cyanobacterial cp12 proteins journal research area most ... · biochemical processes and...
TRANSCRIPT

1
Cyanobacterial CP12 Proteins
Journal research area most appropriate for the paper: Bioenergetics and Photosynthesis or Biochemical Processes and Macromolecular Structures
Corresponding author: Cheryl Kerfeld
Mailing address: Department of Energy Joint Genome Institute, 2800 Mitchell Drive, Walnut Creek, CA 94598. Phone: (925) 296-5691. E-mail: [email protected]
Plant Physiology Preview. Published on November 27, 2012, as DOI:10.1104/pp.112.210542
Copyright 2012 by the American Society of Plant Biologists
www.plantphysiol.orgon July 12, 2018 - Published by Downloaded from Copyright © 2012 American Society of Plant Biologists. All rights reserved.

2
Comparative Analysis of 126 Cyanobacterial Genomes Reveals Evidence of Functional Diversity Among Homologs of the Redox-Regulated CP12 Protein
Desirée N. Stanleya,b, c, Christine A. Rainesd and Cheryl A. Kerfelda,b,e,*
a DOE Joint Genome Institute
b Department of Plant and Microbial Biology – University of California, Berkeley
c Present Address: Department of Biochemistry – University of California, San Francisco
d School of Biological Sciences, University of Essex, Wivenhoe Park, Colchester, UK
e Berkeley Synthetic Biology Institute
www.plantphysiol.orgon July 12, 2018 - Published by Downloaded from Copyright © 2012 American Society of Plant Biologists. All rights reserved.

3
Footnotes:
Financial support:
CAK and DNS acknowledge the support of the NSF (MCB 0851094 and EF1105897). This work of CAR was underpinned by BBSRC grant P19403 and the University of Essex Research Promotion Fund.
Corresponding author: Cheryl Kerfeld ([email protected])
www.plantphysiol.orgon July 12, 2018 - Published by Downloaded from Copyright © 2012 American Society of Plant Biologists. All rights reserved.

4
Abstract
CP12 is found almost universally among photosynthetic organisms where it plays a key role in
regulation of the Calvin cycle by forming a ternary complex with GAPDH (glyceraldehyde 3-
phosphate dehydrogenase) and PRK (phosphoribulokinase). Newly available genomic
sequence data for the phylum Cyanobacteria reveals a heretofore unobserved diversity in
cyanobacterial CP12 proteins. Cyanobacterial CP12 proteins can be classified into eight
different types based on primary structure features. Among these are CP12-CBS (cystathionine-
β-synthase) domain fusions. CBS domains are regulatory modules for a wide range of cellular
activities; many of these bind adenine nucleotides through a conserved motif that is also present
in the CBS domains fused to CP12. In addition, a survey of expression datasets shows that the
CP12 paralogs are differentially regulated. Furthermore, modeling of the cyanobacterial CP12
protein variants based on the recently available three-dimensional structure of the canonical
cyanobacterial CP12 in complex with GAPDH suggests that some of the newly identified
cyanobacterial CP12 types are unlikely to bind to GAPDH. Collectively these data show that, as
is becoming increasingly apparent for plant CP12 proteins, the role of CP12 in cyanobacteria is
likely more complex than previously appreciated, possibly involving other signals in addition to
light. Moreover, our findings substantiate the proposal that this small protein may have multiple
roles in photosynthetic organisms.
www.plantphysiol.orgon July 12, 2018 - Published by Downloaded from Copyright © 2012 American Society of Plant Biologists. All rights reserved.

5
INTRODUCTION
CP12 is a small (~80 amino acid) protein present in most photosynthetic organisms,
including cyanobacteria, diatoms, red and green algae, and higher plants (Pohlmeyer et al.
1996; Wedel and Soll 1998; Oesterhelt et al. 2007; Erales et al. 2008a; Groben et al. 2010). In
eukaryotic organisms CP12 is located in the chloroplast, where the only function thus far
identified is in the regulation of the Calvin cycle in response to changes in light availability by
reversibly binding glyceraldehyde-3-phosphate dehydrogenase (GAPDH) and, subsequently,
phosphoribulokinase (PRK) (Wedel et al. 1997; Graciet et al. 2003a; Graciet et al. 2003b; Marri
et al. 2005a; Erales et al. 2008b; Howard et al. 2008; Marri et al. 2008; Carmo-Silva et al. 2011).
The formation of the GAPDH-PRK-CP12 complex inhibits GAPDH and PRK activity leading to
down-regulation of the Calvin cycle; this has been demonstrated in plants, Chlamydomonas
reinhardtii and cyanobacteria (Wedel et al. 1997; Graciet et al. 2003b; Marri et al. 2005b; Tamoi
et al. 2005). The GAPDH-PRK-CP12 complex dissociates under reducing conditions mediated
by thioredoxin, thereby restoring GAPDH and PRK activity (Lebreton et al. 2003; Marri et al.
2005b; Howard et al. 2008; Marri et al. 2009). Erales et al. (2008b) showed that CP12 binds
aldolase, another Calvin cycle enzyme, implying involvement of CP12 in additional Calvin cycle
regulation. Further studies in higher plants have proposed a role for CP12 outside of Calvin
cycle regulation. CP12 is expressed in non-photosynthetic A. thaliana tissues, and anti-sense
suppression of tobacco CP12 disrupts a variety of developmental processes (Singh et al. 2008;
Howard et al. 2011a; Howard et al. 2011b). Moreover, higher plant genomes encode up to three
forms of CP12, which are differentially expressed, but all have been shown to bind GAPDH and
PRK (Singh et al. 2008; Marri et al. 2010).
In CP12 disulfide bonds between two pairs of cysteine residues create peptide loops. In
vitro analysis of Arabidopsis thaliana CP12 mutants has shown that both pairs of redox-
regulated cysteine residues are required for ternary complex formation: an N-terminal pair for
PRK binding and a C-terminal pair for GAPDH binding (Wedel et al. 1997). The red alga
Galdieria sulphuraria CP12 does not have an N-terminal cysteine pair and, although the
GAPDH-PRK-CP12 complex forms, PRK is not completely inactivated (Oesterhelt et al. 2007).
In the cyanobacterium Synechococcus elongatus PCC7942 (hereafter S. elongatus), the N-
terminal cysteine pair is not required for PRK binding (Tamoi et al. 2005). In the green alga C.
reinhardtii, CP12 and GAPDH have complex interactions centered on the cysteine pairs.
GAPDH can undergo a thiol-disulfide exchange reaction with CP12, cleaving the C-terminal
www.plantphysiol.orgon July 12, 2018 - Published by Downloaded from Copyright © 2012 American Society of Plant Biologists. All rights reserved.

6
disulfide bond (Erales et al. 2009a). CP12 has also been shown to regulate the Calvin cycle in a
redox-independent manner, acting as a specific chaperone for GAPDH to prevent thermal
inactivation and aggregation (Erales et al. 2009b). It is oxidized, but not reduced, CP12 that is
able to complex with GAPDH and PRK (Graciet et al. 2003b; Marri et al. 2009b). Oxidized
CP12 is composed of helix and coil segments and is flexible, whereas reduced CP12 is
unstructured (Graciet et al. 2003a). Interestingly, the bulk of CP12’s primary structure fits the
criteria for intrinsically disordered proteins (IDP). IDPs are conformationally very flexible, which
allows tight regulatory control of important processes, including the cell cycle, and
transcriptional and translational regulation (Wright and Dyson 1999; Tompa 2002). Likewise,
IDPs have been shown to “moonlight”, assuming multiple conformations upon binding their
partners, leading to distinct or even opposing actions (Tompa et al. 2005).
The importance of CP12 in cyanobacteria was demonstrated by analysis of a S.
elongatus insertion mutant which showed that, under both light and dark conditions, the
formation of the GAPDH-PRK-CP12 ternary complex regulated the activities of both enzymes
and thus carbon flow from the Calvin cycle to the oxidative pentose phosphate cycle (Tamoi et
al. 2005). More recently, it was shown that cyanophages of the marine picoplanktonic
cyanobacteria genera Synechococcus and Prochlorococcus encode and express a CP12
protein similar to that of their hosts (Thompson et al. 2011). Cyanophage CP12 expression
results in decreased Calvin cycle activity in infected hosts, leading to an increased level of
NADPH; this is proposed to fuel the phage ribonucleotide reductase for production of phage
nucleotides. Together, these data underscore the key role of CP12 in the regulation of the
Calvin cycle in both redox-dependent and -independent mechanisms in cyanobacteria.
Recently, structures have been determined of both the S. elongatus and A. thaliana
CP12 in complex with GAPDH (Matsumura et al. 2011; Femarni et al. 2012). These structures
provide unprecedented detail of the intermolecular interactions between CP12 and GAPDH and
suggest how PRK may be recruited to the complex. The C-terminal domain of CP12 (residues
53-75 of the S. elongatus CP12) inserts into the active site of GAPDH, while the N-terminus of
CP12 (residues 1-52) remains disordered in the CP12-GAPDH complex and, presumably,
interacts with PRK. In S. elongatus, CP12 binding to GAPDH creates a patch of negative charge
on the binary complex that is proposed to electrostatically attract positive charges on PRK
(Matsumura et al. 2011). However, this may be a feature specific to cyanobacterial CP12
because this negatively charged patch is not present in the A. thaliana structure (Fermani et al.
2012).
www.plantphysiol.orgon July 12, 2018 - Published by Downloaded from Copyright © 2012 American Society of Plant Biologists. All rights reserved.

7
All cyanobacterial genomes sequenced to-date (75 publically available) contain at least
one gene encoding a CP12 or CP12-like protein (except Cyanobacterium sp. UCYN-A and
Moorea producta 3L). Recently, the amount of sequence data available for cyanobacteria more
than doubled as a result of the CyanoGEBA project (Submitted data Shih PM, Wu D, Latifi A,
Axen SD, Fewer DP, Talla E, Calteau A, Cai F, de Marsac T, Rippka R, Herdman M, Sivonen K,
Coursin T, Laurent T, Goodwin L, Nolan M, Davenport KW, Han CS, Rubin EM, Eisen JA,
Woyke T, Gugger M, Kerfeld CA). We report here on the bioinformatic analysis of a large
number of newly available cyanobacterial genome sequences, including members of
Subsections II and V that previously had no sequenced representatives. This analysis has
identified a previously unknown diversity of CP12 homologs among cyanobacteria indicating a
diversity of roles for these proteins, including possibly some outside the Calvin cycle.
RESULTS
Classification of Cyanobacterial CP12 Types and Comparison of Primary Structure
A bioinformatic analysis of 126 cyanobacterial genomes representing the five
morphological subsections revealed that all but UCYN-A and Moorea producta 3L contained
between one and five CP12 paralogs. A total of 274 CP12 genes were identified (Table S1).
They were subdivided into eight groups based on key primary structural features: the N-terminal
cysteine pair, the C-terminal cysteine pair, a core “AWD_VEEL” sequence, and an N-terminal
CBS domain fusion (Fig. 1). Of the 274 cyanobacterial sequences analyzed, 120 resemble the
canonical higher plant-like CP-12 (CP12-N/C; Fig. 2A): CP12-N/C proteins are approximately 80
amino acids long and contain both the N-terminal cysteine pair associated with PRK binding and
the C-terminal cysteine pair required for GAPDH binding. Other types of cyanobacterial CP12s
include a variant lacking both cysteine pairs (CP12-0; eight sequences), CP12 containing only
the N-terminal cysteine pair (CP12-N; two sequences) and CP12 with only the C-terminal
cysteine pair (CP12-C; 53 sequences). Furthermore, the CP12-C group contains a subset of 39
sequences, found only in marine picoplanktonic cyanobacteria, that lack the otherwise
conserved core CP12 “AWD_VEEL” sequence (CP12-C-M; residues 66-73 in CP12-N/C logo).
This core sequence is implicated in GAPDH binding by trypsin-protection experiments (Lebreton
et al. 2006); however, this region of CP12 is not visible in the electron density of either of the
CP12-GAPDH structures (Matsumura et al. 2011; Fermani et al. 2012). Additionally, the N-
terminal Gly24, Ser27, and His46, and C-terminal Pro65 residues are strongly conserved
among all CP12 homologs (including those from eukaryotes), while the Gln41 and Phe57
www.plantphysiol.orgon July 12, 2018 - Published by Downloaded from Copyright © 2012 American Society of Plant Biologists. All rights reserved.

8
residues following the “AWD_VEEL” sequence that have previously been noted as specific to
cyanobacteria (Groben et al. 2010) are not strongly conserved among all cyanobacterial
variants (Fig 2A).
In contrast to any known plant or algal CP12 homologs, a large (92) subset of
cyanobacterial CP12 homologs have a CBS domain fused to the N-terminus (Figs 1 and 2A).
These make up three additional classes of cyanobacterial CP12-like proteins: CP12-N/C-CBS
(14 sequences), CP12-N-CBS (55 sequences), CP12-0-CBS (23 sequences). One of the CP12-
N-CBS proteins was previously described as an ORF4/CP12 fusion in Anabaena variabilis
(Pohlmeyer et al. 1996; Masepohl et al. 1997). As discussed below, CBS domains are typically
associated with regulatory functions and are found in all phylogenetic domains of life (Bateman
1997).
Phylogenetic Distribution of CP12 Types
Phylogenetic analysis of the eight different CP12 types revealed that the distribution of
any individual variant differs among cyanobacterial genomes (Fig. 4). Some genomes have
multiple copies of a given CP12 variant (e.g. Synechococcus sp. PCC 7335 has two copies of
CP12-0 and of CP12-N-CBS). The entire group of marine picoplanktonic cyanobacteria only
possess the CP12-C(M) variant (lacking the “AWD_VEEL” core sequence), occasionally in
multiple copies. With the exception of the marine picoplanktonic cyanobacteria, every genome
in Figure 4 contains at least one copy of the canonical CP12-N/C. A phylogenetic tree of the 274
cyanobacterial CP12 sequences shows each CP12 type clustering in dispersed groups (Fig. 5,
Fig. S2). The primordial cyanobacterium, Gloeobacter violaceus, contains a single canonical
CP12-N/C gene which clusters with the anomalous CP12-C(M)s. The plant and algal eukaryotic
CP12 sequences form two clades that group with the cyanobacterial canonical CP12-N/C type
(Fig. S3). The plant CP12-3 variant forms a cluster distal from the plant CP12-1 and -2 types
and is closer to a distinct group of cyanobacterial CP12-N/Cs.
The remaining cyanobacterial CP12 types are present in clusters of varying sizes. The
lack of clustering of the CP12 variants from any one genome suggests that the duplication,
divergence, and fusion events that gave rise to the different types are very ancient. Recent
duplication events are rare but do seem to have occurred in cases where a genome contains
more than one copy of a CP12 type (for example, the two CP12-N-CBS of Cyanothece sp.
ATCC 51472 appear to be the result of a recent duplication (Fig. S2)).
www.plantphysiol.orgon July 12, 2018 - Published by Downloaded from Copyright © 2012 American Society of Plant Biologists. All rights reserved.

9
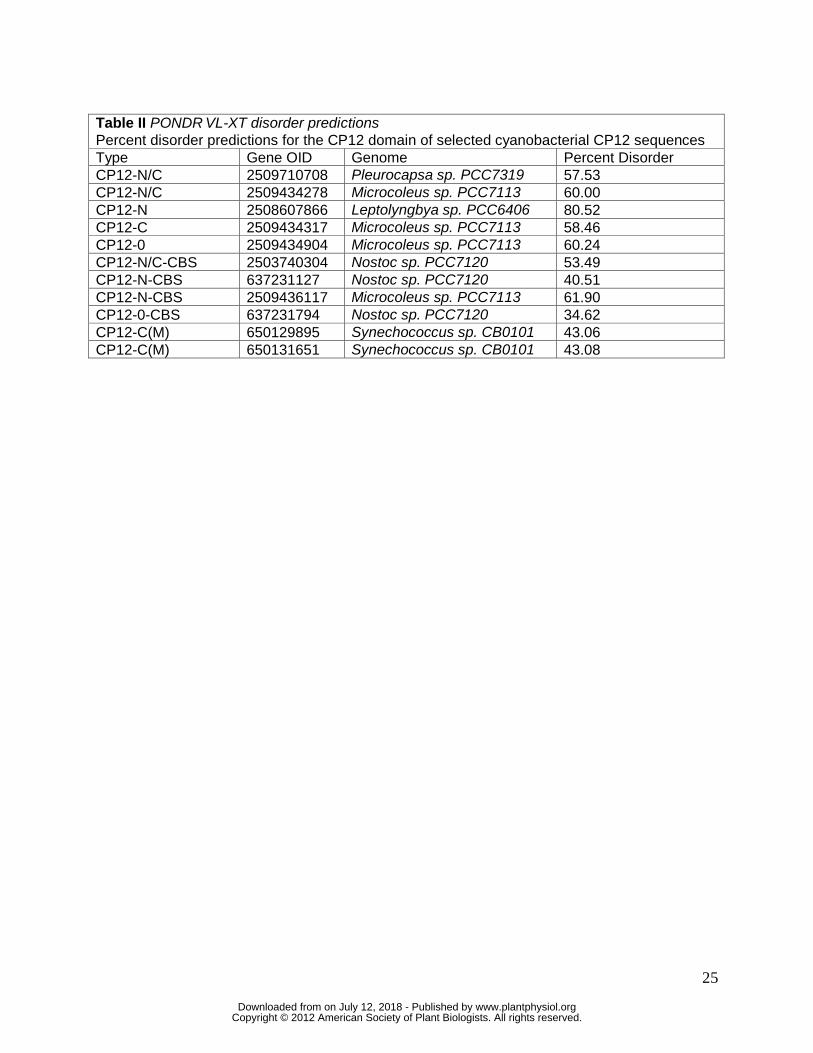
The Predicted Intrinsic Disorder of the Cyanobacterial CP12 Proteins
The majority of CP12 proteins from all photosynthetic organisms are predicted to be
more than 50% disordered, with a few exceptions found in land plants (Marri et al. 2008). A
PONDR VL-XT analysis of the CP12 proteins from five representative cyanobacterial genomes
predicted the expected high level of disorder (ranging from 57.53% to 80.52%) particularly in the
N-terminus of CP12-0, CP12-C, CP12-N and CP12-N/C proteins (Table II, Fig. S1). CP12-C(M)
displayed the same pattern of highest disorder at the N-terminus but the overall predicted
disorder was somewhat lower at 43%. The CP12 domain of the CP12-CBS fusion proteins is
also predicted to be generally less disordered than CP12 proteins lacking the fusion, ranging
from 40.51% to 61.90%.
The Cyanobacterial CP12 Variants in the Context of the CP12-GAPDH Structure
By a combination of sequence comparison and homology modeling using the
coordinates of the S. elongatus CP12-C model from the crystal structure as a template, we
evaluated the potential of the different cyanobacterial CP12 types to form a complex with their
cognate GAPDH, coordinate copper and bind NAD (Matsumura et al. 2011; PDB 3B1J).
Two types of GAPDH are present in cyanobacteria: type 1 contains the residues
identified by Matsumura et al. 2011 as being important for CP12 binding (Thr37, Ser38, Asp39,
Arg80, His182, Arg189, Ser194, His195, Arg196 and the copper-interacting Cys155, Thr156,
His182). A second type (type 2) of GAPDH is also found in cyanobacteria; this paralog contains
the copper-interacting residues, but not the other CP12-binding residues. Each cyanobacterial
genome encodes one copy of type 1 GAPDH and one or two copies of type 2 with two
exceptions: Cyanothece sp. BH68, ATCC 51142 has two type 2 GAPDHs (without the CP12
binding residues) and no type 1 GAPDH and Acaryochloris sp CCMEE 5410 has two of each
GAPDH paralog. There is no apparent correlation between the number of CP12 variants and
GAPDH type or copy number within a given genome.
Homology models for all of the cyanobacterial CP12 variants and the corresponding (i.e.
from the same organism) type1 GAPDH were built, except for CP12-0-CBS (the low sequence
homology between it and the template precluded construction of a homology model). Inter-
protein interactions between the CP12 variant and GAPDH models were examined (Fig. 3).
The CP12-N/C, CP12-C, CP12-C(M), and CP12-C/N-CBS (the variants with the C-
terminal cysteine pair) all have the C-terminal loop residues required for interaction with
www.plantphysiol.orgon July 12, 2018 - Published by Downloaded from Copyright © 2012 American Society of Plant Biologists. All rights reserved.

10
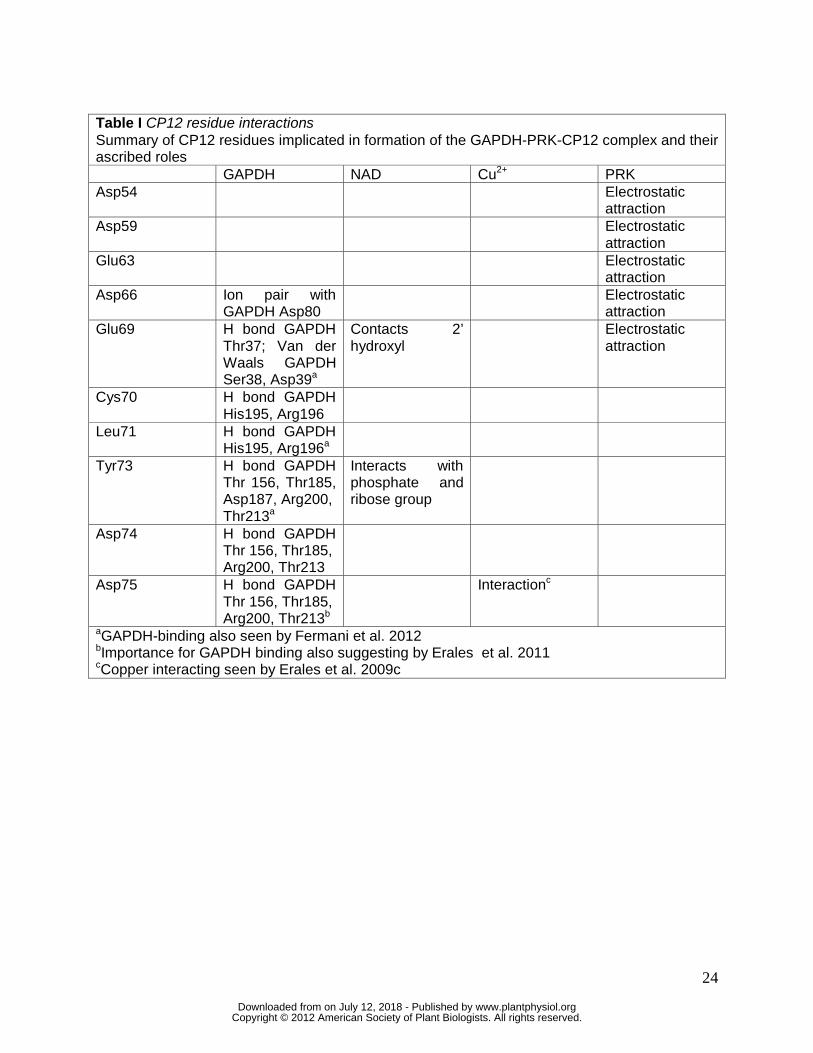
GAPDH, copper and NAD+ (Tyr 73, Asp74, Asp75; Table I). Homology models of
representatives of each of these types show some conformational variation in the disposition of
this three-residue C-terminal loop relative to the solved structure (Fig. 3). At the level of primary
structure, Asp74 and Asp75 are conserved, with a Glu occasionally substituted among some
variants (Fig. 2A). The position of Tyr73 is the most disparate among the models, but flexibility
in the C-terminal loop would likely accommodate positioning compatible with the Tyr73 and
Asp75 NAD and copper interaction. However, modeling with S. elongatus’ other CP12 paralog,
a CP12-N/C, shows substantial differences in the interaction of the C-terminus with GAPDH:
Tyr73 clashes with GAPDH Glu185, and Asp75 is replaced by a Val, suggesting that this
paralog interacts with GAPDH very differently or not at all.
CP12-N, CP12-0 and CP12-N-CBS lack the C-terminal cysteine pair (Fig. 2A) and
therefore the disulfide bond pinning the alpha helix (the ordered section of CP12), in place. In
addition, these variants are missing at least one key interacting C-terminus residue, with the
negative Asp75 most often replaced by a neutral Val (Fig. 2A). Homology modeling shows that
Tyr73 in both CP12-0 and CP12-N-CBS sterically clashes with GAPDH (both at Glu187).
Additionally, CP12-N-CBS Leu75 clashes with GAPDH Thr213 (Fig. 3). For CP12-N, two
residues clash with GAPDH: CP12-N Tyr73 and GAPDH Arg189, and CP12-N Arg71 and
GAPDH Leu186.
DISCUSSION
This study has revealed a previously unknown range of variation among cyanobacterial
CP12 proteins. These data provide strong evidence for a more diverse regulatory role for CP12
and CP12-like proteins in cyanobacteria - in either input signal or output activity - than
previously thought. Direct support for this comes from analysis of a S. elongatus CP12-C knock-
out mutant that showed inhibited growth under dark/light conditions (Tamoi et al. 2005),
suggesting that the CP12 variant (CP12-N/C) present in this organism could not compensate for
the loss of CP12-C.
Consistent with the growing evidence for a multiplicity of roles for CP12, a survey of
numerous expression studies indicates that the CP12 variants we have identified here are
differentially expressed in cyanobacteria (Table S2). For example, the two S. elongatus CP12
proteins, CP12-C and CP12-N/C, in addition to differences in potential interaction with GAPDH
(Fig. 3) show differences in overall expression under constant light and in response to a shift
from high to low atmospheric CO2 (Vijayan et al. 2011; Scharwz et al. 2011). In Synechocystis
www.plantphysiol.orgon July 12, 2018 - Published by Downloaded from Copyright © 2012 American Society of Plant Biologists. All rights reserved.

11
PCC6803, CP12-C expression is upregulated under sulfate deprivation and osmotic stress
conditions and down-regulated in high light (Table S2; Hihara et al. 2001; Shoumskaya et al.
2005; Zhang et al. 2008). In Synechococcus sp. PCC 7002, the ratio of expression of CP12-N-
CBS under dark oxic conditions relative to standard growth conditions is four-fold higher than
that of CP12-N/C (Ludwig and Bryant 2011). Interestingly, in Microcystis aeruginosa PCC7806
mutants lacking the potential to synthesize the toxin microcystin, two of M. aeruginosa’s four
CP12 proteins show differential accumulation: one of its two CP12-N-CBS proteins accumulates
at three-fold the rate of wildtype under light conditions, and its CP12-N/C-CBS accumulates at a
six-fold high rate than wild type under light conditions and three-fold lower rate in the dark
(Zilliges et al. 2011). This study also shows that microcystin plays a role in resistance to
oxidative stress, possibly in conjunction with CP12. In higher plants, the expression of the
CP12-3 like variant is increased under hypoxic conditions and this may indicate a role for this
form of the protein in mediating the shift in metabolism from aerobic to anaerobic (Singh et al.
2008). A microarray analysis of CP12 antisense plants revealed that in roots the most strongly
induced genes were those shown to be involved in hypoxia responses in plants (Liu et al. 2005;
Howard et al. 2011a).
Evidence of differential Interaction of Cyanobacterial CP12 Variants GAPDH and PRK
Previous analysis has shown that the C-terminal loop of CP12 is critical for the
interaction with GAPDH (Matsumura et al. 2011; Erales et al. 2011; Fermani et al. 2012). Glu69
is thought to be important in interactions with PRK, GAPDH and NAD and is particularly well
conserved among cyanobacterial CP12 proteins (Matsumura et al. 2011). Tyr73, which also
interacts with NAD, is conserved among cyanobacterial CP12 variants while Leu71 is not (Fig.
3). Asp75 is conserved in CP12-N/C, CP12-N, CP12-C(M), and CP12-N/C-CBS (Fig. 3). The
presence of the copper-interacting residue in variants with the C-terminal cysteine pairs is
consistent with the proposal that copper binding catalyzes the formation of the disulfide bonds
(Delobel et al. 2005; Erales et al. 2009c). In contrast, the copper-interacting Asp75 is replaced
by Leu (CP12-N-CBS) or by a Val (CP12-0) in CP12 types that do not contain the C-terminal
cysteine pair. Collectively, our survey of residue conservation (Fig. 2) and homology modeling
(Fig. 3) suggests that cyanobacterial CP12-C, CP12-C(M), CP12-N/C and CP12-N/C-CBS could
likely bind GAPDH. However, it is unlikely that CP12-0, CP12-N, CP12-N-CBS and CP12-0-
CBS are able to bind GAPDH, in part because the structure of each of these may be
significantly different from that of the canonical CP12 due to the lack of the disulfide bond that
orients the helical segment (Fig. 1, Fig. 2).
www.plantphysiol.orgon July 12, 2018 - Published by Downloaded from Copyright © 2012 American Society of Plant Biologists. All rights reserved.

12
In contrast to GAPDH, identifying PRK paralogs in cyanobacterial genomes is difficult.
PRK is very similar to another enzyme, uridine kinase, and annotations, COG and pfam
assignments frequently contradict one another, suggesting that such computational-based
functional assignments for this gene product are unreliable. However, it is clear that among the
CP12 variants, the N-terminus, which has been shown to interact with PRK (Groben et al.
2010), is overall less conserved than the GAPDH-interacting C-terminus (Fig. 2A). The CP12-
N/C group contains a conserved N-terminal segment preceding N-terminal cysteine pair that is
not found in the other cyanobacterial CP12 types (Fig. 2A). The function of the N-terminus is
elusive and the N-terminal cysteine pair is not essential for PRK binding in S. elongatus (Tamoi
et al. 2005).
When CP12 is bound to GAPDH, five acidic residues face outward, creating a patch of
negative charge proposed to interact with positively charged residues in PRK (Matsumura et al.
2011). Among the CP12 variants identified here, four of the negatively charged residues are
conserved; the fifth, Asp54, is frequently replaced by a Lys in CP12-N/C and in all three CP12-
CBS variants (Fig. 2A). Changing this final position of the helix from a negative to positively
charged residue will reduce the strength of the negatively charged patch posited to interact with
PRK. The prevalence of this substitution across the CP12-CBS fusion variants suggests that
these proteins have a role other than formation of a CP12-GAPDH-PRK complex.
The Role of the CP12-Associated CBS Domain Pair
An additional layer of complexity in the regulatory role of CP12 proteins is evident from
the widespread occurrence of the three types of CP12-CBS domain fusions. Each type contains
a single CBS domain. CBS domains are regulatory modules usually in two or four tandem
copies per protein. They are found in functionally diverse proteins across all kingdoms of life
including channel proteins, kinases, signal transduction proteins, and membrane proteins
(Bateman 1997). The cyanobacterial genomes that contain the CP12-CBS fusions tend to be
enriched in CBS domains (Table IV). All cyanobacterial genomes sequenced to-date contain
between 5 and 30 CBS domain-containing proteins; by comparison E. coli contains only 9 CBS
domain-containing proteins. Among CBS domain sequences available in the NCBI database,
the cyanobacterial CP12 CBS domain is most similar to that found in a signal transduction
protein in Thermus sp. CCB_US3_UF1 (BLAST, E-value 3e-66, 78% identity).
CBS domain pairs assemble in an anti-parallel arrangement to form what is known as a
tight Bateman module (Ignoul and Eggermount 2005). CBS domain pair-containing proteins
www.plantphysiol.orgon July 12, 2018 - Published by Downloaded from Copyright © 2012 American Society of Plant Biologists. All rights reserved.

13
often form homodimers, with two CBS domain pairs assembling into a CBS module with a total
of four CBS domains (Baykov et al. 2011). A key question raised by our observations is whether
the CP12 CBS domain facilitates homodimerization of two CP12-CBS proteins, or whether the
CBS domain in a given CP12 dimerizes with a CBS domain found on some other protein. The
CBS domains found in the three CP12 variants are present in single copies at the N-terminus
and share high degree of similarity, with a typical pairwise alignment between their CBS
domains being 70 to 75% identical. In order to identify prospective interaction partners (besides
via homodimerization) for the CP12-associated CBS domains, phylogenetic profiling was used
to search for genes conserved specifically among the subset of cyanobacterial genomes
containing a CP12-CBS domain fusion. No candidate genes could be identified that were
restricted to genomes containing CP12-0-CBS, CP12-N-CBS, CP12-N/C-CBS or any
combination of the three types. Searching any one genome using the CBS domain sequence
from a CBS-CP12 fusion encoded in that genome finds the most significant non-CP12 CBS hit
to be a predicted signal transduction protein annotated as insine-5-monophosphate
dehydrogenase-related with an aldolase-type TIM barrel fold (seen in Fischerella PCC9605, E-
value of 4e-06, 26% identity, Rivularia sp. PCC 7116, E-value of 2e-06, 32% identity and Nostoc
PCC7524 E-value of 1e-07, 26% identity). The relatively low sequence homology between the
CP12 CBS domain and any other CBS domain containing protein within a given genome
suggests that the CP12 CBS domain most likely homodimerizes with a second copy of a CP12-
CBS protein.
CBS domains have been shown to bind Mg2+, single-stranded DNA and RNA, and
double stranded DNA (Kery et al. 1998; McLean et al. 2004; Scott et al. 2004; Sharpe et al.
2008; Hattori et al. 2007; Hattori et al. 2009; Aguado-Llera et al. 2010; Feng et al. 2010). A
potential clue to the function of the CP12 CBS domain is found in the primary structure of all
three types of CP12-CBS proteins. Most of the CBS domains characterized to-date bind
adenine nucleotides as regulatory elements for sensing cellular energy status, and the result
can be either inhibitory or activating. This may be the role of the CP12 CBS domains; each
contains two copies of the nucleotide ribose-PO4 binding motif Ghx(T/S)x(T/S)D (Day et al.
2007; Fig. 2).
A total of 34 single and double CBS domain-containing proteins are encoded in the
genomes of Arabidopsis thaliana and Oryza salitica (Kushwaha et al. 2009), but to-date there is
no evidence for a CP12-CBS fusion among eukaryotic CP12 homologs. However, a recent
study of chloroplast CBS proteins in higher plants suggests that they are involved in activation
www.plantphysiol.orgon July 12, 2018 - Published by Downloaded from Copyright © 2012 American Society of Plant Biologists. All rights reserved.

14
of thioredoxins in the ferredoxin-thioredoxin system and that binding AMP in the chloroplast
increases this activation, thereby enabling these molecules to exert a regulatory effect on both
the Calvin cycle and H2O2 levels (Yoo et al. 2011). This is interesting because, as in higher
plants, the formation and breakdown of the PRK-GAPDH complex is mediated by the redox
state of CP12, which is in turned mediated by thioredoxin (Howard et al. 2008; Marri et al.
2009).
Higher plants contain a CP12-GAPDH fusion; the fused GAPDH subunit is known as
GapB. The C-terminal extension (CTE) of GapB consists of the GAPDH-binding module of
CP12 (~30 residues from the C-terminal half of CP12), including the C-terminal cysteine pair.
All cyanobacteria, algae and plants contain GAPDH composed of four GapA subunits, but in
higher plants an A2B2 heterotetramer, composed of two GapA and two GapB subunits, is the
major chloroplast form of GAPDH (Petersen et al. 2006; Howard et al. 2011b, Howard et al.
2011c). GapA and GapB form complexes in response to light, with the CTE playing the
regulatory role of CP12 (Wedel and Soll 1998; Scheibe et al. 2002; Trost et al. 2006). Outside of
the Streptophyta, GapB has only been found in the green alga Ostreococcus: the Ostreococcus
genome does not encode any other CP12, suggesting that the CTE function can replace CP12
redox-regulated activity (Robbens et al. 2007).
CONCLUSION
This study has revealed an unexpected diversity in cyanobacterial CP12 proteins; they
can be classified into 8 distinct types based on primary structural features. The presence of
CBS domain fusions as well as variation in the distribution of key structural features and in
expression patterns among these types, suggest a heretofore unknown layer of regulatory
complexity in CP12 function. The CP12-CBS domain fusions in cyanobacteria functionally
connect CBS domains and CP12. This finding raises the question of whether CBS proteins in
plants, together with CP12, play a redox relay type role acting as metabolic switches. In
cyanobacteria the diversity of CP12 proteins and the CBS-CP12 fusions may have evolved to
cope with the need to trigger different metabolic processes in response to rapidly fluctuating
environments. Perhaps as in other intrinsically disordered proteins (Tompa et al. 2005), some
CP12 variants may “moonlight,” interacting with multiple proteins in different conformations for
multiple functions.
www.plantphysiol.orgon July 12, 2018 - Published by Downloaded from Copyright © 2012 American Society of Plant Biologists. All rights reserved.

15
MATERIALS AND METHODS
Bioinformatics
The Function Search tool in IMG ER (http://img.jgi.doe.gov) was used to survey all
available cyanobacterial genomes for the CP12 pfam (pfam02672). The “blastp” program at the
National Center for Biotechnology website (Altschul et al. 1990; Altschul et al. 1997;
http://www.ncbi.nlm.nih.gov/), was also used to verify the results, using the Synechococcus
PCC6801 and Synechococcus PCC7942 CP12 amino acid sequences as queries.
All CP12 amino acid sequences were aligned using the MUltiple Sequence Comparison
by Log-Expectation (MUSCLE; Edgar 2004). The multiple sequence editor JalView (Waterhouse
et al. 2009) was used to manually curate the alignment and categorize sequences into types
based on primary structure features: the presence or absence of an N-terminal cysteine pair, C-
terminal cysteine pair, a core “AWD_VEEL” sequence, and an N-terminal CBS domain fusion.
Using the MUSCLE alignments, Hidden Markov Model profiles for each variant were
built using hmmer3.0:HMMBUILD (Eddy 1998; Eddy 2008; http://mobyle.pasteur.fr), with the
Henikoff position-based weighting scheme and no effective sequence weighting (enone).
LogoMat-M (Schuster-Böckler et al. 2004) was used to visualize Hidden Markov Model logos for
each variant (http://www.sanger.ac.uk, X-axis = contribution, Y-axis = relative entropy).
The PONDR (Predictor of Naturally Disordered Proteins) server (Romero et al. 1997; Li
et al. 1999, Romero et al. 2001; www.pondr.com) was used to predict the degree of disorder in
the following CP12 proteins: geneOID 2509710708, 2509434278, 2508607866, 2509434317,
2509434904, 2503740304, 637231127, 2509436117, 637231794, 650129895, and 650131651
(Table S1).
The CP12 CBS domain consensus sequence was obtained from loading the MUSCLE
alignment of all cyanobacterial CP12 sequences into JalView (Waterhouse et al. 2009). This
consensus sequence was used as a query to search the Research Collaboratory for Structural
Bioinformatics Protein Data Bank (RCSB PDB) (www.pdb.org; Berman et al. 2000). The CP12
CBS domain consensus sequence was entered into the “blastp” program at the National Center
for Biotechnology website (http://www.ncbi.nlm.nih.gov/), searching the non-redundant protein
database excluding cyanobacterial genomes (Altschul et al. 1990; Altschul et al. 1997). The
www.plantphysiol.orgon July 12, 2018 - Published by Downloaded from Copyright © 2012 American Society of Plant Biologists. All rights reserved.

16
CBS domain from the following genes was used as a query sequence to search its own genome
using the IMG BLAST tool at IMG ER (http://img.jgi.doe.gov): Fischerella PCC9605, geneOID
2509464408 ; Rivularia sp. PCC 7116, geneOID 2510088188; Nostoc PCC7524, geneOID
2509809301.
Phylogenetic profiles were constructed using the “Phylogenetic Profiler: Single Gene”
function in IMG ER (http://img.jgi.doe.gov). Searches were conducted for genes unique to
cyanobacterial genomes possessing all three variants of CP12-CBS fusion proteins, any two
variants of CP12-CBS fusion proteins, or any single variant of CP12-CBS fusion protein.
Cyanobacterial GAPDH and PRK sequences were obtained by using the
Synechococcus PCC7942 GAPDH and PRK amino acid sequences as queries in the IMG
BLAST tool in IMG ER (http://img.jgi.doe.gov; Table S3). The Function Search tool in IMG ER
was also used to search for the GAPDH COG0057.
A phylogenetic tree was constructed of all cyanobacterial CP12s by loading the core
CP12 section of the MUSCLE alignment (the final 84 amino acids) into the “A la carte” analysis
on at www.phylogeny.fr (Dereeper et al. 2008; Dereeper et al. 2010). The alignment was
curated to remove positions with gaps, and a maximum likelihood tree was constructed by
PhyML. The tree generated was visualized using the Interactive Tree of Life (http://itol.embl.de/;
Letunic and Bork 2006; Letunic and Bork 2011).
Expression data was obtained in part via CyanoExpress
(http://cyanoexpress.sysbiolab.eu/; Hernandex-Prieto M and Futschik M 2012).
Structural Comparisons
ESyPred3D Web Server 1.0 (http://www.fundp.ac.be) was used to create homology
models of the following proteins using PDB 3B1J_C as a template: geneOID 641611274,
641610209, 637231127, 637231794, 637231269, 637798767, 2509508362, 2508607866,
2504134271, 637448241 (Lambert et al. 2009; Table S1). PDB 2QH1_A was used as a
template to create a homology model of 2509508362. Homology models were visualized using
The PyMOL Molecular Graphics System, Version 1.5.0.4 Schrödinger, LLC (DeLano WL 2002).
ACKNOWLEDGEMENTS
The authors thank Jeff Cameron and Patrick Shih for helpful discussions and Seth Axen for
helpful discussions and figure preparation.
www.plantphysiol.orgon July 12, 2018 - Published by Downloaded from Copyright © 2012 American Society of Plant Biologists. All rights reserved.

17
LITERATURE CITED
Aguado-Llera D, Oyenarte I, Martínez-Cruz LA, Neira JL (2010) The CBS domain protein MJ0729 of M. jannaschii binds DNA. FEBS Lett. 584: 4485–4489
Altschul SF, Gish W, Miller W, Myers EW, Lipman DJ (1990) Basic local alignment search tool. J. Mol. Bio. 215:403–410
Altschul SF, Madden TL, Schaffer AA, Zhang J, Zhang Z, Miller W, Lipman DJ (1997) Gapped BLAST and PSI-BLAST: a new generation of protein database search programs. Nucleic Acids Res. 25:3389–3402
Bateman A (1997) The structure of a domain common to archaebacteria and the homocystinuria disease protein. Trends Biochem.Sci. 22: 12–13
Baykov AA, Tuominen HK, Lahti R (2011) The CBS domain: a protein module with an emerging prominent role in regulation. ACS Chem. Biol. 6(11):1156-63
Berman HM, Westbrook J, Feng Z, Gilliland G, Bhat TN, Weissig H, Shindyalov IN, Bourne PE (2000) The Protein Data Bank. Nucleic Acids Res. 28: 235-242
Carmo-Silva AE, Marri L, Sparla F, Salvucci ME (2011) Isolation and Compositional Analysis of a CP12-Associated Complex of Calvin Cycle Enzymes from Nicotiana tabacum. Protein Pept Lett. 18(6): 618-624
Day P, Sharff A, Parra L, Cleasby A, Williams M, Horer S, Nar H, Redemann N, Tickle I, Yon J (2007) Structure of a CBS-domain pair from the regulatory gamma 1 subunit of human AMPK in complex with AMP and ZMP. Crystallogr. D Biol. Crystallogr 63: 587-596
DeLano WL (2002) The PyMOL molecular graphics system, version1.3, Schrodinger, LLC
Delobel A, Graciet E, Andreescu S, Gontero B, Halgand F, Laprévote O (2005) Mass spectrometric analysis of the interactions between CP12, a chloroplast protein, and metal ions: a possible regulatory role within a PRK/GAPDH/CP12complex. Rapid Comm. Mass Spectrom. 19(22):3379-88
Dereeper A, Guignon V, Blanc G, Audic S, Buffet S, Chevenet F, Dufayard JF, Guindon S, Lefort V, Lescot M, Claverie JM, Gascuel O (2008) Phylogeny.fr: robust phylogenetic analysis for the non-specialist. Nucleic Acids Res. 36: W465-9
Dereeper A, Audic S, Claverie JM, Blanc G (2010) BLAST-EXPLORER helps you building datasets for phylogenetic analysis. BMC Evol. Biol. 10:8
Eddy SR (1998). Profile hidden Markov models. Bioinformatics, 14:755-763
Eddy SR (2008). A probabilistic model of local sequence alignment that simplifies statistical significance estimation. PLoS Comput. Biol. 4:e1000069
Edgar RC (2004) MUSCLE: multiple sequence alignment with high accuracy and high throughput. Nucleic Acids Res. 32(5):1792-1797
Elvitigala T, Stöckel J, Ghosh BK, Pakrasi HB (2009) Effect of continuous light on diurnal rhythms in Cyanothece sp. ATCC 51142. BMC Genomics 10:226
www.plantphysiol.orgon July 12, 2018 - Published by Downloaded from Copyright © 2012 American Society of Plant Biologists. All rights reserved.

18
Erales J, Gontero B, Maberly SC (2008a) Specificity and function of glyceraldehyde-3-phosphate dehydrogenase in a freshwater diatom, Asterionella formosa (Bacillariophyceae). J Phycol. 44:1455–1464
Erales J, Avilan L, Lebreton S, Gontero B (2008b) Exploring CP12 binding proteins revealed aldolase as a new partner for the phosphoribulokinase/glyceraldehyde 3-phosphate dehydrogenase/CP12 complex—purification and kinetic characterization of this enzyme from Chlamydomonas reinhardtii. FEBS J. 275:1248–1259
Erales J, Lorenzi M, Lebrun R, Fournel A, Etienne E, Courcelle C, Guigliarelli B, Gontero B, Belle V (2009a) A New Function of GAPDH from Chlamydomonas reinhardtii: A Thiol-Disulfide Exchange Reaction with CP12. Biochemistry. 48(25): 6034-6040
Erales J, Lignon S, Gontero B (2009b) CP12 from Chlamydomonas reinhardtii, a Permanent Specific "Chaperone-like" Protein of Glyceraldehyde-3-phosphate Dehydrogenase J. Biol. Chem. 284(19): 12735-12744
Erales J, Lignon S, Gontero B (2009c) Mapping of a copper-binding site and the small CP12 chloroplastic protein of Chlamydomonas reinhardtii using top-down mass spectrometry and site-directed mutagenesis. Biochem J. 419: 75-82
Erales J, Mekhalfi M, Woudstra M, Gontero B (2011) Molecular Mechanism of NADPH-Glyceraldehyde-3-phosphate Dehydrogenase Regulation through the C-Terminus of CP12 in Chlamydomonas reinhardtii. Biochemistry 50(14): 2881-2888
Feng L, Campbell EB, Hsiung Y, MacKinnon, R (2010) Structure of a eukaryotic CLC transporter defines an intermediate state in the transport cycle. Science 330: 635–641
Fermani S, Trivelli X, Sparla F, Thumiger A, Calvaresi M, Marri L, Falini G, Zerbetto F, Trost P (2012) Conformational Selection and Folding-upon-binding of Intrinsically Disordered Protein CP12 Regulate Photosynthetic Enzymes Assembly. J. Biol. Chem. 287(25):21372-83
Flaherty BL, Van Nieuwerburgh F, Head SR, Golden JW (2011) Directional RNA deep sequencing sheds new light on the transcriptional response of Anabaena sp. strain PCC 7120 to combined-nitrogen deprivation. BMC Genomics. 12:332
Graciet E, Lebreton S, Camadro JM, (2003a) Characterization of native and recombinant A(4) glyceraldehydes 3-phosphate dehydrogenase – Kinetic evidence for conformation changes upon association with the small protein CP12. Eur. J. Biochem 270(1): 129-136
Graciet E, Gans P, Wedel N, Lebreton S, Camadro JM, Gontero B (2003b) The Small Protein CP12: A Protein Linker for Supramolecular Complex Assembly. Biochemistry 42, 8163-8170
Groben R, Kaloudas D, Raines CA, Offmann B, Maberly SC, Gontero B (2010) Comparative sequence analysis of CP12, a small protein involved in the formation of a Calvin cycle complex in photosynthetic organisms. Photosynth. Res. 103(3):183-94
Hattori M, Tanaka Y, Fukai S, Ishitani R, Nureki O (2007) Crystal structure of the MgtE Mg2+ transporter. Nature 448: 1072–1075
www.plantphysiol.orgon July 12, 2018 - Published by Downloaded from Copyright © 2012 American Society of Plant Biologists. All rights reserved.

19
Hattori M, Iwase N, Furuya N, Tanaka Y, Tsukazaki T, Ishitani R, Maguire ME, Ito K, Maturana A, Nureki O (2009) Mg 2+-dependent gating of bacterial MgtE channel underlies Mg2+ homeostasis. EMBO J. 28: 3602–3612
Hihara Y, Kamei A, Kanehisa M, Kaplan A, Ikeuchi M (2001) DNA microarray analysis of cyanobacterial gene expression during acclimation to high light. Plant Cell. 13(4): 793-806
Howard TP, Metodiev M, Lloyd JC, Raines CA (2008) Thioredoxin-mediated reversible dissociation of a stromal multiprotein complex in response to changes in light availability. Proc. Natl. Acad. Sci. U. S. A. 105(10): 4056-4061
Howard TP, Upton GJG, Lloyd JC, Raines CA (2011a) Antisense suppression of the small chloroplast protein CP12 in tobacco: a transcriptional viewpoint. Plant Signal. Behav. 6(12) 2026–2030
Howard TP, Fryer MJ, Singh P, Metodiev M, Lytovchenko A, Obata T, Fernie AR, Kruger NJ, Quick WP, Lloyd JC, Raines CA (2011b) Antisense Suppression of the Small Chloroplast Protein CP12 in Tobacco Alters Carbon Partioning and Severely Restricts Growth. Plant Phys. 157(2): 620-631
Howard TP, Lloyd JC, Raines CA (2011c) Inter-species variation in the oligomeric states of the higher plant Calvin cycle enzymes glyceraldehyde-3-phosphate dehydrogenase and phosphoribulokinase J Exp Bot. 62(11): 3799-3805 Ignoul S and Eggermont J (2005) CBS domains: structure, function, and pathology in human proteins. Am. J. Physiol.: Cell Physiol. 289: C1369–1378
Kery V, Poneleit L, Kraus J (1998) Trypsin cleavage of human cystathionine ß-synthase into an evolutionarily conserved active core: structural and functional consequences. Arch. Biochem. Biophys. 355: 222–232
Kushwaha HR, Singh AK, Sopory SK, Singla-Pareek SL, Pareek A (2009) Genome wide expression analysis of CBS domain containing proteins in Arabidopsis thaliana (L.) Heynh and Oryza sativa L. reveals their developmental and stress regulation. BMC Genomics 10: 200
Lambert C, Leonard N, De Bolle X, Depiereux E (2002) ESyPred3D: Prediction of proteins 3D structures. Bioinformatics. 18(9):1250-1256
Leutnic I, Bork P (2006) Interactive Tree of Life (iTOL): an online tool for phylogenetic tree display and annotation. Bioinformatics. 23(1):127-8
Leutnic I, Bork P (2011) Interactive Tree of Life v2: online annotation and display of phylogenetic trees made easy. Nucleic Acid Res. 39(2):W475-8
Lebreton S, Graciet E, Gontero B (2003b) Modulation, via protein-protein interactions, of glyceraldehydes 3-phosphate dehydrogenase activity through redox phosphoribulokinase regulation. J. Biol. Chem. 278(14) 12078-12084
Lebreton S, Andreescu S, Graciet E, Gontero B (2006) Mapping of the interaction site of CP12 with glyceraldehyde-3-phosphate dehydrogenase from Chlamydomonas reinhardtii. FEBS J. 273:3358–3369
www.plantphysiol.orgon July 12, 2018 - Published by Downloaded from Copyright © 2012 American Society of Plant Biologists. All rights reserved.

20
Li X, Romero P, Rani M, Dunker AK, Obradovic Z (1999) Predicting protein disorder for N-, C, and internal regions. Genome Inform. Ser. Workshop Genome Inform. 10: 30-40
Liu F, VanToai T, Moy LP, Bock G, Linford LD, Quackenbush J (2005) Global transcription profiling reveals comprehensive insights into hypoxic response in Arabidopsis. Plant Phys. 137(3): 1115-1129
Ludwig M and Bryant DA (2011) Transcription Profiling of the Model Cyanobacterium Synechococcus sp. Strain PCC 7002 by Next-Gen (SOLiD™) Sequencing of cDNA. Front Microbiol. 2:41
Marri L, Sparla F, Pupillo P, Trost P (2005a) Coordinated gene expression of photosynthetic glyceraldehydes-3-phosphate dehydrogenase, phosphoribulokinase and CP12 in Arabidopsis thaliana. J Exp Bot. 56(409): 73-80
Marri L, Trost P, Pupillo P, Sparla F (2005b) Reconstitution and Properties of the Recombinant Glyceraldehyde-3-Phosphate Dehydrogenase/CP12/Phosphoribulokinase Supramolecular Complex of Arabidopsis. Plant Physiol. 283: 1831-1838
Marri L, Trost P, Trivelli X, Gonnelli L, Pupillo P, Sparla F (2008) Spontaneous assembly of photosynthetic supramolecular complexes as mediated by the intrinsically unstructured protein CP12. J. Biol. Chem. 139(3): 1433-1443
Marri L, Zaffagnini M, Collin V, Issakidis-Bourguet E, Pupillo P, Sparla F, Miginiac-Maslow M, Trost P (2009) Prompt and Easy Activation by Specific Thioredoxins of Cavin Cycle Enzymes of Arabidopsis Thaliana Associated in the GAPDH/CP12/PRK Supramolecular Complex. Mol. Plant 2(2): 259-269
Marri L, Pesaresi A, Valerio C, Lamba D, Pupillo P, Trost P, Sparla F (2010) In vitro characterization of Arabidopsis CP12 isoforms reveals common biochemical and molecular properties. J. Plant Physiol. 167(12): 939-950
Matsumura H, Kai A, Maeda T, Tamoi M, Satoh A, Tamura H, Hirose M, Ogawa T, Kizu N, Wadano A, Inoue T, Shigeoka S (2011) Structure basis for the regulation of glyceraldehyde-3-phosphate dehydrogenase activity via the intrinsically disordered protein CP12. Structure 19(12):1846-54
McLean JE, Hamaguchi N, Belenky P, Mortimer SE, Stanton M, Hedstrom L (2004) Inosine 5’-monophosphate dehydrogenase binds nucleic acids in vitro and in vivo. Biochem. J. 379: 243–251
Masepohl B, Schölisch K, Görlitz K, Kutzki C, Bohme H (1997) The heterocyst-specific fdxH gene product of the cyanobacterium Anabaena sp. PCC 7120 is important but not essential for nitrogen fixation. Mol. Gen. Genet. 253(6):770-6
Hernandex-Prieto M and Futschik M (2012) CyanoEXpress: a web database for interactive exploration and visualization of the integrated transcriptome of cyanobacterium Synechocystis sp. PCC6803. Bioinformation. 8(13):634-638
Oesterhelt C, Klocke S, Holtgrefe S, Linke V, Weber APM, Scheibe R (2007) Redox regulation of chloroplast enzymes in Galdieria sulphuraria in view of eukaryotic evolution . Plant Cell Physiol. 48(9):1359-1373
www.plantphysiol.orgon July 12, 2018 - Published by Downloaded from Copyright © 2012 American Society of Plant Biologists. All rights reserved.

21
Petersen J, Teich R, Becker B, Cerff R, Brinkmann H (2006) The GapA/B Gene Duplication Marks the Origin of Streptophyta (Charophytes and Land Plants) Mol. Biol. Evol. 23(6):1109–1118
Pohlmeyer K, Paap BK, Soll J, Wedel N (1996) CP12: a small nuclear-encoded chloroplast protein provides novel insights into higher-plant GAPDH evolution. Plant Mol. Biol. 32:969–978
Robbens S, Petersen J, Brinkmann H, Rouzé P, Van de Peer Y (2007) Unique regulation of the Calvin cycle in the ultrasmall green alga Ostreococcus. J. Mol. Evol. 64(5):601-4
Romero P, Obradovic Z, Li X, Dunker AK (1997) Sequence data analysis for long disordered regions prediction in the calcineurin family. Genome Inform. Ser. Workshop Genome Inform. 8:110-124
Romero P, Obradovic Z, Li X, Garner E, Brown C, Dunker AK (2001) Sequence complexity of disordered protein. Proteins 42:38-48
Schwarz D, Nodop A, Hüge J, Purfürst S, Forchhammer K, Michel KP, Bauwe H, Kopka J, Hagemann M (2011) Metabolic and transcriptomic phenotyping of inorganic carbon acclimation in the Cyanobacterium Synechococcus elongatus PCC 7942. Plant Physiol. 155(4):1640-55
Scott J, Hawley S, Green K, Anis M, Stewart G, Scullion G, Norman D, Hardie D (2004) CBS domains form energysensing modules whose binding of adenosine ligands is disrupted by disease mutations. J. Clin. Invest. 113: 274–284
Schuster-Böckler B, Schultz J, Rahmann S (2004) HMM Logos for visualization of protein families. BMC Bioinf. 5;7
Sharpe M, Gao C, Kendall S, Baker E, Lott J (2008) The structure and unusual protein chemistry of hypoxic response protein 1,a latency antigen and highly expressed member of the DosR regulon in Mycobacterium tuberculosis. J. Mol. Biol. 383:822–836
Scheibe R, Wedel N, Vetter S, Emmerlich V, Sauermann SM (2002). Co-existence of two regulatory NADP-glyceraldehyde 3-P dehydrogenase complexes in higher plant chloroplasts. Eur. J. Biochem. 269(22): 5617-5624
Shoumskaya MA, Paithoonrangsarid K, Kanesaki Y, Los DA, Zinchenko VV, Tanticharoen M, Suzuki I, Murata N (2005). Identical Hik-Rre systems are involved in perception and transduction of salt signals and hyperosmotic signals but regulate the expression of individual genes to different extents in synechocystis. J. Biol. Chem. 280(22): 21531-8
Singh P, Kaloudas D, Raines CA (2008) Expression analysis of the Arabidopsis CP12 gene family suggests novel roles for these proteins in roots and floral tissues. J. Exp. Bot. 59:3975–3985
Stöckel J, Welsh EA, Liberton M, Kunnvakkam R, Aurora R, Pakrasi HB (2008) Global transcriptomic analysis of Cyanothece 51142 reveals robust diurnal oscillation of central metabolic processes. Proc. Natl. Acad. Sci. U. S. A. 105(16):6156-61
Tamoi M, Miyazaki T, Fukamizo T, Shigeoka S (2005) The Calvin cycle in cyanobacteria is regulated by CP12 via the NAD(H)/NADP(H) ratio under light/dark conditions. Plant J. 42:504–513
www.plantphysiol.orgon July 12, 2018 - Published by Downloaded from Copyright © 2012 American Society of Plant Biologists. All rights reserved.

22
Thompson LR, Zeng Q, Kelly L, Huang KH, Singer AU, Stubbe J, Chisholm SW (2011) Phage auxiliary metabolic genes and the redirection of cyanobacterial host carbon metabolism. Proc. Natl. Acad. Sci. U. S. A. 108(39):E757-64
Tompa P (2002) Intrinsically unstructured proteins. Trends Biochem. Sci 27:527–533
Tompa P, Szasz C, Buday L (2005) Structural disorder throws new light on moonlighting. Trends Biochem. Sci 30:484–489
Trost P, Fermani S, Marri L, Zaffagnini M, Falini G, Scagliarini S, Pupillo P, Sparla F (2006) Thioredoxin-dependent regulation of photosynthetic glyceraldehydes-3-phosphate dehydrogenase: autonomous vs. CP12-dependent mechanisms. Photosynth. Res. 89(2-3): 263-275
Vijayan V, Jain IH, O'Shea EK (2011) A high resolution map of a cyanobacterial transcriptome. Genome Biol. 12(5):R47
Waterhouse AM, Procter JB, Martin DMA, Clamp M, Barton GJ (2009) Jalview Version 2 – a multiple sequence alignment editor and analysis workbench. Bioinformatics 24(9):1189-1191
Wedel N, Soll J, Paap BK (1997) CP12 provides a new mode of light regulation of Calvin cycle activity in higher plants. Proc. Natl. Acad. Sci. U.S.A. 94:10479–10484
Wedel N, Soll J (1998) Evolutionary conserved light regulation of Calvin cycle activity by NADPH-mediated reversible phosphoribulokinase/CP12/glyceraldehyde-3-phosphate dehydrogenase complex dissociation. Proc. Natl. Acad. Sci. U.S.A. 95:9699–9704
Wright P and Dyson H (1999) Intrinsically unstructured proteins: re-assessing the structure-function paradigm. J. Mol. Bio. 293:321-331
Zhang Z, Pendse ND, Phillips KN, Cotner JB, Khodursky A (2008) Gene expression patterns of sulfur starvation in Synechocystis sp. PCC 6803. BMC Genomics. 9: 344-357
Zilliges Y, Kehr J, Meissner S, Ishida K, Mikkat S, Hagemann M, Kaplan A, Börner T, Dittmann E (2011) The Cyanobacterial Hepatotoxin Microcystin Binds to Proteins and Increases the Fitness of Microcystis under Oxidative Stress Conditions. PloS ONE 6: e17615
www.plantphysiol.orgon July 12, 2018 - Published by Downloaded from Copyright © 2012 American Society of Plant Biologists. All rights reserved.

23
Figure Legends
Figure 1 Schematic representation of the eight cyanobacterial CP12 types.
Figure 2 Sequence analysis of cyanobacterial CP12 variants. A) HMM logos of CP12 variants.
Asterisks indicate key residues detailed in panel B. The CBS domains are denoted by a solid
underline; the putative ribose-PO4 binding site Ghx(T/S)x(T/S)D (Day et al. 2007) are denoted
by a dashed underline. The blue helix indicates the segment corresponding to the predicted
alpha helix. B) Average percent of key interacting residues identified by Matsumura et al. 2011
present in each CP12 variant.
Figure 3 Homology models of representative cyanobacterial CP12 and GAPDH proteins in
complex modeled using PDB 3B1J as template (Matsumura et al. 2011). NAD is shown in blue
sticks, copper as an orange sphere, the template CP12 is magenta, the modeled CP12 is
yellow, the template GAPDH is cyan and the modeled GAPDH is red. Residues in the C-
terminal loop are labeled. A) Synechococcus PCC7002 (Identity to template: CP12-N/C 48.0%,
GAPDH 74.33%) B) Synechococcus PCC7942 (Identity to template: CP12-N/C 52.1%, GAPDH
74.11%) C) Gloeocapsa sp. PCC7428 (Identity to template: CP12-C 47.2%, GAPDH 74.11%)
D) Prochlorococcus MIT9313 (Identity to template: CP12-C(M) 36.0%, GAPDH 70.8%) E)
Leptolyngbya PCC6406 (Identity to template: CP12-N 40.0%, GAPDH 74.11%) F) Geitlerinema
PCC7407 (Identity to template: CP12-0 40%, GAPDH 77.98%) G) Fischerella PCC9339
(Identity to template: CP12-N/C-CBS 46.2%, GAPDH 74.41%) H) Nostoc PCC7120 (Identity to
template: CP12-N-CBS 20.7%, GAPDH 76.19%).
Figure 4 Number and type of CP12 variant found in each cyanobacterial genome displayed
phylogenetically (Tree adapted from Shih et al. 2012).
Figure 5 Maximum likelihood gene tree of all cyanobacterial CP12s.
www.plantphysiol.orgon July 12, 2018 - Published by Downloaded from Copyright © 2012 American Society of Plant Biologists. All rights reserved.
24
Table I CP12 residue interactions Summary of CP12 residues implicated in formation of the GAPDH-PRK-CP12 complex and their ascribed roles GAPDH NAD Cu2+ PRK Asp54 Electrostatic
attraction Asp59 Electrostatic
attraction Glu63 Electrostatic
attraction Asp66 Ion pair with
GAPDH Asp80 Electrostatic
attraction Glu69 H bond GAPDH
Thr37; Van der Waals GAPDH Ser38, Asp39a
Contacts 2’ hydroxyl
Electrostatic attraction
Cys70 H bond GAPDH His195, Arg196
Leu71 H bond GAPDH His195, Arg196a
Tyr73 H bond GAPDH Thr 156, Thr185, Asp187, Arg200, Thr213a
Interacts with phosphate and ribose group
Asp74 H bond GAPDH Thr 156, Thr185, Arg200, Thr213
Asp75 H bond GAPDH Thr 156, Thr185, Arg200, Thr213b
Interactionc
aGAPDH-binding also seen by Fermani et al. 2012 bImportance for GAPDH binding also suggesting by Erales et al. 2011 cCopper interacting seen by Erales et al. 2009c
www.plantphysiol.orgon July 12, 2018 - Published by Downloaded from Copyright © 2012 American Society of Plant Biologists. All rights reserved.
25
Table II PONDR VL-XT disorder predictions Percent disorder predictions for the CP12 domain of selected cyanobacterial CP12 sequences Type Gene OID Genome Percent Disorder CP12-N/C 2509710708 Pleurocapsa sp. PCC7319 57.53 CP12-N/C 2509434278 Microcoleus sp. PCC7113 60.00 CP12-N 2508607866 Leptolyngbya sp. PCC6406 80.52 CP12-C 2509434317 Microcoleus sp. PCC7113 58.46 CP12-0 2509434904 Microcoleus sp. PCC7113 60.24 CP12-N/C-CBS 2503740304 Nostoc sp. PCC7120 53.49 CP12-N-CBS 637231127 Nostoc sp. PCC7120 40.51 CP12-N-CBS 2509436117 Microcoleus sp. PCC7113 61.90 CP12-0-CBS 637231794 Nostoc sp. PCC7120 34.62 CP12-C(M) 650129895 Synechococcus sp. CB0101 43.06 CP12-C(M) 650131651 Synechococcus sp. CB0101 43.08
www.plantphysiol.orgon July 12, 2018 - Published by Downloaded from Copyright © 2012 American Society of Plant Biologists. All rights reserved.