:23-28.
AMERASINGHE. F. P., and T. S. B. Alagoda. 1984. Mosquitooviposilion in bamboo traps, with special reference Aedes
albopictus, Aedcs nwalbupictus and Arrnigercs ’iubulbaliis.
Insect Sci Appi, 5(6):493-500.APPERSON, C.. B. Engber, and J. Lcvine. 1989. Relative hiiila-
bility of A<.’. (illwpicms andAe. ac’gypli in North Carolina
support development of Dirofitaria immilis.JAin Mosq Cun-tro! Assoc, 5(3):377~382.
AZZIZAR, R. K. 1980. Studies the breeding habitats and
sonal prevalence of larval population of Aedes ac-gypli (L)and Aedes alhopiclus (Skusc) in Dacca Cily. BMRC Bull.
6(2):45-52.BARRAUD, P. J. 1928. A revision ot the Culicine mosquitoes of
India. Indian Med Res, 15:653-670.BASIO, R., and L. Santos-Basio. 1974. On Philippine mosquitoes.
XIV. Biting cycles of species in their natural foresthabitat, with particular reference to Aede^ alhopiclini. Philipp,/ liiol, 3:155-165.
BAT-MIRIAM, M., and G. B. Craig, Jr. 1966. Mutants in Aedes
alhopictus (Diptera: Culicidae). Movq News. 26(1): 13-22.BAUST, J.. and R. Lee. 1981. Divergent mechanism of frosi-
hardiness in populations of (he gall uy, Eurostu solida-
ginis../ Insect Physiol, 27:485-490.BHAMAN, J., and M. Turell. 199). Transmission of Venezuelan
equine encephalomyelitis virut> by strains otAedes albopiclits(Dipteia: Ciilicidae) collected in North and South America.
Mod Eiitomol, 28(1):16J-164.
41

BEIR, J. C., and G. B. Craig, Jr. 1985. Gregarine parasites atmosquitoes. In: Integrated Mosquito Control Methodologies,Volume 2:167-184. Academic Press, London. 444 pp.
BENTLY, M. D., N. McDanicl. H. P. Lee, B. Sliclil, and M.Yatagai. 1976. Sliidies of Aedes triseriatas oviposiliontractants produced by larvae of Aedes triserialus and Aedes
aitopalpus (Diptcra/’Cnlicidae).yAfct/K;w/i(?/, 13:112-115.BLACK, W., J. A. Fcrrari, K. S. Rai, and D. Spienger. 1987.
Breeding in colonizing species: Aedes albopicltts(Skusc) in the U.S.A. Heredity. 60:173-181.
BLACKMORE, M. 1993. Strain-spccinc differences in suscepti-bilily ofAf. albopiclus infection by mermilhid and filaridnemalodes. Vector Control Bull North Central Stales, 2(1);67-73.
BONNET, D. D.. and D. J. Worcheslci. 1946. The dispersal ofAedes alhopiclus in the lerrilory of Hawaii. Am Trap Medllyg, 26:465-476.
BOSIO, C., R. Thomas, P. Grimsiad, and K. Rai. 1992. Variationin (he efficiency of the vertical transmission of dcnguc-1 vi-
by strains of Ac. albopiclus (Diplera: Culicidae). MedEntomol, 29(6):985-989.
BROWN, A. Vl. A. 1986. Insecticide resistance in mosquitoes: Apragmatic review../ Am Mosq Control Assoc, (2):123-140.
BURKOT, T, P. Oravc.s, R. Paru, and M. Lagog. 1988. Mixedblood feeding by the malaria in the An. punctulatuscomplex (Diptcra: Culicidae). ./ Med Entomol, 25(4):205-213.
CAI.ISHER, C. 1994. Medically important arboviruses of theUnited Slates and Canada, (.’(in Micmblol Kcv, 7(1):89-116.
CALISHER, C., E. Lcvy-Kocnig, C. Milchell, F. Cabrcra, L. Cuc-vas, and J. Peanion. 1981. Enccfalitis equina del la
Republica Dominicana. Bol Oftdna Sanit Panam. 90:19-31.CENTERS FOR DISEASE CONTROL AND PREVENTION
(CDC). 1986a. Aedes albopiclus introduction in Texas.MMWR, 35:141-142.
CENTERS FOR DISEASE CONTROL AND PREVENTION(CDC). 1986b. Aedes alhopicius infestation-United Stales,Brazil. MtfWR, 35(31):493-495.
CENTERS FOR DISEASE CONTROL AND PREVENTION(CDC). Division of Vector-Borne Infectious Diseases. 1993.The Movement of Aedes albopietus in South Florida: PublicHealth Significance. American Mosquito Control Association
Meeting, April 1993, 10 pp.CHAMBERLAIN, R., W. Sudia, and P. Coleman. 1969. Isolations
of arbovirus of the bunyamwcra group (Tensaw virus)from mosquitoes in the Southeastern United States, 1960-1963. Am 1’mp Med Hyg, 18:92-97.
CIIAN, K. L. 1968. Observations Toxorhynchites splendens(Widemann) (Diptcra: Ciilicidae) in Singapore. Mosq News,28( 1):91-95.
CHAN, K. 1.. 1971. Life table studies of Aedes iilbopicliis (Skusc).In: Proceedings of symposium the sterility principle forinsect control eradication. International Atomic EnergyAgency, 131-144.
CIIAN. K. L. 1985. Singapore’s Dengue Haemorrhage b’e.vcrControl Programme: A study the successful control
of Acdcs aegypti and Aedes albopiclus using mainly envi-
ronmental part of integrated control.Southeast Asian Medical Information Center, Tokyo, Japan.
CHAN, K. L. B. C. Ho. and Y. C. Chaii. 1971. Aedes aegypli (L)and Aedes albopicius (Skusc) in Singapore City. 2. Larvalhabitats. Bull World Health Organ, 44:629-633.
CHAN. Y. C.. B. C. Ho, and K. L. CIian. 197I.Aerff.s- aejjypli (L)and Aedes albopiclus (Skusc) in Singapore City. 5. Obser-vations in relation dengue hacmorrhagic lever. Bull WorldHealth Organ, 4:651-658.
CHRISTOPHERS, R. 1960. Aedes aegypli (!.), The Yellow feverMosquito: Ils Life History, Bionomics, and Structure. Cam-bridge University Press, Cambridge.
COLEMAN, P. 1969. Tensaw virus, member of the bun-yamwera arbovirus group from the southern United States.Am Imp Mod Hyg, 18:81-91.
COPELAND, R., and G. Craig. 1990. Cold-hardiness of tree-holemosquitoes in the Great Lakes region of the United Stales.Can Zoo;. 68:1307-1314.
CRAIG, G. 1993. The diaspora of the Asian Tiger Mosquito. In:B. McKnight (ed). Biological Pollution: The Control and Im-pact of Invasive Species. Proceedings of Symposium, Uni-
versity Place Conference, Indiana Univcrsity-Purdue Univer-
sity, Indianapolis. 25-26 Ocl. 1991. Indiana Academy ofScience, pp. 101-120.
CRAVEN, R., D. F.lliason, D. Francy, P. Reilcr, F.. Campos, W.Jakob, C. Bozzi. and C. Moore. 1988. Importation ofAcA’.s-aibopictus into (he USA in used tires from Asia../ Am MosqControl Assoc, 7(4):651-653.
CUI, K. L. 1982. The aulogeny of Aedes albopiclus in GuangzhouAda Entomol Sin, 25(3):256-259.
CULLY, J., P. Heard, W. Wesson, and 0. B. Craig. 1991. Anti-bodies I-<1 Crosse virus in Indiana in chip-munks Aedes albopiclus population.. Am.Mosq Con-rro7A-oc,7:651-653.
CULLY. J., T. Strcil, and P. Heard. 1992. Transmission of LaCrosse virus by four strains oiAedes ulhopictus and fromthe eastern chipmunk (Tamias striatus}. Am Mosq ControlAssor, 8:237-240.
DE BAR1AC. H., and J. Coz. 1979. Scnsibilitc comparee de six
cspeccs differcntes de moustjquc Bacillus thuringiensisisraelensis. Bull WHO, 57:139-141.
DEFOLIART. G. R., D. M. Watts, and P. R. Grimsiad. 1987. Ad-
in mosquito-borne arbovirus/’veclor research. AmwRev Enlomol, 32:479-505.
DEL ROSARIO, A. 1963. Studies the biology of Philippinemosquitoes. Observations the life and behavior ofAerfc-s-albopiclus (Skuse) in the laboratory. Philip? Sci.
DETINOVA, T. S. 1962. Age-grouping Methods in Diplera ofMedical Importance. World Health Organization, Geneva.WHO Monograph Series No. 47.
DINGER, J. E.. W. A. Schuffncr, P. Snijders, and H. H. Swcllcn-grebcl. 1929. Onderzoek gcle koorlsin Nederland.Ncderlandsch Tijdschrift gcngcs Kunde.
DOWL1NG, M. A. C. 1955. An experiment in mosquito controlusing dieldrin dispersed dry fog. Trims R Soc Trap Medffvg,49(6):59(l-601.
EDGERLY, J., M. S. Willey, and T. P. Livdahl. 1993. Themunity ecology of Aedes egg hatching: implications tormosquito invasion. Ecol Entomol, 18:123-128.
ELLIOT, S. A. 1980. Aedes atbopklus in the Solomon and SantaCruz Islands. South 1’acilic. Trails R Soc Trap Med Hyg,74(6)747-748.
42

FAN, W., S. Yu, and T. Cosgriff. 1989. The reemergencc of den-gue in China. Rev Infect Dis, May-June, ll(Suppl 4);847-85;1.
FENG, L.-C. 1937. A critical review of literature regarding (he
records of mosquiloes in China. Peking Nat Hist Bull,12(3)295-296.
FENG, L.-C. 1938. The geographical distribution of mosquitoesin China. 7th Proceedings International Congress of Ento-mology. Berlin, Germany, pp. 1579-1588.
FINNEY-CRAWLEY, J. R. 1985. Future prospects tor commercialdevelopment of nematode agenis for biocontrol. In: Inte-grated Mosquito Control Methodologies, Volume 2:287-304.
Academic Press, I-ondon, 444 pp.FOCKS, D.. S. Linda, 0. B. Craig, W. Ha\ley, and C. Piimpnni.
1994. Aedes albopictus (Diplera: Culicidae): A statisticalmodel of the role of temperature, photopcriod. and geographyin (he induction of egg diapause. ./ Mod Entoinoi, 31:278-286.
FORAITINI, 0. P. 1986. Aedes (Slegomyia) albopictus (Skuse)identification in Brazil. Rev Saude Publka, 20(3):244-245.
FRANCY, D. B., N. Karabalsos. D. M. Wesson, C. G. Moore,J. S:. Lasuick. M. L. Niebylski, T. F. Tsai. and 0. B. Craig.i990a. A arbovirus from Aedes albopictus,, Asian
mosquito established in the United Slates. Science, 250:1738-1740.
FRANCY, D., C. G. Moore, and D. A. Eliason. 1990b. Past, pres-and future oiAt’des albopictus in the United States../ Am
Mofiq Control Assoc, 6:127-132.FUTUYMA. D. 1986. Evolutionary Biology. Sinauer Associates,
Sunderland, Massachusetts.GALLIARD, H, and D. V. Ngu. 1938. Variations saisonnicres de
1’cvohition de Diroftlaria immitis chez Aedes (Slegomyia) al-
hopiclus. Ann Parasil Hum Comp, 16:210-214.
GALLIARD, H., and I. J. Golvan. 1957. Influences dc ccrtains
facleurs nulrilionels hormonaux, des temperatures varia-
ble’;, la croissance des larves d’Aedes (S.) acgyoll, At’des(S.) albopiclus Anopheles (M.) Stephens!. Ann Parasite!,23(5-6):563-579.
GAO, J. Z., Y. Z. Zhao, M. X. Jing, Y. H. Ping, P. Z. Jun, and
H. Nian. 1984. Studies the longevity of caged femalesunder laboratory conditions. Acta Enlomol Sin, 27(2): 182-188.
GERBERG. E. J. 1985. Sequential biocontrol applications in theof Toxoihyncltiles spp. In: Integrated Mosquito Control
Methodologies, Volume 2:33-46. Academic Press, London,444 pp.
GILLET, J. D. 1971. The Mosquito: Its Life, Activities and ImpactHuman Affairs. Douhleday, New York.
GILOTRA, S. K., L. R. Rozcboom, and N. C. Bliallacharya. 1967.Observations possible competitive displacement between
populations of Aedes aegypti (L) and Aedes allwpictus Sktise
in Calcutta. Bull World Health Organ. 37:437-446.
GOMES, A. dc C., 0. Forattini, I. Kakitani, G. Marques. C. dcAzt’vedo Marques. D. Marucci, and M. dc Brito. 1992. Mi-crohabilals at Aedes alhopictus (Skusc) in the Paraiba ValleyRegion of the State of Sao Paulo, Brazil. Rev Saude Publica,
26(2): 108-118.GOULD, D. J., G. A. Mount. J. E. Scanlon. H. R. Ford. and
M. F. Sullivan. 1970. Ecology and control of dengue
island in the Gulf of Thailand. .7 MedEnlomol, 7:499-508.
GRIMSTAD, P. R.. ]. F. Kobayashi, M. Zhang, and G. B. Craig,Jr. 1989. Recently introduced Aedes albopictus in the United
States: Potential of La Crosse virus (Bunyaviridae:California serogroup)../ Am Mosq Control Assoc. 5(3):422-427.
GUBLER, D. J. 1970a. Comparison of reproductive potentials ofAedes (St.) allwpiclus Skuse and Aedes (St.) polynesiefisisMarks. Mosq News. 30:201-208.
GUBLER, D. J. 1970b. Induced sterility in Aedes (Stegomyia)polynesiensis, Marks by cross-insemination with Aedes atlm-
pictus Skiise. .1 Med Enlomol, 7(1): 65-70.GUBLER. D. J. 1970c. Competitive displacement olAcdcs (Ste-
gomyia) polynesiensLs Marks by Aedes (Slegomyia) albo-pictus Skusc in laboratory populations. ./ Med Enlomol, 1:
229-235.GUBLER. D. J. 1971a. Studies the comparative oviposition
behaviour of Aedes (Stegomyia) albopictus and Aedes (Sle-gomyia) polynesiensis. Med Entomol, 8:675-687.
GUBLER. D. J. 1971b. Ecology at Aedes alhopiclus. The Johns
Hopkins University ICMRT Animal Report: 75-80.GUBI.ER. D. J. 1987. Current research dengue. In: K.F. Harris
(ed). Current Topics in Vector Research, pp. 37-56. Springer-Verlag, New York.
GUBLER, D. J., and N. C. Bhallacharya. 1971. Observations
the reproductive history of Aedes (Stegomyia) atbopiclus in
tile laboratory. Mosq News, 30:356-359.GUBLER. D. J., and N. C. Bhallacharya. 1972. Swarming and
mating of Aedes (S.) alboplclus in Mosq News, 32(2):219-223.
GUBLER, D., and L. Rosen. 1976. Variation among geographicstrains of Ac. albopictus in susceptibility infection with
dengue viruses. Am Tmp Med Hyg, 25:318-325.HALSTEAD. S. B. 1980. Dengue haemorrhagic fever: A public
health problem and field for research. Bull World Health
Organ. 58:1-21.
HANSON, S. M. 1991. Cold hardiness of Ac. albopictus eggs.University of Notre Dame. Department of Biological Sciences,Noire Dame, Indiana, August 1991. Doctoral dissertation.
HANSON, S. M., and C. B. Craig. 1994. Cold acclimation, dia-
pause, and geographic origin affect cold hardiness in eggs ofAedes albopictus (Diptcra: Culicidae). ./ Med Enlomol, 31:
192-201.
HARDY, J. L.. L. Roscn, L. D. Kraracr, S. B. Presser, D. Shroyer,and M. S. Turell. 1980. Effect of temperature Iransovarialtransmission of St. Louis encephalitis virus in mosquitoes.Am Tmp Med Hyg, 29:963-968.
HARINASUTA, C.. S. Sucharit, T. Dcesin, K. Surathin, and S.Vulikes. 1970. Bancroftian ftlariasis in Thailand,demic Southeast Asian Trap Med Public Health, 1(2);233-245.
HARRISON, B. A., P. Boonyakanist. and K. Mongkolpanya.1972. Biological observations Aedes sealoi Huang inThailand with notes rural Aedes aegypfi (I.) and other
Stegomyia populations. J Med Entomol, 9(1);1-6.HATCHETT, S. 1946. Winter survival ofAc. aegypti (L) ill Hous-
ton, Texas. Public Health Rep, 61:955-964.
HAWLEY, W. A. 1988. The biology of Aedes alhopiclus. AmMosq Control Assoc, 4(Stippl):l-37.
43

HAWLEY. W. A. 1991. Adaptable immigrant. Afa! Hist Mas.91(7):56-58.
IIAWLEY, W. A.. P. Reiler, C. B. Pumpuni, R. Copcland, and
G. B. Craig, Jr. 1987. Aeilcs alhopiclus in North America:Probable introduction in tires from Northern Asia for North
Asian origin. Science, 236:1114-1116.
IIAWLEY, W. A.. C. Pumpuni, R. Brady. and 0. B. Craig. 1989.
Overwintering survival ofAc. albopiclu.s (Diplcra: Cnlicidae)eggs in Indianit../ Med Enlomol, 26(2):122-129.
HEARD, P., M. Niebylski, D. Francy, and G. B. Craig. 1991.Transmission of newly recognized virus isolated from Ac.albopiclits in Potosi, Missouri../ Med Entumol, 28:601-605.
HIEN, D. S. 1975a. Biology ulAcilcs aegypli (L.. 1762) and Ae-dcs albopidus (Skuse, j8’)5) (Diplcra. Culicidac). 111. Effectof certain environmental conditions the development oflarvae and pupae. Ada Piirasit Pol, 23(46):553-568.
HIEN, D. S. 1975h. Biology of AcaC.s aegypli (L., 1762) and
Andes albopidus (Skusc, 1895) (Diplcra, Culicidac). I. Re-sistance of eggs low humidity. Ada Parasilol Pol. 23(36);395-402.
HIEN, D. S. 1975c. Biology of Aedes acfypli (L., 1762) andAc-des alhopiclus (Skusc, 1895) (Diplcra. Culicidac). II. Effectof environmental conditions the hatching ot larvae. Ac’/a
Parasitol Pol, 23(45):537-552.HIP.N. D. S. 1976a. Biology otAcrfre avgypli (L., 1762) and Ar-
dm atbiipiclus (Skusc, 1895) (Diplcra, Culicidac). V. The
gonolropic cycle and oviposition. Ada Parasil Pol, 24(6):37-55.
HIEN. D. S. 1976b. Biology of At’A’.r aegypli (L., 1762) andAedes albopidus (Skuse, 1895) (Diptera, Culicidac). IV. Thefeeding of females. Ada Parmit Pol, 24(5):27-35.
HO, B. C., K. L. Chan. and Y C. Chan. 1971. Acdcs aegypli (I.)and AcacA’ albopiclits (Skusc) in Singapore City. 3. Popula-tion fluctuations. Bull World Health Organ, 44:635-641.
HO, B. C, K. L. Chan, and Y. C. Chan. 1972. III. Control ofAi’ddi The biology and bionomic olAedes albopidus(Skusc). In: Y C. Chan al. (eds). Vector Control in South-
Asia. Proceedings Isl SEAMEO Workshop, 15-17 Au-
gust 1972, Singapore.HO, B. C, Y. C. Chan. and K. L. Chan. 1973. Field and laboratory
observations landing and biting periodicities of Aide’s al-
bopidus (Skuse). Southeast Asian Trap Afra Public Health,4(2):238-244.
HOBBES, I., E. Hugos, and B. Eichold. 1991. Replacement ofAc. aegypti by At,’, albopidus in Mobile, Alabama. ./ AmMosq Control Assoc, 7(3):488-489.
HONG, 11. K., J. C. Shim, H. K. Shin, and II. Y. Young. 1971.Hibernation studies of forest mosquitoes in Korea, 1971. Ko-
J Enlomol, 1:13-16.HORWATH, K., and J. Duman. 1983. Preparatory adaptations for
winter survival in cold hardy beetles Deftdroides antadettsisand Dcndroidcs com’olur. Coma Physiol. 151:225-232.
HORWATH, K., and .1. Duman. 1986. Thcrmopcriodic involve-in antifreeze protein production in the cold hardy
beetle Dendroides eaftadensis’. implications for pholoperi-odic lime measurement../ lined Physiol. 32:799-806.
IIU, C. R., P. II. Chcn, J. M. Clicn, and 0. Huang. 1989. Electronmicroscopic observation of the sporogonic stage of Plasmo-dium gallinaceum after five antimalarials. Chimg Kuo Yao Li
Husiisn PIM, 10(5):434-437.
HUANG, H. M. 1972. The subgcnus Stegomyia of Aries inSoutheast Asia. The .scutellaris group of species. ConlribAm Eiitoinol lust, 9(1); 108.
mjANO, C. H. 1982. Studies of Japanese cnccphalitics in China.
Adv Virus Rfs. 27:71-101.
I1YLTON, A. 1967. I.ow humidity retention ability in
Erelmapodites chrysogascer and Aedes albopidus. .1 InsectPhysiol. 13:153-157.
HYLTON, A. 1969. Studies longevity of adult Erclinapodilcschrywgaster, Acdcs logoi and Acdcs (Stegomyia) alhopictusfemales../ Med Enlomol, 6(2):)47-149.
IMAT, C., and 0. Macda. 1976. Several factors effecting hatch-
ing Aedes alfwpiclits eggs. Jpn .1 Sanit Zoo/, 27(4):363-372.
1NSTITUT PASTEUR DE GUYANE. 1991. Kiipporl Annue!.French Guiana.
ISIIII, N., A. Nakayama, and Y. Ishii. 1954. Biological observa-
tions (he mosquito, Aedes alhopictus. Yokohama Mod
&i/(.5(4):275-281.JARD1NA, B. 1990. The eradication of Aedes albopidus in In-
diariapolis, Indiana. Am Moscf Confrol Assoc, 6:310-311.JUMALI, S., D. J. Gublcr, S. Nalim. S. Eram, and J. S. Saroso.
1979. Epidemic dengue hemorrhagic fever in rural Indonesia.III. Entomological studies. Am Trap Medltyg, 28:717-724.
KAMBHAMl’ATI, S.. and K. Rai. 1991. Mitochondrial DNAiation within and among populations of the mosquito At’.
alhopiclus. Gemme, 34:288-292.
KAMBHAMPATI, S.. W. Black, and K. Rai. 1991. Geographicorigin of the U.S. and Brazilian Avdfs albopidus inferred
from allozyrnc analysis. Heredity. 67:85-94.KAM1MURA, K. 1968. The distribution and habit of medically
important mosquitoes of Japan. .Ipit Sanit Zool. 27(4):367-372.
KARABATSOS, N. (ed). 1985. International Catalogue of Ar-
hoviruscs Including Certain Other Viruses of Vertebrates, 3rd
ed. American Society of Tropical Medicine and H)gicnc. SanAntonio, Texas.
KAY, B. H., J. A. Miles, D. J. Gublcr, and C. J. Milchcll. 1982.Vectors of Ross River vims: An overview. In: J. S. Macken-zie (ed). Viral Diseases in South-East Asia and the Wesletn
Pacific, pp. 532-536. Academic Press, Australia.
KAY, B. H., W. Ivcs, P. Whclan. P. Barker-Hudson. I. D. Fanning,and E. Marks. 1990. IsAc’are alhopiclus in Australia? Med
Ausl. 153(1);31-34.KHOO. B., D. Sutherland, D. Sprcngcr, D. Dickcrson, and H.
Nguycn. 1988. Susceptibility status ofAc. albopictus three
topically applied adiilticidcs../ Am Mostf Control Assoc, 4:
310-313.KLOWDEN, M. 1993. Mating and nutritional affect the
production ofAc. albopidu.s mosquitoes../ Am Mosq Control
Assoc, 9(2): 169-173.KLOWDEN, M., and G. Chambers. 1992. Reproductive and
abolic differences between Af. aegypti and Ae. albopidus(Diplcra: Culicidae). / Mrd Entomol, 29:467-471.
KNIGHT. J., and J. Bale. 1986. Cold hardiness and overwinteringof the grain aphid Silobion Ecot Enlomol. 11:189-197.
KONISHI, E. 1989. Susceptibility al Aedes alhopiaus and Culcx
trilaeniorhynchmi (Diptera: Culicidac) collected in Miki City,
44

Japan Oirofilana immifis (Spirnrida: Filaridac). Mcd En-lomol. 26(5):420-424.
KOUR1, G. I’., M. (;. Guzman. J. R. Bravo, and C. Triana. 1989.Dengue hemorrhagic fever/dengue shock syndrome; Lessonsfrom the Cuban epidemic, 1981. Hull Woi Id Health Oigan.67(4);375-380.
K.UKAL- 0., J. Duman, and A. Scrianni. 1989. Cold-induced mi-
tochondrial degradation and crvoproteclant synthesis infree/.e-tolerant arctic caterpillars. Comp Physiol, 158;()61-
671.
KUR1HARA, T. 1984. Mosquitoes occurring in the plant axils in
Iropical Asia. Jpn Sanil Zool, 35(1);63-69.I.A CASSE. W. J., and S. Yamaguli. 1950. Mosquilo Fauna of
Japan and Korea, App. //; Organization Function oj Ma-lalia Survey Detachments. Office of (he Surgeon, 8lh U.S.Army, Kyoto. Honshu.
LAIRD. M. 1959. Parasites of Singapore mosquitoes, wilh partic-ular reference the significance of larval epibiontsindex of’ habitat pollution. Ecology. 40(2);206-221.
I.AIRD, M. 1967. A coral island experiment, approachmosquito control. Chronicle; 21:18-26.
LAIRD, R. 1941. Observations mosquito transmission ofP/rtA’-
modium lophurae. Am Hyg, 34(c):163-l()7.LAMBRECIIT, Frank I.., and E. C. C. Van Somcrcn. 1971. Mos-
quitoes of the Chagos Archipelago, Indian Ocean. SoutheastAsian Trap Mrd Public Health, 2(4):483-485.
LEAHY. M., and G. Craig. 1965. Accessory gland substance
stimulant for oviposition in Ac. argypli and Ac. alhopictus.Mosq News, 25:448-452.
I.P.E, R., and D. Denlingcr. 1985. Cold tolerance ill diapausingand nondiapausing stages ol’ tile flesh lly Sarcopliagasipalpis. I’hysiol Enlomol. 10:31)9-315.
LIEN, j, and Y. Lin. 1990. The pathogen of Taiwan mosquitoes,
Coeiomomyce.’f species. Kcio Hsiun^ Hsuelt Ko Hsue-h TsaCh/h, d(7);350-359.
LIEN, S,., and N. D. Lcvinc. 1980. Three species of Asco-
cystis (Apicomplexa, I-eeudinidac) from mosquitoes../ Pro-tozool, 27:147-151.
UNI.EY, J. 1989. Comparative fine or the eggs of.4c.
alhopicias; Ac. aegypll and Ac; bahamensis (Diptera: Culi-
cidae). Med Entomol, 26(6):S 10-521.LIVDAHI, T, and M. Willcy. 1991. Prospects for invasion:
Competition between At; aHwpicliis and native Af. [riseria-
Science, 253;19-191.
L1V1NGSTONE, D.. and K. Krishnamoorlhy. 1982. Studies
the activity patterns of the larvae and adults of Aedes allw-picltts (Sktise) and Aedes vittalus (Bigot) of the scrub junglesof Palghal-Gap. India. .7 Bombay Nat Hist Soc, 82:30-37.
MACDONALD. W. mh.Afdcsacgypli in Malaya. 11. larval and
adult biology. Aim Trap Mrd I’arasilol, 50:399-414.MAK1YA, K. 1973. Population dynamics of mosquitoes in Na-
goya district. A. Larval populations of Aedes ulbopiclus
(Skuse) in cemetery in Nagoya City in 1967. Jpn Sanil
Zool. 24(2); 155-164.MANGIAPICO. J. A. 1971. Chikungunya virus infection and
transmission in five species of mosquito. Am Trap Med
Hyg, 20:642-645.
MARTEN, G. 1984. Impact of the copepod Mcwcyclops leuckarll
pllosa and the green alga Kilchnenetlti trregularies upon lar-
val Acdes albopiclus (Diptera; Culieidac). Kail Soc Vcc-ior
Ecol, 9(1):1-5.MARTEN, G. 1989a. Issues in the development of Cyclops for
mosquito control. In; M. IJren. J. Blok. and H. L. Manderson
(eds). Arhovirt^ Rwarch in Australia, pp. 159-164. Pro-ceedings Fifth Symposium. August 28-September 1. 1989.CSIRD Tropical Animal Science. Brisbane, Australia.
MAR’l’EN, G. 19891). A survey ofcyclopoid copcpods for control
ot Andes albopinus larvae. Bull Soc Vector Ecol. 14(2);232-23(>.
MARTEN, G. 1990a. Evaluation of cyclopoid copcpods for Acilcs
alimpiclits conliol in tires../ Am Mostf Control Aswc. 6:681-688.
MARTEN. G. 1990h. Elimination of Acdt’s albopiclus from tire
piles by introducing Macrocychps alhidtis (Copepoda, Cy-clopidac)../ Am Most/ Control Aswc, 6:689-693.
MARTEN, G., W. Chc, and E. Bordes. 1993. Compatibility of
cyclopoid copepods with mosquito insecticides../ Am Most]Control Aswc, 9(2):150-154.
MATSUZAWA. H., and N. Kilahara. 1966. Some knowledgethe biology of Aide’s albopictus Skuse..//? J Sanil Zool, 7:
232-235"MATT1NGLY. P. F. 1957. Gcnclical aspects of the Avdes mgypti
problem. Taxonomy and bionomics. Ann Tlop Med Para-sitol. 51:392-408.
METSEI.AAR, D.. C. R. Graingcr, K. G. Dei, D. G. Reynolds,M. Pudncy, C. 1. U’akc. R^ M. lukci, R. M. D’Offray. andD. Simpson. 1980. An outbreak of type dengue fever in
the Seychelles, probably transmitted b; Acdes alboplctus(Skuse). Hull World Health Organ, 58:937-943.
MILLER, B., and M. Rallinger. 1988. Aedes aibopictus mosqui-into Brazil: competence for yellow fever and den-
gue virus. Trails It fioc Trap Med Hya, 82:476-477.MILLER. B.. C. Milchcll, and M. Ballingcr. 1989. Replication,
tissue tropisms and transmission of yellow fever virus in Ae-dcs albopKliis. Trans R Soc Trap Uvd Hys, 83(2):252-255.
MITCIIELL, C. 1983. Mosquito competence and arbovi-In: K. F. Harris. Curiem Topics Vector Research,
Vol. 1., pp. 63-92. Pracger, New York.MITCHELL, C. 1991. Vcctoi competence of North and South
American strains of Ac. albopicius for certain arhoviruscs: Areview. Am Mos<j Control Aswc, 7(3):446-451.
MITCHELL, C., and D. Gublcr. 1987. Vector competence of geo-graphic stiains ol Aedes alhopiclus and Aeih", polynefiien’ii’,and certain other Aedes (Slegomyia) mosquitoes for RossRiver virus../ Am Mosq Control Assoc. 3(2):142-147.
MITCHF.LL. C., and B. Miller. 1990. Vertical transmission of
dengue viruses by strains of Ac. albopictu’i recently intro-duced into Brazil. Am Mosif Conlml Aswc, 6(2):251-253.
MITCHELL. C., B. Miller, and D. Gublcr. 1987. Vector compe-of Ac. aihopiftii-i from Houston. Texas for dengue
rolypcs 4, yellow fever and Ross River viruses. ./ Am.Vostj Conlml Assoc. 3(3):460-465.
MITCHELL, C., G. C. Smith, and B. R. Miller. 1990. Vectorcompetence fltAetit’s alhopcccus for newly recogni/.ed Bun-yavirus from mosquitoes collected in Potosi, Missouri../ AmMosq Control Assoc. 6:523-527.
MITCHRLL, C.. M. Nicbylski, G. Smith. N. Karabalsos, D. Mar-tin, J. P. Mutchi, G. B. Craig, and M. Mahler. 1992. Isolation
45

of Eastern equine encephalitis virus from Ac. (tlhopictus in
Florida. Science, 257:526-527.M1YAGI. I. 1972. Feeding habits of Japanese mosquitoes
cold-blooded animals in the laboratory. Trop Med, 14(4):203-217.
M1YAGI. I.. T. Toma, and S. Ilia. 1983. Studies the mosquitoesin Yacyama Islands, Japan. 8. On the mosquitoes collectedin Yonaguinijima. Jpn Sanil Zool. 34(1):1-6.
MOGI, M., and N. Yamanura. 1981. Estimation of the attraction
range of human bail for Aedes albopictus (Diptera, Culi-
cidae) adults and its absolute density by removalmethod applicable population with immigrants. Res PopEcol, 23(2):328-343.
MONATH, T. 1985. Flaviviruscs. Ill: B. N. Fields al. (cds).Virology, pp. 955-1004. Raven Press. New York.
MONATH, T. P. 1986. Aedes alhopictus, exotic mosquitoin the United States. Aim Intern Mcd, 105(3):449-451.
MOORE, C. G., and B. Fisher. 1969. Competition in mosquitoes:Density and species ratio, effects growth, mortality, fe-
cundity and production of growth retardant. Ann Entomol SocAm, 62:1325-1331.
MOORE, C. G., D. Francy. D. Eliason, and T. P. Monath. 19S8.Aedes albopicttts in (he United States: Rapid spread of po-tential disease vector. Am Mosq Control Assoc, 4:356-361.
MORI, A. 1979. Effects of larval density and nutrition
attributes of immature and adult Aedes albopictits. Trap Med,2I(2):85-103.
MORI, A., and Y. Wada. 1977. The gonotrophic cycle of Aedesalbopiclus in the field. Trop Mcd, 19(3-4):141-146.
MORI, A., and Y. Wada. 1978. The seasonal abundance of Aedes
albopiclus in Nagasaki. Trop Med, 20(1):29-37.MORI, A., T. Oda, and Y. Wada. 1981. Studies the egg dia-
pause and overwintering of Aedes albopiclus in Nagasaki.Trop Med. 23(2):79-90.
MUNSTERMANN, L., and D. Wesson. 1990. First record ofAscogregarina taiwanensis (Apicomplcxa: Lccudinidae) inNorth American Aedes albopictus.J Am Mosq Control Assoc,6(2);235-243.
NASCI, R., C. Hare, and F. Willis. 1989. Interspecific matingbetween Louisiana strains of Ae. alhopiclus and Ac. aegypliin the field and the laboratory. Am Mosq Control Assoc,5(3):416-421.
NAWROCK1, S., and W. A. Hawlcy. 1987. Estimation of northernlimits of distribution of Aedes albopictits in North America.
Am Mosq Control Assoc, 3(2):314-317.NENG, W., X. Yan, H. Fuming, and C. Dazong. 1993. Suscepti-
bility of Aedes albopictus from China and mechanism ofDDT resistance. .7 Am Mosq Control Assoc, 9(4):394-397
NEW ORLEANS MOSQUITO CONTROL BOARD. 1987. An-nual Report. City of New Orleans.
NEW ORLEANS MOSQUITO CONTROL BOARD. 1988. An-nual Report. City of New Orleans.
NEW ORLEANS MOSQUITO CONTROL BOARD. 1989. An-nual Report. City of New Orleans.
NEWKIRK. M. R. 1947. Observations Megarhinus splendensWiedemann with reference its value in biological controlof other mosquitoes (Diplera: Culicidae). Am Encoinot Soc,40:522-527.
NIEBYI-SKI, M. L. 1992. Bionomics oCAedes albopiclus (Skusc)in Potosi, Missouri. Department of Biological Sciences,
University of Notre Dame, Notre Dame, Indiana. Doctoral
dissertation.NIEBYLSKI, M. L., II. M. Savage, R. S. Nasci, and G. B. Craig,
Jr. 1994a. Blood hosts of Aedes albopiclus in the UnitedSlates. Am Mosq Control Assoc, 10(3):447-450.
NIEBYLSKI, M. L., and G. B. Craig, Jr. 19941). Dispersal andsurvival oiAedes albopictus scrap lire yard in Missouri.
Am Mosq Control Awe, 10(3):339-343.NIKLASSON. B. 1989. Sindbis and Sindbis-like viruses. In: T. P.
Monath (ed). The Athoviruses: Epidemiology and Ecology,Vol. IV, pp. 167-176. CRC Press, Boca Raton, Florida.
O’MEARA, G.. and A. Gellman. 1991. ’the Asian tiger mosquitoin Florida. Florida Mosquito Control Factsheet. IFAS-Uni-
versity of Florida and Stale of Florida Department of Healthand Rehabilitative Services. Office of Entomology Services,December 1991.
O’MEARA, G., L. Evans, Jr., and A. Gellman. 1992. Reduced
mosquito production in cemetery with copper liners.
Am Mosq Control Assoc, 8:419-420.
O’MEARA, G., A. Octtman, L. Evans, and G. Curtis. 1993. The
spread of Ac. albopictus in Florida. Am Entomol, 39(3): 163-172.
PAN AMERICAN HEALTH ORGANIZATION. 1987. Aedes al-
hopiclus in the Americas: Plan of Action. PAHO, Washing-ton, D.C. Document CE99/15 and Appendix.
PAN AMERICAN HEALTH ORGANIZATION. 1992. Dengueand dengue hacmorrhagic fever in the Americas: Anview of the problem. Epidemic! Bull, 13(1); 1-2.
PANT, C. P., S. Jatanascn, and M. Yasuno. 1973. Prevalence ofAedes aegypli and Aedes albopiclus and observations the
ecology of dengue haemorrhagic fever in several ofThailand. South Asian Trop Med Public Health, 4:113-121.
PAR1GOT, I’.. L. Calhciros, C. Rodopiano, and M. Lima. 1986.Reiacao de Municipios Positives para Aedes albopiclus, parssludo. SUCAM-Brazil, Oclober 1986.
PATTERSON, J., and J. Duman. 1978. The role of the thermal
hysteresis factor in Tenebrio molitor larvae../ Exp Biol, 74:
37-45.
PETERSEN, J., and II. Chapman. 1979. Checklist of mosquitospecies tested against the nemalode parasite Romanomcrmisculkhwrax. Med Enlomol, 15:468-471.
PINHEIRO, F., and J. LeDuc. 1989. Mayaro virus disease. In:T. P. Monalh (ed). The Arhoviruscs: Epidemiology and
Ecology, Vol. Ill, pp. 138-149. CRC Press. Boca Ralon,Florida.
PINHEIRO, F., R. Frcitas, J. Travassos da Rosa, I. B. Gabbay, W.Mcllo, and J. LeDuc. 1981. An outbreak of Mayaro virus in
Belterra, Brazil. Clinical and virological findings. AmImp Med Hyg, 30(3):674-681.
PINHEIRO, F., A. Travassos da Rosa, M. L. Gomes, J. LeDuc,and A. Hoch. 1982. Transmission of Oropouche virus from
hamster by the midge Culicoides paraensis. Science,215:1251-1253.
POLOVODOVA, V. P. 1949. The determination of the physiolog-ical age of female Anopheles by the number of gonotrophiccycles completed. Medskaya Parasil, 10:387-396.
PUMPUNI, C. 1989. Factors influencing pholoperiodic control ofegg diapausc in Aedes tilbopicfus (Skusc). Department of Bi-
ological Sciences, University of Noire Dame, Notre Dame,Indiana. Doctoral dissertation.
46

PUMPUNI, C. B.. J. Knepler, and G. B. Craig, Jr. 1992. Inllucnce
of temperature and larval nutrition (he diapausc inducingphoioperiod of Aedes albopictus. J Am Mosq Control Assoc,8(3):223-227.
01U, R, .H. Zhang, I.. Sliao. X. Li, H. Luo, and Y. Yu. 1981. Sludies
the rapid detection of dengue virus by immunofluorescenee
and radio-immunoassay. Chin MedJ, 94:653-658.RAI, K. S. 1986. Genetics of Andes albopictus. I Am Uosq Con-
trol Assoc. 2(4):429^36.RAI. K. S. 1991. Aedes ulbopiclus in the Amcricas. Aim Rev En-
tomol, 36:459-484.RAI. K. S., D. Pashlcy, and L. Munslermann. 1982. Genetics of
speciation in Aedinc mosquitoes. In: W. Sleincr, W. Tabach-
nick. K. Rai, and S. Narang (cds). Recent Developments in
the Genetics of Insect Disease Vectors, pp. 84-129. Stipes
Publishing, Champaign, Illinois.
RAVAL1.BC, M., A. Vcy, and G. Riba. 1989. Infection at Aedes
albopictus by Tolypocladium cylindrosporum, Invertebr
Palhol, 53(1):7-11.RICKLEFS, R. 1979. Ecology. Chiron Press, New York.
ROBERT, L., and J. Olson. 1989. Susceptibility of female Ac.
ulbopiclus from Texas commonly used adulticidcs. 7 Am
Mo’.q Control Assoc, 5:251-253.
ROBERTS, D. W., K. A. Daoust, and S. P. Wraight. 1983. Bib-
liography pathogens of medically important arthropods:198]. World Health Organization, Geneva. VBC/83.1.
ROSEN, L. 1980. Carbon dioxide sensitivity in mosquitoes in-
fected with sigma, vesicular stomatitis and other rhabdovi-
Science. 207:989-991.
ROSEN. L., L. C. Rozeboom. W. C. Reeves, J. Szugrain, and
D. G. Gublei. 1976. A field trial of competitive displace-of Aedes polynesicnsis by Aedes albopictus Pa-
cific atoll. Am Trap Mod Hyg, 25:906-913.ROSEN. L., R. Tesh, J. C. Lien. and J. H. Cross. 1978. Trans-
ovarial transmission of Japanese encephalities virus byquitoes. Science, 199:909-911.
ROSEN. L., D. Shroycr. R. Tcsh, J. Frcicr, and J. C. Lien. 1983.Transovarial transmission of dengue viruses by mosquitoes:Aedes albopiclus and Aedes uegypli. Am Trap Mc’d Hyg,
32(5):1108-1119.ROSEN. L., L. Roscboom, D. Gnblcr. J. C. Lien, and B. Chan-
iotis. 1985. Comparative susceptibility of mosquito speciesand strains oral and parcntcral infection with dengue and
Japanese encephalitis viruses. Am Trap Mod Hyg. 34(3):603-615.
ROUBAUD, E. 1929. Cycle autogene d’attcntc generationshiv’irnalcs suraetives inapparentes chez Ie moustiquemun, Ciilcx pipiens I..CR Arad Sci (Paris), 188:735-738.
ROZEBOOM, E., and R. Bridges. 1972. Relative population den-
sities of Aedes albopiclus and A. guumensis Guam. Rull
World Health Organ, 46:477-483.ROZEBOOM, L., L. Rosen. and J. Ikda. 1973. Observations
oviposilion by Aedes (S.) albopictits Sknse and A. (S.)polynesiensis Marks in JMrd Entomol. 10(4):397-399.
RUDN1CK, A., and Y. C. Chan. 1965. Dcnguc-2 virus ill naturallyinfected Aedes albopiclus mosquitoes in Singapore. Science,
149:638-639.
RUSSELL, P.. and P. B. Menon. 1942. On the transmission of
Plasmodium gallinaa’um to mosquitoes. Am Trap Mod
Hyg. 22:559-563.
SABATANI, A., V. Raincri, G. Trovato, and M. Coluzzi. 1990.
Ac. itlbopiclus in Italy and possible spread of the species in
the Mediterranean I’arassilologia. 32(3):301-304.SASA, M. 1976. Human Fitiarisis. A Global Survey of Epide-
miology and Control. University of Tokyo Press. 819 pp.SAVAGE, H., V. Ezikc, A. Nwarlko. R. Spicgel. and B. Miller.
1992. First record of breeding populations of Ac. albopictusin continental Africa: Implications for arboviral transmission.Am Mosq Control Assoc. 8(1):101-103.
SAVAGE, H., M. Niebylski, G. Smith, C. Milchell. and G. B.
Craig. 1993. Host-feeding patterns (}t Aedes albopiclus (Dip-Culicidac) temperate North American site../ Mod
Enlomol, 30(1):27-33.SCANLON, J., and S. Esah. 1965. Distribution in altitude of
quitoes in northern Thailand. Mosq News, 26(2); 137-144.SCOLES, G., and G. Craig. 1993. Variation in susceptibility
Dirofilaria immitis among U.S. strains of Aedes albopictus.Vector Control Bull North Central Stales. 2(1);98-103.
SCOTT, T W., and S. C. Weaver. 1989. Eastern equine cnccph-alomyelitis virus: epidemiology and evolution of mosquitotransmission. Adv Virus Res, 37:277-328.
SCOTT, T, L. Lorenz, and S. Weaver. 1990. Susceptibility ofAr.
albopiclus infection with equine encephalomyelitisvirus../ Am Mosq Control Assoc, 6:274-278.
SERVICE, M. W. 1976. Mosquito Ecology: Field Sampling Melh-
ods. John Wiley and Sons, New York-Toronto. 583 pp.SHENG, T, and I. Wu. 1951. An ecological study of mosquitoes
in Wuhan Bull Enlomol Res, 42:527-533.SHOI’E, R. 1985a. Alphaviruscs. In; B. N. Fields al. (eds).
Virology, pp. 931-953. Raven Press, New York.
SHOPE, R. 1985b. Bunyavirus. In: B. N. Fields al. (eds). Vi-
rology, pp. 1055-1079. Raven Press, New York.
SHROYER, D. A. 1986. Aedes atbopictus and alboviruscs: Acise review of the literature. Am Mosq Control Assoc, 2(4):424-428.
SHROYER, D. 1990. Vertical maintenance of dcngue-1 virus in
sequential generations oiAedes albopiclus. Am Mosq Co-troiA.n-oc,6(2):312-314.
S1LER, J., M. Hall. and A. Hilchens. 1926. Dengue; Its history,
epidemiology, mechanism of transmission, etiology, clinical
manifestations, immunity and prevention. Philip? ./ Sci, 29:
1-302.
SIMMONS, J. S., F. H. K. Reynolds, and V. H. Cornell. 1936.
Transmission of the virus of equine encephalomyelitis
through Aedes albopiclus (Skuse). Am Trop Med, 16:289-
302.
SMITH, C. E. G. 1956. The history of dengue in tropical Asia
and its probable relation the mosquito Aedes aegypti. Am
Trap Mod Hyg, 59:3-11.
SMITH, G., and D. Francy. 1991. Laboratory studies of Brazil-
ian strain oiAedes atbopiclus potential vector of Mayaroand Oropouche viruses. Am Mosq Control Assoc, 7(1):89-93.
SOEK1MAN, S., M. Subatyio, S. Adipoetro, H. Yamanishi, and
T. Malsumura. 1984. Comparative studies the biology of
Aedes aegypti (Linnaeus, 1762) and Aedes alhopictus (Skusc,1895) in condition. KMRAnn, 4:143-151.
SOMME, L. 1966. The ctfecl of temperature, anoxia injectionof various substances hemolymph composition and
47

percooling in larvae oi Anagosia kuehnielle (Zcll.). Insect
Physiol, 12:1069-1083.
SOMME, L. 1982. Supercooling and winler survival in terrestrial
arlhropods. Comp Riochem Physiol, 73A(4):519-543.SOTA, T., M. Mogi, and E. Hayamizu. 1992. Seasonal distribution
and habitat selection by Ae. albopictus and Ae. riversi (Dip-Culicidae) in Nonheni Kyushu, Japan. McdEnlomol,
29(2):297-304.SPRENGER, D., and T. Wuithiranyagool. 1986. The discovery
and distribution of Aedes alhopictus in Harris County, Texas.Am Mosq Control Assoc, 2:217-219.
STOjANOViCH, C., and H. G. Scott. 1965. Illustrated KeyAedes Mosquitoes of Vietnam. USDHEW, Centers for Dis-
Control, Atlanta, Georgia. 34 pp.STREIT, T. 1994. Stabilized La Crosse virus infection and
mission in Acdcs allwpictus: urban threat. Doctoral dis-
sertation, University of Notre Dame, Notrc Dame, Indiana.
STREIT, T., and P. Grimsiad. 1990. Vector competence of Ac.
allmpictus for La Crosse encephalitis virus. Proc Ohio MosqControl Assoc. 19:45-50.
SUCHARIT. S., W. Tumrasuin, S. Vutikcs. and S. Viraboonchai.
1978. Interaction between larvae of Ac. aegypti and Ac. al-
bopictus in mixed experimental populations. Southeast Asian
Trap Med Public Health, 9:93-97.
SUDIA, W. D., V F. Newhousc, C. Calisher, and R. Chamberlain.1971a. California group arboviruscs: Isolations from
quitoes in North America. Mosq News. 31(4):576-599.SUDIA, W. D., V. F. Newhousc, and B. E. Henderson. 1971b.
Experimental infection of horses with three strains of Vene-zuelan equine enccphalomyelitis. 11. Experimentalstudies. Am Epidemiol. 93:206-211.
SULLIVAN, M., D. Gould, and S. Maneechai. 1971. Observationsthe host range and feeding preferences of Aedes allw-
pictus (Skuse)../ Med Entomol. 8(6):713-71(>.SUPERINTENDENCE DE CAMPANHAS DE SAUDE PUB-
LICA. 1989. Resumo dos principais morfologicosdifcrenciais do Ac’, aegypti do Ac. alhopjclus. Ministcrio da
Saude. Brasilia. Brazil.
SURTEBS, G. 1966. Aedes (Slegomyia) albopictus: A Summaryof Present Knowledge with Particular Reference Compe-tition with Aedes aegypti. World Health Organization, Ge-
WHO/Veclor Control/66.195.SURTF-ES, G. 1970. Mosquito breeding in the Kucking Area, Sa-
rawak. with special reference the epidemiology of denguefever. Mod Emomol, 7(2):273-276.
SWEENEY, K. 1993. Organophosphorous insecticide susceptibil-ity of mosquitoes in Maryland, 1985-1989. Am Mosq Con-liol Am-oc, 9(1):8-13.
TEMPELIS. C. H., R. 0. Hayes, A. D. Hcss, and W. C. Reeves.1970. Blood-feeding habits of four species of mosquito found
in Hawaii. Am Trap Med ffy^ 19(2):335-34LTESH, R. B. 1975. Multiplication of phleboiomus fever group
arboviruses in mosquitoes after intrathoracic inoculation.
Med Entomol. 12:1-4.
TESH, R. 1980. Experimental studies the tran&ovarial
mission of Kunj’in and San Angelo viruses in mosquitoes.Am Trap Med Hyg, 29(4):657-666.
TESH, R.. and D. Gubler. 1975. Laboratory studies of transovarial
transmission of La Crosse and other arboviruses by Aedes al-
hopictus and Culex falcons. Am Imp Mod Hyg, 24:876-880.
TESH. R. B., D. J. Gubler. and L. Rosen. 1976. Variation amonggeographic strains of Aedes albopictus in susceptibility infec-
tion with Chikungunya virus. Am Trap Med Ilyg, 25:326-335.
TESH, R., and D. A. Shroycr. 1980. The mechanism of arbovirustransovarial transmission in mosquitoes: San Angelo virus in
Aedes a\bopictus. Am Trop Med Hys, 29(1):394-1404.TOMA, T, S. Sakamato, and Migagi. 1982. The seasonal ap-
pearance of Acdcs albopictus Okinawajima, the RyukyuArchipelago, Japan. Mosq News. 42(2):179-185.
TOMORI, 0., and T. H. G. Aitken. 1978. Onmgo virus:
mission studies with Aedes alhopictus and Aedes aegypti(Diptera: Culicidae). Med Entomol, 14:523-526.
TURELL, M. 1988. Horizontal and vertical transmission of vi-
by insect and tick In: T. P. Monath (ed). The
Arboviruses: Epidemiology and Ecology, Vol. I, pp. 127-152. CRC Press, Boca Raton, Florida.
TURELL, M. J.. and J. R. Beaman. 1992. Experimentalmission of Venezuelan equine cncephalomyelilis by strainof Aedes albopictus (Diptera: Culicidae) from New Orleans,Louisiana../ Med Entomol, 29(5):802-805.
TURELL, M. J., C. L. Bailey, and J. R. Beaman. 1988. Vectorcompetence of Houston. Texas, strain Acdcs albopictusfor Rift Valley fever virus. J Am Mosq Control Assoc, 4:94-
96.
TURELL, M. J., G. V. Ludwig, and J. R. Beaman. 1992a. Trans-mission of Venezuelan equine enccphalomyelilis by Acdcs
solicitous and Aedes taeniorhynchus (Dipiera; Culicidae).Med Entomol, 29:62-65.
TURELL, M. J., J. R. Reamaii. and R. F. Tamimiriello. 1992h.
Susceptibility of selected strains ofAi’rfc’A’ aegypti and Aedcv
albopictus (Diptera: Culicidae) Chikungunya virus../ Med
Entomol, 29(1):50-53.TURELL. M. J., J. R. Beaman, and G. W. Neely. 1994. Experi-
mental transmission of equine encephalitis virus b>strains of Acdcs allwpictus and Aedes tuaniorhynchus (Dip-
Culicidae). Med Eiilomol, 31(2):287-2()0.TWOHY, D. W., and L. E. Rozeboom. 1957. A comparison ot
food in autogenous and anauiogenous Cuh-jc pipicnspopulations. Am Hyg, 65:316-324.
UDAKA, M. 1959. Some ecological Aedes alhopictiisSkuse in Foochow. Ada Entomol Sin. 11(4):357-362.
VYTH1LINGAM, I., G. Chiang. and C. Amatachaya. 1992. Adul-
licidal effect of cyfluthrin against mosquitoes of public health
importance in Malaysia. Southeast Asian Trop Med Public
Health, 13{}.):m-\\5.WALKER, D.. R. Copeland, S. Paulson, and I- Munstermann.
1987. Adult survivorship, population density and body size
in sympalric populations of Ac. triseriatu.s and Ae. hender-soni (Dipiera; Culicidae). Med Enlomol, 24:485-493.
WALTON, T.. and M. Grayson. 1989. Venezuelan equine enceph-alomyelitis. In: T. P. Monath (cd). The Arboviruses’ Epide-miology and Ecology. Vol. TV, pp. 203-225. CRC Press,Boca Raton, Florida.
WANG, K. C. 1962. One year’s observation the ecology ofAcdcK alhopiclus Skuse in Foochow. Acta Entomol Sin,
11(4):357-362.WANG, R. L. 196f>. Observations the influence of photopcriod
egg diapausc in Aedes albopictus Skuse. Ada Entomol
Sin, 15(1):75-77.
48

WATTS. D. M.. J. W. LcDuc, C. I.. Bailey, J. M. Dalrymple,and T. P. Gargan II. 1982. Scrological evidence ot James-
Canyon and Keystone virus infection in vertebrates ofthe Delmarva peninsula. Am Trap Mcd Hyg, 31:1245-
1251.
WESSON, D. 1991). Susceptibility organophosphale insecti-
cides in larval Aedes ullmpictas. Am Mosq Control A.’.wc,6:258-264.
WESTAWAY. B. G., M. Brinlon, S. Gaidamovisch, M. IIorzinck,A. Igarashi, L. Kaariaiiicn, D. Lvov, J. Porterlield, P. Rus-sell, and D. Trent. 1985. Flaviviridae. Inicrviroloffy, 24:
183-192.WOMACK, M. 1993. Dislribiition, abundance and bionomics of
Aedes albopicltts in southern Texas. Am Mosq Control A.v-
9:367-369.WORLD HF.ALTII ORGANIZATION. 1980. Dengue in the Peo-
ple Republic of China, South-East Asian and Western Pa-
cific Regions. Dengue Newsletter, 6(1,2):14.WORLD HEALTH ORGANIZATION. 1985. Arlhiopod-borne
and Rodent-borne Viral Diseases. World Health Organiza-tion, Geneva. 119 pp. WHO Technical Report Series 719.
WORLD HEALTH ORGANIZATION. 1986a. Dengue Haemor-rhage Fever: Diagnosis, Treatment and Control. World
Health Organi/ation, Geneva. 58 pp.WORLD HEALTH ORGANIZATION. 1986b. Dengue Hwmar-
rhcigic Fever Control Programme in Singapore: A Case
Study the Successful Control o/Aedes aegypli and Aedes
albopictus Uling Mainly Environmental Measure’, Part
of Integrated Vector Control. World Health Organiy..ition. Ge-
pp. 1-38. WHO/VBC/86.928.XAV1ER, G., D. Ncvcs. and R. Silva. 1991. Biological cycle o[
Af. albopicltts (Diptera: Culicidae) laboratory conditions.
Rev Bra!, Biol, 51(3):647-650.YAMANISHI. H., E. Konishi, T. Sawayama, and T. Malsumiira.
1983. The susceptibility of mosquitoes Chikungunya
virus. Jpii Sanil Zool, 34:229-233.
YAP, H. H. 1975. Preliminary report the colour preference for
oviposilion by Aedes albopietu!, (Skuse) in the field. South-
Asian Trap Med Public Health, (6):451.YOUNG, N.. and K. Johnsoii. 1969. Antigenic variants of Venc/uc-
lan equine encephalitis virus: Theii geographic distribution and
epidemiologic signilicance. Am Epidemiol. 89:286-307.
49



^165144

/^^/ 165144
Fagerstonc, K. A.; Ramey, C. A.; Keith, J. 1995. ABSTRACT. Mongoose management to
protect endangered species in Hawaii. In: The Wildlife Society 2d Annual Conference Abstracts;12-17 September 1995; Portland, OR. Bethcsda, MD; The Wildlife Society: 41-42.
FAGERSTONE, KATHLEEN A., C. A. RAMEY, AND J. KEITH. Mongoosemanagement to protect endangered species in Hawaii. USDA APHIS Denver WildlifeResearch Center, Denver, CO 80225-0266 USA.
Mongooses (Herpesles auropunctatus) were introduced into Hawaii in 1883 to controlintroduced rats and now occur on all main islands except Kauai. Whereas mongooses have not
limited rat populations, they have limited the productivity ofmany ground nesting avian species.They have also restricted the recovery of at least 8 species ofendangered Hawaiian birds byconsuming both their eggs and young. The Denver Wildlife Research Center (DWRC) beganinvestigations in Hawaii in 1984 to develop a management tool for mongoose control. Studies of
mongoose biology and toxicology revealed an extreme susceptibility to the anticoagulantdiphacinone. Using DWRC data, Bell Laboratories, Inc. obtained a state registration in 1991 foruse of diphacinone to control mongoose in bird nesting areas.