characterization and description of novel actinomycete...
TRANSCRIPT

Chapter 3
Characterization and Description
of Novel Actinomycete Taxa

Characterization and Description of Novel Actinomycete Taxa
85
3.1. INTRODUCTION
Of the 10 potentially novel species of actinobacteria recoverd from Hundung
limestone site, three (3) were characterized by polyphasic taxonomy. Polyphasic
taxonomy - a term coined by Colwell (1970) - incorporates all genotypic, phenotypic,
and phylogenetic information. Genotypic information is derived from nucleic acids
present in the cell, whereas phenotypic information is derived from proteins and their
functions, different chemotaxonomic markers, and a wide range of other expressed
factors. These are used for the delineation of taxa at all levels (Murray et al. 1990).
3.2. METHODOLOGY
3.2.1. Phenotypic characterization
The classical phenotypic characteristics of bacteria comprise morphological,
physiological, and biochemical features. Morphological features of a bacterium
include both cellular and colonial characteristics. The biochemical features include
data on presence or absence of various enzymes, diversity of carbon or energy sources
used and metabolic versatility whereas chemotaxonomic features include information
on the various chemical constituents of the cell.
3.2.1.1. Morphological characterization
Microscopy
The axenic actinomycete strain was cultivated aerobically on GM1 (28 °C) for 5
weeks for strain MBRL 34 and SCNA (28 °C) for 2 weeks for strains MBRL 201 and

Characterization and Description of Novel Actinomycete Taxa
86
MBRL 251. Morphology of the spores and mycelia was observed by light microscopy
(Olympus BH2) and scanning electron microscopy (Quanta 200, FEI). Bacterial cells
producing spore-bearing mycelia were cultured by cover-slip techniques. The fully
grown mycelia on the cover-slip were then observed under the light microscope. For
electron microscopy, the mycelia were sputter coated with gold and observed under
scanning electron microscope (SEM) (Williams and Davies 1967).
Growth pattern on various culture media
The actinomycete strain was grown on International Streptomyces Project (ISP) media
(Shirling and Gottlieb, 1966), tryptic soy agar (TSA, Difco), SCNA, Czapek’s Dox
agar (CDA) and Nutrient agar (NA) at 28 °C for 7 d. The colour of aerial and
substrate mycelia and production of soluble pigments was then observed. The colony
colour was determined in accordance with the Inter-Society Colour Council –
National Bureau of Standards (ISCC-NBS) colour charts (Kelly 1964).
3.2.1.2. Biochemical characterization
Aesculin decomposition (Gordon et al. 1974)
Actinomycete cultures were inoculated into aesculin broth, and observed for growth
and blackening of the medium after 2 weeks.
Casein hydrolysis (Leboffe and Pierce 2008)
Actinomycete cultures were spot-inoculated on Skim Milk Agar and incubated
aerobically for 1 d. Formation of halo zones around the growth indicates positive
result.

Characterization and Description of Novel Actinomycete Taxa
87
Catalase activity (Leboffe and Pierce 2008)
A loopful of actinomycete culture was transferred to a microscope slide. One or two
drops of H2O2 (3 %, v/v) was then aseptically placed directly onto the bacteria and
observed for the formation of bubbles.
Citrate utilization (Leboffe and Pierce 2008)
Using an inoculating needle and light inoculum, the Simmons Citrate tube was
stabbed with the test actinomycete and incubated for 4 d. A change in colour of the
medium from green to deep blue indicates a positive reaction.
Gelatin liquefaction (Leboffe and Pierce 2008; Collins et al. 2004)
Nutrient Gelatin medium was stab-inoculated with heavy inoculum of the
actinomycete culture, and was kept incubated for 1 week. The tube was then kept at 4
°C for 1 h and checked for liquefaction.
Indole production (Cappucino and Sherman 2004)
Actinomycete culture was inoculated in Nutrient broth medium and incubated for 2 d.
Formation of red ring on addition of 0.6 ml of Kovac’s reagent indicates production
of indole from tryptophan in the culture medium.
Methyl Red-Voges Proskaeur test (Leboffe and Pierce 2008)
Actinomycete cultures were inoculated into MR-VP broth and incubated at 37 °C for
2-3 d. For Methyl Red test, 3 drops of Methyl Red reagent was added and checked for
red colour formation (positive reaction). For Voges Proskaeur test, 0.6 ml of VP

Characterization and Description of Novel Actinomycete Taxa
88
Reagent A and 0.2 ml of VP Reagent B were added and kept for 10 min to 1 h.
Formation of red colour indicates positive result.
Nitrate reduction (Leboffe and Pierce 2008, Lanyi 1987)
Actinomycete cultures were inoculated into Nitrate Broth. After 2 d of incubation,
eight drops each of Reagent A and Reagent B were added to each tube, and allowed to
stand undisturbed for 10 min. Formation of red colour indicates positive result. A
small amount of zinc powder was added to cultures not producing a colour change.
The development of red colour confirmed negative result for nitrate reduction.
Oxidase activity (Kovacs 1956)
Sterile discs (HiMedia) were impregnated with tetramethyl-p-phenylenediamine
solution (1 %, w/v) and air dried. To the disc, a single colony of the actinomycete
culture was smeared and observed for change into violet colour within 20 s.
Production of acids from carbohydrates (Leboffe and Pierce 2008)
The Phenol red broth is used to test for acid production from carbohydrates (fructose,
glucose, lactose, maltose, mannitol and sucrose). 5 ml each of the medium were
distributed in screw-capped test tubes and autoclaved. The medium was impregnated
with a carbohydrate disc (HiMedia) using a sterilized forcep. The medium was then
inoculated with actinomycete mycelial suspension and incubated for 4 d. The medium
was then checked for change of colour from red to yellow.

Characterization and Description of Novel Actinomycete Taxa
89
Starch hydrolysis (Leboffe and Pierce 2008)
Actinomycete cultures were spot-inoculated on the starch agar plate and incubated
aerobically for 2 d. The cultures were covered with Gram’s iodine and observed for
clearing zones surrounding the cultures. Presence of halo zones indicate positive
result.
Tween hydrolysis (Sierra 1957)
To Tween hydrolysis medium, the actinomycete strain was cross-streaked and
inoculated for 1-2 weeks. The plates were then observed for formation of fuzzy halos
around the actinomycete lawns.
Urea hydrolysis (Leboffe and Pierce 2008)
A heavy inoculum of the actinomycete culture was inoculated on the sterilized
Rustigian and Stuart’s Urea Broth. The tube was inoculated for 4 d, and examined for
change of colour from red to pink. Colour change indicates positive result.
Utilization of sole C source (Shirling and Gottlieb 1966)
Actinomycete cell (or mycelium) suspension was inoculated in the basal medium
containing the carbon source (0.5 %, w/v). The carbon sources used for the test were
adonitol, arabinose, cellobiose, dulcitol, fructose, galactose, inositol, inulin, lactose,
maltose, mannitol, mannose, melibiose, raffinose, rhamnose, ribose, salicin, sodium
malate, sorbose, sorbitol, succinic acid, trehalose, xylitol and xylose. Appearance of
growth was observed after 2 weeks.

Characterization and Description of Novel Actinomycete Taxa
90
Utilization of sole N source (Shirling and Gottlieb 1966)
The basal medium containing Nitrogen source (0.5 %, w/v) was inoculated with
actinomycete suspension and growth was observed after 2 weeks. The nitrogen
sources used in the study were L-alanine, L-arginine hydrochloride, L-asparagine, L-
aspartic acid, L-cysteine, L-cystine, L-glutamic acid, L-glutamine, glycine, L-
histidine hydrochloride, L-isoleucine, L-leucine, L-methionine, L-ornithine,
potassium nitrate, L-phenylalanine, L-proline, L-serine, L-threonine, L-tryptophan, L-
tyrosine and L-valine.
3.2.1.3. Chemotaxonomy
Whole cell amino acid and sugar analyses (Staneck and Roberts 1974; Tang et al.
2009)
Purified cell wall of the actinomycete culture was prepared by hydrolysis of
approximately 3 mg dry cell mass in a sealed ampoule with 0.2 ml HCl (6 M) at 120
°C for 12 h. For determination of the type of diamino pimelic (DAP) acid, 5 l of the
whole cell hydrolysate was spotted at the base of the cellulose TLC sheet (Merck
2330); 1 l meso-DAP (10 mM) was applied separately on the side as the standard
reference. Ascending TLC was performed with the solvent system, Methanol-distilled
water-HCl (6 M)-Pyridine (80:26:4:10 v/v/v/v) for approximately 3.5 h. The
chromatogram was then air-dried and spots were visualized by spraying with
ninhydrin reagent (0.2 % in acetone). For whole cell sugars, the hydrolysis was done
with HCl (0.2 M) at 120 °C for 2 h. 80 l each of the whole cell hydrolysates, 1-
phenyl-3-methyl-5-pyrazolone solution (0.25 M in methanol) and NaOH solution (0.2

Characterization and Description of Novel Actinomycete Taxa
91
M) were mixed and incubated for 30 min at 70 °C. The mixture was then cooled to
room temperature and neutralized with HCl (0.2 M) to pH 7.0, and extracted with
isoamyl acetate. The organic phase was discarded. The extraction process was
continued for three more cycles (except that chloroform was used instead of isoamyl
acetate in the last cycle). The aqueous phase was collected and analyzed in HPLC.
Determination of menaquinones (Collins et al. 1977; Tamaoko et al. 1983)
Approximately 100 mg freeze-dried actinomycete culture was mixed with 20 mL
chloroform-methanol (2:1, v/v) and the suspension was kept incubated under shaking
condition overnight. The biomass was then removed by filtration and the extract
evaporated to dryness under reduced pressure at low temperature (37 °C). Analytical
TLC of quinones was performed in silica gel TLC plate GF254 (Merck) for 20 min
using toluene as the developing solvent. Usually menaquinones have an Rf value of ~
0.7 and was detected after brief irradiation with short-wave ultraviolet light (254 nm).
The menaquinone fraction was then extracted from the gel, dissolved in methanol and
concentrated under reduced pressure. The final extract was then analyzed in HPLC.
Polar lipids analysis (Lechevalier et al. 1977; Minnikin et al. 1984)
In a 50 ml centrifuge tube containing approximately 1 g cell mass, 15 ml methanol
was added and kept in boiling water bath (100 °C) for 5 min. The mixture was cooled
to room temperature, followed by addition of 10 ml chloroform and equal volume of
NaCl solution (2 %, w/v). The tube was vigorously shaken (10 min) and then
centrifuged (8000 rpm, 10 min). The organic phase was dried on a rotary evaporator.
The lipid extract was then dissolved in 200 l chloroform-methanol (2:1, v/v) and

Characterization and Description of Novel Actinomycete Taxa
92
used for chromatographic analysis of the phospholipids. 10 l of the whole cell lipid
extract was applied on one corner of silica gel 60 thin-layer chromatographic plates
(10x10 cm) and developed in two dimensions using the following solvent systems:
Chloroform-methanol-water (65:24:4, v/v/v) in the first direction, and chloroform-
acetic acid-methanol-water (80:18:12:5, v/v/v/v) in the second direction. The
chromatogram was visualized by spraying with molybdatophosphoric acid and
charring at 150 °C for 2 min.
Determination of cellular fatty acids (Sasser 1990)
The five steps involved in the preparation of cells for fatty acid composition analysis
by Gas chromatography is mentioned below:
i. Harvesting – About 40 mg cell mass from a quadrant streaked plate was
harvested using a 4 mm loop and placed in a clean 13x100 culture tube.
ii. Saponification – 1 ml of Reagent 1 was added to the tube. The tube was
securely sealed with teflon lined cap, vortexed briefly and heated in a boiling
water bath for 5 min, and then vigorously vortexed for 5-10 s and returned to
the water bath to complete the 30 min heating.
iii. Methylation – The cooled tube was then uncapped and 2 ml of Reagent 2 was
added. The tube was capped again and briefly vortexed. After vortexing, the
tube was heated for 10 min at 80 °C.
iv. Extraction – 1.25 ml of Reagent 3 was added to the cooled tube followed by
recapping and gentle tumbling on a clinical rotator for about 10 min. The tubes
were uncapped and the aqueous (lower) phase was pipetted out and discarded.

Characterization and Description of Novel Actinomycete Taxa
93
v. Base wash – About 3 ml of Reagent 4 was added to the remaining organic
phase in the tube. The tube was recapped and tumbled for 5 min. Following
uncapping, about 2/3 of the organic phase was pipetted into a GC vial and
capped.
The fatty acid methyl esters (FAMEs) obtained was then analyzed by GC (Agilent
Technologies 7890A GC System) by using the Microbial Identification Software
Package (Sherlock Version 6.1; MIDI database: TSBA6, MIDI Inc).
3.2.1.4. Physiological characterization (Goodfellow 1986)
Growth temperature
Actinomycete cultures were inoculated on TSA and incubated at different
temperatures (5, 15, 28, 37, 42, 50 and 60 °C). Growth was observed at regular
intervals upto the 7th
day.
Growth at different concentration of NaCl
TSA was prepared with different NaCl concentrations (0, 2, 5, 7 and 10 % w/v). The
actinomycete cultures were inoculated on the plates and growth observed at regular
intervals upto the 7th
day.
pH tolerance
pH of TSA was adjusted with different buffer solutions to give a pH range of 4 – 10.
The petri dishes containing the media were inoculated and incubated at 28 °C.
Actinomycete cultures were examined for growth at regular intervals upto the 10th
day.

Characterization and Description of Novel Actinomycete Taxa
94
3.2.2. Genotypic characterization
3.2.2.1. Genomic DNA isolation and PCR amplification of 16S rRNA gene (Li et al.
2007)
The genomic DNA isolation and PCR amplification of 16S rRNA gene were
performed as described in Chapter 2.
3.2.2.2. Cloning of amplified 16S rRNA gene
Purification of amplified 16S rRNA gene
Purification of the amplified 16S rRNA gene product was done using the EasyPure
Quick Gel Extraction Kit (TransGen) as per the instruction manual.
Transformation
The purified PCR products were cloned using the TransGen TA kit (TransGen
Biotech) following the user’s instructions.
Analysis of recombinant clones by colony PCR
The cells were checked for transformation by PCR using the following mixture: 5 l
Buffer for Taq pol (with Mg2+
, 10X), 4 l dNTPs (2.5 mM), 1 l of each M13 +/-
primers (2.5 mM), 0.3 l Taq polymerase (2.5U/l), 37.7 l H2O and 1 l DNA
template.
3.2.2.3. Sequencing of cloned 16S rRNA gene
Sequencing was done by Sangon Sequencing Company, Shanghai, China. Sequences
obtained were then assembled using the SeqMan II (DNASTAR).The almost

Characterization and Description of Novel Actinomycete Taxa
95
complete 16S rRNA gene sequence of the actinomycete strain was submitted to the
EzTaxon-e server database (Kim et al. 2012; http://eztaxon-e.ezbiocloud.net/) and
aligned with the 16S rRNA gene sequences of other closely related species using
CLUSTAL X version 2.1 (Larkin et al., 2007). Phylogenetic analyses were performed
using the software package MEGA version 5.2 (Tamura et al. 2011). Distances (using
distance options according to Kimura’s two-parameter model; Kimura 1983) were
calculated and clustering was performed with the neighbour-joining (NJ) (Saitou and
Nei 1987), Maximum Likelihood (ML) (Felsenstein 1981) and Maximum Parsimony
(MP) (Kluge and Farris 1969) methods. To determine the support of each clade,
bootstrap analysis was performed with 1000 resamplings (Felsenstein 1985).
3.2.2.4. Purification of Genomic DNA for G+C content analysis and DNA-DNA
hybridization studies
Isolation of Genomic DNA
Actinomycete strains were cultured in TSB shake flasks (200 rpm, 37 °C, 7 d). Cell
biomass was collected by centrifugation (5,000 rpm) and washed twice with distilled
water. Approximately 1-2 g cell mass was taken in a 50 ml oak ridge tube, mixed with
7 ml TE buffer (1X) and 0.7 ml EDTA (0.5 M); 0.7 ml lysozyme (50 mg/ml) was then
added and the tube was kept incubated overnight under shaking conditions (200 rpm,
37 °C). This was followed by addition of 0.7 ml SDS (20 %, w/v) and 70 l
proteinase K (5 mg/ml) and the tube was then kept on water bath (55 °C, 2 h) after
thorough mixing. Equal volume of Phenol-Chloroform (1:1, v/v) was then added,
mixed and centrifuged (12,000 rpm, 10 min). The aqueous phase was transferred to a
fresh 50 ml oak ridge tube. The extraction process was repeated 2 more times. The

Characterization and Description of Novel Actinomycete Taxa
96
final aqueous extract was mixed with equal volume of isopropanol and 1/10 volume
of sodium acetate (3 M), and mixed carefully. The precipitated DNA was spooled out
in a fresh eppendorf tube, washed with ethanol (70 %) and dried until no smell of
alcohol was detected. Finally the DNA was dissolved in a minimum volume of TE
buffer (1X).
Purification of Genomic DNA
The DNA obtained by the above method is likely to contain significant amount of
RNA. RNAs were, therefore, degraded using RNA-degrading enzyme, RNase. The
volume of the DNA solution was made up to 500 l with TE (1X) and 15 l RNase A
(400 U/ml) and 150 l RNase T1 (400U/ml) were added to it, mixed and kept
incubated in a shaker (37 °C, 30 min). Extraction was done by adding equal volume
of Phenol-Chloroform-Isoamyl alcohol (25:24:1, v/v/v) followed by slow mixing and
centrifugation at 12,000 rpm. The aqueous phase was carefully transferred to a fresh
eppendorf tube, and the extraction was repeated for another round. The final
extraction was done using chloroform only. To the aqueous extract, an equal volume
of isopropanol was added to precipitate the DNA. The mixture was allowed to stand
for 10 min at room temperature followed by centrifugation. The supernatant was
carefully decanted, and the tube was allowed to dry until no smell of alcohol was
detected. The final purified DNA was then dissolved in 30 l sterile milliQ water.

Characterization and Description of Novel Actinomycete Taxa
97
Determination of purity and concentration of DNA
The purity and concentration of DNA was checked by monitoring the absorbances at
260 and 280 nm. Pure DNA has A260/ A280 between 1.7 and 1.9. For DNA with A260/
A280 ratio below 1.7 or above 1.9, DNA purification was repeated.
DNA concentration in the purified sample was calculated using the relation:
DNA concentration = 50 g/ml x A260 x dilution factor
where, 50 g/ml is the concentration of DNA
corresponding to A260 = 1
3.2.2.5. Determination of G+C content of genomic DNA
DNA enzymolysis
10 l purified DNA was heated for 10 min at 100 °C, followed by immediately
cooling on ice for 5 min. To the denatured DNA, 10 l RNase P1 (63 U/ml) was
added and incubated in water bath (50 °C, 1 h). It was incubated for another 1 h at 37
°C after addition of 10 l alkaline phosphatase (70 U/ml). The final mixture was used
for analysis of G+C content.
Measurement of G+C content (Mesbah et al., 1989)
The hydrolyzed DNA was analyzed in HPLC. The G+C content was calculated
according to the peak area of the deoxynucleotides in the sample chromatogram using
the following formula:
𝐆 + 𝐂 𝐜𝐨𝐧𝐭𝐞𝐧𝐭 (𝐦𝐨𝐥%) = 𝐆+𝐂
𝐆+𝐂+𝐓+𝐀 × 𝟏𝟎𝟎%

Characterization and Description of Novel Actinomycete Taxa
98
3.2.2.6. DNA-DNA hybridization (DDH) studies
Digestion of Genomic DNA
The purified DNA sample was fragmented to ~ 300 bp fragment in a sonicator (5 s
sonication followed by 5 s interval, 3 rounds). The DNA fragment was analyzed by
agarose gel electrophoresis (1 %, w/v) using DNA ladder (100 bp) as the reference.
Hybridization reaction by thermal renaturation method (DeLey et al. 1970)
The study was done using Lambda 35 UV/Vis Spectrophotometer (Perkin Elmer)
equipped with 6+6 Peltier Temperature Programmer (Perkin Elmer) and BG-Chiller
E15 (Baygene Biotech). DNA-DNA relatedness value below 70% is considered the
delineating limit for species determination (Wayne et al. 1987).
3.3. RESULTS
Based on the polyphasic characterization, the strains MBRL 34, MBRL 201 and
MBRL 251 were characterized as Micromonospora kangleipakensis sp. nov.,
Streptomyces manipurensis sp. nov. and Streptomyces hundungensis sp. nov.
respectively. The detailed characterization results of the three novel species are
discussed below.

Characterization and Description of Novel Actinomycete Taxa
99
3.3.1. Micromonospora kangleipakensis sp. nov.
3.3.1.1. Morphological characteristics
Strain MBRL 34T was isolated as a green colour forming colony from the GM1 plate.
The strain forms single spores (~0.87m) on short sporophores in the substrate
mycelium. Figure 3.1 shows its growth morphology on GM1 plates and as observed
under SEM.
Fig. 3.1. Growth morphology of MBRL 34T in GM1 and as observed under SEM after 35 d of
incubation, Bar 5 m.
The strain grew with colours ranging from orange to green (Table 3.1) in most of the
media tested, and no growth (or poor growth) is observed on ISP6, ISP7, NA and
TSA.

Characterization and Description of Novel Actinomycete Taxa
100
Table 3.1. Cultural characteristics of MBRL 34T on various ISP and other
selective media as observed using ISCC-NBS Color Chart
Medium Colour of mycelium
Aerial (Spore mass) Substrate
ISP2
ISP3
ISP4
ISP5
ISP6
ISP7
NA
SCNA
CDA
TSA
GM 1
Orange
Orange
Orange
Orange
NG*
PG#
PG#
Orange
Olive Green
PG#
Olive Green
Orange Yellow
Orange Yellow
Orange
Orange Yellow
-
-
-
Orange Yellow
Deep Green
-
Deep Green
* NG, No Growth; #PG, Poor Growth
3.3.1.2. Biochemical characteristics
The strain hydrolyzed casein, but not aesculin, gelatin, Tween 20, tyrosine and urea. It
was positive for catalase but negative for methyl red, indole production, nitrate
reduction, and citrate utilization tests. The strain utilized inulin and mannose as sole
carbon sources and L-alanine, L-glutamine, L-isoleucine, L-leucine, L-lysine
hydrochloride, L-methionine, L-serine, L-tyrosine and L-valine as sole nitrogen
sources. It could not utilize adonitol, arabinose, cellobiose, dulcitol, fructose,
galactose, inositol, lactose, maltose, mannitol, melibiose, raffinose, rhamnose, salicin,
sorbitol, sucrose, trehalose, xylose, L-arginine hydrochloride, L-asparagine, L-
cysteine, L-cystine, L-glycine, L-histidine, L-phenylalanine, L-proline, L-threonine
and L-tryptophan as either sole carbon or nitrogen sources. Table 3.2 lists the
biochemical characteristics for strain MBRL 34T.

Characterization and Description of Novel Actinomycete Taxa
101
Table 3.2. Biochemical characteristics of strain MBRL 34T
Characteristics Result Characteristics Result
Casein hydrolysis
Catalase activity
Citrate utilization
Gelatin liquefaction
Utilization of sole C sources
Adonitol
Arabinose
Cellobiose
Dulcitol
Fructose
Galactose
Inositol
Inulin
Lactose
Maltose
Mannitol
Mannose
Melibiose
Raffinose
Rhamnose
Salicin
Sorbitol
Trehalose
Xylose
+
+
-
-
-
-
-
-
-
-
-
+
-
-
-
+
-
-
-
-
-
-
-
Indole production
Methyl Red test
Nitrate reduction
Starch hydrolysis
Tween 20 hydrolysis
Urea hydrolysis
Utilization of sole N sources
L-Alanine
L-Arginine hydrochloride
L-Asparagine
L-Cysteine
L-Cystine
L-Glutamine
Glycine
L-Histidine hydrochloride
L-Isoleucine
L-Leucine
L-Methionine
L-Phenylalanine
L-Proline
L-Serine
L-Threonine
L-Tryptophan
L-Valine
-
-
-
-
-
-
+
+
+
-
-
+
-
-
+
+
+
-
-
+
+
-
+
Note: + positive; - negative
3.3.1.3. Chemotaxonomical characteristics
Strain MBRL 34T had meso-DAP as the diagnostic cell wall diamino acid. Galactose
(53.5 %) and glucose (23.1 %) were the main sugars detected in the whole cell
hydrolysates along with small amounts of mannose, xylose, rhamnose and ribose.
MK-10(H6) (36.6 %), MK-10(H2) (24.1 %), MK-11(H4) (16.4 %) and MK-10(H4)
(11.1 %) were the predominant menaquinones detected. The major polar lipids

Characterization and Description of Novel Actinomycete Taxa
102
detected were diphosphatidylglycerol (DPG), phosphatidylethanolamine (PE),
phosphatidylglycerol (PG), phosphatidylinositol (PI) and
phosphatidylinositolmannoside (PIM), with other unknown lipids (UL) (Figure 3.2).
The FAME profile (>1%) contained iso-C16:0 (36.2 %), iso-C15:0 (24.4 %), iso-C17:0
(8.1 %), C17:0 (5.2 %), Summed Feature 9 containing C17:1c and/or 10-methyl C16:0
(4.2%), 10-methyl C17:0 (3.4%), anteiso-C17:0 (2.8%), iso-C18:0 (2.2%), 2-OH C16:1
(2.0%), anteiso-C15:0 (1.9%), C18:0 (1.8%), iso-C14:0 (1.6%) and iso-H C16:1 (1.0%).
Fig. 3.2 Two-dimensional thin-layer chromatogram of polar lipids of strain MBRL 34T
(1, PIM; 2, PI; 3, PG; 4, PE; 5, DPG; 6, UL)
3.3.1.4. Physiological characteristics
The strain grows at the temperature range of 15-37 °C with optimum growth at 28 °C.
It grows over the pH range of 5-7 with optimum at pH 6. It has low tolerance of NaCl
and doesn’t grow at ≥ 2 % NaCl.
1
2
3 4
5
6

Characterization and Description of Novel Actinomycete Taxa
103
3.3.1.5. Genotypic characteristics
The GenBank accession number for the 16S rRNA gene sequence of strain MBRL
34T is JN560152. Comparison of 16S rRNA gene sequences of strain MBRL 34
T and
other members of the genus Micromonospora showed sequence similarities ranging
from 96.36 % to 98.39 % with 43 strains showing similarities above 97%, of which
six had similarities above 98% (M. echinaurantica DSM 43904T, 98.39%; M. eburnea
LK2-10T, 98.37%; M. coerulea DSM 43143
T, 98.25%; M. viridifaciens DSM 43909
T,
98.18%; M. echinofusca DSM 43913T, 98.11% and M. chaiyaphumensis MC5-1
T,
98.02%). In the phylogenetic tree (Figure 3.3) based on the NJ method, supported by
the ML and the MP methods, strain MBRL 34T forms a monophyletic clade with
Micromonospora coerulea DSM 43143T.
The Genomic DNA G+C content for strain MBRL 34T was found to be 73.46 mol%.
The DNA-DNA relatedness value between strain MBRL 34T and Micromonospora
coerulea DSM 43143T was calculated as 53.14% which is below the 70% delineating
limit for species determination.
3.3.1.6. Differentiating characteristics between strain MBRL 34T and DSM 43143
T
Apart from low DNA-DNA reassociation value, the strain MBRL 34T could be
differentiated from the type strain M. coerulea DSM 43143T by several phenotypic
characteristics as shown in Tables 3.3 and 3.4.

Characterization and Description of Novel Actinomycete Taxa
104
Table 3.3. Differential physiological and biochemical characteristics between
strain MBRL 34T and M. coerulea DSM 43143T
Characteristics MBRL 34T DSM 43143T
pH range for growth
Optimum pH for growth
Growth temperature range
Nitrate reduction
Utilization of sole C sources
Adonitol
Cellobiose
Dulcitol
Fructose
Galactose
Inositol
Lactose
Maltose
Mannitol
Melibiose
Raffinose
Utilization of sole N sources
L-Arginine
L-Asparagine
Glycine
L-Histidine
L-Isoleucine
L-Methionine
L-Phenylalanine
L-Proline
L-Threonine
L-Valine
G+C mol %
5-7
6
15-37
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
+
+
-
-
-
+
73.5
6-8
7
28-37
+
+
+
+
+
+
+
+
+
+
+
+
+
+
+
+
-
-
+
+
+
-
72.1

Characterization and Description of Novel Actinomycete Taxa
105
Fig. 3.3. Neighbour-joining tree, based on 16S rRNA gene sequences, showing the
relationships between strain MBRL 34T and all the reported type strains of Micromonospora
species. Catellatospora citrea NBRC 14495T is used as the outgroup. Asterisks indicate branches that
were also recovered using the maximum parsimony tree. Numbers at nodes are levels of bootstrap
support (%) for branch points (1000 resamplings). Bar, 0.005 substitutions per nucleotide position.
M. chokoriensis 2-19/6T (AB241454) M. saelicesensis Lupac 09T (AJ783993)
M. lupini lupac 14NT (AJ783996)
M. mirobrigensis WA201T (AJ626950)
M. siamensis TT2-4T (AB193565)
M. matsumotoense IMSNU 22003T (AF152109)
M. rifamycinica AM105T (AY561829)
M. krabiensis MA-2T (AB196716)
M. carbonacea DSM 43815T (X92606)
M. purpureochromogenes DSM 43821T (X92611) M. coxensis 2-30-b/28T (AB241455)
M. halophytica DSM 43026T (X92601)
M. humi P0402T (GU459068)
M. echinospora ATCC 15837T (U58532)
M. tulbaghiae TVU1T (EU196562)
M. chalcea DSM 43026T (X92594)
M. aurantiaca ATCC 27029T (CP002162)
M. maritima D10-9-5T (HQ704071)
M. marina JSM1-1T (AB196712)
M. sediminicola SH2-13T (AB609325)
M. eburnea LK2-10T (AB107231)
M. narathiwatensis BTG4-1T (AB193559)
M. nigra DSM 43818T (X92609)
M. sagamiensis DSM 43912T (X92624)
M. inyonensis DSM 46123T (X92629)
M. pallida DSM 43817T (X92608)
M. inositola DSM 43819T (X92610)
M. fulviviridis DSM 43906T (X92620)
M. rosaria DSM 803T (X92631)
M. chersina DSM 44151T (X92628)
M. endolithica DSM 44398T (AJ560635)
M. cremea CR30T (FN658654)
M. coriariae NAR01T (AJ784008)
M. echinofusca DSM 43913T (X92625)
M. peucetia DSM 43363T (X92603)
M. citrea DSM 43903T (X92617)
M. chaiyaphumensis MC5-1T (AB196710)
M. auratinigra TT1-11T (AB159779)
M. yangpuensis FXJ6.011T (GU002071)
M. echinaurantiaca DSM 43904T (X92618)
Micromonospora manipurensis MBRL 34T (JN560152)
Micromonospora coerulea DSM 43143T (X92598)
M. viridifaciens DSM 43909T (X92623)
M. equina Y22T (JF912511)
M. pattaloongensis TJ2-2T (AB275607)
M. pisi GUI 15T (AM944497)
M. olivasterospora DSM 43868T (X92613)
M. rhizosphaerae 211018T (FJ261956)
Catellatospora citrea NBRC 14495T (D85477)
97*
91
94*
77*
74
92*
62*
60
96*
50
76*
*
52*
*
*
52
*
*
*
0.005

Characterization and Description of Novel Actinomycete Taxa
106
Table 3.4. Cellular fatty acid compositions of MBRL 34T and M. coerulea DSM 43143T (%)
Fatty acid MBRL 34T DSM 43143T
Straight-chain
C17:0
C18:0
Branched
iso-C14:0
iso-C15:0
anteiso-C15:0
iso-C16:1 H#
iso-C16:0
iso-C17:0
anteiso-C17:0
2-OH C16:1
10-methyl C17:0
iso-C18:0
Summed features*
9
5.2
1.8
1.6
24.4
1.9
1.0
36.2
8.1
2.8
2.0
3.4
2.2
4.2
2.9
1.9
1.1
28.0
1.0
-
27.6
13.7
3.7
3.3
3.0
1.3
6.0
* Summed features represent two or three fatty acids that cannot be separated by the Microbial
Identification System. Summed feature 9 consisted of iso-C17:19c and/or 10-methyl C16:0.
# Indicates that the position and configuration of the double bond is not known.
The morphological, 16S rRNA gene sequence and chemotaxonomical characteristics
indicate the affiliation of the strain MBRL 34T to the genus Micromonospora. The
differences between the strain MBRL 34T and the closest phylogenetic neighbour M.
coerulea DSM 43143T obtained from the results of DDH experiment and differential
phenotypic characteristics indicate that the strain MBRL 34T could be placed as a
novel taxon among the genus Micromonospora, for which the name Micromonospora
kangleikpakensis sp. nov. has been proposed.

Characterization and Description of Novel Actinomycete Taxa
107
3.3.1.7. Etymology of the strain MBRL 34T
Micromonospora kangleipakensis (kang.lei.pak.en´sis. N.L. masc. adj.
kangleipakensis pertaining to Kangleipak, the old name of Manipur, a state in North
East India, the source of the soil from where the type strain was isolated).
The type strain, MBRL 34T has been deposited in the Deutsche Sammlung von
Mikroorganismen und Zellkulturen, Germany (DSM 45612T) and Japan Collection of
Microorganisms, Japan (JCM 17696T).
3.3.2. Streptomyces manipurensis sp. nov.
3.3.2.1. Morphological characteristics
Strain MBRL 201T formed extensive substrate and aerial mycelia bearing long spore
chains (~ 70 spores per chain). At maturity, the strain formed rectiflexible spore
chains. Growth morphology on SCNA and the scanning electron micrograph of the
aerial mycelia for strain MBRL 201T are shown in Figure 3.4. The strain grew well in
all the media tested (Table 3.5).
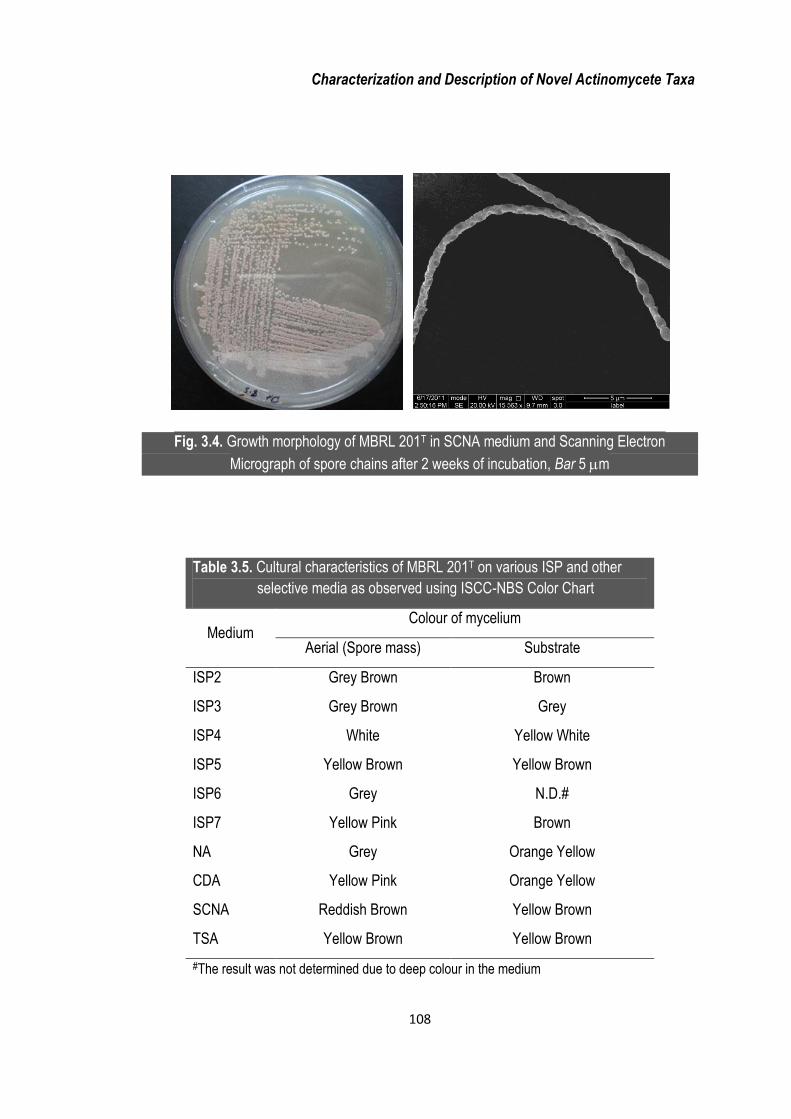
Characterization and Description of Novel Actinomycete Taxa
108
Fig. 3.4. Growth morphology of MBRL 201T in SCNA medium and Scanning Electron
Micrograph of spore chains after 2 weeks of incubation, Bar 5 m
Table 3.5. Cultural characteristics of MBRL 201T on various ISP and other
selective media as observed using ISCC-NBS Color Chart
Medium Colour of mycelium
Aerial (Spore mass) Substrate
ISP2
ISP3
ISP4
ISP5
ISP6
ISP7
NA
CDA
SCNA
TSA
Grey Brown
Grey Brown
White
Yellow Brown
Grey
Yellow Pink
Grey
Yellow Pink
Reddish Brown
Yellow Brown
Brown
Grey
Yellow White
Yellow Brown
N.D.#
Brown
Orange Yellow
Orange Yellow
Yellow Brown
Yellow Brown
#The result was not determined due to deep colour in the medium

Characterization and Description of Novel Actinomycete Taxa
109
3.3.2.2. Biochemical characteristics
The strain could hydrolyze tyrosine, starch (weakly) and Tweens 20, 40, 60 and 80
but not casein and gelatin. The strain was positive for catalase and indole production
tests but negative for methyl red, Voges-Proskauer, citrate utilization and nitrate
reduction tests. It could not produce acid from fructose, glucose, lactose, maltose,
mannitol or sucrose. The strain utilized arabinose, cellobiose, fructose, galactose,
maltose, mannose, raffinose, ribose, sodium malate and succinic acid as sole carbon
sources and L-alanine, L-arginine, L-arginine, L-asparagine, glycine, L-histidine, L-
hydroxyproline, L-leucine, potassium nitrate, proline, L-serine, L-tryptophan and L-
tyrosine as sole nitrogen sources. It could not utilize dulcitol, meso-inositol, lactose,
mannitol, rhamnose, sorbose, trehalose, xylitol, xylose, L-aspartic acid, glutamic acid,
L-methionine, L-ornithine, L-phenylalanine or L-valine as either sole carbon or
nitrogen sources. The biochemical characteristics of strain MBRL 201T are
summarized in Table 3.6.
3.3.2.3. Chemotaxonomical characteristics
Strain MBRL 201T had LL-DAP as the diagnostic cell wall diamino acid. Glucose
was the main sugar detected in the whole cell hydrolysates along with small amounts
of galactose, mannose, rhamnose, ribose and xylose. MK-9(H6) (58.7 %) and MK-
9(H8) (41.3 %) were the predominant menaquinones detected. The major polar lipids
detected were DPG, phosphatidylmethylethanolamine (PME), PG, PE, PI and PIM,
with other ULs and unknown phospholipids (PLs) (Figure 3.5). The FAME profile
(>1%) contained anteiso-C15:0 (38.9 %), iso-C15:0 (19.9 %), anteiso-C17:0 (14.7 %),

Characterization and Description of Novel Actinomycete Taxa
110
C16:0 (7.7 %), iso-C17:0 (6.6 %), iso-C16:0 (6.5 %), iso-C14:0 (1.1 %), and Summed
Feature 9 containing C17:1c and/or 10-methyl C16:0 (1.0 %).
Fig. 3.5. Two-dimensional thin-layer chromatogram of polar lipids of strain MBRL 201T
(1, PIM; 2, PI; 3, PG; 4, PE; 5, PME; 6, DPG; 7-12, ULs; 13-15, PLs)
3.3.2.4. Physiological characteristics
The strain grows between 15-37 °C and pH 6-10, with optimum growth at 28 °C and
pH 8. It can grow in presence of up to 2 % NaCl.

Characterization and Description of Novel Actinomycete Taxa
111
Table 3.6. Biochemical characteristics of strain MBRL 201T
Characteristics Result Characteristics Result
Acid production from carbohydrate
Fructose
Glucose
Lactose
Maltose
Mannitol
Sucrose
Catalase activity
Citrate utilization
Utilization of sole C sources
Arabinose
Cellobiose
Dulcitol
Fructose
Galactose
Inositol
Lactose
Maltose
Mannitol
Mannose
Raffinose
Rhamnose
Ribose
Salicin
Sodium malate
Sorbose
Succinic acid
Trehalose
Xylitol
Xylose
-
-
-
-
-
-
+
+
+
+
-
+
+
-
-
+
-
+
+
-
+
-
+
-
+
-
-
-
Hydrolysis of
Casein
Tween 20
Tween 40
Tween 60
Tween 80
Starch
Gelatin liquefaction
Indole production
Nitrate reduction
Methyl Red test
Voges Proskaeur test
Utilization of sole N sources
L-Alanine
L-Arginine hydrochloride
L-Asparagine
L-Aspartic acid
L-Glutamic acid
Glycine
L-Histidine hydrochloride
L-Leucine
L-Methionine
L-Ornithine
Potassium nitrate
L-Phenylalanine
L-Proline
L-Serine
L-Tryptophan
L-Tyrosine
L-Valine
-
+
+
+
+
w
-
-
-
-
-
+
+
+
-
-
+
+
+
-
-
+
-
+
+
+
+
-
Note: w weakly positive; + positive; - negative,

Characterization and Description of Novel Actinomycete Taxa
112
3.3.2.5. Genotypic characteristics
The GenBank accession number for the 16S rRNA gene sequence of strain MBRL
201T is JN560156. EzTaxon-e sequence similarity results showed that the strain
MBRL 201T
has 16S rRNA gene sequence homologies (>97 %) with 209
Streptomyces type strains. Analysis of EzTaxon-e and phylogenetic NJ tree results
(Figure 3.6) indicated that strain MBRL 201T is closely related to S. virginiae NBRC
12827T and S. cinnamonensis NBRC 15873
T (16S rRNA gene sequence similarities of
99.66% and 99.66%, respectively) and this phylogenetic relationship was also
supported in the tree generated with the MP algorithm.
The Genomic DNA G+C content was found to be 72.9 mol%. Strain MBRL 201T
exhibited low DNA-DNA reassociation values with S. virginiae NBRC 12827T
(44.5±3.1 %) and S. cinnamonensis NBRC 15873T (35.6±2.2 %), thereby indicating
that the whole genomic DNA relatedness values are well below the delineating 70%
cut-off point for species identification. The new species fall in the cluster 39 of the
family Streptomycetaceae as identified by Labeda et al. (2012).
3.3.2.6. Differentiating characteristics among strains MBRL 201T, NBRC 12827
T and
NBRC 15873T
The genotypic and phenotypic features described above suggest that strain MBRL
201T
could be clearly distinguished from its closest phylogenetic relatives. Besides
low DNA-DNA relatedness with the closest phylogenetic neighbours, the strain is
also distinguished from them by several phenotypic properties as listed in Table 3.7.
Therefore, the Hundung strain MBRL 201T is considered to represent a new species

Characterization and Description of Novel Actinomycete Taxa
113
of the genus Streptomyces, for which the name Streptomyces manipurensis sp. nov.
has been proposed.
Fig. 3.6. Neighbour-joining tree, based on 16S rRNA gene sequences, showing the relationship
between strain MBRL 201T and type strains of Streptomyces species. Asterisks indicate
branches that were also recovered using the maximum parsimony tree. Numbers at nodes are
levels of bootstrap support (%) for branch points (1000 resamplings). Bar, 0.001 substitutions
per nucleotide position.
S. xanthophaeus NBRC 12829T (AB184177)
S. spororaveus LMG 20313T (AJ781370)
S. nojiriensis LMG 20094T (AJ781355)
S. cirratus NRRL B-3250T (AY999794)
S. vinaceus NBRC 13425T (AB184394)
S. subrutilus DSM 40445T (X80825)
S. avidinii NBRC 13429T (AB184395)
S. lavendulae subsp. lavendulae NBRC 12789T (AB184146)
S. colombiensis NRRL B-1990T (DQ026646)
S. goshikiensis NBRC 12868T (AB184204)
S. sporoverrucosus NBRC 15458T (AB184684)
S. virginiae NBRC 12827T (AB184175)
S. cinnamonensis NBRC 15873T (AB184707)
Streptomyces manipurensis MBRL 201T (JN560156)
S. yokosukanensis NRRL B-3353T (DQ026652)
S. katrae NBRC 13447T (AB184409)
S. polychromogenes NBRC 13072T (AB184292)
S. racemochromogenes NRRL B-5430T (DQ026656)
S. flavotricini NBRC 12770T (AB184132)
S. globosus LMG 19896T (AJ781330)
S. toxytricini NBRC 12823T (AB184173) 99*
94
99
100
95*
88*
68*
91 56
57
53
0.001
*

Characterization and Description of Novel Actinomycete Taxa
114
Table 3.7. Differential characteristics among strains MBRL 201T, S. virginiae NBRC 12827T and
S. cinnamonensis NBRC 15873T
Characteristics S. manipurensis
MBRL 201T
S. virginiae
NBRC 12827T
S. cinnamonensis
NBRC 15873T
pH range
Optimum pH for growth
Hydrolysis of
Casein
Starch
Tyrosine
Nitrate reduction
Utilization of sole C sources
Arabinose
Cellobiose
Fructose
Galactose
Mannose
Raffinose
Sodium malate
Succinic acid
Trehalose
Utilization of sole N sources
L-Arginine
DL-Methionine
L-Ornithine
L-Phenylalanine
L-Valine
Major fatty acids (>5%)
iso-C15:0
anteiso-C15:0
iso-H C16:1
iso-C16:0
C16:0
Sum in Feature 9
anteiso-C17:19c
iso-C17:0
anteiso-C17:0
G+C mol %
6-10
8
-
w
+
-
+
+
+
+
+
+
+
+
-
+
-
-
-
-
19.9
38.9
-
6.5
7.7
-
-
6.6
14.7
72.9
6-8
7
+
-
-
+
-
+
-
+
+
-
-
-
-
-
-
+
+
+
13.0
27.7
-
24.5
6.2
-
-
-
10.1
72.0
6-9
7
+
+
-
+
-
-
+
-
-
+
-
-
+
+
+
-
+
+
15.9
22.1
5.1
17.1
5.6
6.3
5.4
-
7.8
72.4
Note: +, Positive; - negative; w weakly positive; all the data were from this study.

Characterization and Description of Novel Actinomycete Taxa
115
3.3.2.7. Etymology of the strain MBRL 201T
Streptomyces manipurensis (ma.ni.pur.en´sis. N.L. masc. adj. manipurensis
pertaining to Manipur, a state in North East India, the source of the soil from which
the type strain was isolated).
The type strain, MBRL 201T, has been deposited in the Deutsche Sammlung von
Mikroorganismen und Zellkulturen, Germany (DSM 42029T) and Japan Collection of
Microorganisms, Japan (JCM 17351T).
3.3.3. Streptomyces hundungensis sp. nov.
3.3.3.1. Morphological characteristics
Strain MBRL 251T formed extensive substrate and aerial mycelia bearing long spore
chains (approximately 50 spores per chain). At maturity, the strain formed
rectiflexibile spore chains. Growth morphology of MBRL 251T in the SCNA medium
and scanning electron micrograph showing spore chains in the aerial mycelia are
shown in Figure 3.7. The strain grew well in all the media tested (Table 3.8).

Characterization and Description of Novel Actinomycete Taxa
116
Fig. 3.7. Growth morphology of MBRL 251T in SCNA medium and Scanning Electron
Micrograph of spore chains after 2 weeks of incubation, Bar 5 m.
Table 3.8. Cultural characteristics of MBRL 251T on various ISP and other selective
media as observed using ISCC-NBS Color Chart
Medium Colour of mycelium
Aerial (Spore mass) Substrate
ISP2
ISP3
ISP4
ISP5
ISP6
ISP7
NA
CDA
SCNA
TSA
Grey Brown
Grey Brown
White
Yellow Brown
Grey
Yellow Pink
Grey
Yellow Pink
Reddish Brown
Yellow Brown
Brown
Grey
Yellow White
Grey Yellow Brown
N.D.*
Brown
Orange Yellow
Grey Yellow Brown
Orange Yellow
Yellow Brown
*The result was not determined due to deep colour in the medium.

Characterization and Description of Novel Actinomycete Taxa
117
3.3.3.2. Biochemical characteristics
Strain MBRL 251T could produce acid from fructose and glucose, but not from
lactose, maltose, mannitol or sucrose. The strain was found to hydrolyze tyrosine,
gelatin and Tweens 20, 40, 60 and 80 but not casein and starch. The strain was
positive for catalase, methyl red and indole production and nitrate reduction tests but
negative for Voges-Proskauer and citrate utilization tests. It could utilize arabinose,
cellobiose, fructose, galactose, maltose, mannose, sodium malate, succinic acid and
xylose as sole carbon sources; and L-alanine, L-arginine, glycine, L-histidine, L-
hydroxyproline, DL-methionine, L-ornithine, L-phenylalanine, proline, L-serine and
L-valine as sole nitrogen sources. It could not utilize dulcitol, meso-inositol, lactose,
mannitol, raffinose, rhamnose, sorbose, trehalose, or potassium nitrate as either sole
carbon or nitrogen sources. The biochemical characteristics of strain MBRL 251T are
summarized in Table 3.9.
3.3.3.3. Chemotaxonomical characteristics
Strain MBRL 251T had LL-DAP as the diagnostic cell wall diamino acid. Glucose
(32.9 %) and xylose (32.1 %) were the main sugars detected in the whole cell
hydrolysates along with small amounts of galactose, mannose, rhamnose and ribose.
MK-9(H6) (72.4 %) and MK-9(H8) (27.6 %) were the predominant menaquinones
detected. The major polar lipids detected were DPG, PE, PG, PI and PIM, with other
unknown ULs, PL, aminophospholipid (APL) (Figure 3.8). The FAME profile (>1%)
contained anteiso-C15:0 (35.1 %), iso-C16:0 (21.1 %), anteiso-C17:0 (13.2 %), iso-C14:0
(6.5 %), C16:0 (6.1 %), iso-C15:0 (3.8 %), iso-C17:0 (2.9 %), cyclo-C17:0 (2.8 %), and iso-
C16:1 H (1.7 %).

Characterization and Description of Novel Actinomycete Taxa
118
Fig. 3.8. Two-dimensional thin-layer chromatogram of polar lipids of strain MBRL 251T
1, PIM; 2, PI; 3, PG; 4, PE; 5, DPG; 6-7, ULs; 8, PL; 9, APL.
3.3.3.4. Physiological characteristics
The strain MBRL 251T grows at 15-37 °C and between pH 5-10, with optimum
growth at 28 °C and pH 8. It can grow in presence of up to 7 % NaCl.
3.3.3.5. Genotypic characteristics
The GenBank accession number for the 16S rRNA gene sequence of strain MBRL
251T is JN560156. EzTaxon-e analysis showed that the strain MBRL 251
T shared
close 16S rRNA gene sequence homologies (>97%) with 205 Streptomyces type
strains. Analysis of EzTaxon-e and NJ tree results (Figure 3.9) indicated that strain
MBRL 251T is closely related to S. xanthochromogenes NRRL B-5410
T (99.66 %)
and S. michiganensis NBRC 12797T (99.66 %). The strain MBRL 251
T falls in the
cluster 29 of the family Streptomycetaceae as suggested by Labeda et al. (2012).
1 2
3 4
5
8
6 7
9

Characterization and Description of Novel Actinomycete Taxa
119
Table 3.9. Biochemical characteristics of strain MBRL 251T
Characteristics Result Characteristics Result
Acid production from carbohydrate
Fructose
Glucose
Lactose
Maltose
Mannitol
Sucrose
Catalase activity
Utilization of sole C sources
Arabinose
Cellobiose
Dulcitol
Fructose
Galactose
Inositol
Lactose
Maltose
Mannitol
Mannose
Raffinose
Rhamnose
Sodium malate
Sorbose
Succinic acid
Trehalose
Xylose
+
+
-
-
-
-
+
+
+
-
+
+
-
-
+
-
+
-
-
+
-
+
-
+
Hydrolysis of
Casein
Starch
Tween 20
Tween 40
Tween 60
Tween 80
Tyrosine
Citrate utilization
Gelatin liquefaction
Indole production
Nitrate reduction
Methyl Red test
Voges Proskaeur test
Utilization of sole N sources
L-Alanine
L-Arginine hydrochloride
Glycine
L-Histidine hydrochloride
DL-Methionine
L-Ornithine
Potassium nitrate
L-Phenylalanine
L-Proline
L-Serine
L-Valine
-
-
+
+
+
+
+
-
+
+
+
+
-
+
+
+
+
+
+
-
+
+
+
+
Note: + positive; - negative

Characterization and Description of Novel Actinomycete Taxa
120
Fig. 3.9. Neighbour-joining tree, based on 16S rRNA gene sequences, showing the
relationships between strain MBRL 251T and other type strains of Streptomyces species.
Asterisks indicate branches that were also recovered using the maximum parsimony tree.
Numbers at nodes are levels of bootstrap support (%) for branch points (1000 resamplings).
Bar, 0.002 substitutions per nucleotide position.
The G+C content of the genomic DNA was determined to be 72.3 mol%. DDH
studies for strain MBRL 251T was done with the two closest relatives, S.
xanthochromogenes NBRC 12828T and S. michiganensis NBRC 12797
T. The results
indicated that strain MBRL 251T had low DNA-DNA reassociation values with S.
xanthochromogenes NBRC 12828T (46.6±8.9 %) and S. michiganensis NBRC 12797
T
(40.7±3.7 %), thereby indicating that the whole genomic DNA relatedness values are
well below the delineating 70% cut-off point for species identification.
S. badius NRRL B-2567T (AY999783)
S. globisporus subsp. globisporus NBRC 12867T (AB184203)
S. sindenensis NBRC 3399T (AB184759)
S. parvus NBRC 3388T (AB184756)
S. fimicarius ISP 5322T (AY999784)
S. anulatus NRRL B-2000T (DQ026637)
S. griseorubiginosus NBRC 13047T (AB184276)
S. microflavus NBRC 13062T (AB184284)
S. mauvecolor LMG 20100T (AJ781358)
S. violascens NBRC 12920T (AB184246)
Streptomyces hundungensis MBRL 251T (JN560157)
S. xanthochromogenes NRRL B-5410T (DQ442559)
S. michiganensis NBRC 12797T (AB184153)
S. melanogenes NBRC 12890T (AB184222)
S. noboritoensis NBRC 13065T (AB184287)
S. gobitricini NBRC 15419T (AB184666)
100*
98* 82*
93*
77
60
88*
*
59*
99
68
0.002

Characterization and Description of Novel Actinomycete Taxa
121
3.3.3.6. Differentiating characteristics among strains MBRL 251T, NBRC 12828
T and
NBRC 12797T
The genotypic and phenotypic features described above suggest that strain MBRL
251T
could be clearly distinguished from its closest phylogenetic relatives. Besides
low DNA-DNA relatedness with the closest phylogenetic neighbours, the strain is
also distinguished from them by several phenotypic properties as listed in Table 3.10.
Therefore, the Hundung strain MBRL 251T is considered to represent a new species
of the genus Streptomyces, for which the name Streptomyces hundungensis sp. nov.
has been proposed.
3.3.3.7. Etymology of the strain MBRL 251T
Streptomyces hundungensis (hun.dung.en´sis. N.L. masc. adj. hundungensis
belonging to Hundung in Ukhrul, a hill district in Manipur, India, the source of the
type strain).
The type strain, MBRL 251T has been deposited in Japan Collection of
Microorganism, Japan (JCM 17577T) and Korean Collection for Type Cultures, Korea
(KCTC 29125T).

Characterization and Description of Novel Actinomycete Taxa
122
Table 3.10. Differential characteristics among strains MBRL 251T, S. xanthochromogenes NBRC
12828T and S. michiganensis NBRC 12797T
Characteristics S. hundungensis
MBRL 251T
S. xanthochromogenes
NBRC 12828T
S. michiganensis
NBRC 12797T
pH range
Optimum pH for growth
NaCl tolerance (%, w/v)
Hydrolysis of
Casein
Gelatin
Tyrosine
Catalase
MR
Indole production
Utilization of sole C-sources
Arabinose
Cellobiose
Maltose
Mannitol
Raffinose
Sodium malate
Sorbose
Trehalose
Utilization of sole N-sources
L-Arginine
Potassium nitrate
Major fatty acids (>5%)
iso-C14:0
iso-C15:0
anteiso-C15:0
iso-C16:0
C16:0
anteiso-C17:19c
anteiso-C17:1
G+C mol %
5-10
8
7
-
+
+
w
+
+
+
+
+
-
-
+
-
-
+
-
6.5
-
35.1
21.1
6.1
-
13.2
72.3
6-9
7
2
-
-
-
+
-
-
-
-
-
-
-
-
-
-
-
+
-
5.7
44.2
12.9
-
5.1
12.9
72.3
6-9
7
2
+
+
-
+
+
+
+
+
+
+
+
-
+
+
+
+
-
5.2
40.6
13.2
8.1
-
19.2
71.9
Note: + Positive; w weakly positive; - negative

Characterization and Description of Novel Actinomycete Taxa
123
3.4. DISCUSSION
Limited studies have been done on the microbiology of limestone habitats. Most of
these studies have been focussed on the aspects of microbial biogeography (Groth et
al. 1999; Stomeo et al. 2008; Yamac et al. 2011; Niyomvong et al. 2012; Rule and
Cheeptham 2013). The rich diversity of actinomycetes in limestone habitats has been
demonstrated earlier by the discovery of two novel genera Knoellia and Hoyosella
(Groth et al. 2002; Jurado et al. 2009) from such less explored habitats.
Caves, another limestone ecosystem, with large calcium carbonate deposits, are
considered to be hostile environments for life (Howarth 1993) and are often severely
resource-limited due to absence of light that precludes primary production of organic
materials (Poulson and Lavoie 2000). Microbes, especially the mineral oxidizing
bacteria, play a major role in the dissolution of limestones and other calcareous rocks
for the formation of caves. Microbially influenced dissolution or corrosion of mineral
surfaces can occur through secretion of exoenzymes, organic acids e.g. carbonic acid
(Gillieson 1996; Schwabe et al. 2001) and mineral acids e.g. sulphuric acid (Hill
1987, 1990, 1995, 1996, 2000; Jagnow et al. 2000), and a variety of other mechanisms
(Sand 1997; Jones 2001; Bullen et al. 2008). In addition, bacteria isolated from caves
are shown to be effective in extracellular precipitation of calcium carbonate through a
variety of processes that include ammonification, denitrification, sulphate reduction,
and anaerobic sulphide oxidation (Danielli and Edington 1983; Simkiss and Wilbur
1989; Ehrlich 1996; Castanier et al. 1999; Riding 2000; Engel et al. 2001).
Microorganisms are commonly found fossilized within carbonate speleotherms (Jones
and Motyka 1987; Polyak and Cokendolpher 1992; Jones and Kahle 1995; Melim et

Characterization and Description of Novel Actinomycete Taxa
124
al. 2001). Carbonate pool fingers in Lechuguilla Cave in New Mexico, USA were first
identified by Davis (2000) as potentially biogenic in orgin. Similar results were
found by Baskar et al. (2006) from microcrystalline calcites within stalactites.
Considering the ununsual nature of the limestone habitats and the interaction of
microorganisms in cave formation, it is not surprising to identify novel species from
such habitats. Barton and Jurado (2007) studied the diversity of microbes in
Lechuguilla Cave. They found that 40 out of 83 isolates were possible representatives
of previously uncultivated species. Some of the novel species of actinobacteria
reported from limestone rocks and related habitats are Amycolatopsits halotolerans
sp. nov. (Lee 2006a), Amycolatopsis jejuensis sp. nov. (Lee 2006a), Amycolatopsis
saalfeldensis sp. nov. (Carlsohn et al. 2007a), Catellatospora koreensis sp. nov. (Lee
et al. 2000), Hoyosella altamirensis gen. nov., sp. nov. (Jurado et al. 2009), Knoellia
sinensis gen. nov., sp.nov. (Groth et al. 2002), Knoellia subterranea sp.nov. (Groth et
al. 2002), Kribbella aluminosa sp. nov. (Carlsohn et al. 2007b), Microlunatus
cavernae sp. nov. (Cheng et al. 2013a), Nocardia jejuensis sp. nov. (Lee 2006b) and
Saccharopolyspora cavernae sp. nov. (Cheng et al. 2013b).
This chapter reports the isolation of three novel species from limestone samples. The
isolation of a moderately acidophilic actinomycete Micromonospora kangleipakensis
sp. nov. (Nimaichand et al. 2013a) from the alkaline environment is an interesting
finding. Presence of acidophilic bacteria have also been reported from limestone
caves of the Frasassi Gorge, Italy (Vlasceanu et al. 2000; Macalady et al. 2006). The
formation of weak organic acids from biogenic carbon dioxide or strong mineral acids
during the oxidation of minerals might result in creation of acidic microhabitats

Characterization and Description of Novel Actinomycete Taxa
125
within the alkaline limestone environments. Ruamps et al. (2011) studied microbial
biogeography in different layers of soil and found different microhabitats providing
diverse conditions for microbial growth, activity and survival at the various layers.
This concept of micro-environments within a large environment could be the reason
behind isolation of the strain Micromonospora kangleipakensis strain MBRL 34T. The
isolation of Streptomyces manipurensis sp. nov. (Nimaichand et al. 2012) and
Streptomyces hundungensis sp. nov. (Nimaichand et al. 2013b), and 7 other
potentially novel strains of actinomycetes (among the representative phylotypic
strains) indicate that the Hundung ecosystem (underexplored limestone habitat) could
be a rich source for novel actinomycetes.
REFERENCES
Barton HA, Jurado V (2007) What’s up down there? Microbial diversity in caves. Microbe 2:
132-138
Baskar S, Baskar R, Mauclaire L, McKenzie JA (2006) Microbially induced calcite
precipitation in culture experiments: Possible origin for stalactites in Sahastradhara
Caves, Dehradun, India. Cur Sci 90: 58-64
Bullen HA, Oehrle SA, Bennett AF, Taylor NM, Barton HA (2008) The use of attenuated total
reflectance fourier transformed infrared (ATR-FTIR) spectroscopy to identify
microbial metabolic products on carbonate mineral surfaces. Appl Environ Microbiol
74: 4553-4559
Cappucino JE, Sherman N (2004) In Microbiology: A laboratory manual. New York: Benjamin
Cummings Publication Co

Characterization and Description of Novel Actinomycete Taxa
126
Carlsohn MR, Groth I, Tan GYA, Schűtze B, Saluz HP, Munder T, Yang J, Wink J,
Goodfellow M (2007a) Amycolatopsis saalfeldensis sp. nov., a novel actinomycete
isolated from a medieval alum slate mine. Int J Syst Evol Microbiol 57: 1640-1646
Carlsohn MR, Groth I, Sprőer C, Schűtze B, Saluz HP, Munder T, Stackebrandt E (2007b)
Kribbella aluminosa sp. nov., isolated from a medieval alum slate mine. Int J Syst
Evol Microbiol 57: 1943-1947
Castanier S, Le Métayer-Levrel G, Perthuisot JP (1999) Ca-carbonates precipitation and
limestone genesis – the microbiologist point of view. Sediment Geol 126: 9-23
Cheng J, Chen W, Huo-Zhang B, Nimaichand S, Zhou EM, Lu XH, Klenk HP, Li WJ (2013a)
Microlunatus cavernae sp. nov., a novel actinobacterium isolated from Alu ancient
cave, Yunnan, South-West China. Antonie van Leeuwenhoek 104: 95-101
Cheng J, Zhang YG, Chen W, Li L, Zhang DF, Wang HF, Lu XH, Duan YQ, Li WJ (2013b)
Saccharopolyspora cavernae sp. nov., a novel actinomycete isolated from the
Swallow Cave in Yunnan, south-west China. Antonie van Leeuwenhoek 104: 837-
843
Collins CH, Lyne PM, Grange JM, Falkinham JO (2004) In Microbiological Methods, Eighth
Edition. London: Arnold
Collins MD, Pirouz T, Goodfellow M, Minnikin DE (1977) Distribution of menaquinones in
actinomycetes and corynebacteria. J Gen Microbiol 100: 221-230
Colwell RR (1970) Polyphasic taxonomy of the genus Vibrio: numerical taxonomy of Vibrio
cholera, Vibrio parahaemolyticus, and related Vibrio species. J Bacteriol 104: 410-
433.
Danielli HMC, Edingthon MA (1983) Bacterial calcification in limestone caves. Geomicrobiol J
3: 1-16

Characterization and Description of Novel Actinomycete Taxa
127
Davis DG (2000) Extraordinary features of Lechuguilla Cave, Guadalupe Mountains, New
Mexico. J Cave Karst Stu 62: 147-157
DeLey J, Caltoir H, Reeynaerts A (1970) The quantitative measurement of DNA hybridization
from renaturation rate. Eur J Biochem 12: 133-142
Ehrlich HL (1996) In Geomicrobiology. 3rd
ed. New York: Marcel Dekker Inc
Engel AS, Porter ML, Kinkle BK, Kane TC (2001) Ecological assessment and geological
significance of microbial communities from Cesspool Cave, Virginia. Geomicrobiol J
18: 259-274
Felsenstein J (1981) Evolutionary trees from DNA sequences: a maximum likelihood
approach. J Mol Evol 17: 368-376
Felsenstein J (1985) Confidence limits on phylogenies: an approach using the bootstrap.
Evolution 39: 783-791
Gillieson D (1996) In Caves: Processes, Development, and Managements. Oxford: Blackwell
Publishers Ltd.
Goodfellow M (1986) Genus Rhodococcus Zopf 1891, 28AL
. In Sneath PHA, Mair NS, Sharpe
ME, J. G. Holt JG (Eds.) Bergey’s Manual of Systematic Bacteriology, vol. 2.
Baltimore: Williams & Wilkins pp. 1472-1481
Gordon RE, Barnett DA, Handerhan JE, Pang CHN (1974) Nocardia coeliaca, Nocardia
autotrophica, and the nocardin strain. Int J Syst Bacteriol 24: 54-63
Groth I, Vettermann R, Schuetze B, Schumann P, Saiz-Jimenez C (1999) Actinomycetes in
Karstic caves of northern Spain (Altamira and Tito Bustillo). J Microbiol Methods 36:
115-122

Characterization and Description of Novel Actinomycete Taxa
128
Hill CA (1987) Geology of Carlsbad Cavern and other caves in the Guadalupe Mountains,
New Mexico and Texas. New Mexico Bur Mines Mineral Res Bull 117. New Mexico
Bureau of Mines & Mineral Resources. Socorro, NM
Hill CA (1990) Sulfuric acid speleogenesis of Carlsbad Cavern and its relationship to
hydrocarbons, Delaware Basin, New Mexico and Texas. Am Assoc Petrol Geol Bull
74: 1685-1694
Hill CA (1995) Sulfur redox reactions: hydrocarbons, native sulfur, Mississippi Valley-type
deposits, and sulfuric acid karst in the Delaware Basin, New Mexico and Texas.
Environ Geol 25: 16-23
Hill CA (1996) Geology of the Delaware Basin, Guadalupe, Apache and Glass Mountains,
New Mexico and West Texas. Society of Economic Paleontologists & Mineralogists.
Permian Basin Section. Pub No 96-39. Permian Basin Section-SEPM. Midland, TX
Hill CA (2000) Overview of the geologic history of cave development in the Guadalupe
Mountains, New Mexico. J Cave Karst Stu 62: 60-71
Howarth FG (1993) High-stress subterranean habitats and evolutionary change in cave-
inhabiting arthropods. Am Nat 142: S65-S77
Jagnow DH, Hill CA, Davis DG, Duchene HR, Cunningham KI, Northup DE, Queen JM (2000)
History of the sulfuric acid theory of speleogenesis in the Guadalupe Mountains. J
Cave Karst Stu New Mexico 62: 54-59
Jones B (2001) Microbial activity in caves: A geological perspective. Geomicrobiol J 18: 345-
357
Jones B, Kahle CF (1995) Origin of endogenetic micrite in karst terrains: A case study from
the Cayman Islands. J Sediment Res A65: 283-293
Jones B, Motyka A (1987) Biogenic structures and micrite in stalactites from Grand Cayman
Island, British West Indies. Can J Earth Sci 24: 1402-1411

Characterization and Description of Novel Actinomycete Taxa
129
Jurado V, Kroppenstedt RM, Saiz-Jimenez C, Klenk HP, Mouniee D, Laiz L, Couble A, Potter
G, Boiron P, Rodriguez-Nava V (2009) Hoyosella altamirensis gen. nov., sp. nov., a
new member of the order Actinomycetales isolated from a cave biofilm. Int J Syst Evol
Microbiol 59: 3105-3110
Kelly KL (1964) Inter-Society Color Council – National Bureau of Standards Color-Name
Charts Illustrated with Centroid Colors. US Government Printing Office, Washington
Kim OS, Cho YJ, Lee K, Yoon SH, Kim M, Na H, Park SC, Jeon YS, Lee J, Yi H, Won S,
Chun J (2012) Introducing EzTaxon-e: a prokaryotic 16S rRNA Gene sequence
database with phylotypes that represent uncultured species. Int J Syst Evol Microbiol
62: 716-721
Kimura M (1983) In The Neutral Theory of Molecular Evolution. Cambridge: Cambridge
University Press.
Kluge AG, Farris FS (1969) Quantitative phyletics and the evolution of anurans, Syst Zool 18:
1-32
Kovacs N (1956) Identification of Pseudomonas pyocyanea by the oxidase reaction. Nature
178: 703-70
Lanyi B (1987) Classical and rapid identification methods for medically important bacteria.
Methods Microbiol 19: 1-67
Labeda DP, Goodfellow M, Brown R, Ward AC, Lanoot B, Vanncanneyt M, Swings J, Kim SB,
Liu Z, Chun J, Tamura T, Oguchi A, Kikuchi T, Kikuchi H, Nishii T, Tsuji K,
Yamaguchi Y, Tase A, Takahashi M, Sakane T, Suzuki KI, Hatano K (2012).
Phylogenetic study of the species within the family Streptomycetaceae. Antonie van
Leeuwenhoek 101, 73-104.

Characterization and Description of Novel Actinomycete Taxa
130
Larkin MA, Blackshields G, Brown NP, Chenna R, McGettigan PA, McWilliam H, Valentin F,
Wallace IM, Wilm A, Lopez R, Thompson JD, Gibson TJ, Higgins DG (2007) Clustal
W and Clustal X version 2.0. Bioinformatics 23: 2947-2948
Leboff MJ, Pierce BE (2008) In Microbiology: Laboratory Theory and Application (Ringbound,
Brief Edition), New York: Morton Publishing Company
Lechevalier MP, De Bièvre C, Lechevalier HA (1977) Chemotaxonomy of aerobic
actinomycetes: phospholipid composition. Biochem Syst Ecol 5: 249-260
Lee SD (2006a) Amycolatopsis jejuensis sp. nov. and Amycolatopsis halotolerans sp. nov.,
novel actinomycetes isolated from a natural cave. Int J Syst Evol Microbiol 56: 549-
553
Lee SD (2006b) Nocardia jejuensis sp. nov., a novel actinomycetes isolated from a natural
cave on Jeju Island, Republic of Korea. Int J Syst Evol Microbiol 56: 559-562
Lee SD, Kang SO Hah YC (2000) Catellatospora koreensis sp. nov., a novel actinomycete
isolated from goldmine cave. Int J Syst Evol Microbiol 50: 1103-1111
Li WJ, Xu P, Schumann P, Zhang YQ, Pukall R, Xu LH, Stackebrandt E, Jiang CL (2007)
Georgenia ruanii sp. nov., a novel actinobacterium isolated from forest soil in
Yunnan (China), and emended description of the genus Georgenia. Int J Syst Evol
Microbiol 57: 1424-1428
Macalady JL. Lyon EH, Koffman B, Albertson LK, Meyer K, Galdenzi S, Mariani S (2006)
Dominant microbial populations in limestone-corroding stream biofilms, Frasassin
Cave system, Italy. Appl Environ Microbiol 72: 5596-5609
Melim LA, Shinglman KM, Boston PJ, Northup DE, Spilde MN, Queen JM (2001) Evidence for
microbial involvement in pool finger precipitation, Hidden Cave, New Mexico.
Geomicrobiol J 18: 311-329

Characterization and Description of Novel Actinomycete Taxa
131
Mesbah M, Premachandran U, Whitman WB (1989) Precise measurement of the G+C
content of deoxyribonucleic acid by high-performance liquid chromatography. Int J
Syst Bacteriol 39: 159-167
Minnikin DE, O’Donnell AG, Goodfellow M, Alderson G, Athalye M, Schaal A, Parlett JH
(1984) An integrated procedure for the extraction of bacterial isoprenoid quinones
and polar lipids. J Microbiol Methods 2: 233-241
Murray RGE, Brenner DJ, Colwell RR, De Vos P, Goodfellow M, Grimont PAD, Pfennig N,
Stackebrandt E, Zavarzin GA (1990) Report of the ad hoc committee on approaches
to taxonomy within the Proteobacteria. Int J Sys Bacteriol 40: 213-215
Nimaichand S, Zhu WY, Yang LL, Ming H, Nie GX, Tang SK, Ningthoujam DS, Li WJ (2012)
Streptomyces manipurensis sp. nov., a novel actinomycete isolated from a limestone
deposit site in Manipur, India. Antonie van Leeuwenhoek 102: 133-139
Nimaichand S, Zhang YG, Cheng J, Li L, Zhang DF, Zhou EM, Dong L, Ningthoujam DS, Li
WJ (2013a) Micromonospora kangleipakensis sp. nov., isolated from a sample of
Limestone quarry. Int J Syst Evol Microbiol 63: 4546-4551
Nimaichand S, Tamreihao K, Yang LL, Zhu WY, Zhang YG, Li L, Tang SK, Ningthoujam DS,
Li WJ (2013b) Streptomyces hundungensis sp. nov., a novel actinomycete with
antifungal activity and plant growth promoting traits. J Antibiot 66: 205-209
Niyomvong N, Pathom-aree W, Thamchaipenet A, Duangmal K (2012) Actinomycetes from
Tropical Limestone Caves. Chiang Mai J Sci 39: 373-388
Polyak VJ, Cokendolpher JC (1992) Recovery of microfossils from carbonate speleotherms.
Bull Natl Speleol Soc 54: 66-68
Poulson TL, Lavoie KH (2000) The trophic basis of subsurface ecosystems. In Wilkens H,
Culver DC, Humphreys JW (Eds) Ecosystems of the World 30: Subterranean
Ecosystems. Amsterdam: Elsevier pp. 231-249

Characterization and Description of Novel Actinomycete Taxa
132
Riding R (2000) Microbial carbonates; the geological record of calcified bacterial-algal mats
and biofilms. Sedimentology 47: 179-214
Ruamps LS, Nunan N, Chenu C (2011) Microbial biogeography at the soil pore scale. Soil
Biol Biochem 43: 280-286
Rule D, Cheeptham N (2013) The effects of UV light on the antimicrobial activities of cave
actinomycetes. Int J Speleology 42: 147-153
Saitou N, Nei M (1987) The neighbour-joining method: a new method for reconstructing
phylogenetic trees. Mol Bio Evol 4: 406-425
Sand W (1997) Microbial mechanisms of deterioration of inorganic substrates – A general
mechanistic review. Int Biodeter Biodegrad 40: 183-190
Sasser M (1990) Identification of bacteria by gas chromatography of cellular fatty acids.
USFCC Newsl 20: 16
Schwabe SJ, Herbert RA, Carew JL (2001) A hypothesis for biogenic cave formation: A study
conducted in the Bahamas. In Proceedings of the 13th Symposium on the Geology of
the Bahamas and other Carbonate Regions, pp 141-152
Shirling EB, Gottlieb D (1966) Methods for characterization of Streptomyces species. Int J
Syst Bacteriol 16: 313-340
Sierra GA (1957) A simple method for the detection of lipolytic activity of microorganisms and
some observations on the influence of the contact between cells and fatty
substrates. Antonie van Leeuwenhoek 23: 15-22
Simkiss K, Wilbur KM (1989) In Biomineralization: Cell Biology and Mineral Deposition. San
Diego: Academic Press
Stackebrandt E, Ebers J (2006) Taxonomic parameters revisited: tarnished gold standards.
Microbiol Today 33: 152-155

Characterization and Description of Novel Actinomycete Taxa
133
Staneck JL, Robert GD (1974) Simplified approached to identification of aerobic
actinomycetes by thin-layer chromatography. Appl Microbiol 28: 226-231
Stomeo F, Portillo MC, Gonzalez JM, Laiz L, Saiz-Jimenez C (2008) Pseudonocardia in white
colonizations in two caves with Paleolithic paintings. Int Biodeter Biodegrad 62: 483-
486
Tamaoka J, Katayama-Fujimura Y, Kuraishi H (1983) Analysis of bacterial menaquinone
mixtures by high performance liquid chromatography. J Appl Bacteriol 54: 31-36
Tamura K, Peterson D, Peterson N, Stecher G, Nei M, Kumar S (2011) MEGA5: Molecular
Evolutionary Genetics Analysis Using Maximum Likelihood, Evolutionary Distance,
and Maximum Parsimony Methods. Mol Bio Evol 28: 2731-2739
Tang SK, Wang Y, Chen Y, Lou K, Cao LL, Xu LH, Li WJ (2009) Zhihengliuella alba sp. nov.,
and emended description of the genus Zhihengliuella. Int J Syst Evol Microbiol 59:
2025-2031
Vlasceanu L, Sarbu SM, Engel AS, Kinkle BK (2000) Acidic cave-wall biofilms located in the
Frasassi Gorge, Italy. Geomicrobiol J 17: 125-139
Wayne LG, Brenner DJ, Colwell RR, Grimont PAD, Kandler O, Krichevsky MI, Moore LH,
Moore WEC, Murray RGE et al. (1987) International Committee on Systematic
Bacteriology. Report of the ad hoc committee on reconciliation of approaches to
bacterial systematic. Int J Syst Bacteriol 37: 463-464
Williams ST, Davies FL (1967) Use of a scanning electron microscope for the examination of
actinomycetes. J Gen Microbiol 48: 171-177
Yamac M, Isik K, Sahin N (2011) Numerical classification of streptomycetes isolated from
karstic caves in Turkey. Turk J Biol 35: 473-484