complejo mayor de histocompatibilidad i y iii (1 y 3)
TRANSCRIPT

COMPLEJO MAYOR DE HISTOCOMPATIBILIDAD
C L A S E I Y I I I
Alejandra Rodríguez Estrada
PatologíaDr. Víctor Adolfo Ríos

Diferente con lo que ocurre en el caso de los receptores o anticuerpos de la
célula B, que pueden reconocer el antígeno libre, los receptores de la
célula T solo reconocen antígeno que ah sido procesado y presentado en el contexto de moléculas codificadas por
el complejo mayor de histocompatibilidad (MHC).
Complejo genético que influye en la
capacidad del organismo de
aceptar o rechazar tejido trasplantado de otro miembro
de la misma especie

Las moléculas codificadas por el MHC tienen un papel fundamental
en la determinación de las inmunoreacciones adaptativas.
El conjunto específico de moléculas MHC expresadas por un
individuo influye en el repertorio de antígenos a las cuales las células TH y TC de ese individuo
pueden reaccionar.
Influye en l reacción de un individuo a antigenos de microorganismos infecciosos y por tanto se le ah implicado en la susceptibilidad a enfermedades así como en el desarrollo de autoinmunidad,

Locus genético (brazo corto del cromosoma 6) que codifica para los antígenos de histocompatibilidad
CLASIFICACIÓN
CLASE I
Cadena ligera
Cadena pesada
-2-microglobulina
Glucoproteína transmembran
a
Todas las células excepto los hematíes
Codificados por el locus: HLA-A, -B y -C
CLASE II
Macrófagos y linfocitos B y T estimulados por
antígenos
Control de la respuesta
inmunitaria
Codificado por el locus HLA-D
Codifica proteínas para
procesar antígenos
a1: abierto
a2: plegado
b1 y b2 son
plegadas
Cadena ligera (b)
Cadena pesada (a)
CLASE III
Codifica proteínas del complemento y
citocinas
LPTAPcomo
Moléculas del MHC
Variabilidad genética
Poligénico
Polimórfico
Varios genes para cada clase de molécula
Genes se heredan como
haplotipo
Bloque de genes , poco
entrecruzamiento
Estructural-mente diferente
s
•Marcadores de "lo propio" (el yo inmunológico).•Intervienen en la educación tímica de los linfocitos T •Presentan los péptidos antigénicos para la activación de los linfocitos T
Subpoblación
citotóxica
Regulador, productor de
toxinas
Linfocitos T CD4
Linfocitos T CD8
Reconocidos por
Reconocido por
De la
Se encuentra en Se encuentra en
Función
Posee
es

Desempeñan funciones importantes en el reconocimiento intercelular y la
diferenciación entre lo propio y lo extraño.
Participa en el desarrollo tanto de las inmunoreacciones y la diferenciación entre lo propio y lo extraño.

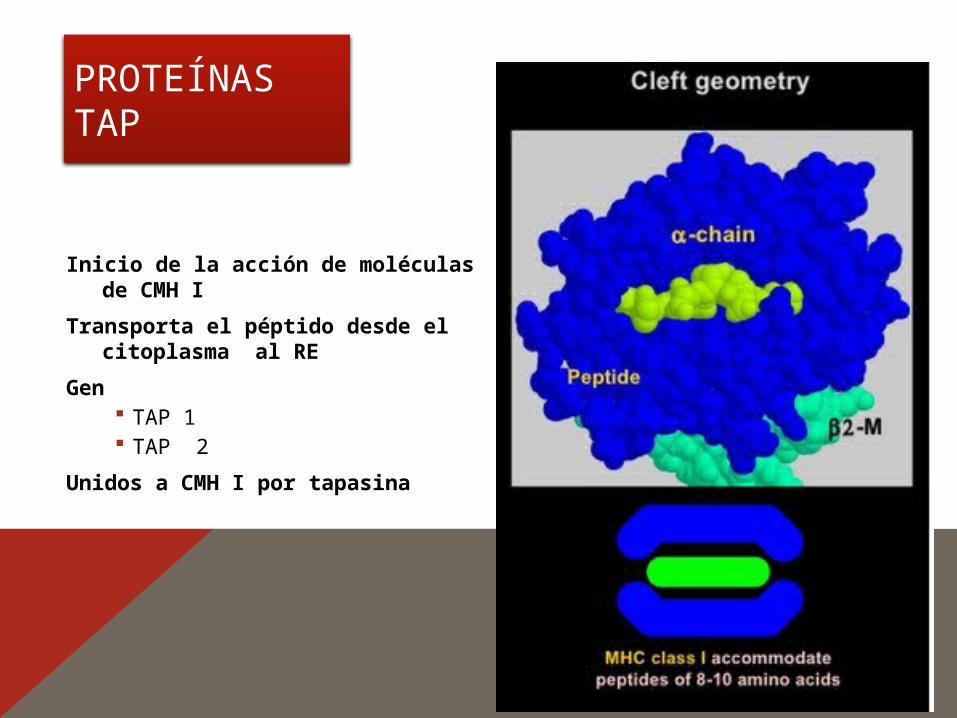
PROTEÍNAS TAP
Inicio de la acción de moléculas de CMH I
Transporta el péptido desde el citoplasma al RE
Gen TAP 1 TAP 2
Unidos a CMH I por tapasina

COMPLEJO MAYOR DEHISTOCOMPATIBILIDAD (MHC)
RECHAZO DE TRANSPLANTES DE TEJIDOS EN RATONES
ANTIGENOLEUCOCITARIOHUMANO (HLA)
Familia de las Ig
Son moléculas que se localizan en la superficie de las células
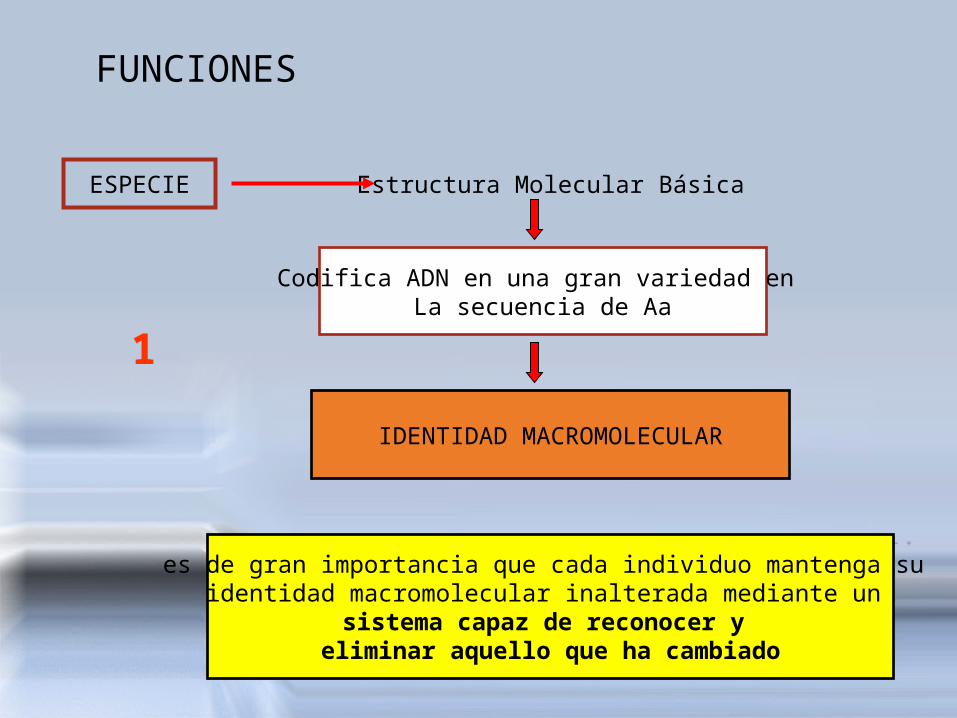
FUNCIONES
ESPECIE Estructura Molecular Básica
Codifica ADN en una gran variedad en La secuencia de Aa
IDENTIDAD MACROMOLECULAR
es de gran importancia que cada individuo mantenga su identidad macromolecular inalterada mediante un
sistema capaz de reconocer y eliminar aquello que ha cambiado
1

Célula normal
LINFOCITO T
No ocurre ataque celular
Molécula MHC clase I
CMH clase I(reconocimiento)
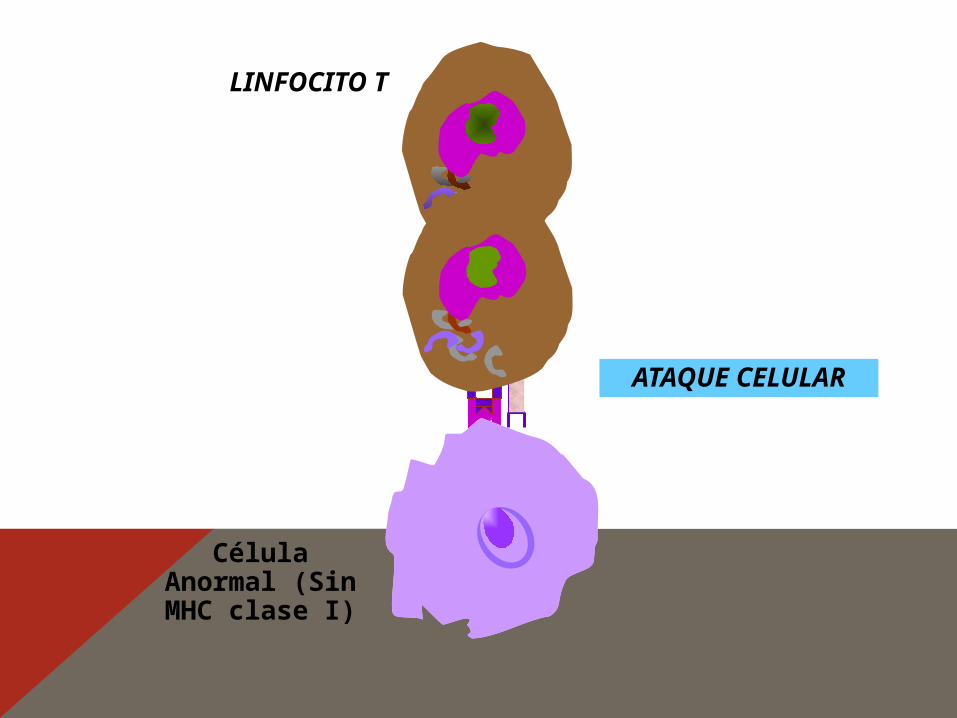
Célula Anormal (Sin MHC clase I)
LINFOCITO T
ATAQUE CELULAR

EL MHC CODIFICA TRES CLASES DE MOLÉCULAS PRINCIPALES
MHC es un conjunto de genes dispuestos dentro de una tira continua de DNA en el cromosoma 6 en
seres humanos. Este se denomina complejo HLA en el ser humano.
Genes MHC
clase I
• Codifican glicoproteínas que se expresan en la superficie de casi todas las células nucleadas. Su principal función es la presentación de antígenos péptidos a células Tc
Genes MHC
clase II
• Codifican glicoproteínas que se expresan sobre todo en células presentadoras de antígeno done presentan péptidos antigénicos procesados a células TH
Genes MHC
clase III
• Codifican, además de otros productos, varias proteínas secretadas que desempeñan funciones inmunitarias, inclusive componentes del sistema de complemento y moléculas relacionadas con la inflamación

Las moléculas MHC clase I fueron las
primeras en descubrirse.
Genes clase I no clásicos
Moléculas clase
uno clásicas
La expresión de los productos génicos no
clásicos se limita a ciertos tipos específicos
de células.

Las moléculas MHC clase I y clase II comparten características estructurales y ambas participan en el procesamiento y la presentación del antígeno. Al contrario las moléculas de clase III a cuyos lados se encuentran las regiones clase I y clase II codifica moléculas críticas para la función inmunitaria pero tienen poco en común como las moléculas clase I o II.

Las moléculas clase I tienen una cadena
pesada de glucoproteína y una cadena ligera proteínica pequeña

LAS FORMAS ALÉLICAS DE LOS GENES DE MHC SE HEREDAN EN GRUPOS UNIDOS LLAMADOS HAPLOTIPOS
Los locus que constituyen el MHC son altamente polimórficos entre las población existen muchas formas alternativas del gen, o alelos, en cada locus. Los genes de los locus de MHC se encuentran muy cerca entre sí.

LAS FORMAS ALÉLICAS DE LOS GENES DE MHC SE HEREDAN EN GRUPOS UNIDOS LLAMADOS HAPLOTIPOS
Halotipo se denomina a cada grupo de alelos. Los alelos se expresan de manera codominante en la misma se expresan productos génicos maternos o paternos.

LAS CEPAS DE RATÓN CONGÉNICO MHC SON IDÉNTICAS EN TODOS LOS LOCUS EXCEPTO EL DEL MHC.
. Las cepas de ratones endogámicas son singénicas idénticas en todos los locus genéticos 2 cepas son congénitas no son genéticamente idénticas excepto en locus o región genética aislada.

MOLÉCULAS Y GENES DEL MHC.

-LAS MOLÉCULAS DE MHC CLASE I Y II.
Son glucoproteinas unidas a la membrana.
Tiene relación en su estructura y función
Se aíslan y purifican
Se determina su estructura tridimensional extracelular con cristalografía de rayos x.
Funcionan como moléculas presentadoras de antígenos
Forman complejos estables con péptidos antigénico (están en la superficie celular para ser reconocida por células T.

-MOLÉCULAS DE MHC CLASE III
Son un grupo de proteínas no relacionadas.
Son diferentes en la estructura y función a las moléculas de clase I y II.

LAS MOLÉCULAS CLASE I TIENEN UNA CADENA PESADA DE GLUCOPROTEÍNA Y UNA CADENA LIGERA PROTEÍNICA PEQUEÑA
*Las moléculas de MHC clase I:
-tienen cadena alfa de 45 kDa, vinculada de forma no covalente con una molécula de micro-globulina beta2 de 12 kDa.
*La cadena alfa:
-Es una glucoproteína transmembrana codificada por genes polimórficos dentro de la región A, B y C del complejo HLA humano y dentro de las regiones K y D/L del complejo H-2 de ratón.
*micro-globulina beta2:
Es una proteína codificada por un gen conservado localizados en cromosomas diferentes.

LAS MOLÉCULAS CLASE I TIENEN UNA CADENA PESADA DE GLUCOPROTEÍNA Y UNA CADENA LIGERA PROTEÍNICA PEQUEÑA
La expresión de la molécula MHC clase I en membrana celular: requiere de vinculación de la cadena alfa con la micro-globulina beta2.
Cadena alfa:
-se fija a la membrana plasmática por su segmento transmembrana hidrófobo y cola citoplasmática hidrófila.
- la cadena alfa de molécula de MHC clase I esta organizada entre dominios externos: alfa(1,2,3):
1) cada uno de estos dominios tiene 90 aminoácidos.
2)el segmento citoplasmático de fijación 30 aminoácidos.

LAS MOLÉCULAS CLASE I TIENEN UNA CADENA PESADA DE GLUCOPROTEÍNA Y UNA CADENA LIGERA PROTEÍNICA PEQUEÑA
Micro-globulina beta2:
-Tienen tamaño y organización parecido al dominio alfa-3
-No contiene una región transmembrana.
- No se une de manera no covalente con la glucoproteína clase I.
*Enzima papaina:
-segmenta la cadena alfa.
-libera la posición extracelular de la molécula [alfa(1, 2, 3) y micro-globulina].

LAS MOLÉCULAS CLASE I TIENEN UNA CADENA PESADA DE GLUCOPROTEÍNA Y UNA CADENA LIGERA PROTEÍNICA PEQUEÑA
*Porción extracelular: -interactúan 2 pares de dominios:
1)Un par de la membrana (alfa 1 y alfa 2)
2)Un par proximal del a membrana (alfa 3 y micro-globulina)
*Dominio alfa 1 y 2.
1)Interactúan para formar una hileras beta antiparalelas con regiones helicoidales alfa larga
2)La estructura forma una hendidura
3)La hendidura de unión de péptido se localiza en la superficie alta de la moléculas MHC clase I y una péptidos de 8 a 10 aminoácidos.

LAS MOLÉCULAS CLASE I TIENEN UNA CADENA PESADA DE GLUCOPROTEÍNA Y UNA CADENA LIGERA PROTEÍNICA PEQUEÑA
* Dominio alfa 3 y micro-globulina beta2:
1) Están organizadas en 2 hojas beta plegadas, formadas por hileras beta antiparalelas de aminoácidos.
2)Posee una estructura similar al de las inmunoglobulinas.
MHC clase I y micro-globulinas son miembros de la superfamilia de inmunoglobulinas.
*Dominio alfa3:
-Están conservados entre las moléculas MHC clase I.
-Contienen una secuencia que interactúa con la molécula de la membrana CD8 de las células Tc.

LAS MOLÉCULAS CLASE I TIENEN UNA CADENA PESADA DE GLUCOPROTEÍNA Y UNA CADENA LIGERA PROTEÍNICA PEQUEÑA.
*Micro-globulina beta2:
-interactúan con el dominio alfa 3 y con aminoácidos de los dominio alfa 1 y 2.
-la interacción de micro-globulina beta y un péptido con una cadena alfa clase I provoca la conformación de moledlas MHC clase I para luego trasportase a la superficie de la célula.
* Células tumorales de Daudi: son incapaces de sintetizar micro-globulinas beta2, por lo tanto producen cadenas alfa de MHC clase I que no se expresan en la superficie de la célula.

LAS MOLÉCULAS CLASE II TIENE CADENAS GLUCOPROTEÍNICAS DISTINTAS
Diferencias entre:
Moléculas clase I Moléculas clase II
Compuesto por un piso de 8 hileras beta antiparalelas y 2 hileras alfa antiparalelas
Carecen de residuo conservados que se unan a residuos terminales de péptidos cortos y forman bolsas abiertas
Presenta mas de una cuenca
Presenta un surco de extremos abiertos

ESQUEMA DE LAS MOLÉCULAS DE MHC CLASE I Y CLASE II QUE MUESTRAN LOS DOMINIOS EXTERNOS, SEGMENTO TRANSMEMBRANA Y LA COLA CITOPLASMÁTICA

LAS DISPOSICIÓN DE AXÓN / INTRÓN DE GENES CLASE I Y II REFLEJA SU ESTRUCTURA DE DOMINIO
Exones separados codifican cada región de las proteínas clase I y II.
a) c/u de los genes clase I ( ratón y hombre) tienen:
-exón líder 5’ codifica un péptido de señal corto.
-5 o 6 Exones codifican los dominios entre células
( alfa 1, 2 y 3)
-Un exón corriente abajo, codifica región transmembrana (Tm)
-1 o 2 Exones de la Terminal 3’codifican los dominios citoplásmicos (c).

LAS DISPOSICIÓN DE AXÓN / INTRÓN DE GENES CLASE I Y II REFLEJA SU ESTRUCTURA DE DOMINIO
B) Genes de MHC clase II ( parecida a clase I)
-Organizadas en exones e intrones que reflejan la estructura de dominio de cadena alfa y beta.
-genes alfa y beta:
1) Codifican moléculas de MCH I.
2) Tienen un exón lider.
3) Tienen exón alfa 1 o beta 1.
4) Tienen exón alfa 2 o beta 3.
5) Tienen exón transmembrana.
6) Tienen exón citoplásmico.

ESQUEMA DE LOS GENES MHC CLASE I (A) Y CLASE II (B)

LAS MOLÉCULAS CLASE I Y II MUESTRAN POLIMORFISMO EN LA REGIÓN QUE SE UNE PÉPTIDOS
Se identifican cientos de variantes de moléculas de MHC clase I y II en el hombre.
Aunque solo se expresa cifras pequeñas de moléculas MHC: 6 moléculas deferentes clase I y 12 moléculas clase II distintas.
Estas cifras deben ser capaz de mostrar una disposición a diferentes péptidos antihigiénicos a células T. y permitir al sistema inmunitario responder de manera especifica a muchos retos de anfígenos diferentes .
“Promiscua” es la unión entre péptido y una molécula que suele tener especificidad amplia.
a) Molécula MHC puede unirse a muchos péptidos distintos
b) Algunos péptidos pueden unirse a varias moléculas de MHC.
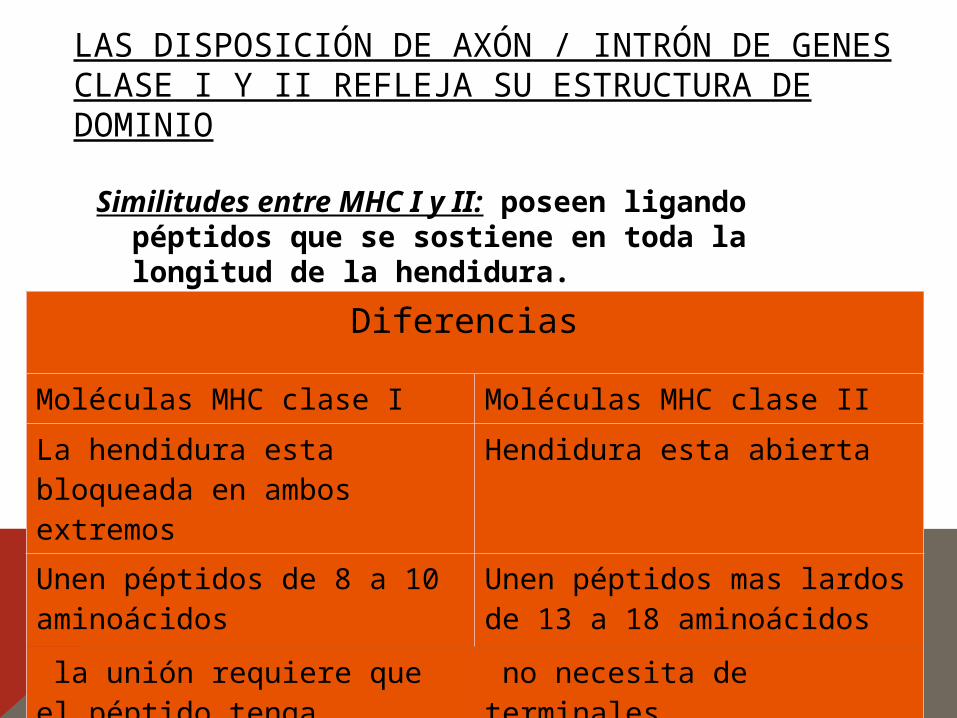
LAS DISPOSICIÓN DE AXÓN / INTRÓN DE GENES CLASE I Y II REFLEJA SU ESTRUCTURA DE DOMINIO
Similitudes entre MHC I y II: poseen ligando péptidos que se sostiene en toda la longitud de la hendidura.
Diferencias
Moléculas MHC clase I Moléculas MHC clase II
La hendidura esta bloqueada en ambos extremos
Hendidura esta abierta
Unen péptidos de 8 a 10 aminoácidos
Unen péptidos mas lardos de 13 a 18 aminoácidos
la unión requiere que el péptido tenga residuos aminoácidos
no necesita de terminales

*INTERACCIÓN MHC CLASE I-PÉPTIDO
*Péptidos:
-se unen a MHC clase I y son presentados a la células TCD8+.
-Se derivan de proteínas intracelulares endógenas.
- Se digieren en el citosol.
-Se transportan del citosol a las cisternas del RE.
- La cisterna de RE interactúan con MHC clase I.
Este es el proceso llamado Vía cistosólica o endógena de procesamiento.

*INTERACCIÓN MHC CLASE I-PÉPTIDO
Con relación al MHC clase I:
- Cada tipo de molécula ( k, D, L, ratones o A, B, C en humanos) se une a un grupo único de péptidos.
- Cada variante alélica de una molécula clase I también se une a un grupo distinto de péptidos.
Porqué sucede esto?
Porque una sola célula nucleada expresa muchas copias ( 10*5) de cada molécula clase I
Las moléculas de MHC clase I expresan muchos péptidos deferentes ( 2.000) en la superficie de una célula nucleada.

MAPA GEONÓMICO DETALLADO DE LOS GENES DEL MHC
MHC abarca:-2.000KB del ADN de ratones.-4.000kb del ADN en hombres.
*Las region clase I humana abarca 2.000kb en l extreo telomerico del complejo de antigenos de histocaompatibilidad leucocitaria ( HLA)
Incluyen cerca de 20 genes. En la región clase I se halan codificando las
moléculas MHC clase I clásica.

*INTERACCIÓN MHC CLASE I-PÉPTIDO
*Se necesita por lo menos 100 complejos péptido-MHC para que una célula sea el blanco ara reconocimiento y lisis por linfocito Tc.
Péptidos nonamericos: se unen a moléculas clase I con una afinidad 100 a 1.000 veces mayor que los péptidos que son mas largos o mas cortos. Son los que mas se unen a la hendidura.
Residuos de fijación: son residuos de aminoácidos que tienen los péptidos y que se presentan en el surco de la molécula MHC clase I . Esta le confiere la capaceada de unión a los péptidos.

INTERACCIÓN MHC CLASE I-PÉPTIDO
Todos los péptidos que se unen con moléculas clase I tienen:
-Fijadores con Terminal carboxilo.
-fijadores con Terminal amino.
Los principales puntos de contacto entre la molécula MHC clase I y péptidos:
-residuo 2 en el extremo Terminal amino.
-residuo 9 en el Terminal carboxilo de nonaméricos.

LAS MOLÉCULAS CLASE I Y CLASE II MUESTRAN DIVERSIDAD DENTRO DE UNA ESPECIE Y SE PRESENTAN MÚLTIPLES FORMAS EN UN INDIVIDUO
El origen de la diversidad para las moléculas de MHC no es el mismo.
La generación de receptores de células T y B cambian con el tiempo.
Las moléculas MHC clase I no cambian están fijas.
Los genes clase II humanos son polimórficos.
La diversidad del MHC dentro de una especie: proviene, del polimorfismo, la presencia de múltiples alelos en un locus genético dentro de la especie.
MHC es poligénico: cuando incluye genes con estructura y función similares pero no idénticas.

*DESEQUILIBRIO DE ENLACE
Se da porque ciertas combinaciones alélicas ocurren con mayor frecuencia en haplotipos HLA que por la combinación aleatoria lo que conduce a una diversidad real menor.
*Importancia funcional de los polimorfismos del MHC.
La localización de estos aminoácidos polimórficos dentro del sitio de unión para antígeno procesado sugiere que las diferencias alélicas contribuyen a las variaciones que se observan en la capacidad de las moléculas de MHC para interactuar con péptido antígeno determinado.

*LAS REGION CLASE I HUMANA ABARCA 2.000KB EN L EXTREO TELOMERICO DEL COMPLEJO DE ANTIGENOS DE HISTOCAOMPATIBILIDAD LEUCOCITARIA ( HLA)
Moleculas MHC clase I clásica: HLA-A; HLA-B; HLA-C.
Los genes clase I no clásica:
1)HLA-E; HLA-F; HLA-G; HLFE; HLA-J y HLA-X.
2)MIC (genes) [MICA Y MICE].
3) Seudogenes que no codifican productos proteicos.
4) Las MICA son muy polimórficos.
Las moléculas MHC clase I clásicas presentan péptidos a células T.