c magnolia - magnolia spp. - plant pathology · climates and may be both evergreen and deciduous....
TRANSCRIPT

© Sarah A. White 2012
CHAPTER 9
Magnolia - Magnolia spp.
Gary W. Knox
University of Florida
William E. Klingeman
University of Tennessee
Mathews L. Paret
University of Florida
Amy F. Fulcher
University of Tennessee

MAGNOLIA BASICS
1. Overview
2. Production
3. Host Plant Resistance
SECTION 1
Introduction The genus* Magnolia encompasses a group of about 240 species of trees and large
shrubs. They are native to southeastern Asia, eastern North America, Central America, the
Caribbean, and parts of South America. Magnolias grow in both temperate and tropical
climates and may be both evergreen and deciduous. These plants characteristically have
showy, fragrant flowers that are white, pink, red, purple, or yellow. The flowers are fol-
lowed by showy red or pink fruits displaying red, orange, or pink seeds, each of which
hangs from the fruit by a thread-like strand.
Magnolia is one of two genera currently recognized in the Magnoliaceae family.
Molecular systematics and re-examination of morphological characteristics resulted in all
genera other than Liriodendron (i.e., Michelia and Manglietia) being combined into Magno-
lia (Figlar and Nooteboom, 2004). The Magnoliaceae family has an ancient taxonomic ori-
gin with fossil remains dating between 36 and 58 million years ago. Surviving magnolia
species represent some of the more primitive flowering plants. Magnolia flowers do not
have true petals and sepals but are composed of petal-like tepals. Flowers do not produce
true nectar but attract pollinating beetles with fragrant, sugary secretions. Beetles of the
Nitidulidae family are the primary pollinators of magnolia flowers, in part because magno-
lias evolved long before bees and other flying pollinators.
Magnolia species are found in a wide range of soils and climates, but they are most
often associated with moist soils in mixed deciduous-coniferous or evergreen woodlands.
In cultivation, most magnolias grow best in moist, well-drained, slightly acidic soils in loca-
tions receiving full sun to light shade. Plants grown in warm or dry climates benefit from
planting locations shaded from afternoon sun, although magnolias can tolerate full sun and
heavier soils.
Magnolias have been harvested for timber and medicinal uses but are usually culti-
vated for their beautiful flowers, fruits, foliage, and plant forms. Asian cultures have long
cultivated several Magnolia species for their flowers. First introduced to Europe and Amer-
ica in 1780, various Asian species were recognized for their free flowering character and
182
© Jeffrey Derr 2012
This chapter is adapted from a review article submitted for publication in the Journal of Environmental Horti-culture in 2012, entitled “Management of Pests, Plant Diseases and Abiotic Disorders of Magnolia Species in the Southeastern U.S.: A Review” by Gary W. Knox, William E. Klingeman, Mathews L. Paret and Amy F. Fulcher.
*For definitions of bold terms, see the digital version of this book at the SNIPM website http://wiki.bugwood.org/SNIPM.

became immensely popular. Intensive breeding programs began hybridizing magnolias to
develop more floriferous and hardy forms with a wider range of flower colors. The hy-
brids and selections produced by these breeding programs, still ongoing, have resulted in
many superior ornamental trees. Many people consider magnolias to be some of the most
popular flowering trees in the United States. Most widely grown, deciduous magnolias
were selected from the early spring-blooming M. denudata, M. liliiflora, Magnolia stellata,
and their hybrids (Callaway, 1994).
Other magnolias are grown for their value as shade trees. The American tree, south-
ern magnolia, M. grandiflora, was introduced to Europe in 1731. This tree quickly became
popular because of its glossy evergreen foliage, large beautiful flowers, and elegant form.
Magnolia grandiflora was also found to be widely adaptable to different climates, soils,
and exposures. Thus, it was the first magnolia to be planted widely as a street or shade tree
and is now grown nearly worldwide wherever suitable climate and soils exist.
Several Magnolia species are widely produced by commercial nurseries in the
southern United States (Table 9.1), and total sales of these flowering deciduous species in
2007 were $18,287,000 (USDA, 2009). Magnolia grandiflora is the most widely grown
magnolia because its cultivars are heavily favored for landscape use. Other species pro-
duced by many growers are M. virginiana and M. stellata cultivars and hybrids. Small
numbers of nurseries produce M. × soulangeana and cultivars derived from complex hy-
brids of these and other deciduous species (i.e., Magnolia 'Jon Jon') and Magnolia figo (=
Michelia figo). New yellow-flowered hybrids derived from M. acuminata are becoming
popular but are not commonly grown. Less widely known natives such as M. ashei, M.
macrophylla, and M. tripetala are rarely produced. Of the more recently introduced Asian
species such as M. laevifolia (=Magnolia dianica=Michelia yunnanensis), M. insignis
(=Manglietia insignis), and M. maudiae (=Michelia maudiae), only the latter is currently
produced. Table 9.2 lists the characteristics of magnolia species and cultivars widely
grown by nurseries in the southeastern United States.
Production
Magnolias may be produced in the field, in containers, by modified field produc-
tion, or by any combination of these methods in the southeastern United States (Knox,
2012). Commercial production of magnolias may start with seeds, particularly for species
that root poorly, for production of native species, or as rootstock for grafting or budding.
The nursery and landscape industries currently emphasize production and use of cultivars;
thus, most magnolias are propagated by cuttings that are rooted to produce a liner (young
transplant). In the southern United States, budding and grafting are only used with culti-
vars of certain genera that are difficult to root, such as M. acuminata hybrids.
Host Plant Resistance
Most magnolias are considered to be "trouble-free" with few pests and diseases (Dirr,
1998). Phytochemicals in Magnolia species show a wide range of biological activity. Phe-
nolics and neolignans are among the magnolia compounds with antimicrobial, nemati-
cidal, and insecticidal properties (Kamikado et al., 1975; Li et al., 2009; Nitao et al., 1991).
Other magnolia compounds act as attractants of natural enemies (Azuma et al., 1997).
Studies comparing host plant resistance among Magnolia species are rare, but Held
(2004) found M. grandiflora, M. ×soulangeana, and M. virginiana were resistant to Japa-
nese beetle (Popillia japonica), as are Magnolia species in general (Klingeman, personal
communication). Studies with the major pest, Neolecanium cornuparvum (Magnolia
scale), found M. acuminata and M. stellata 'Merrill' and 'Royal Star' were highly suscepti-
ble while M. grandiflora 'Little Gem' and M. virginiana were resistant (Vanek and Potter,
2009).
Many other studies included one or two Magnolia species in host plant resistance re-
search but collectively also provide evidence for the relative pest resistance of this genus.
Santamour and Reidel (1993) inoculated seedlings of 23 tree taxa with four common root
knot nematode species (Meloidogyne arenaria races 1 and 2, Meloidogyne hapla, Meloi-
dogyne incognita, and Meloidogyne javanica) and found M. grandiflora was resistant to all
183

four nematode species. In greenhouse tests with 35 taxa, Bernard and Witte (1987) found
M. ×soulangeana 'Alexandrina' would not host Meloidogyne hapla, the primary root knot
nematode of woody ornamentals in Tennessee.
Similarly, M. ×soulangeana exhibited host plant resistance to Lymantria dispar
(gypsy moth) by losing less than 4% of its foliage in a study of gypsy moth feeding and
defoliation of 21 shade and flowering trees (Peterson and Smitley, 1991). In choice test
feeding preference studies with eight woody taxa, M. grandiflora was not consumed by
adult Phyllophaga ephilida (June bug; Coleoptera: Scarabaeidae; Diagne et al., 2006).
Table 9.3 lists insecticides and IRAC codes used within a pesticide rotation plan to
achieve selected insect control against key pests of ornamental plants, including magno-
lias.
184

ABIOTIC & WILDLIFE STRESSORS
1. Winter Burn
2. Cold Damage
3. Flooding Damage
4. Sapsucker Damage
SECTION 2
Abiotic & Wildlife Stressors Winter Burn: Winter burn or scorch (see Abiotic & Wildlife Stressors section
introductory image) is a common problem of evergreen and semi-evergreen magnolias
grown in the upper South. Winter burn is characterized by desiccated leaf margins,
often most extreme on the side exposed to wind or greater sun exposure (Relf and
Appleton, 2009). Magnolias that are evergreen or semi-evergreen continue losing
moisture through their leaves year-round. Magnolias are especially vulnerable to win-
ter burn on warm, sunny days when the ground is frozen and plants cannot take up wa-
ter to replace that which is lost through transpiration. Container liners are more likely
to suffer from scorch than bareroot liners.
In areas of the upper South where winter burn is a problem, a few simple prac-
tices should help reduce the incidence and/or severity of winter burn. Broad-leaved
evergreens should be lined out earlier in the fall than their deciduous counterparts
(Adkins et al., 2010). This allows roots to become more established prior to the onset
of freezing conditions and drying winds. Allowing the root system to establish before
harsh weather will help ensure that the plant is optimally hydrated until the soils
freeze. As with all container liners, evergreen and semi-evergreen magnolias should
be well watered prior to lining out in the field. Anti-desiccants have not been found to
consistently benefit plants during transplanting or lining out (Relf and Appleton,
2009). Growers should plan for regular irrigation in the fall to increase the availability
of water to the newly planted liners.
Cold Damage: Magnolia stellata and M. ×soulangeana are early bloomers,
and in the upper South, flower buds are commonly killed by freezing temperatures.
These magnolias, M. stellata in particular, have a low chill requirement, and therefore
flower buds open very early in the late winter/early spring, often just after a brief
warm spell between freezing temperatures. Flower petals die and turn brown due to
the freeze damage. Flowers of M. stellata and its hybrids are more resistant to freeze
damage than those of M. ×soulangeana. This is purely an aesthetic problem, as the
overall health of the plant is not affected.
185
Winter burn
© Amy Fulcher 2012

Excess nitrogen fertilizer or warm temperatures late into the fall season can
cause plants to remain actively growing too late in the season, and as a result, the ba-
sal portion of the trunk does not sufficiently harden before cold weather (Hartman et
al., 2000). Magnolias are thin-barked
trees and are thus considered prone to
bark cracking. Bark cracking can
be an indication of this type of cold
damage. Yellow-flowered magnolias
can be particularly susceptible to bark
cracks at the base of the trunk (Figure
9.1). Cracked bark usually becomes
evident in the spring but may occur
due to freezing conditions in the late
fall to early spring. Avoid bark
cracking by halting fertigation at the
appropriate time, e.g., September 15
in Middle Tennessee. Also summer
top-dressing should use a fertilizer
that will not continue releasing nitro-
gen too late in the season. Most
controlled-release fertilizers release less nitrogen during colder temperatures. Reduc-
ing irrigation frequency and volume towards the end of the growing season can also
help plants to stop growing and prepare for cold temperatures.
Bark cracking on magnolia also occurs due to frost cracking and sunscald,
which have many similarities (Franklin and Clatterbuck, 2004). Frost cracking and
sunscald both cause vertical cracks through the bark to the wood. Both often happen
on the south or southwest side of a tree since this is where the greatest winter tempera-
ture fluctuations occur. The temperature of bark receiving direct sun exposure is often
68 °F greater than ambient temperatures during the winter (Sinclair and Lyon, 2005);
snow exacerbates the temperature increase. Frost cracking and sunscald are linked
to root injury, aboveground wounds, and pruning cuts. Inadequate hydration is linked
to sunscald but not frost cracks (Harris et al., 2004; Hartman et al., 2000).
Frost cracking occurs when water in the wood expands and contracts as a re-
sult of dramatic temperature fluctuations such as those occurring on warm, sunny win-
ter days with periods of dense clouds or nightfall (Franklin and Clatterbuck, 2004).
Frost cracks are a physical separation of the wood. Frost cracks often close and heal,
but these trees are not marketable. Sunscald occurs when the same dramatic tempera-
ture changes damage or kill the cambium and bark. Often sunscald will not be detect-
able until spring growth resumes (Harris et al., 2004). At this point, the damage ap-
pears as sunken or discolored bark. The bark may then split. If the tree is healthy
enough to resume growth, a callus roll will develop around the wound as the season
progresses. Wrapping the trunks of young trees with a commercial tree wrap made of
insulating paper in November can prevent sunscald by mitigating temperature fluctua-
tions. Trunk wraps must be removed by early spring. Some basal bark cracks have
been observed in conjunction with large pruning cuts near the location of the crack.
[Note: refer to the maple chapter for the section on bark cracking due to herbicide
application.]
Flooding Damage: Flooding damage can occur on evergreen and semi-
evergreen magnolias that are grown in bottomlands. Flooding fills the macrospores
in the soil with water, displacing oxygen-containing air. Root cells need oxygen to
respire and to take up water. Hence, drought-like or winter burn symptoms (desicca-
tion symptoms) typically appear in response to flooding and include marginal leaf ne-
crosis and partial or complete defoliation. Magnolia virginiana is tolerant of saturated
soil.
Sapsucker Damage: Sapsuckers are a type of woodpecker. Sapsuckers peck
at trees and feed on sap, bark, and small insects that are attracted to the sap (Vann and
186
Fig. 9.1
© Amy Fulcher 2012

Robbins, 2008). Sapsucker damage is more common on larger trees in the landscape
than in nurseries, but some large-caliper nursery growers may experience problems
with sapsuckers. Sapsucker damage can be distinguished from trunk-boring insect
damage by the many uniform rows of sapsucker holes forming rings around the trunk
and large branches (Figure 9.2), whereas borer damage occurs more randomly. Sap-
sucker damage is often extensive, creates entry points for disease-causing organisms
and boring insects, and generally weakens the plants, making plants more susceptible
to other types of stresses.
187
© Sarah A. White 2012
Fig. 9.2

INSECT PESTS
1. Magnolia Serpentine Leaf Miner
2. Ambrosia Beetles
3. Scale Insects: Cottony Cushion, Calico, Magnolia, Tuliptree, False Oleander, and Japanese Maple
4. Yellow Poplar Weevil
5. Leaf-Footed Bugs
6. Magnolia Root Borer
7. Thrips
8. Cranberry Rootworm
SECTION 3
Insect Pest Management Magnolia Serpentine Leaf Miner (Phyllocnistis magnoliella): Phyllocnistis magno-
liella (Lepidoptera: Gracillariidae) is widespread throughout the eastern United States, and
larvae feed within leaves of both sweetbay (M. virginiana) and southern magnolias (M.
grandiflora) (Drooz, 1985). After hatching, larvae make 3 to 4 tight spirals before next
feeding across the leaf in a winding, serpentine pattern that ends near the edge of the leaf
(see Insect Pest Management section introductory image). Pupation occurs within the mine
and primarily, but not limited to, leaf edges. Pupae may be the overwintering state of this
caterpillar pest (Dozier, 1920). Affected leaf tissues become necrotic, scorched-looking,
and curled. Larvae are active and mines are apparent on M. virginiana by early July in
Maryland and North Carolina (Baker and Bambara, 1999; Gill, 2010). Adult moths, which
are darkly colored and nondescript, are active in late July in Florida (Dozier, 1920; Moon
and Stiling, 2004), where at least two overlapping generations per year have been reported
(Moon and Stiling, 2004).
Management: Limited work has been done to assess pesticide efficacy against P.
magnoliella on magnolias. Foliar applications of acephate or imidacloprid may be effec-
tive (Table 9.3; Gill, 2010; Smith, 2010). Broad spectrum pesticides may limit natural
enemies in the nursery and landscape. Parasitic wasps Zagrammosoma multilinieatum
and Synpiesis spp. have been reared from parasitized pupae collected in Gainesville, FL
(Dozier, 1920).
Ambrosia Beetles [including Granulate -formerly Asian- Ambrosia Beetle (Xylo-
sandrus crassiusculus), Black Stem Borer (Xylosandrus germanus), and Black Twig
Beetle (Xylosandrus compactus)]: Exotic ambrosia beetles, primarily Xylosandrus crassi-
usculus and Xylosandrus germanus (Coleoptera: Curculionidae), attack over 100 tree spe-
cies and are among the most damaging pests of nursery-grown trees and some shrubs (Hoff-
man, 1941; Oliver and Mannion, 2001; Cole 2008; Adkins et al., 2010; Atkinson et al.,
2011). They are noted as an occasional pest for magnolia (Knox, personal observation).
Female beetles are 0.08-0.12 inches in length, with males being much smaller. Both gen-
ders range in color from reddish-brown to black depending on species (Wood, 1982; Atkin-
188
© Amy Fulcher 2012Magnolia serpentine leaf miner damage

son et al., 2011). Ambrosia beetles become active in early spring (Oliver and Mannion,
2001). Ambrosia beetles bore into host tree trunks and excavate galleries in the heartwood
where adult females deposit eggs, and larvae develop (Hoffman, 1941; Weber and McPher-
son, 1983). Female ambrosia beetles inoculate trees with symbiotic ambrosia fungus on
which the larvae feed (Baker and Norris, 1968; Atkinson et al., 2011). After approxi-
mately 60 days, mature female beetles mate with males and emerge from the galleries to
find new hosts. Although X. crassiusculus and X. germanus are capable of producing three
generations per year in the southern United States, plant injury is primarily inflicted by the
first generation for reasons that are not understood.
Xylosandrus crassiusculus (Figure 9.3) and X. germanus (Figure 9.4) typically attack
small caliper (< 3.5 inch diameter) tree trunks and branches (Dixon et al., 2005). Xylosan-
drus germanus ranges from central Georgia (and possibly north Florida) north to Connecti-
cut (Dixon et al., 2005). Attacks by these borer beetles occur most heavily within 39
inches of the base of the trunk and are rare on smaller diameter branches above the first
scaffold branches (Oliver and Mannion, 2001). Damage is characterized by ‘frass tooth
picks’ that are pushed out of holes as beetles excavate their galleries. The diagnostic value
of these is short-lived as they are easily dislodged by wind, rain, and irrigation exposing a
0.04 inch round entry hole. Infested plants die or become unmarketable from boring dam-
age or infection by a secondary pathogen (Buchanan, 1941).
Xylosandrus compactus (Figure 9.5) is an occasional pest of magnolias and more than
200 other plant species, including red maples, flowering dogwood, and redbuds that serve
as common hosts, as well as many other ornamental plants (Dixon et al., 2005). Injury
may be inflicted on otherwise healthy plants, particularly in twigs less than 0.8 inch diame-
ter, and can limit plant growth and affect aesthetic losses (Dixon et al., 2005). Cankers
may be observed on larger twigs and branches that extend 0.4-8.3 inches away from the
0.03 inch entry holes that indicate the point of attack (Dixon et al., 2005). Populations of
these beetles may be encountered in the southeastern United States along the Coastal Plain
from Florida to North Carolina and west to Texas (Wood, 1982; Dixon et al., 2005). Adult
male and female X. compactus are generally smaller than X. crassiusculus and X. germanus
and are dark brown to black (Dixon et al., 2005). Within excavated twig galleries, females
establish brood galleries either individually (typically in twigs smaller than 0.3 inch diame-
ter) or in groups of up to 20 females (in twigs and branches ranging between 0.3-0.9
inches). Development from egg stage to adulthood takes about 28 days at 77 oF (Dixon et
al., 2005). Adults overwinter within damaged twigs (Wood, 1982) and like X. crassiuscu-
lus, emerge during the first warm days in spring with highest population levels found be-
tween June and September (Dixon et al., 2005).
Management: Effective ambrosia beetle management begins with monitoring adult
flight with ethanol-baited traps, which can be made easily by growers (Ranger et al., 2011).
Optimal trap height to capture first adult flight of X. germanus and X. crassiusculus ap-
pears to be 20 inches and < 67 inches, respectively (Reding et al., 2010). To limit X. com-
pactus populations, prune and destroy infested twigs and branches from affected host
189
Fig. 9.5
Bugwood
Fig. 9.4
Bugwood
Fig. 9.3
Bugwood

plants (Dixon et al., 2005). When adult beetles are captured, growers can protect their trees
by spraying susceptible tree trunks with pyrethroids, such as permethrin or bifenthrin,
every three weeks to prevent beetles from boring into the tree. It is recommended that
growers avoid spraying tree canopies because this wastes insecticide, negatively affects
natural enemies, and results in secondary pest outbreaks (Frank and Sadof, 2011). Once
beetles are inside trees, there is no effective control for either beetle larvae or the associ-
ated fungus, and infested trees should be removed and destroyed (Atkinson et al., 2011).
Cottony Cushion Scale (Icerya purchasi): In the United States, cottony cushion
scales (Homoptera: Margarodidae) are found in the western states, as well as along the
Gulf Coast and eastern states as far north as North Carolina (Johnson and Lyon, 1988).
Winter temperatures of about 10 °F may limit spread of cottony cushion scales in land-
scapes and nurseries. Scales can persist season-long if protected in propagation structures
and greenhouses (Baker and Frank, 2010). Immature cottony cushion scales reduce vigor
of host plants, including boxwood, cypress, hackberry, Magnolia spp., Malus spp., maples,
nandina, oak, pecan, Pittosporum spp., Prunus spp., Pyrus spp., quince, rose, salvia, and
willow, by piercing plant tissues and sucking out sap (Johnson and Lyon, 1988; Baker and
Frank, 2010). Heavily infested plants become chlorotic and often become darkened by ex-
creted honeydew and resultant sooty mold. Leaves and fruit on heavily infested plants
may drop prematurely following environmental or other stressors.
Cottony cushion scales are often obscured by exuded wax that helps to protect them
from predation and direct pesticide contact (Figure 9.6). Beneath the wax, adult females
are about 0.2 inches long and rusty red with black legs and antennae. Male cottony cush-
ion scales are about 0.1 inches long and slender. Adult male scales are uncommon and
when present, are reddish-purple insects that have metallic blue wings (Baker and Frank,
2010). Cottony cushion scales have about two generations per year in their northern range
with three and more generations possible in southern U.S. landscapes.
Female cottony cushion scales can be diagnosed by the presence of 3/8-inch-long
ridged egg sacs formed from waxes extruded from the underside of the female body. The
ovisac contains hundreds of reddish oval eggs that may hatch to crawlers within three
weeks (in summer) to eight weeks (in winter) of being laid. All three nymphal instars are
mobile as crawlers, seeking leaves and twigs to feed on, and upon hatching, can travel up
to 36 inches in 10 minutes (Baker and Frank, 2010). Adult female scales may live 2 to 3
months and may not need to feed after beginning to lay eggs. They are capable of produc-
ing ovisacs and laying eggs after moving away from the host plant.
Management: Cottony cushion scales have relatively few known predators and para-
sites. Vedalia beetles (Rodolia cardinalis) effectively control cottony cushion scales when
temperatures are warm but are susceptible to insecticides and may become non-target casu-
alties following pesticide applications (Baker and Frank, 2010). A parasitic fly from Austra-
lia, Cryptochaetum iceryae, has been introduced to control cottony cushion scale. Whether
the fly has successfully become established or is effective at controlling I. purchasi in mid-
southern states has not been determined (Baker and Frank, 2010).
Dinotefuran is specifically labeled for cottony cushion scale control, and other insecti-
cides are labeled to provide control of scales and scale crawlers. Use of spreader-stickers,
190
Fig. 9.6
Bugwood

including summer horticultural oil that can be used at 2 quarts/100 gallons (4 tsp/gal), is
recommended to improve cottony cushion scale control by helping pesticides penetrate the
waxy ovisac and scale covering. Horticultural oils themselves are moderately toxic to
scales and may also help disperse sooty mold (Baker and Frank, 2010).
Calico Scale (Eulecanium cerasorum): Calico scale adults and nymphs (Homop-
tera: Coccidae) are readily visible on branches and leaves, giving trees a bumpy appearance
under heavy infestation (Johnson and Lyon, 1988). Adults are rounded and globular, about
0.25-0.3 inches long, and at maturity, are checkered in a white and dark brown “calico”
pattern (Figure 9.7). Nymphs are small, cream colored ovals. In addition to magnolias,
including M. stellata, calico scales infest buckeye, dogwood, elm, flowering crabapple,
hackberry, honeylocust, maples, pears, Pyracantha spp., redbud, sweetgum, Virginia
creeper, walnut, Wisteria spp., yellowwood, and Zelkova spp. (Hubbard and Potter, 2005,
2006; Krischik and Davidson, 2007). Scale feeding on phloem sap results in excreted hon-
eydew that accumulates beneath infested trees. Calico scale feeding, as well as the sooty
mold that grows on honeydew, results in reduced tree vigor, premature leaf abscission,
and branch dieback, all of which predispose infested trees to increased environmental
stress (Hubbard and Potter, 2006; Krischik and Davidson, 2007).
Calico scales have one generation per year. Juveniles overwinter as second instar
nymphs on bark. Females can generate up to 4,600 eggs each that remain protected be-
neath the female body until crawlers emerge. Egg laying begins approximately in late
April in Kentucky, and eggs hatch almost synchronously about mid May, following ~818
degree day accumulation (at base about 40 °F; Hubbard and Potter, 2005). Crawlers dis-
perse during the next 2 to 3 weeks to feed on leaf undersides. After feeding from late Sep-
tember to mid-October (Kentucky), nymphs migrate back to bark to overwinter (Hubbard
and Potter, 2005).
Management: Twigs and branches can be inspected for nymphs and adults in spring,
before leaf emergence, and whenever honeydew and sooty mold are detected. When foli-
age is present, inspect the main leaf veins using double-sided tape wrapped around a twig
to monitor tiny cream-colored crawler emergence (Hubbard and Potter, 2006; Krischik and
Davidson, 2007). Prune and destroy any lightly infested branches. Heavy infestations may
require chemical foliar sprays, soil drenches, and granular systemic insecticide treatments.
Contact insecticides may kill beneficial insects and should be timed for application to coin-
cide with crawler emergence. Predators and other biological control organisms, including
green lacewing adults and larvae, may be enhanced by using sticky bands to limit ant ac-
cess to magnolia scales in landscape plants (Vanek and Potter, 2010). Avoid broad-
spectrum insecticides when natural enemies are present and direct sprays toward stems,
branches, and leaf undersides. Biological control agents, including many different preda-
tors and parasitoid species, may suppress calico scales. Curative efficacy by biological
controls in landscapes and nurseries, however, has not been sufficiently tested (Hubbard
and Potter, 2006; Krischik and Davidson, 2007).
Magnolia Scale (Neolecanium cornuparvum): Magnolia scale, Neolecanium cornu-
parvum (Homoptera: Coccidae), is a pest native to the United States that, at about 0.3-0.4
inches (up to 0.5 inches) long, is one of the largest scale insects encountered in U.S. land-
scapes and nurseries (Herrick, 1931; Vanek and Potter, 2010). Magnolia stellata, M. acumi-
nata, M. liliiflora, M. ×soulangeana, and M. grandiflora are the preferred host plants of
191
Fig. 9.7
© Amy Fulcher 2012

this pest (Herrick, 1931; Johnson and Lyon,
1988; Vanek and Potter, 2009). Magnolia
scales produce copious amounts of honey-
dew. When coupled with feeding injury to
magnolias, which removes photosynthates
and induces cell necrosis due to phytotoxic
saliva, heavy scale infestations can cause
branch death, loss of tree vigor, eventual
tree decline, and death (USDA Forest Serv-
ice, 1985; Vanek and Potter, 2009).
Magnolia scales pass through a single
generation per year, overwintering as first
instar nymphs, which are elliptical and slate
gray in color and often gather in masses
along the undersides of 1- to 2-year-old
twigs (Johnson and Lyon, 1988). In New
York, first instars molt approximately in late April or early May, and the second instars
molt during early June (Johnson and Lyon, 1988). By July in Ohio and Kentucky (Boggs
et al., 1998; Townsend, 2005), or August in New York and Pennsylvania (Johnson and
Lyon, 1988; Mazzey and Masiuk, 2011), the large, cream-to-yellow-colored females (Fig-
ure 9.8) give live birth to the next generation of scale nymphs. Newly emerged nymphs
are mobile for a short time and then settle to feed on the semi-hardwood new growth of
host plant twigs.
Management: In small populations, the large female scales can be removed by hand
before crawlers hatch (e.g., before mid-July). Severely infested branches can be pruned
and destroyed. Use horticultural dormant oil sprays when magnolia stems are dormant
and before buds swell and break in April (Mazzey and Masiuk, 2011). When natural ene-
mies including ladybird beetles, predatory mites, and parasitic wasps are present, azadirac-
tin, horticultural “summer” oil, insecticidal soap, and neem oil extract may be good choices
to conserve biological controls. Predators and other biological control organisms, includ-
ing green lacewing adults and larvae, may be enhanced by using sticky bands to limit ant
access to magnolia scales in landscape plants (Vanek and Potter, 2010). Fall foliar and
stem sprays may be made after egg hatch, which occurs around September in Kentucky
(Vanek and Potter, 2010), starting in early October and on 7- to 10-day intervals, once early
instar crawlers are actively feeding and while they remain relatively unprotected by cuticu-
lar waxes. Effective insecticides include acephate, buprofezin, carbaryl, chlorpyrifos, cyflu-
thrin, deltamethrin, dimethoate, dinotefuran, esfenvalerate, imidacloprid, lambda-
cyhalothrin, malathion, permethrin, and thiamethoxam (Townsend, 2005; Mazzey and Ma-
siuk, 2011). Repeat insecticide applications may be necessary regardless of the pesticide
chosen.
Tuliptree Scale (Toumeyella liriodendra): Tuliptree scales (Homoptera: Coccidae)
are significant pests of tulip poplar (Liriodendron tulipifera) in silviculture and landscapes,
as well as Magnolia species and linden in landscapes and nurseries (Hoover, 2006). Tulip-
tree scales feed on all stages of seedling, sapling, and poletimber poplar trees, reduce tree
vigor and health, and cause distortion and death of infested trees (Burns and Donley, 1969).
Tuliptree scales are large: about 0.3 inch in diameter, and are light gray-green to pinkish
orange ovals mottled with black (Figure 9.9; Hoover, 2006). Toumeyella liriodendra over-
winters as second instar nymphs. Nymphs resume feeding in spring, and females give live
birth to up to 3,000 crawlers beginning in August (Hoover, 2006). Mating occurs in June.
Females produce copious amounts of honeydew from June through August (Burns and
Donley, 1969). Nymphs are dark red and about 0.02 inches long and may be dispersed by
crawling, as well as via wind and transfer on plumage of songbirds (Hoover, 2006). There
is only one generation per year (Burns and Donley, 1969; Hoover, 2006).
Management: Dormant-season horticultural oils can be applied to trees and shrubs
while dormant. Summer oil treatments can be made to control eggs, crawlers, and imma-
ture instars on actively growing trees and shrubs. Control with oil is achieved by smother-
192
Fig. 9.8
Bugwood

ing and ostensibly through disruption of cellular respiration. Crawler stages can be treated
with acephate, azadirachtin, buprofezin, carbaryl, chlorpyrifos, cyfluthrin, cyfluthrin plus
imidacloprid, deltamethrin, dinotefuran, horticultural oil, imidacloprid, insecticidal soap,
lambda-cyhalothrin, neem oil extract, and thiamethoxam insecticides. Broad spectrum in-
secticides, including acephate, carbaryl, cyfluthrin, deltamethrin, imidacloprid, and
lambda-cyhalothrin, may result in non-target kill of beneficial arthropods. Tuliptree scales
have several reported predators including a predaceous moth larva (Laetilia coccidivora), a
ladybird beetle (Hyperaspis proba proba), and syrphid fly (Baccha costata; Burns and Don-
ley, 1969).
False Oleander Scale - aka Magnolia White Scale - (Pseudaulacaspis cockerelli):
False oleander scale (Homoptera: Diaspididae) is an armored scale pest of Chinese origin
that is commonly encountered on foliage of many ornamental Magnolia spp., as well as
aucuba, Eleagnus spp., English ivy, flowering dogwood, oleander, and yew plants in land-
scapes and nurseries in Georgia, Florida, and Alabama (Johnson and Lyon, 1988; Deckle,
1976; Hamon and Fasulo, 2007). Foliage of affected plants can be scouted by looking on
both leaf surfaces for the pear-shaped, slightly convex, yellow-brown scale exuviae. The
shiny white female armor (Figure 9.10) may extend 0.08-0.12 inches and is composed of
secreted waxes and cast skins. Males are smaller, at about 0.04 inch long (Deckle, 1976;
Hamon and Fasulo, 2007). Feeding injury may be apparent as chlorotic spots on the upper
leaf surface with scales attached to the leaf under side. Heavy infestations of false oleander
scale may lead to premature leaf drop and loss of host plant vigor (Hamon and Fasulo,
2007). False oleander scales are present year-round where encountered, and controlled
greenhouse studies indicate that a single generation can complete development within
about five weeks (Tippins, 1968).
Management: Armored scale insects such as mature false oleander scale are difficult
to control. Pesticide applications are best timed for crawler emergence. Double-sided
sticky tape can be used to monitor egg hatch and crawler activity. Dead scales do not fall
from plants; thus, to assess pesticide efficacy, crush the waxy covering. When crushed, gut
contents will be extruded from live armored scale insects. Acephate, bifenthrin, diazinon,
fenoxycarb (against immature scales only), horticultural oils, and malathion have been ef-
fective at managing scale insects on ornamental plants, including false oleander scale popu-
lations (Leibee and Savage, 1994). Use of
horticultural oils will conserve natural
enemy populations.
Japanese Maple Scale
(Lopholeucaspis japonica): Japanese ma-
ple scale is a small, oyster shell-shaped,
armored scale believed to have been intro-
duced to the United States from Asia.
Japanese maple scale is currently found
throughout much of the eastern half of the
United States (Gill et al., 2011). Its wide
host plant range includes Magnolia spp.
(Fulcher et al., 2011). Japanese maple
scale infestation can cause branch dieback
193
Fig. 9.9
© Amy Fulcher 2012
Fig. 9.10
Bugwood

progressing to plant death, although this is uncommon. Because Japanese maple scale is
small and easily overlooked, plants are often heavily infested when first noticed. It can
spread over a significant portion of a nursery before the pest is detected, and L. japonica
infestation has caused nursery crop shipments to be rejected.
Japanese maple scale is expected to have two generations a year in the mid-southern
United States. This scale overwinters as an immature nymph on trunks and branches and
matures to an adult in spring (Miller and Davidson, 2005). First generation crawlers
emerge in mid-May and the second generation in early August (Fulcher et al., 2011; Gill et
al., 2011). Japanese maple scale crawlers hatch over an extended period, and first and sec-
ond generations overlap (Fulcher et al., 2011; Gill et al., 2011). Crawlers develop wax
rapidly, starting about three days after hatch (Fulcher et al., 2011). In Kentucky, crawler
hatch coincides with flowering of Syringa reticulata ‘Ivory Silk’ (tree lilac) and Hydrangea
quercifolia (oakleaf hydrangea; Fulcher et al., 2011). In Maryland, the first generation be-
gan at 795 degree days (50 °F base temperature, starting January 1) and peaked at 1,144
degree days, while the second generation started at 2,200 degree days and peaked at 3,037
degree days (Gill et al., 2011).
Japanese maple scale can be difficult to detect because the lavender crawlers are
very small (as are adults) and generally require 16X or greater magnification to detect.
Japanese maple scale is most readily observed on bark of dormant deciduous host plants
and on developing magnolia fruit and leaves. Managers may need to pull aside branches of
densely branched trees and shrubs when scouting. Bodies of adult L. japonica scales have
a white waxy coating, while female body, eggs, and crawlers are lavender.
Management: Japanese maple scale and other armored scales are difficult to control.
Plants with thick canopies can be difficult to successfully treat for scale insects because the
dense foliage interferes with spray penetration. Pruning practices may need to be adjusted
to open canopies and enable greater spray penetration. Placement of water sensitive paper
inside the canopy before spray application can ensure that pesticides are penetrating the
canopy to reach infested portions of the plant. Best pesticide efficacy occurs if applications
are timed for crawler emergence. Double-sided sticky tape can be used to monitor egg
hatch and crawler activity. Dead scales do not fall from plants, and pesticide efficacy must
be assessed. When crushed, live armored scale insects will exude gut contents. This
scale's small size makes this difficult to do in the field. To better assess mortality after pes-
ticide application, clip an infested twig or branch and flip the scale upside down to expose
the body. A hand lens or microscope will show dead scales as shriveled and papery, while
live scales are firm and plump. Acephate, bifenthrin, diazinon, fenoxycarb (against imma-
ture scales only), horticultural oils, and malathion have been effective at managing scale
insects on ornamental plants (Leibee and Savage, 1994). Use of horticultural oils will con-
serve natural enemy populations.
Yellow Poplar Weevil (Odontopus calceatus): Yellow poplar weevils, also called
magnolia leafminer, tuliptree leafminer, and sassafras weevil (Odontopus calceatus, Cole-
optera: Curculionidae), are widespread throughout the eastern United States where the lar-
vae feed on leaves and buds of tulip poplar (L. tulipifera), sassafras, and both sweetbay and
southern magnolia (Burns, 1971; Johnson and Lyon, 1988). Heavy infestations can lead to
multiple stem leaders that emerge from beneath a killed apical terminal and defoliation of
the tree canopy. Weakened and malformed trees may become liabilities in the landscape.
Populations have occasionally reached damaging levels in tulip poplar orchards and ma-
ture trees within the Appalachian Mountain region (Burns, 1971; Heinrichs et al., 1973).
Adult beetles (Figure 9.11) are about 0.1 inches long, rounded, and dark brown or
black. Beetles overwinter in leaf litter beneath host plants and emerge with warm daytime
temperatures to begin feeding in late March and early April, chewing notches and holes in
expanding leaves and buds. In late April and early May, eggs are laid on leaf undersides
along the mid-vein (Burns, 1971). Weevil larvae feed as leafminers, inflicting initial feed-
ing injury in late April through May. Early larval feeding causes blotch mines that may
contain up to 19 white, legless larvae (Burns, 1971). Most larval feeding occurs in May,
after which larvae pupate within the mine. Leaves with mines may look scorched. Adults
194

continue to feed, causing significant injury into summer (Burns, 1971) and until August in
Mississippi (Heinrichs et al., 1973). Odontopus calceatus bears only one brood per year in
the Ohio River Valley (Burns, 1971).
Management: Late spring frosts may kill larvae and adults that have emerged from
overwintering sites. Known natural enemies include two pteromalid wasps, Heterolaccus
hunter and Habrocytus peircei, two eulophid wasps, Horismenus fraternus and Zagrammo-
soma multilineatum, and an ichneumonid wasp, Scambus hispae (Burns, 1971). Insecti-
cides labeled for beetles and caterpillars that feed on leaves may provide some control.
Leaf-Footed Bugs (Leptoglossus fulvicornis and Leptoglossus phyllopus): Ranging
from Massachusetts to Texas, leaf-footed bugs (Heteroptera: Coreidae) feed on and can
damage developing and mature magnolia fruits (Mitchell and Mitchell, 1983). Magnolia
species are the only known breeding hosts. Eggs are laid in linear masses along the midrib
on leaf undersides. Nymphs and adults are active in late summer and fall as fruits develop.
Management: Gryon pennsylvanicum and Anastatus spp. can parasitize eggs but will
not control infestations (Mitchell and Mitchell, 1983). Control of leaf-footed bugs may be
necessary or desirable if growing Magnolia spp. to collect seed for propagation, particu-
larly if the Magnolia spp. in question are rare or endangered. Magnolia breeders should
also be aware of the potential damage inflicted by this pest.
Magnolia Root Borer (Euzophera magnolialis): Magnolia root borer, Euzophera
magnolialis (Lepidoptera: Pyralidae), is a wood boring pest with larvae that feed on mag-
nolia roots 0.2 inch and larger and tunnel about 6 inches up into the trunk above the crown
(Leibee, 1992). The resulting girdling injury can injure and even kill nursery seedlings
(Leibee, 1992; Outcalt, 1990; Popenoe, 2008). Root feeding may be in a spiral, causing
affected roots to have a candy cane-patterned injury (Leibee, 1992). Larval feeding limits
root function, including nutrient and water uptake, thereby inducing initial plant decline.
Adult moths live about 10 days (Leibee, 1992), and adults and larvae are often difficult to
detect. They are more of an economic threat to magnolias under nursery production, rather
than in an urban landscape (Leibee, 1992). Eggs may be deposited on the tree bark (a sin-
gle female can lay about 400 eggs under lab conditions) from the crown up to about 4.5
feet (Leibee, 1992), and larvae tunnel down to feed on tree roots. As larvae mature, they
migrate up toward the crown and begin tunneling beneath bark to pupate. In Florida and
the deep southern United States, E. magnolialis has at least two generations per year, where
adult moth flight activity occurs from February through April and again from June through
August (Leibee, 1992; Popenoe, 2008).
Management: High fertilization and irrigation rates in nurseries may mask plant in-
jury by allowing adventitious roots to overcome damage symptoms and plant stress (Pope-
noe, 2008). If first order lateral, or structural, roots are compromised, root systems may
only persist shallowly once trees are planted in the landscape, yielding poor structural sup-
port to the mature tree. Information about pesticide efficacy against this insect is limited.
The extent to which biological control may be effective is unknown. Root and crown
drenches with a systemic insecticide labeled for borers may provide some control.
195
Fig. 9.11
Bugwood

Thrips (Caliothrips striatus): Caliothrips striatus (Thysanoptera: Thripidae) is a
thrips insect native throughout the eastern United States (Wilson, 1975). Though not
considered a significant pest, Tyler-Julian et al. (2012) identified several Magnoliaceae
species as host plants to C. striatus, including the native L. tulipifera, M. grandiflora
and M. virginiana and the non-native Magnolia delavayi, Magnolia foveolata, M. insig-
nis × M. grandiflora (hybrid), M. laevifolia, Magnolia lotungensis, M. maudiae, and
Magnolia tamaulipana. Magnolias in the affected botanic garden where thrips occurred
exhibited damage caused by C. striatus, that appeared as yellow-brown flecks on the
leaf surface. The irrigated magnolias in the botanic garden are believed to have at-
tracted this thrips from drought-affected native vegetation surrounding the garden (Fi-
glar, personal communication). Adults were sometimes visible on leaves and appeared
black with two white bars. Caliothrips striatus is believed to be capable of using M.
grandiflora both for feeding and for reproduction.
Management: As a minor pest, this thrips will rarely need to be controlled. When
pest populations threaten plant health or aesthetics, pesticides may be used (Table 9.3).
Cranberry Rootworm (Rhadopterus picipes): Cranberry rootworm, Rhadopterus
picipes (Coleoptera: Chrysomelidae), occurs east of the Mississippi River and has an ex-
tremely broad host plant range. In addition to M. virginiana and other magnolia species,
host plants include camellia, cherry laurel, goldenraintree, Ilex crenata, Ilex cornuta, oaks,
photinia, rhododendron, rose, silver maple,
sycamore, sumac, sassafras, and Virginia
creeper. Adult beetles are about 0.2 inch
long, dark brown, and shiny (Figure 9.12).
Adults bear one brood per year and emerge
from late April to mid-May in Mississippi
(Harman, 1931; Oliver and Chapin, 1980;
Johnson and Lyon, 1988). Adults are noc-
turnal feeders, hiding in leaf litter and de-
bris during the day. Adults feed for about 2 weeks after emergence and then seek refuge in
leaf litter where they deposit eggs (Oliver and Chapin, 1980).
Management: Pesticides including carbaryl, imidacloprid, permethrin, and spinosad
may provide control when beetles are actively feeding. Applications should also be di-
rected toward leaf litter and debris beneath the affected plant where nocturnal beetles will
hide. Entomopathogenic nematodes including Heterorhabditis bacteriophora and
Steinernema scarabaei have shown some potential for cranberry rootworm control (Pola-
varapu, 1999; van Tol and Raupp, 2005).
196
Fig. 9.12
© Sridhar Polaverapu 2012

DISEASE & NEMATODE PESTS
1. Bacterial Leaf Spot
2. Verticillium Wilt
3. Phytophthora Root Rot
4. Bacterial Blight
5. Anthracnose
6. Algal Leaf Spot
7. Powdery Mildew
8. Phyllosticta Leaf Spot
9. Pestalotiopsis Leaf Spot
10. Nematodes
SECTION 4
Disease PestsBacterial Leaf Spot (Xanthomonas sp.): Xanthomonas sp. causes bacterial
leaf spot on magnolia. The disease is a serious issue in production under conditions
of high rain and wind or heavy overhead irrigation, all of which lead to easy move-
ment of the bacteria from plant to plant. The pathogen can enter through wounds,
stomata, and hydathodes. Symptoms of bacterial infections vary but usually start
as small necrotic lesions with a yellow halo region. Under ideal environmental con-
ditions, the pathogen can cause severe blighting of the foliage (see Disease Pests
section introductory image).
Management: Manage overhead irrigation carefully to prevent high leaf wet-
ness during nights and overcast conditions. If ideal environmental conditions for
the pathogen persist, the disease can be managed by use of copper-based bacteri-
cides (Table 9.4). The disease is an issue in nursery production due to poor manage-
ment of overhead irrigation systems and is not usually seen in the landscape.
Verticillium Wilt (Verticillium albo-atrum and Verticillium dahlia): Verti-
cillium sp. is a vascular pathogen of magnolia as well as many other ornamental
shrubs and trees. It can cause dieback of leaves and branches, often at one side of
the tree. Infected plants often show vascular discoloration, a characteristic symp-
tom of verticillium wilt. The fungus may kill large areas of cambial tissue and al-
low opportunistic fungi such as Nectria sp. to infect the area and form elongated
cankers (Chartfield et al., 1996). Symptom progression can be slow, but the plants
will completely die in 1 to 2 years. Verticillium sp. survives in soil as microsclero-
tia that can be disseminated by wind, soil movement, and on equipment. The fun-
gus penetrates through root wounds or direct penetration, and root stressors such as
drought favor its development. The disease symptoms are caused by the fungus
plugging the vascular system.
Management: Avoid stressful conditions for the plants including overwater-
ing, drought, and root wounding that will aid Verticillium sp. to infect magnolia.
197
© Mathews Paret 2012
Bacterial leaf spot on magnolia

Sanitize pruners frequently when pruning roots of susceptible species. Fungicides
are not effective in management of verticillium wilt. Verticillium sp. can survive in
soil for many years, so avoid planting susceptible species in areas previously in-
fected by this fungus.
Phytophthora Root Rot (Phytophthora cinnamomi): Phytophthora root
rot symptoms include yellowing of leaves, sudden wilting, premature leaf drop,
slowed shoot growth, limb dieback, and plant death. This disease is a major con-
cern in container production of southern and saucer magnolias (Hagan, 2001).
Phytophthora cinnamomi survives as hyphae in infected roots and as rest-
ing structures known as chlamydospores in crop debris and soil. The oomycete
causal agent is easily and quickly spread by contaminated water in nurseries where
irrigation water is recycled without treatment. Root infection can be common and
severe if container substrates are saturated.
Management: Proper irrigation management and preventive and timely fun-
gicide applications (Table 9.4) are important in managing phytophthora root rot in
container grown magnolias. Avoid conditions that allow water to pool around con-
tainers. Thoroughly wash and disinfect recycled containers before use.
Bacterial Blight (Pseudomonas syringae pv. syringae and Pseudomonas
cichorii): These Pseudomonas sp. cause leaf spots similar to bacterial leaf spot on
magnolia (Miller, 1976; Mullen and Cobb, 1984; Hagan, 2001). Symptoms are
small necrotic spots with a yellow halo (Figure 9.13). The leaf spot symptoms
caused by Pseudomonas sp. are difficult to distinguish from bacterial leaf spot,
caused by Xanthomonas sp. However, Pseudomonas sp. is much more an issue dur-
ing cooler, wetter conditions. Both Pseudomonas species are aggressive pathogens
on southern magnolia.
Management: Manage overhead irrigation carefully to prevent high leaf wet-
ness during nights and overcast conditions. If ideal environmental conditions per-
sist, the disease can be managed by use of copper-based bactericides (Table 9.4).
The disease is an issue in nursery production due to poor management of overhead
irrigation systems and not usually seen in the landscape.
Anthracnose (Colletotrichum spp.): Anthrac-
nose causes large circular spots near margins of mag-
nolia leaves (Figure 9.14). The leaves develop a
burned appearance or angular spots with a yellow
halo, and this can lead to premature leaf drop (Ha-
gan, 2001). Black, blister-like fruiting bodies de-
velop on the upper side of the spot. A pink spore
mass oozes from these fruiting bodies at later stages.
Spores can be spread by splashing water. The fungal
organism can overwinter in dead leaves and branches
during winter. The disease is often reported on south-
ern magnolia.
198
Fig. 9.13
© Mathews Paret 2012
Fig. 9.14
© Mathews Paret 2012

Management: Remove infected leaves at the early stages to prevent any dis-
ease spread. A fungicide management plan is required if the infection becomes se-
vere (Table 9.4).
Algal Leaf Spot (Cephaleuros virescens): The key symptom of algal leaf
spot is the formation of raised blotches on the leaves (Figure 9.15). These spots
develop a velvety appearance, and plant tissues beneath the spots die. Severe infec-
tion can cause premature leaf drop. The disease mostly affects weaker plants in the
nursery or landscape and is commonly seen on southern magnolia. Algal spores
can swim in water on the leaf surface. Thus, continuous rain during warm, windy
conditions serves as a major factor in spread of the algal spores. In addition, the
algae may survive on infected leaves and twigs and result in repeated infection year
after year.
Management: For early crop stage management, removing infected leaves
and plant debris will serve as useful strategies. Overhead irrigation should be man-
aged to avoid long periods of leaf wetness. Crop canopy management is also impor-
tant as dense canopy favors disease conditions.
Powdery Mildew (Microsphaera alni [M. penicillata] and Phyllactinia
corylea [P. guttata]): Two species of fungi cause powdery mildew on magnolia.
The symptoms include white powdery patches on top sections of the leaves (Figure
9.16; Hagan, 2001). The patches can cover the entire leaf during severe infection.
Plant stunting and leaf curling can occur. Powdery mildew fungi overwinter as hy-
phae in dormant buds or as spores on fallen diseased leaves. Spores are spread by
wind in spring to early summer. Powdery mildew can be an issue in production
when days are warm-to-hot and nights are cooler with dew formation on leaves.
Overcrowding plants and lack of aeration are environmental conditions that support
powdery mildew infection. Powdery mildew is commonly found on saucer and star
magnolias.
Management: Plant disease management tools include use of good cultural
practices. Plant canopies should be maintained to allow air movement. Similarly,
plants in production should be spaced to allow air movement around plants. Scout
for the disease during times when days are warm and nights are cool. If the infec-
199
Fig. 9.15
© Mathews Paret 2012
Fig. 9.16
© Amy Fulcher 2012

tion is spotted and these environmental conditions are likely to persist, it will be
necessary to implement a fungicide spray program (Table 9.4).
Phyllosticta Leaf Spot (Phyllosticta magnolia): Phyllosticta leaf spot is an
occasional disease on southern magnolia in the landscape. Symptoms are small
black spots on the upper leaf surface (Hagan, 2001; Pataky, 1997). Borders of these
spots become purple to black, and centers turn off-white and develop black fruiting
bodies. Warm, moist conditions favor this disease. The disease organism is spread
to new leaves by splashing water.
Management: Overhead irrigation should be managed to avoid long periods
of leaf wetness during warm, humid conditions. Removing infected leaves early in
disease progression can prevent spread of phyllosticta leaf spot.
Pestalotiopsis Leaf Spot (Pestalotiopsis sp.): Pestalotiopsis leaf spot of
magnolia causes unique circular spots with a necrotic center and a black border and
usually occurs during cooler weather conditions (Figure 9.17). Fungal spores of the
pathogen are spread by wind and water movement.
Management: Overhead irrigation should be managed to avoid leaf wetness
overnight and during overcast conditions. Maintaining an open canopy is also help-
ful in minimizing conditions favorable for disease development.
Nematodes (Belonolaimus sp. and Meloidogyne sp.): Magnolia species
are hosts of sting nematodes (Belonolaimus species) that cause roots to darken and
rot (Hagan, 2005). Unknown nematodes have been found on some Magnolia spe-
cies (Santamour and Batzli, 1990). Other studies found various Magnolia species
were tolerant or resistant to many root knot nematodes (Meloidogyne species; San-
tamour and Reidel, 1993; Bernard and Witte, 1987).
Management: Nematodes are minor pests of magnolia. Control is not practi-
cal in most situations.
Virus and canker diseases are not documented on Magnolia species. Table
9.4 lists fungicides labeled for Magnolia species.
200
Fig. 9.17
© Mathews Paret 2012

SECTION 5
ReferencesAdkins, C., G. Armel, M. Chappell, J.-H.Chong, S.D. Frank, A. Fulcher, F. Hale, K. Ivors, W. Klingeman, A. LeBude, J. Neal, A. Senesac, S. White, J. Williams-Woodward, and A. Windham. 2010. Pest Management Strategic Plan for Container and Field-Produced Nursery Crops in Georgia, Kentucky, North Carolina, South Carolina and Tennessee. A. Fulcher (ed.). Southern Region IPM Center. http://www.ipmcenters.org/pmsp/pdf/GA-KY-NC-SC-TNnurserycropsPMSP.pdf. Accessed April 19, 2012.
Atkinson, T.H., J.L. Foltz, R.C. Wilkinson, and R.F. Mizell. 2011. Granulate ambro-sia beetle, Xylosandrus crassiusculus (Motchulsky) (Insecta: Coleoptera: Curculi-onidae: Scolytinae). Florida Cooperative Extension Service, IFAS, University of Florida, EENY-131.
Azuma, H., L.B. Thien, M. Toyota, Y. Asakawa and S. Kawano. 1997. Distribution and differential expression of (E)-4,8-dimethyl-1,3,7-nonatriene in leaf and floral volatiles of Magnolia and Liriodendron taxa. Journal of Chemical Ecology 23(11):2467-2478.
Baker, J.M. and S. Bambara. 1999. North Carolina pest news: Ornamentals and turf 14(12).http://ipm.ncsu.edu/current_ipm/99PestNews/99News12/ornament.html. Ac-cessed April 19, 2012.
Baker, J.M. and D.M. Norris. 1968. A complex of fungi mutualistically involved in the nutrition of the ambrosia beetle Xyleborus ferrugineus. Journal of Invertebrate Pathology 11:246-250.
Baker, J.B. and S. Frank. 2010. Cottony cushion scale. North Carolina Coopera-tive Extension ENT/ort-51. http://www.ces.ncsu.edu/depts/ent/notes/O&T/shrubs/note51/note51.html. Accessed April 19, 2012.
Bernard, E.C. and W.T. Witte. 1987. Parsitism of woody ornamentals by Melio-dogyne hapla. Annals of Applied Nematology 1:41-45.
Boggs, J.F., D.J. Shetlar, J.C. Martin, P.J. Bennett, J.A. Chatfield, G.Y. Gao, and C. Carlson. 1998. Insect and mite activity noted in Ohio nurseries and landscapes:
1997. OSU-ARDC, Ornamental Plants Annual Report Research Review special circular 157. http://ohioline.osu.edu/sc157/sc157_6.html. Accessed April 19, 2012.
Buchanan, W.D. 1941. Experiments with an ambrosia beetle Xylosandrus germanus (Blfd.). Journal of Economic Entomology 34:367-369.
Burns, D.P. 1971. Yellow poplar weevil. USDA Forestry Service Forest Pest Leaflet 125.
Burns, D.P. and D.E. Donley. 1969. Biology of the Tuliptree scale, Toumeyella lirio-dendri (Homoptera: Coccidae). Annals of the Entomological Society of America 63:228-235.
Callaway, D.J. 1994. The world of magnolias. Timber Press, Inc., Portland, Oregon.
CDMS. 2011. CDMS Label Search. Crop Data Management Systems, Inc. Ac-cessed October 21, 2011. http://premier.cdms.net/webapls/formsloginRef.asp?/webapls.
Chatfield, J., S. Nameth, and N. Taylor. 1996. Verticillium wilt of landscape trees and shrubs. Ohio State University Extension factsheet. HYG-3053-96.
CDMS. 2011. CDMS Label Search. Crop Data Management Systems, Inc. http://premier.cdms.net/webapls/formsloginRef.asp?/webapls. Accessed April 19, 2012.
Cole, K.W. 2008. Granulate ambrosia beetle. Indiana Department of Natural Re-sources. http://www.in.gov/dnr/entomolo/files/ep-GranulateAmbrosiaBeetleFactsheet.pdf. Accessed April 19, 2012.
Deckle, G.W. 1976. Florida armored scale insects. Florida Department of Agricul-ture Consumer Services, Division of Plant Industry. Arthropods of Florida and Neighboring Land Areas 3:1-345.
Diagne, A.B., R.N. Story, and A.M. Hammond. 2006. Adult Phyllophaga ephilida host plant feeding preference. Florida Entomologist 89:391-395.
201

Dirr, M.A. 1975, rev. 1998. Manual of woody landscape plants. Stipes Publishing L.L.C., Champaign, Illinois.
Dixon, W.N., R.E. Woodruff, and J.L. Foltz. 2005. Black twig borer, Xylosandrus compactus (Eichhoff) (Insecta: Coleoptera: Curculionidae: Scolytinae). Florida Co-operative Extension Service, IFAS, University of Florida, EENY-311.
Dozier, H.L. 1920. An ecological study of hammock and piney woods insects in Florida. Annals of the Entomological Society of America 13:326-380.
Drooz, A.T. 1985. Insects of eastern forests. U.S. Department of Agriculture Forest Service, miscellaneous publication 1426.
Engelbrecht, C. 2005. Host/pest list for diseases on ornamentals in Iowa. School IPM Landscape Pilot Project. http://www.ipm.iastate.edu/ipm/schoolipm/sites/default/files/Host_Disease_Pest_Guide_List.pdf. Accessed April 19, 2012.
Figlar, R.B. and H.P. Nooteboom. 2004. Notes on Magnoliaceae IV. Blumea 49:88-100.
Frank, S.D. and C.S. Sadof. 2011. Reducing insecticide volume and non-target ef-fects of ambrosia beetle management in nurseries. Journal of Economic Entomol-ogy 104:1960-1968.
Franklin, J. and W. Clatterbuck. 2004. Bark splitting on trees. University of Tennes-see Extension publication SP630. https://utextension.tennessee.edu/publications/Documents/SP630.pdf. Accessed April 19, 2012.
Fulcher, A., F. Hale, and M. Halcomb. 2011. Japanese maple scale: An important new insect pest in the nursery and landscape. University of Tennessee Extension publication W277.
Gill, S. 2010. TPM/IPM weekly report for arborists, landscape managers, and nurs-ery managers. http://ipmnet.umd.edu/landscape/lndscpalerts/2010/10jul02l.pdf. Ac-cessed April 19, 2012.
Gill, S., P. Shrewsbury, and J. Davidson. 2011. Japanese maple scale (Lopholeucaspis japonica): A pest of nursery and landscape trees and shrubs. Uni-versity of Maryland Extension fact sheet.
Hagan, A.K. 2001. Magnolia diseases. 59:231-235. In: R.K. Jones and D.M. Ben-son (Eds.). Diseases of woody ornamentals and trees in nurseries. APS Press, St. Paul, MN.
Hagan, A. 2005. Nematode pests of annual and perennial flowers, herbs, woody shrubs, and trees. ANR 689. Alabama Cooperative Extension System, Auburn, AL.
Hamon, A.B. and T.R. Fasulo. 2007. False oleander scale, Pseudalacaspis cocker-elli (Cooley) (Homoptera: Diaspididae). Florida Cooperative Extension Service, IFAS, University of Florida, EENY-149.
Harman, S.W. 1931. The cranberry rootworm as an apple pest. Journal of Economic Entomology 24:180-182.
Harris, R., J. Clark, and N. Matheny. 2004. Arboriculture: integrated management of landscape trees, shrubs and vines, 4th ed. Prentice Hall, Upper Saddle River, NJ.
Hartman, J., T. Pirone, and M.A. Sall. Pirone’s tree maintenance, 7th ed. 2000. Ox-ford University Press, New York, NY.
Heinrichs, E.A., E.E. Burgess, and E.L. Matheny, Jr. 1973. Control of leaf-feeding insects on yellow poplar. Journal of Economic Entomology 66:1240-1241.
Held, D.W. 2004. Relative susceptibility of woody landscape plants to Japanese bee-tle (Coleoptera: Scarabaeidae). Journal of Arboriculture 30:328-335.
Herrick, G.W. 1931. The magnolia scale (Neolecanium cornuparvum Thro). An-nals of the Entomological Society of America 24:302-305.
Hoffmann, C.H. 1941. Biological observations on Xylosandrus germanus (Bldfd). Journal of Economic Entomology 34:38-42.
Hoover, G.A. 2006. Tuliptree scale. Entomological Notes. Penn State Cooperative Extension. http://ento.psu.edu/extension/factsheets/pdf/tuliptreeScale.pdf. Accessed April 19, 2012.
Hubbard, J.L. and D.A. Potter. 2005. Life history and natural enemy associations of calico scale, Eulecanium cerasorum (Homoptera: Coccidae), in Kentucky. Journal of Economic Entomology 98:1202-1212.
Hubbard, J.L. and D.A. Potter. 2006. Managing calico scale (Hemiptera: Coccidae) infestations on landscape trees. Arboriculture and Urban Forestry 32:138-147.Johnson, W.T. and H.H. Lyon. 1988. Insects that feed on trees and shrubs, 2nd ed. Comstock University Press, Ithaca, NY.
202

Kamikado, T., C. Chang, S. Murakoshi, A. Sakurai, and S. Tamura. 1975. Isolation and structure elucidation of growth inhibitors on silkworm larvae from Magnolia kobus DC. Agricultural and Biological Chemistry 39:833-836.
Knox, G.W. 2012. Commercial nursery production of Magnoliaceae in the southern United States. Proceedings of the 2nd International Symposium on the Family Mag-noliaceae (ISFM): 262-269. May 5-8, 2009. Guangzhou, China.
Krischik, V. and J. Davidson. 2007. IPM (integrated pest management) of Midwest landscapes. University of Minnesota Extension Service, Center for Urban Ecology and Sustainability. http://www.entomology.umn.edu/cues/ipmbook.htm. Accessed April 19, 2012.
Leibee, G.L. 1992. Unearthing a magnolia menace. American Nurseryman 173:70-77.
Leibee, G.L. and K.E. Savage. 1994. Insecticidal control of magnolia white scale (false oleander scale) (Homoptera: Diaspididae) on Magnolia. Proceedings of the Florida State Horticural Society 107:226-228.
Li, G.-H., L.-Z. Dang, L.-J. Hong, L.-J. Zheng, F.-F. Liu, L. Li, Y.-J. Liu, and K.-Q. Zhang. 2009. Nematicidal activity of honokiol and magnolol isolated from Magno-lia tripetala. Journal of Phytopathology 157:390-392.
Mazzey, K. and M. Masiuk. 2011. Magnolia scale. Woody ornamental IPM fact-sheet. Penn State University Cooperative Extension http://woodypests.cas.psu.edu/factsheets/InsectFactSheets/pdf/MagnoliaScale.pdf. Accessed April 19, 2012.
Miller, J.W. 1976. Bacterial leaf spot on saucer magnolia. Florida Department of Agriculture and Consumer Services Plant Pathology circular 163.
Miller, D.R. and J.A. Davidson. 2005. Armored scale insect pests of trees and shrubs. Cornell University Press, Ithaca, NY.
Mitchell, P.L. and F.L. Mitchell. 1983. Range extension of Leptoglossus fluvicornis with observations on egg parasitism. The Southwestern Entomologist 8:150-153.
Moon. D.C. and P. Stiling. 2004. The influence of a salinity and nutrient gradient on coastal vs. upland tritrophic complexes. Ecology 85:2709-2716.
Mullen, J.M. and G.S. Cobb. 1984. Leaf spot of southern magnolia caused by Pseu-domonas cichorii. Plant Disease 68:1013-1015.
Nitao, J.K., M.G. Nair, D.L. Thorogood, K.S. Johnson, and J.M. Scriber. 1991. Bio-active neolignans from the leaves of Magnolia virginiana. Phytochemistry 30:2193-2195.
Oliver, A.D. and J.B. Chapin. 1980. The cranberry rootworm: Adult seasonal his-tory and factors affecting its status as a pest of woody ornamentals in Louisiana. Journal of Economic Entomology 73:96-100.
Oliver, J.B. and C.M. Mannion. 2001. Ambrosia beetle (Coleoptera: Scolytidae) species attacking chestnut and captured in ethanol-baited traps in Middle Tennes-see. Environmental Entomology 30:909-918.
Outcalt, K.W. 1990. Magnolia grandiflora L: Southern magnolia. In: Silvics of North America, USDA Forestry Service Agricultural handbook 654, Vol. 2 Hard-woods, Washington, D.C. http://www.na.fs.fed.us/pubs/silvics_manual/volume_2/magnolia/grandiflora.htm. Accessed April 19, 2012.
Pataky, N.R. 1997. Verticillium wilt disease. University of Illinois Extension Plant Disease RPD No. 1010.
Peterson, N.C. and D.R. Smitley. 1991. Susceptibility of selected shade and flower-ing trees to gypsy moth (Lepidoptera: Lymantriidae). Journal of Economic Entomol-ogy 84:587-592.
Plant & Supply Locator. 2011. Plant & Supply Locator, Hutchinson Publishing Cor-poration, Taylors, SC. www.plantlocator.net. Accessed April 19, 2012.
Polavarapu, S. 1999. Insecticidal nematodes for cranberry pest management, pp. 79-90. Proceedings at the National Workshop on Optimal Use of Insecticidal Nema-todes in Pest Management. Polavarapu, S. (ed). Rutgers University, Chatsworth, NJ.
Popenoe, J. 2008. Magnolia, Southern magnolia Magnolia grandiflora. University of Florida F-IFAS Extension. http://cfextension.ifas.ufl.edu/documents/Magnolia.pdf. Accessed April 19, 2012.
Ranger, C.M., M.E. Reding, K.J.K. Gandhi, J.B. Oliver, P.B. Schultz, L. Caňas, and D.A. Herms. 2011. Species dependent influence of (--)-α-Pinene on attraction of ambrosia beetles (Coleoptera: Curculionidae: Scolytinae) to ethanol-baited traps in nursery agroecosystems. Journal of Economic Entomology 104:574-579.
203

Reding, M., J. Oliver, P. Schultz, and C. Ranger. 2010. Monitoring flight activity of ambrosia beetles in ornamental nurseries with ethanol baited traps: influence of trap height on captures. Journal of Environmental Horticulture 28:85-90.
Relf, D. and B. Appleton. 2009. Managing winter injury to trees and shrubs. Vir-ginia Tech Extension publication 426-500. http://pubs.ext.vt.edu/426/426-500/426-500.html. Accessed April 19, 2012.
Santamour, F.S., Jr. and J.M. Batzli. 1990. Host checklist of root-knot nematodes on broad-leaved landscape trees. Journal of Arboriculture 16:162-168.
Santamour, F.S., Jr. and L.G.H. Riedel. 1993. Susceptibility of various landscape trees to root-knot nematodes. Journal of Arboriculture 18:257-259.
Sinclair, W.A. and H. Lyon. 2005. Diseases of trees and shrubs, 2nd ed. Cornell Uni-versity Press, Ithaca, NY.
Smith, H. 2010. Leafminer pests of Connecticut nurseries. Connecticut Agricultural Experiment Station. http://www.ct.gov/caes/lib/caes/documents/publications/fact_sheets/entomology/leafminer_pests_of_connecticut_nurseries.pdf. Accessed April 19, 2012.
Tippins, H.H. 1968. Observations on Phenacaspis [= Pseudacaulaspis] cockerelli (Cooley) (Homoptera: Diaspididae), a pest of ornamtnal plants in Georgia. Journal of the Georgia Entomological Society 3:13-15.
Tyler-Julian, K., O. Demirozer, R. Figlar, T. Skarlinsky, J. Funderburk, and G. Knox. 2012. New host records for Caliothrips striatus Hood (Thysanoptera: Thripi-dae). Florida Entomologist. In press.
Townsend, L. 2005. Magnolia scale. University of Kentucky Cooperative Extension Service Entfact-431.
U.S. Department of Agriculture. 2009. 2007 Census of Agriculture, Washington, DC.
U.S. Department of Agriculture, Forest Service. 1985. Insects of eastern forests. A. T. Drooz (ed.). U.S. Department of Agriculture, miscellaneous publication 1426. Washington, DC. 608 p.
van Tol, R.W.H.M. and M.J. Raupp. 2005. Nursery and tree application. In: Grewal, P.S., R.-U. Ehlers, and D.I. Shapiro-Ilan (eds.), Nematodes as biocontrol agents. CAB Intl. London, UK.
Vann, S. and J. Robbins. 2008. Sapsucker damage on woody plants. University of Arkansas Extension publication FSA7561. http://www.uaex.edu/Other_Areas/publications/PDF/FSA-7561.pdf. Accessed April 19, 2012.
Vanek, S.J. and D.A. Potter. 2009. Host plant resistance among Magnolia spp. and Quercus spp. to soft scale pests. UK Nursery and Landscape Research Report - 2009. UK PR 602. http://www.ca.uky.edu/agc/pubs/pr/pr602/pr602.pdf. Accessed April 19, 2012.
Vanek, S.J. and D.A. Potter. 2010. Ant-exclusion to promote biological control of soft scales (Hemiptera: Coccidae) on woody landscape plants. Environmental Ento-mology 39:1829-1837.
Weber, B.C. and J.E. McPherson. 1983. Life-history of the ambrosia beetle Xylosan-drus germanus (Coleoptera, Scolytidae). Annals of the Entomological Society of America 76:455-462.
Wilson, T.H. 1975. A monograph of the subfamily Panchaetothripinae (Thysanop-tera: Thripidae). Memoirs of the American Entomological Institute 23:1-354.
Wood, S.L. 1982. The bark and ambrosia beetles of North and Central America (Co-leoptera: Scolytidae), a taxonomic monograph. Great Basin Naturalist Memoirs no. 6.
204

SECTION 6
TablesTable 9.1. Number of production nurseries growing various magnolia species reported1 for twelve southern states.
1 Plant & Supply Locator (2011)
205
Species Number of nurseries
Magnolia ×soulangeana and other hybrids not derived from Magnolia acuminata or Magnolia stellata 16
Magnolia acuminata and hybrids 5
Magnolia ashei 2
Magnolia figo 6
Magnolia grandiflora 138
Magnolia macrophylla 1
Magnolia maudiae 1
Magnolia stellata and hybrids 47
Magnolia tripetala 1
Magnolia virginiana (includes var. australis and var. virginiana) 65
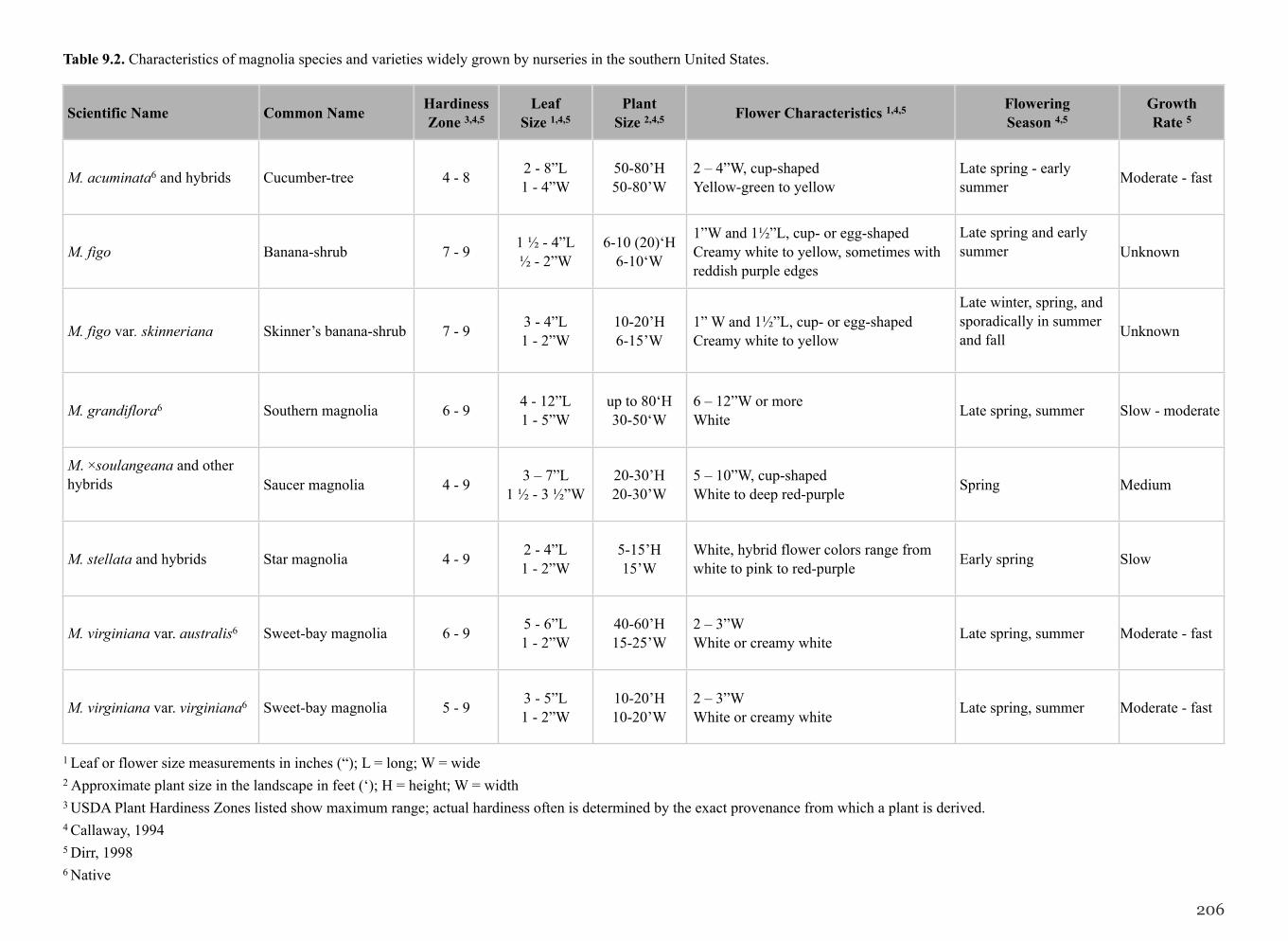
Table 9.2. Characteristics of magnolia species and varieties widely grown by nurseries in the southern United States.
1 Leaf or flower size measurements in inches (“); L = long; W = wide2 Approximate plant size in the landscape in feet (‘); H = height; W = width3 USDA Plant Hardiness Zones listed show maximum range; actual hardiness often is determined by the exact provenance from which a plant is derived. 4 Callaway, 19945 Dirr, 19986 Native
206
Scientific Name Common Name Hardiness Zone 3,4,5
LeafSize 1,4,5
PlantSize 2,4,5 Flower Characteristics 1,4,5 Flowering
Season 4,5GrowthRate 5
M. acuminata6 and hybrids Cucumber-tree 4 - 8 2 - 8”L1 - 4”W
50-80’H50-80’W
2 – 4”W, cup-shapedYellow-green to yellow
Late spring - early summer Moderate - fast
M. figo Banana-shrub 7 - 9 1 ½ - 4”L½ - 2”W
6-10 (20)‘H6-10‘W
1”W and 1½”L, cup- or egg-shapedCreamy white to yellow, sometimes with reddish purple edges
Late spring and early summer Unknown
M. figo var. skinneriana Skinner’s banana-shrub 7 - 9 3 - 4”L1 - 2”W
10-20’H6-15’W
1” W and 1½”L, cup- or egg-shapedCreamy white to yellow
Late winter, spring, and sporadically in summer and fall Unknown
M. grandiflora6 Southern magnolia 6 - 9 4 - 12”L1 - 5”W
up to 80‘H30-50‘W
6 – 12”W or moreWhite Late spring, summer Slow - moderate
M. ×soulangeana and other hybrids Saucer magnolia 4 - 9 3 – 7”L
1 ½ - 3 ½”W20-30’H20-30’W
5 – 10”W, cup-shapedWhite to deep red-purple Spring Medium
M. stellata and hybrids Star magnolia 4 - 9 2 - 4”L1 - 2”W
5-15’H15’W
White, hybrid flower colors range from white to pink to red-purple Early spring Slow
M. virginiana var. australis6 Sweet-bay magnolia 6 - 9 5 - 6”L1 - 2”W
40-60’H15-25’W
2 – 3”WWhite or creamy white Late spring, summer Moderate - fast
M. virginiana var. virginiana6 Sweet-bay magnolia 5 - 9 3 - 5”L1 - 2”W
10-20’H10-20’W
2 – 3”WWhite or creamy white Late spring, summer Moderate - fast

Table 9.3. Pest-directed insecticidal activity and Insecticide Resistance Action Committee (IRAC) codes for use in developing a pesticide rotation plan to manage key pests of magnolia1. Check current products for labeled pesticides, sites for control, and plant2 and pest species.
207
Active Ingredient IRACCode
Foliar Feeders & Leaf Miners
Soft Scales
Armored Scales1
WoodBorers Thrips
abamectin 6 - - - - Y
acephate 1B Y Y Y - Y
acetamiprid 4A Y Y Y - Y
azadirachtin unk Y Y Y - Y
Bacillus thuringiensis 11 Y - - - -
Beauveria bassiana: BotaniGard nr Y Y Y - Y
bifenthrin 3A Y Y Y Y Y
buprofezin 16 - Y Y - -
carbaryl 1A Y Y Y - Y
chlorpyrifos 1B Y Y Y Y Y
clothianidin: Arena 4A Y - - - -
cyfluthrin: Decathlon 3A Y Y Y - Y
cyfluthrin + imidacloprid 3+4A - Y Y - Y
Diazinon3 1B Y Y Y - Y
dimethoate 1B Y Y Y - Y
dinotefuran 4A - Y Y - Y
esfenvalerate 3A Y Y Y - Y
flonicamid PC - Y Y - Y
horticultural oil nr Y Y Y - Y

Table 9.3. (continued) Pest-directed insecticidal activity and Insecticide Resistance Action Committee (IRAC) codes for use in developing a pesticide rotation plan to manage key pests of mag-nolia1. Check current products for labeled pesticides, sites for control, and plant2 and pest species.
nr = not required to have an IRAC codeY = a product with this active ingredient is labeled for use on the pest indicated; - = Not available for use 1 Adapted from Knox, Gary W., William E. Klingeman, Mathews L. Paret and Amy F. Fulcher. Management of Pests, Plant Diseases and Abiotic Disorders of Magnolia Species in the South-
eastern U.S.: A Review. Journal of Environmental Horticulture: In review.2 Check labels carefully to ascertain if any ornamental phytotoxity has been reported and test on a small portion of ornamental plants before spraying the entire nursery crop or range.3 Diazinon is no longer available in the market.
208
Active Ingredient IRACCode
Foliar Feeders & Leaf Miners
Soft Scales
Armored Scales1
WoodBorers Thrips
imidacloprid 4A - Y Y - Y
insecticidal soap nr Y Y Y - Y
kinoprene 7A - Y Y - Y
lambda-cyhalothrin 3A Y Y Y - Y
malathion 1B - Y Y - Y
methiocarb 1A - Y Y - Y
neem oil extract 18B - Y Y - Y
novaluron 15 Y - - - Y
permethrin 3A Y - - Y Y
pyriproxifen 7C - Y Y - -
spinosad 5 Y Y Y - Y
tau-fluvalinate 3A Y Y Y - Y
thiamethoxam 4A - Y Y - Y

Table 9.4. Labeled fungicidal active ingredients and Fungicide Resistance Action Committee (FRAC) Codes to help pest managers integrate plant disease control within a pesticide rotation plan for magnolias1. Efficacy and phytotoxicity data available are largely unavailable for magnolia species and cultivars magnolia2. Check current products for labeled pesticides, sites for con-trol, and plant and pest species.
209
Active IngredientFRAC Code
Plant Pathogen(s) Managed3
azoxystrobin 11 Anthracnose; powdery mildew
Bacillus subtilis strain QST 713 44 Anthracnose; powdery mildew
calcium polysulfides n.l. Powdery mildew
chloropicrin 8B Phytophthora
chlorothalonil + thiophanate methyl: Spectro 90 WDG M5 + 1 Anthracnose; powdery mildew
clove oil + rosemary oil + thyme oil: Sporatec n.l. Anthracnose; powdery mildew
copper M1 Bacterial spot; syringae leaf spot; anthracnose; powdery mildew
corn oil + cotton seed oil + garlic oil: Mildew Cure n.l. Powdery mildew
hydrogen dioxide n.l. Phytophthora
mancozeb M3 Anthracnose
metconazole 3 Anthracnose; powdery mildew
myclobutanil 3 Anthracnose; powdery mildew
neem oil: Trilogy n.l. Anthracnose; powdery mildew
polyoxin D zinc salt 19 Anthracnose; powdery mildew
potassium bicarbonate n.l. Anthracnose; powdery mildew
potassium phosphite n.l. Anthracnose; powdery mildew

Table 9.4. (continued) Labeled fungicidal active ingredients and Fungicide Resistance Action Committee (FRAC) Codes to help pest managers integrate plant disease control within a pesticide rotation plan for magnolias1. Efficacy and phytotoxicity data available are largely unavailable for magnolia species and cultivars magnolia2. Check current products for labeled pesticides, sites for control, and plant and pest species.
n.l. = FRAC code not listed; not required to have a FRAC code, or mode of action unknown1 Adapted from Knox, Gary W., William E. Klingeman, III, Mathews L. Paret and Amy F. Fulcher. Management of Pests, Plant Diseases and Abiotic Disorders of Magnolia Species in the Southeastern U.S.: A Review. Journal of Environmental Horticulture: In review.2 Check labels carefully to ascertain if any ornamental phytotoxity has been reported and test on a small portion of ornamental plants before spraying the entire nursery crop or range.3 Plant disease management information for magnolia derived from the ornamentals category within the CDMS Database (Engelbrecht, 2005; CDMS, 2011).4 "M" category modes of action have multi-site contact activity5 "P"= acts to induce host plant's defense mechanisms
210
Active IngredientFRAC Code
Plant Pathogen(s) Managed3
potassium phosphate n.l. Powdery mildew
potassium phosphate + sulfur: Sanction n.l. + M2 4 Powdery mildew
potassium silicate n.l. Powdery mildew
propiconazole 3 Anthracnose; powdery mildew
Reynoutria sachalinensis extract: Regalia P 5 Anthracnose; powdery mildew
sulfur M2 Powdery mildew
tebuconazole 3 Anthracnose; powdery mildew
thiophanate methyl 1 Anthracnose; powdery mildew
triadimefon 3 Powdery mildew
trifloxystrobin 11 Anthracnose; powdery mildew