deregulated expression of circadian clock and clock-controlled cell cycle genes in chronic...
TRANSCRIPT

Deregulated expression of circadian clock and clock-controlledcell cycle genes in chronic lymphocytic leukemia
Sobia Rana • Mustafa Munawar • Adeela Shahid •
Meera Malik • Hafeez Ullah • Warda Fatima •
Shahida Mohsin • Saqib Mahmood
Received: 15 December 2012 / Accepted: 26 October 2013 / Published online: 5 November 2013
� Springer Science+Business Media Dordrecht 2013
Abstract Circadian rhythms are endogenous and self-
sustained oscillations of multiple biological processes with
approximately 24-h rhythmicity. Circadian genes and their
protein products constitute the molecular components of
the circadian oscillator that form positive/negative feed-
back loops and generate circadian rhythms. The circadian
regulation extends from core clock genes to various clock-
controlled genes that include various cell cycle genes.
Aberrant expression of circadian clock genes, therefore,
may lead to genomic instability and accelerated cellular
proliferation potentially promoting carcinogenesis. The
current study encompasses the investigation of simulta-
neous expression of four circadian clock genes (Bmal1,
Clock, Per1 and Per2) and three clock-controlled cell cycle
genes (Myc, Cyclin D1 and Wee1) at mRNA level and
determination of serum melatonin levels in peripheral
blood samples of 37 CLL (chronic lymphocytic leukemia)
patients and equal number of age- and sex-matched healthy
controls in order to indicate association between deregu-
lated circadian clock and manifestation of CLL. Results
showed significantly down-regulated expression of Bmal1,
Per1, Per2 and Wee1 and significantly up-regulated
expression of Myc and Cyclin D1 (P \ 0.0001) in CLL
patients as compared to healthy controls. When expression
of these genes was compared between shift-workers and
non-shift-workers within the CLL group, the expression
was found more aberrant in shift-workers as compared to
non-shift-workers. However, this difference was found
statistically significant for Myc and Cyclin D1 only
(P \ 0.05). Serum melatonin levels were found signifi-
cantly low (P \ 0.0001) in CLL subjects as compared to
healthy controls whereas melatonin levels were found still
lower in shift-workers as compared to non-shift-workers
within CLL group (P \ 0.01). Our results suggest that
aberrant expression of circadian clock genes can lead to
aberrant expression of their downstream targets that are
involved in cell proliferation and apoptosis and hence may
result in manifestation of CLL. Moreover, shift-work and
low melatonin levels may also contribute in etiology of
CLL by further perturbing of circadian clock.
Keywords Circadian clock � Clock genes �Clock-controlled genes � Shift-work � Melatonin �Chronic lymphocytic leukemia
Introduction
Circadian rhythms are the outward manifestation of an
internal timing system generated by a circadian clock that
is synchronized by the day–night cycle [1]. The circadian
clock proficiently coordinates the homeostatic processes of
living organisms to match imposed 24-h cycles and
S. Rana � A. Shahid
Department of Physiology & Cell Biology, University of Health
Sciences, Lahore, Pakistan
M. Munawar � H. Ullah � S. Mahmood
Department of Allied Health Sciences, University of Health
Sciences, Lahore, Pakistan
M. Malik � S. Mahmood (&)
Department of Human Genetics & Molecular Biology,
University of Health Sciences, Lahore, Pakistan
e-mail: [email protected]; [email protected]
W. Fatima
Department of Microbiology & Molecular Genetics, University
of the Punjab, Lahore, Pakistan
S. Mohsin
Department of Hematology, University of Health Sciences,
Lahore, Pakistan
123
Mol Biol Rep (2014) 41:95–103
DOI 10.1007/s11033-013-2841-7

influences nearly all aspects of physiology and behavior
[2]. The mammalian clock system is hierarchical with a
master clock controlling circadian rhythms located in the
suprachiasmatic nucleus (SCN) and various slave oscilla-
tors present in peripheral organs [3]. The molecular
clockwork is composed of a network of transcriptional–
translational feedback loops that drive rhythmic, *24-h
expression patterns of core clock components [4]. Core
clock components are genes whose protein products are
necessary for the generation and regulation of circadian
rhythms within individual cells throughout the organism
[5]. Some of core clock genes include Bmal1, Crypto-
chrome 1 (Cry1), Cryptochrome 2 (Cry2), Period 1 (Per1),
Period 2 (Per2), Period 3 (Per3), Clock (Clk), and Casein
kinase1e (CK1e) [6]. The identification of the circadian
transcripts has revealed that the transcriptional circadian
regulation extends beyond core clock components to
include various clock-controlled genes (CCGs); genes that
are under the direct or indirect transcriptional control of the
clock transcription factors but are not themselves part of
the clock. Regulation of CCGs is a mechanism by which
the molecular clockwork controls physiological processes.
The CCGs constitute about 10 % of the expressed genes in
a given tissue (SCN or in peripheral tissues) [7]. CCGs may
encode a variety of proteins including key regulators for
cell cycle. It has been shown that expression of several cell
cycle genes, including Wee1, c-Myc, CyclinD1, Gadd45,
and Mdm2; oscillate in a circadian manner [8, 9]. Wee1 [8]
and c-Myc [9] are directly regulated by the molecular clock
via E-box elements at their promoters. E-box or Enhancer
box is a DNA sequence found upstream of some promoter
regions in eukaryotes and is recognized and bound by
transcription factors to initiate gene transcription.
Chronic lymphocytic leukemia (CLL) is characterized
by proliferation and accumulation of morphologically
mature but immunologically dysfunctional lymphocytes in
blood, bone marrow and lymphoid tissues. No specific
genetic alteration has yet been associated with this disease.
In particular, CLL is not associated with reciprocal bal-
anced chromosomal translocations, but rather with specific
deletions [10] suggesting the loss of presently unidentified
tumor suppressor genes. CLL cells have low proliferative
rate and a prolonged life span, suggesting that their primary
alteration may be a defect in apoptosis [11]. Since circa-
dian genes regulate many biological pathways including
cell proliferation and apoptosis, and CLL seems to be an
apoptosis related disorder, their alteration may be directly
involved in the pathophysiology of CLL.
The current study encompasses the investigation of
simultaneous expression of four circadian clock genes
(Bmal1, Clock, Per1 and Per2) and three clock-controlled
cell cycle genes (Myc, Cyclin D1 and Wee1) at mRNA
level by quantitative (real-time) RTPCR and determination
of serum melatonin levels by ELISA in 37 CLL patients
and equal number of age- and sex-matched healthy controls
in order to indicate any association between deregulated
circadian clock and manifestation of CLL.
Materials and methods
Samples and subjects
This study was conducted after obtaining permission from
Institutional Review Board of University of Health Sci-
ences, Lahore. The study involved 37 newly diagnosed
CLL patients (Binet stage A) and equal number of their
age- and sex-matched healthy individuals. The patients
were recruited from the oncology departments of the local
tertiary care hospitals. Clinical diagnosis was based on
standard morphological and immunophenotypic criteria.
The informed consents were obtained from both, the
patients and the healthy controls. Blood samples from CLL
patients were collected before the start of their treatment
regime. In order to rule out the bias of time, collection of
all peripheral blood (PB) samples was carried out between
10:00 and 11:00 AM. Samples were processed within 1 h
of collection for RNA and serum isolation.
Quantitative RTPCR
Total RNA was extracted by using FavorPrepTM Total
RNA Purification Mini Kit (Cat. No. FABRK 100, Fa-
vorgen Biotech Corp., Taiwan). Native agarose gel elec-
trophoresis was performed to assess the overall quality of
total RNA. The quantity of total RNA was estimated using
Nanodrop ND2000 (Thermo Scientific, USA).
The first strand cDNA synthesized was used directly for
amplification by polymerase chain reaction (PCR). The
cDNA sequences of the four circadian clock genes namely
BMAL1 (GenBank accession no. NM_001178), Clock
(GenBank accession no. NM_004898), Per1 (GenBank
accession no. NM_002616), Per2 (GenBank accession no.
NM_022817); three cell cycle genes namely Myc (Gen-
Bank accession no. NM_002467), Cyclin D1 (GenBank
accession no. NM_053056), Wee1 (GenBank accession no.
NM_003390); and GAPDH (GenBank accession no.
NM_002046) were determined. The specific forward and
reverse primers and TaqMan probes were designed using
the free online SciTools of Integrated DNA Technologies
(http://eu.idtdna.com/Scitools/Applications/RealTimePCR/).
The information regarding the designed primers and probes
are summarized in Table 1. The designed primers and
probes (labeled with appropriate fluorescent dyes) were
obtained from Gene LinkTM (Hawthorne, NY, USA).
Expression of GAPDH was also examined by real-time
96 Mol Biol Rep (2014) 41:95–103
123

RT-PCR as the internal control for normalization of target
gene expression. No template control (NTC) and reverse
transcriptase negative (RT-) control were always per-
formed to check reagent and DNA contamination. A
positive control RTPCR was also performed every time
using template RNA and GAPDH primers provided in the
kit to check the fidelity of the RTPCR reaction.
First strand cDNA was synthesized by using Revert-
AidTM First Strand cDNA Synthesis Kit (Cat. No. K1622,
Fermentas, Germany) in a final volume of 20 lL contain-
ing 2 lg RNA. The tube containing RNA, Oligo (dT)18
primer and DEPC-treated water was first incubated for
5 min at 65 �C. Then rest of the components including
reaction buffer, RNase inhibitor, dNTP mix and reverse
transcriptase were added and incubated at 42 �C for
60 min, then the reaction was stopped by heat inactivation
at 70 �C for 5 min. Prior shifting to real-time PCR, reac-
tion was optimized through conventional RTPCR (Fig. 1).
Real-time quantitative PCR was carried out in an iQTM5
Multi-color Real-Time PCR Detection System (Bio-Rad,
USA) using the Maxima Probe qPCR Master Mix (Cat. No.
K0232, Fermentas).
All reactions were carried out in a 25 lL final volume
containing 0.3 lM each primer, 0.2 lM probe and 12.5 lL
29 Maxima Probe qPCR Master Mix. The PCR cycling
parameters were set as follows: 95 �C for 10 min followed
by 40 cycles of PCR reactions at 95 �C for 15 s and 60 �C
for 1 min.
All reactions were run in triplicates. To determine the
inter-assay precision, three replicates of cDNA of each
sample were run on three separate days. Intra-assay
(within-run) precision was determined by calculating
mean, standard deviation (SD) and coefficient of variance
(CV) of the CT values for each sample and for each set of
primers and probe on each day.
ELISA
Enzyme-linked immunosorbent assay (ELISA) was per-
formed to determine melatonin concentrations in the serum
samples of CLL patients and their age- and sex-matched
healthy controls on an automated EIA analyzer (Bio-Rad
Laboratories, Hercules, CA, USA). For this purpose,
Table 1 Oligonucleotide primers and probes for real-time quantitative reverse transcription-polymerase chain reaction analysis of the genes
under consideration
Pimer Sequence Length Location GC (%) Strand Tm (�C) Junction Ampliconsize (bp)
F-GAPDH 50-CATCTTCCAGGAGCGAGAT-30 19 327–345 52.6 ? 60.4 Exon 4–5 136
P-GAPDH 50-CTGCAAATGAGCCCCAGCCTT-30 21 441–421 57.1 - 67.0 Exon 5–6
R-GAPDH 50-GATGACCCTTTTGGCTCC-30 18 462–445 55.6 - 59.5 Exon 6
F-PER1 50-GCAGCCTCGGTTTTCTGA-30 18 3,694–3,711 55.6 ? 61.6 Exon 22 139
P-PER1 50-TGTGATGGCCTGTGTGGACTGT-30 22 3,781–3,802 54.5 ? 67.5 Exon 22–23
R-PER1 50-AGGGTGACCAGGATCTTG-30 18 3,832–3,815 55.6 - 59.9 Exon 23
F-PER2 50-CTGAAGAGGAAATGCGAGT-30 19 2,044–2,062 47.4 ? 59.1 Exon 16 140
P-PER2 50-CCACACGCTGGAGAGGCAGA-30 20 2,125–2,144 65.0 ? 67.4 Exon 16–17
R-PER2 50-GTACCTACTCCCGTGCG-30 17 2,183–2,167 64.7 - 60.2 Exon 17
F-CLK 50-CAGTCTCAAGGAAGCATTGG-30 20 1,583–1,602 50.0 ? 60.2 Exon 16 139
P-CLK 50-TCAGACCCTTCCTCAACACCAACC-30 24 1,674–1,697 54.2 ? 67.3 Exon 16–17
R-CLK 50-AGTGCTCGTATCCGTCG-30 17 1,721–1,705 58.8 - 60.0 Exon 17
F-BMAL1 50-GGAATATGTTTCTCGGCACG-30 20 1,419–1,438 50.0 ? 60.5 Exon 13 137
P-BMAL1 50-CAAAATAGCTGTTGCCCTCTGGTCT-30 25 1,488–1,464 48.0 - 66.3 Exon 13-14
R-BMAL1 50-GTCCTATGTCATCTTGGTGAA-30 21 1,555–1,535 42.9 - 59.2 Exon 14
F-CYCD1 50-CGGTGTCCTACTTCAAATGTG-30 21 331–351 47.6 ? 60.4 Exon 1 140
P-CYCD1 50-TTCCTCGCAGACCTCCAGCAT-30 21 419–399 57.1 - 67.0 Exon 1-2
R-CYCD1 50-GCGGTCCAGGTAGTTCAT-30 17 470–453 55.6 - 60.5 Exon 2
F-WEE1 50-GTGTGAAGAGGCTGGATG-30 18 2,088–2,105 55.6 ? 59.1 Exon 4 136
P-WEE1 50-CTGTTGATGAGCAGAACGCTTTGAGAG-30 27 2,151–2,177 48.1 ? 66.8 Exon 4-5
R-WEE1 50-CCTCGACGGAGTCCTC-30 16 1,243–1,258 68.8 ? 59.1 Exon 2
F-MYC 50-CCTCGACGGAGTCCTC-30 16 1,243–1,258 68.8 ? 59.1 Exon 2 140
P-MYC 50-ATCTTCTTGTTCCTCCTCAGAGTCGC-30 26 1,343–1,318 50.0 ? 66.9 Exon 2-3
R-MYC 50-CTGCCTCTTTTCCACAGAA-30 19 1,382–1,364 47.4 - 59.3 Exon 3
Mol Biol Rep (2014) 41:95–103 97
123

Human Melatonin ELISA Kit (Cat. No. CSB-E08132h,
Cusabio Biotech Co., Ltd., China) was utilized.
Statistical analysis
Relative gene expression levels were calculated by using
Livak method [12]. Student’s t test was applied to make
comparisons between the quantitative variables. P values
B0.05 were regarded as being significantly different. The
graphical output was generated by using GraphPad Prism
version 5.00 for Windows, GraphPad Software, La Jolla,
CA, USA, www.graphpad.com.
Results
Among the 37 CLL patients, 27 (73 %) were males and 10
(27 %) were females. The age of patients ranged from 45
to 85 years with a mean age of 62.81 ± 10.84
(mean ± SD) years. Moreover, average age of male
patients was 62.18 ± 11.69 and that of female patients was
64.50 ± 8.40 years.
Aberrant expression of circadian clock and cell cycle
genes in CLL
The quantitative expression of four circadian clock and
three clock-controlled cell cycle genes determined in the
present study by real time quantitative RTPCR showed
significantly aberrant expression of these genes in CLL
patients as compared to the age- and sex-matched healthy
controls (Table 2). The circadian clock genes namely
hBmal1 (P \ 0.0001), hPer1 (P \ 0.0001), and hPer2
(P \ 0.0001) were found significantly down-regulated in
CLL patients as compared to their healthy controls. The
expression of Clock gene was found up-regulated in CLL
group as compared to healthy controls but in a not
Fig. 1 The electrophoretic gel picture shows the RTPCR product of
reference and target genes resulted from an optimized RTPCR
reaction. Lane 1 contain 100 bp DNA Ladder, Lane 2 contain Gapdh
RTPCR product (136 bp), Lane 3 contain Bmal1 RTPCR product
(137 bp), Lane 4 contains Clk RTPCR product (139 bp), Lane 5
contains Per1 RTPCR product (139 bp), Lane 6 contains Per2 RTPCR
product (140 bp), Lane 7 contains Wee1 RTPCR product (136 bp),
Lane 8 contains CycD1 RTPCR product (140 bp), Lane 9 contains
Myc RTPCR product (140 bp), Lane 10 contains positive control
(PC) (496 bp), Lane 11 contains no template control (NTC), Lane 12
contains RT negative control (RT-ve)
Table 2 The expression levels of the four circadian clock genes and
three cell cycle genes as determined by real-time quantitative reverse
transcription-polymerase chain reaction
Gene Healthy individuals CLL
n 37 37
BMAL1
DCT (BMAL1–GAPDH) -1.91 ± 0.40 1.52 ± 0.28*
Relative expression 1 -3.18
CLOCK
DCT (CLOCK–GAPDH) 4.44 ± 0.34 3.62 ± 0.28
Relative expression 1 2.55
PER1
DCT (PER1–GAPDH) 3.11 ± 0.29 5.96 ± 0.30*
Relative expression 1 -2.62
PER2
DCT (PER2–GAPDH) 2.38 ± 0.35 6.38 ± 0.27*
Relative expression 1 -3.27
MYC
DCT (MYC–GAPDH) 5.32 ± 0.27 2.23 ± 0.19*
Relative expression 1 12.20
CyclinD1
DCT (Cyclin D1–GAPDH) 7.62 ± 0.28 3.22 ± 0.33*
Relative expression 1 26.37
WEE1
DCT (WEE1–GAPDH) 4.13 ± 0.30 6.60 ± 0.18*
Relative expression 1 -1.99
* P \ 0.0001 compared with normal. Results are the mean ± SE.
The level of target gene was normalized to the endogenous reference
glyceraldehyde-3-phosphate dehydrogenase (GAPDH) to obtain the
normalized circadian gene expression (DCT) value for each sample.
The DCT of chronic lymphocytic leukemia cases was first related to
the DCT of healthy individuals to obtain the relative threshold cycle
(DDCT) and then the relative expression levels (2-DDCT) were
calculated. The fold change of gene expression is calculated by taking
2-DDCT for values greater than 1 and -1/2-DDCT for values less than
1. Negative sign (-) with values of fold regulation indicates down-
regulation. Asterisk indicates significant fold change
98 Mol Biol Rep (2014) 41:95–103
123
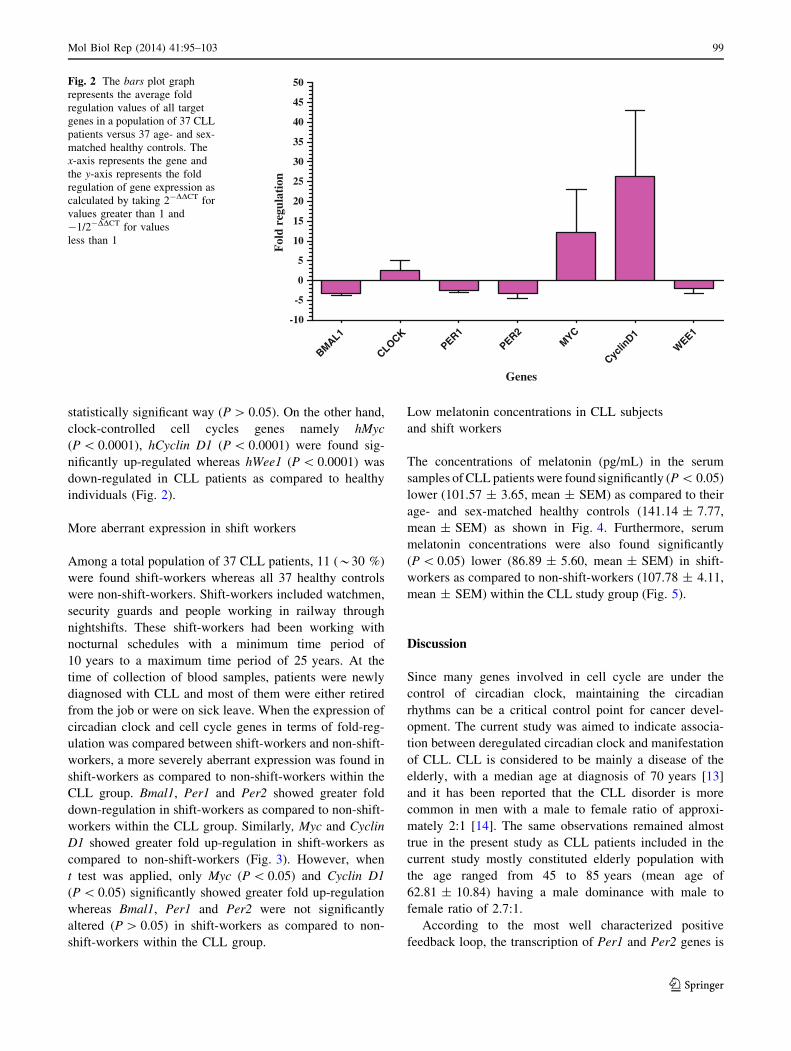
statistically significant way (P [ 0.05). On the other hand,
clock-controlled cell cycles genes namely hMyc
(P \ 0.0001), hCyclin D1 (P \ 0.0001) were found sig-
nificantly up-regulated whereas hWee1 (P \ 0.0001) was
down-regulated in CLL patients as compared to healthy
individuals (Fig. 2).
More aberrant expression in shift workers
Among a total population of 37 CLL patients, 11 (*30 %)
were found shift-workers whereas all 37 healthy controls
were non-shift-workers. Shift-workers included watchmen,
security guards and people working in railway through
nightshifts. These shift-workers had been working with
nocturnal schedules with a minimum time period of
10 years to a maximum time period of 25 years. At the
time of collection of blood samples, patients were newly
diagnosed with CLL and most of them were either retired
from the job or were on sick leave. When the expression of
circadian clock and cell cycle genes in terms of fold-reg-
ulation was compared between shift-workers and non-shift-
workers, a more severely aberrant expression was found in
shift-workers as compared to non-shift-workers within the
CLL group. Bmal1, Per1 and Per2 showed greater fold
down-regulation in shift-workers as compared to non-shift-
workers within the CLL group. Similarly, Myc and Cyclin
D1 showed greater fold up-regulation in shift-workers as
compared to non-shift-workers (Fig. 3). However, when
t test was applied, only Myc (P \ 0.05) and Cyclin D1
(P \ 0.05) significantly showed greater fold up-regulation
whereas Bmal1, Per1 and Per2 were not significantly
altered (P [ 0.05) in shift-workers as compared to non-
shift-workers within the CLL group.
Low melatonin concentrations in CLL subjects
and shift workers
The concentrations of melatonin (pg/mL) in the serum
samples of CLL patients were found significantly (P \ 0.05)
lower (101.57 ± 3.65, mean ± SEM) as compared to their
age- and sex-matched healthy controls (141.14 ± 7.77,
mean ± SEM) as shown in Fig. 4. Furthermore, serum
melatonin concentrations were also found significantly
(P \ 0.05) lower (86.89 ± 5.60, mean ± SEM) in shift-
workers as compared to non-shift-workers (107.78 ± 4.11,
mean ± SEM) within the CLL study group (Fig. 5).
Discussion
Since many genes involved in cell cycle are under the
control of circadian clock, maintaining the circadian
rhythms can be a critical control point for cancer devel-
opment. The current study was aimed to indicate associa-
tion between deregulated circadian clock and manifestation
of CLL. CLL is considered to be mainly a disease of the
elderly, with a median age at diagnosis of 70 years [13]
and it has been reported that the CLL disorder is more
common in men with a male to female ratio of approxi-
mately 2:1 [14]. The same observations remained almost
true in the present study as CLL patients included in the
current study mostly constituted elderly population with
the age ranged from 45 to 85 years (mean age of
62.81 ± 10.84) having a male dominance with male to
female ratio of 2.7:1.
According to the most well characterized positive
feedback loop, the transcription of Per1 and Per2 genes is
BMAL1
CLOCKPER1
PER2MYC
Cyclin
D1W
EE1-10
-5
0
5
10
15
20
25
30
35
40
45
50
Genes
Fol
d re
gula
tion
Fig. 2 The bars plot graph
represents the average fold
regulation values of all target
genes in a population of 37 CLL
patients versus 37 age- and sex-
matched healthy controls. The
x-axis represents the gene and
the y-axis represents the fold
regulation of gene expression as
calculated by taking 2-DDCT for
values greater than 1 and
-1/2-DDCT for values
less than 1
Mol Biol Rep (2014) 41:95–103 99
123

directly activated by BMAL1/CLOCK heterodimers [15].
That is why, in our study, down-regulation of Bmal1 is
accompanied with down-regulation of Per1 and Per2 in
CLL patients as compared to healthy controls. Simulta-
neous down-regulation of Bmal1 and Period genes has also
been reported in chronic myeloid leukemia [16] and head
and neck squamous cell carcinoma [17]. Disturbances in
the periodic expression of Cry1, Per1 and Per2 genes have
also been reported recently [18]. A number of studies
revealed that the loss and deregulation of Per genes is
common in cancer. Diminished expression levels of Per1
and Per2 mRNA have been reported in human colorectal
cancer [19]. Mutation of these 2 core clock genes has also
been identified in breast and colorectal cancers [20]. On the
other hand, over-expression of Per1 or Per2 inhibits cancer
cell growth in culture as well as in animals [21–23]. All
this information indicates tumor suppressive nature of Per1
and Per2 [24].
Myc, Cyclin D1 and Wee1 are clock-controlled cell
cycle genes. In the current study, Myc and Cyclin D1 are
found up-regulated whereas Wee1 is found down-regu-
lated. C-Myc is involved in induction of G0–G1 phase
BMAL1PER1
PER2MYC
Cyclin
D1
WEE1
-20
-10
0
10
20
30
40
50
60
Shift worker
Non-Shift worker
Genes
Fol
d re
gula
tion
CLOCK
Fig. 3 The bars plot graph
shows a comparison of the
average fold regulation values
of study genes in shift-workers
versus non-shift-workers within
CLL population. The x-axis
represents the gene and the
y-axis represents the fold
regulation of gene expression as
calculated by taking 2-DDCT for
values greater than 1 and
-1/2-DDCT for values
less than 1
CLL Pat
ients
Health
y0
20
40
60
80
100
120
140
160
180
CLL PatientsHealthy
Mel
aton
in C
onc.
(pg
/ml)
Fig. 4 The bars plot graph shows a comparison of melatonin
concentrations in the serum samples of CLL patients versus their
age- and sex-matched healthy controls. Melatonin concentrations in
CLL patients were found significantly (P B 0.05) lower as compared
to healthy controls
Shift w
orker
Non-S
hift w
orker
0
20
40
60
80
100
120
140
160
180
Non-Shift worker
Shift worker
Mel
aton
in C
onc.
(pg
/ml)
Fig. 5 The bars plot graph shows a comparison of melatonin
concentrations in the serum samples of shift-workers versus non-
shift-workers within the CLL group. Melatonin concentrations in
shift-workers were found significantly (P B 0.05) lower as compared
to non-shift-workers within the CLL group
100 Mol Biol Rep (2014) 41:95–103
123

transition of the cell. It is an oncogene that functions both
in the stimulation of cell proliferation and in apoptosis. C-
Myc elicits its oncogenic activity by causing immortaliza-
tion, and to a lesser extent the transformation of cells, in
addition to several other mechanisms [25]. Virtually, all
types of human cancer manifest high frequencies of
amplification of the c-Myc gene or over-expression of its
protein product [26]. Cyclin D1 is another oncogene that
drives cell cycle progression (G1-S phase transition); it acts
as a growth factor sensor to integrate extracellular signals
with the cell cycle machinery, though it may also promote
apoptosis [25]. Deregulation of Cyclin D1 gene expression
and increased proliferation are hallmarks of a number of
proliferative diseases, including cancer. Cyclin D1 is over-
expressed in many types of human cancer, with gene
amplification in some cases [27]. WEE1 is a cell cycle
kinase that controls the timing of G2–M transition. It is
activated by ongoing DNA replication or by the presence
of DNA damage and inactivates Cdc2/cyclin B through
phosphorylation resulting in the delay of mitosis or arrest
of the cell cycle at the G2–M interface [28]. In case of
Wee1 inhibition, cells would not undergo cell cycle arrest
and mitosis would continue. Reduced expression of Wee1
has been reported in a number of cancers including colon
carcinoma [29] and non-small-cell lung cancer (NSCLC)
[30]. Wee1 and c-Myc are directly regulated by the
BMAL1:CLOCK heterodimer via the E-box elements at
their promoters [8, 9]. Normally, the binding of
BMAL1:CLOCK to the E-boxes of c-Myc promoter
inhibits the transcription of this gene and the binding of
BMAL1:CLOCK to the E-boxes of Wee1 promoter stim-
ulates the transcription of this gene. Thus, reduced level of
BMAL1 may lead to up-regulation of c-Myc transcription
and down-regulation of Wee1 transcription that seems to be
the case in the current study. It has been indicated that
Cyclin D1 is under circadian control in vivo [9]. Cyclin D1
can be directly reduced [31] or indirectly induced by c-Myc
[32]. Cyclin D1 expression was reported to be arrhythmic
and significantly elevated at most times in the bones of
Per1-/-; Per2m/m mice than in wild-type (wt) mice.
BMAL1/CLOCK was found to inhibit the promoter
activity of c-Myc, a critical regulator of Cyclin D1. Con-
sequently, c-Myc expression was elevated in Per1-/-;
Per2m/m bones at most time points studied and in Per1-/-;
Per2m/m osteoblasts. Thus, one mechanism whereby clock
genes inhibit osteoblast proliferation is the down-regula-
tion of c-Myc expression, although other mechanisms may
exist [33]. In a recent study, down regulation of Bmal1
gene expression was found to accelerate cell proliferation
in vitro and promote tumor growth in mice. Suppressing
Bmal1 expression in murine colon cancer cells (C26) and
fibroblast cells (L929) was reported to cause decreased
apoptosis. Loss of Bmal1 led to the reduced expression of
Per1, Per2, Per3, Wee1 and p53. The expression of p21
and c-Myc was also found up-regulated in certain cell lines
(IEC—intestinal epithelial cells). However, BMAL1 defi-
ciency was reported to increase the protein levels of Cdc2,
Cyclin B1, Cyclin D1 and Cyclin E [34].
Bmal1 epigenetic inactivation contributes to the
development of hematologic malignancies such as diffuse
large B-cell lymphoma and acute lymphocytic and mye-
loid leukemias by disrupting the cellular circadian clock.
Bmal1 epigenetic inactivation impairs the characteristic
circadian clock expression pattern of genes such as c-Myc
with a loss of BMAL1 occupancy in their respective
promoters. Furthermore, the DNA hypermethylation-
associated loss of BMAL1 also prevents the recruitment
of its natural partner, the CLOCK protein, to their com-
mon targets [35]. However, in the current study it is not
known whether the down-regulation of Bmal1 in CLL
was the result of any epigenetic phenomena. Another
study showed that hypermethylation in the Clock pro-
moter was found to reduce breast cancer risk, and these
findings were corroborated by publicly available tissue
array data, which showed lower levels of Clock expres-
sion in healthy controls relative to normal or tumor tissue
from breast cancer patients [36]. In our study, Clock
expression was also found higher in CLL patients as
compared to healthy controls but it was statistically
insignificant (P [ 0.05). Further investigations in a bigger
study population may help to get the more lucid picture
about the expression of Clock gene in CLL.
In current study, among 37 CLL patients, *30 % were
shift-workers. Shift-work has been reported as a risk factor
for cancer in many studies [37]. Women working more
than 20 years of rotating nightshifts were found to have a
significantly increased risk of endometrial cancer [38]. A
significant association between rotating shift-work and
prostate cancer incidence among Japanese male workers
has also been reported [39]. According to another study full
time rotating shift-work was found to be associated with
increased risk of prostate cancer [40]. Non-Hodgkin’s
lymphoma was found to be modestly associated with
nighttime work among men with high exposure [41]. In the
current study, expression levels of the genes were found
more aberrant in shift-workers as compared to non-shift-
workers. Bmal1, Per1 and Per2 were more down-regulated
whereas Myc and Cyclin D1 were more up-regulated in
shift-workers as compared to non-shift-workers within the
CLL group. When serum melatonin concentrations were
compared between CLL patients and healthy controls,
significantly low melatonin levels were found in CLL
patients. Moreover, melatonin levels were found still lower
in shift-workers as compared to non-shift-workers within
the CLL group. Circulating melatonin level can be con-
sidered as a biomarker of circadian disruption and has been
Mol Biol Rep (2014) 41:95–103 101
123

associated with night-shift-work and exposure to light-at-
night in both laboratory-based and field studies [42]. There
is credible evidence that a low level of melatonin is asso-
ciated with an increased risk of prostate [43] and breast
cancer [44].
Conclusion
Our results indicate that deregulated expression of core
clock genes may result in deregulated expression of CCGs
that are involved in cell proliferation and apoptosis and
hence may play a role in etiology of CLL. Furthermore,
shift-work and low melatonin levels may also contribute in
further perturbing of circadian clock and hence in mani-
festation of CLL. Additional research needs to be carried
out to elucidate the mechanisms by which shift-work and
low melatonin levels contribute in perturbing the circadian
clock in CLL.
Acknowledgments This work was supported by an HEC (Higher
Education Commission of Pakistan)-funded Project ‘‘Centre for
Research in Endocrinology and Reproductive Sciences’’ (CRERS) in
University of Health Sciences, Lahore, Pakistan.
References
1. Reppert SM, Weaver DR (2002) Coordination of circadian timing
in mammals. Nature 418:935–941
2. Hastings MH, Reddy AB, Maywood ES (2003) A clockwork
web: circadian timing in brain and periphery, in health and dis-
ease. Nat Rev Neurosci 4:649–661
3. Geyfman M, Andersen B (2009) How the skin can tell time.
J Invest Dermatol 129:1063–1066
4. Shearman LP, Sriram S, Weaver DR, Maywood ES, Chaves I,
Zheng B, Kume K, Lee CC, van der Horst GT, Hastings MH,
Reppert SM (2000) Interacting molecular loops in the mamma-
lian circadian clock. Science 288:1013–1019
5. Takahashi JS (2004) Finding new clock components: past and
future. J Biol Rhythms 19:339–347
6. Fu L, Lee CC (2003) The circadian clock: pacemaker and tumor
suppressor. Nat Rev Cancer 3:350–361
7. Duffield GE (2003) DNA microarray analyses of circadian tim-
ing: the genomic basis of biological time. J Neuroendocrinol
15:991–1002
8. Matsuo T, Yamaguchi S, Mitsui S, Emi A, Shimoda F, Okamura
H (2003) Control mechanism of the circadian clock for timing of
cell division in vivo. Science 302:255–259
9. Fu L, Pelicano H, Liu J, Huang P, Lee C (2002) The circadian
gene Period2 plays an important role in tumor suppression and
DNA damage response in vivo. Cell 111:41–50
10. Dohner H, Stilgenbauer K, Dohner M (1999) Chromosome
aberrations in B-cell chronic lymphocytic leukemia: reassessment
based on molecular cytogenetic analysis. J Mol Med 77:266–281
11. Caligaris-Cappio F, Hamblin TJ (1999) B-cell chronic lympho-
cytic leukemia: a bird of a different feather. J Clin Oncol
17:399–408
12. Livak KJ, Schmittgen TD (2001) Analysis of relative gene
expression data using real-time quantitative PCR and the
2(-delta delta C(T)) method. Methods 25:402–408
13. Smith A, Howell D, Patmore R (2011) Incidence of haemato-
logical malignancy by sub-type: a report from the Haematologi-
cal Malignancy Research Network. Br J Cancer 105:1684–1692
14. Sgambati M, Linet MS, Devesa SS (2001) Chronic lymphocytic
leukemia epidemiological, familial, and genetic aspects. In:
Cheson BD (ed) Chronic lymphoid leukemia s basic and clinical
oncology. Marcel Dekker, New York, pp 33–62
15. Takahashi JS, Hong HK, Ko CH, McDearmon EL (2008) The
genetics of mammalian circadian order and disorder: implications
for physiology and disease. Nat Rev Genet 9:764–775
16. Yang MY, Chang JG, Lin PM, Tang KP, Chen YH, Lin HY, Liu
TC, Hsiao HH, Liu YC, Lin SF (2006) Down-regulation of cir-
cadian clock genes in chronic myeloid leukemia: alternative
methylation pattern of hPER3. Cancer Sci 97:1298–1307
17. Hsu CM, Lin SF, Lu CT, Lin PM, Yang MY (2012) Altered
expression of circadian clock genes in head and neck squamous
cell carcinoma. Tumour Biol 33:149–155
18. Eisele L, Prinz R, Klein-Hitpass L, Nuckel H, Lowinski K,
Thomale J, Moeller LC, Duhrsen U, Durig J (2009) Combined
PER2 and CRY1 expression predicts outcome in chronic lym-
phocytic leukemia. Eur J Haematol 83:320–327
19. Mostafaie N, Kallay E, Sauerzapf E, Bonner E, Kriwanek S,
Cross HS, Huber KR, Krugluger W (2009) Correlated down-
regulation of estrogen receptor beta and the circadian clock
gene Per1 in human colorectal cancer. Mol Carcinog
48:642–647
20. Sjoblom T, Jones S, Wood LD, Parsons DW, Lin J, Barber TD,
Mandelker D, Leary RJ, Ptak J, Silliman N, Szabo S, Buckhaults
P, Farrell C, Meeh P, Markowitz SD, Willis J, Dawson D,
Willson JK, Gazdar AF, Hartigan J, Wu L, Liu C, Parmigiani G,
Park BH, Bachman KE, Papadopoulos N, Vogelstein B, Kinzler
KW, Velculescu VE (2006) The consensus coding sequences of
human breast and colorectal cancers. Science 314:268–274
21. Gery S, Komatsu N, Baldjyan L, Yu A, Koo D, Koeffler HP
(2006) The circadian gene per1 plays an important role in cell
growth and DNA damage control in human cancer cells. Mol Cell
22:375–382
22. Hua H, Wang Y, Wan C, Liu Y, Zhu B, Yang C, Wang X, Wang
Z, Cornelissen Guillaume G, Halberg F (2006) Circadian gene
mPer2 overexpression induces cancer cell apoptosis. Cancer Sci
97:589–596
23. Gery S, Gombart AF, Yi WS, Koeffler C, Hofmann WK, Koeffler
HP (2005) Transcription profiling of C/EBP targets identifies
Per2 as a gene implicated in myeloid leukemia. Blood
106:2827–2836
24. Yang X, Wood PA, Ansell C, Hrushesky WJ (2009) Circadian
time-dependent tumor suppressor function of period genes. Integr
Cancer Ther 8:309–316
25. Liao DJ, Thakur A, Wu J, Biliran H, Sarkar FH (2007) Per-
spectives on c-Myc, Cyclin D1, and their interaction in cancer
formation, progression, and response to chemotherapy. Crit Rev
Oncog 13:93–158
26. Dang CV, O’Donnell KA, Zeller KI, Nguyen T, Osthus RC, Li F
(2006) The c-Myc target gene network. Semin Cancer Biol
16:253–264
27. Fu M, Wang C, Li Z, Sakamaki T, Pestell RG (2004) Mini-
review: Cyclind1: normal and abnormal functions. Endocrinology
145:5439–5447
28. Sancar A, Lindsey-Boltz LA, Unsal-Kacmaz K, Linn S (2004)
Molecular mechanisms of mammalian DNA repair and the DNA
damage checkpoints. Annu Rev Biochem 73:39–85
29. Backert S, Gelos M, Kobalz U, Hanski ML, Bohm C, Mann B,
Lovin N, Gratchev A, Mansmann U, Moyer MP, Riecken EO,
Hanski C (1999) Differential gene expression in colon carcinoma
cells and tissues detected with a cDNA array. Int J Cancer
82:868–874
102 Mol Biol Rep (2014) 41:95–103
123

30. Yoshida T, Tanaka S, Mogi A, Shitara Y, Kuwano H (2004) The
clinical significance of Cyclin B1 and Wee1 expression in non-
small-cell lung cancer. Ann Oncol 15:252–256
31. Mateyak MK, Obaya AJ, Sedivy JM (1999) C-Myc regulates
cyclin D-Cdk4 and -Cdk6 activity but affects cell cycle progression
at multiple independent points. Mol Cell Biol 19:4672–4683
32. Marhin WW, Hei YJ, Chen S, Jiang Z, Gallie BL, Phillips RA,
Penn LZ (1996) Loss of Rb and Myc activation co-operate to
suppress cyclin D1 and contribute to transformation. Oncogene
12:43–52
33. Fu L, Patel MS, Bradley A, Wagner EF, Karsenty G (2005) The
molecular clock mediates leptin-regulated bone formation. Cell
122:803–815
34. Zeng ZL, Wu MW, Sun J, Sun YL, Cai YC, Huang YJ, Xian LJ
(2010) Effects of the biological clock gene Bmal1 on tumour
growth and anti-cancer drug activity. J Biochem 148:319–326
35. Taniguchi H, Fernandez AF, Setien F, Ropero S, Ballestar E,
Villanueva A, Yamamoto H, Imai K, Shinomura Y, Esteller M
(2009) Epigenetic inactivation of the circadian clock gene
BMAL1 in hematologic malignancies. Cancer Res 69:8447–8454
36. Hoffman AE, Yi CH, Zheng T, Stevens RG, Leaderer D, Zhang
Y, Holford TR, Hansen J, Paulson J, Zhu Y (2010) CLOCK in
breast tumorigenesis: evidence from genetic, epigenetic, and
transcriptional profiling analyses. Cancer Res 70:1459–1468
37. Straif K, Baan R, Grosse Y, Secretan B, El Ghissassi F, Bouvard
V, Altieri A, Benbrahim-Tallaa L, Cogliano V (2007) Carcino-
genicity of shift-work, painting, and fire-fighting. Lancet Oncol
8:1065–1066
38. Viswanathan AN, Hankinson SE, Schernhammer ES (2007)
Night shift work and the risk of endometrial cancer. Cancer Res
67:10618–10622
39. Kubo T, Ozasa K, Mikami K, Wakai K, Fujino Y, Watanabe Y,
Miki T, Nakao M, Hayashi K, Suzuki K, Mori M, Washio M,
Sakauchi F, Ito Y, Yoshimura T, Tamakoshi A (2006) Prospec-
tive cohort study of the risk of prostate cancer among rotating-
shift workers: findings from the Japan collaborative cohort study.
Am J Epidemiol 164:549–555
40. Conlon M, Lightfoot N, Kreiger N (2007) Rotating shift work and
risk of prostate cancer. Epidemiology 18:182–183
41. Lahti TA, Partonen T, Kyyronen P, Kauppinen T, Pukkala E
(2008) Night-time work predisposes to non-Hodgkin lymphoma.
Int J Cancer 123:2148–2151
42. Mirick DK, Davis S (2008) Melatonin as a biomarker of cir-
cadian dysregulation. Cancer Epidemiol Biomarkers Prev
17:3306–3313
43. Bartsch C, Bartsch H, Schmidt A, Ilg S, Bichler KH, Fluchter SH
(1992) Melatonin and 6-sulfatoxymelatonin circadian rhythms in
serum and urine of primary prostate cancer patients: evidence for
reduced pineal activity and relevance of urinary determinations.
Clin Chim Acta 209:153–167
44. Schernhammer ES, Hankinson SE (2009) Urinary melatonin
levels and postmenopausal breast cancer risk in the Nurses’
Health Study cohort. Cancer Epidemiol Biomarkers Prev
18:74–79
Mol Biol Rep (2014) 41:95–103 103
123