comparative mitochondrial zygomycetes: bacterial-like rnase p rnas, mobile elements and a close...
TRANSCRIPT

Comparative mitochondrial zygomycetes: bacterial-like Rnase
P RNAs, mobile elements and a close source of the group I intron
invasion in angiosperm
Presented by:Somayeh Haji Kazem Nili Kalyani Rajalingham
Seif et al., (2005) NAR 33(2): 734-744

Summary The mtDNAs from three distantly related
zygomycetes have been sequenced• Rhizopus oryzae (R.oryzae)• Mortierella verticillata (M.verticillata)• Smittium culisetae (S.culisetae)
AIM Analyze comparative mitochondrial genome
• Remediate the lack of data for phylogenetic inferences
• Facilitate biochemical investigations
Seif et al., (2005) NAR 33(2): 734-744

Background Fungi constitute four phylum of highly diverse organisms
Ascomycetes: complete nuclear and mitochondrial sequences of more than dozen are known
Chytrids : few mitochondrial sequences are known
zygomycete:
No mitochondrial gene sequences are known
Nuclear gene sequences are limited (mostly rRNA sequences)
Impossible to determine if zygomycota is a monophyletic taxon
Sequence complete mtDNAs sequencing from several zygomycetes to remediate this situation
Seif et al., (2005) NAR 33(2): 734-744
• Ascomycota • Zygomycota
• Basidiomycota • Chatridiomycota

Research components1) Compare mitochondrial genomes
2) Seven newly identified zygomycete mitochondrial RNase P (mtP-RNAs)
3) Are zygomycetes monophyletic?
4) Group I introns invasion of cox1 gene in angiosperm originated in zygomycete close to Rhizopus Seif et al., (2005) NAR 33(2): 734-744
• Gene content • Genetic code
• Gene organization • Conserved 3’RNA processing site
• Expression • Secondary structure

Introduction Mitochondrial phylogenies
Base on 13 protein sequences Resolves deep divergences in the fungal and animal lineage
Zygomycete mtDNA are of considerable interest for comparative gene expression studies
Zygomycete mtDNA contains rnpB gene Encodes the RNA subunit of RNase P Absent in basidiomycete and chytridiomycete Patchy distribution in ascomycetes No published data on zygomycetes
The RNA subunit of mitochondrial RNase P (mtP-RNA) The enzymatically active part of an endonuclease Has various size and sequences Complicated identification Seif et al., (2005) NAR 33(2): 734-744

Zygomycete mtDNAsShape and Size
Are linear multimeric concatemers in vivo
Mapped as circular molecules
Size ranges between 54-58 kb
Coding regions• R.oryzae: 40.6%• M.verticillata: 43.1%• S.culisetae: 35.3%
Coding regions nad2/nad3 and nad4L/nad5 of R.oryzae overlap by 1 nt
Seif et al., (2005) NAR 33(2): 734-744

Zygomycete mtDNAsComponents
Creator/Presenter: Somayeh Haji Kazem Nili Seif et al., (2005) NAR 33(2): 734-744
The basic fungal set of genes
Full set of tRNAs
RNA component of mitochondrial RNase P
(rnpB)
RNA component of ribosomal protein (rps3) rps3 lacking in R.oryzae
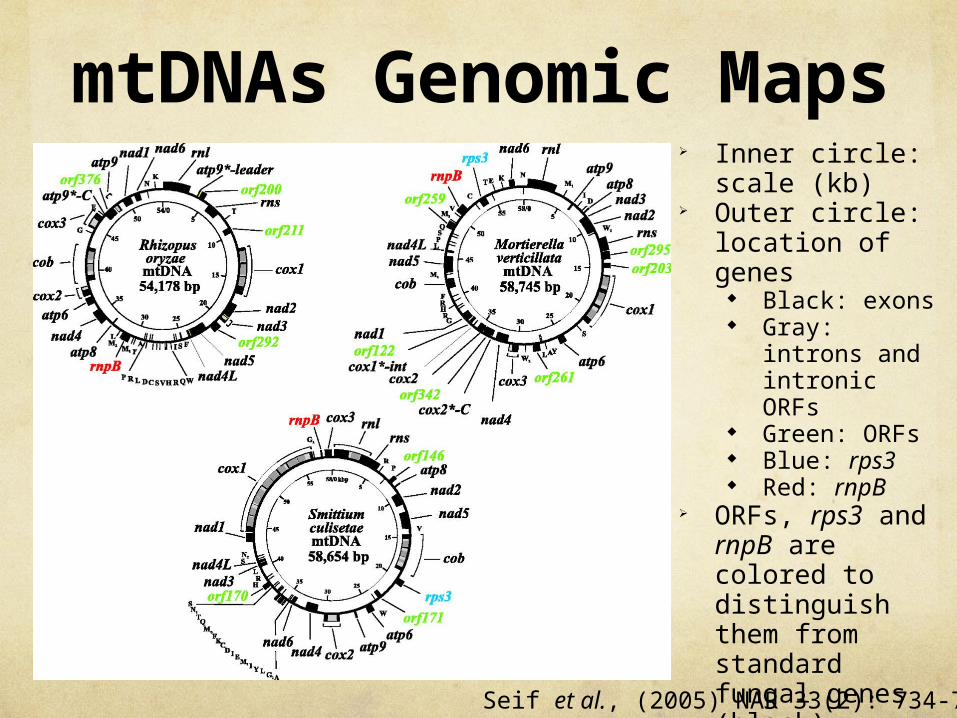
mtDNAs Genomic Maps
Seif et al., (2005) NAR 33(2): 734-744
Inner circle: scale (kb)
Outer circle: location of genes
Black: exons Gray: introns
and intronic ORFs
Green: ORFs Blue: rps3 Red: rnpB
ORFs, rps3 and rnpB are colored to distinguish them from standard fungal genes (black).

Standard Protein Coding Genes
R.oryzae and S.culisetae retain standard translation code for protein coding genes
M.verticillata reassigned two UGA ‘stop’ codons as tryptophan ‘Trp’ on:• nad3• nad4
UGA (Trp) codons are also exist in S.culisetae intronic ORFs:• ORF283 • ORF248
Seif et al., (2005) NAR 33(2): 734-744

UGA (Trp) of Intronic ORFs (283 & 248)
Encoding group I introns homing endonucleases of the LAGLI-DADG type
UGA (Trp) at amino acid position 237 of ORF248• Distinctive motif of this class of endonuclease• Highly conserved • Possibly is a vestige of horizontal intron transfer from a fungus
adapted to this translation code• Closely related to ORF313 of Podospora anserina
S.culisetae and M.verticillata do not encode trnW(uca) • trnW(uca): tRNA recognizing UGA and UGG (Trp) • Assumed UGA codons are inefficiently encoded by trnW(uca)• Alternatively modified or partially edited C in the wobble position of
the anticodon may allow efficient recognition of UGA (Trp)Seif et al., (2005) NAR 33(2): 734-744

tRNA Complete Set Zugomycete mtDNAs encode sufficient tRNA to
recognize all encountered codons
trnI(cau) is absent in R.oryzae• ATA(Ile) codons are absent in standard mitochondrial
genes• ATA(Ile) occur in intronic ORFs
i. The tRNA require for ATA(Ile) translation is imported from cytoplasm to recognize these codons
ii. The intronic ORFs are neither translated nor required for intron splicing
iii. These codon positions are recognized by other tRNAs at low efficiency, resulting amino acid misincorporation
iv. Such unexpected codon usage reflects horizontal intron transfer from species that are adapted to the use of UGA(Trp) and/or ATA(Ile) Seif et al., (2005) NAR 33(2): 734-744

mtP-RNAs Mitochondrial RNA subunit of RNase P
Encoded by rnpB genes
rnpB genes are from outside fungi• Only present in the green alga and various
jakobids• In some ascomycetes • Absent in basidiomycetes and chytridiomycetes • Their presence in all three zygomycetes is
striking
Seif et al., (2005) NAR 33(2): 734-744

mtP-RNA S.culisetae
Close to smallest nrp gene (145 nt) Has highly reduced structure Secondary structure identical to budding yeasts mtP-RNA Perfectly matching the minimum consensus secondary structure
of fungal mtP-RNAs
M.verticillata and R.oryzae Largest nrp gene (980 and 830 bp) Secondary structure identical to bacterial mtP-RNA 3’ end of mtP-RNA in M.verticillata is 9 nt longer than proposed
secondary structure• Elongated by a cytidin-rich stretch sequence
5’ end of mtP-RNA in M.verticillata is and both 3’ and 5’ends of nrp gene in R.oryzae are match the proposed secondary structure and reveal little heterogeneity of mtP-RNA terminiSeif et al., (2005) NAR 33(2): 734-744
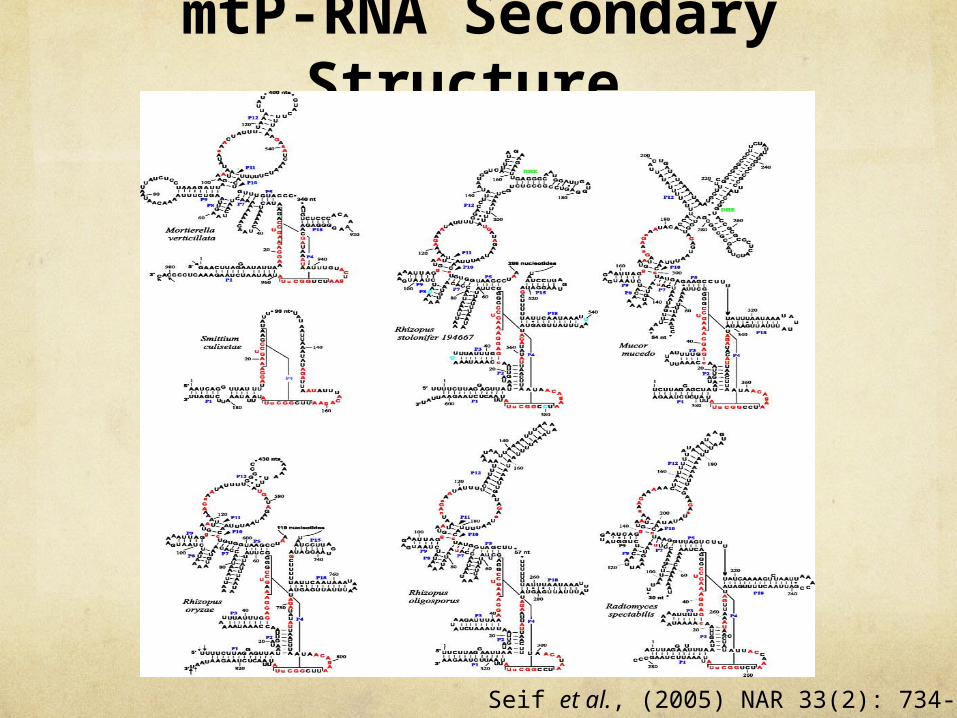
mtP-RNA Secondary Structure
Seif et al., (2005) NAR 33(2): 734-744

mtP-RNA Secondary Structure Zygomycete mtP-RNA structures cover an unprecedented wide range of
intermediate stages in loss of RNA structural elements
R.oryzae mtP-RNA structure The most bacteria like secondary structures Containing almost all structural elements of the bacterial minimum secondary
structure consensus Lacks P13, P14 and P19 helixes (putative-alpha helical structures) If R.oryzae mtP-RNA structure had these sites it was resemble to protist mtP-RNA
M.verticillata mtP-RNA structure Resemble to mtP-RNA of ascomycete (Taphrina deformans) Lacks omnipresent P2 sihelix
S.culisetae mtP-RNA structure The tiny yeast like mtP-RNA molecule Lacks the omnipresent P2 helix
P2 helix absence in M.verticillata and S.culisetae mtP-RNA structure indicates its loss in a common ancestor Seif et al., (2005) NAR 33(2): 734-744

mtP-RNA Insertion Regions The large size of M.verticillata and R.oryzae mtP-RNA is due
to insertions at the J5-15 and J5-18 junctions in the P12 helix
These insertion sequences can be folded into double hairpin structure
mtP-RNA cDNA sequence of R.oryzae were amplified by PCR to determine whether these regions are conserved regions or more variable insertion elements or introns
They are not conserved regions, as their presence in mtP-RNAs pinpoints structural regions are not critical for RNase P activity
They are not introns because the cDNA sequence is identical to the genomic sequence
They are variable insertion elements:• Variations in insertion points and sizes indicates that they have
been acquired recently and independentlySeif et al., (2005) NAR 33(2): 734-744

Conserved C-rich motifs in mRNAs and SSU-rRNA
SSU-rRNA = Small subunit of Ribosomal RNA

Conserved C-rich motifs in mRNAs and SSU-rRNA
SSU-rRNA = Small subunit of Ribosomal RNA
C-rich clusters
- downstream of mt-protein- and SSU-rRNA coding regions
- site of 3′ RNA processing, and are retained in the mature RNA molecules
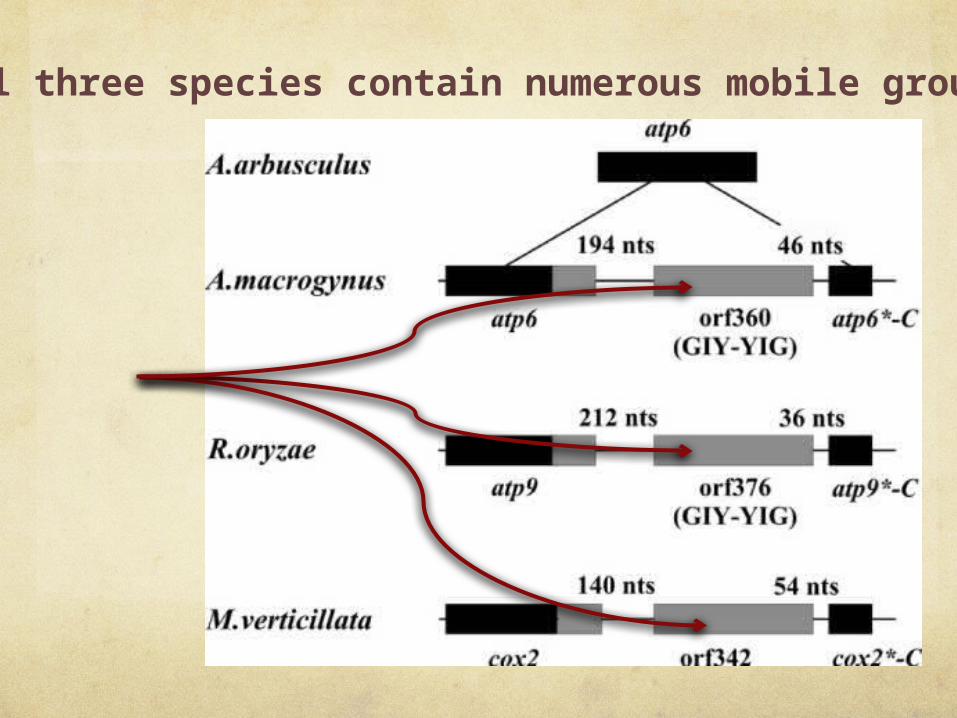
genes of all three species contain numerous mobile group I introns
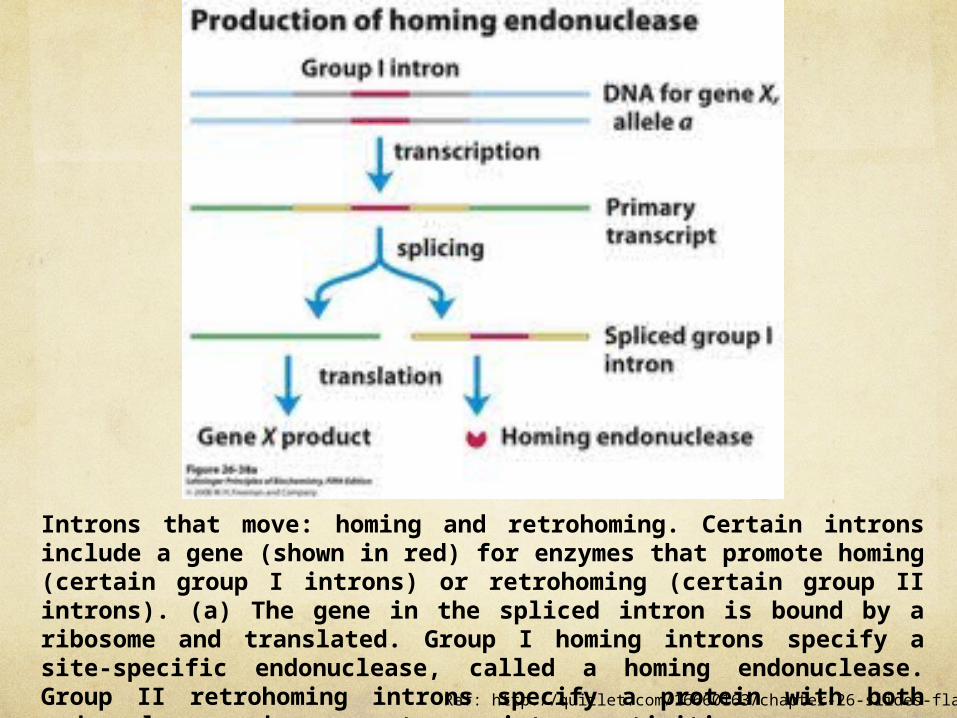
Introns that move: homing and retrohoming. Certain introns include a gene (shown in red) for enzymes that promote homing (certain group I introns) or retrohoming (certain group II introns). (a) The gene in the spliced intron is bound by a ribosome and translated. Group I homing introns specify a site-specific endonuclease, called a homing endonuclease. Group II retrohoming introns specify a protein with both endonuclease and reverse transcriptase activities.
Ref: http://quizlet.com/16660163/chapter-26-slides-flash-cards/
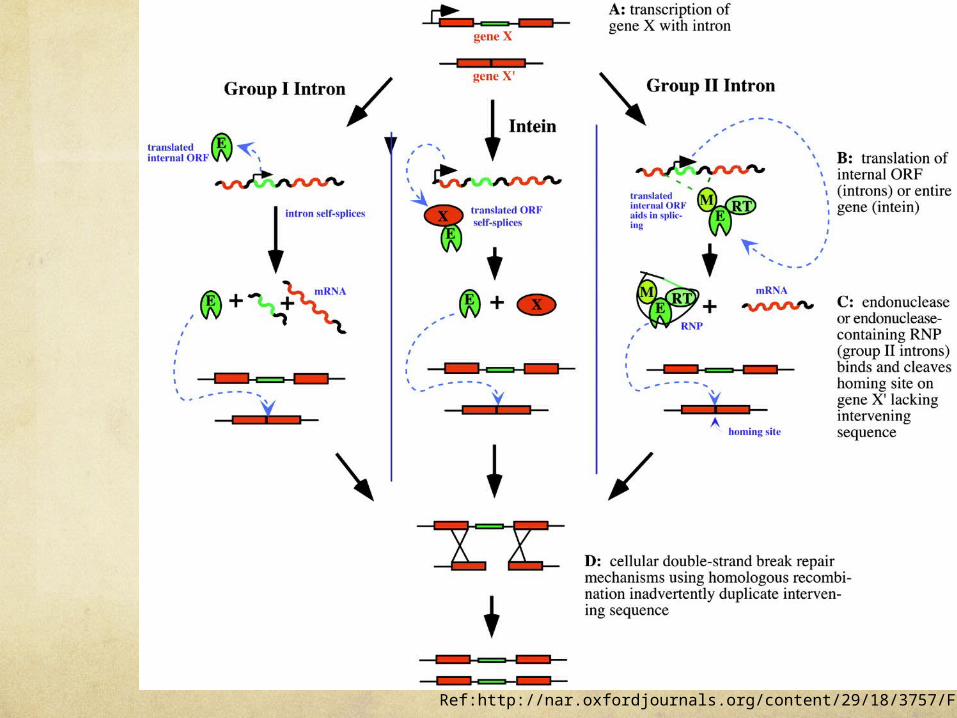
Ref:http://nar.oxfordjournals.org/content/29/18/3757/F1.large.jpg
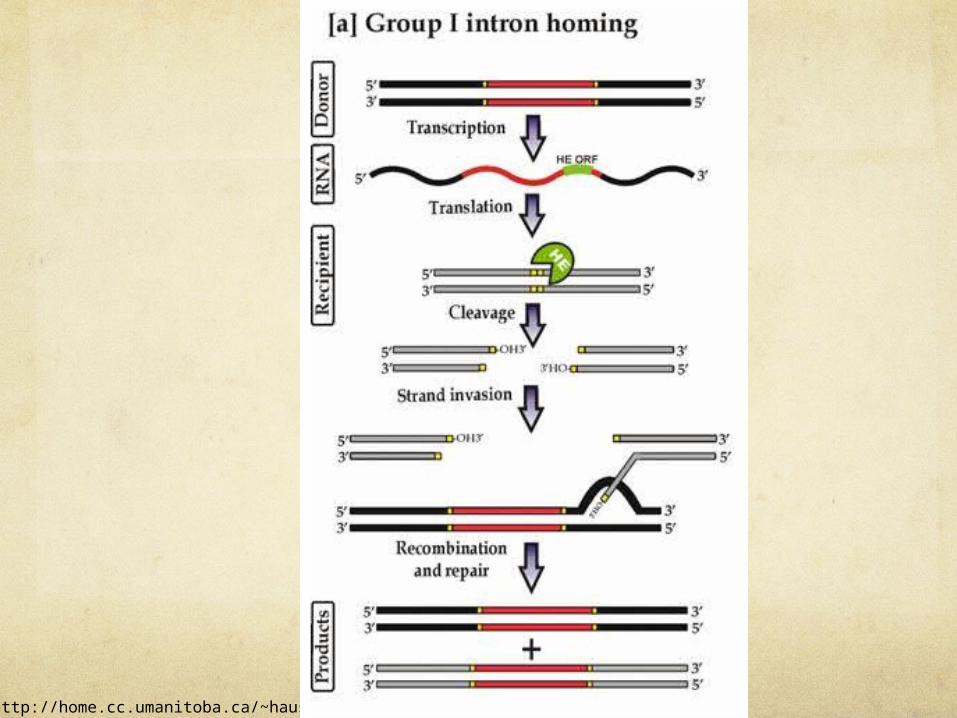
Ref: http://home.cc.umanitoba.ca/~hausnerg/
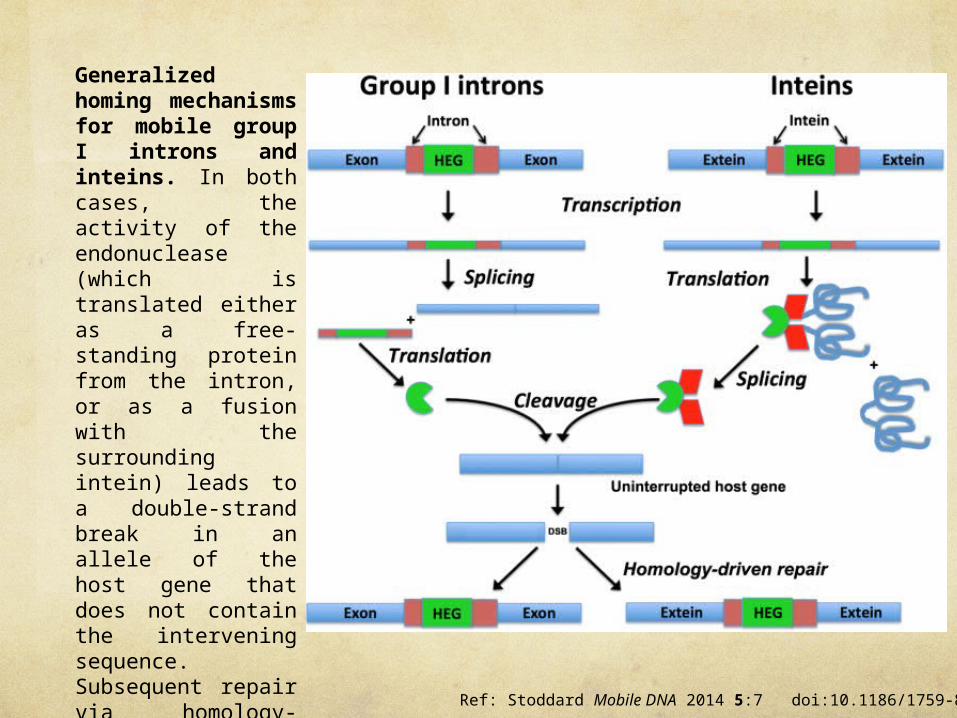
Ref: Stoddard Mobile DNA 2014 5:7 doi:10.1186/1759-8753-5-7/
Generalized homing mechanisms for mobile group I introns and inteins. In both cases, the activity of the endonuclease (which is translated either as a free-standing protein from the intron, or as a fusion with the surrounding intein) leads to a double-strand break in an allele of the host gene that does not contain the intervening sequence. Subsequent repair via homology-driven strand invasion and recombination and DNA replication, using the allele containing the intron or intein (as well as the associated endonuclease coding sequence), completes the homing process. HEG, homing endonuclease gene.
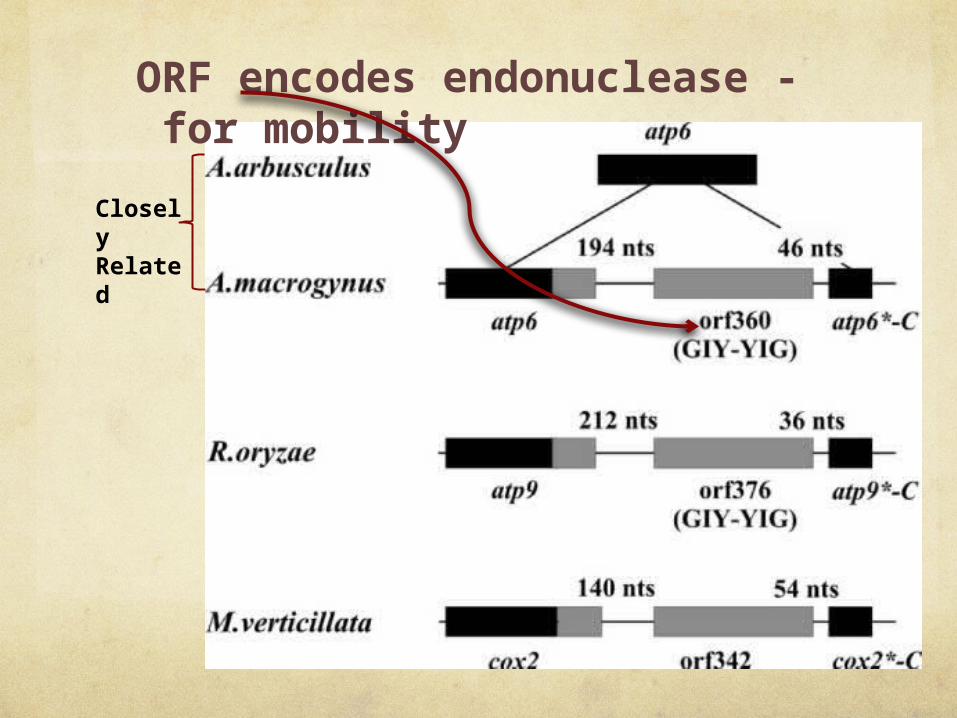
Closely Related
ORF encodes endonuclease - for mobility
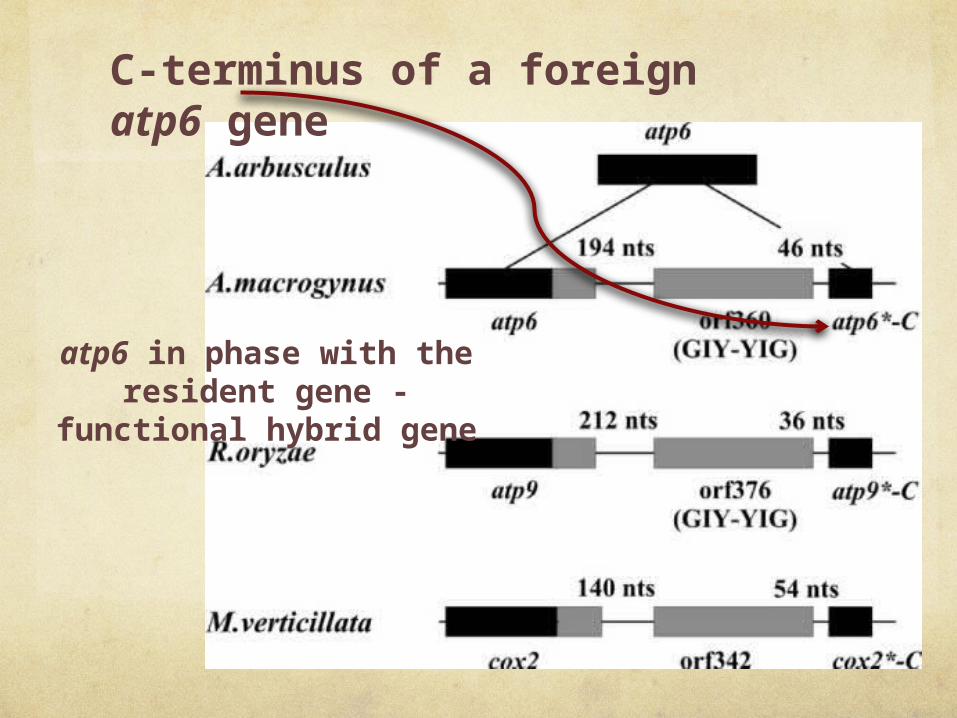
C-terminus of a foreign atp6 gene
atp6 in phase with the resident gene -
functional hybrid gene
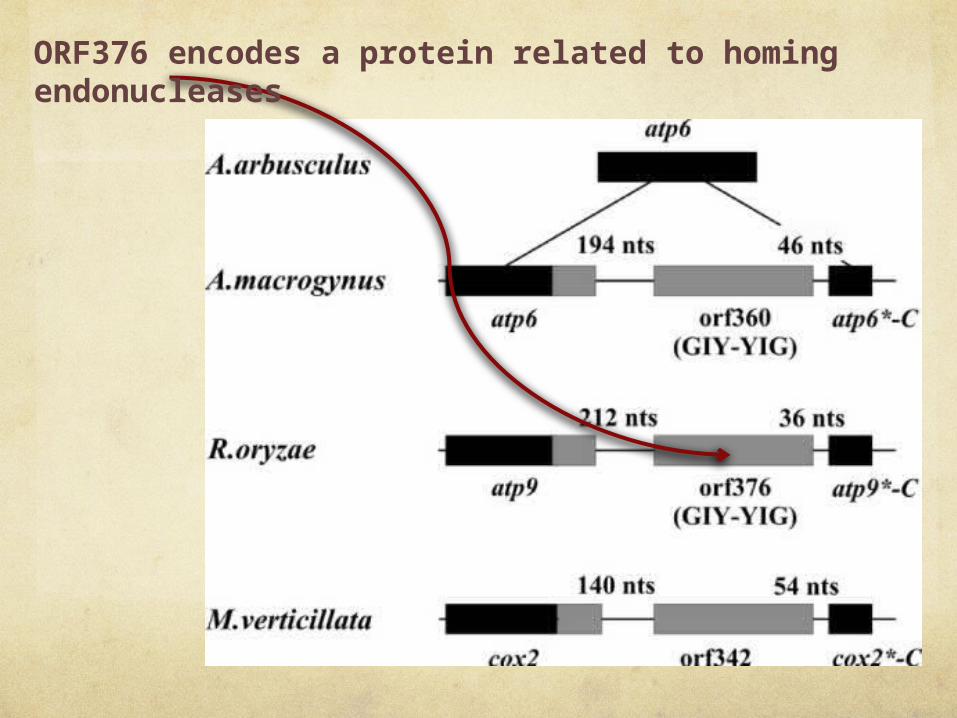
ORF376 encodes a protein related to homing endonucleases
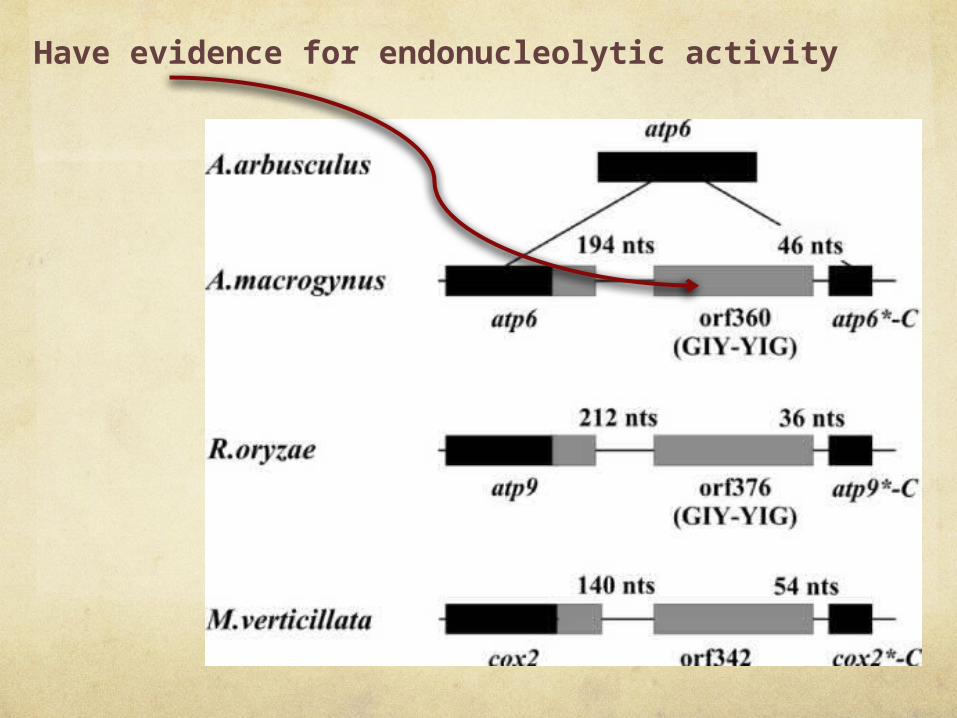
Have evidence for endonucleolytic activity

“we currently do not have biochemical evidence for the endonucleolytic activity”

With 14 introns, the mtDNA of S.culisetae contains the largest number, 9 of which are located in the cox1 gene

In R.oryzae, we identified one intron [cox1-i1(ORF305)], which is most similar to introns inserted at the same positions
of angiosperm cox1 genes (highest BLAST expect value of e−114 with Philodendron oxycardium,
Lamium sp. and Malpighia glabra).

. Because this is the only group I intron in vascular plant mtDNAs, it has most likely been acquired
by lateral transfer.
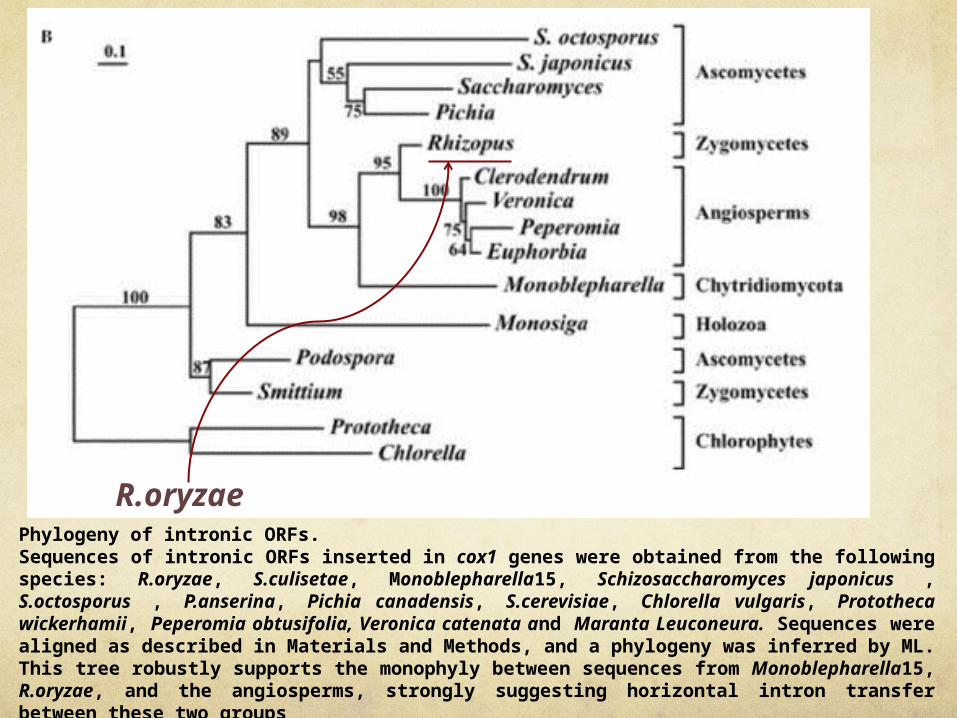
Phylogeny of intronic ORFs. Sequences of intronic ORFs inserted in cox1 genes were obtained from the following species: R.oryzae, S.culisetae, Monoblepharella15, Schizosaccharomyces japonicus , S.octosporus , P.anserina, Pichia canadensis, S.cerevisiae, Chlorella vulgaris, Prototheca wickerhamii, Peperomia obtusifolia, Veronica catenata and Maranta Leuconeura. Sequences were aligned as described in Materials and Methods, and a phylogeny was inferred by ML. This tree robustly supports the monophyly between sequences from Monoblepharella15, R.oryzae, and the angiosperms, strongly suggesting horizontal intron transfer between these two groups
R.oryzae
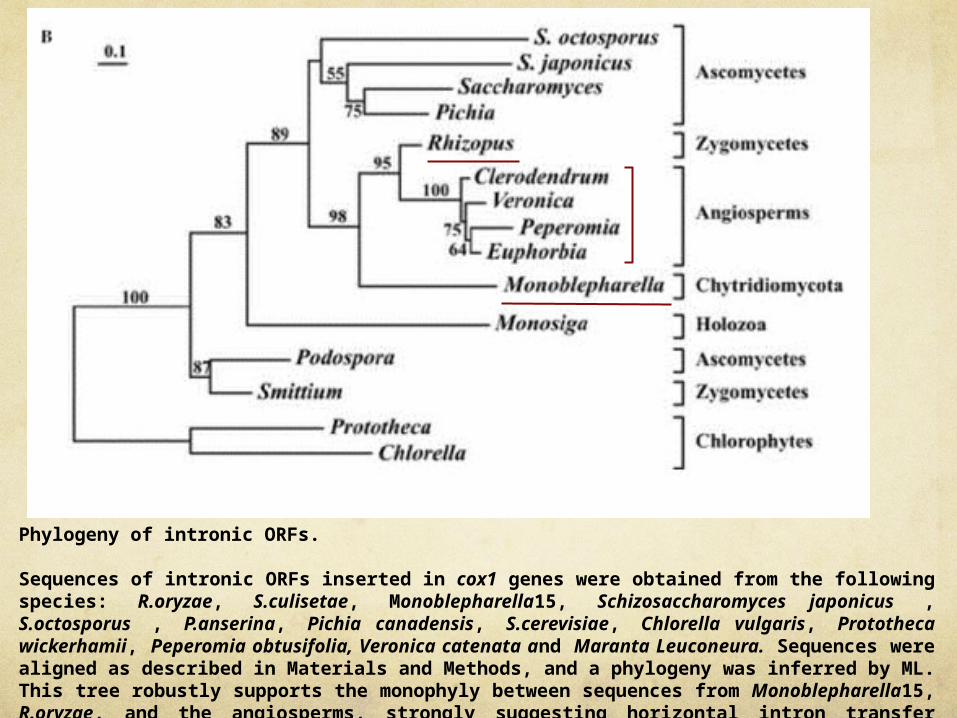
Phylogeny of intronic ORFs.
Sequences of intronic ORFs inserted in cox1 genes were obtained from the following species: R.oryzae, S.culisetae, Monoblepharella15, Schizosaccharomyces japonicus , S.octosporus , P.anserina, Pichia canadensis, S.cerevisiae, Chlorella vulgaris, Prototheca wickerhamii, Peperomia obtusifolia, Veronica catenata and Maranta Leuconeura. Sequences were aligned as described in Materials and Methods, and a phylogeny was inferred by ML. This tree robustly supports the monophyly between sequences from Monoblepharella15, R.oryzae, and the angiosperms, strongly suggesting horizontal intron transfer between these two groups
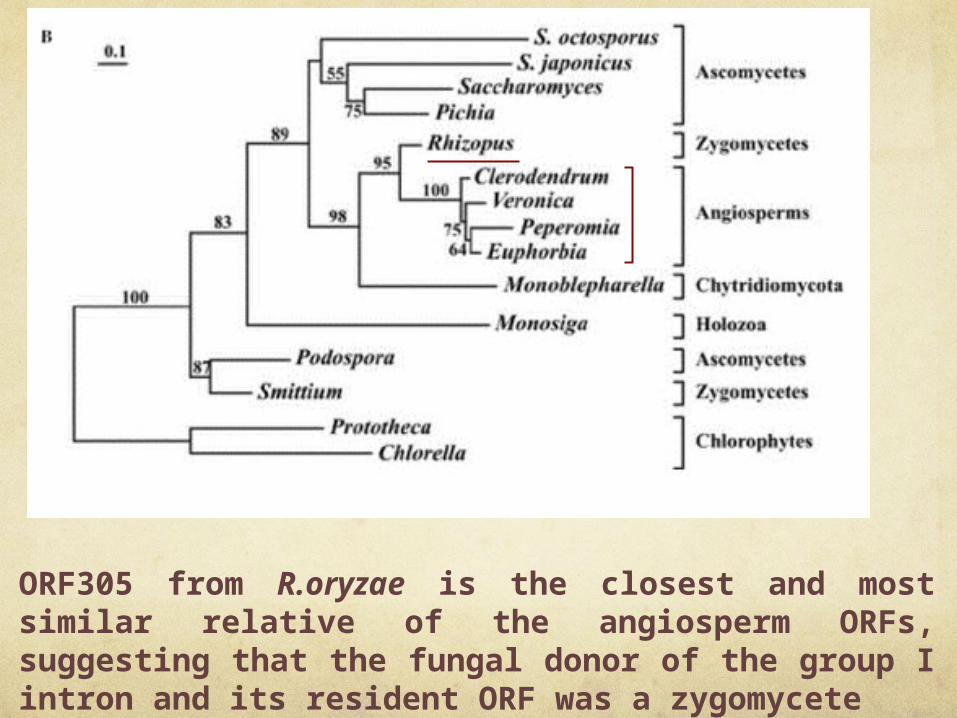
ORF305 from R.oryzae is the closest and most similar relative of the angiosperm ORFs, suggesting that the fungal donor of the group I intron and its resident ORF was a zygomycete
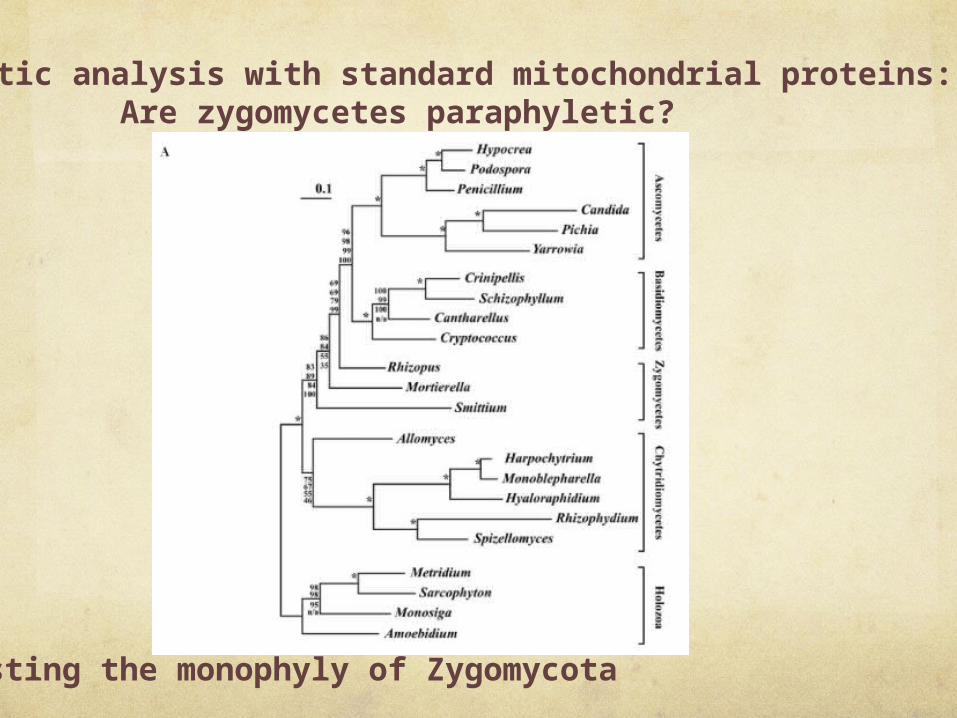
Phylogenetic analysis with standard mitochondrial proteins: Are zygomycetes paraphyletic?
Testing the monophyly of Zygomycota
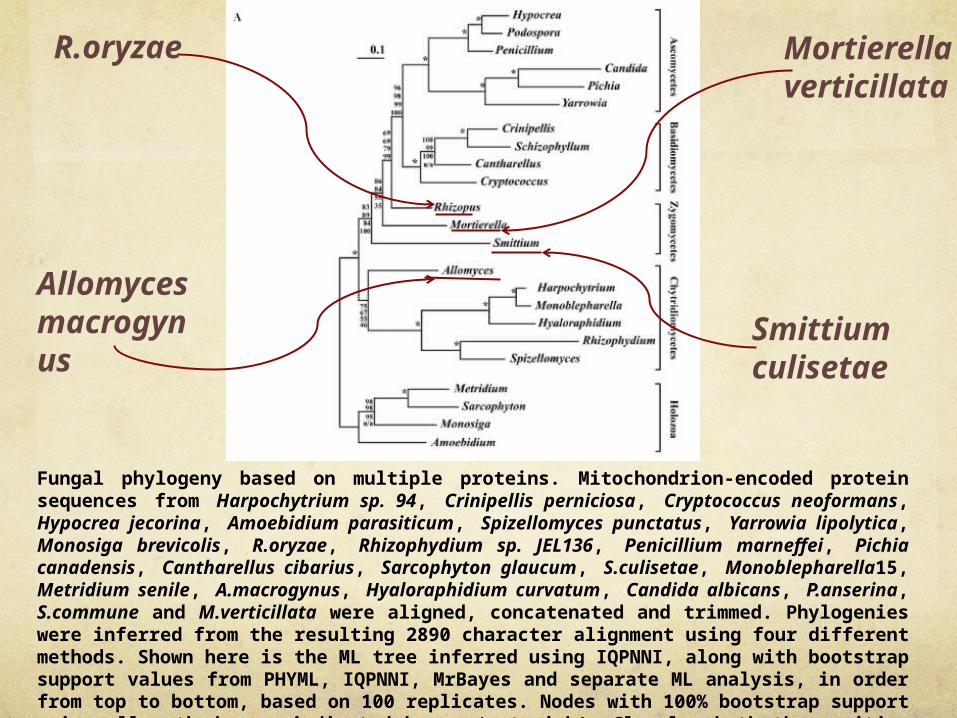
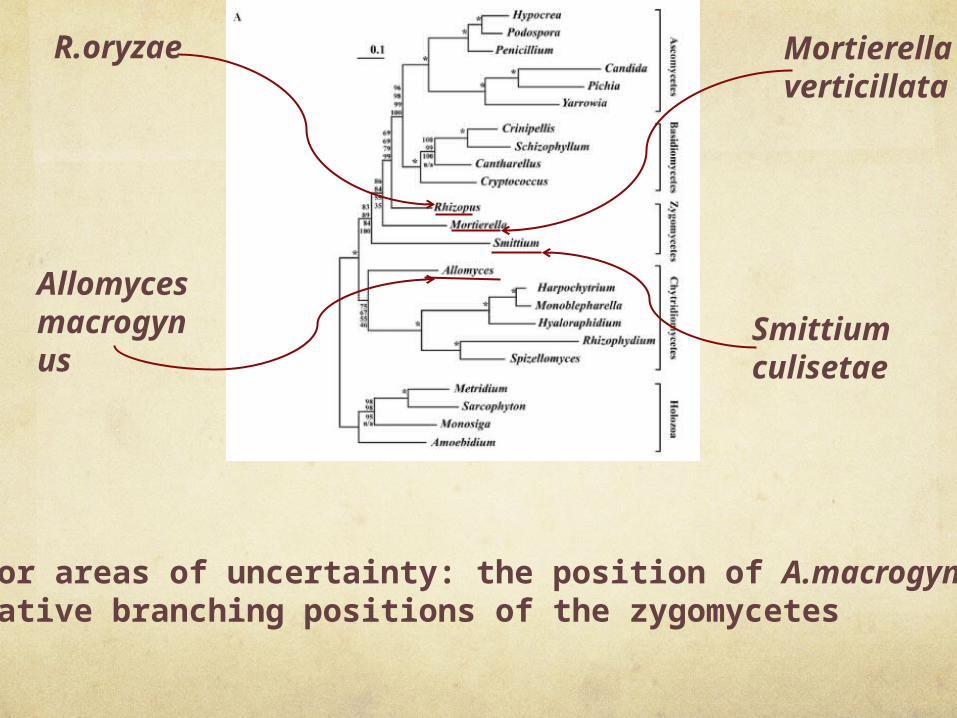
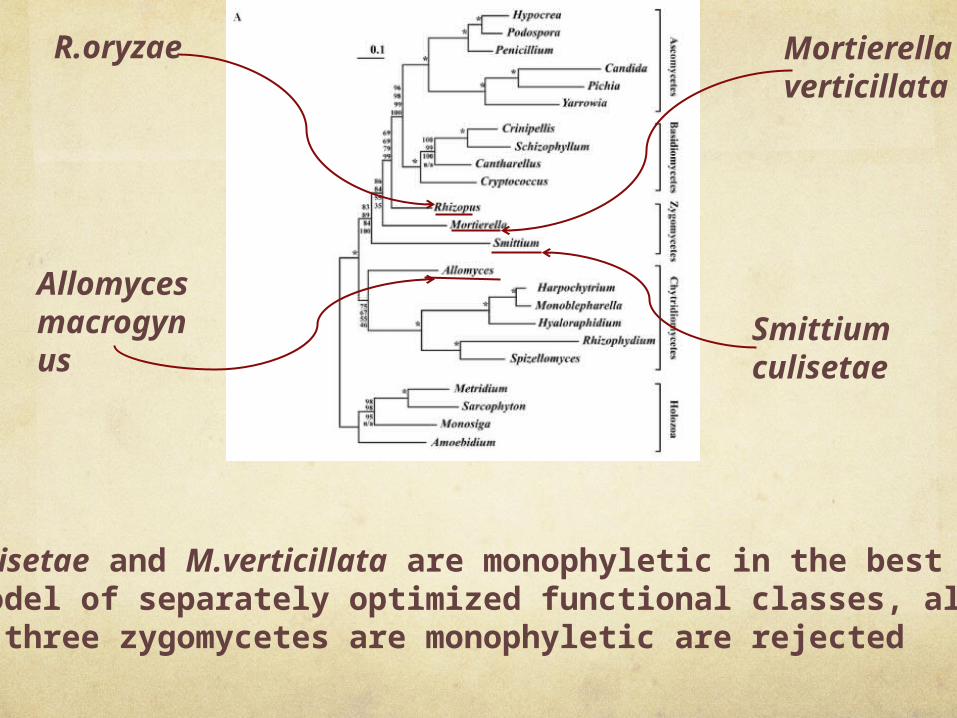
Fungal phylogeny based on multiple proteins. Mitochondrion-encoded protein sequences from Harpochytrium sp. 94, Crinipellis perniciosa, Cryptococcus neoformans, Hypocrea jecorina, Amoebidium parasiticum, Spizellomyces punctatus, Yarrowia lipolytica, Monosiga brevicolis, R.oryzae, Rhizophydium sp. JEL136, Penicillium marneffei, Pichia canadensis, Cantharellus cibarius, Sarcophyton glaucum, S.culisetae, Monoblepharella15, Metridium senile, A.macrogynus, Hyaloraphidium curvatum, Candida albicans, P.anserina, S.commune and M.verticillata were aligned, concatenated and trimmed. Phylogenies were inferred from the resulting 2890 character alignment using four different methods. Shown here is the ML tree inferred using IQPNNI, along with bootstrap support values from PHYML, IQPNNI, MrBayes and separate ML analysis, in order from top to bottom, based on 100 replicates. Nodes with 100% bootstrap support using all methods are indicated by an ‘asterisk’. Clearly, both the position of A.macrogynus and the branching order of the zygomycetes remain unclear, although the topology is robust overall.
Allomyces macrogynus
R.oryzae Mortierella verticillata
Smittium culisetae
Allomyces macrogynus
R.oryzae Mortierella verticillata
Smittium culisetae
two major areas of uncertainty: the position of A.macrogynus and the relative branching positions of the zygomycetes
Allomyces macrogynus
R.oryzae Mortierella verticillata
Smittium culisetae
Although S.culisetae and M.verticillata are monophyletic in the best tree under this model of separately optimized functional classes, all topologies in which all three zygomycetes are monophyletic are rejected

Clearly, these data are insufficient to resolve the phylogeny of the zygomycetes, most likely because these three species diverge deeply
within fungi, and at relatively short distance from each other. In such a situation, two strategies can be used to resolve the
dilemma, addition of more zygomycete and neighboring fungal lineages or addition of more sequence per species

Time to Relax…Joke
So, any questions?