bruce ll a 201102
TRANSCRIPT

Microbiologia Clínica Veterinária VET 3225 ÁREA DE BACTERIOLOGIA 2011-2
UFRGS
Microbiologia Clínica Veterinária VET 3225 ÁREA DE BACTERIOLOGIA 2011-2
UFRGS
Gênero Brucella spp
Prof. Marcos JP Gomes DOENÇA DE BANG, ABORTO CONTAGIOSO, ABORTO INFECCIOSO, ABORTO ENZOÓTICO E "SLINKING OF THE CALF". Homem: FEBRE ONDULANTE, FEBRE DE MALTA OU DO MEDITERRÂNEO, FEBRE MALTESA, FEBRE DE GIBRALTAR ATUALIDADES Atualmente (2011), na “List of Prokaryotic names with Standing in Nomenclature” do pesquisador J.P. Euzéby e o site www.bacterio.cict.fr/b/brucella. Há citação de 10 espécies e nenhuma subespécie neste genro, entretanto existem espécies no gênero Brucella. São elas: Brucella abortus (Schmidt 1901) (Meyer and Shaw 1920).
Brucella canis (Carmichael and Bruner 1968) Brucella ceti (Foster et al. 2007) Brucella inopinata (Scholz et al. 2010) Brucella melitensis (Hughes 1893) (Espécie típica do gênero). Brucella microti (Microtus arvalis Scholz et al. 2008) Brucella neotomae (Neotoma lepida Stoenner and Lackman 1957) Brucella ovis (Buddle 1953). Brucella pinnipedialis (Foster et al. 2007) Brucella suis (Huddleson 1929,)
HISTÓRICO A primeira espécie do gênero Brucella foi isolada, em 1887, pelo Dr. David Bruce, em amostras (baço) colhidas em necropsia de militares que morreram vítimas dessa enfermidade nas costas do Mediterrâneo, chamada de Febre de Malta. O organismo foi mais denominado Micrococcus melitensis e mais tarde denominado de Brucella melitensis. Em 1897, um veterinário dinamarquês chamado Bernard Bang, isolou e denominou de Bacillus abortus e, mais tarde foi conhecida como Brucella abortus de um feto bovino abortado. A B. suis foi isolada, em 1914, por Jacob Traum, de um leitão abortado. Na Nova Zelândia, uma doença genital de ovinos foi associada ao organismo do gênero Brucella spp por M.B. Buddle e Boyes, em 1956. Um organismo, provavelmente idêntico ao isolado na Nova Zelândia foi isolado na Austrália de carneiros infectados por Simmons e Hall, em 1953. A B. neotomae foi isolada, pela primeira vez, em roedores do deserto de Utah em 1957, por Stoenner e Lachaman, nos Estados Unidos. A B. canis foi isolada em 1966, nos EUA e, posteriormente descrita por Carmichael, em 1969. As espécies B. ovis e a B. canis são mais adaptadas aos seus hospedeiros do que a B. abortus, B. melitensis ou a B. suis.
Em 1994, novas espécies do gênero Brucella foram isoladas e identificadas de pinípedes (focas), nas costas da Escócia, por Ross e colaboradores. Em 2007, os novos isolados foram incluídos no gênero Brucella como o nome de B. pinnipedialis (Foster et al.
Generated by Foxit PDF Creator © Foxit Softwarehttp://www.foxitsoftware.com For evaluation only.

Microbiologia Clínica Veterinária VET 3225 ÁREA DE BACTERIOLOGIA 2011-2
UFRGS
Microbiologia Clínica Veterinária VET 3225 ÁREA DE BACTERIOLOGIA 2011-2
UFRGS
2007). Nos Estados Unidos de um cetáceo (golfinho) capturado, por Ewalt e colaboradores, em 1994. Mais tarde, esses isolados foram incluidos no gênero Brucella com o nome de B. ceti (Foster et al. 2007). As amostras isoladas desses mamíferos marinhos e os testes sorológicos neles realizados demonstraram que a infecção brucélica ocorre em um grande numero de espécies marinhas.
Em 2008, nos Estados Unidos (Oregon), Barun Kumar De e colaboradores descreveram as características microbiológicas, bioquímicas e moleculares de uma linhagem de Brucella incomum (BO1) isolada de uma prótese de seio (silicone) de uma senhora com 71 anos de idade com sinais clínicos compatíveis com brucelose. A análise inicial, incluindo teste de susceptibilidade bioquímica, TSA, análise de ácidos graxos e análise molecular baseadas na reassociação do ADN-ADN e presença de múltiplas cópias de IS711 sugeriram que o isolado era uma “brucella like organism”, mas a determinação da espécie, através de dados moleculares baseados no sequencionamento do 16S rARN e na análise sequencial multilocus, demonstraram que a linhagem BO1 era uma cepa incomum, atípica e nova de Brucella, pois não está relacionada as atuais espécies descritas. Em 2010 ela foi incluída na “List of Prokaryotic names with Standing in Nomenclature” com a denominação de B. inopinata (brucela não esperada).
Em 2009, Nos Estados Unidos (Texas), Natalia E. Schlabritz-Loutsevitch e colaboradores descreveram e relataram, pela primeira vez, um novo isolado de Brucella associado a dois casos de morte neonatal em primatas (babuínos). As amostras do útero isoladas foram caracterizadas, utilizando os testes bioquímicos tradicionais, PCR e sequenciamento de multilocus. Os isolados se assemelham morfologicamente a Brucella embora suas caracteristicas não fossem consistentes com qualquer espécie descrita.
No Brasil, a brucelose foi detectada, pela primeira vez, por Gonçalves Carneiro, em 1913, que relatou um caso de brucelose humana. Desde lá, vários inquéritos epidemiológicos revelaram a presença da enfermidade nos animais domésticos em todo o país. A brucelose bovina causada pela B. abortus é a mais prevalente das infecções brucélicas no Brasil, seguida da B. suis em suínos. A B. melitensis e a B. neotomae não foram isoladas/identificadas no país (Poester et al., 2002).
Em 1977, um levantamento nacional estimou a prevalência para cada região do país. Neste estudo a região Norte obteve uma prevalência de 4,1%; a região Nordeste com 2,5%; a região Centro-Oeste com 6,8%; Sudeste com 7,5% e região Sul com 4%. (Brasil, 1977). Em 2001, o Ministério da Agricultura, Pecuária e Abastecimento (MAPA) instituiu o Programa Nacional de Controle e Erradicação de Brucelose e Tuberculose (PNCEBT) que definiu estratégias no controle da brucelose bovina no Brasil (Instrução Normativa 2004). Os inquéritos sorológicos realizados no período compreeendido entre 2001 e 2004, nos 13 estados (BA, ES, GO, MG, MT, PR, SC, RJ, RS, SP, SE, TO e DF) constataram que a infecção está disseminada em todas as áreas estudadas e que a situação é desigual entre os estados e mesmo entre regiões de um mesmo estado. Detectou-se uma tendência de crescimento da brucelose bovina no país (Centro-Oeste / Norte) com elevação nos estados com muita tradição na pecuária de corte. Mato Grosso apresentou aumento da prevalência quando comparado ao último levantamento da situação nacional realizado em 1975 (Anselmo e Pavez, 1977). Outros estados diminuíram sua prevalência, especialmente os
Generated by Foxit PDF Creator © Foxit Softwarehttp://www.foxitsoftware.com For evaluation only.

Microbiologia Clínica Veterinária VET 3225 ÁREA DE BACTERIOLOGIA 2011-2
UFRGS
Microbiologia Clínica Veterinária VET 3225 ÁREA DE BACTERIOLOGIA 2011-2
UFRGS
estados de Minas Gerais e Santa Catarina que apresentou os menores índices do Brasil (Lage et al. 2008). CARACTERÍSTICAS GERAIS DO GÊNERO Cocos, cocobastonetes ou bastonetes curtos com 0,5-0,7 de diâmetro e 0,6-1,5µm de comprimento; arranjos individuais, aos pares, cadeias curtas ou mesmo em pequenos grupos; acapsulados; não possuem coloração bipolar verdadeira; não possuem esporos; Gram negativas; imóveis; não flagelados; aeróbias, possuindo metabolismo do tipo respiratório; possuem sistema citocromo, e como aceptor final de elétrons, oxigênio ou nitrato e, sendo assim nitrato redutase é produzida. Muitas amostras requerem CO2 suplementar para seu crescimento, especialmente no cultivo primário. As colônias, em agar dextrose ou outro meio sem sangue são transparentes, elevadas, convexas com bordos inteiros, lisos e com superfície brilhante. Possuem cor de mel quando iluminadas com luz transmitida. Variantes rugosas de espécies lisas existem. Existem também espécies rugosas estáveis com uma preferência por determinados hospedeiros (caninos e ovinos).
A temperatura ótima de crescimento é de 37ºC. O crescimento ocorre entre 20ºC a 40ºC e o pH ótimo é entre 6,6 a 7,4; catalase positiva; geralmente oxidase positiva; quimiorganotróficas. A maioria das cepas requer meios de cultivo seletivo e complexo, contendo aminoácidos, tiamina, nicotinamida e íons de magnésio. Algumas cepas podem ser induzidas ao crescimento em meio mínimo, contendo sais de amônio como única fonte de nitrogênio. O crescimento é promovido pela adição de soro ou sangue, entretanto hemina (Fator X) e nicotinamida adenina dinucleotídio (NAD Fator V) não são essenciais.
Produz ácido, mas não de carboidratos em meios convencionais, exceto para a B. neotomae. Não produzem indol; não liquefaz a gelatina ou soro coagulado; não lisam hemácias; não produzem metil carbinol (Teste de Voges-Proskauer); vermelho de metila negativo; possuem antígenos intracelulares específicos para o gênero; são parasitos intracelulares facultativos. São agentes infecciosos transmissíveis para muitas espécies animais, e o homem. TAXONOMIA Classicamente, o gênero Brucella continha seis espécies, mas continua evoluindo. Atualmente, novas espécies foram incluídas. Cada uma das espécies possui seus hospedeiros preferenciais: B. abortus (bovinos); B. melitensis (caprinos e ovinos); B. suis (suínos); B. canis (caninos); B. ovis (ovinos); B. neotomae (rato do deserto, Neotomae lepida); B. microti (camundongo do campo, Microtus arvalis); B. ceti (cetáceos); B. pinnipedialis (penípedes) e a B. inopinata (homem). Todas são importantes patógenos para os animais (domésticos e silvestres) e o homem, causando uma doença que é denominada genericamente de brucelose. Exceto a B. neotomae e a B. ovis, todas as demais são capazes de infectar o homem. Novas espécies como B. pennipedialis dos penípedes; a B. ceti dos cetáceos e a B. microti do roedor Microtus arvalis, e a B. innopinata isolada do homem foram incluídas recentemente no gênero. As alterações causadas pela brucelose são encontradas nos órgãos reprodutores e sistema microcítico fagocitário (retículo-endotelial). As lesões no trato reprodutor incluem
Generated by Foxit PDF Creator © Foxit Softwarehttp://www.foxitsoftware.com For evaluation only.

Microbiologia Clínica Veterinária VET 3225 ÁREA DE BACTERIOLOGIA 2011-2
UFRGS
Microbiologia Clínica Veterinária VET 3225 ÁREA DE BACTERIOLOGIA 2011-2
UFRGS
placentites e/ou abortamentos (bovinos, ovinos, suínos e caprinos) que levam à infertilidade, associadas às perdas econômicas importantes.
A facilidade com que algumas espécies do gênero Brucella podem ser transmitidas (direta ou indiretamente) aos animais e ao homem mostra a importância do controle desta enfermidade.
As características morfotintoriais e culturais não são suficientes para diferenciar as espécies novas e clássicas, assim como os seus vários biovares. Pode não ser correto associar o isolamento de uma espécie ou biotipo a um hospedeiro, muito embora, cada espécie tenha certa predileção por certos hospedeiros, conforme o quadro I. Quadro I. As principais espécies de Brucella spp e seus hospedeiros preferenciais
Espécies Hospedeiro Preferencial Outras espécies _______________________________________________________________________________ Bovinos B. abortus B. melitensis** B. suis
Ovinos B. melitensis** B. abortus B. ovis (Epididimite) Caprinos B. melitensis** B. abortus
Eqüinos B abortus B. suis
Suínos B. suis B. melitensis** B. abortus
Caninos B. canis B. abortus B. melitensis** B. suis Homem B. abortus B. canis B. melitensis** B. suis B. inopinata** ?
B. ceti** B.pinnipedialis** ? Roedores B. neotomae** ?
(Neotoma lepida)
Camundongo campo B. microti** ? (Microtus arvalis)
Cetáceos B. ceti** ?
Penípedes B. pinnipedialis** ?
** Não há registro dessas espécies do gênero Brucella no Brasil, até o presente momento.
Generated by Foxit PDF Creator © Foxit Softwarehttp://www.foxitsoftware.com For evaluation only.

Microbiologia Clínica Veterinária VET 3225 ÁREA DE BACTERIOLOGIA 2011-2
UFRGS
Microbiologia Clínica Veterinária VET 3225 ÁREA DE BACTERIOLOGIA 2011-2
UFRGS
DIAGNÓSTICO LABORATORIAL O diagnóstico laboratorial é complexo, pois exige equipamento, meio e pessoal
qualificado. As características diferenciais entre diferentes espécies clássicas e seus biovares do gênero Brucella é mostrado no Quadro II abaixo Quadro II. Características diferenciais entre espécies/biovares clássicas do gênero Brucella __________________________________________________________________________________
C r e s c i m e n t o em c o r a n t e s A g l u t i n a ç ã o do s o r o Espécies Biovar CO2 H2S Tionina Fucsina básica A M R _________________________________________________________________________________ B. melitensis 1 n n + + n + n 2 n n + + + n n 3 n n + + + + n B. abortus 1 (+) + n + + n n 2 (+) + n n + n n 3 (+) + + + + n n 4 (+) + n (+) n + n 5 n n + + n + n 6 n n + + + n n 7 +/- + + + + + n 9 n + + + n + n B. suis 1 n + + n + n n 2 n n + n + n n 3 n n + + + n n 4 n n + (-) + + n 5 n n + n n + n B. canis n n + (n) n n + B. ovis + n + n n n + B. neotomae n + (n) n + n n
Obs. * n = negativo- + = positivo
Generated by Foxit PDF Creator © Foxit Softwarehttp://www.foxitsoftware.com For evaluation only.

Microbiologia Clínica Veterinária VET 3225 ÁREA DE BACTERIOLOGIA 2011-2
UFRGS
Microbiologia Clínica Veterinária VET 3225 ÁREA DE BACTERIOLOGIA 2011-2
UFRGS
Brucella abortus Prof. Marcos JP. Gomes BRUCELOSE BOVINA
INTRODUÇÃO O isolamento de Brucella abortus foi obtido nas diversas populações bovinas distribuídas pelo mundo, embora sejam raros em países onde programas de controle e erradicação foram bem sucedidos. MORFOLOGIA E COLORAÇÃO A B. abortus é um bastonete curto ou cocobacilo, medindo 0,5 a 0,7 por 0,6 a 1,5µm. Os bastonetes são tão curtos que podem ser confundidos com cocos. Eles estão presentes em arranjos individuais ou cadeias curtas. A B. abortus por ser um parasito intracelular facultativa e frequentemente encontrada em aglomerados ou em esfregaços de exsudatos. A B. abortus é Gram negativa, corando-se, com alguma dificuldade, pelos corantes comuns. Ela não é álcool ácido resistente, mas pode resistir à descoloração com alguns ácidos fracos; esta propriedade confere a base para algumas colorações especiais tais como a coloração de Köster em que o organismo cora-se de vermelho vivo. B. abortus não é móvel, não forma esporos e não possui cápsula bem desenvolvida. Entretanto, a presença desta cápsula pode ser demonstrada em cepas isoladas, utilizando-se corantes especiais. RESISTÊNCIA E SOBREVIVÊNCIA As espécies são inativadas pela pasteurização entre 10 e 15 segundos; são destruídas rapidamente pelos desinfetantes comuns como o cresol 3%; hidróxido de sódio a 2%; compostos de ortofenóis 3-5%; mercuriais e álcool 70%. O crescimento de B. abortus fora da célula dos mamíferos hospedeiros não tem importância na epidemiologia da doença, pois ela não se multiplica fora dos animais e somente persiste no ambiente. A viabilidade dela fora do hospedeiro é influenciada pelas condições ambientais. A sua viabilidade é aumentada em temperatura mais amena e umidade, sendo diminuída quando há alta temperatura, luz solar direta e dessecamento, como é demonstrado no Quadro II.
A resistência fora do corpo do hospedeiro é de aproximadamente: 5 dias à temperatura ambiente; 30-37 dias quando secas lentamente no solo; 75 dias no feto abortado em clima temperado. O tempo de sobrevivência nas fezes parece ter importância, especialmente em muitos sistemas produtivos. O tempo de sobrevivência das fezes líquidas varia, sendo dependente da temperatura de estocagem. Na temperatura de 45–50 ºC a sobrevivência da B. abortus é de 4 horas, enquanto que na temperatura de 15ºC é de aproximadamente 8 meses.
Generated by Foxit PDF Creator © Foxit Softwarehttp://www.foxitsoftware.com For evaluation only.

Microbiologia Clínica Veterinária VET 3225 ÁREA DE BACTERIOLOGIA 2011-2
UFRGS
Microbiologia Clínica Veterinária VET 3225 ÁREA DE BACTERIOLOGIA 2011-2
UFRGS
Quadro II. Sobrevivência da B. abortus, segundo o ambiente e o tempo Ambiente Tempo Luz direta 4,5 h Solo Seco 4 dias Úmido 66 dias Frio 151 – 185 dias Fezes Fluidas 8 –240 dias Altas Temperaturas 2 dias Urina 5 dias Água Tratada 5 – 114 dias Poluída 30 – 150 dias Feto à sombra 180 dias
OUTRAS FONTES DE INFECÇÃO O papel dos touros na transmissão da B. abortus tem sido objeto de investigações. A B. abortus pode causar orquite, epididimite e vesiculite seminal e a localização nesses lugares resulta na eliminação do agente pelo sêmen. Seu potencial de transmissão está associado ao método de reprodução (monta natural, I.A.) e, freqüentemente touros infectados permanecem férteis. Equídeos domésticos são susceptíveis à infecção com B. abortus havendo pouca evidência que eles possuam um papel significante na epidemiologia da doença. A B. abortus geralmente localiza-se na bursa, tendões, músculos e articulações assim como tecidos e trato reprodutivo. O achado clínico clássico é o abscesso fistulado denominado “Mal das cernelhas” ou “Mal das Cruzes”. CARACTERISTICAS CULTURAIS E BIOQUÍMICAS O crescimento pode ser aeróbio, mas muitas cepas necessitam tensão de CO2 para o seu crescimento, especialmente no isolamento primário (B. abortus e B. ovis). A B. abortus é catalase e oxidase positivas, produzindo H2S de proteínas ou peptídeos ricos em aminoácidos e enxofre. Geralmente produz urease.
Os biovares foram classificados segundo os testes: 1)suscetibilidade aos corantes; 2)necessidade de CO2; 3)produção de H2S e 4)presença do antígeno de superfície A ou M.
Generated by Foxit PDF Creator © Foxit Softwarehttp://www.foxitsoftware.com For evaluation only.

Microbiologia Clínica Veterinária VET 3225 ÁREA DE BACTERIOLOGIA 2011-2
UFRGS
Microbiologia Clínica Veterinária VET 3225 ÁREA DE BACTERIOLOGIA 2011-2
UFRGS
A B. abortus não é hemolítica, não liquefaz a gelatina e não produz ácido a partir da glicose ou outro carboidrato, possuindo um padrão de oxidação de substrato. O crescimento de B. abortus é incrementado pela adição de sangue ou soro. Meios com composição complexa como soro dextrose agar ou Albimi Brucella agar ou caldo devem ser utilizados no isolamento primário e/ou na manutenção de cepas.
Antimicrobianos podem ser adicionados ao meio básico (polimixina B 5.000 UI/L; bacitracina 25.000 UI/L; ciclohexamida 100 mg/L; ácido nalidíxico 5 mg/L; nistatina 100.000UI/L e vancomicina 20 mg/L), inibindo o crescimento de contaminantes em amostras de placenta, secreções vaginais, leite ou fluido do abomaso de fetos abortados. As colônias da B. abortus no isolamento primário são de crescimento lento e raramente visível antes das 48 horas. Elas atingem o crescimento máximo após 5 a 7 dias a 37ºC. A colônia isolada pode ser lisa, caracterizado por serem convexas; redondas com o bordo inteiro; podem estar dissociadas ou rugosas, caracterizadas por serem colônias chatas, grandes com aparência granular e opacas. O crescimento é esparso no meio fluido. A quantidade de C+G do ADN é de 57%. A B. abortus mostra 100% de homologia com as outras brucelas, exceto a B. ovis. FAGOTIPAGEM A fagotipagem é utilizada para identificar diferentes espécies de Brucella spp, incluindo as amostras de B. abortus. Os fagos utilizados na rotina são Tb, Wb, Fi, Bk2 e R. A diluição do fago é conhecida como RTD (“Routine Test Dilution”) que corresponde a 104-105 unidades formadoras de placas por mililitro (UFP/mL). O RTD é o número mínimo de fagos responsável pela lise confluente numa cepa propagante. Os procedimentos para fagotipagem foram descritos em detalhes por Corbel e Hendy (1985). A fagotipagem é uma ferramenta rápida e segura na identificação de espécies do gênero Brucella. ANTÍGENOS A parede celular da B. abortus consiste de uma camada externa de lipopolissacarídio na qual a cadeia de polissacarídeos fica exposta. A cadeia de polissacarídeo possui os antígenos de superfície principais (A ou M), os quais estão envolvidos na aglutinação. Estes antígenos da B. abortus nas cepas lisas estão relacionados com antígenos de superfície encontrados na Yersinia enterocolitica 09, sendo fonte de confusão na interpretação de testes sorológicos para a brucelose.
As proteínas da parede celular estão agrupadas em 3 categorias de acordo com o peso molecular (SDS-PAGE). As proteínas possuem: a) 8.000 a 94.000D; b) 35.000 a 40.000D (porinas) e c) 25.000 a 30.000D. O antígeno protéico provavelmente estimule a reação de hipersensibilidade retardada, provavelmente uma porina. Os antígenos envolvidos na resposta celular não estão totalmente identificados. EPIZOOTIOLOGIA A B. abortus é um agente intracelular facultativo de bovinos e outras espécies, sendo transmitida pela ingestão do alimento ou secreções contaminadas. A transmissão venérea é possível, mas pouco comum. A transmissão congênita ou intramamária pode ocorrer. Equinos, ovinos e cães podem infectar-se, mas a transmissão, nestas espécies, para
Generated by Foxit PDF Creator © Foxit Softwarehttp://www.foxitsoftware.com For evaluation only.

Microbiologia Clínica Veterinária VET 3225 ÁREA DE BACTERIOLOGIA 2011-2
UFRGS
Microbiologia Clínica Veterinária VET 3225 ÁREA DE BACTERIOLOGIA 2011-2
UFRGS
o hospedeiro primário é pouco provável. A doença é encontrada em todas as regiões criatórias de bovino no mundo, exceto nas áreas onde os programas de erradicação tiveram sucesso.
A B. abortus não é resistente à luz solar e à dissecação, sobrevivendo mais no inverno do que no verão. Sobrevive no leite, mas é destruída pela pasteurização.
A brucelose bovina é endêmica no Brasil, estando presente e diagnosticada em todos os estados, entretanto há grandes variações entre as estimativas dos estados. O Ministério da Agricultura (MA), em 1975, num levantamento soroepidemiológico estimou a prevalência da infecção brucélica em bovinos em 4,0% na região Sul; 7,5% na região Sudeste; 6,8% na região Centro-Oeste; 2,0% na região Nordeste e 4,1% na região Norte (Anselmo e Pavez, 1977; Poester et al., 2002). Outros estudos, realizados principalmente na década de 80 não detectaram mudanças significativas comparadas à estimativa da década anterior. No começo da década de 1990, Minas Gerais iniciou uma campanha de vacinação compulsória de terneiras com a B19 em todo o estado. Além de Minas Gerais, o Rio Grande do Sul era o único estado que possuía um programa de vacinação ativo, mas com menores índices de cobertura vacinal (Paulin e Ferreira Neto, 2003).
No Rio Grande do Sul, os levantamentos regionais revelaram que a situação pouco mudou nos últimos anos. O programa bem sucedido de vacinação contra a brucelose bovina fez baixar a prevalência de 2% para 0,6% em 1986. O último levantamento, realizado em 2006, revelou uma prevalência ao redor de 1% com variações, segundo a região do Estado. Segundo os dados oficiais, a prevalência da brucelose bovina no Brasil variou entre 4 e 5%, no período de 1989-1998 (Brasil, 2001).
Os estudos brasileiros mostraram que foram isoladas e identificadas a B. abortus biovares 1, 2, 3 e 6 e a B. suis (Garcia-Garrillo, 1987; Langenegger et al., 1975). Giorgi et al. (1972), em São Paulo, isolaram 23 amostras de B. abortus e B. suis de bovinos, suínos e equinos. No Rio Grande do Sul, Poester (1974; 1977) isolou B.abortus biotipo 1 e B. suis de bovinos, equinos e suínos. No Rio de Janeiro, Langenegger et al. (1975), isolaram e identificaram 10 amostras B. abortus, sendo 4 cepas de B. abortus biotipo 1 e 6 do biotipo 3 de bovinos em matadouro. A maioria dos casos de brucelose humana está ligada a atividade profissional do trabalhador e essencialmente relacionada com o magarefe.
A prevalência da infecção humana ou animal muito provavelmente esteja subestimado, pois são reduzidos os trabalhos disponíveis. Em Minas Gerais, Godoy et al. (1977) estimaram prevalência de 0,28% de reagentes dentre 9.360 amostras de doadores de sangue. Barufa (1978), no Rio Grande do Sul, evidenciaram que a enfermidade era mais frequente em pessoas da zona rural; mais frequente no homem do que na mulher e que o consumo de queijo não pasteurizado seria a fonte de infecção para o homem. Nos Estados Unidos, 85% das infecções em bovinos são causadas pelo biótipo 1; as remanescentes pelos biótipos 2 e 4. As propriedades de maior tamanho provavelmente possuem maiores infecções maiores pela manutenção de animais, latentemente infectados. O uso de áreas de pastejo comunitárias pode ser um fator na transmissão de B. abortus entre as propriedades. Há casos de infecção por B. abortus em várias espécies silvestres como bisões, cervídeos e alces. O convívio destas espécies com bovinos aumenta a possibilidade de infecções cruzadas, representando uma via pouco frequente. Na África, a
Generated by Foxit PDF Creator © Foxit Softwarehttp://www.foxitsoftware.com For evaluation only.

Microbiologia Clínica Veterinária VET 3225 ÁREA DE BACTERIOLOGIA 2011-2
UFRGS
Microbiologia Clínica Veterinária VET 3225 ÁREA DE BACTERIOLOGIA 2011-2
UFRGS
situação é bem diferente, ocorrendo casos de brucelose bovina devido à transmissão, através dos animais silvestres. Os animais pré-púberes são geralmente resistentes à infecção, mas há um aumento da suscetibilidade à infecção com o amadurecimento sexual e prenhez. A infecção da bezerra (terneira) pode ocorrer no útero da mãe ou pela ingestão de leite contaminado. Animais expostos podem desenvolver infecções latentes que não são detectáveis, através de testes sorológicos. A freqüência de tais infecções está estimada em 2 a 3 % dos animais expostos. Os animais com infecção latente podem permitir a transmissão da infecção ao produto e disseminar a brucelose em uma propriedade (tida como) indene. Nos machos, os casos de epididimites e orquites são relativamente comuns, podendo assim, transmitir a brucelose, através do sêmen contaminado. A principal porta de entrada da B. abortus é a mucosa oral de terneiros ou terneiras que ingerem leite contaminado; nasofaringe e a mucosa conjuntival e, mais raramente, o trato genital de machos e de fêmeas. Sob condições experimentais, o organismo penetra na pele íntegra de cobaias, suínos e bovinos. Após a penetração a B. abortus dirige-se ao linfonodo regional e, posteriormente à corrente sanguínea. A fase de bacteremia resulta na disseminação do organismo ao úbere, útero e linfonodos associados.
A B. abortus é um microrganismo intracelular facultativo, podendo sobreviver e multiplicar-se em macrófagos e células epiteliais. A sobrevivência nas células fagocíticas do hospedeiro, em parte, está relacionada à falha do organismo em estimular um nível efetivo de desgranulação após a entrada do agente. Este efeito é mais aparente em B. abortus com variantes lisas do que nas rugosas. A B. abortus possui uma resistência natural à morte intraleucocitária. O agente prolifera maciçamente em células com altos níveis de eritritol, como aquelas encontradas no trato genital de fêmeas prenhe e macho. As bactérias penetram nas células epiteliais do córion e proliferam, produzindo placentite e endometrite com ulceração da camada de revestimento do útero.
As lesões no feto incluem: edema, congestão pulmonar, hemorragias do epicárdio e da cápsula esplênica. A morte fetal segue, mas não é certo que seja devido à endotoxinas da B. abortus ou interferência da função placentária. A presença dos organismos induz inflamação das membranas, interferindo com a circulação do feto e pode explicar o porquê do aborto. O feto morre por falta de aporte nutricional. O organismo é encontrado no estômago e pulmões do feto abortado. O aborto, geralmente ocorre no terço final da prenhez. Após o parto ou aborto a B. abortus está presente nas descargas uterinas por poucos dias, sendo gradualmente eliminada do trato reprodutivo. A infecção pode ser mantida no sistema microcítico fagocitário e úbere. Grande número de bactérias é eliminado no leite e fonte de infecção para bezerros (terneiros) e para o homem. A maioria dos infectados permanece portador pela vida, eliminando o organismo no exsudato e no leite, após cada parto. A B. abortus pode ser detectada nos linfonodos do trato digestivo e baço de animais infectados, podendo ser isolada do sangue e de higromas do joelho. O higroma da articulação do joelho possui alta correlação com abortos causados por B. abortus em animais no continente africano, onde um grande número de organismos virulentos está presente no fluido. B. abortus pode infectar equinos em menor frequência do que em bovinos. Nos equinos, as localizações preferenciais são: bursas, articulações ou bainhas tendíneas, e tem
Generated by Foxit PDF Creator © Foxit Softwarehttp://www.foxitsoftware.com For evaluation only.

Microbiologia Clínica Veterinária VET 3225 ÁREA DE BACTERIOLOGIA 2011-2
UFRGS
Microbiologia Clínica Veterinária VET 3225 ÁREA DE BACTERIOLOGIA 2011-2
UFRGS
sido encontrada em bursites supra-atlantal, bursites supra-espinosa e em cernelhas fistulosas (Mal das cernelhas). Esta bactéria pode também infectar ovinos, caprinos e suínos, mas numa frequência menor. IMUNIDADE A B. abortus é parasito intracelular facultativo, podendo escapar do efeito bactericida do anticorpo ou complemento presente no plasma. A imunidade protetora depende principalmente da resposta celular em que a atividade lítica dos macrófagos é iniciada, após a ativação das linfocinas pelos linfócitos T. A opsonização pelos anticorpos facilita a entrada da bactéria nos macrófagos, aumentando a possibilidade da morte bacteriana intracelular.
As bactérias multiplicam-se mais lentamente nos macrófagos de animais vacinados do que em animais controle não vacinados. O anticorpo humoral é pouco correlacionado com a imunidade protetora. Fêmeas vacinadas com B19, quando (bezerras) terneiras, mostram-se resistentes ao desafio, após os títulos caírem a níveis abaixo dos detectáveis. Entretanto, grandes doses de soro hiperimune podem interromper a difusão de Brucella spp nos animais infectados. Após a infecção, aglutininas da classe IgM são as primeiras imunoglobulinas a aparecerem no plasma, atingindo o seu pique em 2 semanas. Os anticorpos IgG aparecem um pouco mais tarde, superando os títulos de IgM em 4 a 6 semanas e, permanecendo como anticorpos dominantes. Bovinos infectados possuem altos títulos de IgG. Estes anticorpos aglutinantes não possuem atividade opsonizante e nenhum efeito sobre a eliminação do organismo. Em bezerras vacinadas com a B 19, a produção de IgM aumenta rapidamente. O anticorpo IgG1 eleva-se mais lentamente, não atingindo altos níveis, nem persistem por muito tempo. Os anticorpos de IgG1 são muito mais baixos nos animais vacinados do que dos animais naturalmente infectados. DIAGNÓSTICO A infecção causada pela B. abortus pode ser detectada, através do: a) diagnóstico bacteriológico; b) diagnóstico sorológico e c) diagnóstico molecular. Diagnóstico bacteriológico O isolamento da B. abortus é obtido pelo cultivo no meio base (agar Triptose ou agar Albimi) com a adição de soro e ATMs. O cultivo é incubado à 37ºC em uma atmosfera de 10 % de CO2, durante, pelo menos, 4 dias. As amostras podem ser obtidas de feto abortado; placenta; exsudato uterino; leite e abscessos. A prova biológica consiste na inoculação de tecidos ou fluidos macerados em cobaias, devendo ser sacrificados 3 a 6 semanas, mais tarde. O soro é testado para a presença de anticorpos e os órgãos como o baço, fígado, linfonodos regionais e testículos devem ser cultivados para o re-isolamento de B. abortus. O exame direto de tecidos pode ser realizado pela IF (Imunofluorescência) ou pela imunoistoquímica. Estas técnicas podem ser importantes, especialmente em amostras contaminadas (membranas fetais, cotilédones, secreções vaginais ou fetais), apesar de ser pouco utilizada na prática.
Generated by Foxit PDF Creator © Foxit Softwarehttp://www.foxitsoftware.com For evaluation only.

Microbiologia Clínica Veterinária VET 3225 ÁREA DE BACTERIOLOGIA 2011-2
UFRGS
Microbiologia Clínica Veterinária VET 3225 ÁREA DE BACTERIOLOGIA 2011-2
UFRGS
Diagnóstico sorológico Os testes sorológicos para diagnóstico da brucelose tiveram inicio, em 1897, com o
desenvolvimento do teste de aglutinação de Wright (Wright e Smith, 1897). Problemas de reações sorológicas positivas resultantes de exposição a outras bactérias ou reações cruzadas foram detectados. Desde então, melhores testes têm sido desenvolvidos e muitos destes são utilizados mundialmente. Raras enfermidades infecciosas apresentam tanta variedade diagnóstica como a brucelose. Estes métodos incluem testes aplicados no soro, no sangue total, no muco vaginal, no plasma seminal, no leite dessorado ou no leite total. O diagnóstico da brucelose, de um modo geral, enfrenta situações específicas e próprias que podem interferir com o desempenho dos testes aplicados (sensibilidade e especificidade). Reações inespecíficas são atribuídas a anticorpos naturais ou a produtos catabólitos de organismos que compartilham antígenos de estruturas semelhantes às da Brucella. Reações cruzadas com outros microrganismos como: Salmonella urbana, Escherichia coli O:116 ou O:157, Stenotrophomonas maltophilia e Yersinia enterocolitica O:9 são alguns desses exemplos.
A membrana externa da brucela lisa é composta de fosfolipídios, proteínas e LPS-S. A maioria dos testes sorológicos, particularmente aqueles que utilizam suspensão de bactérias íntegras como o teste de soro-aglutinação lento (SAL) em tubo; o teste do Antígeno Acidificado Tamponado (AAT) e o teste de Fixação do Complemento (FC); a maioria dos testes de ELISA e o Teste do Anel em Leite (TAL) foram desenvolvidos para detectar anticorpos contra a cadeia O do LPS-S.
Estes testes são úteis em levantamentos, campanhas em larga escala e em programas sanitários de controle e erradicação com fins comerciais, muito embora outros testes como ELISA não sejam oficialmente reconhecidos no Brasil, no contexto do Programa Nacional de Controle e Erradicação da Brucelose e Tuberculose (PNCEBT). Alguns destes testes são aplicados como teste de triagem por serem baratos, simples, com alta sensibilidade, seguidos de testes confirmatórios mais específicos.
Teste do Antígeno Acidificado Tamponado (AAT)
O teste do AAT é uma modificação do teste de aglutinação em placa. O antígeno é corado pelo Rosa de Bengala e tamponado a um pH de 3,65. Neste pH as “aglutininas não específicas” são inativadas o a IgG dos animais infectados, aglutinam fortemente. Iguais volumes de soro e antígeno (30μl) são misturados; agitados por 4 minutos e observados em caixa com iluminação indireta. O teste é barato e simples de realizar. Apesar de melhorar a especificidade num pH ácido, resultados falso-positivos podem ocorrer, geralmente devido a presença de IgM devido a vacinação pela B19.
Generated by Foxit PDF Creator © Foxit Softwarehttp://www.foxitsoftware.com For evaluation only.

Microbiologia Clínica Veterinária VET 3225 ÁREA DE BACTERIOLOGIA 2011-2
UFRGS
Microbiologia Clínica Veterinária VET 3225 ÁREA DE BACTERIOLOGIA 2011-2
UFRGS
1) B. abortus ; 2) B. canis ; 3) B. ceti ; 4) B. melitensis ; 5) B. microti* ; 6) B. neotomae ; 7) B. ovis ; 8) B. pinnipedialis ; 9) B. suis biovar 1 ; 10) B. suis biovar 2 ; 11) B. suis biovar 3 ; 12) B. suis biovar 4 ; 13) B. suis biovar 5
1 2 3 4 5 6 7 8 9 10 11 12 13
Urease +**** + + + + + - + + + + + +
VP - - - - + - - - - - - - -
Fago Tb** (Tbilisi)
+ - - - - d - -
***** - - - - -
Fago Wb** (Weybridge)
+ - + - + + - + + + + + +
Fago Iz** (Izatnagar)
+ - + + + - + + + + + +
Fago** R/C - + - - - + - - - - - -
L-Alanina*** + - - + - d - d - d - -
L-Arabinose***
+ - + - + - - + + - - -
L-Arginina*** - + - - - - - + + + + +
L-Asparagina***
+ - - + + + - - d - - d
Meso-Eritritol***
+ d d + + - + + + + + +
D-Galactose***
+ - + - + - - + + - - -
L-Acido glutâmico***
+ + + + + + + d + + + +
L-Lysina*** - d - - - - - + d d d d
DL-Ornitina***
- + - - - - - + + + + +
D-Ribose*** + + + - d - + + + + + +
D-xilose*** - - + - - - - + d d d d
Hospedeiro preferencial
Bov Can Cet Cap Mar Nle Ov Pin Sui Sui, Leb
Sui Ren Ren
1 2 3 4 5 6 7 8 9 10 11 12 13
1) B. abortus ; 2) B. canis ; 3) B. ceti ; 4) B. melitensis ; 5) B. microti* ; 6) B. neotomae ; 7) B. ovis ; 8) B. pinnipedialis ; 9) B. suis biovar 1 ; 10) B. suis biovar 2 ; 11) B. suis biovar 3 ; 12) B. suis biovar 4 ; 13) B. suis biovar 5
Generated by Foxit PDF Creator © Foxit Softwarehttp://www.foxitsoftware.com For evaluation only.

Microbiologia Clínica Veterinária VET 3225 ÁREA DE BACTERIOLOGIA 2011-2
UFRGS
Microbiologia Clínica Veterinária VET 3225 ÁREA DE BACTERIOLOGIA 2011-2
UFRGS
Fixação do Complemento Este teste é considerado como o teste confirmatório na detecção sorológica de
anticorpos de animais infectados. Ele foi modificado, padronizado e adaptado ao sistema de microplacas (Alton et al. 1988; Anon, 2000). Ao contrário do teste SAL, os títulos não diminuem quando a doença torna-se crônica. Os resultados são expressos em unidades internacionais (UI) e, apesar de não estar padronizado no Brasil, o ponto de corte é definido por alguns países em 20 UI que é aplicado rigorosamente onde a B19 não tiver sido utilizada por muitos anos. A sua aplicação é restrita em países que aplicam a B19 como a África do Sul que frequentemente tem problemas com um número inaceitável de falso-positivos, pois a vacinação induz a títulos significantes. É necessário treinamento e experiência para certificar como livres de brucelose, quando rebanhos ou animais individualmente são positivos ao teste.
Os títulos vacinais tendem a declinar mais rápido do que aqueles devido à infecção com cepas de campo. O declínio de títulos é também dependente da idade de vacinação dos animais. Existe uma dificuldade na diferenciação das reações vacinais das causadas por cepas selvagens, não existindo nenhuma prova sorológica capaz de diferenciar títulos vacinais de infecciosos. A vacinação de novilhas e animais adultos pode resultar em muita confusão na interpretação de testes laboratoriais, sendo essencial o registro de vacinações e de datas de nascimento para permitir a correta interpretação dos resultados do testes de FC. Este teste é preconizado em bovinos para comércio internacional. Teste de Soroaglutinação lenta (SAL)
O teste de soro-aglutinação em tubo (SAL) foi, no passado, utilizado nos programas de erradicação da brucelose em muitos países. O antígeno e as condições do teste foram padronizados internacionalmente. O teste é realizado em pequenos tubos e com diluições seriadas do soro. A aglutinação completa na diluição 1:100 ou maior é considerada positiva. Mais tarde, este teste foi adaptado como teste de SAR (soro aglutinação rápida) e hoje não é mais utilizado.
Em alguns países, o teste de SAL foi, e ainda é utilizado, associado ao teste do 2-mercaptoetanol (2ME), como teste confirmatório nos programas de erradicação e controle da brucelose. Ele é considerado como de alta especificidade quando associado ao 2ME e alguns autores desencorajam o seu uso isoladamente, especialmente para propósitos comerciais.
Aglutininas não específicas no soro são diminuídas pela adição de EDTA, sem a redução nos títulos de B. abortus de animais infectados. O teste de SAL-EDTA é um teste muito mais específico e particularmente útil na detecção de infecções novas, particularmente naquelas com duas semanas de curso como demonstradas em condições experimentais, mas sua utilidade em rebanhos que estão cronicamente infectados é mais limitada, pois alguns animais infectados poderiam ser classificados como negativos pelo teste, pois a infecção está na fase crônica. O teste de SAL é ainda muito útil como teste suplementar para indicar os níveis de anticorpos IgM, a imunoglobulina após a vacinação com a B19, mas não serve como indicador de infecção ou vacinação.
Generated by Foxit PDF Creator © Foxit Softwarehttp://www.foxitsoftware.com For evaluation only.

Microbiologia Clínica Veterinária VET 3225 ÁREA DE BACTERIOLOGIA 2011-2
UFRGS
Microbiologia Clínica Veterinária VET 3225 ÁREA DE BACTERIOLOGIA 2011-2
UFRGS
O teste do 2-Mercaptoetanol (2ME) É um teste utilizado em paralelo com a prova lenta que por sua seletividade apenas
para IgG é empregado como teste complementar aos testes de triagem. Os anticorpos da classe IgM degradam-se em presença de substâncias, contendo radicais tiol como o 2ME. Desse modo, as amostras de soro com predomínio de IgM apresentam-se negativas ao 2ME e positivos à prova lenta. Este mesmo resultado pode ser interpretados como resultados inconclusivo. Se os 2 testes forem positivos com predomínio de IgG, eles podem indicar infecção. O teste de ELISA indireto (ELISAi)
O teste de ELISA indireto (ELISAi) possui uma sensibilidade semelhante à do AAT, porém menor que a FC. O teste ELISA foi o teste substituto do AAT por estar padronizado e sua leitura feita por espectrofotômetro ao invés de visual. Deve-se ter cuidado ao empregar-se esta técnica em rebanhos vacinados com B19, pois muitos resultados falso-positivos podem ocorrer. O teste tem sido indicado para o comércio internacional de bovinos pela OIE (Anon, 2000). O teste de ELISA competitivo (ELISAc) A base deste teste foi o uso de anticorpo monoclonal seletivo (Mab) que compete com o anticorpo a ser testado. O ELISAc utiliza um Mab específico para um dos epitopos da B. abortus (cadeia O) e que por isso possui maior especificidade do que o ELISA indireto. O ELISAc foi capaz de eliminar em parte, os problemas de reações pós-vacinais (B19) ou com outras bactérias. Infelizmente, o ELISAc resolve apenas parcialmente este problema. Ainda persistem os anticorpos após a infecção com Yersinia enterocolitica O:9 e pela vacinação com B19. Entretanto, a atividade do anticorpo residual devido à vacinação ou reação cruzada foi menos persistente neste teste do que com os outros testes. É recomendado como teste alternativo no comércio internacional de bovinos pela OIE (Anon, 2000) Ensaio da Polarização Fluorescente (EPF)
O ensaio de polarização fluorescente é uma técnica rápida e simples para mensurar a interação antígeno-anticorpo, sendo útil tanto no laboratório quanto em trabalhos de campo. O mecanismo do teste é baseado na velocidade de rotação aleatória das moléculas em solução. Moléculas menores apresentam maior velocidade e vice-versa. O antígeno, conjugado com fluorcromo de baixo peso molecular (um fragmento do polissacarídeo O da B abortus) é adicionado ao soro ou outro fluido a ser testado. Se o anticorpo estiver presente, há a ligação com o antígeno marcado, causando uma diminuição de taxa de rotação que pode ser medida por um equipamento de luz polarizada. O EPF tem apresentado bons resultados em trabalhos experimentais. Este teste é recomendado como uma alternativa para o mercado internacional de bovinos, pela OIE. Teste do Anel em Leite (TAL)
O teste do anel em leite (TAL) foi utilizado na detecção de anticorpos no leite. O desenvolvimento de reação positiva depende de duas reações:
Generated by Foxit PDF Creator © Foxit Softwarehttp://www.foxitsoftware.com For evaluation only.

Microbiologia Clínica Veterinária VET 3225 ÁREA DE BACTERIOLOGIA 2011-2
UFRGS
Microbiologia Clínica Veterinária VET 3225 ÁREA DE BACTERIOLOGIA 2011-2
UFRGS
a) Glóbulos de gordura no leite são aglutinados pelos anticorpos no leite (aglutininas/glóbulos de gordura)
b) Brucelas coradas pela hematoxilina (antígeno) que são adicionadas ao leite e que formam um complexo antígeno/anticorpo e glóbulos de gordura que ascendem à superfície, formando uma camada colorida no topo do tubo de teste.
O teste foi utilizado com teste de detriagem pela sensibilidade e versatilidade, pois pode detectar a infecção brucélica nas amostras de leite (tarro ou de tanques) de rebanhos com animais infectados, nos indivíduos infectados de um rebanho ou no monitoramento de rebanhos livres. A sensibilidade é reduzida quando aplicada em rebanhos grandes com poucos reagentes. Entretanto, a perda na sensibilidade em rebanhos grandes com 150 ou mais animais pode ser contrabalançada pelo aumento na proporção do leite em relação ao antígeno adicionado à amostra. Apesar de sua reduzida sensibilidade em grandes rebanhos, o TAL tem tido sucesso no monitoramento de rebanhos leiteiros livres de brucelose. Após um teste positivo no tarro, as vacas que contribuíram com esse leite devem ser individualmente testadas pela sorologia, na identificação das fêmeas infectadas.
O TAL tem desvantagens como resultados falso-positivos incluindo 1-Prevalência alta de mastites; 2-Proporção alta de vacas no início e fim de lactação; 3-Vacinação recente com B19 (3-4 meses); 4-Leite congelado. As amostras de leite podem ser preservadas para o TAL pela adição de 0,5 mL de
solução de formalina (7,5 mL de 37% de formaldeído em 1 litro de água destilada) para 10 ml de leite se o teste for realizado muito longe.
No TAL o antígeno é uma suspensão morta de B. abortus, coradas pela hematoxilina. Leite fresco misturado ao antígeno na proporção de 1 mL do leite para cada gota de antígeno. A mistura é incubada a 37ºC, durante 1 hora. Caso seja positiva, a reação apresentará agrupamentos de organismos na porção superficial, levados pelos glóbulos de gordura. Assim, um teste positivo é evidenciado pela formação de um anel de coloração violeta azulada na porção superior do tubo. A duração e temperatura na qual as amostras são estocadas podem causar reações falso-positivas (em particular temperaturas maiores de 45º C por 5 min). As amostras pasteurizadas não devem ser testadas pelo TAL.
Vários países têm substituído o TAL pelo ELISAi para o leite. Embora essa técnica não esteja padronizada, ela é recomendada no comércio de bovinos pela OIE. Diagnóstico molecular (PCR)
Numerosos ensaios, utilizando o PCR já foram desenvolvidos na identificação de espécies do gênero Brucella spp, especialmente nos estudos epidemiológicos. Várias estratégias têm sido exploradas na diferenciação de espécies de Brucella, incluindo “locus specific multiplexing” como o baseado no IS711 (sequência de inserção); PCR-RFLP no locus omp2. Infelizmente, não há técnica robusta o suficiente que permita a diferenciação entre os diferentes biótipos. Inicialmente, esses ensaios foram dirigidos ao ADN purificado de isolados cultivados, mas tão logo novas linhagens foram identificadas (no leite e queijo), novos avanços permitiram a melhoria dessa técnica diagnóstica em laboratórios como a remoção de inibidores da PCR.
Generated by Foxit PDF Creator © Foxit Softwarehttp://www.foxitsoftware.com For evaluation only.

Microbiologia Clínica Veterinária VET 3225 ÁREA DE BACTERIOLOGIA 2011-2
UFRGS
Microbiologia Clínica Veterinária VET 3225 ÁREA DE BACTERIOLOGIA 2011-2
UFRGS
TESTE DE SENSIBILIDADE AOS ANTIMICROBIANOS A B. abortus é sensível às gentamicinas, canamicinas, tetraciclinas e rifampicinas. A tetraciclina é associada à estreptomicina no tratamento de brucelose humana. A combinação de cotrimoxazol à rifampicina ou tetraciclina e estreptomicina à rifampicina também é utilizada. A localização intracelular do organismo requer uma terapia prolongada. Bovinos não devem ser tratados com ATMs. VACINAS Há dois imunógenos importantes no controle da infecção por B. abortus: a vacina elaborada com a amostra B19 e a vacina elaborada com a amostra RB51. As 2 vacinas são vivas. A cepa 19 consiste de uma linhagem viável, caracterizada pela baixa virulência (cobaias e bovinos) e excelente propriedade imunizante. Esta cepa possui grande estabilidade. A sua virulência não foi alterada desde 1930, ano em que foi isolada pela primeira vez. A cepa B19 é uma amostra lisa de B. abortus, levemente patogênica para cobaias. Vacas prenhes podem abortar pela inoculação da vacina B19 e, nesse caso, o organismo vacinal pode ser demonstrado com facilidade nas membranas fetais e feto. A linhagem vacinal, raramente é eliminada pelo leite. Ela pode causar infecção no homem, geralmente de forma leve, com período de recuperação mais curto do que infecções por cepas virulentas. Esta vacina deve por isso ser manipulada com cuidado. As terneiras (bezerras) devem ser vacinadas entre 4 e 8 meses de idade. A vacinação nessa idade é recomendada para evitar a persistência de aglutininas e problemas no diagnóstico mais tarde. A cepa B19 protege da infecção por B. abortus cerca de 70% das vacas ou por 4 a 5 gestações, sendo mais efetiva na proteção de animais jovens de cria quando aplicada sob bases populacionais. Animais adultos vacinados com a B19 são protegidos, mas desenvolvem aglutininas, persistentemente. Há evidências de que se reduzirmos a dose vacinal haveria uma menor persistência de aglutininas, mas esta prática não é recomendada no Brasil. A vacinação de vacas no período de prenhez inicial com grandes doses (60 bilhões) de B19 produz uma alta probabilidade de infecção uterina. O risco é menor quando a dose é reduzida (300 milhões). A vacina elaborada com a cepa rugosa 45/20 de MacEwen não é mais empregada em nosso meio, mas foi usada no Brasil e amplamente utilizada na Irlanda e outros países europeus no controle da brucelose bovina, no passado. É uma vacina produzida com a bactéria morta e com adjuvante oleoso. CONTROLE E PREVENÇÃO As perdas econômicas advindas da brucelose, juntamente com o perigo de infecção humana, impuseram o programa de controle e erradicação da doença. No Brasil, a perda econômica com a brucelose bovina foi estimada, em 1971, como algo em torno de 32 milhões de dólares anuais (Poester et al., 2002). É muito provável que esses prejuízos estejam subestimados e urge que façamos um levantamento da prevalência da brucelose em todos os estados do Brasil. Assim poderíamos inferir nos prejuízos diretos e indiretos da brucelose nos bovinos e em outros animais de produção.
Generated by Foxit PDF Creator © Foxit Softwarehttp://www.foxitsoftware.com For evaluation only.

Microbiologia Clínica Veterinária VET 3225 ÁREA DE BACTERIOLOGIA 2011-2
UFRGS
Microbiologia Clínica Veterinária VET 3225 ÁREA DE BACTERIOLOGIA 2011-2
UFRGS
Os princípios incorporados no PNCEBT foram dependentes das condições locais, manejo e do número de animais envolvidos. 1) Animais infectados devem ser detectados, marcados e eliminados da propriedade. A detecção é realizada pela sorologia ou ainda utilizando o Teste do Anel no Leite como teste de triagem, seguido pelo teste de aglutinação de cada amostra animal com sacrifício dos reagentes. 2) A vacinação obrigatória com B19 de terneiras (bezerras) entre 4-8 meses deve aumentar a resistência dos animais. É importante esclarecer que no RS é aconselhado fazer a vacinação de animais um pouco mais cedo, pois algumas raças européias ciclam com 5-6 meses de idade. 3) Princípios gerais de higiene são impostos na prevenção da infecção ou re-introdução da infecção. A doença desaparece dentro de 2 anos e, ao final de 5 anos, dos animais com infecção crônica, se forem tomadas essas medidas,. VACINA RB51 A amostra RB51 é uma amostra rugosa, estável e que não contém cadeia O no LPS da parede celular e comporta-se bioquimicamente como a linhagem lisa 2308 em sua utilização do eritritol, da qual foi originada. A RB51 induz a formação de anticorpos às proteínas da membrana externa, mas não contra a cadeia O. Por esta razão pode ser aplicada em animais com qualquer idade, pois não interfere com os testes sorológicos. Ela possui reduzida virulência para camundongos por ter um período curto de "clearence" no baço, conferindo por isso imunidade ao camundongo imunizado e desafiado com B. abortus 2308. Esta vacina está sendo utilizada em vários países, inclusive no Brasil, pois não interfere nos testes sorológicos tradicionais para detecção de anticorpos contra o LPS (lipopolissacarídio) da cadeia O. A vigilância de uma propriedade deve ser mantida por meio de testes sorológicos periódicos; pelos testes nos animais comprados; pela ocorrência de sinais clínicos compatíveis com a enfermidade e pelo exame sorológico de animais enviados ao matadouro.
Generated by Foxit PDF Creator © Foxit Softwarehttp://www.foxitsoftware.com For evaluation only.
Microbiologia Clínica Veterinária VET 3225 ÁREA DE BACTERIOLOGIA 2011-2
UFRGS
Microbiologia Clínica Veterinária VET 3225 ÁREA DE BACTERIOLOGIA 2011-2
UFRGS
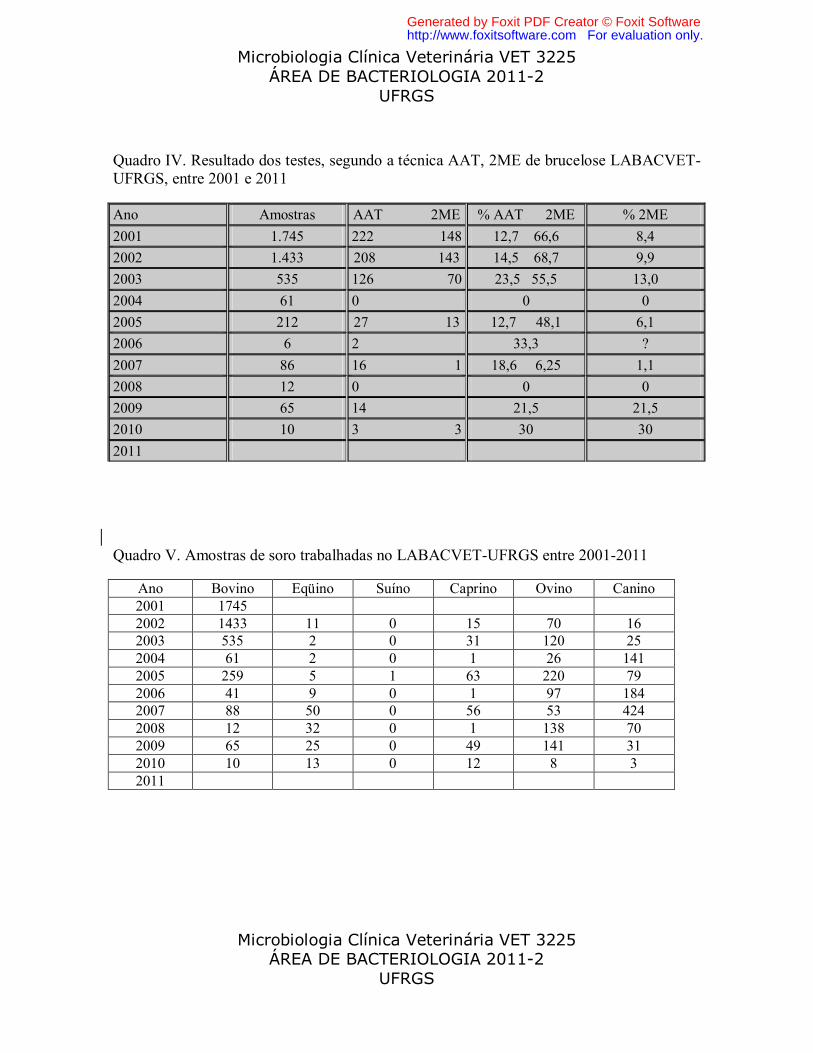
Quadro IV. Resultado dos testes, segundo a técnica AAT, 2ME de brucelose LABACVET-UFRGS, entre 2001 e 2011
Ano Amostras AAT 2ME % AAT 2ME % 2ME
2001 1.745 222 148 12,7 66,6 8,4
2002 1.433 208 143 14,5 68,7 9,9
2003 535 126 70 23,5 55,5 13,0
2004 61 0 0 0
2005 212 27 13 12,7 48,1 6,1
2006 6 2 33,3 ?
2007 86 16 1 18,6 6,25 1,1
2008 12 0 0 0
2009 65 14 21,5 21,5
2010 10 3 3 30 30
2011
Quadro V. Amostras de soro trabalhadas no LABACVET-UFRGS entre 2001-2011
Ano Bovino Eqüino Suíno Caprino Ovino Canino 2001 1745 2002 1433 11 0 15 70 16 2003 535 2 0 31 120 25 2004 61 2 0 1 26 141 2005 259 5 1 63 220 79 2006 41 9 0 1 97 184 2007 88 50 0 56 53 424 2008 12 32 0 1 138 70 2009 65 25 0 49 141 31 2010 10 13 0 12 8 3 2011
Generated by Foxit PDF Creator © Foxit Softwarehttp://www.foxitsoftware.com For evaluation only.

Microbiologia Clínica Veterinária VET 3225 ÁREA DE BACTERIOLOGIA 2011-2
UFRGS
Microbiologia Clínica Veterinária VET 3225 ÁREA DE BACTERIOLOGIA 2011-2
UFRGS
Brucella ovis Prof. Marcos JP Gomes BRUCELOSE OVINA ou EPIDIDIMITE OVINA
INTRODUÇÃO A Brucella ovis causa uma enfermidade crônica nos ovinos e, caracterizada principalmente por alterações testiculares com conseqüências sobre a fertilidade dos carneiros, abortos ocasionais nas ovelhas e aumento da mortalidade de perinatal em cordeiros. HISTÓRICO A B. ovis foi isolada, pela primeira vez, em 1952, por McFarlane e colaboradores, na Nova Zelândia. Simmons e Hall, em 1953, na Austrália isolaram e descreveram o organismo como semelhante a “brucella like organism”. Buddle e Boyes (1953) consideraram-na como uma mutante da Brucella melitensis. Buddle propôs os nomes de Brucella ovis, tendo como base os antígenos de superfície comuns entre a amostra e as amostras rugosas de B. abortus e B. melitensis. Propôs também o nome da doença como “epididimite infecciosas dos carneiros”. A homologia do DNA recentemente propôs a existência de uma única espécie, a B. melitensis, no gênero Brucella e as demais espécies (de hoje) seriam biovares. Assim teríamos B. melitensis biovar Ovis. Esta proposta, no entanto, não se concretizou na prática. DISTRIBUIÇÃO A infecção é cosmopolita, especialmente nos países onde há criação de ovinos. A infecção é maior quando detectada pela primeira vez, podendo variar entre 20 a 60% dos carneiros e em 45 a 75% dos rebanhos testados. A prevalência é baixa, nos países onde há programas de controle, entretanto a erradicação é difícil de ser alcançada.
A B. ovis produz doença clínica ou subclinica em ovinos que é caracterizada por lesões genitais em carneiros e placentite nas ovelhas. A principal consequência da doença é a redução na fertilidade dos machos, abortos esporádicos na fêmea e aumento da mortalidade perinatal. A doença tem sido relatada na Argentina, Austrália, Brasil, Canadá Chile, França, Alemanha, Hungria, México, Nova Zelândia, Peru, Romenia, Russia, República Eslovaca, África do Sul, Espanha, Uruguai, EUA, mas provavelmente ela ocorra na maioria dos paises produtores de ovinos. IMPACTO ECONÔMICO O impacto econômico é difícil de ser quantificado. No Brasil, sabemos que a enfermidade causa prejuízo direto e indireto sobre a produtividade dos rebanhos infectados, especialmente sobre a fertilidade de reprodutores ovinos (machos e fêmeas). Recentemente, as autoridades governamentais brasileuras elaboraram um programa nacional de controle da brucelose ovina para o Brasil, mas ainda não foi efetivamente implantado.
Generated by Foxit PDF Creator © Foxit Softwarehttp://www.foxitsoftware.com For evaluation only.

Microbiologia Clínica Veterinária VET 3225 ÁREA DE BACTERIOLOGIA 2011-2
UFRGS
Microbiologia Clínica Veterinária VET 3225 ÁREA DE BACTERIOLOGIA 2011-2
UFRGS
CARACTERÍSTICAS MORFOLÓGICAS ECULTURAIS A B. ovis é um cocobastonete ou bastonete Gram negativo; não encapsulado; não forma esporos; possui tamanho de 0,5 a 0,7 de diâmetro e 0,7 a 1,2 μm de comprimento; corados pela técnica de Stamp ou de Köster; cresce bem em meios base (Trypticase Soya Agar, Blood Agar Base, Columbia Agar) enriquecidos com 5 a 10% de sangue ou soro ou seletivamente isolada no meio de Thayer-Martin modificado. O crescimento necessita 10 a 20% de CO2 no cultivo primário, embora existam cepas CO2 independentes. As colônias tornam-se visíveis após 3 a 5 dias de incubação a temperatura de 34–37ºC. As amostras podem crescer a temperatura de 26ºC, mas as colônias aparecem em 6-10 dias. Colônias não são hemolíticas, são circulares, convexas, com bordos inteiros. São positivas ao teste da acriflavina para colônias rugosas. CARACTERÍSTICAS BIOQUIMICAS
Perderam a atividade da urease e não reduzem nitrato a nitrito. São catalase positivas e oxidases negativas. H2S negativas. Crescem nas concentrações de fucsina básica e tionina. Oxidam: L-alanina, D-alanina, L-asparagina, D-asparagina, Ácido d-glutâmico, DL-serina, e adonitol. Não oxidam: L-arabinose, D-galactose, D-glicose, D-ribose, meso-eritritol, D-xilose, L-arginina, DL-citrulina, DL-ornitina e L-lisina. FAGOTIPAGEM
A B. ovis não é lisada pelos fagos: Tibilisi, Weybridge, M51-S708, Firenze, BK, MC/75, D ou grupo R. No entanto é lisada pelo fago R/O, tanto na RTD (dose rotina de teste) como 104 RTD. Não há biotipos reconhecidos. TRANSMISSÃO A principal forma de transmissão é por via venérea. A transmissão macho para macho é possível em que machos infectados e animais susceptíveis compartilham o mesmo espaço. Sodomia é outro meio de transmissão. A maioria dos casos ocorre após a estação de monta ou acasalamento. Indubitavelmente existem mecanismos complexos que não estão totalmente conhecidos, mas a transmissão venérea passiva de macho infectado para macho suscetível via fêmea infectada parece ser a mais importante forma de transmissão para manter e difundir a doença. A probabilidade de infecção depende principalmente da via, da dose e das características intrínsecas do animal, tais como idade e raça.
A infecção experimental no carneiro pode ser obtida de muitas formas: oral, intravenosa, intratesticular, conjuntival, intraprepucial subcutânea, através de escarificações da pele, intraretal, e intranasal. Embora não haja estudos comparativos, os melhores resultados foram obtidos, através da inoculação intraconjuntival, intraprepucial ou por ambas simultaneamente. Doses de 5 X 108 – 1010 UFC de B. ovis são suficientes para obter as taxas de infecção próximas a 100%. Se a idade afeta a susceptibilidade à infecção, isto é alvo de controvérsia. A infecção tem sido demonstrada em carneiros com 4 meses de idade, sugerindo que animais na puberdade ou logo após a ela são susceptíveis a B. ovis. Embora a transmissão venérea pareça ser a principal forma de difusão, os animais adultos são mais susceptíveis à infecção natural. Alem disso, a incidência de alterações testiculares e a
Generated by Foxit PDF Creator © Foxit Softwarehttp://www.foxitsoftware.com For evaluation only.

Microbiologia Clínica Veterinária VET 3225 ÁREA DE BACTERIOLOGIA 2011-2
UFRGS
Microbiologia Clínica Veterinária VET 3225 ÁREA DE BACTERIOLOGIA 2011-2
UFRGS
brucelose aumentam com a idade, estando relacionadas à experiência sexual dos animais. Não há informações publicadas sobre o efeito da idade sobre a suscetibilidade à infecção sobre condições experimentais. SUSCEPTIBILIDADE AOS HOSPEDEIROS As ovelhas, ao contrário dos carneiros, parecem ser resistentes à infecção. Poucas fêmeas adquirem a infecção ativa com aborto e morte perinatal mesmo quando cobertas por machos infectados. O mesmo acontece com a infecção experimental de fêmeas prenhes. A infecção que ocorreu na primeira prenhez, dificilmente acompanha a seguinte. Cordeiros nascidos de mães infectadas, dificilmente tornam-se infectados, mesmo ingerindo leite infectado. Estas evidências demonstram que o papel das fêmeas na transmissão ativa da infecção é menos importante. Há muitas referências sugerindo que a susceptibilidade pode variar entre as raças de carneiros. A raça Merina australiana parece ser menos infectada pela B. ovis do que as raças britânicas no mesmo ambiente. A mesma observação tem sido feita, comparando raças importadas com raças nativas. As raças nativas da Espanha e a raças derivadas da raça Merina são mais resistentes à brucelose ovina do que as raças européias importadas. Embora a resistência genética à doença possa ser importante, a suscetibilidade pode estar relacionada à taxa de crescimento, precocidade e atividade sexual. INFECÇÃO EM OUTRAS ESPÉCIES A B. ovis parece infectar exclusivamente ovinos, acomentendo essencialmente carneiros. Não há relatos do isolamento da B. ovis no homem. Há referências da infecção experimental de outras espécies animais. A inoculação experimental da B. ovis no macho caprino leva à colonização genital e extragenital e posteriormente ao desenvolvimento de lesões semelhantes àquelas observadas nos carneiros. O manejo extensivo em que caprinos e ovinos coabitam pode facilitar a transmissão de ovinos para caprinos e vice-versa. Entretanto, o isolamento de B. ovis de casos naturais, em caprinos ainda não foi relatado. A infecção tem sido também reproduzida nos cervos silvestres. Animais de laboratório têm sido infectados por inúmeras vias com doses variando entre 104 a 1011 UFC, mas com sucesso variável. Não há um modelo animal experimental na pesquisa com B. ovis, embora coelhos, ratos, gerbil, hamster, camundongo e cobaias tenham sido utilizados. PATOGENIA
Há um longo período de latência antes de surgirem os sinais clínicos evidentes, assim como a infecção causada por outras espécies de brucelas. Sob condições experimentais, o agente permanece confinado aos linfonodos próximos ao local de entrada por 2 a 3 semanas e então ocorre bacteremia.
Nos carneiros experimentalmente infectados, a bactéria tem sido isolada do fígado, rins, baço, testículo, epidídimo, vesícula seminal, glândula bulbouretral, ampolas e linfonodos ilíaco, pré-escapulares, pré-crural, submaxilar, parotídio e retrofaríngeo. Os órgãos-alvo são epidídimo e glândulas sexuais acessórias com eliminação do agente,
Generated by Foxit PDF Creator © Foxit Softwarehttp://www.foxitsoftware.com For evaluation only.

Microbiologia Clínica Veterinária VET 3225 ÁREA DE BACTERIOLOGIA 2011-2
UFRGS
Microbiologia Clínica Veterinária VET 3225 ÁREA DE BACTERIOLOGIA 2011-2
UFRGS
através do sêmen, na maioria dos carneiros. Nos animais reagentes, geralmente o cultivo é negativo, mas a bacteria pode estar localizada em outros órgãos. Essa hipótese é consistente com o isolamento da B. ovis no baço e linfonodo ilíaco de animais experimentalmente infectados e que não foram isoladas dos órgãos genitais e glândulas sexuais acessórias. A localização no epidídimo é acompanhada por edema perivascular e infiltração dos linfócitos, monócitos e neutrófilos. Logo após a inflamação do epitélio tubular segue-se uma hiperplasia papilar e degeneração hidrópica local com formação de cistos intra-epiteliais. A destruição epitelial, tanto pela bactéria quanto pela reação inflamatória leva a um extravasamento de espermatozóides. A resposta do hospedeiro aos espermatozóides extravasados leva a formação de um granuloma espermárico que pode bloquear o epidídimo com posterior degeneração e fibrose.
Na fêmea, a patogenia da B. ovis não é bem conhecida. Fêmeas ovinas experimentalmente expostas a B. ovis tanto antes da monta quanto no final da gestação não abortam. Somente femeas expostas no início ou na metade da prenhez desenvolvem infecção, podendo eventualmente abortar. A bactéria localiza-se na placenta e alcança o feto, através dos vasos do córion, com a progressão da infecção. Embora o aborto não seja frequente, as fêmeas infectadas desenvolvem placentite, interferindo na nutrição fetal e nascimento de cordeiros fracos. A reação inflamatória e imunológica da B. ovis ao feto é similar àquelas observadas no feto bovino causado pela B. abortus. PATOLOGIA NO MACHO
Carneiros infectados não desenvolvem epididimite palpável, mesmo quando a B. ovis está associada à epididimite ovina, Em levantamento conduzido com 267 carneiros soropositivos, somente 125 (46,8%) mostraram alterações testiculares palpáveis. Uma proporção importante de carneiros infectados não evidencia lesões escrotais.
Na maioria dos casos, a localização testicular é unilateral e a cauda do epidídimo o local mais comum. Alterações na cabeça e no corpo do epidídimo são frequentes. A atrofia testicular e aumento da cauda do epidídimo são características da fase crônica da doença. A aparência macroscópica dos testículos geralmente é normal, mas pode-se perceber granulomas e calcificação. O epidídimo afetado parece firme, mostrando superfície esbranquiçada ao corte em consequência da proliferação de tecido conectivo. Frequentemente são encontradas no tecido conectivo, abscessos, contendo substância cremosa ou caseosa. Hemorragia e inflamação exsudativa na túnica vaginal são achados frequentes como resultado da ruptura da lesão básica (espermatocele) do epidídimo. A organização deste exsudato leva a formação de adesões entre estas duas camadas da tunica vaginal. As vesículas seminais estão aumentadas com ductos dilatados com conteúdo fluido ao corte. Nenhuma alteração macroscópica pode ser observada na glândula bulbo-uretral, próstata e ampolas.
LESÕES NO MACHO O exame microscópico dos epidídimos infectados mostra edema intersticial, fibrose e infiltrado perivascular de linfócitos e plasmócitos. Os ductos epididimários mostram hiperplasia epitelial com cistos intraepiteliais, contendo neutrófilos e restos celulares.
Generated by Foxit PDF Creator © Foxit Softwarehttp://www.foxitsoftware.com For evaluation only.

Microbiologia Clínica Veterinária VET 3225 ÁREA DE BACTERIOLOGIA 2011-2
UFRGS
Microbiologia Clínica Veterinária VET 3225 ÁREA DE BACTERIOLOGIA 2011-2
UFRGS
Granuloma espermático circundado por linfócitos, células gigantes e epitelióides são achados frequentes.
A atrofia testicular deriva do processo regressivo do epitélio testicular e suspenção da espermatogênese. Pode também ocorrer proliferação do tecido conectivo intertubular, granuloma espermático extratubular, necrose e calcificação dos ductos seminíferos.
As principais alterações das vesículas seminais incluem a infiltração de linfócitos e plasmócitos; fibrose e hiperplasia epitelial difusa com cistos intraepiteliais, contendo neutrófilos.
A inflamação das ampolas está associada com a epididimite. Áreas focais de hiperplasia com cistos intraepiteliais vazios e acúmulo de neutrófilos no lúmen das dilatações do epitélio são geralmente observadas. Alem disso, acúmulo de células redondas e fibrose podem ser visualizados na lâmina própria.
O exame microscópico da próstata e da glândula bulbouretral revela uma discreta infiltração de células redondas e hipertrofia glandular focal. Formações papiliformes e concreções (corpora amilacea) são frequentes nessas duas glândulas acessórias. Nas ovelhas infectadas observa-se um exsudato purulento, variando de uma pequena quantidade sobre a superfície da membrana corioalantóide intacta até uma grande quantidade na área interplacentoma. O exsudato contém bactérias, macrófagos, neutrófilos e células epiteliais de descamação. Fibrose, espessamento e edema da membrana corioalantóide são observadas nos casos mais graves. Os cotilédones podem evidenciar vários graus de necrose. Células do epitélio coriônico podem estar aumentadas e conter bactérias. Necrose focal epitélio intercotiledonário e coriônico são frequentemente observados. Lesões nas artérias são comuns com trombos fibrinosos no interior de vasos; aumento de células endoteliais e proliferativas, na túnica intima. Não há lesões patognomônicas na infecção brucélica. ISOLAMENTO DE AMOSTRAS As amostras para o isolamento de B. ovis de animais vivos inclui: sêmen, suabe vaginal e leite. O sêmen pode ser colhido facilmente em suábios tomados da cavidade prepucial, apos a eletroejaculação. Se o eletroejaculador não é disponível, podemos coletar da vagina de fêmeas livre da infecção, imediatamente após a monta natural. A melhor técnica de diagnóstico direto é o isolamento bacteriano em meio de cultivo adequado. Amostras de sêmen, suábio vaginal, ou leite, devem ser semeados diretamente em placas, contendo meios adequados; incubadas a 37°C numa atmosfera de 5-10% de CO2. Os tecidos devem ser macerados e triturados com pequena quantidade de salina estéril ou PBS antes de serem semeado. A amostra de sêmen menos contaminada pode ser obtida pela exposição e limpeza do pênis e o ejaculado colhido em frasco estéril.
Nos animais mortos, a colheita de amostras para isolamento da B. ovis inclui nos machos, o epididimo, vesículas seminais, ampolas seminais, linfonodos inguinais e nas fêmeas, o útero, linfonodos ilíacos e supramamário. Entretanto para obter melhores resultados deveremos incluir outros órgãos e linfonodos como: baço, linfonodos cranial, pré-escapular, prefemural e testiculares. Cordeiros mortos e placenta devem ser examinados. As amostras preferenciais devem colhidas como o fluido do abomaso e pulmão. As amostras para cultivo devem ser refrigeradas e transportadas para o laboratório
Generated by Foxit PDF Creator © Foxit Softwarehttp://www.foxitsoftware.com For evaluation only.

Microbiologia Clínica Veterinária VET 3225 ÁREA DE BACTERIOLOGIA 2011-2
UFRGS
Microbiologia Clínica Veterinária VET 3225 ÁREA DE BACTERIOLOGIA 2011-2
UFRGS
o mais rápido possível após a coleta. A bactéria permanece viável por até 72 horas a temperatura ambiente e a sobrevivência pode ser aumentada a 4ºC ou preferencialmente pelo congelamento das amostras teciduais.
Esfregaços vaginais e de sêmen podem ser corados pela técnica de Stamp onde cocobastonetes característicos podem ser evidenciados na maioria dos infectados. O exame de amostras coradas pelo Stamp (trato genital do carneiro, linfonodo inguinal, placenta, conteúdo do abomaso e pulmão de feto) pode permitir o diagnóstico presuntivo rápido. Entretanto outras bactérias com morfologia similar ou características tintoriais semelhantes (B. melitensis, Coxiella burnetii e Chlamydophila spp) podem estar presentes nas amostras, tornando o diagnóstico difícil para profissionais menos experientes. A microscopia deve ser confirmada pelo cultivo do agente. IDENTIFICAÇÃO DO AGENTE A existência de lesão macroscópica (epididimite uni ou bilateral) no carneiro pode ser indicativa de infecção, mas exames laboratoriais são necessários para confirmar a doença, através de métodos diretos e indiretos.
Esfregaços diretos podem ser examinados, através da coloração de Gram ou Stamp e a presença de cocobastonetes demonstrada em muitos animais infectados. Entretanto, outras bactérias com características morfotintoriais semelhantes (B. melitensis, Chlamydophila abortus) podem estar presentes. Exame Direto
O diagnóstico direto é realizado, através do isolamento da B. ovis no sêmen, nos tecidos do carneiro ou ainda nas secreções ou leite de fêmeas em meio de cultivo seletivo. As técnicas de biologia molecular como PCR e eletroforese de campo pulsante têm sido aplicadas, entretanto as técnicas indiretas baseadas nos testes sorológicos são as preferidas no diagnóstico de rotina. O sêmen do carneiro pode ser obtido facilmente por meio do eletroejaculador. O sêmen pode ser colhido para bacteriológico em sacos plásticos ou por meio de suábio tomado da cavidade prepucial após a eletroejaculação. Cultivo
No isolamento da B. ovis, as amostras de sêmen são semeadas diretamente em plascas de AS apropriadas (Thayer-Martin); incubadas a 10% de CO2.
A colheita e transporte de amostras é uma tarefa trabalhosa, pois alguns animais infectados não excretam ou a eliminam intermitentemente e, por isso, o bacteriológico de sêmen não é prático e nem útil no diagnóstico da infecção, especialmente em programas de controle e erradicação de grande escala. Isolamento e identificação As colônias da B. ovis não são hemolíticas; são circulares, convexas; possuem bordos inteiros; são do tipo rugoso quando examinadas pela luz oblíqua; positivas ao teste de acriflavina. A B. ovis perdeu a atividade de urease; falha na redução de nitrato para nitrito; catalase positiva; oxidade negativa; não produz H2S; não cresçe na presença de
Generated by Foxit PDF Creator © Foxit Softwarehttp://www.foxitsoftware.com For evaluation only.

Microbiologia Clínica Veterinária VET 3225 ÁREA DE BACTERIOLOGIA 2011-2
UFRGS
Microbiologia Clínica Veterinária VET 3225 ÁREA DE BACTERIOLOGIA 2011-2
UFRGS
violeta de metila, mas geralmente cresce na presença de concentrações de fucsina básica e tionina. A maioria dos laboratórios de diagnóstico veterinário não está equipada na identificação completa da B. ovis, sendo necessária a adoção de um protocolo prático e presuntivo na identificação. A maioria das B. ovis pode ser identicada, tendo como base as caracteristicas de crescimento, observação direta, utilizando a luz refletida oblíqua, coloração de Stamp, Gram, catalase, oxidase, urease e teste de acriflavina. Entretanto, a identificação definitiva deve ser realizada no laboratório de referência com experiência na identificação e tipificação de Brucella spp. Recentemente, um método de eletroforese de campo pulsante foi desenvolvido para o gênero Brucella spp e com essa técnica é possível diferenciar B. ovis das outras espécies. Fagotipagem Os cultivos não são lisadas pelos bacteriófagos do grupo Tbilisi, Weybridge e Iz, no teste de diluição rotineiro (RTD) ou 104 RTD, enquanto são lisados pelo bacteriófago R/C. Diagnóstico sorológico Teste de Imunodifusão em gelose de agar (IDGA), Teste de Fixação do Complemento (FC) e, ensaio imunoenzimático (ELISA), utilizando antígenos de superfície solúveis da B. ovis, podem ser utilizados. Alguns testes de ELISA, que utilizam proteínas recombinantes e monoclonais, estão sendo testadas em pesquisas de campo. As sensibilidades dos testes IDGA e ELISA são semelhantes e o teste de ELISA apresenta maior sensibilidade do que o teste de FC. A combinação da IDGA e ELISA apresenta os melhores resultados em termos de sensibilidade, embora o teste de IDGA seja mais prático e barato. O teste recomendado para o comércio internacional permanece sendo o teste de FC. O LPS da parede celular é o principal antígeno de superfície na maioria das bactérias Gram negativas, sendo verdade para as brucelas lisas. Entretanto a B. ovis é rugosa e, portanto sua superficie celular difere das demais brucelas lisas. A principal diferença na estrutura da parede celular entre as brucelas lisas e rugosas é a ausência de cadeia O no LPS das amostras rugosas. O LPS da B. ovis assim como o LPS das mutantes rugosas de B. abortus e B. melitensis podem ser extraídas com o uso de solventes orgânicos. A técnica de éter-de-petróleo-cloforfórmio-fenol (PCP), originalmente desenvolvida para mutantes rugosas de enterobactérias é a técnica de escolha para obter LPS de B. ovis, uma vez que ela produz antígeno essencialmente livre de proteínas e ácidos nucléicos. Entretanto a grande hidrofobicidade do LPS purificado representa um problema para o seu uso em muitos testes sorológicos. A estrutura do LPS da B. ovis não tem sido estudada e conhecida. Embora ela contenha açúcar presente na estrutura núcleo-lipídio A do LPS de outras brucelas, o LPS purificado da B. ovis revela somente uma reação de identidade parcial com o LPS dos mutantes rugosos de B. abortus e B. melitensis ou com o LPS rugoso presente no antígeno extraído pelo calor da B. canis, sugerindo a presença de determinantes antigênicos espécie-específicos no LPS da B. ovis.
Generated by Foxit PDF Creator © Foxit Softwarehttp://www.foxitsoftware.com For evaluation only.

Microbiologia Clínica Veterinária VET 3225 ÁREA DE BACTERIOLOGIA 2011-2
UFRGS
Microbiologia Clínica Veterinária VET 3225 ÁREA DE BACTERIOLOGIA 2011-2
UFRGS
Antígenos O tratamento das brucelas rugosas pela técnica da salina aquecida (método da salina
quente), produz extratos antigênicos solúveis cujo componente principal precipita com o soro das brucelas rugosas. Por essa razão, o antígeno termorresistente tem sido referido como o antígeno rugoso específico ou quando obtido da B. ovis ele é chamado antígeno específico da B. ovis. Entretanto, a caracterização química do antígeno termorresistente da B. ovis mostrou que são enriquecidos com LPS rugoso, proteínas da membrana externa e outros componentes externos da membrana. Assim, o antígeno termorresistentes contém determinantes do LPS específicos para a B. ovis, mas outros componentes antigênicos adicionais. O antígeno termorresistente, devido a sua solubilidade em água e alta concentração em epítopos na superfície celular é o melhor antígeno para o diagnóstico e têm sido amplamente utilizados para o diagnóstico sorológico da infecção causada pela B. ovis.
A B. ovis cepa REO 198 é CO2 independente e por isso, recomendada como fonte de antígeno termorresistente para os testes sorológicos. Esta cepa foi obtida no Institute National de la Recherche Agronomique (INRA) Laboratoire de Pathologie Infectieuse et Immunologie, Nouzilly, França. Meios sólidos são satisfatórios para o crescimento da B. ovis REO 198. O ideal seria incluírem-se várias cepas ou linhagens na elaboração do antígeno.
O antigeno deve ser preparado como segue: 1) Crescimento exponencial de uma cepa de preferência aeróbia (B. ovis REO 198)
em frascos de Tripticase soja em incubador orbital a 37°C e a 150 rpm; ou em garrafas Roux de Tripticase Soja Agar ou outro meio adequado, com 5% de soro ou em fermentador descrito para B. abortus, mas com a adição de 5% de soro ao meio.
2) As células são lavadas 2 vezes e então suspensas em salina a 0,85% (12 g células-peso seco) ou (30 g de células em 150 ml).
3) A suspensão celular é autoclavada a 120°C por 15-30 minutos. 4) Após o resfriamento, a suspensão é centrifugada (15.000 X G; 4°C; 15 minutos) e
o sobrenadante é filtrado e dializado em água destilada (3 x 100 volumes) a 4°C por pelo menos 2 dias.
5) O fluido dializado é ultracentrifugado (100.000 X G; 4°C; 6-8 hours). 6) O sedimento é ressuspenso em uma pequena quantidade de água destilada e
liofilizado. 7) O antigeno é reconstituído em água destilada (para uso em IDGA) ou em tampão
salina-veronal (para uso no teste de FC) ou em tampão carbonato-bicarbonado (para uso em ELISA).
8) A titulação é feita contra os soros-controle positivos e negativos. Ensaio imunoenzimático (ELISAi)
Há inúmeras variações do ensaio imunoenzimático. O teste descrito aqui é um teste de ELISA indireto, utilizando o ABTS (2,2’-Azino-bis-[3-ethylbenzothiazoline-6-sulphonic acid]) como cromógeno. Os testes são realizados em placas de ELISA com fundo chato com 96 orificios. Os reagentes e as diluições são realizadas em PBS pH 7,2 com a adição
Generated by Foxit PDF Creator © Foxit Softwarehttp://www.foxitsoftware.com For evaluation only.

Microbiologia Clínica Veterinária VET 3225 ÁREA DE BACTERIOLOGIA 2011-2
UFRGS
Microbiologia Clínica Veterinária VET 3225 ÁREA DE BACTERIOLOGIA 2011-2
UFRGS
de 0,05% de Tween 20 (PBST). As diluições do antígeno são realizadas com tampão carbonato-bicarbonato com pH 9,6.
As placas são lavadas com PBST, após a fixação do antígeno. O antígeno termorresistente e o conjugado são titulados e as diluições são selecionadas para dar a melhor relação entre o soro padrão positivo e negativo. Secundariamente os anticorpos (anti-ovino IgG [H+L cadeias]) são geralmente conjugadas à peroxidase, embora outras enzimas possam ser utilizadas. Se o conjugado com peroxidase é utilizado, o cromógeno (ABTS) é diluído em tampão substrato pH 4 (composto de ácido cítrico trisódico e ácido cítrico). A isso é adicionada água oxigenada (H2O2), e as placas incubadas por 15-60 minutos a temperatura ambiente. A reação é bloqueada com 1 mM de azida sódica e a mudança de cor é lida em espectrofotômetro com filtro de 405-414 nm.
O antígeno utilizado no ELISA é o termorresistente estoque (1 mg/ml em tampão de sensibilização) titulado com diferentes diluições do antígeno, de conjugado e de substrato, contra um soro padrão ou contra diluições seriais de um painel de soros que são positivos e negativos para B. ovis, visando a determinação da diluição mais adequada (geralmente 5-10 µg/ml). As placas de ELISA são sensibilizadas com 100 µl de uma predeterminada diluição de antígeno em tampão carbonato, pH 9,6 para cada poço. As placas são incubadas por 2 horas a 37°C ou “overnigh” a 4°C. Posteriormente, elas são lavadas 4 vezes para remover o antígeno não ligado e, em seguida, são secas com batidinhas firmes com elas viradas para baixo sobre toalhas absorventes. As placas sensibilizadas podem ser utilizadas imediatamente ou estocadas a 4°C (a estabilidade nessas condições é adequada por pelo menos 1 mes).
Diluir os soros controles positivos e negativos a 1/200 pela adição de 10 µl do soro a 2 ml PBST. Adicionar um volume de 100 µl da amostra em duplicata na microplaca. As placas são cobertas, incubadas a 37°C por 1 hora; lavadas 3 vezes com tampão de lavagem.
O conjugado titulado e diluído em PBST é adicionado (100 µl) a cada poço e as placas cobertas e incubadas por 1 hora a 37°C. Após a incubação, as placas são lavadas novamente 3 vezes com PBST.
Solução de ABTS no tampão é adicionada (100 µl/orifício); as placas são incubadas por 15-60 minutos a temperatura ambiente com agitação continua.
A absorvância é lida automaticamente no espectrofotômetro com o comprimento de onda entre 405 a 414 nm. Os valores de absorvância podem ser expressos em percentagens de absorvância média dos controles positivos. Os valores de absorvância podem ser expressos como percentuais de absorvância médios do controle positivo transformados em unidades ELISA calculados, tanto manualmente quanto pelo programa de computador que calcula a curva de uma curva-padrão construída com uma série de resultados de diluições do controle positivo. O limiar deve ser calculado, testando uma quantidade grande da população ovina livre da infecção e a sensibilidade do teste sendo controlados na população de animais infectados pela B. ovis. Estudos comparativos evidenciaram que o teste de ELISA possui melhor sensibilidade que a IDGA ou a FC. A existência de alguns soros ELISA-negativos e IDGA positivos faz com que a combinação de IDGA com ELISA melhore a especificidade. Entretanto a combinação do teste de FC e ELISA ou CF e IDGA não melhoram a
Generated by Foxit PDF Creator © Foxit Softwarehttp://www.foxitsoftware.com For evaluation only.

Microbiologia Clínica Veterinária VET 3225 ÁREA DE BACTERIOLOGIA 2011-2
UFRGS
Microbiologia Clínica Veterinária VET 3225 ÁREA DE BACTERIOLOGIA 2011-2
UFRGS
sensibilidade do teste de ELISA sozinho. O teste de FC possui outras desvantagens, tais como:
a) Complexidade, b) Obrigatoriedade de inativação das amostras de soros; c) Atividade anticomplementária de alguns soros; d) Dificuldade de realização com soros hemolisados e e) Fenômeno de pró-zona.
A IDGA é o teste mais prático no diagnóstico de rotina em laboratórios menos especializados por sua sensibilidade, simplicidade e fácil interpretação.
Generated by Foxit PDF Creator © Foxit Softwarehttp://www.foxitsoftware.com For evaluation only.

Microbiologia Clínica Veterinária VET 3225 ÁREA DE BACTERIOLOGIA 2011-2
UFRGS
Microbiologia Clínica Veterinária VET 3225 ÁREA DE BACTERIOLOGIA 2011-2
UFRGS
Brucella canis Prof. Marcos JP Gomes BRUCELOSE CANINA
INTRODUÇÃO A brucelose canina era uma enfermidade associada à B. abortus, B. suis e a B.
melitensis, antes do isolamento da B. canis, em 1966. A fonte de infecção em quase todos os casos era o consumo de animais domésticos infectados ou pelo contato de cães com derivados destes. Em contraste à forma esporádica da brucelose canina, há outra, extremamente infecciosa para os cães, causada por uma nova espécie de Brucella. Em 1966, Carmichael, nos Estados Unidos, durante uma investigação sobre a causa de abortos em cães da raça Beagle, isolou um cocobastonete Gram negativo de vários fetos abortados. Mais tarde, várias outras observações deram suporte à inclusão deste organismo no gênero Brucella, sendo denominada B. canis. AGENTE-INFECÇÃO A B. canis é o agente etiológico da brucelose canina; enfermidade caracterizada por manifestações clínicas variadas. A maioria dos cães infectados não apresenta sinais clínicos perceptíveis, mostrando-se clinicamente normais.
As manifestações clínicas indicativas da enfermidade não são exuberantes, mas associadas com o trato reprodutor.
O sinal clínico primário na fêmea infectada é o aborto, podendo ser acompanhado ou não de mortes embrionárias ou natimortos.
Nos machos, os sinais clínicos mais comuns são: epididimite, orquite e/ou atrofia testicular e dermatite de escroto. Outras manifestações podem incluir: linfadenites e esplenites, lesões oculares, discoespondilites e osteomielites. A contaminação ocorre, através das mucosas (oral, vaginal ou ocular). A bactéria é fagocitada e será transportada por macrófagos e granulócitos para os linfonodos regionais e sistema circulatório.
Bacteremia aparece entre 1 a 4 semanas, após a infecção, podendo durar seis meses e, algumas vezes, podendo persistir por 60 meses. A fase de bacteremia é acompanhada de sinais clínicos como febre discreta, linfadenopatia discreta e astenia de inconstante a moderada. A fase de bacteremia pode levar lesões inflamatórias nos discos intervertebrais (discoespondilite), no globo ocular (uveíte) e nos rins (glomerulonefrite), mas o mais importante, é que o agente atinge o trato reprodutos de machos e fêmeas prenhas.
Nos machos, causa espermatozóides anormais, levando à esterilidade, após cinco semanas pós-infecção. Outros sinais como orquite, epididimite, hipertrofia prostática, edema do escroto podem ser detectados. Nas formas crônicas, há atrofia de um ou ambos os testículos.
Na fêmea prenha, a infecção geralmente leva a morte embrionária entre os dias 10 e 20 após a concepção ou ao abortamento entre 45 e 59 dias de gestação. O aborto é seguido de secreção vaginal que pode persistir de 1 a 6 semanas. Geralmente a cadela pare filhotes
Generated by Foxit PDF Creator © Foxit Softwarehttp://www.foxitsoftware.com For evaluation only.

Microbiologia Clínica Veterinária VET 3225 ÁREA DE BACTERIOLOGIA 2011-2
UFRGS
Microbiologia Clínica Veterinária VET 3225 ÁREA DE BACTERIOLOGIA 2011-2
UFRGS
mortos ou filhotes que sobrevivem apenas algumas horas ou dias. Raramente, a prenhez chega a termo.
As fontes de infecção são representadas por sêmen, urina, fetos abortados ou secreção vaginal. Os animais são infectados por via venérea ou pela ingestão de material infectado. Infecção iatrogênica (transfusão de sangue, uso de material de injecção contaminado) é possível. ZOONOSE
Os técnicos (laboratório, os proprietários de animais, veterinários) podem ser infectados, apesar de o homem ser relativamente resistente a B. canis. Os sinais clínicos (febre, calafrios, fadiga, perda de peso, linfadenopatia) são mais moderados do que aqueles observados em outras espécies e, em muitos casos, são completamente assintomáticos. As complicações são raras (artrite, meningite, endocardite), mas foram descritas.
A presença da brucelose canina nos centros urbanos brasileiros é preocupante, pois representa risco à saúde pública, pelo estreito convívio ou contato com os seus “familiares”, tanto pela população de cães infectados e não domiciliados quanto por aqueles cães infectados dentro de criatórios. Estudos epidemiológicos dirigidos à brucelose canina nestes centros urbanos forneceram dados importantes quanto a sua apresentação, distribuição, comportamento da enfermidade venérea para os caninos, mas também como uma zoonose, especialmente para criadores, proprietários, tratadores e clínicos veterinários. IMPACTO ECONÔMICO O impacto econômico decorrente da infecção pela B. canis nos criatórios é complexo e difícil de estimar, pois as perdas incluem principalmente a queda brusca no número de filhotes, eliminação de reprodutores infectados (machos e fêmeas) da reprodução; perda de material genético superior; gastos com a reposição de animais hígidos; os gastos com diagnóstico, tratamento, desinfecção e prevenção da infecção; gastos com monitoramento do criatório e a perda de confiança entre quem os criam e quem os compram. Talvez essas dificuldades sejam, em parte, razão para que nenhuma medida seja tomada por parte da sociedade interessada somente no comercio de animais a despeito do bem estar animal. É hora de estimar os prejuízos causados com a brucelose canina em criatórios, pois a infecção causada pela B. canis acomete um grande número de animais (canis), onde ela ocorre sob a forma silenciosa ou sob a forma de pequenos surtos, envolvendo animais com alto valor zootécnico. EPIDEMIOLOGIA A enfermidade foi registrada nos Estados Unidos; em países da Europa; no Japão, Madagascar e México. Na América do Sul, especialmente no Peru, Argentina e Brasil. Estudos epidemiológicos mostraram que a prevalência da infecção causada pela B. canis é variável, dependendo da área geográfica e do tipo de teste aplicado. Estudos sorológicos obtidos têm permitido valiosa informação sobre a enfermidade na população canina bem como na população humana, representando, deste modo, mais um risco à saúde pública.
Generated by Foxit PDF Creator © Foxit Softwarehttp://www.foxitsoftware.com For evaluation only.

Microbiologia Clínica Veterinária VET 3225 ÁREA DE BACTERIOLOGIA 2011-2
UFRGS
Microbiologia Clínica Veterinária VET 3225 ÁREA DE BACTERIOLOGIA 2011-2
UFRGS
BRUCELOSE CANINA NO BRASIL No Brasil, os primeiros relatos ocorreram simultaneamente em São Paulo e no Rio Grande do Sul, em 1976. Sandoval e colaboradores estimaram a infecção em 3,61%, dentre as 221 amostras de soro de cães apreendidos no centro de zoonoses da prefeitura municipal de São Paulo. Wald e Fernandes (1976-7), no Rio Grande do Sul, estimaram a prevalência da brucelose canina em 12% dentre as 192 amostras testadas de cães atendidos aleatoriamente pelo ambulatório do Hospital de Clínicas Veterinárias, em Porto Alegre. Pereira Filho e colaboradores, em 1978, na Bahia quantificaram a enfermidade em 1,43%, das 1.393 amostras de soro canino testadas em Salvador. Larsson, em 1979, obteve um percentual de 7,0% num levantamento realizado na população canina da cidade de São Paulo. Germano e colaboradores, em 1987, em Campinas, obtiveram um percentual de 5,4% de reagentes. Schlemper e Vaz, em 1990, em Santa Catarina, estimaram em 6,0% entre as 334 amostras de soro examinadas de cães na região do Planalto Catarinense. Magalhães Neto e colaboradores em 1992, em Pelotas, obtiveram uma prevalência de 22,7% entre 304 amostras de soro canino. Poester ecolaboradores, em 1994, estimaram a infecção em 7,4 % das 95 amostras da zona urbana de Uruguaiana. O isolamento da B. canis é considerado como o "padrão-ouro" no diagnóstico da brucelose canina. Ela é chave importante para a aplicação de medidas no diagnóstico, controle e prevenção da infecção/doença. ISOLAMENTO NO MUNDO E BRASIL O isolamento da B. canis pode ser obtido do sangue periférico de animais com títulos altos de anticorpos (Moore e Kakuk, 1969).
Serikawa et al., (1978) isolaram B. canis da urina e da próstata de cães infectados. Ueda et al., (1974) isolaram B. canis do baço, epidídimo, próstata e linfonodos poplíteos e ilíacos. Harris et al., (1974) isolaram B. canis do sangue, fluido cerebrospinal e de vários
órgãos de fêmeas com sinais neurológicos. Henderson et al., (1974), nos Estados Unidos, isolaram a B. canis de 3 cães com
lesões de osteomielite vertebral na região tóraco-lombar (discoespondilites). Riecke e Rhoades (1974) e Seagusa et al., (1977) isolaram B.canis de lesões
oculares. Schoeb e Morton (1978) isolaram o agente da brucelose canina de úlcera exsudativa
escrotal. No Brasil, Fernandes e Wald em (1976-7) isolaram, pela primeira vez, no Brasil a
B. canis do humor aquoso de cão da raça Boxer, com lesões oculares. Em 1977, em Minas Gerais, Godoy e colaboradores isolaram a B. canis em
hemocultura de uma fêmea que havia abortado recentemente. Larsson e Costa (1980), em São Paulo, isolaram três amostras de B. canis, sendo 1
amostra de fêmea com histórico de infertilidade, e as outras duas de uma fêmea e de um macho sem sinais clínicos.
Vargas et al., (1996), em Santa Maria, isolaram B. canis da placenta, de fetos abortados e neonatos de um canil com reprodutores caninos de várias origens.
Generated by Foxit PDF Creator © Foxit Softwarehttp://www.foxitsoftware.com For evaluation only.

Microbiologia Clínica Veterinária VET 3225 ÁREA DE BACTERIOLOGIA 2011-2
UFRGS
Microbiologia Clínica Veterinária VET 3225 ÁREA DE BACTERIOLOGIA 2011-2
UFRGS
Gomes et al., (1999), em Porto Alegre, isolaram B. canis do epidídimo e testículo de um cão com epididimite e orquite clínica.
No Rio de Janeiro, Ferreira e colaboradores (2003) isolaram e identificaram B. canis de dois cães com sinais clínicos de brucelose canina de um canil comercial. DIAGNÓSTICO LABORATORIAL O diagnóstico laboratorial da brucelose canina é uma tarefa difícil e complexa, incluindo as técnicas de isolamento, segundo evidencia a Tabela 1.
O diagnóstico sorológico é uma probabilidade diagnóstica, pois dependem da relação agente-hospediero e o tempo de infecção. Eles incluem o teste de fixação do complemento (FC), soroaglutinação lenta (SAL), soroaglutinação rápida (SAR), imunofluorescência (IF), o teste de ELISA e imunodifusão (IDGA). A imunodifusão em gelose de agar (IDGA) é a técnica mais utilizada no diagnóstico laboratorial da infecção por ser de fácil realização, rápida e barata. Além disso, esta técnica possibilita utilizar, tanto o antígeno superficial rugoso da B. ovis quanto da B. canis e da RB51. HEMOCULTIVO
O hemocultivo da B. canis pode ser obtid através da seguinte técnica: 1- Um volume de 0,1 mL de sangue, colhido com heparina ou citrato de sódio é semeado em placas de agar sangue (Brucella agar, Triptose agar); incubadas a temperatura de 37°C em atmosfera normal. 2- Um volume de 5 mL de sangue é semeado em 10 mL de caldo. O caldo inoculado deve ser congelado de 12 a 24 h a -20°C. Posteriormente, a amostra congelada deve ser descongelada e incubada por 6 dias a 37°C. Após esse tempo, a amostra é repicada em 2 meios em paralelo (seletivos e não seletivo).
Os meios de Caldo Brucella soro; Triptose soro; Kuzdas e Morse tornam-se seletivo pela adição de cicloheximida (100 mg/L), bacitracina (25 000 U/L) e polimixina B (6 000 U/L), dão bons resultados. SÊMEN e URINA
Sêmen e urina devem ser cultivados por 4-7 dias em caldo seletivo e posteriormente em gelose. Outras amostras clínicas eventualmente são trituradas em stomacher; inoculadas em paralelo em meios seletivos e não seletivos.
As placas devem ser conservadas por 30 dias; examinada periodicamente em estereoscópio com luz obliqua. As colônias suspeitas devem ser submetidas ao exame bacterioscópico, teste de oxidase e tipo respiratório. A identificação precisa da espécie está reservada aos laboratórios especializados que pode pesquisar a aglutinação com antissoro preparado com uma cepa de Brucella em fase R (geralmente uma cepa de Brucella ovis) posteriormente se estudará as características bioquímicas; lisotipia e oxidação de açúcares e aminoácidos.
Generated by Foxit PDF Creator © Foxit Softwarehttp://www.foxitsoftware.com For evaluation only.

Microbiologia Clínica Veterinária VET 3225 ÁREA DE BACTERIOLOGIA 2011-2
UFRGS
Microbiologia Clínica Veterinária VET 3225 ÁREA DE BACTERIOLOGIA 2011-2
UFRGS
Tabela 1 Resultados Esperados no isolamento da B. canis em amostras clinicas
Amostra(s) Isolamento Pós-Infecção (PI)
Fluidos (vaginal placenta, aborto)
Positivo
Sêmen Positivo entre 3 a 11 semanas PI
Positivo ou negativo em 12 a 60 semanas PI
Negativo para além de 60 semanas PI
Sangue Positivo entre 5 a 30 semanas PI
80% positivo em 6 a 12 meses PI
50 – 80% positivo em 24 a 48 meses PI
25 – 50% positivo em 48 a 58 meses PI
0 – 25% positivo para além 58 meses PI
Urina Geralmente positivo (geralmente ♂) entre 8 - 30 semanas PI
Epidídimo 50 – 100% positivo entre 35 - 60 semanas PI
Negativo além de 24 meses PI
Próstata Geralmente positivo até 64 semanas PI
Linfonodo, baço, medula óssea
Geralmente positivo quando o animal está na fase de bacteremia
Generated by Foxit PDF Creator © Foxit Softwarehttp://www.foxitsoftware.com For evaluation only.

Microbiologia Clínica Veterinária VET 3225 ÁREA DE BACTERIOLOGIA 2011-2
UFRGS
Microbiologia Clínica Veterinária VET 3225 ÁREA DE BACTERIOLOGIA 2011-2
UFRGS
CONTROLE E PREVENÇÃO O controle torna-se um dilema, quando a brucelose foi diagnosticada num criatório,
A eliminação da infecção demanda tempo e custos, mas a doença pode apresentar uma crise emocional ou quando cães de valor genético são acometidos.
A prevenção é particularmente importante nos canis. Ao contrário da brucelose nos outros animais domésticos, a brucelose canina não é uma doença de notificação obrigatória e sua prevalência está baseada em limitados estudos sorológicos. A incidência da enfermidade nos cães é pouco diferente de quando ela foi reconhecida nos anos sessenta; com exceção de alguns criatórios comerciais e organizações de criadores de cães ou clubes de caça que instituíram medidas preventivas de maneira privada.
A prevenção da reprodução em canis/criatórios deverá incluir um plano de monitoramento alem de cuidados sanitários e de higiene.
Os testes sorológicos incluem vantagens e desvantagens que devem ser conhecidas pelos clínicos veterinários conforme tabela 2.
1-Os testes sorológicos devem ser realizados duas vezes com intervalos de um mês em todos os cães introduzidos em um canil de reprodução.
2-As cadelas de um criatório/canil devem ser testadas várias semanas antes do cio esperado. Se outros testes alem do SAT-2ME são necessários para certificação de livres de brucelose, haveria tempo para aplicar aqueles procedimentos antes do período do cio.
3-Nenhum animal novo deve ser introduzido na reprodução até que tenham sido considerados negativos a dois testes com um mês de intervalo.
4-Testes anuais deverão ser aplicados em todos os animais ou quando houver alterações reprodutivas ou abortos.
5-Há fortes suspeitas de brucelose canina nos casos de aborto até que seja provado o contrário.
6-Cães com testes de triagem positivos ou inconclusivos devem ser isolados e avaliados por hemocultura e sorologia, utilizando testes mais específicos como AGID, utilizando antígeno de proteínas citoplasmáticas.
7-Todo cão infectado deve ser removido do canil e eliminado. Em alguns casos, há recusa por parte dos proprietários e, nesses casos, há indicação da castração concomitantemente à terapia com antimicrobianos e acompanhamento do caso, em pelo menos, três meses.
Generated by Foxit PDF Creator © Foxit Softwarehttp://www.foxitsoftware.com For evaluation only.

Microbiologia Clínica Veterinária VET 3225 ÁREA DE BACTERIOLOGIA 2011-2
UFRGS
Microbiologia Clínica Veterinária VET 3225 ÁREA DE BACTERIOLOGIA 2011-2
UFRGS
Tabela. 2 Comportamento dos testes aplicados à Brucelose canina por B. canis
Teste Positivar Resultados
Persistência Positividade
Vantagens e Desvantagens
Aglutinação rápida em lâmina (antígeno corado com Rosa Bengala), após tratamento do soro com 2-ME
3 a 4 semanas PI
Até 3 meses pós bacteremia
Simples, rápido, sensível, poucos falsos negativos.
Muitos falsos positivos (teste com resultados positivos devem ser confirmados por outra técnica).
Aglutinação lenta em tubos
3-6 semanas PI
Até 3 meses pós bacteremia
Técnica semi-quantitativa.
Numerosos falsos +, inutilisável com soros hemolisados, fenômeno de zona
possível.
Aglutinação lenta em tubos na presença de 2-ME
1,2-2 meses PI
Até 3 meses pós bacteremia
Técnica semi-quantitativa, + especifica que aglutinação lenta em tubos.
Inutilisável com soros hemolisados, fenômeno de zona possivel, necessita
48 h de incubação
IDGA (antígeno LPS)
1,5-2,5 meses PI
Até 4 meses pós bacteremia
Sensível, + específico que os testes anteriores.
Técnica complexa, interpretação delicada
IDGA (antígenos citoplasmáticos,
de natureza proteíca)
2-3 meses PI
Até 3 anos pós bacteremia
O teste mais específico (detectar animais infectados por outras espécies do gênero Brucella, mas não apresenta
reação cruzada com outros gêneros bacterianos), detecta infectado
cronicamente.
Teste complexo, sensíbilidade menor que o anterior
Generated by Foxit PDF Creator © Foxit Softwarehttp://www.foxitsoftware.com For evaluation only.

Microbiologia Clínica Veterinária VET 3225 ÁREA DE BACTERIOLOGIA 2011-2
UFRGS
Microbiologia Clínica Veterinária VET 3225 ÁREA DE BACTERIOLOGIA 2011-2
UFRGS
VACINAÇÃO Não existe vacina e a profilaxia baseia-se sobre as medidas de ordem sanitárias tais
como isolamento do agente; eutanásia dos infectados; desinfecção dos locais; quarentena e vigilância serológica antes da introdução em uma criação.
O tratamento com antimicrobiano não é 100% eficaz. Ele geralmente está associado ao uso de tetraciclinas a aminosídio durante um mês. SENSIBILIDADE AOS ANTIMICROBIANOS
A B. canis “in vitro” é sensível às tetraciclinas; ao cloranfenicol; aos aminosídios; à rifampicina, às fluoroquinolonas e às sulfamidas. Muitas linhagens apresentam resistência cruzadas aos macrolidios. Quadro I. Percentual de reagentes contra a B. canis, na IDGA, (antígeno LPS) no LABACVET-UFRGS
Ano Nº Amostras Reagentes %
1994 47 5 10,63
1995 29 2 6,89
1996 130 7 5,38
1997 72 4 5,55
1998 108 15 13,88
1999 34 8 23,52
2000 ? ? ?
2001 76 02 2,63
2002 35 0 00
2003 27 5 19,23
2004 145 29 20,00
2005 79 12 15,10
2006 184 23 12,50
2007 424 93 22,00
2008 70 4 5,70
2009 31 1 3,2
2010 3 1 33,3
2011
PERSPECTIVAS A preocupação em conhecer dados sobre brucelose canina nas diferentes faculdades de veterinária do país tem como objetivos: 1) Obter dados quanto à prevalência de brucelose canina em cães atendidos pelo serviço de atendimento policlínico nas instituições de ensino superior, especialmente por não haver muitos registros. 2) Isolar a B. canis em amostras clínicas bem como aplicar testes sorológicos no monitoramento e controle da infecção. 3) Prestação de serviço especializado à comunidade acadêmica pelo treinamento de técnicos, estudantes e outros profissionais 4)Garantir segurança dos acadêmicos e professores quando do manuseio desses animais em aulas práticas de clinica médica e cirurgia.
Generated by Foxit PDF Creator © Foxit Softwarehttp://www.foxitsoftware.com For evaluation only.

Microbiologia Clínica Veterinária VET 3225 ÁREA DE BACTERIOLOGIA 2011-2
UFRGS
Microbiologia Clínica Veterinária VET 3225 ÁREA DE BACTERIOLOGIA 2011-2
UFRGS
Brucelose nos Mamíferos Marinhos Prof. Marcos JP Gomes SINONÍMIA: Brucella ceti : "Brucella cetaceae". Brucella pinnipedialis: "Brucella pinnipediae". CONSIDERAÇÕES GERAIS Os estudos de hibridização do ADN-ADN têm mostrado que o gênero Brucella constitui uma única espécie genômica. Segundo as regras de prioridade, Verger e colaboradores propuseram que o nome de B. melitensis, B. abortus, B. canis, B. neotomae, B. ovis e B. suis deveriam ser considerados como simples biovares da Brucella melitensis. Esta proposição respeitou (e respeita hoje) os critérios utilizados para a definição de uma espécie bacteriana apesar de que em 1988, o subcomitê de taxonomia de o gênero Brucella ter considerado a existência de uma única espécie no gênero Brucella, este mesmo comitê admite o uso da nomenclatura anterior bem como o uso corrente em trabalhos baseados na taxonomia. A redução do gênero Brucella a uma única espécie não é adotada por parte da grande maioria dos bacteriologistas e compreende parte dos especialistas do gênero Brucella. Depois da reunião de 11 septembro de 2003, o subcomitê de taxonomia de Brucella decidiu por unanimidade rever a situação anterior e de reconhecimento à existência de novas espécies dentro do gênero Brucella. O reconhecimento dessas espécies é justificado por estudos fenotípicos, pela epidemiologia, pela importância dessas bactérias dentro da Saúde Animal e Humana e pela utilização potencial delas em guerra bacteriológica. Classicamente, as espécies do gênero Brucella foram isoladas de mamíferos terrestres. Mais tarde, a partir de 1994, amostras do gênero Brucella spp foram isoladas de mamíferos marinhos. As características fenotípicas, os estudos de hibridização, sequenciamento do ARNr 16S e o sequenciamento do gene recA mostraram que essas cepas ou linhagens pertenciam ao gênero Brucella. O estudo do metabolismo oxidativo permitiu individualizar as amostras isoladas de mamíferos marinhos, permitindo que Jahans e colaboradores propusessem a espécie "Brucella maris". Posteriormente, a ribotipagem (estudo dos fragmentos de restrição dos genes codantes para o ARNr obtido pela enzima HindIII), a presença do segmento de inserção IS711 em aval do gene bp26 e a presença de “motifs” específicos dentro da sequência dos genes omp2 confirmaram que as cepas isoladas de mamíferos marinhos eram distintas daquelas isoladas de mamíferos terrestres. As linhagens isoladas de mamíferos marinhos podem ser diferenciadas em 2 grupos. As cepas isoladas principalmente de penípedes possuem uma cópia dos genes omp2a e omp2b enquanto que as cepas isoladas principalmente de cetáceos possuem 2 cópias do gene omp2b. Assim, em 2001, Cloeckaert e colaboradores propuseram a nomenclatura de "Brucella pinnipediae" para as cepas principalmente isoladas de penípedes e a nomenclatura de "Brucella cetaceae" para as linhagens isoladas principalmente de cetáceos. A técnica de IRS-PCR (Interspersed Repetitive Sequence-PCR) permitiu
Generated by Foxit PDF Creator © Foxit Softwarehttp://www.foxitsoftware.com For evaluation only.

Microbiologia Clínica Veterinária VET 3225 ÁREA DE BACTERIOLOGIA 2011-2
UFRGS
Microbiologia Clínica Veterinária VET 3225 ÁREA DE BACTERIOLOGIA 2011-2
UFRGS
igualmente reconhecer a existencia de 2 grupos de cepas. A nomenclatura de "Brucella cetaceae" e da "Brucella pinnipediae" não é correta. Em 2007, Foster e colaboradores propuseram a nomenclatura de B. ceti (para as cepas isoladas de cetáceos) e de B. pinnipedialis (para cepas isoladas de penípedes). Essa nomenclatura foi validada e publicada em 08 novembro de 2007.
A Brucella foi cultivada ou detectada com técnicas de DNA em pinípedes e muitas espécies de cetáceos, incluindo focas comuns foca do porto (Phoca vitulina), foca anelada (Phoca hispida), focas harpa (Phoca groenlandica), foca de capuz (Cystophora cristata), foca cinzenta (Halichoerus grypus), boto (Phocoena phocoena), curto-bico golfinhos comuns (Delphinus delphis), golfinhos listrados (Stenella coeruleoalba), golfinhos-roaz (Tursiops truncatus), os golfinhos de lado branco do Atlântico (Lagenorhynchus acutus), golfinho de bico branco (Lagenorhynchus albirostris), os golfinhos de Maui (Cephalorhynchus hectori Maui), golfinhos-cabeça-branca ou golfinho de Hector (Cephalorhynchus hectori), a baleia minke (Balaenoptera acutorostrata) e uma lontra europeia (Lutra lutra). CARACTERÍSTICAS GERAIS As novas espécies B. ceti e B. pinnipedialis são constituidas de cocos, cocobastonetes e/ou de bacilos curtos; Gram negativos; com 0,5 a 0,7µm de diâmetro e de 0,6 a 1,5µm de comprimento; se apresentam de maneira isolada ou agrupada em curtas cadeias ou em pequenos filamentos; possuem os antígenos do gênero Brucella; são imóveis e desprovidas de flagelos, aeróbias; catalase, oxidase nitrato redutase e urease positivos; indol negativas; H2S negativas; incapazes de liquefazer a gelatina e não acidificam os açúcares contidos em meios convencionais. Crescimento é obtido em temperatura compreendida entre 20 e 40°C (temperatura ótima 37 °C) e pH ótimo compreendido entre 6,6 e 7,4. No AS com sangue ovino, as colônias são visíveis após 3 a 4 dias de incubação a 37°C. Elas são convexas, circulares, de contorno regular, não hemolítica e com 0,5 a 1,0 mm de tamanho. O crescimento é estimulado pela adição de soro ou sangue. No meio de gelose glicose-soro, as colônias são transparentes, convexas, lisas, brilhantes, de contornos regulares e apresentam uma coloração mel pálida quando examinadas sob iluminação oblíqua. B. ceti é capaz de crescer em 3 a 4 dias em meio Farrell enquanto que o crescimento da B. pinnipedialis é retardado (7 a 10 dias) ou não observado. A grande parte das cepas da B. ceti é cultivada na ausência de CO2 enquanto que o CO2 é necessário para a grande maioria das amostras da B. pinnipedialis. As características que permitem diferenciar a B. ceti e B. pinnipedialis das outras espécies estão contidas no Quadro 1.
Generated by Foxit PDF Creator © Foxit Softwarehttp://www.foxitsoftware.com For evaluation only.

Microbiologia Clínica Veterinária VET 3225 ÁREA DE BACTERIOLOGIA 2011-2
UFRGS
Microbiologia Clínica Veterinária VET 3225 ÁREA DE BACTERIOLOGIA 2011-2
UFRGS
Quadro 1. Diferenciação entre espécies do gênero Brucella spp
1) B. abortus ; 2) B. canis ; 3) B. ceti ; 4) B. melitensis ; 5) B. microti* ; 6) B. neotomae ; 7) B. ovis ; 8) B. pinnipedialis ; 9) B. suis biovar 1 ; 10) B. suis biovar 2 ; 11) B. suis biovar 3 ; 12) B. suis biovar 4 ; 13) B. suis biovar 5
1 2 3 4 5 6 7 8 9 10 11 12 13
Urease +**** + + + + + - + + + + + +
VP - - - - + - - - - - - - -
Fago Tb** (Tbilisi)
+ - - - - d - -***** - - - - -
Fago Wb** (Weybridge)
+ - + - + + - + + + + + +
Fago Iz** (Izatnagar)
+ - + + + - + + + + + +
Fago** R/C - + - - - + - - - - - -
L-Alanina*** + - - + - d - d - d - -
L-Arabinose*** + - + - + - - + + - - -
L-Arginina*** - + - - - - - + + + + +
L-Asparagina***
+ - - + + + - - d - - d
meso-Eritritol***
+ d d + + - + + + + + +
D-Galactose*** + - + - + - - + + - - -
L-Ácido glutâmico***
+ + + + + + + d + + + +
L-Lisina*** - d - - - - - + d d d d
DL-Ornitina*** - + - - - - - + + + + +
D-Ribose*** + + + - d - + + + + + +
D-Xilose*** - - + - - - - + d d d d
Hospedeiros preferenciais
Bov Can Cet Pr Cd Neo Ov Penip Sui Sui, Leb
Sui Re Sui
1 2 3 4 5 6 7 8 9 10 11 12 13
1) B. abortus ; 2) B. canis ; 3) B. ceti ; 4) B. melitensis ; 5) B. microti* ; 6) B. neotomae ; 7) B. ovis ; 8) B. pinnipedialis ; 9) B. suis biovar 1 ; 10) B. suis biovar 2 ; 11) B. suis biovar 3 ; 12) B. suis biovar 4 ; 13) B. suis biovar 5 * : Outras caract. permitem diferenciar a B. microti das outras espécies do gênero Brucella estão contidas na tabela II ** : Líticos na diluição corrente. *** : Oxidação **** : A cepa de referencia 544 assim como qualquer outra dá resultado negativo. ***** : Todas as amostras da Brucella pinnipedialis são lisadas pelo fago Tb. B: Bovinos; Ca: Cd dos campos (Microtus arvalis); Can: Caninos; Ce: Cetáceos; Pr: Pequenos Ruminantes; Ov: Ovinos Ne: Neotomos; Sui: Suinos; Leb: Lebres; Re: Renas.
Generated by Foxit PDF Creator © Foxit Softwarehttp://www.foxitsoftware.com For evaluation only.

Microbiologia Clínica Veterinária VET 3225 ÁREA DE BACTERIOLOGIA 2011-2
UFRGS
Microbiologia Clínica Veterinária VET 3225 ÁREA DE BACTERIOLOGIA 2011-2
UFRGS
SINAIS CLÍNICOS As alterações reprodutivas como aborto, em fêmeas; orquite/epididimite, lesões granulares, em machos, são sinais primários da brucelose nos animais terrestres. A avaliação da fertilidade em animais marinhos de vida livre é difícil, entretanto nos Estados Unidos, em 1999, Miller e colaboradores relataram casos de aborto em dois golfinhos (Tursiops truncatus) capturados. O DIAGNÓSTICO DIFERENCIAL
Em golfinhos com meningite, o diagnóstico diferencial inclui parasitismo (Nasitrema spp), infecção estafilocócica, herpes e infecções por Morbillivirus. Outras doenças que causam aborto, orquite, epididimite, abscessos e doenças sistêmicas devem ser consideradas e evidência de infecção por Brucella. ESTUDOS MOLECULARES Os estudos moleculares evidenciaram que as cepas dos mamíferos marinhos diferem das dos mamíferos terrestres e mesmo entre as linhagens que acometem os cetáceos e os pinípedes. Algum tempo atrás, pesquisadores sugeriram a criação de uma nova espécie denominada B. maris, entretanto ela não foi aceita. Mais recentemente, duas novas espécies foram sugeridas; a B. cetaceae e a B pinnipediae, as quais ainda não foram oficialmente aceitas nem rejeitadas.
Em 2007, Foster e colaboradores sugeriram os nomes de B. ceti e B. pennipedialis para as brucelas de cetáceos e de focas, respectivamente (Foster et al 2007). ZOONOSE A brucelose é uma importante zoonose para o homem causando a grande variedade de sinais e sintomas clínicos, incluindo febre ondulante, fadiga, prostração, dor articular, mialgia, depressão e anorexia. Frequentemente ocorrem sequelas e períodos de recrudescência, após o episódio inicial de infecção. A brucela pode ser transmitida de animais para o homem pelo contato direto com animais infectados; pela ingestão de produtos infectados e pela inalação de aerossóis. Quatro espécies do gênero Brucella (classif. atual) são causas primárias de infecção no homem. A B. melitensis é bastante infecciosa, sendo transmitida por caprinos e ovinos. A B. abortus é transmitida por bovinos. A B. suis transmitida por suínos e a B. canis transmitida por cães. As outras espécies de Brucella infectam raramente ou não infectam o homem. Há alguns poucos relatos na literatura relativos à infecção humana causada por linhagens de Brucella de mamíferos marinhos.
Brew et al. 1999 registraram o caso ocorrido em um laboratorista que adquiriu os sinais clínicos compatíveis com brucelose. A infecção foi confirmada pelo isolamento, testes diagnósticos sorológicos, PCR, RFLP de Brucella de origem marinha ().
Sohn et al. (2003) relataram em dois pacientes peruanos, os quais foram diagnosticados como portadores de neurobrucelose. O quadro clínico foi confirmado pelos
Generated by Foxit PDF Creator © Foxit Softwarehttp://www.foxitsoftware.com For evaluation only.

Microbiologia Clínica Veterinária VET 3225 ÁREA DE BACTERIOLOGIA 2011-2
UFRGS
Microbiologia Clínica Veterinária VET 3225 ÁREA DE BACTERIOLOGIA 2011-2
UFRGS
testes diagnósticos (isolamento, PCR, sequenciamento de DNA) como infectados naturalmente por linhagens de Brucella de mamíferos marinhos. Na Nova Zelândia, McDonald e colaboradores, em 2006, relataram um caso em um homem de 43 anos, morando em Auckland - Nova Zelandia, o qual apresentou sinais de osteomielite espinal por 2 semanas com sinais de febre, rigor e fraqueza lombar. Os testes aplicados reagiram tanto para a B. suis quanto B. melitensis. As amostras foram então enviadas ao laboratório de Referência internacional que o identificou como relacionado a Brucella originária dos Estados Unidos de golfinhos (Tursiops truncatus) e da foca comum (Phoca vitulina). EPIDEMIOLOGIA – PREVALÊNCIAS Há na literatura relatos em muitos lugares e em diversas ocasiões de isolamento de Brucella e comprovação sorológica em muitos mamíferos marinhos, especialmente no Hemisfério Norte. A prevalência da detecção de anticorpos contra Brucella nos animais marinhos variou, segundo diversos autores entre 0 a 38% dos cetáceos, dos penípedes e mustelídeos (Jepson et al. 1997; Tryland et al. 1999; Calle at al. 2002; Hanni et al. 2003; Maratea et al. 2003; Ohishi et al 2003; Nielsen et al 2005). Um grande estudo, envolvendo 1.855 penípedes dos EUA e 1.386 penípedes e cetáceos do Atlântico Norte revelaram que 3,1 e 8,2% tinham sorologia positiva para brucelas marinhas, respectivamente (Tryland et al 1999; Nielsen et al 2001). Amostras de Brucella foram isoladas em 31% (54/175) dos mamíferos marinhos provindos de diferentes origens (Forbes et al 1993; Ewalt et al 1994; Ross et al 1994; Foster et al 1996; Clavareau et al 1998; Miller et al 1999; Tryland et al 1999; Gonzalez et al 2002; Maratea et al 2003; Tryland et al 2005).
Há poucos relatos de sorologia positiva em penípedes e cetáceos no Hemisfério Sul. Retamal e colaboradores, na Antártica, em 2000, estimaram em 3,5% (6/17) das amostras de penípedes positivas para brucelose. Van Bressem e colaboradores, em 2001 quantificaram em 55, 2% (32/58) das amostras reagentes à prova de brucelose nos cetáceos examinados, nas costas peruanas do Pacífico Sul. Na Austrália, Dawson (2005) detectou reações sorológicas positivas em 3 espécies incluindo 75% (9/12) dos leões marinhos (Neophoca cinerea). Na Nova Zelândia, Mackereth e colaboradores, em 2005 não detectaram reagentes positivos em 1001 leões marinhos da Nova Zelândia (Arctocephalus
hookeri). LESÕES Pequenas alterações patológicas têm sido relatadas nos primeiros estudos nos animais marinhos encalhados no leste do Atlântico Norte apesar do sucesso no isolamento de agentes cepas pertencentes ao gênero Brucella. Contrariamente com as lesões granulomatosas e caseosas observadas e relatadas nas gônadas da baleia minque (B. acutorostrata) no Oeste do Pacífico. Lesões granulomatosas caseosas testiculares foram constatadas em 31% (11/35) e 38% (35/93) em machos da baleia minque do Pacífico Norte capturados em 2000 e 2001 respectivamente.
Generated by Foxit PDF Creator © Foxit Softwarehttp://www.foxitsoftware.com For evaluation only.

Microbiologia Clínica Veterinária VET 3225 ÁREA DE BACTERIOLOGIA 2011-2
UFRGS
Microbiologia Clínica Veterinária VET 3225 ÁREA DE BACTERIOLOGIA 2011-2
UFRGS
COLETA DE AMOSTRAS As amostras são isoladas de pulmões, do baço, do fígado, do líquido peritoneal, do coração, dos rins, do epidídimo, dos ovários, do leite, de abscesso, dos linfonodos e do sangue. Em alguns animais, nenhuma patologia pode ser associada à presença do agente, enquanto em outros animais pode-se notar epididimite discoespondilite, meningite, lesões purulentas, necrose hepática, necrose esplênica, necrose dos linfonodos, peritonite, mamite e endometrite. O diagnóstico presuntivo da infecção pode ser evidenciado através da presença de anticorpos anti-Brucella em outras espécies: golfinho sombra (Lagenorhynchus obscurus), marsopa espinosa (Phocoena spinipinnis), baleia piloto (Globicephala melas), baleia comum (Balaenoptera physalus), baleia boreal (Balaenoptera borealis), foca de Weddell (Leptonychotes weddellii), morça do Atlantico (Odobenus rosmarus rosmarus), foca de Kerguelen (Arctocephalus gazella) . Estudo realizado em animais vivos nas costas da Escócia mostrou que aproximadamente 49% das focas comuns e 33% dos botos eram soropositivos. A ingestão de mamíferos marinhos ou alimentos contaminados por esses animais podem representar perigo para os animais silvestres e domésticos. Os ursos polares consomem grandes quantidades de focas e um estudo mostrou que 5,4% dos ursos polares apresentarm anticorpos anti-Brucella. As carcaças dos mamíferos marinhos encontrados nas praias podem representar um risco para os animais que crculam naquele ambiente. A inoculação experimental por via intravenosa de cepas de Brucella isoladas de mamíferos marinhos em bovinos provocou abortos e a mesma amostra pode ser reisolada do feto. Por sua vez, a inoculação por via intraconjuntival não provoca soroconversão. Em cabras prenhes, a inoculação por via conjuntival provocou uma soroconversão transitória. DIAGNÓSTICO LABORATORIAL O isolamento é realizado em paralelo em meio de Farrell (incubado pelo menos por 14 dias após a conclusão do cultivo) e em AS incubado a 37 °C em atmosfera enriquecida com 10% de CO2. O meio de Farrell deve conter concentrações menores de bacitracina e/ou de ácido nalidíxico, pois favorecem o cultivo da Brucella pinnipedialis. O aspecto da colônia; o exame microscópico após a coloração de Gram e a pesquisa de anticorpos aglutinantes em lâmina com soro anti-Brucella abortus permitem orientação do diagnóstico. O diagnóstico da espécie é mais complexo, devendo ser confiado a laboratórios especializados. O diagnóstico sorológico emprega os testes clássicos utilizados para o diagnóstico de brucelose dos mamíferos terrestres (AAT, FC, ELISA, etc.). Embora esses testes ainda não estejam validados. CONTROLE
Métodos específicos de controle não foram estabelecidos para a brucelose em mamíferos marinhos. Princípios gerais de controle de infecção, incluindo o isolamento, desinfecção e higiene devem ser utilizadas com os animais infectados em instalações
Generated by Foxit PDF Creator © Foxit Softwarehttp://www.foxitsoftware.com For evaluation only.

Microbiologia Clínica Veterinária VET 3225 ÁREA DE BACTERIOLOGIA 2011-2
UFRGS
Microbiologia Clínica Veterinária VET 3225 ÁREA DE BACTERIOLOGIA 2011-2
UFRGS
marítimas de mamíferos. Alguns autores sugerem que os centros envolvidos na reabilitação de mamíferos marinhos devem rotineiramente selecionar animais para Brucella spp.
Generated by Foxit PDF Creator © Foxit Softwarehttp://www.foxitsoftware.com For evaluation only.

Microbiologia Clínica Veterinária VET 3225 ÁREA DE BACTERIOLOGIA 2011-2
UFRGS
Microbiologia Clínica Veterinária VET 3225 ÁREA DE BACTERIOLOGIA 2011-2
UFRGS
Brucella microti Prof. Marcos JP. Gomes INTRODUÇÃO Atualmente, além das oito espécies descritas, há mais outra espécie que foi adicionada às isoladas. A nova espécie chama-se B. microti, espécie isolada do camundongo do campo (Microtus arvalis). Este roedor do campo provoca grandes prejuízos às colheitas de cereais na Europa e reservatório para outros agentes como Leptospira interrogans, Francisella tularensis e Babesia microti. Em 1996, a população de camundongos do sul da Morávia (República Tcheca) foi objeto de um levantamento para melhor conhecer a dinâmica das populações. Entre 1999-2003, os pequenos roedores foram vítimas de uma epidemia com grande mortalidade. As observações microscópicas das lesões permitiram demonstrar cocobastonetes Gram negativos.
O exame bacteriológico efetuado em quatro animais capturados, em setembro de 2000, permitiu isolar em cultivo puro, oito amostras nas quais cinco pareciam assemelhar-se com amostras do gênero Ochrobactrum spp. Duas dessas amostras, as amostras CCM 4915 e CCM 4916, foram de fato submetidas a um estudo bacteriológico mais completo, sendo identificadas como cepas de Ochrobactrum intermedium. O O. intermedium é um germe do solo, raramente responsável por infecção em vertebrados, sendo pouco provável que o O. intermedium fosse a origem das epidemias observadas nos camundongos. As amostras CCM 4915 e CCM 4916 foram submetidas a um estudo taxonômico. O sequenciamento dos genes rrs, recA, omp2a e omp2b, assim como a hibridização ADN-ADN permitiu colocar as duas cepas dentro do gênero Brucella. A sequência dos genes rrs (1422 pb) e recA (897 pb) apresentaram 100% de semelhança com as seqüências das Brucella spp. As sequências dos genes omp2a (1104 pb) apresentaram 100 % de semelhança com a sequência da cepa 6516/98 da B. pinnipedialis, enquanto que a sequencia dos genes omp2b apresentou 99% de semelhança com a cepa B3196 da B. abortus e 99 % de similitude com a linhagem B. suis 1330. A técnica de AMOS (AMOS para AbortusMelitensisOvisSuis) permite detectar e diferenciar B. abortus, B. melitensis, B. ovis e B. suis, permitiu obter um fragmento de 2.000 pb ausente nas outras Brucella spp. A amplificação da região situada em direção ao gene bp26 conduz a obtenção de um amplicon de 1029 pb, característico das Brucella spp isoladas de mamíferos terrestres. A técnica de MLVA [multilocus VNTR (variable-number-tandem-repeats) analysis] efetuadas nas amostras CCM 4915 e CCM 4916 e em 424 amostras de Brucella spp revelaram que as cepas isoladas de roedores formavam um grupo distinto das outras espécies do gênero Brucella. As características fenotípicas permitem diferenciar as cepas CCM 4915 e CCM 4916 das outras espécies do gênero Brucella. Assim, em fevereiro de 2008, Scholz e colaboradores sugeriram o nome de B. microti para esta bactéria.
Generated by Foxit PDF Creator © Foxit Softwarehttp://www.foxitsoftware.com For evaluation only.

Microbiologia Clínica Veterinária VET 3225 ÁREA DE BACTERIOLOGIA 2011-2
UFRGS
Microbiologia Clínica Veterinária VET 3225 ÁREA DE BACTERIOLOGIA 2011-2
UFRGS
CARACTERÍSTICAS GERAIS As cepas de B. microti são cocobastonetes ou de bastonetes curtos, Gram negativos, com 0,5 a 0,8 µm de diametro por 0,6 a 1,4 µm de comprimento, e apresentam-se isoladas ou agrupadas em filamentos irregulares, aeróbias, imóveis, não esporuladas, oxidase positiva, catalase positiva, metabolismo oxidativo e aglutinação pelo soro anti-M diluído a 1/80. CARACTERÍSTICAS BIOQUÍMICAS Apresentam resposta positiva aos testes:
Redução dos nitratos (com formação de gas), Redução dos nitritos, Urease, VP, Fenilalanine desaminase, assimilação (galeria API 20 NE) de Adipato, L-arabinose, N-acetil-glucosamina, D-glicose, Maltose e D-manose, Oxidação (utilização do meio OF, Difco), Frutose, Glucose, Maltose e Xilose. Apresentam resposta negativa aos testes
ADH, LDC, ODC, Produção de H2S, Citrato de Simmons, ONPG, hidrólise do ADN, do amido, da caseina, da esculina, da gelatina, da lecitina, da tirosina, Assimilação (galeries API 20 NE) do caprato, do citrato, do gluconato, do malato, do D-manitol e do fenil-acetato. Apresentam resposta positiva no API ZYM
Fosfatase ácida, fosfatase alcalina, leucina arilamidase, valina arilamidase, cistina arilamidase, alpha-glicosidase e esterase lipase. Apresentam resposta negativa no API ZYM
Esterase, lipase, alpha-galactosidase, beta-galactosidase, beta-glucuronidase, beta-glucosidase, alfa-manosidase, alfa-fucosidase, N-acetil-beta-glucosaminidase. Outras características fenotípicas podem ser estudadas utilizando um sistema miniaturizado, denominado Micronaut (Merlin microbiological diagnostics company) o kit de identificação utilizado por Scholz e colaboradores não é relatado no catálogo da (Merlin microbiological diagnostics company). Os autores não deram nenhum detalhe sobre este kit, citando apenas uma publicação de Neubauer et al. 2000. A B. microti é cultivada na presença de tionina (diluida 1/25.000) e na presença de fucsina básica (diluida 1/50.000). Nenhum crescimento é evidenciado em caldo contendo 6,5% de NaCl. Os cultivos são lisados pelos fagos Tbilisi, F1 e F25 a 10.000 RTD (Routine Test Dilution, Dilution) e pelo fago Weybridge na diluição de rotina. O cultivo é facilmente obtido no agar nutritivo incubado na temperatura entre 25 e 42°C, em atmosfera aeróbia não enriquecida em CO2. Após 24 a 48 horas de incubação a 28-37°C, as colônias são lisas, transparentes, beges, ligeiramente côncavas de bordos regulares e com diâmetro de 1 a 2 mm. No AS, as colônias não são hemolíticas. Após 72h
Generated by Foxit PDF Creator © Foxit Softwarehttp://www.foxitsoftware.com For evaluation only.

Microbiologia Clínica Veterinária VET 3225 ÁREA DE BACTERIOLOGIA 2011-2
UFRGS
Microbiologia Clínica Veterinária VET 3225 ÁREA DE BACTERIOLOGIA 2011-2
UFRGS
de incubação a 37°C, as colônias possuem um tamanho compreendido entre 6 e 9 mm e de coloração amarronzada. A B. microti é sensível a gentamicina, tobramicina, ofloxacina e a associação de trimetoprima-sulfametoxazol. Resistência é observada para a colistina, piperacilina, da ceftazidima e tazobactam. PATOGENICIDADE A infecção dos camundongos provoca caquexia, edema de extremidades dos membros, artrites, orquites e adenites. Esses abscessos são, algumas vezes, sobre os membros e sua ruptura causa perfurações na pele. Na necropsia, há presença de abscessos peritoniais, esplenomegalia, hepatomegalia pouco pronunciada e hemorragias intestinais moderadas. A inoculação por via subcutânea de 2,5 x 107 microrganismos em camundongos de laboratório (Cd SPF da linhagem ICR) provoca, em 4 a 69 dias, morte de 50 % dos animais. Os camundongos mortos apresentam abscesso, uma ligeira esplenomegalia, um exsudato peritonial contendo bactérias Gram negativas observadas nas lesões. Patogenicidade para o homem ou para outras espécies animais não foi ainda documentada. DIAGNÓSTICO B. microti é facilmente isolada das lesões em agar nutritivo e no TSA. A identificação é delicada, pois a B. microti pode ser confundida com uma espécie do gênero Ochrobactrum, especialmente quando se utiliza a galeria API 20 NE conduzindo a uma identificação errônea de Ochrobactrum anthropi. De qualquer forma a ausência de motilidade permite distinguir facilmente B. microti de Ochrobactrum spp.
BIBLIOGRAFIA RECOMENDADA PARA O GÊNERO
Aguirre, NP.; Vanzini, VR.; Torion de Echaide, S.; Valentín, BS.; De Lucca, G.; Aufranc, C.; Canal, A. & Nielsen, K. Antibody dynamics in holstein friesian heifers vaccinated with Brucella abortus strain 19, using seven serological tests. J. Immunoassay Immunochemistry , v. 23, p. 471-478, 2002. Alton, GG.; Jones, LM.; Angus, RD.; Verger, JM. Tecniques for the Brucellosis Laboratory. 1º Edição Paris: Institut National de la Recherche Agronomique (INRA), 190 p. 1988. Anon, Joint FAO/WHO Expert Committee on Brucellosis. WHO Technical Report Series 740, WHO. Genebra, 1986. Anon, Manual of Standards for diagnostic Tests and Vaccines. 3ª Ed., Paris: OIE. 2000. Web address: http://www.oie.int/eng/normes/manual/A-00048htm. Azevedo, SS.; Vasconcellos, SA.; Alves, CJ. Brucelose canina por Brucella canis. Revista CRMV Ano X n.31, jan/fev/mar/abr p. 38-46, 2004 Blasco, JM. Brucella ovis. In: Animal Brucellosis, Nielsen K.; Duncan JR., eds. CRC Press, Boca Raton, Florida, USA, 351-378, 1990. Brew, SD.; Perrett, JA.; Stack, AP.; MacMillan, AP.; Staunton, NJ. Human exposure to Brucella recovered from a sea mammal. Vet. Rec., v. 144, p. 483, 1999.
Generated by Foxit PDF Creator © Foxit Softwarehttp://www.foxitsoftware.com For evaluation only.

Microbiologia Clínica Veterinária VET 3225 ÁREA DE BACTERIOLOGIA 2011-2
UFRGS
Microbiologia Clínica Veterinária VET 3225 ÁREA DE BACTERIOLOGIA 2011-2
UFRGS
Bricker, BJ. PCR as a diagnostic tool for brucellosis. Vet. Microbiol., v. 90, p. 435-446, 2002. Bricker, BJ.; Ewalt, DR.; Macmillan, AP.; Foster, G.; Brew, S. Molecular characterization of Brucella strains isolated from marine mammals. J. Clin. Microbiol. v. 38, p. 1258-1262, 2000. Bricker, BJ.; Ewalt, DR.; MacMillan, AP.; Foster, G.; Brew, S. Molecular characterization of Brucella strains isolated from marine mammals. J. Clin. Microbiol. v. 38, p. 1258-1262, 2000. Brown, GM.; Ranger, CR.; Kelley, DJ. Selective media for the isolation of Brucella ovis. Cornell Vet., v. 61, p. 265-280, 1971. Buddle, MB. Studies on Brucella ovis n. sp., a cause of genital disease of sheep in New Zealand and Australia. J. Hyg., v. 54, p. 351-364, 1956. Bulgin, MS.; Anderson, BC. Association of sexual experience with isolation of various bacteria in cases of ovine epididymitis. J. Am. Vet. Med. Assoc., v. 182, p. 372-374, 1983. Burgess, GW.; McDowell, JW. Escherichia coli epididymitis and seminal vesiculitis in a ram. Aust. Vet. J., v. 57, p. 479-480, 1981. Burgess, GW.; Norris, MJ. Evaluation of the cold complement fixation test for diagnosis of ovine brucellosis. Aust. Vet. J., v. 59, p. 23-25, 1982. Calle, PP.; Seagars, DJ.; McClave, C.; Senne, D.; House, C.; House, JA. Viral and bacterial serology of free-ranging Pacific walrus. J. Wildl. Dis., v. 38, p. 93-100, 2002. Cargill, C.; Lee, K.; Clarke, I. Use of an enzyme-liked immunoabsorbent assay in a bovine brucellosis eradication programme. Aust. Vet J., v. 62, p. 49-52, 1985. Carmichael, LE. Abortions in 200 Beagles. JAVMA., v. 149, n. 8, p. 1126, 1966. Carmichael, LE. Canine Brucellosis: An annoted review with selected cautionary comments. Theriogenol., v. 6, nº 2-3, p. 105-116, 1976. Carmichael, LE. ; Greene, GE. Canine brucellosis. In: Infectious diseases of the dog and cat. Greene, CE, (Ed), W.B. Saunders Company, p. 573-584, 1990. Carmichael, LE.; Bruner, DW. Characteristics of a new species of Brucella responsible for infectious canine abortions. Cornell Vet., v. 58, p. 579-592, 1968. Carmichael, LE.; Bruner, DW. Characteristics of a newly recognized species of Brucella responsible for infectious canine abortions. Cornell Veterinarian, v. 58, p. 579-592, 1968. Carmichael, LE.; Kenney, RM. Canine abortion caused by Brucella canis. JAVMA., v. 152, p. 605-616, 1968. Carmichael, LE.; Kenney, RM. Canine Brucellosis: The Clinical Disease, Pathogenesis, and Immune Response. JAVMA., v.56, n. 12, p. 726-1734, 1970. Carmichael, LE.; Zoha, SJ.; Flores-Castro, R. Problems in the serodiagnosis of canine brucellosis : dog responses to cell wall and internal antigens of Brucella canis. Develop. Biol. Standard, v. 56, p. 371-383, 1983. Clavareau, C.; Wellemans, V.; Walravens, K.; Tryland, M.; Verger, JM.; Grayon, M.; Cloeckaert, A.; Letesson, JJ.; Godfroid, J. Phenotypic and molecular characterization of a Brucella strain isolated from a minke whale (Balaenoptera acutorostrata). Microbiol., v. 144, p. 3267-3273, 1998.
Generated by Foxit PDF Creator © Foxit Softwarehttp://www.foxitsoftware.com For evaluation only.

Microbiologia Clínica Veterinária VET 3225 ÁREA DE BACTERIOLOGIA 2011-2
UFRGS
Microbiologia Clínica Veterinária VET 3225 ÁREA DE BACTERIOLOGIA 2011-2
UFRGS
Cloeckaert, A.; Grayon, M.; Grépinet, O. An IS711 element downstream of the bp26 gene is a specific marker of Brucella spp. isolated from marine mammals. Clin. Diagn. Lab. Immunol., v. 7, p. 835-839, 2000. Cloeckaert, A.; Grayon, M.; Grépinet, O.; Boumedine, KS. Classification of Brucella strains isolated from marine mammals by infrequent restriction site-PCR and development of specific PCR identification tests. Microbes Infect., v. 5, p. 593-602, 2003. Cloeckaert, A.; Verger, JM.; Grayon, M.; Grépinet, O. Restriction site polymorphism of the genes encoding the major 25 kDa and 36 kDa outer-membrane proteins of brucella. Microbiol., v. 141, p. 2111-2121, 1995. Cloeckaert, A.; Verger, JM.; Grayon, M.; Paquet, JY.; Garin-Bastuji, B.; Foster, G.; Godfroid, J. Classification of Brucella spp. isolated from marine mammals by DNA polymorphism at the omp2 locus. Microb. Infect., v. 3, p. 729-738, 2001. Corbel MJ.; Gill, KPW.; Thomas, EL. Methods for the identification of Brucella. Ministry of Agriculture, Fisheries and Food, UK, ADAS, RCV 22, 1978. Corbel, MJ.; Moriyón, I. International Committee on Systematic Bacteriology Subcommittee on the Taxonomy of Brucella. Minutes of the Meeting, 5 and 7 July 1994, Prague, Czech Republic. Int. J. Syst. Evol. Microbiol., v. 56, p. 1169-1170, 2006. Corbel, MJ.; Thomas, EL. Use of phage for the identification of Brucella canis and Brucella ovis cultures. Res. Vet. Sci., v. 38, p. 35-40, 1985. Currier, RW.; Raithel, WF.; Martin, RJ.; Potter, ME. Canine Brucellosis. JAVMA., v. 180, n. 2, p. 132-133, 1982. Da Costa, M.; Guillou, JP.; Garin-Bastuji, B.; Thiebaud, M.; Dubray, G. Specificity of six genes sequences for the detection of the genus Brucella by DNA amplification. J. Appl Bacteriol., v. 81, p. 267-275, 1996. De Long, WJ.; Waldhalm, DG. ; Hall, RF. Bacterial isolates associated with epididymitis in rams from Idaho and eastern Oregon flocks. Am. J. Vet. Res., v. 40, p. 101-102, 1979. De, BK; Stauffer, L; Koylass, MS; Sharp, SE; Gee, JE; Helsel, LO; Steigerwalt, AG; Vega, R; Clark, TA; Daneshvar, MI; Wilkins, PP.; Whatmore, AM. Novel Brucella Strain (BO1) Associated with a Prosthetic Breast Implant Infection. J. Clin. Microbiol., v. 46, n. 1, p. 43-49, 2008. Diaz, R.; Bosseray, N. Identification d’un composé antigénique spécifique de la phase rugueuse (R) des Brucella. Ann. Rech. Vet., v. 4, p. 283-292, 1973. Diaz, R.; Jones, LM.; Wilson, JB. Antigenic Relationship of the Gram negative Organism causing Canine Abortion to Smooth and Rough Brucellae. J. Bact., v. 95, p, 618-624, 1968. Ekdahl, MO.; Money, DF.; Martin, CA. Some aspects of epididymitis of rams in New Zealand. N. Z. Vet. J., v. 16, p. 81-82, 1968. Englehardt, CEM. Incidencia de Brucella canis em perros en el distrito de Chiclayo. Tesis Universidad Nacional Pedro Ruiz Gallo. Peru, 46p. 1974. Ewalt, D.; Payeur, J.; Martin, B.; Cummins, D.; Miller, W. Characteristics of a Brucella species from a bottlenose dolphin (Tursiops truncatus). J. Vet. Diagn. Investig., v. 6, p. 448-452, 1994.
Generated by Foxit PDF Creator © Foxit Softwarehttp://www.foxitsoftware.com For evaluation only.

Microbiologia Clínica Veterinária VET 3225 ÁREA DE BACTERIOLOGIA 2011-2
UFRGS
Microbiologia Clínica Veterinária VET 3225 ÁREA DE BACTERIOLOGIA 2011-2
UFRGS
Fernandes, JCT.; WALD, VB.; JOBIM, GB. Isolamento de Brucella canis do humor aquoso de um cão com lesões oculares. Arq. Fac. Vet. UFRGS., v. 4-5, p. 109-113, 1976/7. Flores-Castro, R.; Segura, R. A Serological and Bacteriological Survey of Canine Brucellosis in México. Cornell Vet., v. 66, p. 347-352, 1976. Forbes, LB.; Nielsen, O.; Measures, L.; Ewalt, DR. Brucellosis in ringed seals and harp seals from Canada. J. Wildl. Dis., v. 36, p. 595-598, 2000. Foster, G.; Jahans, KL.; Reid, RJ.; Ross, HM. Isolation of Brucella species from cetaceans, seals and an otter. Vet. Rec., v. 138, p. 583-586, 1996. Foster, G.; MacMillan, AP.; Godfroid, J.; Howie, F.; Ross, HM.; Cloeckaert, A.; Reid, RJ.; Brew, S.; Patterson, IAP. A review of Brucella sp. infection of sea mammals with particular emphasis on isolates from Scotland. Vet. Microbiol., v. 90, p. 563-580, 2002. Foster, G.; Osterman, BS.; Godfroid, J.; Jacques, I.; Cloeckaert, A. Brucella ceti sp. nov. and Brucella pinnipedialis sp. nov. for Brucella strains with cetaceans and seals as their preferred hosts. Int. J. Syst. Evol. Microbiol., v. 57, p. 2688-2693, 2007. Garcia-Carrillo, C. Animal and Human Brucellosis in the Americas. Paris: Office International des Epizooties, 299p. 1990. Gargani, G.; López-Merino, A. International Committee on Systematic Bacteriology Subcommittee on the taxonomy of Brucella. Correspondence Report (Interim Report), 1991-1993. Int. J. Syst. Evol. Microbiol., v. 56, p. 1167-1168, 2006. Gargani, G.; López-Merino, A. International Committee on Systematic Bacteriology Subcommittee on the taxonomy of Brucella. Correspondence Report (Interim Report), 1991-1993. Int. J. Syst. Evol. Microbiol., v. 56, p. 1167-1168, 2006. Garin, B.; Trap, D.; Gaumont, R. Assessment of the EDTA seoagglutination test for the diagnosis of bovine brucellosis. Vet Rec., v. 117, p. 444-445, 1985. Germano, PML.; Vasconcellos, SA.; Ishizuka, MM.; Passos, EC.; Erbolato, EB. Prevalência de infeção por Brucella canis em cães da cidade de Campinas-SP., Brasil. Rev.Fac. Med. Vet. Zootec., v. 24, n. 1, p. 27-34, 1987. Gleiser, CA.; Sheldom, WG.; Van Hoosier Jr, GL.; Hill, WA. Pathologic changes in dogs infected with a Brucella canis. Lab. Anim. Sci., v. 21, p. 540-545, 1971. Godfroid, J.; Cloeckaert, A.; Liautard, J.; Kohler, S.; Fretin, D.; Walravens, K.; Garin-Bastuji, B.; Letesson, J. From the discovery of the Malta fever's agent to the discovery of a marine mammal reservoir, brucellosis has continuously been a re-emerging zoonosis. Vet. Res., v. 36, p. 313-326, 2005. Godfroid, J.; Kasbohrer, A. Brucellosis in the European Union and Norway at the turn of the twenty-first century. Vet Microbiol., v. 90, p. 135-145, 2002. Godfroid, J.; Saegerman, C.; Welleman, V.; Walravens, K.; Letesson, JJ.; Tibor, A.; Mcmillan, A.; Spencer, S.; Sanna, M.; Bakker, D.; Pouillot, R.; Garin-Bastuiji, B. How to substantiate eradication of bovine brucellosis when aspecific serological reactions occur in the course of brucellosis testing. Vet. Microbiol., v. 90, p. 461-477, 2002. Godoy, AM.; Neves, J.; Peres, JN.; Barg, L. Sobre um caso de infecção humana por Brucella canis em laboratorio. Arq. Esc. Vet. UFMG., v. 31, n. 2, p. 141-145, 1979. Godoy, AM.; Peres, JN.; Barg, L. Isolamento de Brucella canis em Minas Gerais, Brasil. Arq. Esc. Vet. UFMG., v. 29, n. 1, p. 35-42, 1977.
Generated by Foxit PDF Creator © Foxit Softwarehttp://www.foxitsoftware.com For evaluation only.

Microbiologia Clínica Veterinária VET 3225 ÁREA DE BACTERIOLOGIA 2011-2
UFRGS
Microbiologia Clínica Veterinária VET 3225 ÁREA DE BACTERIOLOGIA 2011-2
UFRGS
Gonzalez, L.; Patterson, IA.; Reid, RJ.; Foster, G.; Barberan, M.; Blasco, JM.; Kennedy, S.; Howie, FE.; Godfroid, J.; MacMillan, AP.; Schock, A.; Buxton, D. Chronic meningoencephalitis associated with Brucella sp. infection in live-stranded striped dolphins (Stenella coeruleoalba). J. Comp. Pathol., v. 126, p. 147-152, 2002. Gyles, CL.; Thoen, CO. Pathogenesis of Bacterial Infections in Animals. 2.ª Ed., Ames: Iowa State University Press, 331p. 1993. Hagiwara, MK. Brucelose Canina Experimental: Estudos Bacteriológicos, Sorológicos e Anatomopatológicos. Arq. Bras. Med. Vet. Zootec., v. 36, n. 2, p. 141-156, 1984. Hanni, KD.; Mazet, JAK.; Gulland, FMD.; Estes, J.; Staedler, M.; Murray, MJ.; Miller, M.; Jessup, DA. Clinical pathology and assessment of pathogen exposure in Southern and Alaskan sea otters. J. Wildl. Dis., v. 39, p. 837-850, 2003. Harris, AM.; Horton, ML.; Letscher, RM.; McConnell, EE.; New, AL. Brucella canis-an occult disease in a research canine colony. Lab. Anim. Sci., v. 24, p. 796-799, 1974. Henderson, RA.; Horlein, BF.; Kramer, TT.; Meyer, ME. Discospondylitis in Three Dogs Infected with Brucella canis. JAVMA., v.65, n. 5, p. 451-455, 1974. Herr, S.; Te Brugge, LA. & Guiney, MCM. The value of microtitre serum agglutination test as a second screening test in bovine brucellosis. Onderstepoort J. Vet. Res., v. 49, p. 23-28, 1982. Herr, S.; Te Brugge, LA. Profiles of serological reactions following adult cow inoculation with standard dose Brucella abortus strain 19 vaccine. J. South Africa Vet. Assoc., v. 56, p. 93-96, 1985. Herr, S.; Williamson, CC.: Prigge, RE.; Van Wyk, A. The relationship between the microtitration serum agglutination test and complement fixation tests in bovine brucellosis serology. Onderstepoort J. Vet. Res., v. 53, p. 199-200, 1986. Hosgood, G.; Eilts, BE. Diskoespondylitis associated with Brucella canis infection in dogs: 14 cases (1980-1991). JAVMA., v. 201, nº 8, p.1253-1257, 1992. Hubálek, Z.; Scholz, HC.; Sedlácek, I.; Melzer, F.; Sanogo, YO.; Nesvadbová, J. Brucellosis of the common vole (Microtus arvalis). Vector Borne Zoonotic Dis., v. 7, p. 679-688, 2007. Huddleson, IF. The differentiation of the species of the genus Brucella. Michigan State College Agricultural Experimental Station Technical Bulletin. V. 100, p. 1-16, 1929. Hughes, ML. The natural history of certain fevers occurring in the Mediterrranean. Mediterranean Nature, v. 2, p. 325-327, 1893. ICSB, Subcommittee on the Taxonomy of Brucella: Report of the Meeting, 5 September 1986, Manchester, England. Int. J. Syst. Bacteriol., v. 38, p. 450-452, 1988. Jacques, I.; Grayon, M.; Verger, JM. Oxidative metabolic profiles of Brucella strains isolated from marine mammals: contribution to their species classification. FEMS Microbiol. Lett., v. 270, p. 245-249, 2007. Jahans, KL.; Foster, G.; Broughton, ES. The characterisation of Brucella strains isolated from marine mammals. Vet. Microbiol., v. 57, p. 373-382, 1997. Jepson, PD.; Brew, S.; MacMillan, AP.; Baker, JR.; Barnett, J.; Kirkwood, JK.; Kuiken, T.; Robinson, IR.; Simpson, VR. Antibodies to brucella in marine mammals around the coast of England and Wales. Vet. Rec., v. 141, p. 513-515, 1997.
Generated by Foxit PDF Creator © Foxit Softwarehttp://www.foxitsoftware.com For evaluation only.

Microbiologia Clínica Veterinária VET 3225 ÁREA DE BACTERIOLOGIA 2011-2
UFRGS
Microbiologia Clínica Veterinária VET 3225 ÁREA DE BACTERIOLOGIA 2011-2
UFRGS
Johnson, CA. Clinical signs and diagnosis of Brucella canis infection. Comp. Cont. Educ. Pract. Vet., v. 14, p. 763-772, 1992. Jones, LM.; Zanardi, M.; Leong, D.; Wilson, JB. Taxonomic position in the Genus Brucella of the causative Agent of Canine Abortion. J. Bact., v. 95, p. 625-630, 1968. Larsson, MHMA.; Costa, E.O. Isolation of Brucella canis. Int. J. Zoon., v. 7, p. 125-130, 1980. Larsson, MHMA.; Larsson, CE.; Costa, EO.; Guerra, JL.; Hagiwara, MK. Brucelose canina Experimental: Estudos Bacteriológicos, Sorológico e Anatomopatológico. Arq. Bras. Med. Vet. Zoot., v. 36, n. 2, p. 141-156, 1984. Livingstone, CW.; Hardy, WT. Isolation of Actinobacillus seminis from ovine epididymitis. Am. J. Vet. Res., v. 25, p. 660-663, 1964. Magalhães Neto, A.; Cruz, FW.; Santos, AF.; Gil-Turnes, C.; Aleixo, JAG.; Martins, LFS.; Brodt, CS.; Gurvitz, R. Prevalência de humanos e caninos reatores à Brucella rugosa no Município de Pelotas, RS. In: XIº Congresso Estadual de Medicina Veterinária, Gramado, RS. Anais. SOVERGS, 1992, 150 p. p.92, 1992. Mantur, BG.; Amarnath, SK.; Shinde, RS. Review of clinical and laboratory features of human brucellosis. Indian J. Med. Microbiol., v. 25, p. 188-202, 2007. Maratea, J.; Ewalt, DR.; Frasca Jr, S.; Dunn, JL.; De Guise, S.; Szkudlarek, L.; St. Aubin, DJ.; French, RA.; De Guise, S. Evidence of Brucella sp. infection in marine mammals stranded along the coast of Southern. N. Engl. J. Zoo Wildl. Med., v. 34, p. 256-261. 2003. Marin, CM.; Alabart, JL.; Blasco, JM. Effect of antibiotics contained in two Brucella selective media on growth of B. abortus, B. melitensis and B. ovis. J. Clin. Microbiol., v. 34, p. 426-428, 1996. Marin, CM.; Barberan, M.; Jimenez de Bagues, MP.; Blasco, JM. Comparison of subcutaneous and conjunctival routes of Rev. 1 vaccination for the prophylaxis of Brucella ovis infection in rams. Res. Vet. Sci., v. 48, p. 209-215, 1990. Marin, CM.; Jimenez de Bagues, MP.; Blasco, JM.; Gamazo, C.; Moriyon, I.; Diaz, R. Comparison of three serological tests for Brucella ovis infection of rams using different antigenic extracts. Vet. Rec., v. 125, p. 504-508, 1989. Mateudeantonio, EM.; Martin, M. In vivo efficacy of several antimicrobial combinations against Brucella canis and Brucella melitensis strains isolated from dogs. Vet. Microbiol., v. 45, p. 1-10, 1995. McDonald, WL.; Jamaludin, R.; Mackereth, G.; Hansen, M.; Humphrey, S.; Short, P.; Taylor, T.; Swingler, J.; Dawson, CE.; Whatmore, AM.; Stubberfield, E.; Perrett, LL.; Simmons, G. Characterization of a Brucella sp. strain as a marine-mammal type despite isolation from a patient with spinal osteomyelitis in New Zealand. J. Clin. Microbiol., v. 44, p. 4363-4370, 2006. Michaux-Charachon, S.; Bourg, G.; Jumas-Bilak, E.; Guigue-Talet, P.; Allardet-Servent, A.; O’Callahan, D.; Ramuz, M. Genome structure and phylogeny in genus Brucella. J. Bacteriol., v. 179, p. 3244-3249, 1997. Miller, W.; Adams, L.; Ficht, T.; Cheville, N.; Payeur, J.; Harley, D.; House, C.; Ridgway, S. Brucella-induced abortions and infection in bottlenose dolphins (Tursiops truncatus). J. Zoo Wildl. Med., v. 30, p. 100-110, 1999.
Generated by Foxit PDF Creator © Foxit Softwarehttp://www.foxitsoftware.com For evaluation only.

Microbiologia Clínica Veterinária VET 3225 ÁREA DE BACTERIOLOGIA 2011-2
UFRGS
Microbiologia Clínica Veterinária VET 3225 ÁREA DE BACTERIOLOGIA 2011-2
UFRGS
Moore, JA.; Bennett, M. A Previously Undescribed Organism Associated with Canine Abortion. Vet Rec., v. 80, p. 604-605, 1967. Moore, JA.; Kakuk, TJ. Male dogs Naturally Infected with Brucella canis. JAVMA., v. 155, n. 8, p. 1352-1358, 1969. Morrow, DA. Current Therapy in Theriogenology 2. Philadelphia: W.B. Sauders Company, 1143p. 1986. Myers, DM.; Jones, LM.; Varela-Diaz, V. Studies of antigens for complement fixation and gel diffusion tests in the diagnosis of infections caused by Brucella ovis and other Brucella. Appl. Microbiol., v. 23, p. 894-902, 1972. Myers, DM.; Varela-diaz, V. Sorological and Bacteriological detection of Brucella canis infection of stray dogs in Moreno, Argentina. Cornell Vet., v. 70, p. 258-265, 1980. Myers, DM.; Varela-Diaz, VM.; Coltorti, E. A Comparative Sensitivity of Tube Agglutination Tests for the Detection of Brucella canis Antibodies in Experimental Infected Dogs. Appl. Microbiol., v. 28, n. 1, p. 1-4, 1974. Nicoletti, P. Further studies on the use of antibiotics in canine brucellosis. Comp. Cont. Educ. Pract. Vet., v. 13, p. 944-947, 1991. Nicoletti, P.; Chase, A. The use of antibiotics to control canine brucellosis. Comp. Cont. Educ. Pract. Vet., v. 9, p. 1063-1066, 1987. Nielsen, K. Diagnosis of brucellosis by serology. Vet. Microbiol., v. 90, p. 447-459, 2002. Nielsen, K. The serological response of cattle immunized with Yersinia enterocolitica O:9 or O;16 to Yersinia and Brucella abortus antigen in enzyme immunoassay. Vet Immunol. Immunopathol., v. 24, p. 373-382, 1990. Nielsen, K.; Duncan, JR. Animal Brucellosis. Boca Raton: CRC Press Inc., 453p. 2000. Nielsen, K.; Gall, D. Fluorescence polarization assay for the diagnosis of brucellosis: A review J. Immunoassay and Immunochem., v. 22, p. 183-201, 2001. Nielsen, K.; Kelly, L.; Gall, D.; Nicoletti, P.; Kelly, W. Improved competitive enzyme immunoassay for the diagnosis of bovine brucellosis. Vet. Immunol. Immunopathol., v. 46, p. 285-291, 1995. Nielsen, O.; Nielsen, K.; Braun, R.; Kelly, L. A comparison of four serologic assays in screening for brucella exposure in Hawaiian monk seals. J. Wildl. Dis., v. 41, p. 126-133, 2005. Nielsen, O.; Stewart, REA.; Nielsen, K.; Measures, L.; Duignan, P. Serologic survey of Brucella spp. antibodies in some marine mammals of North America. J. Wildl. Dis., v. 37, p. 89-100, 2001. Ohishi, K.; Takishita, K.; Kawato, M.; Zenitani, R.; Bando, T.; Fujise, Y.; Goto, Y.; Yamamoto, S.; Maruyama, T. Chimeric structure of omp2 of Brucella from Pacific common minke whales (Balaenoptera acutorostrata). Microbiol. Immunol., v. 49, p. 789-793, 2005. Ohishi, K.; Zenitani, R.; Bando, T.; Goto, Y.; Uchida, K.; Maruyama, T.; Yamamoto, S.; Miyazaki, N.; Fujise, Y. Pathological and serological evidence of brucella-infection in baleen whales (Mysticeti) in the western North Pacific. Comp. Immunol. Microbiol. Infect. Dis., v. 26, p. 125-136, 2003.
Generated by Foxit PDF Creator © Foxit Softwarehttp://www.foxitsoftware.com For evaluation only.

Microbiologia Clínica Veterinária VET 3225 ÁREA DE BACTERIOLOGIA 2011-2
UFRGS
Microbiologia Clínica Veterinária VET 3225 ÁREA DE BACTERIOLOGIA 2011-2
UFRGS
Osterman, B.; Moriyón, I. International Committee on Systematics of Prokaryotes Subcommittee on the taxonomy of Brucella. Minutes of the meeting, 17 September 2003, Pamplona, Spain. Int. J. Syst. Evol. Microbiol., v. 56, p. 1173-1175, 2006. Paulin, LM.; Ferreira Neto, JS. O Combate a Brucelose Bovina: Situacao Brasileira. Jaboticabal: Funep, 154p. 2003. Pereira Filho, MP.; Silva, JAH.; Rocha, JVN. Estudo da incidência da Brucelose canina na zona metropolitana de Salvador. In: Congresso Brasileiro de Medicina Veterinária,16 Salvador, 1978. Anais. Salvador, Doc. Bras. Med. Vet.; Soc. Med. Vet. Bahia, 1978 p.32. Perrett, LL.; Brew, SD.; Stack, JA.; MacMillan, AP.; Bashiruddin, JB. Experimental assessment of the pathogenicity of Brucella strains from marine mammals for pregnant sheep. Sm. Rum. Res., v. 51, p. 221-228, 2004. Poester, FP.; Gonçalves, VSP.; Lage, AP. Brucellosis in Brazil. Vet Microbiol., v. 90, p. 55-62, 2002. Quinn, J.; Carter, ME.; Markey, B.; Carter, GR. Clinical Veterinary Microbiology. Edimburgh: MOSBY Harcourt Publishers Limited, 648p. 1999. Quinn, PJ.; Markey, BK.; Carter, ME.; Donnelly, WJ.; Leonard, FC. Microbiologia Veterinária e Doenças Infecciosas. Porto Alegre: Artmed, 512p. 2005. Radostits, OM.; Gay, CC.; Blood, DC.; Hinchcliff, KW. Veterinary Medicine: A Textebook of the Diseases of Cattle Sheep, Pigs, Goats and Horses. 9ª Ed., Philadelphia: W.B.Sauders, 1877p. 2000. Radostits, OM.; Leslie, KE.; Fetrow, J. Herd Health: Food Animal Producton Medicine. 2ª Ed., Philadelphia: W.B. Sauders Company, 631p. 1994. Ramacciotti, F. Primer aislamiento de "Brucella canis" en humano por hemocultivo efectuado en la Republica Argentina. Rev. Med. Vet. Arg., v. 61, p. 49-54, 1980. Rebhun, WC.; Guard, C.; Richards, CM. Diseases of Dairy Cattle. Baltimore: Williams & Wilkins, 530p. 1995. Retamal, P.; Blank, O.; Abalos, P.; Torres, D. Detection of anti-brucella antibodies in pinnipeds from the Antarctic territory. Vet. Rec., v. 146, p. 166-167, 2000. Rhyan, JC.; Gidlewski, T.; Ewalt, DR.; Hennager, SG.; Lambourne, DM.; Olsen, SC. Seroconversion and abortion in cattle experimentally infected with Brucella sp. isolated from a Pacific harbor seal (Phoca vitulina richardsi). J. Vet. Diagn. Investig., v. 13, p. 379-382, 2001. Riecke, JA.; Rhoades, HE. Brucella canis Isolated from the Eye of a Dog. JAVMA., v. 166, n. 6, p. 583-584. 1975. Riezu-Boj, JI.; Moriyon, I.; Blasco, JM.; Marin, CM.; Diaz, R. Comparison of lipopolysaccharide and outer membrane protein-lipopolysaccharide extracts in an enzyme-linked immunosorbent assay for the diagnosis of Brucella ovis infection. J. Clin. Microbiol., v. 23, p. 938-942, 1986. Ris DR.; Hamel, KL.; Long, DL. Comparison of an enzyme-linked immunospecific assay (ELISA) with the cold complement fixation test for the serodiagnosis of Brucella ovis infection. N. Z. Vet. J., v. 32, p. 18-20, 1984. Rodolakis, A.; Bernard, K. Isolement de Chlamydia des organes génitaux de béliers atteints d’épididymite. Bull. Acad. Vet. Fr., v. 50, p. 65-70, 1977.
Generated by Foxit PDF Creator © Foxit Softwarehttp://www.foxitsoftware.com For evaluation only.

Microbiologia Clínica Veterinária VET 3225 ÁREA DE BACTERIOLOGIA 2011-2
UFRGS
Microbiologia Clínica Veterinária VET 3225 ÁREA DE BACTERIOLOGIA 2011-2
UFRGS
Ross, HM.; Foster, G.; Reid, RJ.; Jahans, KL.; MacMillan, AP. Brucella species infection in sea mammals. Vet. Rec., v. 134, p. 359, 1994. Rylatt, DB.; Wyat, DM.; Bundesen, PG. A competitive enzyme immunoassay for the detection of bovine antibodies to Brucella abortus using monoclonal antibodies. Vet Immunol. Immunopathol., v. 8, p. 261-271, 1985. Saegerman, C.; VO, TK.; De Waele, L.; Gilson, D.; Bastian, A.; Dubray, G.; Flanagan, P.; Limet, JN; Letesson, JJ.; Godfroid, J. Diagnosis of bovine brucellosis by skin test and evaluation of its performance. Vet. Rec., v. 145, p. 2140218, 1999. Sandoval, LA.; Ribeiro, LOC.; Amaral, LBS.; Feitosa, MH.; Bazan, JM. Incidência da brucelose canina na cidade de São Paulo. Biol., v. 42, p. 128-132, 1976. Santos, JM.; Verstreate, DR.; Perera, VY.; Winter, AJ. Outer membrane proteins from rough strains of four Brucella species. Infect. Immun., v. 46, p. 188-194, 1984. Schlabritz-Loutsevitch, NE; Whatmore, AM; Quance, CR; Koylass, MS; Cummins, LB; Dick Jr, EJD; Snider, CL; Cappelli, D; Ebersole, JL; Nathanielsz, PW.; Gene B. Hubbard, GB. A novel Brucella isolate in association with two cases of stillbirth in non-human primates – first report. J. Med. Primatol., v. 38, p. 70–73, 2009. Schlemper, SRM.; VAZ, AK. Inquérito sorológico para Brucelose canina por Brucella canis na Região do Planalto Catarinense, Brasil. Rev. Bras. Med. Vet., v.12, p. 8-12, 1990. Schoeb, TR.; Morton, R. Scrotal and Testicular Changes in Canine Brucellosis: A case Report. JAVMA., v. 172, nº 5, p. 598-600, 1978. Scholz, HC.; Hubalek, Z.; Sedláček, I.; Vergnaud, G.; Tomaso, H.; Al Dahouk, S.; Melzer, F.; Kämpfer, P.; Neubauer, H.; Cloeckaert, A.; Maquart, M.; Zygmunt, MS.; Whatmore, AM.; Falsen, E.; Bahn, P.; Göllner, C.; Pfeffer, M.; Huber, B.; Busse, HJ.; Nöckler, K. Brucella microti sp. nov., isolated from the common vole Microtus arvalis. Int. J. Syst. Evol. Microbiol., v. 58, p. 375-382, 2008. Seagusa, J.; Ueda, K.; Goto, Y.; Fujiwara, K. Ocular lesions in experimental Canine Brucellosis. Jap. J. vet. Sci., v. 39, p. 181-185, 1977. Searson, JE. Sensitivity and specificity of two microtitre complement fixation tests for the diagnosis of Brucella ovis infection in rams. Aust. Vet. J., v. 58, p. 5-7, 1982. Serikawa, T.; Muraguchi, T.; Nakao, N. Significance of urine culture for detecting infection with Brucella canis in dogs. Jap. J. vet. Sci., v. 40, p.353-355, 1978. Sohn, A.; Probert, W.; Glaser, C.; Gupta, N.; Bollen, A.; Wong, J.; Grace, E.; McDonald, W. Human neurobrucellosis with intracerebral granuloma caused by a marine mammal Brucella spp. Emerg. Infect. Dis., v. 9, p. 485-488, 2003. Spencer, TL.; Burgess, GW. Enzyme-linked immunosorbent assay for Brucella ovis specific antibody in ram sera. Res. Vet. Sci., v. 36, p. 194-198, 1984. Stackebrandt, E.; Frederiksen, W.; Garrity, G.M. Grimont, PAD.; Kämpfer, P.; Maiden, MCJ.; Nesme, X.; Rossello-Mora, R.; Swings, J.; Trüper, HG.; Vauterin, L.; Ward, AC.; Whitman, WB. Report of the ad hoc committee for the re-evaluation of the species definition in bacteriology. Int. J. Syst. Evol. Microbiol., v. 52, p. 1043-1047, 2002. Stoenner, HG.; Lackman, DB. A new species of Brucella isolated from the desert wood rat, Neotoma lepida Thomas. Am. J.Vet. Res., v. 18, p. 947-951, 1957. Sutherland, SS. An enzyme-liked immunosorbent assay for the detection of Brucella abortus in cattle usinag monoclonal antibodies. Aust. Vet J., v. 62, p. 264-268, 1985
Generated by Foxit PDF Creator © Foxit Softwarehttp://www.foxitsoftware.com For evaluation only.

Microbiologia Clínica Veterinária VET 3225 ÁREA DE BACTERIOLOGIA 2011-2
UFRGS
Microbiologia Clínica Veterinária VET 3225 ÁREA DE BACTERIOLOGIA 2011-2
UFRGS
Sutherland, SS. Evaluation of the enzyme-linked immunosorbent assay in the detection pf cattle infected with Brucella abortus. Vet Microbiol., v. 10, p. 23-32, 1984. Swenson, RM.; Carmichael, LE.; Cundy, KR. Human infection with Brucella canis. Ann. Int. Med., v. 76, p. 435-438, 1972. Terakado, TE.; Ueda, NH.; Sugawara, H.; Isayama, Y. Drug susceptibility of Brucella canis isolated from dogs. Jap. J. Vet. Sci., v. 40, p. 291-295, 1978. Thoen, CO.; Steele, JH. Mycobacterium bovis Infection in Animals and Humans Ed. Thoen CO.; Steele, JH. Ames: Iowa State University Press, 355p, 1995. Timoney, JF.; Gillespie, JH.; Scott, FW.; Barlough, JE. Hagan and Bruner’s Microbiology and Infectious Disease of Domestic Animals. 8ª Ed. Ithaca: Cornell University Press, 951p. 1992. Tryland, M.; Derocher, AE.; Wiig, Y.; Godfroid, J. Brucella sp. antibodies in polar bears from Svalbard and the Barents Sea. J. Wildl. Dis., v. 37, p. 523-531, 2001. Tryland, M.; Kleivane, L.; Alfredsson, A.; Kjeld, M.; Arnason, A.; Stuen, S.; Godfroid, J. Evidence of brucella infection in marine mammals in the North Atlantic Ocean. Vet. Rec., v. 144, p. 588-592, 1999. Tryland, M.; Sørensen, K.; Godfroid, J. Prevalence of Brucella pinnipediae in healthy hooded seals (Cystophora cristata) from the North Atlantic Ocean and ringed seals (Phoca hispida) from Svalbard. Vet. Microbiol., v. 105, p. 103-111, 2005. Ueda, K.; Magaribuchi, T.; Saegusa, J.; Urano, K.; Itoh, K.; Kiuchi, Y.; Fujiwara, K. Spontaneous Brucella canis infection in beagles: Bacteriological and Serological Studies. Jap. J. vet. Sci., v. 36, p. 381-389, 1974. Van Bressem, M.; Van Waerebeek, K.; Raga, J.; Godfroid, J.; Brew, S.; MacMillan, A. Serological evidence of Brucella species infection in odontocetes from the south Pacific and the Mediterranean. Vet. Rec., p. 148, p. 657-661, 2001. Vargas, AC.; Lazzari, A.; Dutra, V.; Poester, FP. Brucelose canina: Relato de caso. Ciência Rural, v. 26, nº2, p. 305-308, 1996. Verger, JM.; Gâté, M.; Piechaud, M.; Chatelain, R.; Amisse, J.; Blancou, J. Isolement de Brucella suis biotype 5 à Madagascar, chez une chienne. Validity du nom d’espece Brucella canis . Ann. Microbiol., v. 126 A, p.57-74, 1975. Verger, JM.; Grimont, F.; Grimont, PAD. ; Grayon, M. Brucella, a monospecific genus as shown by deoxyribonucleic acid hybridization. Int. J. Syst. Bacteriol., v. 35, p. 292-295, 1985. Wald, VB.; Fernandes, JCT. Sorologia da Brucelose canina no Município de Porto Alegre, RS. Arq. Fac. Vet. UFRGS., v. 4-5, p. 92-95, 1976-7. Wayne, LG.; Brenner, DJ.; Colwell, RR.; Grimont, PAD.; Kandler, O.; Krichevsky, MI.; Moore, LH.; Moore, WEC.; Murray, RGE.; Stackebrandt, E.; Starr, MP.; Trüper, HG. Report of the ad hoc committee on reconciliation of approaches to bacterial systematics. Int. J. Syst. Bacteriol., v. 37, p. 463-464, 1987. Webb, RF. Clinical findings and pathological changes in Histophilus ovis infections of sheep. Res. Vet. Sci., v. 35, p. 30-34, 1983. Webb, RF.; Quinn, CA.; Cockram, FA.; Husband, AJ. Evaluation of procedures for the diagnosis of Brucella ovis infection in rams. Aust. Vet. J., v. 56, p. 172-175, 1980.
Generated by Foxit PDF Creator © Foxit Softwarehttp://www.foxitsoftware.com For evaluation only.

Microbiologia Clínica Veterinária VET 3225 ÁREA DE BACTERIOLOGIA 2011-2
UFRGS
Microbiologia Clínica Veterinária VET 3225 ÁREA DE BACTERIOLOGIA 2011-2
UFRGS
Williamson, P.; Nairn, ME. Lesions caused by Corynebacterium pseudotuberculosis in the scrotum of rams. Aust. Vet. J., v. 56, p. 496-498, 1980. Worthington, RW.; Stevenson, BJ.; de Lisle, GW. Serology and semen culture for the diagnosis of Brucella ovis infection in chronically infected rams. N.Z. Vet. J., v. 33, p. 84-86, 1985. Worthington, RW.; Weddell, W.; Penrose, ME. A comparison of three serological tests for the diagnosis of B. ovis infection in rams. N.Z. Vet. J., v. 32, p. 58-60,1984. Wright, AE.; Smith, F. On the application of the serum test to differential diagnosis of thyphoid and Malta fever. Lancet, v. 1, p. 656-659, 1897. Yamauchi, C.; Suzuki, T.; Nomura, T. Canine brucellosis in a beagle breeding colony. Jap. vet. Sci., v. 36, p. 175-182, 1974.
Generated by Foxit PDF Creator © Foxit Softwarehttp://www.foxitsoftware.com For evaluation only.