conference review creating porcine biomedical models through
TRANSCRIPT

Comparative and Functional GenomicsComp Funct Genom 2004; 5: 262–267.Published online in Wiley InterScience (www.interscience.wiley.com). DOI: 10.1002/cfg.404
Conference Review
Creating porcine biomedical models throughrecombineering
Margarita M. Rogatcheva1, Laurie A. Rund1, Kelly S. Swanson1, Brandy M. Marron1, Jonathan E. Beever1,Christopher M. Counter3 and Lawrence B. Schook1,2*1Department of Animal Sciences, University of Illinois, Urbana, IL 61801, USA2Department of Veterinary Pathobiology, University of Illinois, Urbana, IL 61801, USA3Department of Pharmacology and Oncology, Duke University Medical Center, Durham, NC, USA
*Correspondence to:Lawrence B. Schook, Departmentof Animal Science and VeterinaryBiology, University of Illinois, 329Edward R. Madigan Laboratory,1201 W. Gregory Dr., Urbana, IL61801, USA.E-mail: [email protected]
Received: 16 February 2004Accepted: 17 February 2004
AbstractRecent advances in genomics provide genetic information from humans and othermammals (mouse, rat, dog and primates) traditionally used as models as wellas new candidates (pigs and cattle). In addition, linked enabling technologies,such as transgenesis and animal cloning, provide innovative ways to design andperform experiments to dissect complex biological systems. Exploitation of genomicinformation overcomes the traditional need to choose naturally occurring models.Thus, investigators can utilize emerging genomic knowledge and tools to createrelevant animal models. This approach is referred to as reverse genetics. In contrastto ‘forward genetics’, in which gene(s) responsible for a particular phenotypeare identified by positional cloning (phenotype to genotype), the ‘reverse genetics’approach determines the function of a gene and predicts the phenotype of acell, tissue, or organism (genotype to phenotype). The convergence of classicaland reverse genetics, along with genomics, provides a working definition of a‘genetic model’ organism (3). The recent construction of phenotypic maps definingquantitative trait loci (QTL) in various domesticated species provides insights intohow allelic variations contribute to phenotypic diversity. Targeted chromosomalregions are characterized by the construction of bacterial artificial chromosome(BAC) contigs to isolate and characterize genes contributing towards phenotypicvariation. Recombineering provides a powerful methodology to harvest geneticinformation responsible for phenotype. Linking recombineering with gene-targetedhomologous recombination, coupled with nuclear transfer (NT) technology canprovide ‘clones’ of genetically modified animals. Copyright 2004 John Wiley &Sons, Ltd.
Keywords: genomics; animal models; porcine; recombineering; BAC
Introduction
Comparative genomics provides an important cri-terion for creating animal models relevant to dis-secting human diseases. Resolving complex humandiseases is difficult (e.g. time course of dis-ease onset in animals, expenses associated withhuman clinical experiments, ethical issues), andthus appropriate biomedical models must be devel-oped and validated. Biomedical models are defined
as ‘surrogates for a human being, or a human bio-logic system, that can be used to understand nor-mal and abnormal function from gene to phenotypeand to provide a basis for preventive or therapeu-tic intervention in human diseases’ [10,18]. In thepast, researchers have used two main approachesto study human diseases. In one strategy, a humanclinical disease is fully characterized and then themost appropriate animal model is chosen, basedon criteria such as anatomical and/or physiological
Copyright 2004 John Wiley & Sons, Ltd.

Creating porcine biomedical models through recombineering 263
characteristics (biological relevance), cost and ani-mal husbandry required. Another tactic has beento characterize naturally occurring or induced (bychemical or radiation exposure) mutant animals(most commonly the rat or mouse) and identifywhich human disease they resemble.
Integrating gene discovery and functional analy-sis is key to harvesting the ‘genomic promise’. Val-idation of identified genes associated with QTL isessential. Because traditional validation approachesare time consuming, expensive and complicatedby the diverse genetic backgrounds of non-inbredbreeding stock, new methods of directly testing thecausal effect of a given gene for a given phenotypeare required. Because complex traits are polygenic(i.e. controlled by more than one genetic compo-nent), the ability to use a somatic cell target priorto NT to produce multiple gene substitutions into asingle genetic background is essential for validat-ing genetic interactions. Finally, by using somaticcell genomics, an ex vivo platform for expressionprofiling prior to NT, the number and associatedcosts of transgenic animals may be reduced.
Over the past decade, tremendous progress hasbeen made with regard to the mapping and char-acterization of the swine genome. Moderate- tohigh-resolution genetic linkage maps containinghighly polymorphic loci (Type II) have beenproduced using independent mapping populations[11,15]. Additionally, physical mapping methods,such as somatic cell hybrid analysis [12,19],in situ hybridization and ZOO-FISH [1] have beenemployed to enrich the Type I marker map andto perform comparative analysis with map-richspecies such as the human and mouse. To date,over 5000 mapped loci are catalogued for thepig genome (http://www.thearkdb.org). Swinewhole-genome radiation hybrid (WG-RH) mapsalso have been generated [6], resulting in yetanother rapid increase in the number of locimapped. Even more recently, the swine genomicscommunity has acquired access to resources suchas BAC libraries [5] to facilitate the productionof high-resolution physical maps in specific chro-mosomal regions [13,14], and the construction ofsequence-ready mapping resources for the porcinegenome.
In current model systems (fruit fly, yeast, round-worm and mouse), functional genomics is sup-ported by the ability to develop congenic inbred
lines, cloning and creating mutants by either dele-tion or substitution of specific genes [17]. In pigsand cattle, there are neither inbred lines nor embry-onic stem cell (ES) lines to create gain-of-function(GOF, knock-in) or loss-of-function (LOF, knock-out) lines. In addition, the relatively long gestationtime and costs of creating large breeding herds tomap polygenic traits devoid of various backgroundgenes is not cost-effective with respect to validatingQTL through breeding. Thus, there is an importantneed to develop in vitro correlates for transcrip-tion profiling (functional genomics and proteomics)similar to that developed for the worm, fly andmouse. This discovery platform must also allowcapture of sequence information from compara-tive genomes (creating models to validate hypothe-ses with respect to gene–gene interactions asso-ciated with multigenic traits). It is also importantthat such an experimental system should allow theintrogression of ‘alleles’ into germplasm, overcom-ing the current limitation of complicated targetingand deletion constructs and manipulation involvedin constructing large pieces of DNA for homol-ogous recombination (HR). Such a model for anintegrated approach to build, test and refine cellu-lar pathways resulting from specific perturbations(analysed by DNA microarrays) has recently beendeveloped using Saccharomyces cerevisiae [17].
‘Recombineering’, or ‘chromosomal engineer-ing’, permits directed genetic modification ofgenomic DNA and links gene discovery of a givenphenotype with functional analysis (directly test-ing cause and effect) [2]. Recombineering providesa rapid method to genetically manipulate largeDNA inserts cloned into BACs [16]. BACs haveproved useful for cloning and maintenance of largeDNA fragments in a recA− genetic backgroundthat prevents genomic rearrangement. However,such host backgrounds also prevent the manipula-tion of insert DNA using conventional homologousrecombination techniques [9]. Recently, a numberof approaches involving modification of the hostbacterium have been developed to permit BACmanipulation. These have included inducible pro-moters to permit transient expression of bacterialrecE and recT genes or other analogous bacte-riophage lambda (λ) genes (exo and beta) [20].Recently an Escherichia coli strain harbouring adefective λ-prophage has been developed that pro-motes high BAC recombination frequencies.
Copyright 2004 John Wiley & Sons, Ltd. Comp Funct Genom 2004; 5: 262–267.

264 M. M. Rogatcheva et al.
In order to make this system suitable for BACmanipulation, the E. coli strain DY380 was gener-ated by introducing the λ prophage into the BAChost strain DH10B [16]. The λ prophage provides arapid single-step method to generate subtle changesin any gene in BAC clones using oligonucleotidesas targeting vectors. By using a PCR-based selec-tive amplification screen to identify targeted clones,this system enables the generation of single-basechanges, deletions (up to 1.93 kb), and the insertionof unique sequences in different regions of a BACcontaining Brca2. This system has since been usedto generate BAC transgenic mice using a Brca2-deficient genetic background [7]. In addition, theability to insert the fusion tag FLAG, that consistsof eight amino acids, including an enterokinase-cleavage site [4], into the Brca2 gene provided aunique method to monitor gene expression.
In order for this experimental approach to besuccessful, DNA sequence information of rele-vant genomic regions containing genes of interestmust be accessible. In our laboratory, we have tar-geted genomic sequencing of chromosomal regionsthat either contribute to resolving complex pheno-types (QTL) or act as biomedical research mod-els. Regions of interest include the porcine MHCclass I, myostatin, neurofibromatosis (NF-1) andthe ataxia-telangiectasia (AT) genes. In this review,we describe how we have utilized the porcine BAC403O5 (from the RPCI-44 library) that contains themyostatin gene to demonstrate our ability to intro-duce small and large insertions and to monitor theexpression of recombineered genes in transfectedfibroblasts.
Genetic modifications of BACs
Using the recombinogenic E. coli strain DY380,we have rapidly generated multiple genetic mod-ifications in BAC clones, harbouring the porcinemyostatin gene [13]. PCR-derived ssDNA frag-ments were used to delete a 68 bp fragment fromexon 3 of the porcine myostatin gene. A deletionin this region disrupts gene expression and blockssynthesis of the myostatin protein, resulting in adouble-muscling phenotype. In order to create atargeted deletion, a 140 bp ssDNA targeting vectorwas constructed using a 100 bp synthetic template(50 bp flanking the deletion site in both directions)and 40 bp oligo 5′ and 3′ primers each of 20 bp
flanking the 100 bp template and 20 bp overlap-ping the template. In the first set of experimentswe used PCR screening with primers, designedusing BAC sequencing information, flanking thedeletion site of the gene. Upon analysis follow-ing recombineering, we identified PCR pools hav-ing both wild-type and recombineered product anddemonstrated a recombineering deletion efficiencyof 1 recombinant/470 electroporated cells. This isa lower targeting efficiency than was obtained ina similar experiment by Swaminathan et al. [16],where they introduced a deletion into exon 11 ofthe murine Brca2 gene with a 1/120 targeting fre-quency. Thus, we re-screened our pools using themismatch amplification mutation assay (MAMA-PCR) and the annealing temperature was adjustedso that a PCR product would only be amplifiedfrom BACs having a deletion, whereas a wild-typeBAC would not be amplified. This approach per-mitted the detection of 1 recombinant/170 electro-porated cells. DNA sequencing of the recombinantclones confirmed that the targeted deletion occurredin the desired location.
Recombineering inserts into porcine BACs
The ability to generate small insertions is a keystep in using the recombineering methodology todevelop an animal model. Experiments were there-fore designed to determine the targeting precisionof inserting specific short sequences into a particu-lar BAC region. The 24 bp sequence encoding theFLAG octapeptide, which when attached to theC-terminus of a protein can be used as a tag to dis-tinguish between normal and modified protein andmonitor protein movement within a cell as wellas for protein purification by affinity chromatogra-phy, was utilized [13]. An oligonucleotide targetingvector (164 bp) to attach the FLAG tag at theC-terminus of myostatin was synthesized by PCRusing two synthetic 94 bp oligonucleotides, whichserved as template and primer simultaneously. Thetargeting vector thus had 70 bp arms homologousto target regions flanking both the 5′ and 3′ end ofthe insertion site in the BAC encoding myostatin,and the 24 bp FLAG sequence. The recombi-nants were identified in cultured pools and then asindividual colonies by FLAG-specific PCR, witha 1/260 targeting efficiency. DNA sequencing ofthree PCR-positive individual clones showed thatthe targeting was specific and that no mutations
Copyright 2004 John Wiley & Sons, Ltd. Comp Funct Genom 2004; 5: 262–267.
Creating porcine biomedical models through recombineering 265
were induced in sequences flanking the insertionsite (500 bp in both directions).
Recombineered target expression in somaticcellsAnother vital component in developing a recom-bineering platform is the ability to use the recom-bineered BACs to introduce targeted changes in
fibroblast and other somatic cells. These studieswere designed to demonstrate (a) that larger, com-plex gene constructs could be developed and(b) that transient gene expression could be mon-itored following BAC transfection into somaticcells. This approach requires either a selectionmarker such as an antibiotic resistance gene oranother selective marker such as green fluorescent
BAC Target for Recombineering
Myostatin Exon 3 95 bp deletion TGA
70 bp homologyupstream of
70 bp homologydownstream
insertion
EGFPpEGFP-N1 vector
PCR amplificationDpnl digestion of template DNA
Targeting Vector(1,772 bp)
120
96
64
32
0
100 101 102 103 104
GFP
Region Count % MeanTotal 1,173,125 100 15
2,6260.01162R3
EGFP
FS
C
B
A
Copyright 2004 John Wiley & Sons, Ltd. Comp Funct Genom 2004; 5: 262–267.

266 M. M. Rogatcheva et al.
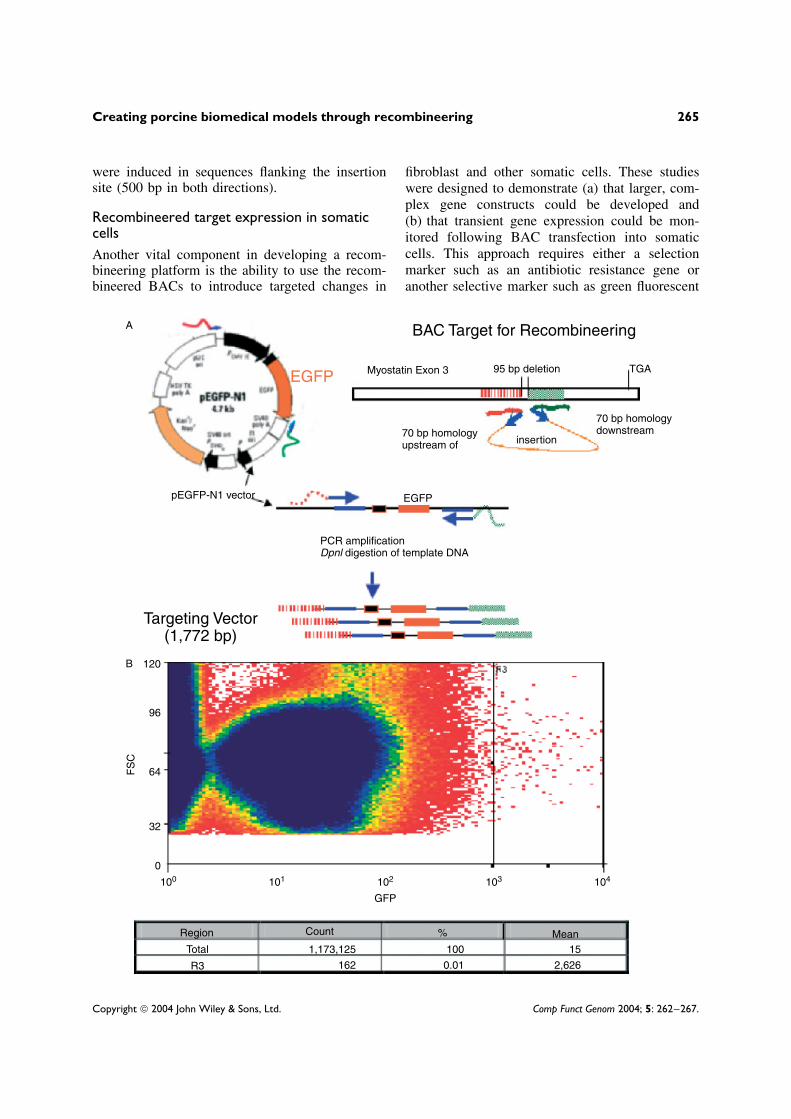
Figure 1. (a) Constructing a targeting vector for EGFP insertion. The targeting vector was constructed using 70 bparms homologous to sequences upstream and downstream of the insertion site (within exon 3 of the porcine myostatingene). The pEGFP-N1 vector (Clontech) was used to amplify the targeting cassette containing the human cytomegalovirusimmediate early promoter. The resulting recombineered vector was PCR-amplified and then treated with DpnI. The DpnIrestriction enzyme cleaves the methylated GATC DNA of the pEGFP-N1 vector, thus preventing contamination with theEGFP expression vector in the recombineered preparation. As illustrated, the targeting vector used for recombineeringis approximately 1.7 kb. (b) EGFP gene expression monitored by flow analysis of transfected porcine fetal fibroblasts.The targeting vector was used to insert EGFP into the myostatin containing BAC (175 kb). E. coli DY380 clonescontaining the modified myostatin–EGFP (recombineered BACs) were grown overnight and BAC DNA was extractedusing a Nucleobond AX kit. Porcine fetal fibroblasts (8HY17F) were then electroporated with BAC DNA (10 µg).Electroporations using different conditions for EGFP expression were used to establish the amount of BAC and the optimaltime of expression. Thus, cells were grown for 3 days and then sorted by flow cytometry to estimate the percentage ofcells transfected and the level of transient gene expression
protein. We have chosen the enhanced green flu-orescent protein (EGFP) as a selection marker(Figure 1a). During the screening of transfectedmammalian cells, EGFP expression can be usedin conjunction with flow cytometric cell sortingto select for homologous recombinants. Using avector containing the EGFP and its promoter andwithout a polyA tail, we constructed a 1592 bpsequence coding for EGFP. A 1772 bp targetingvector was amplified from the plasmid pEGFP-N1(Clontech) using 90 bp PCR primers. Specific PCRamplification using pooled cells was able to detectrecombinants in 4/190 pools, yielding a targetingefficiency for the large targeting cassette (EGFP)of 1/775. Although a higher targeting efficiency isobserved for small oligonucleotide cassettes (dele-tion and FLAG), it is still promising that sucha large insert could be incorporated in a singlestep without the requirement of additional antibi-otic resistance or other selection markers.
The recombineered BAC (EGFP–myostatinfusion gene) was then used to transfect fetal porcinefibroblasts to demonstrate the ability of recom-bineered genes to be expressed. As shown inFigure 1b, we were able to demonstrate the expres-sion of EGFP by fetal fibroblasts as measured byflow cytometry. This study showed the efficiencyof transient expression of recombineered myostatinBAC (approximately 170 kb) to be around 0.01%,which is consistent with previous studies [8]. Theseresults suggest that insertions longer than 1000 bpcan be successfully recombineered into a geneof interest. Constructs generated by this approachcould also be used to develop specifically modifiedsomatic cells for conditional expression, GOF orLOF studies.
Discussion
This review summarizes our development of asomatic cell technology platform that integrates therecombineering of genomic DNA with the subse-quent targeted gene manipulation of somatic celllines. This platform provides a novel approachfor dissecting physiological pathways using definedgenetic systems in vitro while avoiding the costof validation in non-inbred animals and ‘geneticnoise’ from various background genes. The use ofgene-targeted fibroblasts provides a suitable nucleardonor for use in nuclear transfer to generate uniqueswine germplasm for generating biomedical mod-els. The targeting efficiencies observed in this studyfor FLAG insertion and deletions are comparableto those observed using a mouse model [7,16]. Ourattention is now focused on developing site-specificrecombination systems to permit targeted allelicsubstitution in porcine BACs. This will permit us totarget gene function in somatic fibroblast lines andevaluate downstream events prior to in vivo stud-ies. As previously reported by Montigny et al. [8],we were able to demonstrate that BAC DNA purityand concentration did affect transfection efficiency.Furthermore, although the BAC construct used herewas extremely large (>175 kb), we were able todemonstrate transfection and expression profilesconsistent with other studies [8].
Acknowledgements
This work was partially supported by grants from theUSDA–National Research Initiative (2002-35205-12712),the USDA Cooperative State Research Service (AG2002-34480-11828) and the USDA Agricultural Research Service(Agreement No. 58-5438-2-313). The authors wish torecognize the contributions and assistance of E. Forsberg,
Copyright 2004 John Wiley & Sons, Ltd. Comp Funct Genom 2004; 5: 262–267.

Creating porcine biomedical models through recombineering 267
Infigen. The authors also acknowledge the support andassistance of N. Copeland and N. Jenkins (NCI).
References
1. Chowdhary BP, Raudsepp T, Fronicke L, Scherthan H. 1998.Emerging patterns of comparative genome organization insome mammalian species as revealed by Zoo-FISH. GenomeRes 8: 577–589.
2. Copeland NG, Jenkins NA, Court DL. 2001. Recombineering:a powerful new tool for mouse functional genomics. NatureRev 2: 769–779.
3. Dow JAT, Davies SA. 2003. Integrative physiology andfunctional genomics of epithelial function in a genetic modelorganism. Physiol Rev 83: 687–729.
4. Einhauer A, Jungbauer A. 2001. The FLAG peptide, aversatile fusion tag for the purification of recombinantproteins. J Biochem Biophys Meth 49: 455–465.
5. Fahrenkrug SC, Rohrer GA, Freking BA, et al. 2001. Aporcine BAC library with tenfold genome coverage: a resourcefor physical and genetic map integration. Mamm Genome 12:472–474.
6. Hawken RJ, Murtaugh J, Flickinger GH, et al. 1999. A firstgeneration porcine whole-genome radiation hybrid map.Mamm Genome 10: 824–830.
7. Lee EC, Yu D, Martinez de Velasco J, et al. 2001. A highlyefficient Escherichia coli-based chromosome engineeringsystem adapted for recombinogenic targeting and subcloningof BAC DNA. Genomics 73: 56–65.
8. Montigny WJ, Phelps SF, Illenye S, Heintz NH. 2003.Parameters influencing high-efficiency transfection of bacterialartificial chromosomes into cultured mammalian cells.BioTechniques 35: 796–807.
9. Muyrers JPP, Zhang Y, Benes V, et al. 2000. Point mutationof bacterial artificial chromosomes by ET recombination.EMBO Rep 1: 239–243.
10. NRC (National Research Council). 1998. Biomedical Modelsand Resources: Current Needs and Future Opportunities.National Academy Press: Washington, DC.
11. Paszek AA, Wilkie PJ, Flickinger GH, et al. 1999. Intervalmapping of growth in divergent swine cross. Mamm Genome10: 117–122.
12. Rettenberger G, Klett C, Zechner U, et al. 1995. Visualizationof the conservation of synteny between humans and pigs byheterologous chromosomal painting. Genomics 26: 372–378.
13. Rogatcheva MB, Rund LA, Beever JE, Counter CM,Schook LB. 2002. Recombineering pig BACs for somatic cellgenomics. Int Soc Animal Genet A022.
14. Rogel-Gaillard C, Bourgeaux N, Billault A, Vaiman M,Chardon P. 1999. Construction of a swine BAC library:application to the characterization and mapping of porcine typeC endoviral elements. Cytogenet Cell Genet 85: 205–211.
15. Rohrer GA, Alexander LJ, Hu Z, et al. 1996. A comprehen-sive map of the porcine genome. Genome Res 6: 371–391.
16. Swaminathan S, Ellis HM, Waters LS, et al. 2001. Rapidengineering of bacterial artificial chromosomes usingoligonucleotides. Genesis 29: 14–21.
17. Trey R, Vesteinn T, Ranish JA, et al. 2001. Integratedgenomic and proteomic analyses of a systemically perturbedmetabolic network. Science 292: 929–934.
18. Tumbleson M, Schook LB. 1996. Advances in Swine inBiomedical Research. Kluwer Academic: New York.
19. Yerle M, Echard G, Robic A, et al. 1996. A somatic cellhybrid panel for pig regional gene mapping characterized bymolecular cytogenetics. Cytogenet Cell Genet 73: 194–202.
20. Zhang Y, Buchholz F, Muyrers JPP, Stewart AF. 1998. Anew logic for DNA engineering using recombination inEscherichia coli . Nature Genet 20: 123–128.
Copyright 2004 John Wiley & Sons, Ltd. Comp Funct Genom 2004; 5: 262–267.

Submit your manuscripts athttp://www.hindawi.com
Hindawi Publishing Corporationhttp://www.hindawi.com Volume 2014
Anatomy Research International
PeptidesInternational Journal of
Hindawi Publishing Corporationhttp://www.hindawi.com Volume 2014
Hindawi Publishing Corporation http://www.hindawi.com
International Journal of
Volume 2014
Zoology
Hindawi Publishing Corporationhttp://www.hindawi.com Volume 2014
Molecular Biology International
GenomicsInternational Journal of
Hindawi Publishing Corporationhttp://www.hindawi.com Volume 2014
The Scientific World JournalHindawi Publishing Corporation http://www.hindawi.com Volume 2014
Hindawi Publishing Corporationhttp://www.hindawi.com Volume 2014
BioinformaticsAdvances in
Marine BiologyJournal of
Hindawi Publishing Corporationhttp://www.hindawi.com Volume 2014
Hindawi Publishing Corporationhttp://www.hindawi.com Volume 2014
Signal TransductionJournal of
Hindawi Publishing Corporationhttp://www.hindawi.com Volume 2014
BioMed Research International
Evolutionary BiologyInternational Journal of
Hindawi Publishing Corporationhttp://www.hindawi.com Volume 2014
Hindawi Publishing Corporationhttp://www.hindawi.com Volume 2014
Biochemistry Research International
ArchaeaHindawi Publishing Corporationhttp://www.hindawi.com Volume 2014
Hindawi Publishing Corporationhttp://www.hindawi.com Volume 2014
Genetics Research International
Hindawi Publishing Corporationhttp://www.hindawi.com Volume 2014
Advances in
Virolog y
Hindawi Publishing Corporationhttp://www.hindawi.com
Nucleic AcidsJournal of
Volume 2014
Stem CellsInternational
Hindawi Publishing Corporationhttp://www.hindawi.com Volume 2014
Hindawi Publishing Corporationhttp://www.hindawi.com Volume 2014
Enzyme Research
Hindawi Publishing Corporationhttp://www.hindawi.com Volume 2014
International Journal of
Microbiology