cell, vol. 108, 475–487, february 22, 2002, copyright … · cooperation between complexes review...
TRANSCRIPT

Cell, Vol. 108, 475–487, February 22, 2002, Copyright 2002 by Cell Press
ReviewCooperation between Complexesthat Regulate ChromatinStructure and Transcription
repressors, mediator complexes, and general transcrip-tion factors are also required to ensure proper regulation.
The conceptual problem discussed in this review, howchromatin-modifying complexes interact with eachother to generate specific template structures, is funda-
Geeta J. Narlikar, Hua-Ying Fan,and Robert E. Kingston1
Department of Molecular BiologyMassachusetts General HospitalBoston, Massachusetts 02114Department of Genetics mental to all known nuclear processes such as DNA
replication, recombination, and repair. Since each ofHarvard Medical SchoolBoston, Massachusetts 02115 these processes functions on chromatin, regulation of
the accessibility of chromatin structure can play a rolein these nuclear processes much as it does in tran-scription.
Two major classes of complexes regulate accessibil-Chromatin structure creates barriers for each stepity of the template to DNA binding factors. ATP-depen-in eukaryotic transcription. Here we discuss how thedent complexes can move nucleosome positions,activities of two major classes of chromatin-modifyingthereby exposing or occluding DNA sequences, and cancomplexes, ATP-dependent remodeling complexescreate conformations where DNA is accessible on theand HAT or HDAC complexes, might be coordinatedsurface of the histone octamer. The major questionsto create a DNA template that is accessible to theconcerning this class of complexes are the relative im-general transcription apparatus.portance, in biological systems, of “sliding” nucleo-somes (as opposed to catalyzing conformational changesof the nucleosome) and the nature and stability of the
No individual factor is capable of playing a dominant remodeled state of the nucleosome.role in generating the immense specificity required to The other class of complexes, which covalently mod-regulate transcription in eukaryotes. Distinct multipro- ify nucleosomes, can add or remove many chemicaltein complexes are needed to modulate higher-order moieties; acetylation, phosphorylation, and methylationchromatin structure, to bind to promoters, to bind to of histone N termini have received the most recent atten-enhancers, to communicate between activators and re- tion. The chemical and enzymatic activities of many ofpressors and sites of transcription initiation, to modify these complexes are well understood. The major ques-nucleosomal structure, and to generate transcripts. tions concern how these covalent modifications impactEach of these complexes might be a key player in regu- the structure of the template and the ability of otherlating a given gene. A major challenge is to determine complexes (e.g., those in the transcription machinery)how all of these complexes work together to ensure to function. For example, numerous acetyltransferaseproper regulation. complexes have been associated with transcription acti-
The scope of this problem is too large to be covered vation; what is it about the action of these complexesin a single review. The goal here is to discuss a segment and specific acetylation patterns of histone tails thatof this problem: how do ATP-dependent remodeling and leads to increased transcription?histone-modifying complexes work together to modify We will start with a discussion of the nature and capa-chromatin structure and regulate function of the tran- bilities of each individual class of complexes. We willscription machinery? ATP-dependent remodeling com- then consider how these classes of complexes can coor-plexes use energy to modify chromatin structure in a dinate with each other and with the transcription ma-noncovalent manner, while histone-modifying com- chinery to create robust and specific regulation.plexes add or remove covalent modifications from his-tone tails. ATP-Dependent Remodeling Complexes
In principle, all reactions that involve DNA can be regu- ATP-dependent remodeling complexes use ATP hydro-lated by altering DNA packaging and hence DNA accessi- lysis to increase the accessibility of nucleosomal DNA,bility. Transcription requires that the DNA be accessible to which is a fundamental requirement for several steps insequence-specific transcription factors and RNA poly- transcription. These complexes can be divided into threemerase and requires the melting and reformation of the main classes based on the identity of their catalyticdouble helix throughout the length of the transcript. Chro- ATPase subunit (Figure 1). These ATPase subunits dis-matin structure, including the structure imposed by the play homology only within the ATPase domain and con-nucleosome, impedes all steps required for transcription. tain different additional domains. Further, each ATPaseThus, in the simplest case, repression can be achieved subunit forms complexes with different additional pro-by creating a stable, inaccessible chromatin structure, and teins (Figure 1). Our knowledge of the composition ofactivation can be achieved by creating an accessible chro- these complexes and their biochemical capabilities ismatin structure. While regulating chromatin structure is constantly growing (Figures 1 and 2), but there are stillnecessary for regulating gene expression, it is not suffi- some key unanswered questions that are important forcient: the functions of sequence-specific activators and understanding how these complexes function in vivo:
what are the mechanisms by which these different com-plexes increase access to nucleosomal DNA; do the large1Correspondence: [email protected]
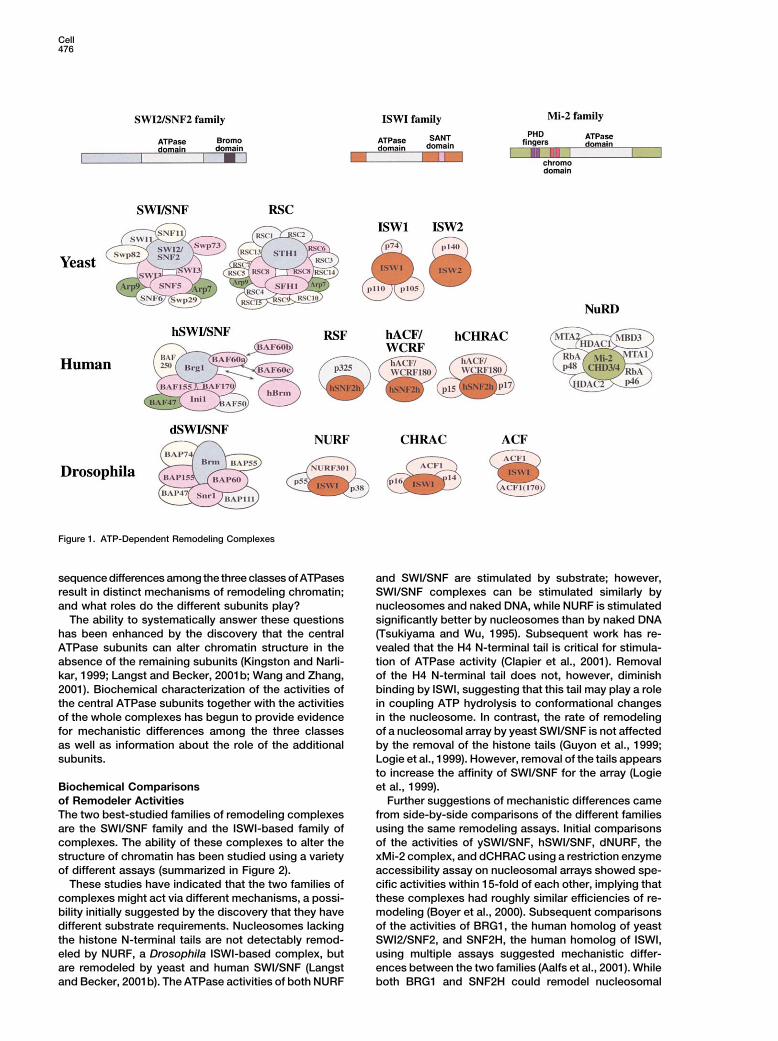
Cell476
Figure 1. ATP-Dependent Remodeling Complexes
sequence differences among the three classes of ATPases and SWI/SNF are stimulated by substrate; however,SWI/SNF complexes can be stimulated similarly byresult in distinct mechanisms of remodeling chromatin;
and what roles do the different subunits play? nucleosomes and naked DNA, while NURF is stimulatedsignificantly better by nucleosomes than by naked DNAThe ability to systematically answer these questions
has been enhanced by the discovery that the central (Tsukiyama and Wu, 1995). Subsequent work has re-vealed that the H4 N-terminal tail is critical for stimula-ATPase subunits can alter chromatin structure in the
absence of the remaining subunits (Kingston and Narli- tion of ATPase activity (Clapier et al., 2001). Removalof the H4 N-terminal tail does not, however, diminishkar, 1999; Langst and Becker, 2001b; Wang and Zhang,
2001). Biochemical characterization of the activities of binding by ISWI, suggesting that this tail may play a rolein coupling ATP hydrolysis to conformational changesthe central ATPase subunits together with the activities
of the whole complexes has begun to provide evidence in the nucleosome. In contrast, the rate of remodelingof a nucleosomal array by yeast SWI/SNF is not affectedfor mechanistic differences among the three classes
as well as information about the role of the additional by the removal of the histone tails (Guyon et al., 1999;Logie et al., 1999). However, removal of the tails appearssubunits.to increase the affinity of SWI/SNF for the array (Logieet al., 1999).Biochemical Comparisons
of Remodeler Activities Further suggestions of mechanistic differences camefrom side-by-side comparisons of the different familiesThe two best-studied families of remodeling complexes
are the SWI/SNF family and the ISWI-based family of using the same remodeling assays. Initial comparisonsof the activities of ySWI/SNF, hSWI/SNF, dNURF, thecomplexes. The ability of these complexes to alter the
structure of chromatin has been studied using a variety xMi-2 complex, and dCHRAC using a restriction enzymeaccessibility assay on nucleosomal arrays showed spe-of different assays (summarized in Figure 2).
These studies have indicated that the two families of cific activities within 15-fold of each other, implying thatthese complexes had roughly similar efficiencies of re-complexes might act via different mechanisms, a possi-
bility initially suggested by the discovery that they have modeling (Boyer et al., 2000). Subsequent comparisonsof the activities of BRG1, the human homolog of yeastdifferent substrate requirements. Nucleosomes lacking
the histone N-terminal tails are not detectably remod- SWI2/SNF2, and SNF2H, the human homolog of ISWI,using multiple assays suggested mechanistic differ-eled by NURF, a Drosophila ISWI-based complex, but
are remodeled by yeast and human SWI/SNF (Langst ences between the two families (Aalfs et al., 2001). Whileboth BRG1 and SNF2H could remodel nucleosomaland Becker, 2001b). The ATPase activities of both NURF
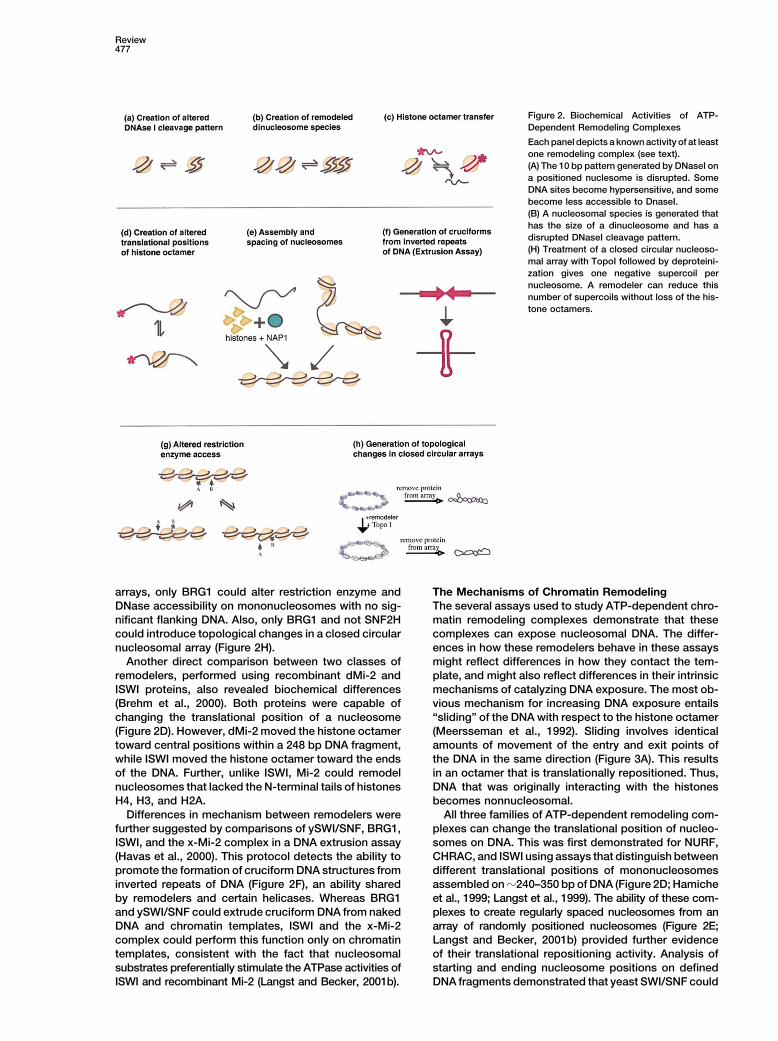
Review477
Figure 2. Biochemical Activities of ATP-Dependent Remodeling Complexes
Each panel depicts a known activity of at leastone remodeling complex (see text).(A) The 10 bp pattern generated by DNaseI ona positioned nuclesome is disrupted. SomeDNA sites become hypersensitive, and somebecome less accessible to DnaseI.(B) A nucleosomal species is generated thathas the size of a dinucleosome and has adisrupted DNaseI cleavage pattern.(H) Treatment of a closed circular nucleoso-mal array with TopoI followed by deproteini-zation gives one negative supercoil pernucleosome. A remodeler can reduce thisnumber of supercoils without loss of the his-tone octamers.
arrays, only BRG1 could alter restriction enzyme and The Mechanisms of Chromatin RemodelingThe several assays used to study ATP-dependent chro-DNase accessibility on mononucleosomes with no sig-
nificant flanking DNA. Also, only BRG1 and not SNF2H matin remodeling complexes demonstrate that thesecomplexes can expose nucleosomal DNA. The differ-could introduce topological changes in a closed circular
nucleosomal array (Figure 2H). ences in how these remodelers behave in these assaysmight reflect differences in how they contact the tem-Another direct comparison between two classes of
remodelers, performed using recombinant dMi-2 and plate, and might also reflect differences in their intrinsicmechanisms of catalyzing DNA exposure. The most ob-ISWI proteins, also revealed biochemical differences
(Brehm et al., 2000). Both proteins were capable of vious mechanism for increasing DNA exposure entails“sliding” of the DNA with respect to the histone octamerchanging the translational position of a nucleosome
(Figure 2D). However, dMi-2 moved the histone octamer (Meersseman et al., 1992). Sliding involves identicalamounts of movement of the entry and exit points oftoward central positions within a 248 bp DNA fragment,
while ISWI moved the histone octamer toward the ends the DNA in the same direction (Figure 3A). This resultsin an octamer that is translationally repositioned. Thus,of the DNA. Further, unlike ISWI, Mi-2 could remodel
nucleosomes that lacked the N-terminal tails of histones DNA that was originally interacting with the histonesbecomes nonnucleosomal.H4, H3, and H2A.
Differences in mechanism between remodelers were All three families of ATP-dependent remodeling com-plexes can change the translational position of nucleo-further suggested by comparisons of ySWI/SNF, BRG1,
ISWI, and the x-Mi-2 complex in a DNA extrusion assay somes on DNA. This was first demonstrated for NURF,CHRAC, and ISWI using assays that distinguish between(Havas et al., 2000). This protocol detects the ability to
promote the formation of cruciform DNA structures from different translational positions of mononucleosomesassembled on �240–350 bp of DNA (Figure 2D; Hamicheinverted repeats of DNA (Figure 2F), an ability shared
by remodelers and certain helicases. Whereas BRG1 et al., 1999; Langst et al., 1999). The ability of these com-plexes to create regularly spaced nucleosomes from anand ySWI/SNF could extrude cruciform DNA from naked
DNA and chromatin templates, ISWI and the x-Mi-2 array of randomly positioned nucleosomes (Figure 2E;Langst and Becker, 2001b) provided further evidencecomplex could perform this function only on chromatin
templates, consistent with the fact that nucleosomal of their translational repositioning activity. Analysis ofstarting and ending nucleosome positions on definedsubstrates preferentially stimulate the ATPase activities of
ISWI and recombinant Mi-2 (Langst and Becker, 2001b). DNA fragments demonstrated that yeast SWI/SNF could

Cell478
have flanking DNA onto which the histone octamer canslide. Restriction sites that are closer to the center ofthe DNA are exposed with similar or faster rates thansites that are close to the DNA ends, contrary to theresult expected from sliding of the histone octamer (Nar-likar et al., 2001). Further, site-specific crosslinking ofthe DNA to the histone octamer, which is expected toprevent sliding of the DNA, does not prevent remodelingby hSWI/SNF (Lee et al., 1999).
Another argument that sliding is not the sole mecha-nism for remodeling comes from studies showing thathSWI/SNF and ySWI/SNF can introduce topologicalchanges in closed circular nucleosomal arrays (Guyonet al., 2001; Jaskelioff et al., 2000; Kwon et al., 1994).Translational repositioning of histone octamers is notexpected to cause stable topological changes becausenucleosomes in a standard conformation have been
Figure 3. Two Models for the Mechanism of ATP-Dependent shown to adopt a topology that does not vary signifi-Nucleosome Remodeling cantly. Any transient changes in the twist or writhe ofThe structures are depicted for the intermediate, and products in linker DNA caused by the movement of nucleosomes(B) are hypothetical and could involve changes in the conformation would be expected to resolve rapidly on the uncon-of DNA, histones, or both. strained templates that have been used to study topo-
logical changes. In contrast, the topological changesintroduced by hSWI/SNF are stable on a timescale ofalso move nucleosomes in cis to a new positionhours, suggesting that SWI/SNF alters the conformation(Whitehouse et al., 1999). In separate studies performedof the nucleosome in a manner that kinetically traps theusing nucleosomal arrays, SWI/SNF action blocked cer-topological changes of the remodeled products (Guyontain restriction enzyme sites in linker regions, implyinget al., 2001).that SWI/SNF was able to reposition nucleosomes over
These experiments suggest that a classical slidingthese previously accessible sites (Jaskelioff et al., 2000).mechanism (Figure 3A) is not obligatory for remodelingFurther, when arrays of nucleosomes were visualized byby SWI/SNF. Yet SWI/SNF is known to be able to transla-atomic force microscopy before and after remodeling bytionally reposition nucleosomes (Jaskelioff et al., 2000;human SWI/SNF, they showed clear changes in the distri-Whitehouse et al., 1999). A simple hypothesis to resolvebution of nucleosome positions (Schnitzler et al., 2001).these observations is that translational repositioning byThe fact that all remodeling complexes can causeSWI/SNF is accomplished by a mechanism that is dis-changes in the translational position of nucleosomestinct from the classical sliding mechanism and that
has led to speculation that this is achieved by a commonallows exposure of DNA within the region bound by
mechanism that entails sliding. However, other mecha-a histone octamer. Any mechanism that involves the
nisms can also result in changes in translational posi-formation of an altered conformation of the nucleosome,
tion: a conformationally altered nucleosome could col- accomplished by changes in the DNA path, the shapelapse to a canonical nucleosome structure that has an of the histone octamer, or both, is consistent with allaltered position (Figure 3B), or there could be partial or characteristics for SWI/SNF remodeling. For example,complete release of the histone octamer followed by one such model (Figure 3B) proposes that the energyrebinding at a new location (Lorch et al., 1999; Studitsky of ATP hydrolysis is used to generate a high-energyet al., 1994). One reason to invoke mechanisms for re- intermediate that stochastically collapses to several dis-modeling other than sliding is that sliding cannot explain tinct final remodeled nucleosomal states, some of whichhow substantial tracts of DNA can be made accessible involve translational movement and some of which doin regions of tightly spaced nucleosomes. Sliding mech- not. Models of this type are also consistent with theanisms require, by definition, translational repositioning known ability of SWI/SNF complexes to form stably re-to expose DNA. Thus, sliding will not increase the modeled dinucleosome-like structures (Figure 2B) and toamount of exposed DNA; it will simply change the loca- promote the transfer of histone octamers (Figure 2C), bothtion of the exposed DNA. Mechanisms that could expose of which could occur via the high-energy intermediate.DNA sequences within the boundaries of the histone In contrast to SWI/SNF family complexes, all of theoctamer, therefore, without a requirement for transla- data for ISWI-based complexes are consistent with slid-tional repositioning (e.g., Figure 3B) would facilitate DNA ing of the DNA being their main mechanism. First, aexposure in regions of closely packed nucleosomes. timecourse analysis of the repositioning of nucleosomesInitial characterization of products of the remodeling by NURF was consistent with gradual movement of thereaction suggests that SWI/SNF has the ability to cause histones along the DNA (Hamiche et al., 1999). Second,conformational changes that expose nucleosomal DNA all the products generated by ISWI-based complexes inon the surface of the histone octamer. the sliding, assembly, or spacing assays have so far shown
SWI/SNF family members can increase the DNase characteristics of canonical nucleosomes. Third, SNF2H,and restriction enzyme sensitivity of DNA sites within a the human homolog of ISWI, shows at least 100-fold lowermononucleosome (Kingston and Narlikar, 1999). This is activity on mononucleosomes without flanking DNA than
on nucleosomal arrays, even though it binds mononucleo-achieved even though the mononucleosomes do not

Review479
somes with nanomolar affinity (Aalfs et al., 2001). This has as well as substrate specificity. The Acf1 subunit in theACF complex increases the ability of ISWI to assembleled to speculation that SNF2H requires flanking DNA onto
which it can slide the histone octamer. chromatin from histones and DNA (Figure 2E) by �30-fold (Ito et al., 1999). Consistent with this result, ACFIt is therefore possible that SWI/SNF and ISWI-based
complexes expose nucleosomal DNA by different mech- and CHRAC, both of which contain Acf1, are 10-foldbetter than ISWI alone at translational repositioning ofanisms. A mechanism based on classical sliding will
lead to identical amounts of movement of the entry and the histone octamer (Figure 2D) (Langst and Becker,2001b). Acf1 also alters the predominant product: ISWIexit points of the DNA in the same direction (Figure 3A).
One way to generate such movement involves twisting alone moves the histone octamer to the ends of thefragments of DNA that are used in the positioning analy-of the double helix “into” the nucleosome at the entry
point, with the twist propagating through the nucleo- sis, but it moves the octamer predominantly to the centerof the DNA when Acf1 is present. In contrast, NURFsome to the exit point (“twist diffusion;” Luger et al.,
1997). An alternative mechanism, consistent with the moves the histone octamer to positions that are interme-diate between the center and the ends of the DNA (Ha-characteristics of SWI/SNF function, results in minor
movements of the DNA at the entry and exit points to- miche et al., 1999). This effect appears to require only theNURF301 subunit in addition to ISWI (Xiao et al., 2001).ward each other (Figure 3B) with either both or only
one DNA end involved in the movement. This might be The extra subunits in CHRAC confer a different substratespecificity; unlike ISWI alone, CHRAC can translationallycoupled to changes in histone-DNA or histone-histone
contacts that result in stable exposure of a segment of reposition nucleosomes that lack H3 and H2A N-terminaltails (Clapier et al., 2001).DNA, possibly in the form of a DNA loop (Figure 3B).
The ability of SWI/SNF and ISWI to extrude DNA from One explanation for the different outcomes in thesesliding assays is based on the hypothesis that ISWIinverted repeats (Figure 2F) suggested that both these
complexes can introduce changes in superhelicity by acting alone can generate rapid movement of the his-tone octamer. In the absence of the remaining subunits,twisting the DNA (Havas et al., 2000). It has been pro-
posed that the ability to cause these changes in super- the octamer predominantly adopts the most thermody-namically stable position, which is often at the ends ofhelicity contributes directly to disruption of histone-DNA
contacts (Gavin et al., 2001). Recent work with ISWI the DNA. The additional subunits may exert their effectby binding to the DNA ends and forcing the histoneshows, however, that introduction of single-strand nicks
into nucleosomal DNA, which is predicted to dissipate octamer to adopt intermediate positions. Whether theseobservations are relevant to function in vivo, where thereany superhelicity, does not inhibit remodeling (Langst
and Becker, 2001a). In fact, one specifically positioned are very few if any ends of double-stranded DNA, isunclear. Alternatively, additional subunits might causenick caused a small increase in the ability of ISWI to slide
the histone octamer. Thus, generation of superhelicity changes in position because they directly alter funda-mental properties of ISWI mechanism.might not be part of the mechanism of ISWI action,
but might be a consequence of a related event (e.g., Less is known about the role of the additional subunitsin the SWI/SNF and Mi-2 based complexes. Initial worktransient unpairing of basepairs) that leads to disruption
of histone-DNA contacts. suggests that Mi-2 activity can be enhanced by up totwo orders of magnitude in the NuRD complex (WangWhile there are clues about the ways in which ATP-
dependent remodeling complexes alter chromatin and Zhang, 2001), and that SWI/SNF components canincrease BRG1 function under conditions of limiting en-structure, we are still in the early stages of understand-
ing the molecular details of remodeling mechanisms. zyme and substrate concentrations (Kingston and Narli-kar, 1999). These effects could be caused by changesFurther work, including detailed kinetic and thermody-
namic analyses of these enzymes coupled with struc- in substrate binding and/or enzymatic activity.tural information on the products, will be required tounderstand the molecular strategies used by this class Implications for In Vivo Functions of ATP-Dependentof molecular motors and to elucidate differences and Chromatin Remodelerssimilarities in function. At the simplest level, all ATP-dependent remodeling
complexes help regulate transcription by regulating theaccess of nucleosomal DNA to various transcription fac-Roles of the Remaining Subunits
The discovery that the central ATPase subunit alone has tors. However, the challenges of exposing DNA differaccording to local nucleosome organization: some pro-remodeling activity raised obvious questions about the
roles of the remaining subunits of remodeling com- moter regions contain regularly spaced nucleosomes,while others have gaps in nucleosome organization. Atplexes. In principle, the remaining subunits can perform
two distinct types of functions. They can modulate the promoters where multiple closely spaced nucleosomesocclude different factor binding sites, the DNA has toremodeling activity of the ATPase subunit, or they can
be involved in targeting of the remodeling complex to be made accessible at several sites without significantlyaltering the translational position of the histone octa-specific promoters directly or via interactions with tran-
scriptional activators. Very little is currently known about mers. This is different from the task of creating DNAaccess in regions of low nucleosome density, wherethe roles of these subunits in targeting; more is known
concerning the mechanistic effects, particularly with there is sufficient space for moving the nucleosomesand DNA can be exposed by creating nucleosome-freeISWI-based complexes.
In the ISWI-based complexes, the remaining subunits regions. Exposing DNA from densely packed nucleo-somes also differs from the essential task, often theaffect both the efficiency and outcome of remodeling

Cell480
responsibility of ATP-dependent remodelers, of creating Other modifications, such as methylation, ubiquitina-tion, and phosphorylation have also been discoveredappropriately spaced nucleosomes throughout the ge-and shown to be crucial in regulation of transcriptionnome following replication. As described above, ATP-(Berger, 2001). Here, we briefly summarize the vast liter-dependent remodeling can in theory be accomplishedature on functional consequences of acetylation andby sliding mechanisms, or instead by mechanisms thatdeacetylation and the nature of the complexes that per-do not require translational movement of the histoneform these modifications. We then discuss the regula-octamer. It appears that nature has evolved differenttory interactions between these complexes and theclasses of ATP-dependent remodelers that use differentATP-dependent remodeling complexes and transcrip-mechanisms, each suited to a specific biological task.tion machinery.The proposed ability of SWI/SNF-based complexes
to expose nucleosomal DNA without requiring slidingFunctional Consequences of Modificationsof the histone octamer would allow it to function atThere are multiple mechanisms by which acetylation ofpromoters, such as the mouse mammary tumor virusthe histone tails might facilitate transcription. In vitro,promoter (Deroo and Archer, 2001), which have closelyhistone acetylation has been shown to enhance the ac-spaced nucleosomes. On the other hand, the ability tocessibility of DNA to restriction enzymes and transcrip-translationally reposition nucleosomes would allowtion factors (Lee et al., 1993; Anderson et al., 2001;ISWI-based complexes to establish specifically spacedSewack et al., 2001). This might be caused in part bynucleosomal structures at various promoters. Indeed,the lowered positive charge on acetylated N termini andISWI-based complexes have been implicated in estab-a consequent lowered stability of interaction with DNA.lishing X chromosome structure in male Drosophila andHistone acetylation can also decrease compaction ofnucleosome positions at the hsp26, hsp70, and yeastnucleosomal arrays by disrupting internucleosomal in-early meiotic gene promoters (Deuring et al., 2000). Anteractions made via the histone tails (Tse et al., 1998).important goal over the coming years will be to developA third possibility is that acetylated residues in differenttechniques to determine whether or not a nucleosomecombinations can be involved in recruiting additionalhas been remodeled, and how it has been remodeled,transcription factors. When other tail modifications areon a given segment of DNA in vivo.taken into account, the combinatorial possibilities ex-pand rapidly. This has led to the hypothesis that certainCovalent Modificationscombinations of modifications in one or more tails actHyperacetylation of lysines in the N-terminal tails of thesequentially or concomitantly to form a histone codecore histones was proposed to be involved in activation(Strahl and Allis, 2000). Recognition of this code viaof transcription almost 40 years ago and has subse-binding of specific regulatory proteins is proposed toquently been strongly correlated with active genes (All-lead to additional downstream events.frey et al., 1964). The relatively recent discovery and
characterization of complexes that can add and removeHistone Acetyltransferase Complexesacetyl groups and the development of stringent antibod-There are two main classes of histone acetyltransfer-ies to specifically acetylated histone tails has helpedases (HATs), the type A nuclear HATs and the type B
provide a more comprehensive picture of these initialcytoplasmic HATs. Here we focus on nuclear regulatory
correlations.complexes, which mainly contain type A HATs. Three
Studies of bulk acetylation levels have shown that up families of Type A HATs have been identified (Figure 4).to 13 of the 30 tail lysine residues in a histone octamer All of them share a highly conserved motif containingare acetylated (Roth et al., 2001). Genetic studies in an Acetyl-CoA binding site, and several of these HATscombination with ChIP studies suggest that this steady- have been shown to have activity on histones in vitro.state level of acetylation is maintained by the opposing However, when in vivo they always seem to act as partactions of histone acetyltransferase (HAT) and histone of large complexes.deacetylase (HDAC) complexes (Reid et al., 2000; Vogel- The different complexes have different subunit com-auer et al., 2000). It is believed that targeting of HAT positions and different histone specificities (Table 1).and HDAC complexes to promoter regions then creates Correspondingly, these complexes appear to be in-specific patterns of hyper- and hypoacetylation in a back- volved in distinct biological functions (Roth et al., 2001).ground of global acetylation that correlate with transcrip- For example, genetic and biochemical studies implicatetion activation and repression, respectively. Indeed, ChIP the Spt3, 7, and 8 proteins, which are unique to thestudies using antibodies to specifically acetylated histone SAGA complex, in stabilizing TBP binding to the TATAtails have shown that increased acetylation in promoter box, suggesting that SAGA might function as a coactiva-proximal regions of specific genes correlates with re- tor at the site of initiation in addition to its acetylationcruitment of HAT complexes and increased gene ex- activity. In contrast, Spt16, a subunit of NuA3, or itspression (Kuo et al., 2000). Hypoacetylation at specific mammalian homologs have been implicated in tran-promoters has analogously been correlated with recruit- scriptional elongation and replication, events that mightment of HDAC complexes to repressed genes (Khochbin require more large-scale acetylation over several kb. Aet al., 2001). The discovery that certain HAT complexes distinct role for NuA4 is suggested based on its HAT,contained general transcription factors and that certain EsaI, which, unlike GCN5, is essential in yeast and isHDAC complexes contained known transcriptional re- homologous to MOF, the HAT implicated in 2-fold upreg-pressors further strengthened the correlations between ulation of the Drosophila male X chromosome. Mice thathyperacetylation and activation and between hypoacet- are homozygous for deletion of the HAT proteins p300,
CBP, PCAF, or GCN5 exhibit distinct developmental de-ylation and repression.

Review481
Table 2. Subunits in the Sin3 and NuRD Complexes
Sin3 Complex NuRD Complex
HDAC1 HDAC1HDAC2 HDAC2RbAp46 RbAp46RbAp48 RbAp48
Mi-2�, Mi-2�
MTA1MTA2MBD3p66
Sin3SAP18SAP30
Adapted from Ahringer, 2000.
target the complexes to distinct genes. Indeed, whereasSAGA and NuA4 can be functionally recruited by theglucocorticoid receptor and acidic activators such asGal4-VP16 (Brown et al., 2000; Roth et al., 2001), thedistinct HAT complexes ADA and NuA3 do not appear tointeract with these activators, leading to the speculationthat these might be involved in more global, nontargetedacetylation events. The subunits may also differentiallymodulate HAT activity; GCN5-containing complexeshave different substrate specificities than isolated GCN5(Brownell et al., 1996; Grant et al., 1999). Finally, there aremany examples where sequence-specific transcriptionfactors directly affect HAT activity (Roth et al., 2001) and
Figure 4. Representative Members of the Three Different HAT Familieswhere adjacent histone modifications such as phos-phorylation and methylation regulate HAT activity (Ber-ger, 2001; Zhang and Reinberg, 2001).fects, further suggesting differences in function of these
highly related HAT subunits.Histone Deacetylase ComplexesThe different functions of HAT complexes are likelyHDAC proteins identified to date fall in three maincaused by differences in the histone residues that areclasses (reviewed in Khochbin et al., 2001). The Sin3acetylated and differences in targeting. Each complexcomplex and NuRD complex contain members of thecontains a specific set of non-HAT subunits, which mightclass I HDAC family, HDAC1 and HDAC2. Complexesinteract with different sequence-specific activators thatcontaining members of class II HDACs have yet to bepurified. Two complexes containing a member of classIII HDACs, Sir2, have been recently identified and showTable 1. Some Representative HAT Complexesdistinct deacetylase activities. Sir2 is involved in hetero-
yADA ySAGA hPCAF yNuA3 yNuA4 chromatin silencing at silent mating loci, telomeres, andGCN5 GCN5 hPCAF Sas3 Esa1 ribosomal DNA (reviewed in Moazed, 2001).
Ada1 The histone specificities of the class I HDAC familyAda2 Ada2 hAda2 complexes are just beginning to be characterized. Re-Ada3 Ada3 hAda3
cent in vivo studies using highly specific antibodies sug-Ada5/Spt20gest that the yeast homolog of HDAC1, Rpd3, deacety-Spt3 hSpt3 Spt16lates all sites except lysine 16 on histone 4, a site thatSpt7
Spt8 is strongly linked to heterochromatic silencing and aTra1 PAF400 Tra1 site whose acetylation by dMOF is linked to activationTAFII90 PAF65� TAFII30 of the Drosophila male X chromosome (Roth et al., 2001;TAFII61/68 TAF15/20
Suka et al., 2001). This is consistent with Rdp3 havingTAFII60 PAF65�distinct functions from the Sir 2 deacetylases. Further,TAFII23/25 TAF30this suggests that Rpd3 does not act antagonisticallyTAFII17/20 TAF31
Sin4 to EsaI, the yeast homolog of dMOF (Figure 4).Ahc1 Analogous to the HAT complexes, the different sub-
Act3/ARP unit compositions of Sin3 and NuRD suggest that theyAct1
participate in distinct biological functions (Table 2). TheEpl1biochemical activities of the Sin3 and NuRD complexesEaf3have not yet been compared side by side, but it is possi-
Adapted from Roth et al., 2001.ble that quantitative comparison of their deacetylase
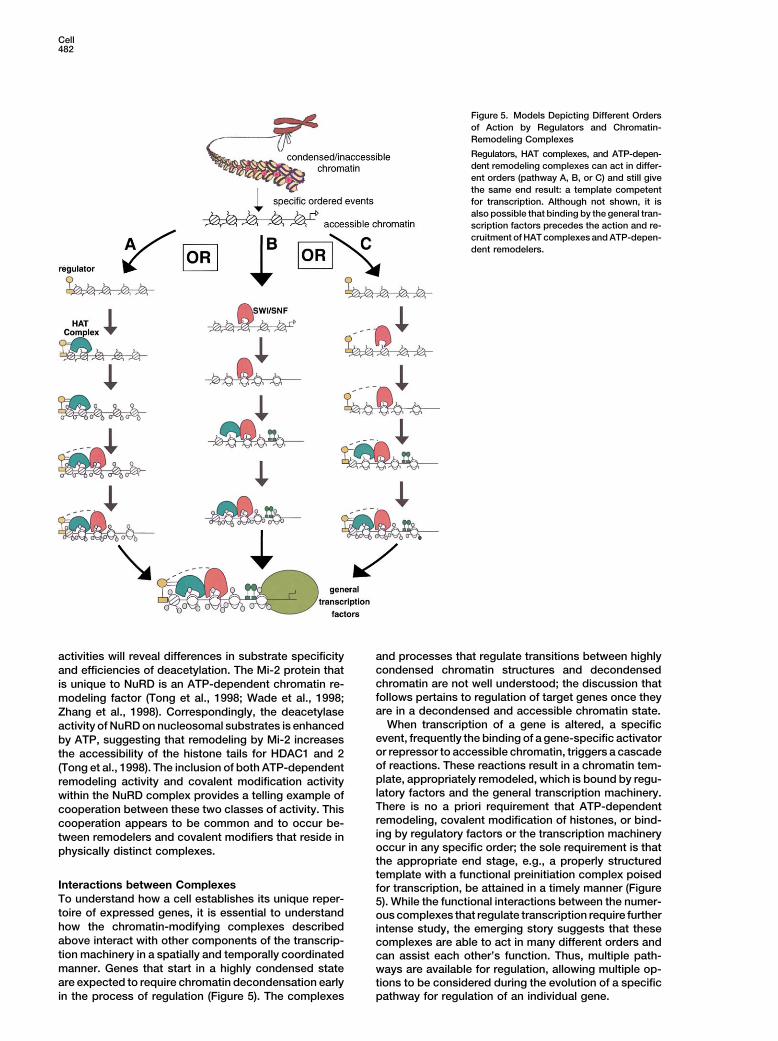
Cell482
Figure 5. Models Depicting Different Ordersof Action by Regulators and Chromatin-Remodeling Complexes
Regulators, HAT complexes, and ATP-depen-dent remodeling complexes can act in differ-ent orders (pathway A, B, or C) and still givethe same end result: a template competentfor transcription. Although not shown, it isalso possible that binding by the general tran-scription factors precedes the action and re-cruitment of HAT complexes and ATP-depen-dent remodelers.
activities will reveal differences in substrate specificity and processes that regulate transitions between highlycondensed chromatin structures and decondensedand efficiencies of deacetylation. The Mi-2 protein thatchromatin are not well understood; the discussion thatis unique to NuRD is an ATP-dependent chromatin re-follows pertains to regulation of target genes once theymodeling factor (Tong et al., 1998; Wade et al., 1998;are in a decondensed and accessible chromatin state.Zhang et al., 1998). Correspondingly, the deacetylase
When transcription of a gene is altered, a specificactivity of NuRD on nucleosomal substrates is enhancedevent, frequently the binding of a gene-specific activatorby ATP, suggesting that remodeling by Mi-2 increasesor repressor to accessible chromatin, triggers a cascadethe accessibility of the histone tails for HDAC1 and 2of reactions. These reactions result in a chromatin tem-(Tong et al., 1998). The inclusion of both ATP-dependentplate, appropriately remodeled, which is bound by regu-remodeling activity and covalent modification activitylatory factors and the general transcription machinery.within the NuRD complex provides a telling example ofThere is no a priori requirement that ATP-dependentcooperation between these two classes of activity. Thisremodeling, covalent modification of histones, or bind-cooperation appears to be common and to occur be-ing by regulatory factors or the transcription machinerytween remodelers and covalent modifiers that reside inoccur in any specific order; the sole requirement is thatphysically distinct complexes.the appropriate end stage, e.g., a properly structuredtemplate with a functional preinitiation complex poised
Interactions between Complexes for transcription, be attained in a timely manner (FigureTo understand how a cell establishes its unique reper- 5). While the functional interactions between the numer-toire of expressed genes, it is essential to understand ous complexes that regulate transcription require furtherhow the chromatin-modifying complexes described intense study, the emerging story suggests that theseabove interact with other components of the transcrip- complexes are able to act in many different orders andtion machinery in a spatially and temporally coordinated can assist each other’s function. Thus, multiple path-manner. Genes that start in a highly condensed state ways are available for regulation, allowing multiple op-are expected to require chromatin decondensation early tions to be considered during the evolution of a specific
pathway for regulation of an individual gene.in the process of regulation (Figure 5). The complexes

Review483
Regulation of any biological reaction can be accom- moters (Deroo and Archer, 2001; Hassan et al., 2001a)In addition to nuclear receptors, transcription activatorsplished kinetically, by regulating the rate of a rate-
determining step, or thermodynamically, by regulating the as divergent as erythroid Kruppel-like factor, C/EBP�,c-Myc, MyoD, HSF1, and EBNA2 have also been foundaffinity of an enzyme for its substrate. For example,
prokaryotic transcription initiation can be regulated, to recruit SWI/SNF to specific promoters and, in somecases, to activate gene expression (Hassan et al., 2001a;among other mechanisms, by changing the rate of pro-
moter DNA melting and by modulating the stability of Peterson and Workman, 2000; Sullivan et al., 2001).Although the molecular details of the association be-RNA polymerase binding. While eukaryotic transcription
can be regulated at many more steps, similar concepts tween SWI/SNF and transcription activators remainlargely unknown, there is evidence that points to theapply. Several previous reviews have addressed how
ATP-dependent remodeling complexes and histone importance of specific features of transcription activa-tion domains. A study of the chimeric Gal4-VP16 acidicmodifiers can individually contribute to transcription by
increasing transcription factor access to DNA. Here we transcription activator has shown that targeting is re-duced or eliminated by mutations that disrupt its acidicexplore an additional level of regulation, in which ATP-
dependent remodelers, histone modifiers, and tran- residues (Hassan et al., 2001a; Peterson and Workman,2000). Additionally, hydrophobic residues within thescription factors can kinetically and/or thermodynami-
cally modulate each other’s activities in the context of transcription activation domain of the human heat shockfactor HSF1 have been shown to be important for medi-activation or repression on different genes.ating SWI/SNF recruitment (Sullivan et al., 2001).
Nuclear HAT complexes and transcription coactiva-How Are Chromatin-Modifying Complexestors with intrinsic HAT activities have also been found toRecruited to Specific DNA Loci?interact with transcription activators (Roth et al., 2001).While both ATP-dependent remodeling and histoneSimilar to SWI/SNF targeting, the direct contact betweenmodifying complexes can bind nonspecifically to DNA,an activator and a HAT complex will contribute to ana wide variety of data implies that the activity of theseincreased affinity for a particular DNA region, thus facili-complexes is regulated both spatially and temporally.tating recruitment to specific DNA loci. For example, theFor example, genome-wide expression studies per-transactivation domain of GR has been shown to interactformed with mutants of ySwi/Snf subunits have revealedwith CBP, a protein with intrinsic HAT activity (Deroothat this complex is involved in the expression of onlyand Archer, 2001). As described above for SWI/SNF,6% of all yeast genes (Holstege et al., 1998; Sudarsanamit has been demonstrated that transcription activatorset al., 2000). Similarly, Gcn5p-containing HAT com-containing acidic domains can directly interact withplexes seem to be required for the expression of onlySAGA and NuA4 and target these HAT complexes to5% of yeast genes (Holstege et al., 1998). How is thisspecific promoters (Hassan et al., 2001a; Peterson andspecificity achieved? Accumulating evidence indicatesWorkman, 2000). Recently, Tra1p, a common subunitthat at least some sequence-specific transcription fac-of SAGA and NuA4, has been shown to mediate thetors bind directly to both ATP-dependent chromatin-interaction of these complexes with acidic transcriptionremodeling and histone acetyltransferase complexes toactivators (Brown et al., 2001).“target” these activities to specific locations.
Direct interactions between activators and these com-plexes can have three functional consequences that Repressors Can Target ATP-Dependentmay affect the overall rate of chromatin remodeling. Remodelers and HDACsThese interactions can specifically increase the affinity In contrast to ySWI/SNF, which can be targeted by acidicof the remodeling complex for a given DNA region due activators, Isw2p has been shown to be targeted toto the contacts between the activator and the complex. promoters of early mitotic genes by the transcriptionalThis would increase overall activity by increasing the repressor UME6 (Goldmark et al., 2000). In further con-local concentration of the remodeling complex. Targeting trast to the examples discussed above, the recruitmentcan also regulate remodeling by increasing the rate at of Isw2p leads to the formation of inaccessible chroma-which the remodeling complex binds to the chromatin tin structure proximal to the Ume6p binding site and,template, if the binding event is rate limiting. Finally, consequently, represses gene expression (Goldmark etinteractions with the specific DNA binding factors may al., 2000). The Sin3-Rpd3 complex, a member of thedirectly affect the activity of the remodeling complex. yeast HDAC family, is one of the most extensively stud-Many studies have provided examples of the first role ied histone deacetylase complexes. This complex hasof targeting with recent data providing initial examples been shown to be targeted to specific promoters by theof the third role. same transcription repressor Ume6p, resulting in local
histone deacetylation and transcription repression (Faz-zio et al., 2001).Transcription Activators Target SWI/SNF
and HATs Another example demonstrating the generality of tran-scription repressor-mediated recruitment is the tar-A widely accepted model has SWI/SNF targeted to spe-
cific promoters by direct interactions with sequence- geting of the NuRD complex, which contains an ATP-dependent chromatin remodeler, Mi-2, and a histonespecific transcription activators. Examples of targeting
exist both in vivo and in vitro, from yeast to man. As deacetylase (Khochbin et al., 2001; Kingston and Narli-kar, 1999). Upon T cell activation, the DNA binding pro-an illustration, human SWI/SNF participates in nuclear
receptor-mediated transcriptional regulation, and the tein Ikaros recruits NuRD to regions of heterochromatin(Kim et al., 1999). It has been proposed that this recruit-glucocorticoid receptor (GR) and the estrogen receptor
have been shown to recruit SWI/SNF to responsive pro- ment either maintains an inactive chromatin state or

Cell484
converts an accessible chromatin conformation to an and p300/CBP function interdependently; the interde-pendence between ATP-dependent remodelers and his-inaccessible structure. The transcription repressor,tone-modifying enzymes could be a general feature.Kap-1, can also target NuRD to specific promoters to
Analogous to the effects of sequence-specific activa-repress gene expression (Schultz et al., 2001). Anothertors, these two types of complexes could help each othercandidate for this type of recruitment is the hunchbackin multiple ways. Direct physical interactions between theprotein of Drosophila, which associates with the Dro-ATP-dependent remodelers and chromatin modifiers couldsophila Mi-2 homolog (Kehle et al., 1998).increase their affinity for the chromatin template. SuchDNA microarray analyses, biochemical studies, anddirect interactions could also affect the activities of eachgenetic studies indicate that yeast and human SWI/SNFcomplex. Finally, alteration of the chromatin templateare also involved in gene repression. The transcriptionby one complex could make it a better substrate for thecorepressors Hir1p and Hir2p of yeast can recruit ySwi/other complex. For example, remodeling of the nucleo-Snf to a responsive promoter and repress gene expres-somes by ATP-dependent remodelers may increase thesion (Dimova et al., 1999), and in Drosophila and mam-accessibility of the histone N termini for acetylation ormals SWI/SNF has been found to participate in Rb-E2Fdeacetylation. Alternatively, ATP-dependent remodel-repression pathways (Zhang and Dean, 2001). Evidenceers might bind more strongly to, or dissociate moreconnecting SWI/SNF and deacetylation came from bio-slowly from, nucleosomes having N termini acetylatedchemical studies which have shown that componentsat specific positions. Acetylation states of histones mayof the Sin3-HDAC complex copurify with a BRG1-con-directly affect the kinetics of ATP-dependent remodel-taining SWI/SNF complex, and mSin3 interacts withers’ activities. Thus far, few biochemical analyses havecomponents of SWI/SNF in vitro (Sif et al., 2001).afforded the temporal resolution necessary to resolveIt is therefore clear that sequence-specific DNA bind-these issues, and detailed quantitative studies will being regulatory factors can directly target ATP-dependentrequired in the future to dissect out the different waysremodeling complexes, HAT complexes, and HDACin which HAT complexes and ATP-dependent remodelingcomplexes to specific locations. In certain instances,complexes can affect each other’s activities.there is concerted recruitment of a remodeler and HDAC
Two studies raise the possibility that acetyltrasferaseactivity. In many cases, targeting has mainly a thermody-complexes might stabilize the interaction of yeast SWI/namic effect, with the direct contacts between the gene-SNF complexes with the template. Work performed us-specific regulatory protein and the chromatin-modifyinging an altered PHO5 promoter in yeast suggested thatcomplex contributing to increased binding energy. DNAthe Gcn5p-containing SAGA complex might serve twobinding factors can also have a kinetic effect by directlyfunctions (Syntichaki et al., 2000). The catalytic Gcn5paffecting the activity of a remodeling complex. Factorssubunit first acetylates a promoter region, and then thesuch as Twist and E1a have been shown to directly alterbromodomain of Gcn5p is proposed to stabilize theacetyltransferase activity of p300 (Roth et al., 2001). Tobinding of Swi/Snf to the newly hyperacetylated nucleo-date, there is no example of an activator or repressorsomes. Biochemical analysis provides a somewhat dif-directly altering the rate of chromatin remodeling, al-ferent story that supports the basic notion that acetyl-though it is early days and such kinetic effects are likelytransferases can stabilize binding of yeast SWI/SNF. Itto be uncovered. These targeting mechanisms likelywas shown using an in vitro system that SWI/SNF isserve to initiate a cascade of events at a given promoterpreferentially bound on an acetylated template, implyingthat results in local alteration of chromatin structure tothat acetylation stabilizes SWI/SNF association (Hassanfacilitate formation of an active or repressed state.et al., 2001b).
ISWI family ATP-dependent remodeling complexes canHow Do Chromatin Remodelers and HATs and the also interact with covalent modifying complexes. AlthoughTranscription Machinery Function Together? Isw2p and Sin3p-Rpd3p have unique biochemical activi-There is strong evidence to support the common-sense ties, genetic studies and genome-wide microarray anal-notion that ATP-dependent remodelers and covalent yses indicate that ISW2 and SIN3-RPD3 function syner-modifiers work together to regulate gene expression. A gistically to regulate gene expression (Fazzio et al., 2001;functional link between ATP-dependent remodeling and Goldmark et al., 2000). In an in vitro transcription system,HAT complexes was first suggested by genetic studies p300 and P/CAF enhanced NURF-mediated transcrip-in yeast (Pollard and Peterson, 1997; Roberts and Win- tional activation from a chromatin template (Mizuguchiston, 1997). Mutations in subunits of the SAGA complex et al., 2001). And yet, acetylated histones were not found(excluding GCN5) were lethal in combination with muta- to increase the amount of NURF-remodeled template,tions in the ySwi/Snf complex, though none of the single as judged by the steady-state accessibility of restrictionmutants showed any severe growth defects, indicating enzymes or micrococcal nuclease (Mizuguchi et al.,a synergistic interaction between components of these 2001). It remains possible, however, that acetylatedtwo complexes. In mammalian cells, ChIP experiments chromatin might have changed the rate of remodelinghave found that both BRG1 and p300/CBP are present catalyzed by NURF.on Estrogen receptor-responsive promoters following es- The importance of changing the rate of remodeling istrogen treatment of MCF7 cells, consistent with cooper- demonstrated by a study of the PHO5 promoter (Bar-ative interactions (DiRenzo et al., 2000; Shang et al., baric et al., 2001). It had been accepted that Gcn5p has2000). Increasing levels of histone acetylation, by treat- little effect on transcription activation of the yeast PHO5ing cells with an HDAC inhibitor while simultaneously gene; however, Barbaric et al. showed that whereas theoverexpressing BRG1, synergistically enhances ER- steady-state level of the PHO5 message is not altered
in gcn5 mutants, the rate of nucleosome remodeling atresponsive gene expression. This suggests that BRG1

Review485
the PHO5 locus is decreased (Barbaric et al., 2001). ture, including changes in nucleosome position (Lee andGarrard, 1991).Thus, HATs can increase the rate of gene induction,
without affecting steady-state expression levels, appar- Chromatin-modifying/remodeling complexes can fa-cilitate specific steps in transcription. ATP-dependentently by stimulating the rate of remodeling.remodeling complexes can increase binding by gene-specific activators and by components, such as TBP,
Is There a General Order of Recruitment? of the preinitiation complex (Burns and Peterson, 1997;It appears that there is no obligate order for function of Cote et al., 1994; Imbalzano et al., 1994). Acetylation ofATP-dependent remodelers and covalent modifiers that the template can also increase binding of transcriptionis general for all promoters. Rather, it seems that each factors (Sewack et al., 2001). Both HAT complexes andindividual promoter will work using a set order of action ATP-dependent remodeling complexes can significantlyby these complexes that differs from promoter to pro- increase the rate of overall transcription from nucleoso-moter. On the yeast HO promoter, the recruitment of mal templates in defined in vitro transcription systemsATP-dependent chromatin remodelers precedes that of (Ikeda et al., 1999; Neely et al., 1999).HAT complexes (Cosma et al., 1999; Krebs et al., 1999). It appears that each gene has “chosen” a specific path-The reverse order of complex recruitment has been ob- way to achieve appropriate regulation from a menu ofserved on the IFN-� promoter and for retinoic acid- different possible pathways. This is consistent with recentinduced transcription (Agalioti et al., 2000; Dilworth et biochemical studies that emphasize the requirement foral., 2000). The precise order seems to depend upon the specific chromatin-modifying complexes in an individualnature of the promoter, the complement of transcription system. For example, activation of the �-globin locusfactors present, and the chromatin structure in which via action of the transcription factor EKLF requires athe promoter resides. specific SWI/SNF-family remodeling complex (Hassan
Indeed, for a particular gene, the requirement for spe- et al., 2001a). Transcriptional activation in vitro of a spe-cific chromatin remodelers may vary. This is best re- cific promoter by hormone receptors in a defined in vitrovealed by the cell-cycle dependence of ATP-dependent system remarkably requires one particular SWI/SNFremodeling and HAT activities in yeast. Activation of family complex, while a different SWI/SNF family com-the inducible GAL1 promoter during interphase requires plex will not function (Lemon et al., 2001). It appears thatGcn5p but not ySwi/Snf. Induction of this gene in late these very specific requirements for a given complex onmitosis, however, requires both ySwi/Snf and the Gcn5p a given promoter reflect the particular pathway that hascomplex. This indicates that ATP-dependent remodel- evolved to work on that promoter, not a requirementing complexes are needed to assist HAT-dependent that is general for the activator involved or for othergene expression when chromatin is highly condensed promoters.(Krebs et al., 2000). This small subset of examples serves to make the
point that there is no obligate order for function of thetranscription machinery with respect to the chromatin-Many Paths Can Lead to the Same Place;modifying machinery. The sole requirement is that theCoordination with the Transcription Machineryend point, which is the structure of the template andThe analysis described above implies that ATP-depen-association of appropriate components of the generaldent remodelers and chromatin-modifying complexestranscription machinery, be reached in a timely mannercan work in any order, and that these two types of(Figure 5). Each component is capable of functioning incomplexes can directly influence each other’s activity.the absence of the other components. What is apparentFor example, acetylation can enhance ATP-dependentfrom the studies described above, and is the key toremodeling, and remodeling can increase the rate of cova-allowing different pathways to occur on different genes,lent changes such as deacetylation (DiRenzo et al., 2000;is that each step is able to facilitate another step. Re-Guschin et al., 2000). While space constraints preclude amodeling can facilitate transcription factor binding, anddetailed discussion of interactions involving the generalfactor binding can facilitate remodeling. Thus, each playertranscription machinery, we describe below a few exam-can help the other, and a pathway can be chosen thatples to make the point that the components of the gen-offers the most parsimonious solution to the problem oferal transcription machinery can assist in chromatin re-achieving specific and robust transcriptional regulation.modeling, and that chromatin-remodeling complexes
can increase function of the transcription machinery.AcknowledgmentsThus, the notion that there is no obligate order of action
for ATP-dependent remodeling and covalent modificationWe apologize to our colleagues whose primary papers could not
can be extended to state that there is no obligate order be cited due to space constraints. We thank I. King, J. Dennis, R.of action for chromatin-modifying complexes and com- Lake, and D. Reinberg for comments.plexes in the general transcription machinery.
ReferencesTranscription factors and regulatory complexes canbind chromatin prior to recruitment of chromatin-modi-
Aalfs, J.D., Narlikar, G.J., and Kingston, R.E. (2001). Functional dif-fying complexes, and are capable of altering chromatinferences between the human ATP-dependent nucleosome remodel-
structure. Biochemical studies have shown that binding ing proteins BRG1 and SNF2H. J. Biol. Chem. 276, 34270–34278.of activators such as GAL4 can displace nucleosomes
Agalioti, T., Lomvardas, S., Parekh, B., Yie, J., Maniatis, T., andin cis and in trans to create nucleosome-free regions Thanos, D. (2000). Ordered recruitment of chromatin modifying and(Workman and Kingston, 1992). Elongation by RNA poly- general transcription factors to the IFN-beta promoter. Cell 103,
667–678.merase can also cause changes in the chromatin struc-

Cell486
Ahringer, J. (2000). NuRD and SIN3 histone deacetylase complexes Gavin, I., Horn, P.J., and Peterson, C.L. (2001). SWI/SNF chromatinremodeling requires changes in DNA topology. Mol. Cell 7, 97–104.in development. Trends Genet. 16, 351–356.
Goldmark, J.P., Fazzio, T.G., Estep, P.W., Church, G.M., and Tsuki-Allfrey, V.G., Faulkner, R., and Mirsky, A.E. (1964). Acetylation andyama, T. (2000). The Isw2 chromatin remodeling complex repressesmethylation of histones and their possible role in the regulation ofearly meiotic genes upon recruitment by Ume6p. Cell 103, 423–433.RNA synthesis. Proc. Natl. Acad. Sci. USA 51, 786–794.
Grant, P.A., Eberharter, A., John, S., Cook, R.G., Turner, B.M., andAnderson, J.D., Lowary, P.T., and Widom, J. (2001). Effects of his-Workman, J.L. (1999). Expanded lysine acetylation specificity oftone acetylation on the equilibrium accessibility of nucleosomal DNAGcn5 in native complexes. J. Biol. Chem. 274, 5895–5900.target sites. J. Mol. Biol. 307, 977–985.
Guschin, D., Wade, P.A., Kikyo, N., and Wolffe, A.P. (2000). ATP-Barbaric, S., Walker, J., Schmid, A., Svejstrup, J.Q., and Horz, W.dependent histone octamer mobilization and histone deacetylation(2001). Increasing the rate of chromatin remodeling and gene activa-mediated by the Mi-2 chromatin remodeling complex. Biochemistrytion—a novel role for the histone acetyltransferase Gcn5. EMBO J.39, 5238–5245.20, 4944–4951.
Guyon, J.R., Narlikar, G.J., Sif, S., and Kingston, R.E. (1999). StableBerger, S.L. (2001). An embarrassment of niches: the many covalentremodeling of tailless nucleosomes by the human SWI-SNF com-modifications of histones in transcriptional regulation. Oncogeneplex. Mol. Cell. Biol. 19, 2088–2097.20, 3007–3013.
Guyon, J.R., Narlikar, G.J., Sullivan, E.K., and Kingston, R.E. (2001).Boyer, L.A., Logie, C., Bonte, E., Becker, P.B., Wade, P.A., Wolffe,Stability of a human SWI-SNF remodeled nucleosomal array. Mol.A.P., Wu, C., Imbalzano, A.N., and Peterson, C.L. (2000). FunctionalCell. Biol. 21, 1132–1144.delineation of three groups of the ATP-dependent family of chroma-
tin remodeling enzymes. J. Biol. Chem. 275, 18864–18870. Hamiche, A., Sandaltzopoulos, R., Gdula, D.A., and Wu, C. (1999).ATP-dependent histone octamer sliding mediated by the chromatinBrehm, A., Langst, G., Kehle, J., Clapier, C.R., Imhof, A., Eberharter,remodeling complex NURF. Cell 97, 833–842.A., Muller, J., and Becker, P.B. (2000). dMi-2 and ISWI chromatin
remodelling factors have distinct nucleosome binding and mobiliza- Hassan, A.H., Neely, K.E., Vignali, M., Reese, J.C., and Workman,tion properties. EMBO J. 19, 4332–4341. J.L. (2001a). Promoter targeting of chromatin-modifying complexes.
Front. Biosci. 6, D1054–D1064.Brown, C.E., Lechner, T., Howe, L., and Workman, J.L. (2000). Themany HATs of transcription coactivators. Trends Biochem. Sci. 25, Hassan, A.H., Neely, K.E., and Workman, J.L. (2001b). Histone ace-15–19. tyltransferase complexes stabilize swi/snf binding to promoter
nucleosomes. Cell 104, 817–827.Brown, C.E., Howe, L., Sousa, K., Alley, S.C., Carrozza, M.J., Tan,S., and Workman, J.L. (2001). Recruitment of HAT complexes by Havas, K., Flaus, A., Phelan, M., Kingston, R., Wade, P.A., Lilley,direct activator interactions with the ATM-related Tra1 subunit. Sci- D.M., and Owen-Hughes, T. (2000). Generation of superhelical tor-ence 292, 2333–2337. sion by ATP-dependent chromatin remodeling activities. Cell 103,
1133–1142.Brownell, J.E., Zhou, J., Ranalli, T., Kobayashi, R., Edmondson, D.G.,Roth, S.Y., and Allis, C.D. (1996). Tetrahymena histone acetyltrans- Holstege, F.C., Jennings, E.G., Wyrick, J.J., Lee, T.I., Hengartner,ferase A: a homolog to yeast Gcn5p linking histone acetylation to C.J., Green, M.R., Golub, T.R., Lander, E.S., and Young, R.A. (1998).gene activation. Cell 84, 843–851. Dissecting the regulatory circuitry of a eukaryotic genome. Cell 95,
717–728.Burns, L.G., and Peterson, C.L. (1997). The yeast SWI-SNF complexfacilitates binding of a transcriptional activator to nucleosomal sites Ikeda, K., Steger, D.J., Eberharter, A., and Workman, J.L. (1999).in vivo. Mol. Cell. Biol. 17, 4811–4819. Activation domain-specific and general transcription stimulation by
native histone acetyltransferase complexes. Mol. Cell. Biol. 19,Clapier, C.R., Langst, G., Corona, D.F., Becker, P.B., and Nightin-855–863.gale, K.P. (2001). Critical role for the histone H4 N terminus in nucleo-
some remodeling by ISWI. Mol. Cell. Biol. 21, 875–883. Imbalzano, A.N., Kwon, H., Green, M.R., and Kingston, R.E. (1994).Facilitated binding of TATA-binding protein to nucleosomal DNA.Cosma, M.P., Tanaka, T., and Nasmyth, K. (1999). Ordered recruit-Nature 370, 481–485.ment of transcription and chromatin remodeling factors to a cell
cycle and developmentally regulated promoter. Cell 97, 299–311. Ito, T., Levenstein, M.E., Fyodorov, D.V., Kutach, A.K., Kobayashi,R., and Kadonaga, J.T. (1999). ACF consists of two subunits, Acf1Cote, J., Quinn, J., Workman, J.L., and Peterson, C.L. (1994). Stimu-and ISWI, that function cooperatively in the ATP-dependent cataly-lation of GAL4 derivative binding to nucleosomal DNA by the yeastsis of chromatin assembly. Genes Dev. 13, 1529–1539.SWI/SNF complex. Science 265, 53–60.Jaskelioff, M., Gavin, I.M., Peterson, C.L., and Logie, C. (2000). SWI-Deroo, B.J., and Archer, T.K. (2001). Glucocorticoid receptor-medi-SNF-mediated nucleosome remodeling: role of histone octamer mo-ated chromatin remodeling in vivo. Oncogene 20, 3039–3046.bility in the persistence of the remodeled state. Mol. Cell. Biol. 20,
Deuring, R., Fanti, L., Armstrong, J.A., Sarte, M., Papoulas, O., Pres- 3058–3068.tel, M., Daubresse, G., Verardo, M., Moseley, S.L., Berloco, M., et
Kehle, J., Beuchle, D., Treuheit, S., Christen, B., Kennison, J.A.,al. (2000). The ISWI chromatin-remodeling protein is required forBienz, M., and Muller, J. (1998). dMi-2, a hunchback-interactinggene expression and the maintenance of higher order chromatinprotein that functions in polycomb repression. Science 282, 1897–structure in vivo. Mol. Cell 5, 355–365.1900.
Dilworth, F.J., Fromental-Ramain, C., Yamamoto, K., and Chambon,Khochbin, S., Verdel, A., Lemercier, C., and Seigneurin-Berny, D.P. (2000). ATP-driven chromatin remodeling activity and histone(2001). Functional significance of histone deacetylase diversity.acetyltransferases act sequentially during transactivation by RAR/Curr. Opin. Genet. Dev. 11, 162–166.RXR in vitro. Mol. Cell 6, 1049–1058.Kim, J., Sif, S., Jones, B., Jackson, A., Koipally, J., Heller, E., Wi-Dimova, D., Nackerdien, Z., Furgeson, S., Eguchi, S., and Osley,nandy, S., Viel, A., Sawyer, A., Ikeda, T., et al. (1999). Ikaros DNA-M.A. (1999). A role for transcriptional repressors in targeting thebinding proteins direct formation of chromatin remodeling com-yeast Swi/Snf complex. Mol. Cell 4, 75–83.plexes in lymphocytes. Immunity 10, 345–355.
DiRenzo, J., Shang, Y., Phelan, M., Sif, S., Myers, M., Kingston, R.,Kingston, R.E., and Narlikar, G.J. (1999). ATP-dependent remodelingand Brown, M. (2000). BRG-1 is recruited to estrogen-responsiveand acetylation as regulators of chromatin fluidity. Genes Dev. 13,promoters and cooperates with factors involved in histone acetyla-2339–2352.tion. Mol. Cell. Biol. 20, 7541–7549.Krebs, J.E., Kuo, M.H., Allis, C.D., and Peterson, C.L. (1999). CellFazzio, T.G., Kooperberg, C., Goldmark, J.P., Neal, C., Basom, R.,cycle-regulated histone acetylation required for expression of theDelrow, J., and Tsukiyama, T. (2001). Widespread collaboration ofyeast HO gene. Genes Dev. 13, 1412–1421.Isw2 and Sin3-Rpd3 chromatin remodeling complexes in transcrip-
tional repression. Mol. Cell. Biol. 21, 6450–6460. Krebs, J.E., Fry, C.J., Samuels, M.L., and Peterson, C.L. (2000).

Review487
Global role for chromatin remodeling enzymes in mitotic gene ex- mono- and polynucleosomes by atomic force microscopy employingcarbon nanotube tips. Mol. Cell. Biol. 21, 8504–8511.pression. Cell 102, 587–598.
Schultz, D.C., Friedman, J.R., and Rauscher, F.J., 3rd. (2001). Tar-Kuo, M.H., vom Baur, E., Struhl, K., and Allis, C.D. (2000). Gcn4geting histone deacetylase complexes via KRAB-zinc finger pro-activator targets Gcn5 histone acetyltransferase to specific promot-teins: the PHD and bromodomains of KAP-1 form a cooperative uniters independently of transcription. Mol. Cell 6, 1309–1320.that recruits a novel isoform of the Mi-2alpha subunit of NuRD.Kwon, H., Imbalzano, A.N., Khavari, P.A., Kingston, R.E., and Green,Genes Dev. 15, 428–443.M.R. (1994). Nucleosome disruption and enhancement of activatorSewack, G.F., Ellis, T.W., and Hansen, U. (2001). Binding of TATAbinding by a human SW1/SNF complex. Nature 370, 477–481.binding protein to a naturally positioned nucleosome is facilitatedLangst, G., and Becker, P.B. (2001a). ISWI induces nucleosomeby histone acetylation. Mol. Cell. Biol. 21, 1404–1415.sliding on nicked DNA. Mol. Cell 8, 1085–1092.Shang, Y., Hu, X., DiRenzo, J., Lazar, M.A., and Brown, M. (2000).Langst, G., and Becker, P.B. (2001b). Nucleosome mobilization andCofactor dynamics and sufficiency in estrogen receptor-regulated
positioning by ISWI-containing chromatin- remodeling factors. J.transcription. Cell 103, 843–852.
Cell Sci. 114, 2561–2568.Sif, S., Saurin, A.J., Imbalzano, A.N., and Kingston, R.E. (2001). Purifi-
Langst, G., Bonte, E.J., Corona, D.F., and Becker, P.B. (1999). cation and characterization of mSin3A-containing Brg1 and hBrmNucleosome movement by CHRAC and ISWI without disruption or chromatin remodeling complexes. Genes Dev. 15, 603–618.trans-displacement of the histone octamer. Cell 97, 843–852.
Strahl, B.D., and Allis, C.D. (2000). The language of covalent histoneLee, M.S., and Garrard, W.T. (1991). Transcription-induced nucleo- modifications. Nature 403, 41–45.some ‘splitting’: an underlying structure for DNase I sensitive chro-
Studitsky, V.M., Clark, D.J., and Felsenfeld, G. (1994). A histonematin. EMBO J. 10, 607–615.octamer can step around a transcribing polymerase without leaving
Lee, D.Y., Hayes, J.J., Pruss, D., and Wolffe, A.P. (1993). A positive the template. Cell 76, 371–382.role for histone acetylation in transcription factor access to nucleo-
Sudarsanam, P., Iyer, V.R., Brown, P.O., and Winston, F. (2000).somal DNA. Cell 72, 73–84.
Whole-genome expression analysis of snf/swi mutants of Saccharo-Lee, K.M., Sif, S., Kingston, R.E., and Hayes, J.J. (1999). hSWI/SNF myces cerevisiae. Proc. Natl. Acad. Sci. USA 97, 3364–3369.disrupts interactions between the H2A N-terminal tail and nucleoso- Suka, N., Suka, Y., Carmen, A.A., Wu, J., and Grunstein, M. (2001).mal DNA. Biochemistry 38, 8423–8429. Highly specific antibodies determine histone acetylation site usageLemon, B., Inouye, C., King, D.S., and Tjian, R. (2001). Selectivity of in yeast heterochromatin and euchromatin. Mol. Cell 8, 473–479.chromatin-remodeling cofactors for ligand-activated transcription. Sullivan, E.K., Weirich, C.S., Guyon, J.R., Sif, S., and Kingston, R.E.Nature 414, 924–928. (2001). Transcriptional activation domains of human heat shock fac-Logie, C., Tse, C., Hansen, J.C., and Peterson, C.L. (1999). The tor 1 recruit human SWI/SNF. Mol. Cell. Biol. 21, 5826–5837.core histone N-terminal domains are required for multiple rounds Syntichaki, P., Topalidou, I., and Thireos, G. (2000). The Gcn5 bromo-of catalytic chromatin remodeling by the SWI/SNF and RSC com- domain co-ordinates nucleosome remodelling. Nature 404, 414–417.plexes. Biochemistry 38, 2514–2522.
Tong, J.K., Hassig, C.A., Schnitzler, G.R., Kingston, R.E., andLorch, Y., Zhang, M., and Kornberg, R.D. (1999). Histone octamer Schreiber, S.L. (1998). Chromatin deacetylation by an ATP-depen-transfer by a chromatin-remodeling complex. Cell 96, 389–392. dent nucleosome remodelling complex. Nature 395, 917–921.Luger, K., Mader, A.W., Richmond, R.K., Sargent, D.F., and Rich- Tse, C., Sera, T., Wolffe, A.P., and Hansen, J.C. (1998). Disruptionmond, T.J. (1997). Crystal structure of the nucleosome core particle of higher-order folding by core histone acetylation dramatically en-at 2.8 A resolution. Nature 389, 251–260. hances transcription of nucleosomal arrays by RNA polymerase III.
Mol. Cell. Biol. 18, 4629–4638.Meersseman, G., Pennings, S., and Bradbury, E.M. (1992). Mobilenucleosomes—a general behavior. EMBO J. 11, 2951–2959. Tsukiyama, T., and Wu, C. (1995). Purification and properties of an
ATP-dependent nucleosome remodeling factor. Cell 83, 1011–1020.Mizuguchi, G., Vassilev, A., Tsukiyama, T., Nakatani, Y., and Wu, C.(2001). ATP-dependent nucleosome remodeling and histone hyper- Vogelauer, M., Wu, J., Suka, N., and Grunstein, M. (2000). Globalacetylation synergistically facilitate transcription of chromatin. J. histone acetylation and deacetylation in yeast. Nature 408, 495–498.Biol. Chem. 276, 14773–14783. Wade, P.A., Jones, P.L., Vermaak, D., and Wolffe, A.P. (1998). AMoazed, D. (2001). Common themes in mechanisms of gene silenc- multiple subunit Mi-2 histone deacetylase from Xenopus laevis co-
fractionates with an associated Snf2 superfamily ATPase. Curr. Biol.ing. Mol. Cell 8, 489–498.8, 843–846.Narlikar, G.J., Phelan, M.L., and Kingston, R.E. (2001). GenerationWang, H.B., and Zhang, Y. (2001). Mi2, an auto-antigen for dermato-and interconversion of multiple distinct nucleosomal states as amyositis, is an ATP-dependent nucleosome remodeling factor. Nu-mechanism for catalyzing chromatin fluidity. Mol. Cell 8, 1219–1230.cleic Acids Res. 29, 2517–2521.Neely, K.E., Hassan, A.H., Wallberg, A.E., Steger, D.J., Cairns, B.R.,Whitehouse, I., Flaus, A., Cairns, B.R., White, M.F., Workman, J.L.,Wright, A.P., and Workman, J.L. (1999). Activation domain-mediatedand Owen-Hughes, T. (1999). Nucleosome mobilization catalysedtargeting of the SWI/SNF complex to promoters stimulates tran-by the yeast SWI/SNF complex. Nature 400, 784–787.scription from nucleosome arrays. Mol. Cell 4, 649–655.Workman, J.L., and Kingston, R.E. (1992). Nucleosome core dis-Peterson, C.L., and Workman, J.L. (2000). Promoter targeting andplacement in vitro via a metastable transcription factor-nucleosomechromatin remodeling by the SWI/SNF complex. Curr. Opin. Genet.complex. Science 258, 1780–1784.Dev. 10, 187–192.Xiao, H., Sandaltzopoulos, R., Wang, H.M., Hamiche, A., Ranallo,Pollard, K.J., and Peterson, C.L. (1997). Role for ADA/GCN5 prod-R., Lee, K.M., Fu, D., and Wu, C. (2001). Dual functions of largestucts in antagonizing chromatin-mediated transcriptional repression.NURF subunit NURF301 in nucleosome sliding and transcriptionMol. Cell. Biol. 17, 6212–6222.factor interactions. Mol. Cell 8, 531–543.
Reid, J.L., Iyer, V.R., Brown, P.O., and Struhl, K. (2000). CoordinateZhang, H.S., and Dean, D.C. (2001). Rb-mediated chromatin struc-regulation of yeast ribosomal protein genes is associated with tar-ture regulation and transcriptional repression. Oncogene 20, 3134–geted recruitment of Esa1 histone acetylase. Mol. Cell 6, 1297–1307.3138.
Roberts, S.M., and Winston, F. (1997). Essential functional interac-Zhang, Y., and Reinberg, D. (2001). Transcription regulation by his-
tions of SAGA, a Saccharomyces cerevisiae complex of Spt, Ada, tone methylation: interplay between different covalent modificationsand Gcn5 proteins, with the Snf/Swi and Srb/mediator complexes. of the core histone tails. Genes Dev. 15, 2343–2360.Genetics 147, 451–465.
Zhang, Y., LeRoy, G., Seelig, H.P., Lane, W.S., and Reinberg, D.Roth, S.Y., Denu, J.M., and Allis, C.D. (2001). Histone acetyltransfer- (1998). The dermatomyositis-specific autoantigen Mi2 is a compo-ases. Annu. Rev. Biochem. 70, 81–120. nent of a complex containing histone deacetylase and nucleosomeSchnitzler, G.R., Cheung, C.L., Hafner, J.H., Saurin, A.J., Kingston, R.E., remodeling activities. Cell 95, 279–289.and Lieber, C.M. (2001). Direct imaging of human SWI/SNF-remodeled