catheter-associated urinary tract infection by...
TRANSCRIPT

1
Catheter-associated urinary tract infection by 1 Pseudomonas aeruginosa is mediated by 2 exopolysaccharide independent biofilms 3 4 Stephanie J. Cole*, Angela R. Records*, Mona W. Orr, Sara B. Linden, 5 Vincent T. Lee 6 7 Department of Cell Biology and Molecular Genetics, Maryland Pathogen 8 Research Institute, University of Maryland at College Park, College Park, MD, 9 USA 10 11 *Authors contributed equally to this work 12 13 For correspondence: [email protected] 14 15 16 17 Short title: Pseudomonas catheter-associated urinary tract infection 18 19 20 21 22
IAI Accepts, published online ahead of print on 3 March 2014Infect. Immun. doi:10.1128/IAI.01652-14Copyright © 2014, American Society for Microbiology. All Rights Reserved.
on June 25, 2018 by guesthttp://iai.asm
.org/D
ownloaded from

2
ABSTRACT (248 words) 23 Pseudomonas aeruginosa is an opportunistic human pathogen that is especially 24 adept at forming surface-associated biofilms. P. aeruginosa causes catheter-associated 25 urinary tract infections (CAUTI) through biofilm formation on the surface of indwelling 26 catheters. P. aeruginosa encodes for three extracellular polysaccharides, PEL, PSL and 27 alginate, and utilizes PEL and PSL polysaccharides to form biofilms in vitro; however, 28 the requirement of these polysaccharides during in vivo infections is not well understood. 29 Here we show in a murine model of CAUTI that PAO1, a strain harboring both pel and 30 psl genes, and PA14, a strain harboring only pel genes, form biofilms on the implanted 31 catheters. To determine the requirement of exopolysaccharide during in vivo biofilm 32 infections, we tested isogenic mutants lacking the pel, psl, and alg operons and showed 33 that PA14 mutants lacking these operons can successfully form biofilms on catheters in 34 the CAUTI model. To determine the host factor(s) that induce the ∆pelD mutant to form 35 biofilm, we tested mouse, human and artificial urine and show that urine can induce 36 biofilm formation by the PA14 ∆pelD mutant. By testing the major constituents of urine, 37 we show that urea can induce a pel-, psl-, and alg-independent biofilm. These pel-, psl-, 38 and alg-independent biofilms are mediated by the release of extracellular DNA. 39 Treatment of biofilms formed in urea with DNase I reduced the biofilm indicating 40 extracellular DNA supports biofilm formation. Our results indicate that the opportunistic 41 pathogen P. aeruginosa utilizes a distinct program to form biofilms that are independent 42 of exopolysaccharides during CAUTI. 43 44
on June 25, 2018 by guesthttp://iai.asm
.org/D
ownloaded from

3
INTRODUCTION 45 Pseudomonas aeruginosa causes biofilm-mediated infections including catheter-46 associated urinary tract infection (CAUTI), ventilator-associated pneumonia, infections 47 related to mechanical heart valves, stents, grafts, sutures, and contact lens-associated 48 corneal infections (1, 2). P. aeruginosa is responsible for 12% of all nosocomial urinary 49 tract infections (UTI), making it the third most common organism after E. coli and 50 enterococci isolated from UTI patients in the hospital setting (3). Nosocomial UTIs are 51 catheter-associated, and the development of bacteriuria is directly related to the duration 52 of catheterization (4, 5). Between 15 and 25% of hospital patients are catheterized for two 53 to four days during their stays, while many nursing home patients remain catheterized for 54 months or years (5, 6). Catheter-associated bacteriuria leads to increased length of 55 hospital stay, causing an estimated 900,000 additional hospital days per year (5); and 56 complications associated with nosocomial UTIs cause or contribute to an estimated 7,500 57 deaths per year (7). Biofilms are a serious problem as they are often refractory to 58 antibiotic therapy. Antibiotic therapy can eliminate planktonic bacteria, but bacteria 59 within biofilms survive antibiotic treatment (8-10). When antibiotic treatment ends, the 60 biofilm can again shed planktonic cells resulting in recurrent acute infection. This cycle 61 of infection is difficult to stop and often requires the removal of the contaminated device 62 to eliminate the bacterial biofilm (9, 11). However, removing the contaminated device is 63 only a temporary solution as replacement with a new device yet again provides a surface 64 for biofilm formation. Using a suitable animal model to investigate the bacterial factors 65 contributing to chronic infections will provide insights and potential novel target for 66 therapeutic intervention. 67
on June 25, 2018 by guesthttp://iai.asm
.org/D
ownloaded from

4
P. aeruginosa biofilms consist of an extracellular matrix that includes 68 polysaccharides, proteinaceous components and extracellular DNA (eDNA) (12-16). 69 Non-mucoid strains of P. aeruginosa produce biofilms that are independent of alginate 70 biosynthesis (17, 18). These biofilms can colonize solid surfaces and form mushroom 71 structures in flow cells (19), rings in culture tubes and microtiter plates (20), or pellicles 72 at the air-liquid interface (21). The main polysaccharide components include the PEL and 73 PSL exopolysaccharides produced by the proteins encoded by the pel and psl genes, 74 respectively (21-23). Mutations in either pel or psl genes result in bacteria that produce 75 less biofilm (11, 24, 25). The genome of P. aeruginosa strain PA14 lacks pslA-C genes 76 and does not produce the PSL polysaccharide. A PA14 pel operon mutant cannot produce 77 the PEL polysaccharide and fails to form biofilm (21). In addition to the requirement of 78 the pel and psl operons, the requirement of eDNA has been demonstrated by the ability of 79 DNase I to reduce biofilm formation (13). Numerous other studies have demonstrated the 80 participation of eDNA in P. aeruginosa biofilms (26, 27). Despite having identified a 81 number of pseudomonal factors that contribute to biofilm formation in vitro, little is 82 known about the contribution of each of these biofilm components during an animal 83 model of CAUTI. 84 The relevance of biofilm genes identified from in vitro systems during animal 85 infections has not been sufficiently studied because few models for chronic infections 86 exist. One model system involves inserting an implant or catheter containing bacterial 87 biofilm into either the lung (28, 29) or bladder of an animal to mimic catheter induced 88 chronic pneumonia or urinary tract infection (UTI) (30, 31), respectively. An advantage 89 of the UTI model is that the biofilm formation and disseminating infection events can be 90
on June 25, 2018 by guesthttp://iai.asm
.org/D
ownloaded from

5
separated. Animals infected either with a catheter containing pre-formed biofilm or 91 inoculated after the implantation of a sterile catheter yielded biofilm-based chronic 92 infections (30). Here we use the murine model of CAUTI to test the contribution of 93 extracellular polysaccharides to pseudomonal biofilm-mediated infection. We employed 94 P. aeruginosa strains PAO1 and PA14 and their isogenic mutants lacking the pel, psl and 95 alg operons encoding the biosynthetic genes for the PEL and PSL exopolysaccharides. 96 Using these strains, we show that both PEL, PSL and alginate exopolysaccharides are 97 dispensable for biofilm formation on the catheter and for further dissemination into the 98 kidneys. The PA14 ∆pelD mutant can also form biofilm on the catheter during mixed 99 infections with the parental PA14 strain. The PA14 ∆pelD mutant participates in in vitro 100 mixed biofilms with PA14 indicating that the ∆pelD mutant can utilize the PEL 101 polysaccharide produced by PA14. The ability of exopolysaccharide-deficient 102 pseudomonas to form biofilm can be induced in vitro by mouse and human urine as well 103 as urea, a major component of urine. Exposure of P. aeruginosa to urea induces a 104 subpopulation of cells to round and lyse. The released eDNA can mediate biofilm 105 formation of both PA14 and the ∆pelD mutant strains. Addition of DNase I degrades 106 eDNA and reduces biofilm formation of both PA14 and ∆pelD in the presence of urea. 107 Together, these data demonstrate that P. aeruginosa has a mechanism independent of 108 exopolysaccharides to mediate biofilm formation on catheters during CAUTI. 109
110
on June 25, 2018 by guesthttp://iai.asm
.org/D
ownloaded from

6
MATERIALS AND METHODS 111 Bacterial strains and culture conditions 112 Strains and plasmids used in this study are listed in Table S1. Strains were grown 113 in LB broth. PA14-lux and PA14 ∆pelD-lux were constructed by mating recombination 114 of the pCTX-lac-lux into the att site followed by Flp based recombination to remove the 115 residual pCTX vector sequence (32-34). Plasmid pMMB-PA3702 was maintained in 15 116 µg ml-1 gentamycin, and 1 mM isopropylthiogalactose (IPTG) was used to induce gene 117 expression. 118 119 In vitro catheter biofilm assay 120
One-centimeter-long pieces of PE50 catheter material (polyethylene; outside 121 diameter, 0.965 mm; inside diameter, 0.58 mm; Braintree Scientific, Inc.) were sterilized 122 in 70% ethanol and allowed to air dry. Catheters were filled from end-to-end with 123 bacterial suspensions (109 CFU/ml) in either PBS with 1% tryptone (PBS-T); a 1:1 124 mixture of PBS-T and mouse urine; a mixture containing 10% PBS-T and 90% mouse 125 urine; or PBS-T supplemented with 165 mM urea or 330 mM urea. Catheters were placed 126 in sterile Petri plates and incubated in a humid chamber for 1-16 hours. After incubation, 127 liquid was wicked from tubing via capillary action on a paper towel. The biofilms were 128 stained with crystal violet and the unbound dye was washed away. The crystal violet was 129 dissolved in 30% acetic acid and quantitated spectrophotometrically by reading 130 absorbance 595 nm. Data was analyzed using Prism Graphpad software with statistical 131 significance determined using Student’s t-test, P < 0.05. 132 133
on June 25, 2018 by guesthttp://iai.asm
.org/D
ownloaded from

7
Murine model of CAUTI 134 The CAUTI mouse model established by Kadurugamuwa et al. (30) was modified 135
and is outlined below. All experimental animal procedures were performed in accordance 136 with protocols approved by IACUC at University of Maryland at College Park. Segments 137 of PE50 catheter material (polyethylene; length, either 1 or 5 cm; outside diameter, 0.965 138 mm; inside diameter, 0.58 mm; Braintree Scientific, Inc.) were sterilized in 70% ethanol 139 and allowed to air dry. CF-1 female mice (Charles River, Wilmington, Mass) weighing 140 26-35 g were anesthetized with ketamine (100 mg/kg) and xylazine (10 mg/kg) and 141 placed on their backs. The periurethral areas were cleansed with 70% ethanol. For each 142 mouse, a 1-cm catheter piece was loaded onto a sterile 7-cm metal stylet followed by a 5-143 cm piece of tubing. The stylet was placed in the urethral opening, and the tubing was 144 advanced over the stylet, into the bladder until the 1-cm catheter piece was placed into 145 the bladder. The metal stylet was removed, allowing the 5-cm tubing to serve as a conduit 146 for inoculation of the bladder. Using an insulin syringe, 35 µl of the designated bacterial 147 suspension was injected transurethrally, resulting in a final inoculum concentration of 106 148 CFU. The tubing was subsequently removed, while the 1-cm catheter was allowed to 149 remain in the bladder. Data was analyzed using Prism Graphpad software with statistical 150 significance determined using one-way ANOVA, P < 0.05. 151 152 Collection of human urine 153
Urine was collected from volunteers in compliance with the Institutional Review 154 Board at University of Maryland, College Park. No personal identifiable information was 155
on June 25, 2018 by guesthttp://iai.asm
.org/D
ownloaded from

8
collected about participants at the time of urine collection. Urine samples were 156 immediately sterile filtered with 0.22 µM filter and frozen at -20˚C until use. 157 158 Scanning Electron Microscopy 159
Samples were obtained either from bacteria grown in vitro in the catheter or from 160 catheters dissected from infected animals in vivo. In vitro, bacteria were grown in 161 catheters with LB broth with and without 0.5 M urea and incubated for 2 days at 30˚C. In 162 vivo samples, including bladders and catheters, were dissected from animals infected with 163 the indicated strains. All catheters and bladders were washed with sterile PBS and fixed 164 with 2% glutaraldehyde in PBS overnight at 4˚C, post-fixed in 1% OsO4 in PBS and 165 dehydrated through an ethanol series. The samples were critical point dried, mounted on 166 stubs and evaporative-coated with gold/palladium alloy. Stubs were mounted in Hitachi 167 S-4700 SEM to obtain micrographs. 168 169 Fluorescence Microscopy 170
PA14 wild-type and PA14 ∆pelD were grown overnight in LB. The overnight 171 culture was diluted 1:100 into LB and 2 μL were transferred onto a sterile coverslip and 172 covered with LB-1% agarose pads containing 5 mM MgCl2, 0.001 mg/mL of propidium 173 iodide and either 0 M urea, 0.5 M urea, or 0.1% SDS. Bacteria were incubated for 8 hrs 174 in a humidified chamber and imaged using a Zeiss Axio Observer.Z1 inverted fluorescent 175 microscope. Phase contrast was used to image bacteria and eDNA was visualized via 176 propidium iodide staining. 177 178
on June 25, 2018 by guesthttp://iai.asm
.org/D
ownloaded from

9
RESULTS 179 PEL, PSL and alginate polysaccharides are not required in a murine model of 180 CAUTI 181
P. aeruginosa encodes three exopolysaccharide biosynthesis operons, pel, psl and 182 alg operons, that produce PEL, PSL and alginate, respectively. Biofilms formed in vitro 183 require two extracellular polysaccharides, PEL and PSL, synthesized by (21-23, 26). We 184 hypothesized that one or more of the PSL, PEL and alginate exopolysaccharides are 185 required for biofilm-mediated CAUTI in vivo. To test this hypothesis, we adapted a 186 mouse model of urinary tract infection utilized in previous studies (30, 35). Mice were 187 catheterized with 5-mm pieces of catheter tubing and subjected to transurethral challenge 188 with P. aeruginosa. Mice were sacrificed at 14 days post-inoculation, and the bladder, 189 kidneys, liver, and spleen of each mouse were harvested and homogenized. The 190 homogenates were plated for determination of colony forming units (CFUs) on both LB 191 agar and LB agar supplemented with triclosan, which is selective for Pseudomonas. To 192 confirm that the catheterization process itself did not negatively affect the mice, several 193 mice were catheterized in the absence of bacteria. The organs from these mice were 194 visually healthy and were free of bacteria (data not shown). Importantly, neither PA14 195 nor the PA14 ∆pelD strain was able to colonize mouse tissue in the absence of a catheter 196 after two weeks as determined by CFU counts (data not shown). This underscores the 197 importance of the catheter as a surface for initial colonization by the bacteria. To 198 determine the requirement for PEL, PSL and alginate polysaccharides, mice were 199 implanted with catheters and challenged with 106 CFU of PA14, PA14 ∆pelD, PA14 200 ∆pelA-G or PA14 ∆pelA-G ∆algD-A. No non-pseudomonads were recovered at the end 201
on June 25, 2018 by guesthttp://iai.asm
.org/D
ownloaded from

10
of the two-week CAUTI. The majority of animals exhibited chronic colonization by P. 202 aeruginosa in the bladder after inoculation with PA14 (9/10 animals), PA14 ∆pelD 203 (12/13 animals), PA14 ∆pelA-G (12/12 animals), or PA14 ∆pelA-G ∆algD-A (7/8 204 animals) (Fig. 1). Furthermore, the organs harvested from mice inoculated with the PA14 205 ∆pelD, PA14 ∆pelA-G and PA14 ∆pelA-G ∆algD-A strains harbored bacterial numbers 206 similar to mice infected with PA14 parental strain (Fig. 1A). To confirm these results for 207 the PA14 strain, we also tested another commonly used laboratory strain PAO1 and its 208 isogenic ∆pel ∆psl ∆alg mutant lacking the exopolysaccharides. Both PAO1 (12/12 209 animals) and PAO1 ∆pel ∆psl ∆alg (6/10 animals) colonized the bladder in the murine 210 model of CAUTI, but the average bacterial load of PAO1 ∆pel ∆psl ∆alg was less than 211 that of the parental PAO1 strain (Fig. S1A). Nonetheless, the PAO1 ∆pel ∆psl ∆alg was 212 able to colonize the bladder indicating that the exopolysaccharides are not absolutely 213 required for biofilm formation in vivo. Of note, all of the kidneys with P. aeruginosa 214 infections were from animals that also had bacteria in the bladder; this suggests that the 215 kidney infections were the result of bacterial dissemination from the bladder. Only a few 216 animals in each inoculation group had detectible CFU in the spleen and liver (Figs. S1B 217 and S1C), indicating that the murine CAUTI is a localized and chronic model of P. 218 aeruginosa infection that is restricted primarily to the urinary tract. These results 219 demonstrate P. aeruginosa lacking the exopolysaccharide genes are able to colonize the 220 bladder of mice and indicate that P. aeruginosa establishes CAUTI independent of all 221 three exopolysaccharides. 222 223
on June 25, 2018 by guesthttp://iai.asm
.org/D
ownloaded from

11
Scanning electron microscopy (SEM) of catheters reveals biofilm formation 224 independent of extracellular polysaccharide genes in vivo 225
We asked whether the bacteria within the infected bladder were located on the 226 catheter, the bladder epithelium or both surfaces. To test this, bladders and associated 227 catheters were removed from infected mice, sectioned and processed for SEM. The 228 interior of the catheter isolated from mice infected with PAO1 had visible bacteria that 229 formed elongated rods many of which were septated in the interior surface of the catheter 230 (Fig. 2A). We also asked whether the strain PA14 ∆pelD pMMB-PA3702, which lacks 231 both PEL and PSL polysaccharides, can also colonize the interior of implanted catheters. 232 The pMMB-PA3702 was used to elevate the levels of cyclic-di-GMP which enhances the 233 in vitro biofilm phenotype between wild-type and ∆pelD mutant strains (36). We also 234 observed PA14 ∆pelD pMMB-PA3702 in the interior of the catheter embedded in a 235 matrix material (Fig. 2B). This is in contrast to the uniform short rods that P. aeruginosa 236 forms when grown in vitro biofilms (Fig. S2), which suggests that P. aeruginosa 237 responds differently to the in vivo environment. The bacteria were encased in a granular 238 matrix but it is not apparent by SEM whether this matrix is of host or bacteria origin. The 239 external surface of the catheter was devoid of bacteria (Fig. S3C). We performed SEM 240 on the bladder epithelium and were unable to detect bacteria on the surface of the bladder 241 (Fig. S3) indicating that the interior of the catheter is the main reservoir of P. aeruginosa. 242 These results show that the primary reservoir of P. aeruginosa during CAUTI is biofilms 243 on the luminal surface of catheters. For the remainder of this report, PA14 and its 244 isogenic pel mutants were used to study the requirement of PEL and PSL during biofilm 245 formation in vitro and in vivo. 246
on June 25, 2018 by guesthttp://iai.asm
.org/D
ownloaded from

12
247 Competition studies reveal that neither PA14 nor PA14 ∆pelD is at a selective 248 advantage in vivo 249
Our in vivo studies suggest that PelD is dispensable for biofilm formation on 250 catheters in the mouse. One possibility is that the ΔpelD mutant strain occupies a niche 251 that would be otherwise occupied by the parental PA14 strain during CAUTI. We tested 252 this hypothesis by competing parent PA14 and the isogenic ∆pelD strain in the murine 253 model of CAUTI. Modified versions of both PA14 and PA14 ∆pelD were prepared by 254 incorporation of a constitutive lux marker at the chromosomal ctx phage attachment site 255 (att) of each strain. The PA14-lux and PA14 ∆pelD-lux colonies were easily identifiable 256 due to bioluminescence resulting from constitutive expression of the lux gene. Bacteria 257 were mixed in a 1:1 ratio in two different combinations: PA14 + PA14 ∆pelD-lux or 258 PA14 ∆pelD + PA14-lux. Each combination was tested in the murine model of CAUTI in 259 groups of five mice, as described above, with 106 CFU. At 14 days post-inoculation, mice 260 were sacrificed and the bladder and kidneys of each mouse were harvested, homogenized 261 and enumerated by plating serial dilution of the tissue homogenates and counting the 262 numbers of luminescent (Lux+) and non-luminescent (Lux-) colonies. Colony counts from 263 homogenized bladders and kidneys revealed that the competitive index for PA14 ∆pelD-264 lux/PA14 was 0.63±0.42 and for PA14 ∆pelD/PA14-lux was 1.95±1.57 (Fig. 3). These 265 results indicate that there is no competitive disadvantage for PEL and PSL 266 polysaccharide-deficient bacteria in the murine model of CAUTI. Since the ∆pelD mutant 267 can form biofilms on catheters equally well when inoculated alone and during 268 coinfections with PA14, we asked whether the PA14 strain could promote the ability of 269
on June 25, 2018 by guesthttp://iai.asm
.org/D
ownloaded from

13
the ∆pelD strain to participate in mixed biofilms in vitro. Using the same lux-labeled 270 strains in mixed biofilms in vitro, we found that the ∆pelD strain can participate in mixed 271 biofilms in a 1:1 ratio to the parental PA14 strain (Fig. S4). The simplest explanation for 272 these results is that the ∆pelD mutant can utilize a product produced by the parental 273 PA14, presumably the PEL polysaccharide, to participate in the biofilm and suggests that 274 PEL is likely a public good that is shared by pseudomonads in a biofilm. Taken together, 275 these results show that exopolysaccharide-deficient P. aeruginosa can cause CAUTI 276 alone or in combination with wild-type P. aeruginosa. 277 278 PA14 ∆pelD and PA14 ∆pelA-G retain phenotype and genotype after murine model 279 of CAUTI 280
Since pel mutants alone can infect mice in the catheter model of infection, we 281 asked whether this ability is due to a heritable suppressor mutation. To test if genetic 282 mutations caused a permanent change, P. aeruginosa strains were isolated after urinary 283 tract infection with PA14, PA14 ∆pelD, PA14 ∆pelA-G or PA14 ∆pelA-G ∆algD-A. Each 284 of the isolates were tested for their ability to form biofilm in 96 well polystyrene plates 285 and subsequently genotyped for pelD. Of the thirty-six PA14 ∆pelD isolates collected, all 286 isolates remained ∆pelD and were unable to form biofilms. Similarly, sixteen PA14 287 ∆pelA-G mutants, that lack the entire pel operon, were unable to form biofilms after 288 isolation from mouse and lacked the pelD gene (Table 1). Of the twenty-four PA14 289 isolates, all isolates retained the wild-type pelD allele and the ability to form biofilm. 290 Similarly, all 42 of the isolates from PA14 ∆pelA-G ∆algD-A infected mice retained both 291 ∆pelA-G and ∆algD-A mutations. Our results differ from other chronic pseudomonal 292
on June 25, 2018 by guesthttp://iai.asm
.org/D
ownloaded from

14
infections, such as cystic fibrosis infections, in which mutations are known to occur (37). 293 These results suggest that there is not sufficient selective pressure to alter the pel locus, 294 the alg locus or biofilm formation in vivo during the two-week CAUTI. 295 296 PA14 ∆pelD forms biofilm in vitro in the presence of urine 297 Our murine CAUTI studies revealed that infection with PA14 ∆pelD alone 298 resulted in persistent biofilm formation. This suggests that the host environment allows 299 PA14 to bypass the requirement for pelD during biofilm formation in vivo. We reasoned 300 that urine represents one of the major differences between the in vitro biofilm conditions 301 and the mouse model. Therefore, we conducted in vitro catheter studies in the presence of 302 mouse urine. To enhance the difference between the PA14 and PA14 ∆pelD strains, we 303 overexpressed the WspR diguanylate cyclase (pMMB-PA3702) that activates the 304 transcription of the pel operon and the production of the PEL polysaccharide. Catheters 305 were filled with suspensions of either of the two strains in the presence of PBS-T, a 1:1 306 mixture of PBS-T and mouse urine, or a mixture containing 10% PBS-T and 90% mouse 307 urine. The catheters were incubated at 37°C overnight, and bacterial biofilm formation 308 was quantified by the crystal violet assay. PA14 pMMB-PA3702 formed biofilms on 309 catheters in PBS-T (OD595 of 0.17 ± 0.01), while the ∆pelD pMMB-PA3702 strain did 310 not (OD595 of 0.0035 ± 0.001) (Fig. 4A). In the presence of 50% mouse urine, PA14 311 biofilm formation was similar to that in PBS-T (OD595 of 0.177 ± 0.001). However, the 312 OD595 for the ∆pelD strain was 0.023 ± 0.009, a 6.6-fold increase as compared to PA14 313 ∆pelD biofilm formation in the presence of PBS-T alone. PA14 ∆pelD biofilm formation 314 was further increased in the presence of 90% urine (OD595 of 0.075 ± 0.004; a 21.6-fold 315
on June 25, 2018 by guesthttp://iai.asm
.org/D
ownloaded from

15
increase over PBS-T levels). Interestingly, biofilm formation by PA14 decreased in the 316 presence of 90% urine (OD595 of 0.069 ± 0.001; a 2.5-fold decrease from PBS-T levels). 317 Despite this reduction, PA14 still formed biofilm in urea as compared to the PA14 ∆pelD 318 strain in the absence of urea. 319
To determine if the induction of PA14 ∆pelD biofilm development applies to 320 human urine, we conducted a biofilm assay in 1x PBS-T supplemented with 25%, 50% or 321 75% human urine. We performed an in vitro biofilm experiment using PA14 and PA14 322 ∆pelD strains. Similar to the results from mouse urine, human urine enhanced the OD595 323 of PA14 ∆pelD pMMB-PA3702 in 50% (OD595 of 0.20 ± 0.05) and 75% (OD595 of 0.20 ± 324 0.03) human urine compared to the PBS-T control (OD595 of 0.09 ± 0.03) (Fig. 4B). 325 Unlike mouse urine, the addition of human urine enhanced PA14 pMMB-PA3702 biofilm 326 formation. With the addition of either 25% or 50% human urine there was a three-fold 327 increase in OD595 (3.73 ± 0.25 and 3.30 ± 0.23, respectively) compared to the PBS-T 328 control (OD595 of 1.10 ± 0.05). Furthermore, the addition of 75% human urine resulted in 329 a four-fold increase in biofilm formation (OD595 of 4.39 ± 0.44) (Fig. 4C). Urine is a 330 complex medium consisting of more than sixty different compounds that compose 99% 331 of urine (38). We tested the ability of chemically defined artificial urine media (AUM) to 332 induce biofilm formation by the PA14 ∆pelD strain (39). The biofilm formed by PA14 333 ∆pelD in AUM (OD595 of 0.68 ± 0.23) increased 6-fold as compared to growth in LB 334 (OD595
of 0.11 ± 0.05) (Fig. 4D). Based on this data, we conclude that there are 335 components in both human and mouse urine that alter the program in vitro biofilm 336 development to a biofilm that is formed independent of exopolysaccharides. 337 338
on June 25, 2018 by guesthttp://iai.asm
.org/D
ownloaded from

16
Urea promotes polysaccharide-independent biofilms in vitro 339 In order to identify a specific component of urine that induces PA14 ∆pelD 340
pMMB-PA3702 biofilm formation, we performed biofilm assays in LB supplemented 341 with biologically relevant amounts of six of the most concentrated nitrogen and carbon 342 containing compounds identified in human urine. These compounds included urea (500 343 mM), citric acid (2 mM), creatinine (17.7 mM), glycine (49.4 mM), histidine (1.2 mM), 344 and glutamic acid (1.1 mM). Uric acid (1.7 mM) was not included in these studies as it 345 was insoluble in LB. Our biofilm results showed that urea was the only compound able to 346 significantly enhance biofilm formation by the ∆pelD mutant strain (OD595 of 0.43 ± 347 0.06) compared to the control (OD595 of 0.26 ± 0.06) (Fig. 5A). Interestingly, PA14 348 pMMB-PA3702 biofilm formation decreased in the presence of 0.5 M urea (OD595 of 349 0.28 ± 0.001) compared to the control (OD595 of 0.99 ± 0.11) (Fig. 5B). Although 350 reduced, the parental PA14 strain still formed biofilm in urea as compared to the PA14 351 ∆pelD strain in the absence of urea. We determined the concentration of urea in the 352 pooled mouse and human urine to be 786 mM and 243 mM, respectively (Supplemental 353 Materials and Methods). To determine the dose response of PA14 ∆pelD to urea, we 354 repeated our biofilm assays with 0.165 M urea or 0.330 M urea in catheters. In PBS-T, 355 PA14 formed biofilm on the catheters (OD595 of 0.53 ± 0.03), while the ∆pelD strain did 356 not (OD595 of 0.011 ± 0.007) (Fig. 5C). As was the case with the mouse urine, increasing 357 concentrations of urea reduced the PA14 biofilm formation on catheters to 0.52 ± 0.05 358 and 0.441 ± 0.009 in the presence of 0.165 M and 0.330 M, respectively (Fig. 5C). 359 Biofilm formation by the ∆pelD strain was enhanced by the presence of urea. In the 360 presence of 0.165 M and 0.330 M, the OD595 readings for PA14 ∆pelD were 0.0071 ± 361
on June 25, 2018 by guesthttp://iai.asm
.org/D
ownloaded from

17
0.0004 and 0.25 ± 0.07, respectively. These results indicate that urea, the main 362 nitrogenous waste component in mammalian urine, alters the program of biofilm 363 formation by P. aeruginosa to an exopolysaccharide-independent biofilm. 364 365 Urea promotes polysaccharide-independent biofilms on catheters in vitro 366 Results from the crystal violet assays for PA14 and PA14 ∆pelD suggest that the 367 ∆pelD mutant bacteria can form biofilm in the presence of urine and urea. We asked 368 whether these in vitro biofilms on catheters are morphologically similar when visualized 369 by SEM. Catheters incubated in vitro with either PA14 or PA14 ∆pelD in the presence or 370 absence of 0.5 M urea were washed, fixed, evaporative-coated and images were revealed 371 by SEM. Representative micrographs are shown in Fig. 6. As expected, the catheters 372 inoculated with wild-type PA14 in PBS-T contained a layer of bacteria that covered most 373 of the surface of the catheter (Fig. 6A). Greater magnification of the image reveals that 374 the wild-type PA14 have extracellular filaments that connect the individual cells to form 375 a biofilm (Fig. 6A, inset). Also expected, PA14 ∆pelD cannot form biofilm on catheters 376 in PBS-T just as the mutant is unable to form biofilm on plastic and glass materials in 377 vitro (Fig. 6B). In contrast, treatment in the presence of urea caused a reduction in the 378 coverage of the PA14 biofilm on catheters, which agrees with the results from crystal 379 violet assays (Fig. 6C). Urea treatment enabled the PA14 ∆pelD mutant to form biofilm 380 on the catheter surface (Fig. 6D). Increased magnification shows that the biofilm formed 381 by the PA14 ∆pelD mutant in the presence of urea appears to have similar filamentous 382 material that connects the bacteria (Fig. 6D, inset). Together, these results support that 383
on June 25, 2018 by guesthttp://iai.asm
.org/D
ownloaded from

18
urea alters the morphology of P. aeruginosa biofilm to a heterogeneous form that is 384 independent of exopolysaccharides. 385 Some clinical isolates of P. aeruginosa from human urinary tract infections 386 demonstrate a mucoid phenotype (40, 41). We asked whether urine or urea would induce 387 the production of alginate in vitro. We quantitated the amount of uronic acids present in 388 cultures grown with and without added urine or urea. We found that neither the parental 389 PA14 nor the PA14 ∆pelD mutant make detectable levels of alginate in response to urea 390 (Table S2). We then asked if urine or urea would affect alginate production from a 391 mucoid clinical isolate (CF27). We found that urea and urine inhibited the production of 392 alginate from CF27, suggesting that the biofilms formed during CAUTI are occurring 393 independent of alginate biosynthesis (Table S2). These results corroborate our findings 394 that PA14 ∆pelA-G ∆algD-A is capable of causing CAUTI similar to the parental PA14 395 strain. 396 397 Urea effects on biofilm formation by PA14 and PA14 ∆pelD are mediated by cell 398 death 399 One possible explanation for the ability of the ∆pelD mutant bacteria to form 400 biofilm in the presence of urea is that some cells lysed and released eDNA. To test this 401 possibility, we placed either PA14 or the ∆pelD mutant under agarose pads containing 402 propidium iodide to stain eDNA. In the absence of additional chemicals, neither PA14 403 nor PA14 ∆pelD released eDNA after 8 hours of growth under a LB agarose pad (Fig. 7A 404 and 7B). When placed under a LB agarose pad containing 0.5 M urea, the urea had little 405 effect on P. aeruginosa growth and replication. Exposure to 0.5 M urea led to cell 406
on June 25, 2018 by guesthttp://iai.asm
.org/D
ownloaded from

19
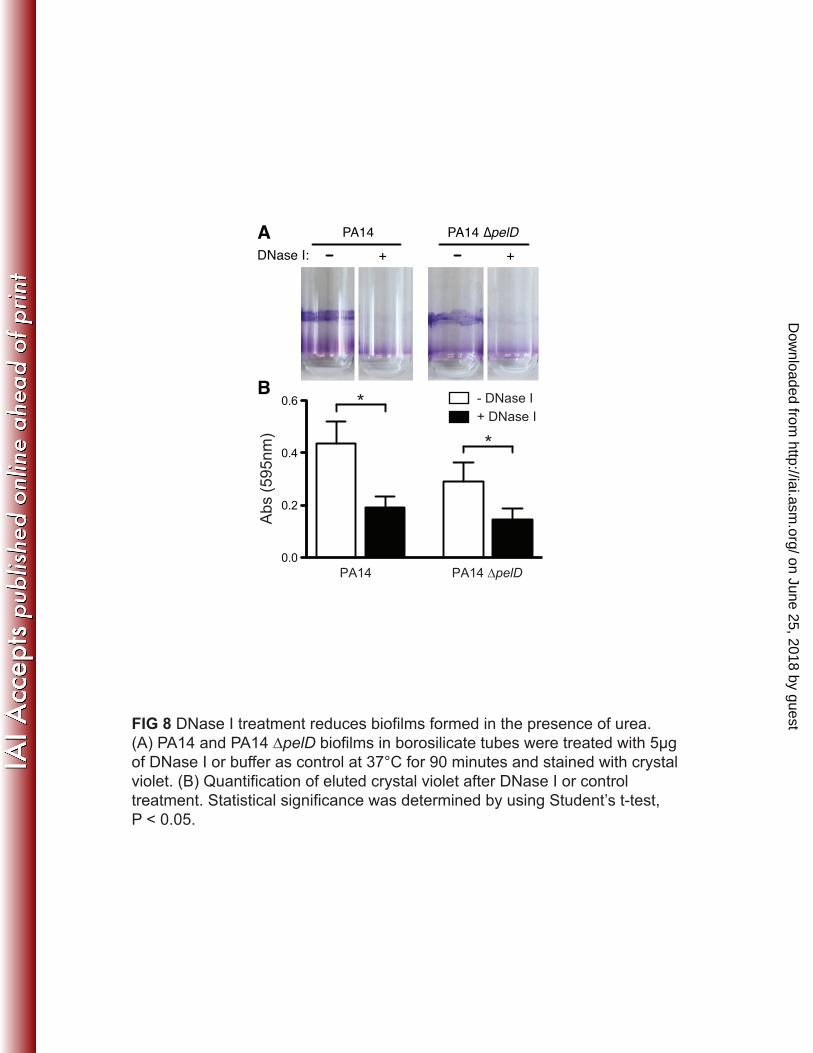
rounding in a subset of cells and their inability to exclude propidium iodide in both PA14 407 and the ∆pelD mutant (Fig. 7A and 7B and insets). Quantitation of these images 408 revealed that 0.07% of PA14 and 0.28% of PA14 ∆pelD had rounded or lysed. When 409 placed under a LB agarose pad with 0.1% SDS, P. aeruginosa also grew to high density 410 indicating that not all cells were lysed. Nonetheless, SDS led to the release of eDNA in a 411 subset of cells for both PA14 and ∆pelD strains (Fig. 7A and 7B). We asked if the lysis 412 of pseudomonads by urea is due to inhibition of growth. We performed growth curves of 413 PA14 and PA14 ∆pelD in various concentrations of urea and urine. We found that the 414 bacteria can reach the same numbers of CFU/ml in overnight cultures. Although the final 415 CFU/ml was not affected in overnight cultures, higher concentrations of urea (0.4 and 0.5 416 M) reduced the growth rate of PA14 (Fig. S5A-5C). The reduction in growth was not due 417 to the urease genes as transposon mutants of ureB, ureC, ureE, and ureG have similar 418 growth curves as PA14 (Fig. S5D). Together, these results support the hypothesis that the 419 ∆pelD mutant lacking extracellular polysaccharide can be induced to form biofilm via 420 release of eDNA from a subpopulation of rounded and lysed cells. 421 422 eDNA allows for biofilm formation independent of exopolysaccharides 423 To test our hypothesis that eDNA is a matrix component that contributes to 424 biofilm formation in the absence of PEL and PSL exopolysaccharides, we asked if DNase 425 I treatment reduces urea induced biofilms. PA14 and PA14 ∆pelD were grown in the 426 presence of 0.5 M urea for forty-eight hours to develop a biofilm. These biofilms were 427 washed and treated with buffer or buffer containing DNase I. Following DNase I 428 treatment, biofilms were washed a second time and immediately stained with crystal 429
on June 25, 2018 by guesthttp://iai.asm
.org/D
ownloaded from

20
violet (Fig. 8A). Significantly less pellicle was left in tubes treated with DNase I 430 compared to controls for both PA14 and PA14 ∆pelD. This suggests that eDNA is a key 431 component in the matrix that maintains biofilm structure. However, exopolysaccharides 432 still play a role in these biofilms formed in urea, as PA14 biofilms were more robust than 433 PA14 ∆pelD biofilms (Fig. 8B). While DNase I treatment significantly reduced biofilms 434 of both PA14 and PA14 ∆pelD, there were still residual biofilms remaining (Fig. 8B) 435 suggesting that there are additional factors other than eDNA that are contributing to 436 biofilm formation in the presence of urea. These results indicate that eDNA from lysed 437 bacteria is a major component of urea-mediated biofilms in vitro, which allows PA14 438 ∆pelD mutants to form biofilms independent of both PEL and PSL exopolysaccharides. 439 In addition, exposure to urea induces the parental PA14 to produce a DNase I-sensitive 440 biofilm. 441 442 443 on June 25, 2018 by guest
http://iai.asm.org/
Dow
nloaded from

21
DISCUSSION 444 445 Exopolysaccharides in P. aeruginosa biofilm-mediated infections 446
Previous investigations of the role of P. aeruginosa biofilms during infection 447 were assessed by two primary methods: 1. Analysis of tissue and isolates from infected 448 patients and 2. Animal-infection models. Wound infection biopsies of patients were 449 stained with P. aeruginosa- and Staphylococcus aureus-specific peptide-nucleic acid 450 stains and analyzed by confocal microscopy (42). These studies revealed that S. aureus 451 occupies the wound surface while P. aeruginosa resides deeper within the tissue beneath 452 the wound site (42). As both P. aeruginosa and S. aureus are detected in microcolonies 453 that resemble biofilms, these observations indicate that biofilm formation may be relevant 454 in wound infections. Recent studies sequencing bacteria isolated from cystic fibrosis (CF) 455 patients demonstrated that genetic mutations in P. aeruginosa lead to defective quorum 456 signaling, increased production of PEL and PSL polysaccharides, and the appearance of 457 small colony variants (SCV) (18, 37, 43). In previous studies and our current study, we 458 and others have shown that P. aeruginosa forms biofilms during CAUTIs (30). Taken 459 together, these studies show that P. aeruginosa biofilms participate in a number of 460 different infection sites including wounds, the lungs of CF patients, and the urinary tract. 461
The question of the matrix components involved in P. aeruginosa biofilm 462 formation during infection has been addressed using a number of animal model systems. 463 These studies reveal a diverse role for exopolysaccharide in different infection models. In 464 the murine subcutaneous catheter implant model, the ∆yfiR mutant, which has an SCV 465 phenotype and promotes biofilm formation via induction of pel and psl genes, had a 466
on June 25, 2018 by guesthttp://iai.asm
.org/D
ownloaded from

22
competitive disadvantage in the catheter and surrounding tissue as compared to the PAO1 467 parental strain (44). The authors attributed this defect to the reduced growth rate of the 468 bacteria. The direct contribution of the pel and psl genes in an animal infection model 469 was also assessed. In an acute pneumonia model of competitive infection, the ∆psl 470 mutant had a 3-fold decrease in competitive index as compared to the parental PAO1 471 strain at the 12-hour time point suggesting that extracellular polysaccharides can 472 contribute to acute and chronic infections (45). In a Drosophila model of oral P. 473 aeruginosa infection, both the PAO1 parent and the ∆rsmA mutant, which is 474 constitutively induced for pel and psl operons, formed biofilm in the fly crop (46). 475 However, the ∆rsmA mutant had a slower progression of lethality as compared to the 476 parental PAO1 suggesting reduced virulence (46). In the Drosophila model, the PEL 477 polysaccharide is required for biofilm formation, as a pelB insertion mutant does not form 478 microcolonies and larger aggregates (46). However, mutations in the gacAS, retS, and 479 rsm pathway prevent the expression of the type III secretion system, a potent system for 480 the delivery of cytotoxic effector molecules for acute infections, suggesting that biofilm 481 formation and lethality are reciprocally regulated by this pathway (46, 47). In a chinchilla 482 model of otitis media, parental PAO1 and an isogenic ∆wspA mutant, with constitutive 483 overproduction of c-di-GMP and induction of pel and psl genes, both colonized the bullae 484 (48). However, the ∆wspA mutant strain showed delayed pathogenesis suggesting 485 constitutive biofilm formation reduced the progress of disease symptoms. Further studies 486 that compared the ∆wspA strain with ∆wspA∆psl and ∆wspA∆pel revealed that neither 487 polysaccharide alone altered the outcome of the infection (48). However, eDNA was 488 visualized by staining with propidium iodide suggesting that eDNA may also serve as a 489
on June 25, 2018 by guesthttp://iai.asm
.org/D
ownloaded from

23
matrix component during the infection (48). Similarly, studies in Drosophila model of 490 infection have revealed another alternative pseudomonal biofilm pathway that is 491 regulated by PprAB two component regulatory system (49). Our studies using P. 492 aeruginosa PA14 and PAO1 strains lacking PEL, PSL, and alginate polysaccharides in 493 the murine model of CAUTI suggest that these exopolysaccharides are dispensable for 494 CAUTIs as eDNA can compensate for biofilm formation. Our results indicate that P. 495 aeruginosa utilizes a distinct program to form biofilms during CAUTI of mammalian 496 hosts and that eDNA can act as the primary matrix component of these biofilms. 497 Furthermore, P. aeruginosa biofilms form in response to different local host conditions 498 depending on the site of infection. 499 500 Genetic alterations in biofilms formed in vitro and in vivo 501
Some intriguing possible explanations for the ability of the ∆pel mutant to 502 participate in biofilm formation include either the acquisition of the necessary genes in 503 trans via horizontal gene transfer or the acquisition of a second site suppressor mutation. 504 Horizontal gene transfer is a likely possibility, as P. aeruginosa is known to have large 505 number of regions of genome plasticity on the chromosome (50). Mutagenesis of P. 506 aeruginosa in chronic infections such as those in cystic fibrosis patients is well known 507 and has been demonstrated in sequencing studies of longitudinally collected strains from 508 a single patient (37). In the mixed biofilm assays, we found that the PA14 strain 509 maintains its ability to form biofilm in vitro while the ∆pelD strain retains its inability to 510 form biofilm. These results suggest that the ability of ∆pelD mutant to participate in 511 mixed biofilm is likely due to the expression of a surface protein that specifically binds to 512
on June 25, 2018 by guesthttp://iai.asm
.org/D
ownloaded from

24
the PEL polysaccharide produced by the parental PA14 strain. Bacterial isolates from the 513 mice infected with either PA14 or the ∆pelD strain retained their respective pel locus and 514 their ability or inability to form in vitro biofilm. These results suggest that during a 14-515 day infection, P. aeruginosa has not begun to mutate the pel genes suggesting these genes 516 are not under negative selection during CAUTI. Extending the infection time may reveal 517 whether P. aeruginosa will accumulate mutations in chronic, non-CF infections. Future 518 studies in other chronic models of P. aeruginosa infection will reveal whether the PEL 519 and PSL polysaccharides are required for biofilm infections at other anatomical locations. 520 In addition to the requirement for PEL and PSL polysaccharide in vitro, our results also 521 suggest that P. aeruginosa mutants that are unable to produce PEL and PSL can 522 nonetheless participate in biofilm formed by wild-type bacteria. This raises an important 523 question as to the evolutionary pressure to prevent P. aeruginosa from cheating and 524 losing the pel genes. Likely, there are environmental conditions in which P. aeruginosa 525 form planktonic single-cells that have to be able to form biofilm without assistance from 526 other bacteria. Those cells that lost the ability to form biofilm from individual planktonic 527 cells would be at a great disadvantage. Another alternative is that participation in the 528 biofilm alone is not sufficient to confer the same physiological advantages as a PEL 529 polysaccharide-producing bacteria. Future experiments will delineate between these 530 possibilities. 531 532 Urea-mediated exopolysaccharide-independent biofilms 533
The urinary tract is a dynamic environment with physical, chemical and cellular 534 changes that can influence CAUTI. The bladder alone must expand and contract in order 535
on June 25, 2018 by guesthttp://iai.asm
.org/D
ownloaded from

25
to store and expel urine. The concentrations of components that make up urine are 536 constantly changing in response to the diet and hydration level of the host over the course 537 of a day (51). The cells of the bladder epithelium are repeated being turned over, a 538 process that occurs approximately every six weeks in guinea pigs (52). Also, during 539 infection, immune cells such as neutrophils can infiltrate into the lumen of the bladder. 540 Here, we demonstrate that human urine can induce P. aeruginosa biofilm formation and 541 is an important environmental factor in P. aeruginosa mediated CAUTI. Furthermore, to 542 determine a component in urine that enhanced biofilm formation in the absence of 543 exopolysaccharide, we individually tested the most abundant nitrogen and carbon-544 containing compounds detected in human urine. This does not exclude the possibility that 545 other non-nitrogen or non-carbon compounds enhance P. aeruginosa biofilm formation. 546 Of the components tested, only urea, the primary solute of urine, was able to induce P. 547 aeruginosa biofilm formation in the absence of exopolysaccharides. Urea may trigger a 548 number of possible changes to P. aeruginosa to promote biofilm formation by altering 549 the existing cell envelope or production of additional extracellular matrix. Here we show 550 that urea induced the release of eDNA from a subset of bacteria to enhance biofilm 551 formation of both parental and exopolysaccharide-deficient P. aeruginosa. These results 552 suggest that P. aeruginosa has a genetic program to respond to host urine that permits the 553 formation of an alternative eDNA-based biofilm. What is the basis for this urea-554 dependent activation of DNA release? Several studies have established that quorum 555 sensing induces eDNA release (53-55). In one study, PAO1 released increasing amounts 556 of eDNA when the culture reached stationary phase (55). Supporting the requirement of 557 the quorum sensing pathway, addition of the quorum sensing inhibitor furanone C30 558
on June 25, 2018 by guesthttp://iai.asm
.org/D
ownloaded from

26
prevented eDNA release (55). PAO1∆lasI, ∆rhlI, and ∆pqsA mutants released less eDNA 559 than parental PAO1 and formed thinner biofilms that lacked characteristic mushroom 560 formations (55). Furthermore, the reduction in eDNA release by these quorum mutants 561 could be complemented by supplementing media with exogenously added AHLs, 3-oxo-562 C12-HSL and C4-HSL (54, 55). Together, these studies demonstrate that the quorum 563 sensing system activates the release of eDNA to promote biofilm formation in vitro. In 564 our system, urine and urea can trigger eDNA release through the quorum-regulated 565 process or a novel mechanism that is independent of quorum regulation. In future studies 566 we will resolve the influence urea has on quorum sensing and in turn establish the role 567 quorum sensing plays during CAUTI. 568 569 ACKNOWLEDGMENTS 570
We thank Dr. Timothy Maugel at the Ultrastructure Laboratory at University of 571 Maryland at College Park for assistance with the SEM. We thank Drs. Kevin McIver, 572 David Mosser and members of the Lee laboratory for critical reading of the manuscript. 573 574
on June 25, 2018 by guesthttp://iai.asm
.org/D
ownloaded from

27
REFERENCES 575 576 1. Costerton JW, Stewart PS, Greenberg EP. 1999. Bacterial biofilms: a 577 common cause of persistent infections. Science 284:1318-1322. 578 2. Donlan RM, Costerton JW. 2002. Biofilms: survival mechanisms of clinically 579 relevant microorganisms. Clin Microbiol Rev 15:167-193. 580 3. Kunin CM. 1994. Infections of the urinary tract due to Pseudomonas 581
aeruginosa, p. 237-256. In Baltch AL, Smith RP (ed.), Pseudomonas aeruginosa 582 infections and treatment. Marcel Dekker, Inc., New York, NY. 583 4. Johnson DE, Lockatell CV, Hall-Craggs M, Warren JW. 1991. Mouse models 584 of short- and long-term foreign body in the urinary bladder: analogies to the 585 bladder segment of urinary catheters. Lab Anim Sci 41:451-455. 586 5. Warren JW. 2001. Catheter-associated urinary tract infections. Int J 587 Antimicrob Agents 17:299-303. 588 6. Warren JW, Steinberg L, Hebel JR, Tenney JH. 1989. The prevalence of 589 urethral catheterization in Maryland nursing homes. Arch Intern Med 590 149:1535-1537. 591 7. Stamm WE. 1991. Catheter-associated urinary tract infections: 592 epidemiology, pathogenesis, and prevention. Am J Med 91:65S-71S. 593 8. Colvin KM, Gordon VD, Murakami K, Borlee BR, Wozniak DJ, Wong GC, 594 Parsek MR. 2011. The pel polysaccharide can serve a structural and 595 protective role in the biofilm matrix of Pseudomonas aeruginosa. PLoS 596 pathogens 7:e1001264. 597 9. Costerton JW, Lewandowski Z, Caldwell DE, Korber DR, Lappin-Scott 598 HM. 1995. Microbial biofilms. Annu Rev Microbiol 49:711-745. 599 10. Anderson GG, O'Toole GA. 2008. Innate and induced resistance mechanisms 600 of bacterial biofilms. Curr Top Microbiol Immunol 322:85-105. 601 11. Stewart PS, Costerton JW. 2001. Antibiotic resistance of bacteria in biofilms. 602 Lancet 358:135-138. 603 12. Sutherland IW. 2001. The biofilm matrix--an immobilized but dynamic 604 microbial environment. Trends Microbiol 9:222-227. 605 13. Whitchurch CB, Tolker-Nielsen T, Ragas PC, Mattick JS. 2002. 606 Extracellular DNA required for bacterial biofilm formation. Science 607 295:1487. 608 14. Vallet I, Olson JW, Lory S, Lazdunski A, Filloux A. 2001. The 609 chaperone/usher pathways of Pseudomonas aeruginosa: identification of 610 fimbrial gene clusters (cup) and their involvement in biofilm formation. Proc 611 Natl Acad Sci U S A 98:6911-6916. 612 15. Vallet I, Diggle SP, Stacey RE, Camara M, Ventre I, Lory S, Lazdunski A, 613 Williams P, Filloux A. 2004. Biofilm formation in Pseudomonas aeruginosa: 614 fimbrial cup gene clusters are controlled by the transcriptional regulator 615 MvaT. J Bacteriol 186:2880-2890. 616
on June 25, 2018 by guesthttp://iai.asm
.org/D
ownloaded from

28
16. Kulasekara HD, Ventre I, Kulasekara BR, Lazdunski A, Filloux A, Lory S. 617 2005. A novel two-component system controls the expression of 618 Pseudomonas aeruginosa fimbrial cup genes. Mol Microbiol 55:368-380. 619 17. Wozniak DJ, Wyckoff TJ, Starkey M, Keyser R, Azadi P, O'Toole GA, 620 Parsek MR. 2003. Alginate is not a significant component of the extracellular 621 polysaccharide matrix of PA14 and PAO1 Pseudomonas aeruginosa biofilms. 622 Proc Natl Acad Sci U S A 100:7907-7912. 623 18. Starkey M, Hickman JH, Ma L, Zhang N, De Long S, Hinz A, Palacios S, 624 Manoil C, Kirisits MJ, Starner TD, Wozniak DJ, Harwood CS, Parsek MR. 625 2009. Pseudomonas aeruginosa rugose small-colony variants have 626 adaptations that likely promote persistence in the cystic fibrosis lung. J 627 Bacteriol 191:3492-3503. 628 19. Lawrence JR, Korber DR, Hoyle BD, Costerton JW, Caldwell DE. 1991. 629 Optical sectioning of microbial biofilms. J Bacteriol 173:6558-6567. 630 20. O'Toole GA, Kolter R. 1998. Flagellar and twitching motility are necessary 631 for Pseudomonas aeruginosa biofilm development. Mol Microbiol 30:295-632 304. 633 21. Friedman L, Kolter R. 2004. Genes involved in matrix formation in 634 Pseudomonas aeruginosa PA14 biofilms. Mol Microbiol 51:675-690. 635 22. Friedman L, Kolter R. 2004. Two genetic loci produce distinct 636 carbohydrate-rich structural components of the Pseudomonas aeruginosa 637 biofilm matrix. J Bacteriol 186:4457-4465. 638 23. Jackson KD, Starkey M, Kremer S, Parsek MR, Wozniak DJ. 2004. 639 Identification of psl, a locus encoding a potential exopolysaccharide that is 640 essential for Pseudomonas aeruginosa PAO1 biofilm formation. J Bacteriol 641 186:4466-4475. 642 24. Fux CA, Costerton JW, Stewart PS, Stoodley P. 2005. Survival strategies of 643 infectious biofilms. Trends Microbiol 13:34-40. 644 25. Stewart PS. 2001. Multicellular resistance: biofilms. Trends Microbiol 9:204. 645 26. Matsukawa M, Greenberg EP. 2004. Putative exopolysaccharide synthesis 646 genes influence Pseudomonas aeruginosa biofilm development. J Bacteriol 647 186:4449-4456. 648 27. Nemoto K, Hirota K, Murakami K, Taniguti K, Murata H, Viducic D, 649 Miyake Y. 2003. Effect of Varidase (streptodornase) on biofilm formed by 650 Pseudomonas aeruginosa. Chemotherapy 49:121-125. 651 28. Yanagihara K, Tomono K, Imamura Y, Kaneko Y, Kuroki M, Sawai T, 652 Miyazaki Y, Hirakata Y, Mukae H, Kadota J, Kohno S. 2002. Effect of 653 clarithromycin on chronic respiratory infection caused by Pseudomonas 654 aeruginosa with biofilm formation in an experimental murine model. J 655 Antimicrob Chemother 49:867-870. 656 29. Yanagihara K, Tomono K, Sawai T, Kuroki M, Kaneko Y, Ohno H, 657 Higashiyama Y, Miyazaki Y, Hirakata Y, Maesaki S, Kadota J, Tashiro T, 658 Kohno S. 2000. Combination therapy for chronic Pseudomonas aeruginosa 659 respiratory infection associated with biofilm formation. J Antimicrob 660 Chemother 46:69-72. 661
on June 25, 2018 by guesthttp://iai.asm
.org/D
ownloaded from

29
30. Kadurugamuwa JL, Modi K, Yu J, Francis KP, Purchio T, Contag PR. 2005. 662 Noninvasive biophotonic imaging for monitoring of catheter-associated 663 urinary tract infections and therapy in mice. Infect Immun 73:3878-3887. 664 31. Kadurugamuwa JL, Sin L, Albert E, Yu J, Francis K, DeBoer M, Rubin M, 665 Bellinger-Kawahara C, Parr Jr TR, Jr., Contag PR. 2003. Direct continuous 666 method for monitoring biofilm infection in a mouse model. Infect Immun 667 71:882-890. 668 32. Hoang TT, Karkhoff-Schweizer RR, Kutchma AJ, Schweizer HP. 1998. A 669 broad-host-range Flp-FRT recombination system for site-specific excision of 670 chromosomally-located DNA sequences: application for isolation of 671 unmarked Pseudomonas aeruginosa mutants. Gene 212:77-86. 672 33. DiGiandomenico A, Rao J, Harcher K, Zaidi TS, Gardner J, Neely AN, Pier 673 GB, Goldberg JB. 2007. Intranasal immunization with heterologously 674 expressed polysaccharide protects against multiple Pseudomonas aeruginosa 675 infections. Proc Natl Acad Sci U S A 104:4624-4629. 676 34. Becher A, Schweizer HP. 2000. Integration-proficient Pseudomonas 677 aeruginosa vectors for isolation of single-copy chromosomal lacZ and lux 678 gene fusions. BioTechniques 29:948-950, 952. 679 35. Kurosaka Y, Ishida Y, Yamamura E, Takase H, Otani T, Kumon H. 2001. A 680 non-surgical rat model of foreign body-associated urinary tract infection 681 with Pseudomonas aeruginosa. Microbiol Immunol 45:9-15. 682 36. Lee VT, Matewish JM, Kessler JL, Hyodo M, Hayakawa Y, Lory S. 2007. A 683 cyclic-di-GMP receptor required for bacterial exopolysaccharide production. 684 Mol Microbiol 65:1474-1484. 685 37. Smith EE, Buckley DG, Wu Z, Saenphimmachak C, Hoffman LR, D'Argenio 686 DA, Miller SI, Ramsey BW, Speert DP, Moskowitz SM, Burns JL, Kaul R, 687 Olson MV. 2006. Genetic adaptation by Pseudomonas aeruginosa to the 688 airways of cystic fibrosis patients. Proc Natl Acad Sci U S A 103:8487-8492. 689 38. Putnam DF. 1971. Composition and Concentrative Properties of Human 690 Urine. NASA Contractor Report. 691 39. Brooks T, Keevil CW. 1997. A simple artificial urine for the growth of 692 urinary pathogens. Lett Appl Microbiol 24:203-206. 693 40. Odugbemi T, Anandan N, Lina OF. 1992. Urinary tract pathogens with a 694 special reference to mucoid Pseudomonas aeruginosa at Aflaj General 695 Hospital. Ann Saudi Med 12:581-582. 696 41. Suman E, Blat KG, Hegde BK. 1993. Mucoid Pseudomonas aeruginosa in 697 urinary tract infection. Trop Doct 23:180-181. 698 42. Fazli M, Bjarnsholt T, Kirketerp-Moller K, Jorgensen B, Andersen AS, 699 Krogfelt KA, Givskov M, Tolker-Nielsen T. 2009. Nonrandom distribution 700 of Pseudomonas aeruginosa and Staphylococcus aureus in chronic wounds. J 701 Clin Microbiol 47:4084-4089. 702 43. Bjarnsholt T, Jensen PO, Jakobsen TH, Phipps R, Nielsen AK, Rybtke MT, 703 Tolker-Nielsen T, Givskov M, Hoiby N, Ciofu O. 2010. Quorum sensing and 704 virulence of Pseudomonas aeruginosa during lung infection of cystic fibrosis 705 patients. PloS one 5:e10115. 706
on June 25, 2018 by guesthttp://iai.asm
.org/D
ownloaded from

30
44. Malone JG, Jaeger T, Spangler C, Ritz D, Spang A, Arrieumerlou C, Kaever 707 V, Landmann R, Jenal U. 2010. YfiBNR mediates cyclic di-GMP dependent 708 small colony variant formation and persistence in Pseudomonas aeruginosa. 709 PLoS pathogens 6:e1000804. 710 45. Mishra M, Byrd MS, Sergeant S, Azad AK, Parsek MR, McPhail L, 711 Schlesinger LS, Wozniak DJ. 2012. Pseudomonas aeruginosa Psl 712 polysaccharide reduces neutrophil phagocytosis and the oxidative response 713 by limiting complement-mediated opsonization. Cell Microbiol 14:95-106. 714 46. Mulcahy H, O'Callaghan J, O'Grady EP, Macia MD, Borrell N, Gomez C, 715 Casey PG, Hill C, Adams C, Gahan CG, Oliver A, O'Gara F. 2007. 716 Pseudomonas aeruginosa RsmA plays an important role during murine 717 infection by influencing colonization, virulence, persistence and pulmonary 718 inflammation. Infect Immun. 719 47. Goodman AL, Kulasekara B, Rietsch A, Boyd D, Smith RS, Lory S. 2004. A 720 signaling network reciprocally regulates genes associated with acute 721 infection and chronic persistence in Pseudomonas aeruginosa. Dev Cell 722 7:745-754. 723 48. Byrd MS, Pang B, Hong W, Waligora EA, Juneau RA, Armbruster CE, 724 Weimer KE, Murrah K, Mann EE, Lu H, Sprinkle A, Parsek MR, Kock ND, 725 Wozniak DJ, Swords WE. 2011. Direct Evaluation of Pseudomonas 726 aeruginosa Biofilm Mediators in a Chronic Infection Model. Infect Immun 727 79:3087-3095. 728 49. de Bentzmann S, Giraud C, Bernard CS, Calderon V, Ewald F, Plesiat P, 729 Nguyen C, Grunwald D, Attree I, Jeannot K, Fauvarque MO, Bordi C. 2012. 730 Unique biofilm signature, drug susceptibility and decreased virulence in 731 Drosophila through the Pseudomonas aeruginosa two-component system 732 PprAB. PLoS pathogens 8:e1003052. 733 50. Mathee K, Narasimhan G, Valdes C, Qiu X, Matewish JM, Koehrsen M, 734 Rokas A, Yandava CN, Engels R, Zeng E, Olavarietta R, Doud M, Smith RS, 735 Montgomery P, White JR, Godfrey PA, Kodira C, Birren B, Galagan JE, 736 Lory S. 2008. Dynamics of Pseudomonas aeruginosa genome evolution. Proc 737 Natl Acad Sci U S A 105:3100-3105. 738 51. Lewis SA. 2000. Everything you wanted to know about the bladder 739 epithelium but were afraid to ask. Am J Physiol Renal Physiol 278:F867-874. 740 52. Martin BF. 1972. Cell replacement and differentiation in transitional 741 epithelium: a histological and autoradiographic study of the guinea-pig 742 bladder and ureter. J Anat 112:433-455. 743 53. Davies DG, Parsek MR, Pearson JP, Iglewski BH, Costerton JW, 744 Greenberg EP. 1998. The involvement of cell-to-cell signals in the 745 development of a bacterial biofilm. Science 280:295-298. 746 54. Nakamura S, Higashiyama Y, Izumikawa K, Seki M, Kakeya H, Yamamoto 747 Y, Yanagihara K, Miyazaki Y, Mizuta Y, Kohno S. 2008. The roles of the 748 quorum-sensing system in the release of extracellular DNA, 749 lipopolysaccharide, and membrane vesicles from Pseudomonas aeruginosa. 750 Jpn J Infect Dis 61:375-378. 751
on June 25, 2018 by guesthttp://iai.asm
.org/D
ownloaded from

31
55. Allesen-Holm M, Barken KB, Yang L, Klausen M, Webb JS, Kjelleberg S, 752 Molin S, Givskov M, Tolker-Nielsen T. 2006. A characterization of DNA 753 release in Pseudomonas aeruginosa cultures and biofilms. Mol Microbiol 754 59:1114-1128. 755 756 757
on June 25, 2018 by guesthttp://iai.asm
.org/D
ownloaded from

32
FIG. LEGENDS 758 759 FIG 1 P. aeruginosa colonization of catheters during CAUTI is independent of PEL, PSL 760 and alginate exopolysaccharides. Indicated organs from mice infected with PA14, PA14 761 ∆pelD, PA14 ∆pelA-G or PA14 ∆pelA-G ∆algD-A in the murine model of CAUTI were 762 harvested, homogenized and plated for CFU counts. Each symbol represents an 763 individual mouse. Bars represent means. Points at 100 CFU per macerated organ 764 represent samples below the limit of detection. ns, differences not significant as 765 determined by one-way ANOVA. 766 767 FIG 2 Scanning electron micrographs of catheters isolated from mouse bladders. 768 Catheters were isolated from mice infected with either (A) PAO1 or (B) PA14 ∆pelD 769 pMMB-PA3702 and visualized by SEM. Bar length equals 5 µm. The asterisks indicate 770 divided bacteria and the arrowheads indicate filamentous bacteria. 771 772 FIG 3 PA14 and PA14 ∆pelD do not compete in the murine CAUTI model. Mice were 773 inoculated with either PA14 and PA14 ∆pelD-lux or PA14-lux and PA14 ∆pelD. Each 774 symbol represents the ratio of PA14 ΔpelD/PA14 in either the bladder or kidneys of an 775 individual mouse. Bars represent means. 776 777 FIG 4 Mouse and human urine induce biofilm formation by PA14 ∆pelD. (A) 778 Quantification of crystal violet eluted from catheters after staining for colonization by 779 PA14 pMMB-PA3702 or PA14 ∆pelD pMMB-PA3702 in the presence of PBS-T with or 780
on June 25, 2018 by guesthttp://iai.asm
.org/D
ownloaded from

33
without indicated amounts of mouse urine. (B) Quantification of PA14 ∆pelD pMMB-781 PA3702 or (C) PA14 pMMB-PA3702 biofilms in PBS-T with or without indicated 782 amounts of human urine. (D) PA14 ∆pelD biofilm formation in LB or artificial urine 783 media (AUM). Statistical significance was determined by using Student’s t-test, P < 0.05. 784 785 FIG 5 Urea induces biofilm formation by PA14 ∆pelD. 786 Quantification of crystal violet biofilm assays for colonization of either (A) PA14 787 pMMB-PA3702 (closed bars) or (B) PA14 ∆pelD pMMB-PA3702 (open bars) in the 788 presence of LB or LB supplemented with urea, citric acid, creatinine, glycine, histidine, 789 or glutamic acid. (C) Biofilm assays of catheters inoculated with either PA14 pMMB-790 PA3702 (closed bars) or PA14 ∆pelD pMMB-PA3702 (open bars) in PBS-T with or 791 without urea at the indicated concentrations. Statistical significance was determined by 792 using Student’s t-test, P < 0.05. 793 794 FIG 6 Scanning electron microscopy reveals biofilm formation by PA14 ∆pelD pMMB-795 PA3702 in the presence of urea. (A and C) PA14 pMMB-PA3702 and (B and D) PA14 796 ∆pelD pMMB-PA3702 were visualized after incubation for 16 h on catheter tubing in the 797 absence or presence of 0.5 M urea. Inset photos taken at higher magnification. Bar length 798 equals 5 µm. 799 800 FIG 7 Urea causes release of DNA from a subset of P. aeruginosa cells. 801 (A) PA14 or (B) PA14 ∆pelD overnight cultures were subcultured 1:100 and placed 802 under agarose pads containing LB with the indicated concentrations of urea or SDS. 803
on June 25, 2018 by guesthttp://iai.asm
.org/D
ownloaded from

34
Samples were incubated for 8 hours at 37˚C in a humidified chamber. Cells were imaged 804 by phase contrast microscopy and eDNA was detected by propidium iodide staining 805 (red). Insets show magnified images of rounded cells (indicated by arrowheads) that stain 806 positive with propidium iodide. 807 808 FIG 8 DNase I treatment reduces biofilms formed in the presence of urea. (A) PA14 and 809 PA14 ∆pelD biofilms in borosilicate tubes were treated with 5µg of DNase I or buffer as 810 control at 37°C for 90 minutes and stained with crystal violet. (B) Quantification of 811 eluted crystal violet after DNase I or control treatment. Statistical significance was 812 determined by using Student’s t-test, P < 0.05. 813 814 815
on June 25, 2018 by guesthttp://iai.asm
.org/D
ownloaded from

35
Table 1. Pseudomonas strains maintain genetic identity after murine UTI. 816 Isolatesa PCR of pelDb
Biofilm
Positivec
Biofilm
Negatived
PA14 (24) WT 24 0
∆pelD 0 0
PA14 ∆pelD (36) WT 0 0
∆pelD 0 36
PA14 ∆pelA-G (16) WT 0 0
∆pelD 0 16
PA14 ∆pelA-G ∆algD-A (42) WT 0 0
∆pelD 0 42
817 a Strains were isolated from catheters used for murine UTIs. 818 b Isolates were genotyped using primers specific to the pelD gene, vl491 (5’-819 AGTTAACGGAGTGGGCCCACACACTGTTCT-3’, vl494 (5’-820 AAAGCTTTGTGTTCGGTCATGTCCAGTATCT-3’). 821 c Biofilm positive isolates had an OD595 ≥0.5 with an average OD595 of 1.35 ± 0.36. 822 d Biofilm negative isolates had an OD595 <0.5 with an average OD595 of 0.09 ± 0.08. 823 824 on June 25, 2018 by guest
http://iai.asm.org/
Dow
nloaded from

0
1
2
3
4
5
6
7
8
9
10
Kidneys Bladder
Lo
g C
FU
pe
r o
rga
n1
0
ns ns
PA14
PA14 ∆pelA-G
PA14 ∆pelD
PA14 ∆pelA-G ∆algD-A
FIG 1 P. aeruginosa colonization of catheters during CAUTI is independent of PEL, PSL and alginate
exopolysaccharides. Indicated organs from mice infected with PA14, PA14 ∆pelD, PA14 ∆pelA-G
or PA14 ∆pelA-G ∆algD-A in the murine model of CAUTI were harvested, homogenized and plated for
CFU counts. Each symbol represents an individual mouse. Bars represent means. Points at 100 CFU
per macerated organ represent samples below the limit of detection. ns, differences not significant as
determined by one-way ANOVA.
on June 25, 2018 by guesthttp://iai.asm
.org/D
ownloaded from

5 µm
A
B
5 µm
*
*
*
*
FIG 2 Scanning electron micrographs of catheters isolated from mouse bladders.
Catheters were isolated from mice infected with either (A) PAO1 or (B) PA14 ∆pelD
pMMB-PA3702 and visualized by SEM. Bar length equals 5 µm. The asterisks indicate
divided bacteria and the arrowheads indicate filamentous bacteria.
on June 25, 2018 by guesthttp://iai.asm
.org/D
ownloaded from

Bladder Kidney
Co
mp
etitive
in
de
x
-2
-1
0
1
PA14 ∆pelD-lux
PA14
PA14 ∆pelD
PA14-lux
FIG 3 PA14 and PA14 ∆pelD do not compete in the murine CAUTI model.
Mice were inoculated with either PA14 and PA14 ∆pelD-lux or PA14-lux and PA14 ∆pelD.
Each symbol represents the ratio of PA14 ΔpelD/PA14 in either the bladder or kidneys of
an individual mouse. Bars represent means.
on June 25, 2018 by guesthttp://iai.asm
.org/D
ownloaded from

0.0
0.1
0.2
0.3
25%
50%
75%
PB
S-T
Human Urine
**
Ab
s (
59
5 n
m)
BA PA14 PA14∆pelD
0.00
0.05
0.10
0.15
0.20
PBS-T 50% 90%
Urine
Ab
s (
59
5 n
m)
0
2
4
6
C
Ab
s (
59
5 n
m)
** *
25%
50%
75%
PB
S-T
Human Urine
0.0
0.2
0.4
0.6
0.8
1.0
LB AUM
*
D
Ab
s (
59
5 n
m)
FIG 4 Mouse and human urine induce biofilm formation by PA14 ∆pelD.
(A) Quantification of crystal violet eluted from catheters after staining for colonization
by PA14 pMMB-PA3702 or PA14 ∆pelD pMMB-PA3702 in the presence of PBS-T
with or without indicated amounts of mouse urine. (B) Quantification of PA14 ∆pelD
pMMB-PA3702 or (C) PA14 pMMB-PA3702 biofilms in PBS-T with or without
indicated amounts of human urine. (D) PA14 ∆pelD biofilm formation in LB or
artificial urine media (AUM).
on June 25, 2018 by guesthttp://iai.asm
.org/D
ownloaded from

FIG 5 Urea induces biofilm formation by PA14 ∆pelD.
Quantification of crystal violet biofilm assays for colonization of either (A) PA14 pMMB-PA3702
(closed bars) or (B) PA14 ∆pelD pMMB-PA3702 (open bars) in the presence of LB or LB
supplemented with urea, citric acid, creatinine, glycine, histidine, or glutamic acid. (C) Biofilm
assays of catheters inoculated with either PA14 pMMB-PA3702 (closed bars) or PA14 ∆pelD
pMMB-PA3702 (open bars) in PBS-T with or without urea at the indicated concentrations.
Statistical significance was determined by using Student’s t-test, P < 0.05.
C
A
Co
ntr
ol
Ure
a
Citri
c A
cid
Cre
atin
ine
Gly
cin
e
His
tid
ine
Glu
tam
ic A
cid
0.0
0.5
1.0
1.5
*
Ab
s (
59
5 n
m)
B
0.0
0.2
0.4
0.6*
Ab
s (
59
5 n
m)
Co
ntr
ol
Ure
a
Citri
c A
cid
Cre
atin
ine
Gly
cin
e
His
tid
ine
Glu
tam
ic A
cid
0.0
0.2
0.4
0.6
PBS-T 0.165 0.330
Urea (M)
Ab
s (
59
5 n
m)
on June 25, 2018 by guesthttp://iai.asm
.org/D
ownloaded from

5μm
5μm
PA14 PA14 ∆pelD
PBS-T
PBS-T
+
Urea
A B
C D
5μm
5μm
5μm
5μm
5μm
FIG 6 Scanning electron microscopy reveals biofilm formation by PA14 ∆pelD pMMB-PA3702 in the presence
of urea. (A and C) PA14 pMMB-PA3702 and (B and D) PA14 ∆pelD pMMB-PA3702 were visualized after incubation
for 16 h on catheter tubing in the absence or presence of 0.5 M urea. Inset photos taken at higher magnification.
Bar length equals 5 µm.
on June 25, 2018 by guesthttp://iai.asm
.org/D
ownloaded from

Phase Contrast Propidium Iodide OverlayA. PA14
Phase Contrast Propidium Iodide Overlay
B. PA14 ∆pelD
0.1%
SDS
0.5M
Urea
0M
Urea
0.1%
SDS
0M
Urea
0.5M
Urea
FIG 7 Urea causes release of DNA from a subset of P. aeruginosa cells.
(A) PA14 or (B) PA14 ∆pelD overnight cultures were subcultured 1:100 and placed under agarose
pads containing LB with the indicated concentrations of urea or SDS. Samples were incubated for
8 hours at 37˚C in a humidified chamber. Cells were imaged by phase contrast microscopy and
eDNA was detected by propidium iodide staining (red). Insets show magnified images of rounded
cells (indicated by arrowheads) that stain positive with propidium iodide.
on June 25, 2018 by guesthttp://iai.asm
.org/D
ownloaded from
PA14
DNase I:
A
B
0.0
0.2
0.4
0.6 - DNase I
+ DNase I
PA14 PA14 ∆pelD
*
*
Ab
s(5
95
nm
)+
PA14 ∆pelD
+
FIG 8 DNase I treatment reduces biofilms formed in the presence of urea.
(A) PA14 and PA14 ∆pelD biofilms in borosilicate tubes were treated with 5µg
of DNase I or buffer as control at 37°C for 90 minutes and stained with crystal
violet. (B) Quantification of eluted crystal violet after DNase I or control
treatment. Statistical significance was determined by using Student’s t-test,
P < 0.05.
on June 25, 2018 by guesthttp://iai.asm
.org/D
ownloaded from