cryptogam spores and other non-pollen microfossils as sources
TRANSCRIPT

Ann. Bot. Fennici 39: 1–14 ISSN 0003-455XHelsinki 8 March 2002 © Finnish Zoological and Botanical Publishing Board 2002
Cryptogam spores and other non-pollenmicrofossils as sources of palaeoecologicalinformation: case-studies from Spain
José S. Carrión & Cristina Navarro
Carrión, J. S. &, Navarro, C., Departamento de Biología Vegetal (Botánica),Facultad de Biología, Universidad de Murcia, 30100 Murcia, Spain (e-mail:[email protected])
Received 17 May 2001, accepted 10 August 2001
Carrión, J. S. & Navarro, C. 2002: Cryptogam spores and other non-pollenmicrofossils as sources of palaeoecological information: case-studies fromSpain. — Ann. Bot. Fennici 39: 1–14.
Four examples from Mediterranean Spain are used to show that records of non-pollenpalynomorphs (algal and fungal spores and cysts, charophytes and other microfossilsof unknown biological origin) are worthwhile in addition to pollen analysis for studiesof palaeoecological reconstruction. In the lacustrine sequence of Cañada de la Cruz,the stratigraphy of palaeolimnological indicators is compatible with climatic controlof vegetation stages at the response scales of decades to centuries. The sequence ofNavarrés provides evidence of millennial-scale change in the trophic conditions of apeat-forming basin, which parallels local and vegetation changes since the last glacialto the Holocene under the influence of fire events, climate change and human activity.The sequence of Villaverde shows out-of-phase relationships of vegetational andlimnological developments. The sequence of San Rafael offers an example ofsynchronous variation of xerophytic pollen and microfossils indicative of temporarymarsh desiccation, in concordance with regional evidence of aridification since themid-Holocene.
Key words: algal cysts, fungi, Holocene, palaeobotany, palaeoecology, palaeolimnol-ogy, Quaternary, Spain, spores
Introduction
The combination of pollen analysis with therecords of cryptogam spores and other non-
pollen palynomorphs has shown to be valuablefor detailed reconstructions of the changing lo-cal environment of Quaternary sites from north-ern Europe (e.g., Van Geel et al. 1989). In lake

Carrión & Navarro • ANN. BOT. FENNICI Vol. 392
sediments, for instance, long-term rapid changesin the microfossil record may be limnologicalresponses via variations in the flux of water,nutrients and particulate material to the site fromits drainage basin (Moore 1986). In studies ofpalaeoclimatic reconstruction, it is of paramountimportance to elucidate whether limnologicalchanges correlate to events of vegetation change.To judge from the published record, however,research on non-pollen palynomorphs has formedlittle or no part of studies of the majority ofQuaternary palynologists from southern Europe.Moreover, the synchronous history of vegetationtypes and “extra” palynomorphs is still a ratherunexplored field of study in the Mediterraneanarea. This paper offers examples of the valuableuse of this approach in deposits from southernSpain.
Methods
The selected case studies correspond to the lateQuaternary palaeoecological sequences of Caña-da de la Cruz (Jaén), Navarrés (Valencia), Vil-laverde (Albacete), and San Rafael (Almería) inthe Mediterranean region of Spain (Fig. 1). Thesection of Navarrés was obtained by means ofmetal boxes, the sharp edges of which werepushed into an exposed profile and subsequentlycut out. The three remaining basins were sam-pled with piston corers. Extraction of pollen andspores from the sediment matrix followed thestandard procedure described in Moore et al.(1991), which, by using hydrofluoric acid, pre-
cludes the preservation of microfossils possess-ing silicified walls such as diatoms. Mineralseparation with zinc chloride density 2.0 wasused for all the samples. Exotic Lycopodiumtablets of a known concentration were added tocalculate pollen concentrations.
Identification of palynomorphs was aided bythe use of the reference collections at Universi-dad de Murcia and Universiteit van Amsterdam.For the nomenclature of non-pollen microfossilsof unknown or controversial biological origin,we have followed the sequence established byvan Geel et al. (1989), in which the typesindicated by a number can be considered asprovisionally, but not formally, named form-taxa (Figs. 2–4). Relevant information on thesetypes can be found in references listed inTable 1. Identification criteria for Characeaegyrogonites followed Haas (1994).
�
��������
��������������
��� �� ���
�������������
��� ������������
�
�
�
�
��������
�� ���!�
�"����
#�$�
���� ����
%�&��
�
�
�
Fig. 1. Location of the studied sites.
Fig. 2. LM and SEM micrographs of palynomorphs.— 1 and 2: Closterium, ¥1600. — 3: Botryococcus,¥704. — 4, 5, 7: Zygnema-type, ¥1760. — 6: Spiro-gyra-type, ¥1600. — 8: Rivularia-type heterocyst,¥1950. — 9: Chara hispida gyrogonite, ¥110 (SEM)
ANN. BOT. FENNICI Vol. 39 • Cryptogam spores and palaeoecology 3
Percentage pollen diagrams of selected taxawere elaborated using Tilia 1.12 and TiliaGraph1.18 programs (Grimm 1992). Between ca. 400and 1500 total palynomorphs were counted foreach sample. Hydro- and hygrophytic pollen,and non-pollen microfossils were excluded fromthe pollen sum. In Navarrés and Villaverde,these diagrams also include concentrations ofcharcoal particles and Characeae gyrogonites.The total number of charcoal particles in themicroscope slides can be taken as an indicationof regional fire occurrence, while macrocharcoalpeaks can be rather indicative of local events offire (Mehringer et al. 1977). The delimitation ofpercentage zone boundaries was aided by strati-graphically constrained incremental sum-of-squares analysis (CONISS analysis, Grimm1992) using a square-root transformation and
chord-distance dissimilarity measure for all pol-len and spores taxa. In Villaverde, a distinctionwas made between “pollen” (VP) and “micro-fossil” (VM) zones, the latter including thestratigraphic variation of non-pollen microfossilsand basin-associated pollen. The number of zoneswas finally determined using the method ofBennett (1996) and the delimitation of VP andVM assemblage zone boundaries was obtainedfrom an optimal division of the sequence usingthe information content criterion, after explora-tion of four other methods (binary splitting bysums-of-squares, binary splitting by informationcontent, optimal splitting by sum-of-squares,and constrained cluster analysis). All chronolo-gies were established on the basis of radiocar-bon dates obtained from bulk organic sediment,dated by the AMS method where necessary
Fig. 3. LM micrographs of palynomorphs. — 1: The-caphora, ¥1760. — 2 and 3: Sordariaceae, ¥1760.— 4: Gelasinospora, ¥1760. — 5: Pteridophyta tri-lete, ¥1600. — 6: Type 179, ¥1760. — 7 and 8:Type 984, ¥1760. — 9 and 10: Type 983, ¥1760. —11: Type 985, ¥1760
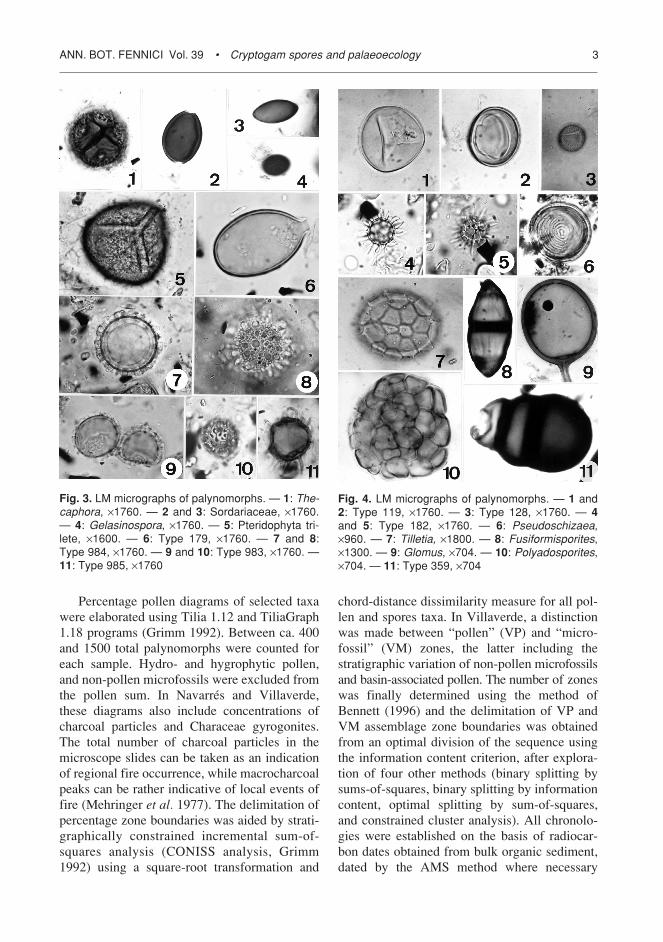
Fig. 4. LM micrographs of palynomorphs. — 1 and2: Type 119, ¥1760. — 3: Type 128, ¥1760. — 4and 5: Type 182, ¥1760. — 6: Pseudoschizaea,¥960. — 7: Tilletia, ¥1800. — 8: Fusiformisporites,¥1300. — 9: Glomus, ¥704. — 10: Polyadosporites,¥704. — 11: Type 359, ¥704

Carrión & Navarro • ANN. BOT. FENNICI Vol. 394
Tab
le 1
. Non
-pol
len
mic
rofo
ssils
sel
ecte
d fo
r th
is s
tudy
. Eco
logi
cal i
ndic
atio
n, b
iolo
gica
l and
taxo
nom
ic a
ffini
ties
and
refe
renc
e lit
erat
ure.
——
——
——
——
——
——
——
——
——
——
——
——
——
——
——
——
——
——
——
——
——
——
——
——
——
——
——
——
——
——
——
——
——
——
——
Mic
rofo
ssil
type
Indi
catio
nT
axon
omic
/bio
logi
cal a
ffini
ties
Lite
ratu
re—
——
——
——
——
——
——
——
——
——
——
——
——
——
——
——
——
——
——
——
——
——
——
——
——
——
——
——
——
——
——
——
——
——
——
—G
lom
us (
T. 2
07)
Col
onis
atio
n of
lake
mar
gins
, roo
tG
lom
acea
e ch
lam
ydos
pore
sV
an G
eel e
t al.
(198
9), C
arrió
n et
al.
(199
5,ac
tivity
1999
)F
usifo
rmis
porit
esO
rgan
ic m
atte
r de
com
posi
tion
Unk
now
n. A
scos
pore
s of
e.g
. Nec
tria
,E
lsik
(19
80, 1
983)
, Car
rión
& v
an G
eel (
1999
)H
erpo
tric
hia,
Par
odie
lla, C
erio
phor
a ?P
olya
dosp
orite
s (T
. 367
)O
rgan
ic m
atte
r de
com
posi
tion
Col
onie
s of
thin
-wal
led
fung
al c
ells
?Ja
rzen
& E
lsik
(19
86),
Van
Gee
l et a
l . 19
81)
Till
etia
(T
. 27)
Fun
gal a
ctiv
ityT
illet
ia s
pore
sV
an G
eel (
1972
), N
avar
ro e
t al.
(200
0)T
heca
phor
a (T
. 364
)F
unga
l act
ivity
, agr
icul
tura
l pra
ctic
es?
The
caph
ora
basi
dios
pore
sV
an G
eel e
t al.
(198
1)G
elas
inos
pora
(T
.1)
Dec
ompo
sing
act
ivity
Gel
asin
ospo
ra a
scos
pore
sLu
ndqv
ist (
1972
), V
an G
eel (
1978
)T
ype
359
Fun
gal a
ctiv
ityS
epta
te c
onid
iaV
an G
eel e
t al.
(198
1)S
orda
riace
ae (
T.5
5)E
utro
phic
con
ditio
ns, d
ung
inpu
tS
orda
riace
ae a
scos
pore
sV
an G
eel (
1978
), V
an G
eel e
t al.
(198
9),
Car
rión
et a
l. (1
999,
200
0b),
Nav
arro
et a
l. (2
000)
Zyg
nem
atac
eae
Sha
llow
sta
gnan
t, ox
ygen
-ric
h w
ater
s,Z
ygos
pore
s an
d ap
lano
spor
es o
fV
an G
eel e
t al.
(198
9)(S
piro
gyra
, Zyg
nem
a,la
ke m
argi
nsZ
ygne
mat
acea
eM
ouge
otia
- typ
es)
Riv
ular
ia-t
ype
(T.1
70)
Incr
ease
d al
kalin
isat
ion
and
Cya
noph
yta
hete
rocy
sts
Van
Gee
l et a
l. (1
983)
conc
entr
atio
n of
oxi
diza
ble
orga
nics
inw
ater
bod
ies
Clo
ster
ium
Olig
o-m
esot
roph
ic, t
empe
rate
to c
old
Zyg
ospo
res
of s
ever
al s
peci
es o
fV
an G
eel e
t al.
(198
1)id
iosp
orum
type
(T
. 60)
wat
erC
lost
eriu
mB
otry
ococ
cus
Ope
n w
ater
Col
onie
s of
Bot
ryoc
occu
sM
ateu
s (1
992)
Pse
udos
chiz
aea
Rel
ativ
ely
war
m c
limat
e w
ith lo
cal
Unk
now
n, R
estin
g st
ruct
ure
of a
ny o
fS
cott
(199
2)se
ason
al d
ryin
ga
wid
e va
riety
of o
rgan
ism
sT
ype
128
Mes
otro
phic
, col
d to
tem
pera
te o
pen
Unk
now
n, A
lgal
cys
t or
spor
e?, S
imila
rK
rutz
sch
(197
0), P
als
et a
l. (1
980)
, Van
Gee
lw
ater
to S
igm
opol
lis h
ispi
dus
(Hed
lund
196
5)et
al.
(198
3)T
ype
119
Lacu
strin
e en
viro
nmen
t with
aqu
atic
Unk
now
nP
als
et a
l. (1
980)
angi
ospe
rms
Typ
e 17
9S
tagn
ant o
pen
wat
er, e
utro
phic
Unk
now
n, P
hore
tic c
ysts
of e
pibi
ontic
Van
Gee
l et a
l. (1
989)
, Wol
owsk
i (19
98)
cond
ition
sor
par
asiti
c C
opep
oda,
Eug
leno
phyt
a?T
ype
182
Sta
gnan
t sha
llow
ope
n w
ater
Unk
now
n, A
lgal
orig
in?
van
Gee
l et a
l. (1
983)
Typ
e 98
3S
hallo
w fr
eshw
ater
Unk
now
n, E
ncys
tmen
t pha
se o
f alg
aeS
trot
her
(199
6), C
arrió
n &
van
Gee
l (19
99)
(e.g
. des
mid
s)?
Typ
e 98
4S
hallo
w fr
eshw
ater
Unk
now
n, E
uast
rum
zyg
ospo
res?
Car
rión
& v
an G
eel (
1999
)T
ype
985
Sha
llow
fres
hwat
erU
nkno
wn,
Alg
al o
rigin
?C
arrió
n &
van
Gee
l (19
99)
——
——
——
——
——
——
——
——
——
——
——
——
——
——
——
——
——
——
——
——
——
——
——
——
——
——
——
——
——
——
——
——
——
——
——

ANN. BOT. FENNICI Vol. 39 • Cryptogam spores and palaeoecology 5
because of low carbon content in the organicextracts.
Palaeoecological sequences
Cañada de la Cruz (Fig. 5)
The Cañada de la Cruz (38∞04´N, 2∞42´W,1595 m a.s.l.) is a shallow lake situated in ahigh-altitude mountain valley within the Sierrade Segura, Betic Cordillera (Fig. 1). It extendsup to ca. 0.25 ¥ 2.7 km during high-rainfallyears, but normally dries up for 6 to 8 monthsper year, being fed by direct precipitation andsurface runoff from the catchment. Pollen andspores were only found in the uppermost 128 cmof a sediment core raised from a central point inthe dry lake. The depositional context is a com-pact bed of brown calcareous clays overlying asmall-scale river deposit of calcreted conglom-erates. The pollen sequence has provided in-sights into the vegetation history of the regionsince ca. 8320 yr BP. Abrupt changes in abun-dance of the pollen dominants are shown atabout 7770 yr BP, 3370 yr BP, 2630 yr BP,1525 yr BP, and 790 yr BP (Fig. 5). Thissequence has been crucial to demonstrate thatpollen may be a sensitive indicator of Holoceneclimate changes at the multi-decadal to centuryscale in Mediterranean context (Carrión et al.2001b).
Pollen curves generally correlate to patternsof non-pollen palynomorphs. Several facts sup-port the view that the stratigraphy of palaeolim-nological indicators is compatible with climaticcontrol of vegetation stages. Firstly, the ecotonebetween pine forest and xerophytic grassland-scrub is known to be sensitive to thermal lapserate, wind intensity, summer length, and rainfalldistribution (Valle et al. 1989). Secondly, thecorrelation between Poaceae, Artemisia, Cheno-podiaceae, Juniperus, and Ephedra nebrodensissuggests cold arid conditions for the stages inwhich pollen of these taxa increase in value,namely at ca. 8320–7770 yr BP, 3370–2630 yr BP,and to a lesser extent, 1525–790 yr BP. Declinesof these herbaceous assemblages correspond withincreases in value of pollen of mesophilous(Quercus, Pinus pinaster, Fraxinus, Acer, Erica
arborea, Alnus, Betula, Corylus, Arbutus, Ul-mus) and thermophilous (Pistacia, Cistus, Phil-lyrea, Ephedra fragilis, Olea) taxa. Thus, themontane climate must have been relativelywarmer and wetter during the periods of ca.7770–3370 yr BP, 2630–1525 yr BP, and790 yr BP to present. During these stages,invasion of high-elevation grassland-scrub bypines is consistent with upward displacement ofdeciduous forests, and expansion of evergreenforest and scrub (Carrión et al. 2001b). Thirdly,zones C1, C3, and, to a lesser extent, C5,characterized by Closterium and Type 128, andincreased minerogenic influx, may representstages with comparatively poor-nutrient water inthe context of low temperatures, and short sum-mers leading to treeless catchment vegetation. Incontrast, zones C2, C4, and C6, with character-istic Zygnemataceae-dominated assemblages,decomposing fungi, and hygrophyte pollenwould represent moderate eutrophication andtemporary colonisation of the lake by marginalvegetation within a more forested landscape andmilder climate. Finally, Polygonum dumetorumtype may also indicate shifts to longer snow-freeperiods and mesic summer conditions. The ab-sence of both peaty layers and preserved mac-roremains suggests, however, that prolongeddessication of the basin, organic matter decom-position and mineralization occurred throughoutthe sequence. The highest values of the pelagicBotryococcus suggest that water levels mayhave reached their maxima during C2, in con-cordance with spread of deciduous oaks. How-ever, water levels would have diminished soonthereafter, as the subsequent peak of Zygne-mataceae suggests.
Navarrés (Fig. 6)
The Canal de Navarrés is a tectonic, endorheicvalley (39∞06´N, 0∞41´W, 225 m a.s.l.), 1 to2 km wide, running NW–SE in the limits of theIberian Cordillera, province of Valencia, easternSpain (Fig. 1). A sedimentary sequence of 25 mdepth was preliminarly reported (Dupré et al.1998) describing 10 lithostratigraphic units inthe valley deposit, of which only the topmostlayer (above 250 cm depth) was peaty and

Carrión & Navarro • ANN. BOT. FENNICI Vol. 396
Fig
. 5.
Pal
aeoe
colo
gica
l seq
uenc
e of
Cañ
ada
de la
Cru
z (J
aén)
. A
P:
sum
of
arbo
real
pol
len.
“D
ecid
uous
tre
es”:
dec
iduo
us Q
uerc
us,
Fra
xinu
s, A
cer,
Aln
us,
Bet
ula,
Cor
ylus
, U
lmus
, S
alix
. “M
edite
rran
ean
fore
st-s
crub
”: P
inus
pin
aste
r , e
verg
reen
Que
rcus
, E
rica
arbo
rea,
Arb
utus
, E
phed
ra f
ragi
lis,
Pis
taci
a, C
istu
s,P
hilly
rea.
“X
erop
hyte
s”:
Poa
ceae
, Ju
nipe
rus,
Art
emis
ia,
Eph
edra
neb
rode
nsis
, C
heno
podi
acea
e, A
ster
acea
e. H
ydro
-hyg
roph
ytes
: P
olyg
onum
dum
etor
umty
pe,
Cyp
erac
eae,
Typ
ha.
Cro
sses
ind
icat
e pa
lyno
mor
ph f
requ
enci
es b
elow
2%
. A
ges
in u
ncal
ibra
ted
radi
ocar
bon
year
s B
P.
Hyg
roph
yte
polle
n an
d no
n-po
llen
paly
nom
orph
s ex
clud
ed fr
om p
olle
n su
m. Z
onat
ion
follo
ws
Grim
m (
1992
)
�
��
��
��
���
����
���
�� �
������������
���
�����
��
��
��
�
�
��
��
��
���
��
��
��
��
��
��
��
��
��
��
��
��
��
��
��
��
��
��
��
��
��
���
���
!"#
�!$"�$!%
��&
����
���
'()*
)+���
�+
#�(
'����,-
�,-.*��+�/+��)0
1��*�
&�'�
�* �*
-�-�
2&3
�*/&(
�*�
&��+
4�%$�#��
�����
���
������"�
����
������ !���
����
�����+
��',5
.)-3
'
�&��
���
�' -*5*3'�,5�,-3�+
6�3��,�'*-�,-3�+
���7��
���7��
����7��
����7��
��8�7��
��8�7���
88��7��
����7��
����7��
�8��7��
����7��
88��7��
88��78�

ANN. BOT. FENNICI Vol. 39 • Cryptogam spores and palaeoecology 7
Fig
. 6.
Pal
aeoe
colo
gica
l se
quen
ce o
f N
avar
rés
(Val
enci
a).
Cro
sses
ind
icat
e pa
lyno
mor
ph f
requ
enci
es b
elow
2%
. A
ges
in u
ncal
ibra
ted
radi
ocar
bon
year
s.Z
onat
ion
follo
ws
Grim
m (
1992
)
��
��
���
���
���
��
��
��
�
� ��
����
���
���
���
��
��
�
��
����
���
��
��
���
����
���
��
�
�
��
!"#$
#%�
��
�
�
���
&'(
)*
��
+�
��,
��
��
���
���
, �
��-�
.�
�
���
���
��
�. �
&��!"
$�'�
&/
��.
0
�
� ���
��
���
����
���
�
��
+�
��
�,��
��,
.�
�
�-
�
����
0��
��
�!)
�$1$�! /1 '/���%
����&/&!$� '/���%
� ��
�23
� ��
���
���
4
�
�����
�
���
�(%
��"%
*
� ��
��3
�.
��
��
���
���
��
��
����
��
.��.
��
�.
� ��
.��.
���
��
0.
�
�.
���
����
����
.�
�
���
��
5 �
�0
���
��
��.
���
��
0
���
�.
��
,�5
���
-.
�� .
�.
��
�
���
��
���
����
���
+�
��
��
��
��
6��6
.�
0��
�.
���
��
� ��
3��
�����
����
���
���
���$�78���
���
��
� ��
3��
� ��
3��
0��
�.
�,
��
�.
��
(9��
�:)
*
�,
��
�,
�����
�
���
��
��
��
��
��
�/�&! 1�%8���
����
�����;��
��3��;���
�����;���
�2�3�;���
�����;+��
�����;��
+���;��
��+��;+�
+���;2�
�3��;��
3�+�;��
��+�;���
=

Carrión & Navarro • ANN. BOT. FENNICI Vol. 398
polliniferous. This upper unit was formed undercontinuous sedimentation processes in a shallowlacustrine environment. Several pollen phaseshave been reported from the study of this section(Carrión & Dupré 1997, Dupré et al. 1998), thelast one covering the period from ca. 30 900 to3160 yr BP (Carrión & van Geel 1999).
The last glacial vegetation (N3PA-N3PB) ischaracterized by Pinus–Artemisia–Ephedra as-semblages (Fig. 6). This dominance is interrupt-ed between ca. 30 260 and 27 890 uncalibrated14C yr BP by the development of Quercus, Pinuspinaster, deciduous trees and Mediterraneanshrubs, suggesting the proximity of glacial refu-gia and an expansion of their vegetation underinfluence of a milder climate. The palynologicalsequence of N3PB is interrupted between 166and 145 cm. This hiatus seems to correspondwith the last glacial maximum, during whichconditions were not favourable for biotic preser-vation in wide parts of the basin. A YoungerDryas signal is indicated by increases of Artemi-sia and Ephedra around 10 340 BP. There is noimmediate response of Quercus to the lategla-cial and Holocene climatic ameliorations andPinus continues to dominate the landscape untildrastic drop at ca. 5930 BP, presumably bysevere fire events, which may be linked with theinstallation of Neolithic settlement in the vicini-ties (Carrión & van Geel 1999). The suggestionof fire derives from the sharp increase of char-coal particles prior to the vegetation change,especially at 69 cm (dated at ca. 6310 BP).
Above 145 cm, there is evidence of a changein the trophic conditions, shallow water andplant advancement into the lake, as shown bythe curves of Cyperaceae, some fungal types(Type 359 and Glomus) and Zygnema type.Still, the occurrence of Rivularia and the contin-uous curves of Closterium type and Botryococ-cus along zone N3PB indicate the presence ofopen water. Type 128 also occurs continuously.During N3PC, Type 128 and Closterium dimin-ish and Botryococcus and zygnemataceous zy-gospores (Zygnema first and Spirogyra after-wards) increase. During N3PD, there are indica-tors of increased colonization of the lake mar-gins and peat decomposition (e.g. pollen ofCyperaceae, Typha, Apium, spores of terrestrialferns, fungal types like Gelasinospora, Fusi-
formisporites, Polyadosporites, Glomus). Hu-man influence in the landscape is suggested bythe curve of Plantago. Since it frequently para-sites grasses, Thecaphora, might also be relatedwith agricultural practices. In addition, there areindicators of dung input such as spores of Sord-ariaceae.
The algal succession observed since the Late-glacial is remarkable, and is characterized by thefollowing assemblages: (N3PB) Types 128–Clos-terium–Botryococcus, (N3PC) Zygnema–Spiro-gyra–Botryococcus, (N3PD) Zygnema–Spirogy-ra–Mougeotia–Rivularia–Chara. This successionmay be certainly related with a trophic change toincreased alkalinization and concentration of oxi-dizable organic compounds in relatively warmwater. Types 179, 983, 984, and 985 parallel thisdistribution, with the very abundant Type 983showing a peak just when maxima for Quercusand Chara hispida are recorded. This Type mustbe related with a shallow freshwater lake environ-ment. Finally there is an important increase ofangiosperm seeds, notably Juncus articulatus,which suggests partial infilling.
Villaverde (Fig. 7)
The site (2∞22´W, 38∞48´N, 870 m a.s.l.) is atuffaceous peat deposit developed on an oldlake-bed adjacent to the Cubillo River, 57 kmwest of the city of Albacete (Fig. 1). A sedimentcore (total depth 550 cm) was obtained with apiston corer at the eastern part of the fan. Thepollen stratigraphy at Villaverde has been usedto infer past vegetation changes from ca. 8700 to1230 uncalibrated 14C yr BP within a well-defined catchment area that represents boundaryconditions for semi-arid, plateau and mountainvegetation (Carrión et al. 2001a). At the millen-nial scale, climate appears to have been a majorcontrol of the species pool. The picture is of agenerally wet mid Holocene stage (VP3–VP5)characterized by spread of mesophilous vegeta-tion, anteceded and followed by drier condi-tions, which are inferred from a higher abun-dance of xerophytes, pines, evergreen oaks, andMediterranean scrub (Fig. 7).
The lithology comprises pale-brown organicmarls (VM1, VM3, VM5) and black organic

ANN. BOT. FENNICI Vol. 39 • Cryptogam spores and palaeoecology 9
Fig
. 7. P
alae
oeco
logi
cal s
eque
nce
of V
illav
erde
. VP
: pol
len
stra
tigra
phy,
VM
: str
atig
raph
y of
oth
er m
icro
foss
ils. V
P a
nd V
M z
ones
are
out
-of-
phas
e be
low
c.
350
cm d
epth
. AP
: sum
of a
rbor
eal p
olle
n. “
Xer
ophy
tes’
’: A
rtem
isia
, Che
nopo
diac
eae,
Eph
edra
frag
ilis ,
and
Ast
erac
eae.
“O
ther
mes
ophi
lous
tree
s’’:
Bet
ula,
Fra
xinu
s , S
alix
, C
oryl
us,
Aln
us,
Ulm
us,
Ace
r, a
nd S
orbu
s . “
Oth
er M
edite
rran
ean’
’: P
ista
cia ,
Phi
llyre
a , E
rica
arbo
rea ,
Ole
a, C
istu
s , R
ham
nus ,
and
Myr
tus.
Cro
sses
indi
cate
pal
ynom
orph
fre
quen
cies
bel
ow 2
%.
Age
s in
unc
alib
rate
d ra
dioc
arbo
n ye
ars.
Del
imita
tion
of V
P a
nd V
M z
one
boun
darie
s fo
llow
s B
enne
tt(1
996)
���
���
�����
��
�
���
���
����
���������
���
���
����
������
�����
���
��
�
���
���
���
���
� �
���
���
���
���
���
���
���
���
���
����
��
���
� �
!��
! �
"��
" �
#��
# �
��
�
�$�
�%��
��������
&��
���'
���
(� �
�!)
(� �
��*
�++�
�(�
,�-�(�
.
���/�
�(�
,
�� �������
�++�
(� ��)
!
(� �
�0* �
/1�2�
�/�
3�-.�,
�(�����
�2�
�(�
�/�
,
��
����
���
����
��
� �
���
���
���
���
���
���
���
�� �����
(� �
*)
(� �
*)#
�������4/3
���
�����
���
��
�
�%'�
�%�
����5%6
�
!�
!�
78�
#�
!�
!�
#�
!�
8�
!�
!�
!�
!�
!�
���
!�
"�
8�
!�
�2�
���� ��/(
��
-2�
,��
�++�
!�
2�����%�����%��
1�%�������%��
#�
�!"�9"
�8)�9 �
!!8�98�
!8 �9#
"!#�9 �
�#�98�
880�98�
)0!�9)�
-��'���������:��
����;�
���<����'
����������
���<=��>��?�����
����������
��:��@
��� �
���
���<����������
��:��@
��� �
���
��������=
���:��@
@���:��<
����������
��� �
���
��:��@���
@�����::����
��:���
���������

Carrión & Navarro • ANN. BOT. FENNICI Vol. 3910
sediments of varying states of humification(“peats”: VM6, VM7) or decomposition (“sapro-pels”: VM2, VM4) (Fig. 7). Characeae mineraldetritus is the main component of plant macro-fossils throughout the core from VM2 upwards,although the detritus component is better pre-served in marl layers. This detritus consists ofbranchlets, fragments of axial internodes withthe typical diplostichous-anisostichous cortex,clusters of spine cells attached to axial frag-ments, and gyrogonites. The variations of thelithology suggest fluctuating water levels withthe deposition of marls during periods of higherlake levels and the deposition of peats associat-ed with advances of littoral emergent vegetationduring lower lake levels. Overall, charophyte-rich facies throughout the core confirms thepresence of permanent, albeit relatively shallow,water conditions. The abundance of Chara hisp-ida gyrogonites in VM3 and VM5 indicates insitu deposition as well as submersion for at leastthree months, which is the time required for theplant to complete a full cycle of growth includ-ing calcification of the reproductive organs(Soulié-Märsche 1993). Associated microfossilassemblages (VM3, VM5) fit into this picture.Similarly to Navarrés (Fig. 6), the associationClosterium–Type 128 may be characteristic ofmesotrophic stages in lake hydroseres of theregion. Type 119 and pollen of aquatic an-giosperms (e.g., Potamogeton, Myriophyllum)correlate to this association. Physico-chemicalprecipitation, in addition to biogenic, must havetaken place during the initial VM1 stage becausethose lime marls are not associated with charo-phyte-rich facies, and gyrogonites are almostabsent. Higher flow velocities would have pre-vented biofilm colonisation and Chara growth.
During short periods in VM2, and stagesVM4, VM6, and VM7, there must have beenincreased organic input associated with loweredwater levels. Increased turbidity would havediminished the reproductive capability of aquat-ic angiosperms and charophytes carpeting thebottom of the lake. Characteristic microfossilassemblages include Zygnematales, Rivularia,Types 182, 179, Glomus, and Polyadosporites.Together these palynomorphs suggest advancesof peat- and sapropel-producing marsh environ-ments, areas of temporary stagnant water and
lake eutrophication. In comparison with thesapropelic VM2 and VM4 zones, the peats VM6and VM7 appear to be formed under moreterrestrial conditions, as is further indicated bythe occurrence of Equisetum spores, Cyperace-ae, Pseudoschizaea cysts, Polyadosporites, Glomus,and spore Types 984 and 985.
While remaining clear that these biologicaland geological changes must be coupled withwater level variation, it is difficult to pinpointthe prime source of the hydrological eventssince there must have been alterations in localdrainage patterns caused by karst dynamics andgroundwater input. Similar karstic lake systemsshow heavily fissured bedrock and concentratedflows of groundwater. These landscapes mayindeed show great sensitivity to disturbance, andsmall-scale events can result in basin-widechange in regime (Goudie et al. 1993). Thiswould explain why there is no good correlationbetween limnological and pollen events (Fig. 7),and suggests that this basin is not a suitablemodel for multi-proxy palaeoclimate reconstruc-tion studies. In any case, there might be arelationship between the last millennia peat hu-mification and infilling, and the climate changesuggested by the pollen record through the in-crease of xerophytes.
San Rafael (Fig. 8)
A sediment core (depth 1900 cm) composed oforganic clays and silts was obtained from theeastern edge of a marshland (36∞20´N, 2∞12´W,10 m a.s.l.) in the Campo de Dalías, Almeríaprovince, the most arid region of Spain (Fig. 1).The pollen record covers the last ca. 17 000years (Fig. 8). It demonstrates that temperatetrees (e.g. deciduous Quercus, Betula, Corylus,Alnus, Fraxinus, Acer, Abies) and Mediterrane-an scrub (e.g. evergreen Quercus, Pistacia, Phil-lyrea, Myrtus, Chamaerops, Buxus) persisted insmall quantities in the southeastern littoral rang-es during full glacial times (SRA-SRB), with nopalynological evidence of xerophytization ofvegetation (Pantaleón-Cano 1997). In contrast,Artemisia reaches a first maximum from lategla-cial to early Holocene (SRC-SRD). As in theformer sequences, the optimum of deciduous

ANN. BOT. FENNICI Vol. 39 • Cryptogam spores and palaeoecology 11
trees and Mediterranean scrub (ca. 7500–4500 yr BP,SRE) occurs later than mesocratic early-Holo-cene phase of forest development in north-westEurope (Birks 1986), and coincides with thehighest percentage values of basin-associatedhygrophytes (Cyperaceae, Typha).
Percentage curves of Asteraceae, Pseudo-schizaea and trilete spores (mainly ferns) markthe boundaries of the mesophytic phase, thuscoinciding with important changes in the plantcover (Fig. 8). Pseudoschizaea increases havebeen shown to correlate with decreased pollenconcentrations and increased minerogenic input
within this and other regional sequences ob-tained from alluvial sediments (Pantaleón-Canoet al. 1996). In concordance with the highlyresistant trilete spores and composite pollen, theabundance of Pseudoschizaea cysts may be re-lated with increased oxydization and averagedesiccation of the basin, the alternation of wet-dry phases, or the prolongation of summerdrought (Scott 1992). The synchronous increas-es of Artemisia and Pseudoschizaea togetherwith mesophyte depletion after ca. 4500 yr BPcan be viewed as the consequence of enhancedregional aridity, which would have been critical-
Fig. 8. Palaeoecological sequence of San Rafael (Almería) (Pantaleón-Cano 1997). Ages in uncalibratedradiocarbon years. Zonation follows Grimm (1992).
�
���
���
���
���
���
���
���
���
��
����
����
����
����
����
����
����
����
����
���
��
����
������
����������� ����
!��"���#�� ����
$�% &���$
���'������ $
� �$� $
% '$
�����%��(�)��*����+,��#�-
� �����'�.$ $
���
���
��
��
���
���
��
�� �
��/ ��� �� �� �� ��� �� �� �� �� ������
$�% ���� $
��*)0+�1-
��1#�(�"��2( �02#"��3�"�)2)��# �02#"��
���� 4 ��
���� 4 ���
���� 4 ��
��� 4 ��
�� 4 ��
����� 4 ���

Carrión & Navarro • ANN. BOT. FENNICI Vol. 3912
ly manifested in the pollen record of Villaverdeat 4500, 3240, 2200, and 1700 yr BP (Fig. 5), inCañada de la Cruz after 3370 yr BP and1525 yr BP (Fig. 3), and Navarrés after3160 yr BP, when peatbog formation ceased(Fig. 6). Other palynological data from Mediter-ranean Spain coincide to suggest sub-humidforest declines after ca. 4500 yr BP (Dupré1988, Riera 1993, Burjachs et al. 1997, Yll et al.1997, Carrión et al. 1999, 2000a). Several pol-len records from elsewhere in the Mediterraneanshow progressive matorralization during the lastfour millennia at the expense of sub-humidforests (Bottema & van Zeist 1991, Willis 1994).The arid tendency is also supported by palae-oanthracological information relative to speciescomposition (Badal et al. 1994), and wood anat-omy (Terral & Arnold-Simard 1996), geomor-phological evidence of badlands (Wise et al.1982), and sedimentological evidence of chang-es in the hydrological regimes of lakes of tropi-cal and Mediterranean north Africa (Lamb et al.1995, Gasse 2000).
Concluding remarks
The occurrence of characteristic microfossil as-semblages of the Late Quaternary in lacustrinesystems of Mediterranean Spain has been shown.Regarding the water level, water temperature,and trophic stage, the alternation of stages domi-nated by Zygnemataceae zygospores and Clos-terium–Type 128 spores can be particularlymeaningful. Fungal types such as Polyadosporites,Fusiformisporites, Glomus, and Gelasinosporaare often abundant during the advances in mar-ginal vegetation and formation of peaty layers.Pseudoschizaea cysts occur in parallel with des-iccation phases or enhanced summer drought.
The correlation of these palynofacies withevents of vegetation change as reflected in thepollen record is a reliable indicator of broad-scale environmental change. The most strikingexample derives from the sequence of Cañadade la Cruz, where a sensitive response of Holo-cene vegetation correlates to palaeolimnologicalindicators. The sequence of Navarrés providesevidence of millennial-scale change in both veg-etation and the trophic conditions of a peat-
forming basin since the last glacial to the Holo-cene under the influence of fire events, climatechange and human activity. The sequence ofVillaverde shows certain discordance betweenvegetational and limnological developments, butthere are palynomorphs whose variation fits intothe overall scheme of climate control for bothsystems. The sequence of San Rafael offers anexample of synchronous variation of xerophyticpollen and microfossils indicative of temporarymarsh desiccation, which agrees with broaderevidence of aridification since the mid Holo-cene.
Acknowledgements
This research was funded by the Spanish Ministerio deEducación y Ciencia through the project ARDACHO(Ref. BOS2000-0149). A DGICYT fellowship (PR95-214) awarded to J. S. Carrión for research at the Hugo deVries Laboratorium during 1996 under the supervision ofDr. Bas van Geel was equally crucial. J. Pantaleón kindlyprovided the database relative to the site of San Rafael.
References
Badal, E., Bernabeu, J. & Vernet, J. L. 1994: Vegetationchanges and human action from the Neolithic to theBronze Age (7000–4000 B.P.) in Alicante, Spain,based on charcoal analysis. — Veg. Hist. Arch. 3:155–166.
Bennett, K. D. 1996: Determination of the number ofzones in biostratigraphical sequence. — New Phytol.132: 155–170.
Birks, J. H. B. 1986: Late Quaternary biotic changes interrestrial and lacustrine environments, with particu-lar reference to north-west Europe. — In: Berglund,B. E. (ed.), Handbook of Holocene palaeoecologyand palaeohydrology: 3–65. Chichester, Wiley.
Bottema, S. & van Zeist, W. 1991: Late Quaternaryvegetation of the Near East. — Beih. Tübinger AtlasDes Vorderen Orients, Reihe A (Naturwissench-schaften) 18: 1–156.
Burjachs, F., Giralt, S., Roca, J. R., Seret, G. & Julià, R.1997: Palinología holocénica y desertización en elMediterráneo occidental. — In: Ibáñez, J. J., Valero,B. L. & Machado, C. (eds.), El paisaje mediterráneoa través del espacio y del tiempo. Implicaciones en ladesertificación: 379–394. Logroño, Geoforma Edi-tores.
Carrión, J. S. & Dupré, M. 1997: Late Quaternary vegeta-tional history at Navarrés, eastern Spain. A two-coreapproach. — New Phytol. 134: 177–191.

ANN. BOT. FENNICI Vol. 39 • Cryptogam spores and palaeoecology 13
Carrión, J. S., Scott, L. & Vogel, J. 1999: Twentiethcentury changes in montane vegetation in the easternFree State, South Africa, derived from palynology ofhyrax dung middens. — J. Quat. Sci. 14: 1–16.
Carrión, J. S. & van Geel, B. 1999: Fine-resolution UpperWeichselian and Holocene palynological record fromNavarrés (Valencia, Spain) and a discussion aboutfactors of Mediterranean forest succession. — Rev.Palaeobot. Palynol. 106: 209–236.
Carrión, J. S., Andrade, A., Bennett, K. D., Munuera,M. & Navarro, C. 2001a: Crossing forest thresh-olds: inertia and collapse in a Holocene pollensequence from south-central Spain. — Holocene11:635–653.
Carrión, J. S., Dupré, M., Fumanal, M. P. & Montes, R.1995: A palaeoenvironmental study in semi-arid south-eastern Spain: the palynological and sedimentologi-cal sequence at Perneras Cave (Lorca, Murcia). — J.Arch. Sci. 22: 355–367.
Carrión, J. S., Munuera, M., Dupré, M. & Andrade, A.2001b: Abrupt vegetation changes in the SeguraMountains of southern Spain throughout the Holo-cene. — J. Ecol. 89: 783–797.
Carrión, J. S., Munuera, M., Navarro, C. & Sáez, F.2000a: Paleoclimas e historia de la vegetación cua-ternaria en España a través del análisis polínico.Viejas falacias y nuevos paradigmas. — Complutum11: 115–142.
Carrión, J. S., Scott, L., Huffman, T. & Dreyer, C. 2000b:Pollen analysis of Iron Age cow dung in southernAfrica. — Veg. Hist. Arch. 9: 239–249.
Dupré, M. 1988: Palinología y paleoambiente. Nuevosdatos polínicos. Referencias. — Servicio de Investi-gación Prehistórica, Valencia. 188 pp.
Dupré, M., Carrión, J. S., Fumanal, M. P., La Roca, N.,Martínez-Gallego, J. & Usera, J. 1998: Evolution andpalaeoenvironmental study of a semiendorheic areain eastern Spain (Navarrés, Valencia). — Il Quater-nario 11: 95–105.
Elsik, W. C. 1980: The utility of fungal spores in margin-al marine strata of the late Cenozoic, northern Gulf ofMexico. — IV Internat. Palynol. Conference, Luc-know 2: 436–443.
Elsik, W. C. 1983: Annotated glossary of fungal paly-nomorphs. — AASP Contr. Series 11: 1–36.
Gasse, F. 2000: Hydrological changes in the Africantropics since the last glacial maximum. — Quat. Sci.Rev. 19: 189–211.
Goudie, A. S., Viles, H. A. & Pentecost, A. 1993: Thelate-Holocene tufa decline in Europe. — Holocene 3:181–186.
Grimm, E. 1992: Tilia version 1.12. — Illinois StateMus., Res. Coll. Center, Springfield, USA.
Haas, J. N. 1994: First identification key for charophyteoospores from central Europe. — Europ. J. Phycol.29: 227–235.
Jarzen, D. M. & Elsik, W. C. 1986: Fungal palynomorphsrecovered from recent river deposits, Luangwa Val-
ley, Zambia. — Palynology 10: 35–60.Krutzsch, W. 1970: Atlas der mittel und jungtertiären
dispersen Sporen und Pollen sowie der Mikroplank-tonformen des nördlichen Mitteleuropas. — Fischer,Jena. 149 pp.
Lamb, H. F., Gasse, F., Benkaddour, A., El Hamouti, N.,van der Kaars, S., Perkins, W. T., Pearce, N. J. &Roberts, C. N. 1995: Relation between century-scaleHolocene arid intervals in tropical and temperatezones. — Nature 373: 134–137.
Lundqvist, N. 1972: Nordic Sordariaceae s. lat. — Symb.Bot. 20: 332–335.
Mateus, J. E. 1992: Holocene and present-day ecosys-tems of the Carvalhal Region, southwest Portugal. —Ph.D. thesis, Univ. Utrecht. 184 pp.
Mehringer, P. J., Arno, S. F. & Petersen, K. L. 1977:Postglacial history of Lost Trail Pass Bog, BitterrootMountains, Montana. — Arctic Alp. Res. 9: 345–368.
Moore, P. D. 1986: Hydrological changes in mires. — In:Berglund, B. E. (ed.), Handbook of Holocene palae-oecology and palaeohydrology: 91–107. Chichester,Wiley.
Moore, P. D., Webb, J. A. & Collinson, M. E. 1991:Pollen analysis. 2nd ed. — Blackwell Sci. Publ.,Oxford. 216 pp.
Navarro, C., Carrión, J. S., Navarro, J., Munuera, M. &Prieto, A. R. 2000: An experimental approach to thepalynology of cave deposits. — J. Quat. Sci. 15:603–619.
Pals, J. P., van Geel, B. & Delfos, A. 1980: Paleoecologi-cal studies in the Klokkeweel Bog near Hoogkarspel(Prov. of Noord-Holland). — Rev. Palaeobot. Paly-nol. 30: 371–418.
Pantaleón-Cano, J. 1997: Estudi palinològic de sedimentslitorals de la provincia d´Almería. Transformacionsdel paisatge vegetal dins un territori semiàrid. —Ph.D. thesis, Universidad Autónoma de Barcelona.189 pp.
Pantaleón-Cano, J., Yll, E., Pérez-Obiol, R. & Roure, J.M. 1996: Las concentraciones polínicas en mediossemiáridos. Su importancia en la interpretación de laevolución del paisaje. — In: Ramil-Rego, P., Fernán-dez, C. & Rodríguez, R. (eds.), Biogeografía pleisto-cena-holocena de la Península Ibérica: 215–226.Xunta de Galicia.
Riera, S. 1993: Changements de la composition forestièredans la plaine de Barcelone pendant l’Holocene(littoral méditerranéen de la Péninsule Iberique). —Palynosciences 2: 133–146.
Scott, L. 1992: Environmental implications and origin ofmicroscopic Pseudoschizaea Thiergart and Franz exR. Potonié emend. in sediments. — J. Biogeogr. 19:349–354.
Soulié-Märsche, I. 1993: Diversity of Quaternary aquaticenvironments in Africa as shown by fossil charo-phytes. — In: Thorweine, U. & Schandelmeier, H.(eds.), Geoscientific research in northeast Africa:575–579. Balkema, Rotterdam.

Carrión & Navarro • ANN. BOT. FENNICI Vol. 3914
Strother, P. K. 1996: Acritarchs. — In: Jansonius, J. &McGregor, D. C. (eds.), Palynology: principles andapplications 1: 81–106. Amer. Ass. Stratigr. Palynol-ogists Found., Texas.
Terral, J. F. & Arnold-Simard, G. 1996: Beginnings ofolive cultivation in eastern Spain in relation to Holo-cene bioclimatic changes. — Quat. Res. 46: 176–85.
Valle, F., Gómez-Mercado, F., Mota, J. F. & Díaz de laGuardia, C. 1989: Parque Natural de Cazorla, Segu-ra y Las Villas. Guía botánico-ecológica. — Rueda,Madrid. 354 pp.
Van Geel, B. 1972: Palynology of a section from theraised peat bog “Wietmarscher Moor” with specialreference to fungal remains. — Acta Bot. Neerl. 21:261–284.
Van Geel, B. 1978: A palaeoecological study of Holo-cene peat bog sections in Germany and The Nether-lands, based on the analysis of pollen, spores andmacro- and microscopic remains of fungi, algae,cormophytes and animals. — Rev. Palaeobot. Paly-nol. 25: 1–120.
Van Geel, B., Bohncke, S. J. P. & Dee, H. 1981: Apalaeoecological study of an Upper Late Glacial andHolocene sequence from “De Borchert”, The Nether-
lands. — Rev. Palaeobot. Palynol. 31: 367–448.Van Geel, B., Hallewas, D. P. & Pals, J. P. 1983: A Late
Holocene deposit under the Westfriese Zeedijk nearEnkhuizen (Prov. of Noord-Holland, The Nether-lands): Palaeoecological and archaeological aspects.— Rev. Palaeobot. Palynol. 38: 269–335.
Van Geel, B., Coope, G. R. & Van der Hammen, T. 1989:Palaeoecology and stratigraphy of the Lateglacialtype section at Usselo (The Netherlands). — Rev.Palaeobot. Palynol. 60: 25–129.
Willis, K. J. 1994: The vegetational history of the Bal-kans. — Quat. Sci. Rev. 13: 769–788.
Wise, S. M., Thornes, J. B. & Gilman, A. 1982: How oldare the badlands? A case-study from south-east Spain.— In: Bryan, R. & Yair, A. (eds.), Badland geomor-phology and piping: 259–278. Geobooks, Norwich.
Wolowski, K. 1998: Taxonomic and environmental stud-ies on euglenophytes of the Kraków–Czestochowaupland (southern Poland). — Fragm. Flor. Geobot.6: 1–192.
Yll, E. I., Pérez-Obiol, R., Pantaleón-Cano, J. & Roure, J.M. 1997: Palynological evidence for climatic changeand human activity during the Holocene on Minorca(Balearic Islands). — Quat. Res. 48: 339–347.