cloning, expression, and characterization of tnfsf14 (light) gene in mefugu, takifugu obscurus
TRANSCRIPT

Cloning, expression, and characterization of TNFSF14 (LIGHT)gene in mefugu, Takifugu obscurus
Chunlan Li • Yuefen Shen • DingFang Liang •
Fei Yin • Hongxin Ai • Boqing Sun •
Shikang Lin • Shuangquan Zhang
Received: 25 December 2012 / Accepted: 21 March 2013 / Published online: 29 March 2013
� Springer Science+Business Media New York 2013
Abstract LIGHT/TNFSF14 is a member of the tumor
necrosis factor ligand superfamily, which plays important
roles in inflammatory and immune responses. In this study,
the cDNA of mefugu (Takifugu obscures) LIGHT (desig-
nated as fLIGHT) was amplified from spleen by reverse
transcription polymerase chain reaction. The open reading
frame of fLIGHT covers 765 bp and translates into a
254-aa protein. The predicted three-dimensional (3D)
structure of the soluble LIGHT of mefugu (designated as
fsLIGHT) monomer analyzed by comparative protein
modeling revealed that it was very similar to its human
counterpart. Real-time quantitative PCR analysis indicated
that LIGHT is constitutively expressed in various tissues in
mefugu. The soluble LIGHT binding of green fluorescent
protein (GFP) (designated as GFP/fsLIGHT) had been
cloned into the pET28a vector; SDS-PAGE and western
blotting analysis confirmed that the recombinant protein
pET28a-GFP/fsLIGHT was efficiently expressed in Esch-
erichia coli BL21 (DE3). After purification, the GFP/
fsLIGHT fusion protein obtained similar fluorescence
spectrum to those of GFP. Laser scanning confocal
microscopy analysis showed GFP/fsLIGHT could bind to
its receptors. In view of our basic research, LIGHT may be
a potential immunologic factor for enhancing immuno-
logical efficacy in fish, and our results might provide a
platform for further study into the effects of LIGHT.
Keywords GFP/fsLIGHT � Real-time quantitative PCR �3D structure � Fluorescence spectrum � Confocal laser
Introduction
Members of the tumor necrosis factor (TNF) family and their
receptors are important regulators of the immune system [1].
This family includes cytokines that regulate various cellular
responses, including proliferation, differentiation, inflamma-
tion, and cell death [2]. So the TNF superfamily consists of
many membrane-bound and soluble proteins with proin-
flammatory effects on innate and adaptive immune responses
[3]. LIGHT (lymphotoxin-related inducible ligand that com-
petes with herpes simplex virus glycoprotein D for herpesvi-
rus entry mediator on T cells) is a recently identified member
of TNF superfamily [4]. It is mainly expressed on activated T
cells, natural killer cells, immature dendritic cells (DCs), and
some tumor cell lines. Constitutive expression of LIGHT
results in T cell-mediated inflammation that causes tissue
destruction [5], whereas the absence of LIGHT leads to the
deficient T cell responses [6, 7]. It is reported that LIGHT may
bind to three receptors: HVEM, lymphotoxin-b receptor
(LTbR), and decoy receptor 3(DcR3)/TNF receptor 6 [8].
LTbR is prominent on epithelial cells but is absent in T and B
lymphocytes [9], and HVEM is expressed on T, B, and other
hematopoietic cells [4], which is a new member of TNFR
family involved mainly in T cell activation [10]. LIGHT is
also capable of binding to a soluble non-signaling receptor,
C. Li � Y. Shen � D. Liang � F. Yin � H. Ai � S. Zhang (&)
Jiangsu Province Key Laboratory for Molecular and Medical
Biotechnology, Life Sciences College, Nanjing Normal
University, Nanjing 210046, China
e-mail: [email protected]
C. Li � Y. Shen � S. Zhang
Jiangsu Province Key Laboratory for Aquatic Crustacean
Diseases, Life Sciences College, Nanjing Normal University,
Nanjing 210046, China
D. Liang � B. Sun � S. Lin � S. Zhang
Jiangsu Province Key Laboratory of Supramolecular Medicinal
Materials and Technology, Life Sciences College,
Nanjing Normal University, Nanjing 210046, China
123
Mol Cell Biochem (2013) 379:87–96
DOI 10.1007/s11010-013-1630-x

decoy receptor 3(DcR3), which is implicated in T cell
development and homeostasis [11, 12], DC maturation [13],
malignant glioma progression, and immune escape [14].
LIGHT is a 29-kDa type II transmembrane protein
containing an intracellular N-terminal domain, a short
transmembrane segment, an extracellular stalk, and a
C-terminal THD (TNF homology domain) that can fold
into a b-sandwich jelly roll structure that interacts with
adjacent monomers to form a bell-shaped homotrimer [15,
16]. An alternatively spliced LIGHT isoform has been
described as encoding a non-glycosylated molecule form
that lacks the transmembrane region and thus is not dis-
played on the cell surface. The third form of LIGHT is
soluble because metalloprotease activity can cleave LIGHT
and release it from the cell surface [17]. Recent studies
have shown that soluble LIGHT (sLIGHT) promotes ath-
erogenesis by inducing pro-inflammatory cytokines and
matrix metalloproteinases in macrophage/foam cells and
endothelial cells [18].
Mefugu (Takifugu obscurus) is a teleost fish of high
commercial value. It is mainly found in the East China Sea,
the South China Sea, the inland waters in China and the
Korean Peninsula. It lives in the bottom layer of inshore
and inland waters, and grows 20–40 cm in length. Because
of the relatively small size and simple organization of its
genome, Takifugu has become model organism to under-
stand genome architecture, organization, and function [19].
In this study, we cloned the mefugu LIGHT gene and
determined its genomic structure. Furthermore, phyloge-
netic analysis, tissue distribution, predicted three-dimen-
sional structure, and protein expression of the LIGHT gene
from mefugu were conducted.
Materials and methods
Animals and cell preparations
Mefugu were purchased from Jiangsu Zhongyang Group
(Jiangsu, China). Mice were obtained from Nanjing
Qinglongshan Animal Center, Jiangsu, China. All animal
usage was conducted according to the protocol approved
by Institutional Animal Care and Use Committee of Nan-
jing Normal University. The mefugu splenocytes were
separated from spleen homogenate using Lymphocyte
Separation Medium (BD Pharmingen, USA) according to
the manual. The mefugu splenocytes were maintained in
DMEM supplemented with 10 % fetal bovine serum
(FBS), 1 % glutamine, and 1 % penicillin/streptomycin
(P/S) at 37 �C. Mouse T cells were isolated from spleen
using T cell-specific anti-Mouse CD90.2 Magnetic Parti-
cles (BD bioscience). The mouse T cells were maintained
in RPMI1640 medium with penicillin/streptomycin
(Gibco-BRL, USA) supplemented with 10 % FCS at 37 �C
in an atmosphere of 5 % CO2.
Tissue sampling, RNA isolation, and RT-PCR
Various tissues including heart, liver, spleen, gill, kidney,
and intestine from mefugu were collected and immediately
snap frozen in liquid nitrogen and stored at -80 �C until
use. Total RNA was prepared with an RNAprep pure
Tissue Kit (Tiangen Biotech Co. Ltd., Beijing, China)
following the manufacturer’s instruction. A first-strand
cDNA was synthesized from 1 lg RNA isolated from
spleen using Reverse Transcriptase M-MLV (Takara,
Japan) according to the manufacturer’s protocol. A pair of
primers, F1 and R1 (Table 1) was designed and synthe-
sized according to alignments of the predicted sequence of
Tetraodon nigroviridis and Takifugu rubripes. PCR was
conducted with F1 and R1 under the following conditions:
94 �C for 5 min, 30 cycles (94 �C/30 s; 60 �C/30 s; 72 �C/
1 min), and finally 72 �C for 10 min. PCR product was
cloned into pMD19-T vector (Takara) and sequenced.
Bioinformatics analysis
The sequence of open reading frame of fLIGHT was used
to search for similarity using BLAST at web servers of the
National Center of Biotechnology Information (http://blast.
ncbi.nlm.nih.gov/). For the exact localization of the exon–
intron boundaries, the mRNA-to-genomic alignment pro-
gram Spidey was used. The deduced amino acid sequence
was analyzed with the Expert Protein Analysis System
(http://www.expasy.org/). Multiple sequence alignment
was performed using the ClustalW (version 1.83) [20]. The
phylogenetic tree was constructed from the ClustalW-
generated alignments using the neighbor-joining method
and the MEGA4.0 program [21]. The predicted 3D
(three-dimensional) structure of the soluble mature region
of mefugu LIGHT (fsLIGHT) was determined by
Table 1 Primer sequences used in this study
Primer 50–30
F1 GTTGGAGCTGTCCACACCNA
R1 GCTCNAAACNGGGTNNGAG
QL1 TTGAATGCTGCAGAGGAGTG
QL2 CCTGTTTGCAGGTGGAAAAT
QG1 CACCTCCAAGAAGGTGGAAA
QG2 CTCTCGTGGAAAACGGTGAT
A1 CGCCATATG GTGAGCAAGGGCGAGGA
A2 GCTGCCACCTCCACCCTTGTACAGC TCGTC
B1 GGTGGAGGTGGCAGCATCCAAAACCGGCCA
B2 CCCAAGCTTTCACGGGGCTATCATAAAGGCT
88 Mol Cell Biochem (2013) 379:87–96
123

comparative protein modeling on the I-TASSER sever
(http://zhanglab.ccmb.med.umich.edu/ I-TASSER), then
visualized and manipulated with the RasWin molecular
graphics program (RasMol, version 2.7.2). Real-time quan-
tification PCR primer pairs were designed manually based on
Primer3 Input [22].
Real-time quantitative PCR (qPCR) analysis
Tissue distribution of fLIGHT was studied by real-time
qPCR. First-strand cDNAs were synthesized from RNA
obtained from various tissues as described above. A pair of
primers QL1 and QL2 (Table 1) was used to amplify a
PCR product of 197 bp. A constitutively expressed gene,
the glyceraldehyde phosphate dehydrogenase (GAPDH),
was used as an internal control to verify the real-time qPCR
reaction. Two primers QG1 and QG2 (Table 1) were used
to amplify a 171 bp fragment of mefugu GAPDH cDNA.
The program of PCR was 94 �C for 5 min, followed by 40
cycles of 94 �C for 30 s, 65 �C for 30 s, 72 �C for 12 s.
DEPC-water for the replacement of cDNA template was
used as a negative control. The SYBR Green RT-PCR
assay was carried out as previously described in detail [23].
Construction of expression vector pET28a-GFP/
fsLIGHT
The DNA encoding mefugu LIGHT soluble domain (amino
acids 101–253) was amplified by RT-PCR. To obtain GFP/
fsLIGHT fusion gene first, the method of overlapping
polymerase chain reaction (overlap PCR) was used. Four
primers for PCR were designed based on the green fluo-
rescent protein (GFP) and the fsLIGHT encoding sequence
as follows: A1 (sense), A2 (anti-sense), B1 (sense), and B2
(anti-sense) (Table 1). The primer A2 contained a (Gly4-
Ser) linker encoding sequence (Italic and Bold). For
overlap PCR, the primers A2 and B1 had an overlapping
complementary sequence shown with Italic and Bold,
resulting in one fusion cassette containing GFP (Gly4Ser),
linker, and fsLIGHT-encoding sequences. The A1 and the
B2 were introduced with NdeI and HindIII restriction sites,
respectively, for subsequent cloning. The A1 and the A2
were used to amplify the cDNA sequence of GFP without
the stop codon. The B1 and the B2 were used to amplify
the cDNA sequence of fsLIGHT, with pMD19T-fLIGHT
above as template. After the first round of PCR using A1/
A2 and B1/B2, respectively, gel purification was used for
the two products. The second round of PCR was performed
using the two resulting PCR products as templates and A1/
B2 as primers to obtain the cDNA encoding GFP/(Gly4-
Ser)/fsLIGHT fusion protein. Following digestion with
NdeI and HindIII, the PCR product was subcloned into the
pET28a expression vector (Novagen), forming a sequence
encoding a fusion protein of pET28a-GFP/fsLIGHT and an
NH2-terminal His6-tag. The recombinant plasmid was
named as pET28a-GFP/fsLIGHT.
Expression, purification of recombinant GFP/fsLIGHT
and western blotting analysis
The constructed recombinant plasmid pET28a-GFP/fsLIGHT
was transformed into Escherichia coli BL21 (DE3) cells. The
bacteria were cultured in 600 ml Luria–Bertani media with
vigorous shaking (220 rpm) and 30 mg/ml kanamycin at
37 �C to the optical density of OD600 & 0.6. Then, the protein
production was induced with 0.1 mM IPTG (isopropyl b-a-
thiogalactoside) at 16 �C for a further 24 h and shaking speed
was set to 135 rpm/min. The soluble proteins in the super-
natant were collected by refrigerated centrifugation after hy-
persound quassation. Finally, the target proteins were purified
with His-Bind Columns (Qiagen, Germany) according to the
manual. The expression of His6-tagged pET28a-GFP/
fsLIGHT was analyzed by SDS-PAGE and western blotting
with an anti-His6-tag mouse antibody (Invitrogen, USA).
Purified pET28a-GFP/fsLIGHT was used for further studies.
Measurement of fluorescence spectra
The fluorescence spectra of GFP/fsLIGHT and GFP were
measured using the fluorescence spectrophotometer LS50B
(Perkin Elmer, USA). Emission spectra were measured at
510 nm and excitation spectra were measured at 488 nm.
Laser scanning confocal microscopic analysis
The purified splenic lymphocytes from mefugu and mouse
splenic T cells (107 cell/ml) were cultured in RPMI 1640
containing 10 % FBS. The cells were incubated for 90 min
at 37 �C with/without GFP/fsLIGHT (15 lg/ml), and then
washed three times with PBS buffer (pH 7.2). Free GFP
(15 lg/ml) was used as a control protein. The images were
captured on an LSM 510 confocal microscope with a
cooled MicroMax CCD camera.
Results
Molecular cloning and analysis of the genomic
structure of fLIGHT
In this study, the fLIGHT was identified from the spleen
cDNA library of mefugu. The open reading frame (ORF) of
this EST clone was obtained by RT-PCR with primers F1
and R1 (GenBank accession no. JX023508). The 765 bp
ORF encodes a 254-amino acid protein with a putative
molecular weight of 27.7 kDa, and the isoelectric point pI
Mol Cell Biochem (2013) 379:87–96 89
123

is 8.78 (Fig. 1). The nucleotide and predicted amino acid
sequence of the ORF of fLIGHT are shown in Fig. 1. To
obtain the genomic DNA of mefugu LIGHT, the publicly
available mefugu genome database at the ensemble
(http://www.ensembl.org/Takifugu_rubripes/blastview) was
screened using the full-length cDNA sequence of fLIGHT.
The mefugu LIGHT gene spans 1,691 bp at chromosomal
scaffold_194. The gene consists of six exons (Table 2),
Exon 1 of fLIGHT encodes the first 66 amino acids of the
polypeptide, which comprises the entire cytoplasmic tail (aa
1–38), transmembrane domain (aa 39–61), and the begin-
ning of the extracellular stalk region. The remaining exons
encode amino acids 67–89, 90–99,100–111,112–191, and
192–254, respectively, which make up the stalk region and
the trimerization domain. They all belong to the extracel-
lular domain. All exon–intron splice junction sequences
basically conformed to the canonical GT–AG rule (Bold),
which is similar to that of other animals.
Fig. 1 The sequence of the
fLIGHT gene (GenBank
accession no. JX023508). The
entire deduced amino acid
sequence is depicted in single
letter code beneath the
corresponding nucleotide
sequence. The stop codon TGA
is with an asterisk
Table 2 Intron/exon arrangement of fLIGHT
50End 30End Size (bp) Encodes (aa) Size (bp)
Exon1 ATGT- -GAGG 199 1–66 66
Intron1 GTAA- -ACAG 515
Exon2 CGTT- -ACAG 68 67–89 23
Intron2 GTT- -TCAG 112
Exon3 GGGG- -AAAG 31 90–99 10
Intron3 GTAC- -GTGG 147
Exon4 AACA- -ATAG 36 100–111 12
Intron4 GTCA- -GTAG 83
Exon5 GCTC- -AAAG 238 112–191 80
Intron5 GTTA- -CTAG 79
Exon6 TTCT- -TGA stop 193 192–254 63
Fig. 2 a Amino acid sequence alignment of LIGHT from several
species: Fo_LIGHT (Takifugu obscurus, JX023508), Hs_LIGHT (Homosapiens, NM_003807), Mm_LIGHT (Mus musculus, NM_019418.2),
and Om_LIGHT (Oncorhynchus mykiss, NP_001118039.1). Multiple
alignments were first performed using ClustalW program. Asteriskidentical residues among the four sequences. Substitution said to be
conservative or semi-conservative by ClustalWare are with semicolonand end period, respectively. The transmembrane domain was unlined.
The thick black solid line below LIGHT sequences is the conserved TNF
homology domain (THD). The TNF homology domain (THD) that is
typically composed of 10 b-strands (designated A, A0, B0, B, C, D, E, F, G,
and H),which are in blue. The light grey is the potential cleave site.
b Comparison of the predicted mefugu LIGHT protein sequences from
our study and Ensembl
c
90 Mol Cell Biochem (2013) 379:87–96
123

Mol Cell Biochem (2013) 379:87–96 91
123
Sequence comparison analysis
The fLIGHT comprises 254 amino acids, containing a
predicted transmembrane domain (TMD) of 21 amino
acids and a extracellular domain which include a cleave
site (Gln101, Ile102) can be cleaved form a soluble protein.
To study the evolutionary relationship of mefugu LIGHT
with other known LIGHT proteins, amino acid sequences
comparison was performed using the ClustalW software
(Fig. 2a). By analysis of the primary structure, fLIGHT
protein is a trimerization polypeptide, the receptor-binding
domain includes a site for N-linked glycosylation (Asn79)
[24]. Based on structural homology with LTa [25] and
supportive evidence from modeling and biochemical
analysis, the trimerization domain of LIGHT folds into an
anti-parallel b sandwich, wherein the receptor-binding sites
are formed from adjacent subunits [26]. A search of the
genome revealed the presence of a T. rubripes LIGHT
cDNA (Ensembl: scaffold_194: 420,541-422,231) and its
genomic DNA sequence. The fLIGHT sequence differs by
four amino acids from the sequence of T. rubripes in
Ensembl (Fig. 2b).
Three-dimensional modeling and phylogenetic analysis
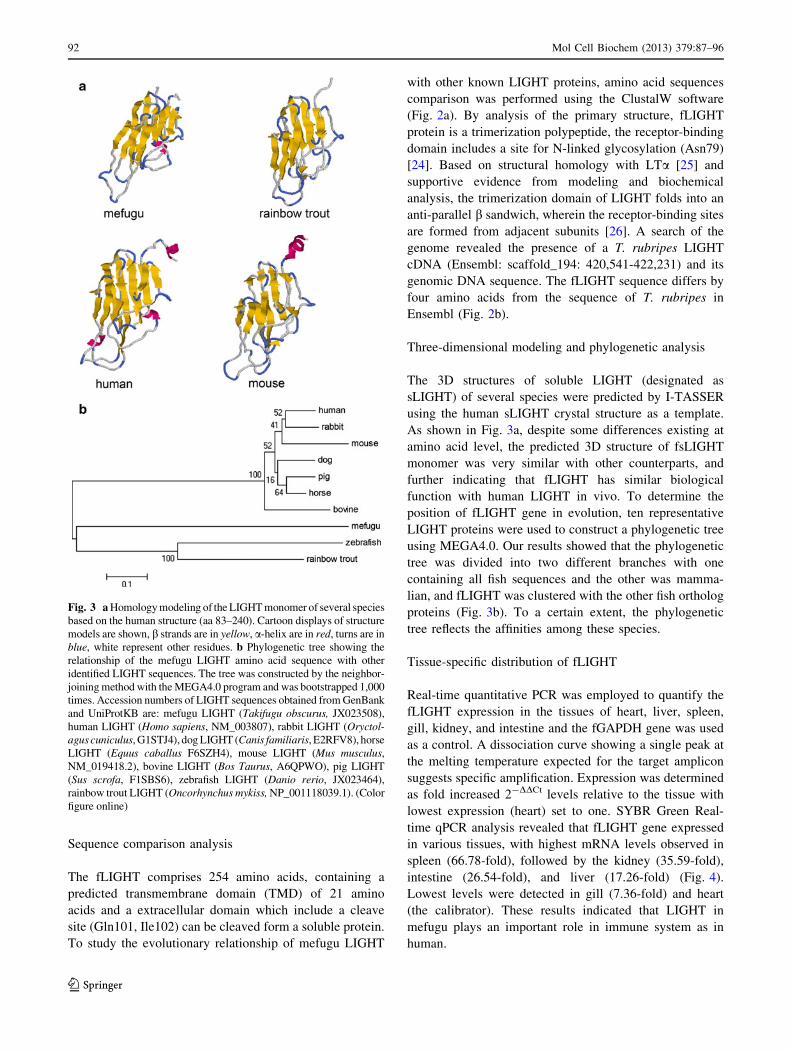
The 3D structures of soluble LIGHT (designated as
sLIGHT) of several species were predicted by I-TASSER
using the human sLIGHT crystal structure as a template.
As shown in Fig. 3a, despite some differences existing at
amino acid level, the predicted 3D structure of fsLIGHT
monomer was very similar with other counterparts, and
further indicating that fLIGHT has similar biological
function with human LIGHT in vivo. To determine the
position of fLIGHT gene in evolution, ten representative
LIGHT proteins were used to construct a phylogenetic tree
using MEGA4.0. Our results showed that the phylogenetic
tree was divided into two different branches with one
containing all fish sequences and the other was mamma-
lian, and fLIGHT was clustered with the other fish ortholog
proteins (Fig. 3b). To a certain extent, the phylogenetic
tree reflects the affinities among these species.
Tissue-specific distribution of fLIGHT
Real-time quantitative PCR was employed to quantify the
fLIGHT expression in the tissues of heart, liver, spleen,
gill, kidney, and intestine and the fGAPDH gene was used
as a control. A dissociation curve showing a single peak at
the melting temperature expected for the target amplicon
suggests specific amplification. Expression was determined
as fold increased 2-DDCt levels relative to the tissue with
lowest expression (heart) set to one. SYBR Green Real-
time qPCR analysis revealed that fLIGHT gene expressed
in various tissues, with highest mRNA levels observed in
spleen (66.78-fold), followed by the kidney (35.59-fold),
intestine (26.54-fold), and liver (17.26-fold) (Fig. 4).
Lowest levels were detected in gill (7.36-fold) and heart
(the calibrator). These results indicated that LIGHT in
mefugu plays an important role in immune system as in
human.
Fig. 3 a Homology modeling of the LIGHT monomer of several species
based on the human structure (aa 83–240). Cartoon displays of structure
models are shown, b strands are in yellow, a-helix are in red, turns are in
blue, white represent other residues. b Phylogenetic tree showing the
relationship of the mefugu LIGHT amino acid sequence with other
identified LIGHT sequences. The tree was constructed by the neighbor-
joining method with the MEGA4.0 program and was bootstrapped 1,000
times. Accession numbers of LIGHT sequences obtained from GenBank
and UniProtKB are: mefugu LIGHT (Takifugu obscurus, JX023508),
human LIGHT (Homo sapiens, NM_003807), rabbit LIGHT (Oryctol-agus cuniculus, G1STJ4), dog LIGHT (Canis familiaris, E2RFV8), horse
LIGHT (Equus caballus F6SZH4), mouse LIGHT (Mus musculus,NM_019418.2), bovine LIGHT (Bos Taurus, A6QPWO), pig LIGHT
(Sus scrofa, F1SBS6), zebrafish LIGHT (Danio rerio, JX023464),
rainbow trout LIGHT (Oncorhynchus mykiss, NP_001118039.1). (Color
figure online)
92 Mol Cell Biochem (2013) 379:87–96
123
Expression and purification of the GFP/fsLLIGHT
protein
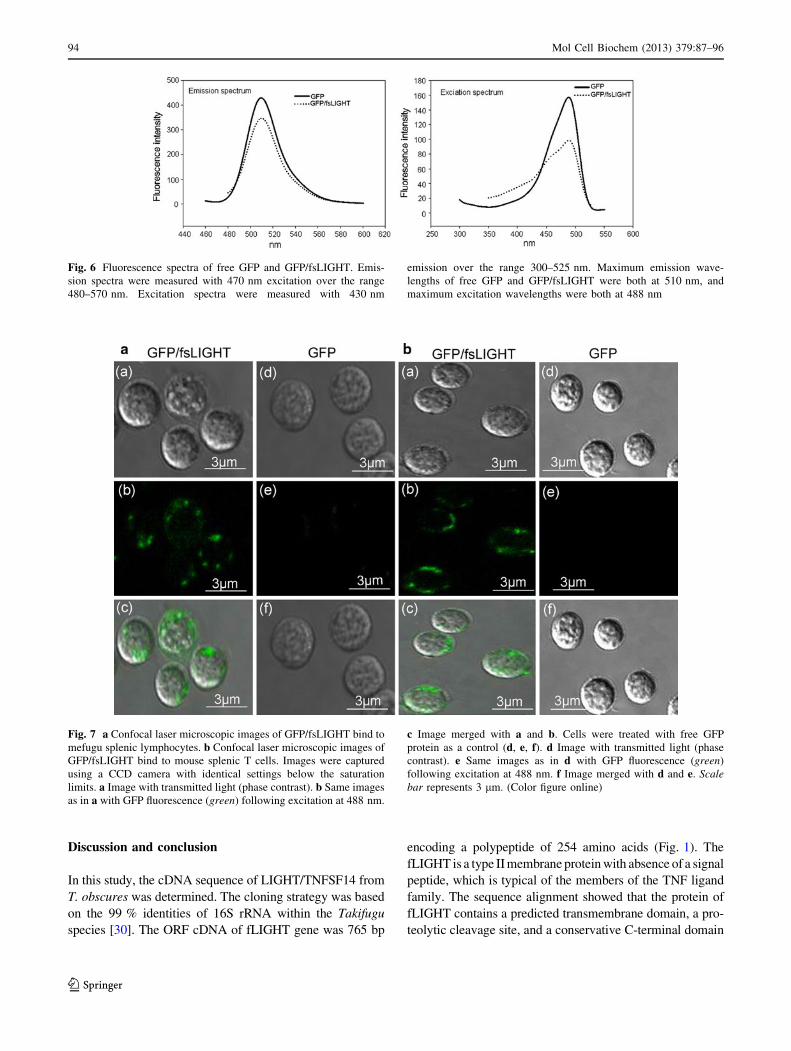
The GFP/fsLIGHT plasmids were transformed into E. coli
BL21 (DE3), and the protein was efficiently expressed and
purified under optimal conditions. According to the SDS-
PAGE analysis, after IPTG induction, 44.4 kDa band (Fig. 5,
lane 3) corresponding to the expectant size of the fusion
protein was observed. There was almost no basal expression of
soluble fusions without IPTG induction (Fig. 5, lane 2). Sol-
uble recombinant GFP/fsLIGHT was successfully purified
with His-Bind columns (Fig. 5, lane 4). The expressed GFP/
fsLIGHT was recognized by western blotting using anti-His6-
tag monoclonal antibody (Fig. 5, lane 5).
Fluorescence activity
GFP is a spontaneously fluorescent protein isolated from
marine organisms, such as certain jellyfish and sea pansies [27].
The fluorophore of GFP originates from an internal Ser-Tyr-
Gly sequence which is post-translationally modified to a 4-(p-
hydroxybenzylidene)-imidazolidin-5-one structure [28]. When
GFP is fused to the N-terminus of over-expressed globular
proteins, over-expression in soluble form allows GFP to fold
correctly. To verify the correct folding of the GFP domain, the
purified soluble GFP/fsLIGHT was tested for its fluorescence
activity with comparison to free GFP. As determined by the
measurement of fluorescence spectra using the florescence
spectrophotometer, fluorescence properties of GFP/fsLIGHT
were quite similar to free GFP. Their maximum excitation
wavelengths were both at 488 nm, and the maximum emission
wavelengths were both at 510 nm. However, the fluorescence
intensity exhibited weaker than free GFP (Fig. 6). It indicates
no shift in excitation and emission spectra of GFP/fsLIGHT
fusion protein compared to free GFP. The fluorophore is pro-
tected from collisions with fluorescence quenchers [29], so the
decrease of fluorescence in chimera might be explained by one
mechanism: the fluorophore is formed but the fluorescence is
quenched by fsLIGHT, which could absorb the inducing or
induced radiation.
Recombinant GFP/fsLIGHT binds to mefugu
lymphocytes or mouse splenic T cells
GFP is widely used as a fluorescent probe, no need for
substrates and co-factors, being active as a monomer, and
exhibiting stability in a broad range of buffers and tem-
peratures. Localization of GFP fusion proteins can be
analyzed in live bacteria and eukaryotic cells. In view of
this, laser scanning confocal microscopy (LSCM) provided
a visibly clear image of the distribution of GFP/fsLIGHT
binding to mefugu lymphocytes and mouse splenic T cells
by detection of fluorescent signal of GFP. However, cells
treated with free GFP protein did not release any fluores-
cence signals (Fig. 7). The findings indicate that fsLIGHT
has the binding ability to mefugu lymphocytes and mouse
splenic T cells.
Fig. 4 Expression analysis of mefugu LIGHT in various tissues.
Real-time qPCR analysis of the expression of fLIGHT. Mean mRNA
levels in six tissues, heart, liver, spleen, gill, kidney, and small
intestine, were analyzed. Data are 2-DDCt levels calculated relative to
the tissue with lowest expression (heart) set to 1, normalized against
GAPDH mRNA levels. Vertical bars the mean ± SE (N = 3)
Fig. 5 SDS-PAGE analysis of recombinant GFP/fsLIGHT expressed
in E. coli BL21 (DE3). Molecular weight marker is in the left lane.
Lane 1 the pure GFP protein. Lane 2 Cell lysates of bacteria
transformed with pET28a-GFP/fsLIGHT without IPTG induction.
Lane 3 cell lysates of bacteria transformed with pET28a-GFP/
fsLIGHT under IPTG (0.1 mM) induction. Lane 4 the GFP/fsLIGHT
fusion protein purified by immobilized metal affinity chromatogra-
phy. Lane 5 western blot analysis of purified GFP/fsLIGHT using
mAb against His6-tag. The arrow the location of GFP/fsLIGHT
fusion protein
Mol Cell Biochem (2013) 379:87–96 93
123
Discussion and conclusion
In this study, the cDNA sequence of LIGHT/TNFSF14 from
T. obscures was determined. The cloning strategy was based
on the 99 % identities of 16S rRNA within the Takifugu
species [30]. The ORF cDNA of fLIGHT gene was 765 bp
encoding a polypeptide of 254 amino acids (Fig. 1). The
fLIGHT is a type II membrane protein with absence of a signal
peptide, which is typical of the members of the TNF ligand
family. The sequence alignment showed that the protein of
fLIGHT contains a predicted transmembrane domain, a pro-
teolytic cleavage site, and a conservative C-terminal domain
Fig. 6 Fluorescence spectra of free GFP and GFP/fsLIGHT. Emis-
sion spectra were measured with 470 nm excitation over the range
480–570 nm. Excitation spectra were measured with 430 nm
emission over the range 300–525 nm. Maximum emission wave-
lengths of free GFP and GFP/fsLIGHT were both at 510 nm, and
maximum excitation wavelengths were both at 488 nm
Fig. 7 a Confocal laser microscopic images of GFP/fsLIGHT bind to
mefugu splenic lymphocytes. b Confocal laser microscopic images of
GFP/fsLIGHT bind to mouse splenic T cells. Images were captured
using a CCD camera with identical settings below the saturation
limits. a Image with transmitted light (phase contrast). b Same images
as in a with GFP fluorescence (green) following excitation at 488 nm.
c Image merged with a and b. Cells were treated with free GFP
protein as a control (d, e, f). d Image with transmitted light (phase
contrast). e Same images as in d with GFP fluorescence (green)
following excitation at 488 nm. f Image merged with d and e. Scalebar represents 3 lm. (Color figure online)
94 Mol Cell Biochem (2013) 379:87–96
123

known as the THD like other LIGHTs (Fig. 2a). The THD is
typically composed of ten b-strands (designated as A, A0, B0,B, C, D, E, F, G, and H), wherein the receptor-binding sites are
formed. The four amino acid replacements may indicate that
the Takifugu species diversified very recently (Fig. 2b). As
shown in Fig. 3a, the predicted 3D structure of fsLIGHT is
very similar to its human counterpart, indicating that mefugu
LIGHT may have similar biological function in vivo with
human LIGHT. Phylogenetic tree analysis showed that
fLIGHT protein is grouped primarily with other teleost
LIGHT molecules (Fig. 3b). It is consistent with the previous
studies such as Glenney et al. [31]. The real-time quantitative
PCR results showed that fLIGHT was constitutively expres-
sed in all detected tissues, with high mRNA level observed in
fish-related immune organs, especially in the lymphoid organ
spleen (Fig. 4), indicating that fLIGHT could play important
immunological roles.
To determine the activity of the fsLIGHT, we con-
structed fusion protein consisting of GFP and fsLIGHT.
First, we expressed recombinant GFP/fsLIGHT in E. coli
BL21 (DE3) using the pET28a plasmid. The protein effi-
ciently expressed and was mostly found in soluble region
under hypothermia induction. Then, the soluble recombi-
nant GFP/fsLIGHT was successfully purified using His-
Bind columns. Western blot analysis of GFP/fsLIGHT
purified from BL21 (DE3) revealed a prominent band at
44.4 kDa, indicative of GFP/fsLIGHT soluble expression.
Fluorescence analysis showed that fluorescence properties
of GFP/fsLIGHT were quite similar to free GFP despite the
fluorescence intensity exhibited weaker than free GFP
(Fig. 6). It indicates that fsLIGHT do not damage the
correct folding of the GFP fluorophore.
LSCM was used to determine receptor-binding activity.
It was found that GFP/fsLIGHT could be able to bind to the
receptor-positive cell lines, including mouse LIGHT
receptors (Fig. 7). This result suggested that, due to the
highly conserved nature of LIGHT, functional cross-reac-
tivity exists among different species.
In conclusion, this is the first report of cloning and
molecular structure research of mefugu LIGHT. It provides
information regarding the molecular evolution of fish,
mRNA expression in various tissues, the fused protein
construction, expression, and receptor-binding activity.
LIGHT play important roles in the immune system, so
LIGHT protein may be developed as fish biologics for
mefugu, which is a very important economic fish. More-
over, this study may be useful for further research into
LIGHT using mefugu as a model system.
Acknowledgments This study was funded by a grant from The
National Science Foundation of China (No. 30271093) and a project
funded by the Priority Academic Program Development (PAPD) of
Jiangsu Higher Education Institution (No. 164320H106).
References
1. Locksley RM, Killeen N, Lenardo MJ (2001) The TNF and TNF
receptor superfamilies: integrating mammalian biology. Cell
104:487–501
2. Smith CA, Farrah T, Goodwin RG (1994) The TNF receptor
superfamily of cellular and viral proteins: activation, costimula-
tion, and death. Cell 76:959–962
3. Doherty TA, Soroosh P, Khorram N, Fukuyama S, Rosenthal P,
Cho JY, Norris PS, Choi H, Scheu S, Pfeffer K, Zuraw BL, Ware
CF, Broide DH, Croft M (2011) The tumor necrosis factor family
member LIGHT is a target for asthmatic airway remodeling. Nat
Med 17:596–603. doi:10.1038/nm.2356
4. Mauri DN, Ebner R, Montgomery RI, Kochel KD, Cheung TC,
Yu GL, Ruben S, Murphy M, Eisenberg RJ, Cohen GH, Spear
PG, Ware CF (1998) LIGHT, a new member of the TNF super-
family, and lymphotoxic a are ligands for herpesvirus entry
mediator. Immunity 8:21–30
5. Shaikh RB, Santee S, Granger SW, Butrovich K, Cheung T,
Kronenberg M, Cheroutre H, Ware CF (2001) Constitutive
expression of LIGHT on T cells leads to lymphocyte activation,
inflammation, and tissue destruction. J. Immunol 167:6330–6337
6. Tamada K, Shimozaki K, Chapoval AI, Zhai Y, Su J, Chen SF,
Hsieh SL, Nagata S, Ni J, Chen L (2000) LIGHT, a TNF-like
molecule, costimulates T cell proliferation and is required for
dendritic cell-mediated allogeneic T cell response. J. Immunol
164:4105–4110
7. Tamada K, Shimozaki K, Chapoval AI, Zhu G, Sica G, Flies D,
Boone T, Hsu H, Fu YX, Nagata S, Ni J, Chen L (2000) Mod-
ulation of T-cell-mediated immunity in tumor and graftversus-
host-disease models through the LIGHT co-stimulatory pathway.
Nat Med 6:283–289. doi:10.1038/73136
8. Hu G, Liu Y, Li H, Zhao D, Yang L, Shen J, Hong X, Cao X,
Wang Q (2010) Adenovirus-mediated LIGHT gene modification
in murine B-cell lymphoma elicits a potent antitumor effect. Cell
Mol Immunol 7:296–305. doi:10.1038/cmi.2010.15
9. Crowe PD, VanArsdale TL, Walter BN, Ware CF, Hession C,
Ehrenfels B, Browning JL, Din WS, Goodwin RG, Smith CA
(1994) A lymphotoxin-beta-specific receptor. Science
264:707–710. doi:10.1126/science.8171323
10. Kwon BS, Tan KB, Ni J, Lee KO, Kim KK, Kim YJ, Wang S,
Gentz R, Yu GL, Harrop J, Lyn SD, Silverman C, Porter TG,
Truneh A, Young PR (1997) A newly identified member of the
tumor necrosis factor receptor superfamily with a wide tissue
distribution and involvement in lymphocyte activation. J Biol
Chem 272:14272–14276. doi:10.1074/jbc.272.22.14272
11. Migone TS, Zhang J, Luo X, Zhuang L, Chen C, Hu B, Hong JS,
Perry JW, Chen SF, Zhou JX, Cho YH, Ullrich S, Kanakaraj P,
Carrell J, Boyd E, Olsen HS, Hu G, Pukac L, Liu D, Ni J, Kim S,
Gentz R, Feng P, Moore PA, Ruben SM, Wei P (2002) TL1A is a
TNF-like ligand for DR3 and TR6/DcR3 and functions as a T cell
costimulator. Immunity 16:479–492. doi:10.1016/S1074-7613
(02)00283-2
12. Shi G, Luo H, Wan X, Salcedo TW, Zhang J, Wu J (2002) Mouse T
cells receive costimulatory signals from LIGHT, a TNF family
member. Blood 100:3279–3286. doi:10.1182/blood-2002-05-1404
13. Hsu TL, Chang YC, Chen SJ, Liu YJ, Chiu AW, Chio CC, Chen
L, Hsieh SL (2002) Modulation of dendritic cell differentiation
and maturation by decoy receptor 3. J Immunol 168:4846–4853
14. Roth W, Isenmann S, Nakamura M, Platten M, Wick W, Kleihues
P, Bahr M, Ohgaki H, Ashkenazi A, Weller M (2001) Soluble
decoy receptor 3 is expressed by malignant gliomas and sup-
presses CD95 ligand-induced apoptosis and chemotaxis. Cancer
Res 61:2759–2765
Mol Cell Biochem (2013) 379:87–96 95
123

15. Harrop JA, McDonnell PC, Brigham-Burke M, Lyn SD, Minton
J, Tan KB, Dede K, Spampanato J, Silverman C, Hensley P,
DiPrinzio R, Emery JG, Deen K, Eichman C, Chabot-Fletcher M,
Truneh A, Young PR (1998) Herpesvirus entry mediator ligand
(HVEM-L), a novel ligand for HVEM/TR2, stimulates prolifer-
ation of T cells and inhibits HT29 cell growth. J Biol Chem
73:27548–27556. doi:10.1074/jbc.273.42.27548
16. Bodmer JL, Schneider P, Tschopp J (2002) The molecular
architecture of the TNF superfamily. Trends Biochem Sci
27:19–26. doi:10.1016/S0968-0004(01)01995-8
17. Granger SW, Rickert S (2003) LIGHT–HVEM signaling and the
regulation of T cell-mediated immunity. Cytokine Growth F R
14:289–296
18. Otterdal K, Smith C, Øie E, Pedersen TM, Yndestad A, Stang E,
Endresen K, Solum NO, Aukrust P, Damas JK (2006) Platelet-derived
LIGHT induces inflammatory responses in endothelial cells and
monocytes. Blood 108:928–935. doi:10.1182/blood-2005-09-010629
19. Aparicio S, Chapman J, Stupka E et al (2002) Whole-genome
shotgun assembly and analysis of the genome of Fugu rubripes.
Science 297:1301–1310. doi:10.1126/science.1072104
20. Thompson JD, Higgins DG, Gibson TJ (1994) Clustal W:
improving the sensitivity of progressive multiple sequence
alignment through sequence weighting, position-specific gap
penalties and weight matrix choice. Nucleic Acids Res
22:4673–4680. doi:10.1093/nar/22.22.4673
21. Tamura K, Dudley J, Nei M, Kumar S (2007) MEGA4: molecular
evolutionary genetics analysis (MEGA) software version 4.0.
Mol Biol Evol 24:1596–1599. doi:10.1093/molbev/msm092
22. Rozen S, Skaletsky HJ (2000) Primer3 on the WWW for general
users and for biologist programmers. In: Krawetz S, Misener S
(eds) Bioinformatics methods and protocols: methods in molec-
ular biology. Humana, Totowa, pp 365–386
23. Guan ZB, Shui Y, Zhang SQ (2007) Two related ligands of the
TNF family, BAFF and APRIL, in rabbit: molecular cloning, 3D
modeling, and tissue distribution. Cytokine 39:192–200
24. Granger SW, Butrovich KD, Houshmand P, Edwards WR, Ware
CF (2001) Genomic characterization of LIGHT reveals linkage to
an immune response locus on chromosome 19p13.3 and distinct
isoforms generated by alternate splicing or proteolysis. J Immu-
nol 167:5122–5128
25. Eck MJ, Sprang SR (1989) The structure of tumor necrosis fac-
tor-a at 2.6 A resolution. Implications for receptor binding. J Biol
Chem 264:17595–17604
26. Rooney IA, Butrovich KD, Glass AA, Borboroglu S, Benedict
CA, Whitbeck JC, Cohen GH, Eisenberg RJ, Ware CF (2000)
The lymphotoxin-b receptor is necessary and sufficient for
LIGHT-mediated apoptosis of tumor cells. J Biol Chem
275:14307–14315. doi:10.1074/jbc.275.19.14307
27. George N, Phillips JR (1997) Structure and dynamics of green
fluorescent protein. Curr Opin Struc Biol 7:821–827
28. Cody CW, Prasher DC, Westler WM, Prendergast FG, Ward WW
(1993) Chemical structure of the hexapeptide chromophore of the
Aequorea green-fluorescent protein. Biochemistry 32:1212–1218.
doi:10.1021/bi00056a003
29. Rao BDN, Kemple MD, Prendergast FG (1980) Proton nuclear
magnetic resonance and fluorescence spectroscopic studies of
segmental mobility in aequorin and a green fluorescent protein
from Aequorea forskalea. Biophys J 32:630–632
30. Kato A, Doi H, Nakada T, Sakai H, Hirose S (2005) Takifuguobscurus is a euryhaline fugu species very close to Takifugurubripes and suitable for studying osmoregulation. BMC Physiol
5:1–11. doi:10.1186/1472-6793-5-18
31. Glenney GW, Wiens GD (2007) Early diversification of the TNF
superfamily in teleosts: genomic characterization and expression
analysis. J Immunol 178:7955–7973
96 Mol Cell Biochem (2013) 379:87–96
123