designing and engineering of dna-vaccine construction encoding multiple ctl-epitopes of major hiv-1...
TRANSCRIPT

Vaccine 22 (2004) 1672–1682
Designing and engineering of DNA-vaccine construction encodingmultiple CTL-epitopes of major HIV-1 antigens
Sergei I. Bazhana,∗, Pavel A. Belavina, Sergei V. Seregina, Nadezhda K. Danilyuka,Irina N. Babkinaa, Larisa I. Karpenkoa, Nadezhda A. Nekrasovaa, Leonid R. Lebedeva,
George M. Ignatyeva, Alexander P. Agafonova, Vera A. Poryvaevab,Irina V. Abornevab, Alexander A. Ilyicheva
a The State Research Center of Virology and Biotechnology Vector, 630559 Koltsovo, Novosibirsk Region, Russiab The Joint-Stock Company Vector Best, 630559 Koltsovo, Novosibirsk Region, Russia
Abstract
A synthetic T cell immunogen (TCI) has been designed as a candidate DNA-based vaccine against Human immunodeficiency virus(HIV)-1 using cytotoxic T lymphocytes (CD8+ CTL) and T-helper lymphocytes (CD4+ Th) epitopes retrieved from theLos AlamosHIV Molecular Immunology Database. The protein 392 amino acids in length contains about eighty CTL-epitopes, many of which areoverlapping and are totally restricted by ten different HLA class I molecules. To be able to detect CTL responses induced by a DNA vaccinein experimental animals, additional epitopes, restricted by mouse andMacaque rhesusmajor histocompatibility complex (MHC) class Imolecules, were included in the target immunogen. The gene encoding the TCI protein was assembled, cloned into vector plasmids andexpressed in a prokaryotic and a eukaryotic system. The presence of HIV-1 protein fragments in the immunogen structure was ascertainedby ELISA and immunoblotting using panels of HIV-1-positive sera and monoclonal antibodies to p24. It has been demonstrated thatDNA vaccine can induce both specific T cell responses (CTL and blast transformation) and specific antibodies in mice immunized withpcDNA-TCI.© 2004 Elsevier Ltd. All rights reserved.
Keywords:Human immunodeficiency virus (HIV); Cytotoxic T lymphocytes (CTL); T-helper lymphocytes (Th); HIV CTL-epitopes; Designing apoly-CTL-epitope-based immunogen; Synthetic gene expression; DNA vaccine; Polymerase chain reaction (PCR); Immunogenicity
1. Introduction
One of the promising approaches to the development ofa new generation of effective and safe vaccines is based onidentification of T and B cell epitopes in virus proteins foruse as a basis for synthetic polyepitope vaccines[1,2]. Itis expected that these vaccines will have none of the manydrawbacks that are inherent to the those based on native viralproteins or live attenuated or whole inactivated pathogens.In particular, synthetic polyepitope vaccines will have noregions that induce immunopathology or inhibit protectiveimmunity.
To provide protection effectively, a vaccine should beable to induce both humoral and cell immune responses.Although new promising approaches to the induction ofneutralizing antibodies are still being developed[3], the
∗ Corresponding author. Tel.:+7-3832-366479; fax:+7-3832-366479.E-mail address:[email protected] (S.I. Bazhan).
mainstream is the induction of cell immunity[4–8]. And thatis for a reason, because convincing evidence exists that theresponses of cytotoxic T lymphocytes (CD8+ CTL) associ-ated with HIV infection are important mediators of antiviralimmunity and, therefore, induction of HIV-specific CTLscould be an important component of an effective vaccineagainst HIV-1[9,10]. For example, a reliable negative corre-lation has been found between the frequency of HIV-specificCTLs and the amount of virus RNA in the blood ofHIV-infected individuals[11]. It is hypothesized that CTLsare capable of protecting the host against HIV infection, be-cause they would kill HIV-infected cells before they couldproduce new virions[12] and, furthermore, CTLs wouldrelease chemokines, which inhibited HIV infection[13,14].
This work describes the engineering of a syntheticpoly-CTL-epitope T cell immunogen (TCI), which is con-sidered to be a candidate for use as an effective and safeDNA vaccine against HIV-1. In summary, we have: (i)designed a poly-CTL-epitope immunogen; (ii) synthesized
0264-410X/$ – see front matter © 2004 Elsevier Ltd. All rights reserved.doi:10.1016/j.vaccine.2003.09.048

S.I. Bazhan et al. / Vaccine 22 (2004) 1672–1682 1673
the gene encoding the target protein; (iii) produced recom-binant plasmids for the target gene to be expressed in aprokaryotic and a eukaryotic system; (iv) obtained proofthat the expression products have antigen specificity andimmunogenicity.
2. Materials and methods
2.1. Bacterial strains, plasmids, enzymes, oligonucleotides
We used the bacterial strainsEscherichia coliJM109,AD494(DE3) (Novagen, USA) and XL1-blue (Vector, Rus-sia); vector plasmids pFH123[15]; pcDNA3.1/Myc-His(-)/lacz (Invitrogen, USA), pET32a (Novagen, USA), andpGEX-4T-1 (Pharmacia, Sweden); restriction endonucle-ases, T4-DNA-ligase, Taq-DNA-polymerase (Sibenzyme,Russia); oligonucleotides (Vector, Russia), HIV-1 cDNA[16]; a diagnostic test system HIV-Express, a panel of hu-man sera against HIV-1 and MABs against the EPFRDYV-DRFYKTLR determinant located in HIV-1 protein p24(Joint-Stock Company Vector Best, Russia). Genetic en-gineering procedures were as described in[17]. Sequenc-ing of DNA fragments as part of recombinant plasmidswas performed according to Sanger and Coulson[18].Oligonucleotide primers for PCR and reaction conditionswere determined using the OLIGO software program[19].
2.2. Expression of the TCI gene in E. coli cells
To be able to ascertain the expression of the TCIgene in E. coli cells, recombinant plasmids pET-TCIand pGEX-TCI were assembled by cloning this genein the form of the BamHI-SalI fragment of plasmidpFH-TCI into vector plasmids pET32a/BamHI-SalI andpGEX-4T-1/BamHI-SalI, respectively. CompetentE. coliJM109/pET-TCI andE. coli XL1-blue/pGEX-TCI cellswere transformed with these plasmids. The presence of thetarget chimeric TRN-TCI and GST-TCI proteins inE. colicells induced by IPTG was ascertained by electrophoresisunder denaturing conditions[20]. The chimeric TRN-TCIand GST-TCI proteins were isolated from bacterial cells byaffinity chromatography on Ni-NTA agarose (QIAexpressProtein Purification System, QIAGEN) as recommendedby the manufacturer. The protein was obtained at 90–95%purity.
2.3. Polyclonal antibodies to the TRN-TCI protein
Mongrel rabbits were immunized i.m. three times at a2-week interval with the purified TRN-TCI protein at a doseof 100�g per animal. Antibody titers were measured oncea week. Higher-titer sera were collected and used for im-munoblotting of recombinant proteins.
2.4. Assessment of HIV antigenicity of the GST-TCIand TRN-TCI recombinant proteins by ELISA andimmunoblotting
ELISA was performed using 96-well polystyrene plates.Protein solution (1 mg/ml) was sorbed in a volume of 100�lper well at a dilution of 1:100, 1:200, and 1:500. Frag-ments of HIV-1 proteins supplied as part of commercialtest-systems (Vector-Best Ltd., Russia) were used as a pos-itive control, recombinant GST and TRN proteins and son-icatedE. coli cells were used as a negative control.
Immunoblotting was performed using MABs 29F2 and30A6 to HIV-1 p24 protein and polyclonal anti-TRN-TCIrabbit antibodies. The immune response was ascertained us-ing anti-mouse IgG rabbit antibodies conjugated to alkalinephosphatase and anti-rabbit IgG goat antibodies conjugatedto alkaline phosphatase.
2.5. Construction of a plasmid for cloning the TCI geneinto a eukaryotic system
Vector pcDNA3.1/Myc-His(-)/lacz (Invitrogen, USA) wasused for cloning the TCI gene in a eukaryotic system. Inorder to achieve the optimal expression of the target TCIgene, first, the plasmid was deleted for the plasmid frag-ment containing theE. coli lacZgene region, secondly, theKozak consensus sequence was included before the ATGstart codon. The resulting recombinant plasmid pcDNA-TCIwas used for DNA immunization and for assessing the im-munogenicity of the vaccine construct.
2.6. Immunization of mice
The immunogenicity of the vaccine construct was as-sessed using BALB/c mice, weighing 10–12 g, from Vector’sanimal breeding facilities. Immunization with plasmid con-struct pcDNA-TCI was performed by i.m. injection in therear paws with no adjuvant used. Two doses of plasmid DNAwere used: 20 and 100�g. Reimmunization was performed 1month later. In order to measure antibody titers (ELISA), theproliferative activity of lymphocytes (blast transformation),and CTL levels (ELISPOT), blood and splenocyte sampleswere collected from four mice 7, 10, 14, 21, 30 and 48 daysafter immunization. Samples from four mice were pooledand assayed by ELISA and blast transformation. ELISPOTassay was carried out for splenocyte samples taken from ev-ery mouse individually. All the work with animals met theprotocols on animal use approved by the IACUC of the SRCVB Vector (NIH OLAW registration no. A5505-01).
2.7. Measuring the proliferative activity of T lymphocytes
Antigen-specific T cell proliferation was assessed by blasttransformation in the splenocytes of immunized and controlanimals using a routine technique[21]. The TCI protein in a

1674 S.I. Bazhan et al. / Vaccine 22 (2004) 1672–1682
concentration of 2�g/ml and its peptides N15 (DRVIEVVQ-GAYRAIR) and N16 (KQIINMWQEVGKAMYA) wereused as the specific antigens. In a murine system, thesepeptides are multiple class I molecules, which can be pre-sented to CTL. The EHEC antigen (uninfectedE. colistrain O:H157) in a concentration of 2�g/ml was used asthe nonspecific antigen (negative control). Concanavalin A(5�g/ml, Sigma, USA) was used as the mitogen. The pro-liferative activity of the splenocytes was estimated as thestimulation index calculated as the ratio of mean absoluteDeltaOD of stimulated to non-stimulated splenocytes.
2.8. Measuring antibody titers by ELISA
ELISA was run with the TCI protein, a mixture of therecombinant Gag and Env proteins and HIV-1 lysate as theantigens in a 96-well CORNING plate. Fifty�l of the anti-gen in a concentration of 1�g per well was adsorbed for15 h at 4◦C. Titrations were performed at serum dilutionsfrom 1:10 to 1:2560. Anti-mouse IgG antibodies conjugatedto horseradish peroxidase (Sigma, USA) was used.
2.9. Measuring CTL by ELISPOT
Anti-INF-� MABs were sorbed in a concentration of5�g/ml per well in a 96-well ImmunoSpot M200 plate.Upon incubation for 12 h at 4◦C, each well was washedtwice with PBS, after which RPMI 1640 growth mediumcontaining 10% fetal bovine serum was added for 2 h.Splenocytes from immunized animals in a concentration of106 ml−1 served as the effector cells. Production of INF-�by cell suspension was stimulated using 1�g/ml TCI pro-tein and peptides N15 (DRVIEVVQGAYRAIR) and N16(KQIINMWQEVGKAMYA); non-specific products wereidentified using the EHEC peptide (negative control).
3. Results
3.1. Design of a synthetic immunogen containing multipleCTL-epitopes of the major HIV-1 antigens
As is known, HIV-1 induces CD8+ CTL responses bypresenting its antigens together with major histocompatibil-ity complex (MHC) class I molecules on the surface of in-fected cells. In so doing, the viral antigens are not recog-nized by specific T lymphocytes as full-length proteins butshort peptides (8–12 amino acids) associated with specificMHC class I molecules[22,23]. These short antigenic epi-topes originate from cytoplasmic viral proteins as a resultof proteasome-mediated processing[24,25]. Since peptidesbind to MHC molecules specifically, only certain peptidescan be presented by a particular type of MHC molecules.
The amino acid sequences of the CTL-epitopes impli-cated in the induction of HIV-1-specific CTL responses
in infected individuals are present in theLos Alamos HIVMolecular Immunology Database. In order to be chosen forthe CTL-immunogen design, the epitopes were supposed tomeet the following eligibility criteria:
1. They induce both CD8+ CTL and CD4+ Th and representthree major HIV-1 subtypes-A, B, and C—occurring inRussia, Western Europe, and the USA.
2. They belong to the major viral proteins-antigens: Env,Gag, Pol, and Nef.
3. All the CD8+ CTL-epitopes are restricted by 10 dif-ferent optimally selected HLA class I molecules. As isknown, this is sufficient for covering the genetic diver-sity of MHC class I antigens in whatever geographicalregion[6,26,27].
4. They may not induce autoimmune antibodies. To meetthis criterion, the HIV-1 protein regions that have localhomology with human proteins had been identified andthe corresponding epitopes excluded from consideration[28].
As turned out, most of the successful CTL-epitopes aremapped in the native viral proteins as overlapping peptides,which are located in several continuous regions of the Env,Gag, Pol, and Nef proteins. We included these antigenicallyactive regions in the target T cell immunogen. As we havefound out, not only do these regions contain T cell epitopes,but also B cell epitopes, which overlap with them (data notshown). Based on this finding, the TCI protein can be rec-ognized by HIV-1-positive sera.
A general schematic of the design of the TCI immunogenis presented inFig. 1. As can be seen, the target protein is anarrangement of p17, p24, gp120, gp41, Pol, and Nef proteinfragments depicted inFig. 1. The order of the fragments is ofno importance, because the target immunogen is presented tothe immune system in the form of short peptides associatedwith MHC class I molecules (see above). The TCI proteinis 392 amino acids in length and contains more than eightyCD8+ CTL and CD4+ Th epitopes restricted by HLA class Iand II alleles. To be able to study CTL responses induced bya DNA vaccine in experimental animals, additional epitopes,which were restricted by mouse andMacaque rhesusMHCclass I molecules, were included in the target immunogen(Fig. 1).
3.2. Assembly of a synthetic gene
The TCI gene was synthesized by its assembling fromthree synthetic DNA fragments (blocks 1–3). A generalschematic of the process is presented inFig. 2. The pri-mary structure of the 1176 bp TCI gene and the correspond-ing amino acid sequence are presented inFig. 3. Fragments(blocks) 1–3 were synthesized by combining a matrix anda matrix-free technique using PCR. Either the long HIV-1cDNA (HIV-1 strain EVK [16]) regions containing the se-lected overlapping CTL-epitopes of a particular protein orpartially complementary oligonucleotide primers were used

S.I. Bazhan et al. / Vaccine 22 (2004) 1672–1682 1675
Fig. 1. Design of the CTL immunogen, a candidate for use as an anti-HIV-1 vaccine: a general schematic. Bar patterns indicate the polyepitope CTLimmunogen and the origin of the sequences. The positions of individual epitopes and their MHC restrictions (HLA-A, B, Cw—human; H-2a, b, d, f, k,p, u, q—mouse; Mamu-A∗01—Macaca mulatta) are depicted as lines below the CTL immunogen. Th stands for helper epitopes.
as the matrix. The resulting TCI DNA fragments 1–3 werecloned into vector plasmid pFH123[15] using theBamHIandSalI restriction sites inE. coli XL1-blue cells. This plas-mid carries a specific polylinker with a site forFokI restric-tase, which allows DNA to be produced with unique “sticky”5′ ends after intermediate cloning. As a result, we had re-combinant plasmids pFH-I, pFH-II, and pFH-III containingthe target TCI fragments.
In order to produce plasmid pFH-TCI containing the tar-get TCI gene, we used vector pFH123/BamHI-SalI describedabove and three TCI gene fragments isolated from plas-mids pFH/I (BamHI-FokI), pFH/II (FokI-FokI), and pFH/III(FokI-SalI). Here the structural changes in the 5′ ends of theTCI fragments 1–3 allowed these fragments to be easily un-ambiguously assembled into a full-length TCI gene at thefinal cloning stage. The authenticity of the synthesized TCIgene was confirmed by sequencing.
3.3. Immunochemical study of the TCI gene product
In order to ensure that the target gene does express HIV-1epitopes, the TCI gene was cloned into expression vectorspGEX-2T and pET-32a inE. coli cells. The immunogenicproperties of the synthesized TCI protein associated bothwith GST and TRN-buffer proteins were examined byELISA using a panel of HIV-1-positive human sera (10samples) and a panel of HIV-1-negative human sera (10blood samples). Both chimeric proteins were absolutely de-tectable on the positive serum panel; however they had no
binding activity to any of the negative sera. Therefore, theTCI protein has HIV-1 antigen activity indeed.
The antigenic properties of the TCI protein were alsoexamined by immunoblotting (Fig. 4) using MABs 29F2 and30A6, which bind to the EPFRDYVDRFYKTLR epitope ofHIV-1 p24 and TCI. As can be seen, the expression productsof plasmid pET-TCI (the TRN-TCI protein) and plasmidpGEX-TCI (the GST-TCI protein) bind to MABs to HIV-1p24 and, therefore, these proteins and the p24 protein havea shared epitope (Fig. 4B). This conclusion is supported bythe results presented inFig. 4C. The serum of the rabbitimmunized with the TCI-TRN protein is reactive not onlywith both recombinant proteins, TRN-TCI and GST-TCI,but also recognizes the native HIV-1 protein p24.
3.4. Study of the immunogenicity of plasmid DNAencoding the TCI protein
In order to assess the immunogenicity of plasmid DNAencoding poly-CTL-epitope immunogen, TCI, recombinantplasmid pcDNA-TCI was used. The following parametersof the immune response were measured: the CTL levels, theproliferative activity of the lymphocytes, and the antibodytiters.
3.4.1. Quantification of cytotoxic T lymphocytes inimmunized animals
CTL responses were assessed by quantification ofIFN-�-producing lymphocytes revealed by an ELISPOT

1676 S.I. Bazhan et al. / Vaccine 22 (2004) 1672–1682
Fig. 2. A schematic set-up of the synthesis and assembly of TCI gene. PCR stages (vertical arrows) are numbered by Roman numerals.Oligonucleotides—primers (horizontal arrows) are numbered by Arabic numerals. Alternating black and gray bars within TCI fragments stand for thesequences of HIV-1 cDNA encoding fragments of viral proteins that were selected for designing the TCI immunogen.

S.I. Bazhan et al. / Vaccine 22 (2004) 1672–1682 1677
Fig. 3. Nucleotide and amino acid sequences of the TCI gene and TCI-encoded protein.
assay. Results are presented inFig. 5 and Table 1. Basedon analysis of the immunogenicity of the constructs, thefollowing conclusions were made.
First, plasmid DNA, which encodes the TCI protein,can induce specific CTLs in immunized animals in re-
sponse to in vitro stimulation of splenocytes by bothfull-length TCI and separate epitopes (peptides N15 andN16).
Secondly, a dose-dependent effect is observed after im-munization with pcDNA-TCI. An earlier, stronger and more

1678 S.I. Bazhan et al. / Vaccine 22 (2004) 1672–1682
Fig. 4. Electrophoresis and immunoblotting of the recombinant GST-TCI and TRN-TCI proteins: (A) electrophoresis in 13% polyacrylamide gel; (B)immunoblotting using MABs to p24 protein; (C) immunoblotting using the serum of a rabbit immunized with the TCI-TRN protein. (1) lysate ofE. colicells carrying plasmid pGEX-TCI; (2) purified p24 protein of HIV-1; (3) lysate ofE. coli cells carrying plasmid pET-TCI; (4) lysate ofE. coli cellswithout plasmid; (5) molecular weight markers.
prolonged CTL response is induced by plasmid DNA in adose of 100 than 20�g.
Reimmunization of animals, which were boosted by plas-mid pcDNA-TCI in a dose of 50�g, enhances markedlyCTL responses. That was even more obvious with the an-imals primed with the same plasmid in a dose of 100�g(Table 1).
Table 1Cytotoxic activity of splenocytes elicited from mice after single or prime-boost immunization with plasmid pcDNA-TCI
Days after immunization Immunization schedules Number of�-IFN-producing cells (M ± m)a
Specific antigensb
TCI N15 N16
14 20 mkg (single) 21.40± 5.83 14.60± 1.63 19.0± 4.58100 mkg (single) 26.00± 3.45 17.80± 1.02 21.6± 3.47P < – – –
21 20 mkg (single) 2.75± 1.11 2.75± 1.03 1.25± 0.75100 mkg (single) 27.50± 11.64 11.0± 4.85 17.75± 7.31P < 0.070 – 0.065
30 20 mkg (single) 6.60± 1.21 3.60± 0.25 3.00± 0.63100 mkg (single) 8.20± 1.50 2.20± 0.20 2.40± 0.51P < – – –
48 20 mkg (single) 12.50± 1.44 7.25± 2.72 5.25± 0.48100 mkg (single) 13.67± 3.18 7.33± 0.88 5.67± 1.67P < – – –
48 20 mkg (single) 12.50± 1.44 7.25± 2.72 5.25± 0.4820 mkg (prime)+ 50 mkg (boost) 26.25± 6.30 17.75± 4.59 15.75± 3.96P < 0.077 0.096 0.039
48 100 mkg (single) 13.67± 3.18 7.33± 0.88 5.67± 1.67100 mkg (prime)+ 50 mkg (boost) 26.25± 2.39 21.25± 2.05 18.75± 1.49P < 0.023 0.003 0.021
48 20 mkg (prime)+50 mkg (boost) 26.25± 6.30 17.75± 4.59 15.75± 3.96100 mkg (prime)+ 50 mkg (boost) 26.25± 2.39 21.25± 2.05 18.75± 1.49P < – – –
a ELISPOT assay was carried out for splenocyte samples taken from every mouse individually (n = 4). The results are expressed as the mean (M ±m)numbers of IFN� secreting cells (spots) per 106 splenocytes, where (m) represent the standard errors of means.
b TCI protein, peptides N15 and N16 were used for in vitro restimulated of splenocytes.
3.4.2. Measuring the proliferative activity of splenocytesin immunized animals
The proliferative response of lymphocytes was assessedby blast transformation. Analysis of experimental resultsindicates that the splenocytes from immunized animalsdisplay an increased proliferative activity toward TCI andpeptides N15 and N16 compared to the control animals
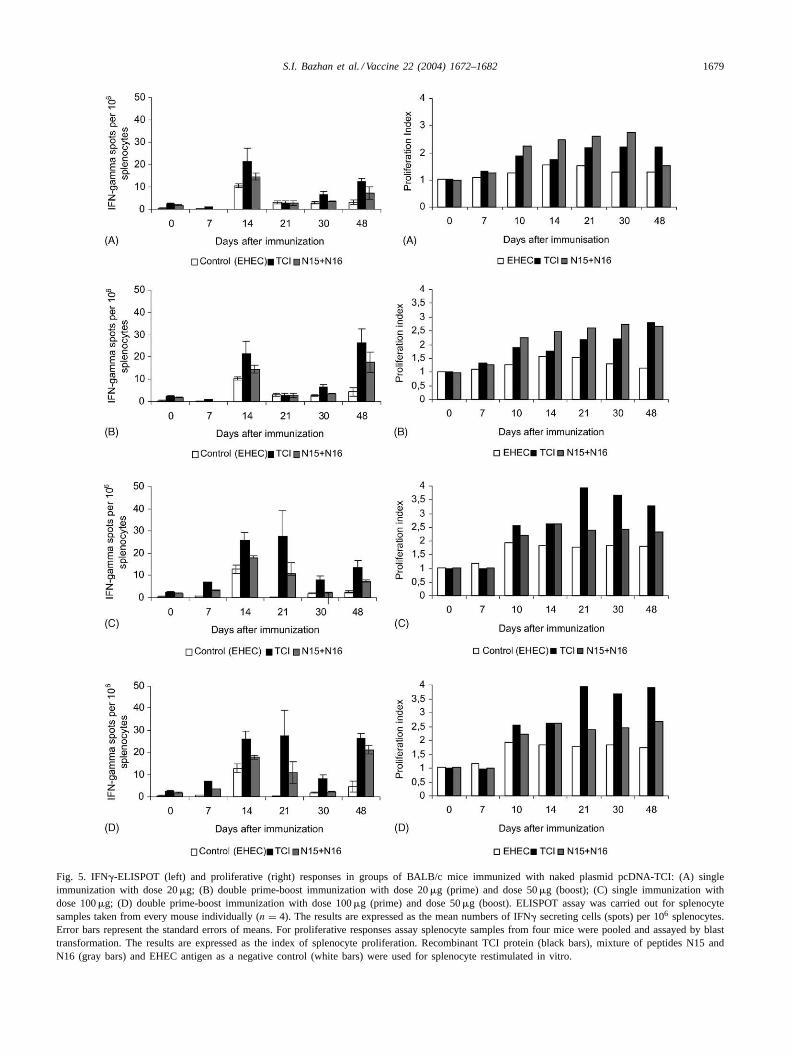
S.I. Bazhan et al. / Vaccine 22 (2004) 1672–1682 1679
Fig. 5. IFN�-ELISPOT (left) and proliferative (right) responses in groups of BALB/c mice immunized with naked plasmid pcDNA-TCI: (A) singleimmunization with dose 20�g; (B) double prime-boost immunization with dose 20�g (prime) and dose 50�g (boost); (C) single immunization withdose 100�g; (D) double prime-boost immunization with dose 100�g (prime) and dose 50�g (boost). ELISPOT assay was carried out for splenocytesamples taken from every mouse individually (n = 4). The results are expressed as the mean numbers of IFN� secreting cells (spots) per 106 splenocytes.Error bars represent the standard errors of means. For proliferative responses assay splenocyte samples from four mice were pooled and assayed by blasttransformation. The results are expressed as the index of splenocyte proliferation. Recombinant TCI protein (black bars), mixture of peptides N15 andN16 (gray bars) and EHEC antigen as a negative control (white bars) were used for splenocyte restimulated in vitro.
1680 S.I. Bazhan et al. / Vaccine 22 (2004) 1672–1682
immunized with vector plasmid pcDNA 3.1 (Fig. 5). Aswith CTL responses, an increase in immunization dose orreimmunization enhances the proliferative activity of thesplenocytes. It should be noted that there was no prolifer-ative response in the immunized animals at any point ofobservation during in vitro stimulation of lymphocytes by a
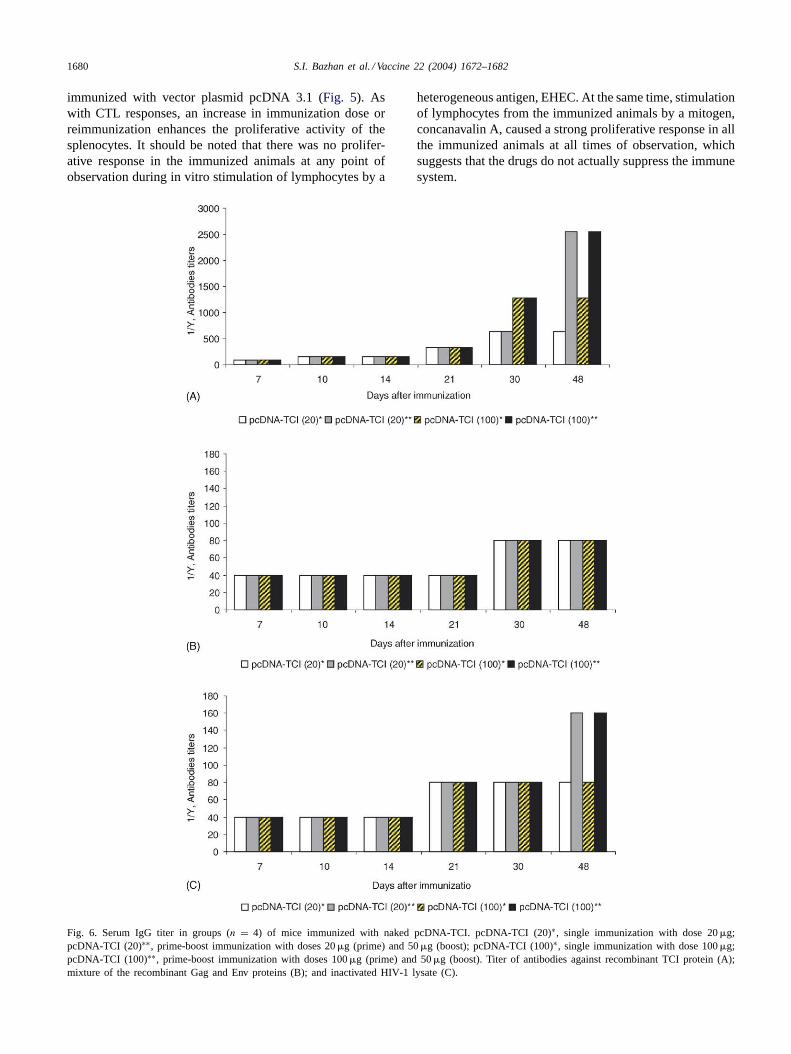
Fig. 6. Serum IgG titer in groups (n = 4) of mice immunized with naked pcDNA-TCI. pcDNA-TCI (20)∗, single immunization with dose 20�g;pcDNA-TCI (20)∗∗, prime-boost immunization with doses 20�g (prime) and 50�g (boost); pcDNA-TCI (100)∗, single immunization with dose 100�g;pcDNA-TCI (100)∗∗, prime-boost immunization with doses 100�g (prime) and 50�g (boost). Titer of antibodies against recombinant TCI protein (A);mixture of the recombinant Gag and Env proteins (B); and inactivated HIV-1 lysate (C).
heterogeneous antigen, EHEC. At the same time, stimulationof lymphocytes from the immunized animals by a mitogen,concanavalin A, caused a strong proliferative response in allthe immunized animals at all times of observation, whichsuggests that the drugs do not actually suppress the immunesystem.

S.I. Bazhan et al. / Vaccine 22 (2004) 1672–1682 1681
3.4.3. Study of antibody titers in the blood sera ofimmunized animals
After DNA immunization with recombinant plasmidpcDNA-TCI, antibody titers were assessed by ELISA usinga number of antigens, including the TCI protein, a mixtureof recombinant proteins, Gag and Env, and HIV-1 viruslysate. Based on the results obtained (Fig. 6), the followingconclusions were made:
1. Immunization of mice with plasmid pcDNA-TCI triggersthe production of specific antibodies that bind both to TCIand the recombinant proteins, Env and Gag, and HIV-1lysate.
2. Titers of the antibodies to all the antigens began to in-crease on day 7–10 post immunization and reached theirmaximum by the end of the observation period (48–55days).
3. In reimmunized animals, antibody titers were about twotimes higher the value revealed in animals followingone-time immunization.
4. Discussion
The successful development of a vaccine against HIV-1is believed by many investigators to be associated with thedevelopment of synthetic vaccine constructs inducing CTLresponses to multiple HIV-1 CTL-epitopes[6–8,29,30]. De-livery of poly-CTL-epitope DNA-based vaccines is normallyperformed using no-protein techniques such as attenuatedviral vectors[29,31], bacterial vector systems[32], or plas-mid DNA [6–8,33]. Recently the safety and potential im-munogenicity of an HIV-directed DNA-based vaccine con-taining envandrev genes was demonstrated[35], a findingthat should encourage further studies. Therefore a consid-erable amount of effort is now being invested into the de-velopment of new synthetic vaccine constructs that warrantan efficient expression and processing of target immuno-gens in the course of DNA-immunization[30,34]. At thesame time, an effective vaccine, besides having the abil-ity to induce high-level cytotoxic T lymphocyte responsesto the selected epitopes, should also warrant the inductionof immune responses to quite a few CTL-epitopes with aparticular HLA specificity. The synthetic poly-CTL-epitopeDNA-based vaccines that are known to date encode about20 HIV-1 CTL-epitopes and as such cannot warrant that avaccine be highly effective against HIV-1.
The DNA-vaccine construct, which was developed interms of the effort being reported, encodes a 392 a.a. protein.The target TCI immunogen contains about 80 CTL-epitopes(both CD8+ CTL and CD4+ Th), which are all togetherrestricted by 10 different HLA-alleles. To be able to studyCTL responses induced by a DNA vaccine in experimentalanimals, additional epitopes, which were mouse andM.rhesusMHC class I molecules, were included in the targetimmunogen.
The most popular strategy for designing polyepitope con-structs is via assembling the epitopes into a linearly orderedbeads-like structure[4,5,34]. We propose an alternative strat-egy, which is via assembling the epitopes into overlappingpeptides[6] in accordance with their location in native virusproteins whenever possible. The sequence of the TCI im-munogen was designed so that its fragments are in the formof clusters of overlapping CTL-epitopes, which correspondto quite long and continuous amino acid sequences of thenative viral proteins Env, Gag, Pol, and Nef. Overlappingallowed us to minimize the total length of the immunogen.
The synthesis of the TCI gene was performed by a com-bined technique using PCR with specially prepared “hybrid”oligonucleotides primers and HIV-1 cDNA. In order to as-certain whether the synthetic TCI protein was immunoac-tive toward HIV-1-positive sera, the gene encoding all theepitopes was cloned into two expression plasmids inE. colicells. The presence of HIV-1 protein fragments in the im-munogen structure was ascertained by ELISA using panelsof HIV-1-positive sera and monoclonal antibodies to p24and immunoblotting.
In order to assess the immunogenicity of the DNA-vaccineconstruct, the gene encoding poly-CTL-epitope immuno-gen, TCI, was cloned into plasmid pcDNA-TCI. Results ob-tained indicate that plasmid pcDNA-TCI can induce bothspecific T cell responses (CTL and blast transformation)and specific antibodies in immunized animals. The use of afull-length TCI protein and separate epitopes (peptides N15and N16) for in vitro re-stimulation of splenocytes indicatethat DNA-immunization incorporates all the stages requiredfor delivery of the target immunogen to the immune system:expression of the TCI gene, processing of the target protein,and presentation of the resulting peptides (determinants) andMHC class I molecules to CD8+ lymphocytes (CTL). It hasbeen demonstrated that specific antibodies bind to both theTCI protein and native and recombinant HIV-1 proteins. Asa results of DNA immunization, first, the TCI gene is ex-pressed in vivo; secondly, the immunochemical propertiesof TCI gene products are identical to those of the recombi-nant TCI protein; and, finally, antibodies to the TCI proteinrecognize HIV-1 proteins–antigens.
It should be noted that in the non-reimmunized animalsboth humoral and cellular responses stop increasing or evendecrease, whereas in the reimmunized animals secondaryresponses have a clear-cut tendency to increase, which isindicative of the formation of specific T and B memory cells.
Acknowledgements
This work was supported by grants from Russia’s StateScientific and Technical Programs “New Generation Vac-cines and Future Medical Diagnostics” and “First Prioritiesin Science and Technology” subprogram “Protection againstPathogens”. We are thankful to Vladimir Filonenko for trans-lating this manuscript from Russian into English.

1682 S.I. Bazhan et al. / Vaccine 22 (2004) 1672–1682
References
[1] Eroshkin AM, Karginova EA, Gileva IP, Lomakin AS, Lebedev LR,Kamyinina TP, et al. Design of four-helical protein as a candidatefor HIV vaccine. Protein Eng 1995;8:167–73.
[2] Loktev VB, Ilyichev AA, Eroshkin AM, Karpenko LI, Pokrovsky AG,Svyatchenko VA, et al. Design of immunogens as components of anew generation of molecular vaccines. J Biotechnol 1996;44:129–37.
[3] Binley JM, Sanders RW, Clas B, Schuelke N, Master A, Guo Y, etal. A recombinant human immunodeficiency virus type 1 envelopeglycoprotein complex stabilized by an intermolecular disulfide bondbetween the gp120 and gp41subunits is an antigenic mimic of thetrimeric virion-associated structure. J Virol 2000;74:627–43.
[4] Thomson S, Khanna R, Gardner J, Burrows SR, Coupar B,Moss DJ, et al. Minimal epitopes expressed in a recombinantpolyepitope protein are processed and presented to CD8+ cytotoxicT cells: Implications for vaccine design. Proc Natl Acad Sci USA1995;92:5845–9.
[5] Thomson S, Elliott SL, Sherritt M, Sproat KW, Coupar BE, ScalzoAA, et al. Recombinant polyepitope vaccines for the delivery ofmultiple CD8+ cytotoxic T cells epitopes. J Immunol 1996;157:822–6.
[6] Hanke T, Schneider J, Gilbert SC, Hill AVS, McMichael A. DNAmulti-CTL epitope vaccines for HIV and Plasmodium falciparum:immunogenicity in mice. Vaccine 1998;16:426–36.
[7] Hanke T, Blanchard TJ, Schneider J, Ogg GS, Tan R, Becker M, et al.Immunogenecities of intravenous and intramuscular administrationof modified vaccinia virus Ankara-based multi-CTL epitope vaccinefor human immunodeficiency virus type 1 in mice. J Gen Virol1998;79:83–90.
[8] Hanke T, Blanchard TJ, Schneider J, Hannan CM, Becker M, GilbertSC, et al. Enhancement of MHC class I-restricted peptide-specificT cell induction by a DNA prime MVA boost vaccination regime.Vaccine 1998;16:439–45.
[9] Rowland-Jones S, Sutton J, Ariyoshi K, Dong T, Gotch F, McAdamS, et al. HIV-specific cytotoxic T-cells in HIV-exposed but uninfectedGambian women. Nat Med 1995;1:59–64.
[10] Letvin NL. Progress in the development of an HIV-1 vaccine. Science1998;280:1875–80.
[11] Ogg GS, Jin X, Bonhoeffer S, Dunbar PR, Nowak MA, Monard S,et al. Quantitation of HIV-1-specific cytotoxic T lymphocytes andplasma load of viral RNA. Science 1998;279:2103–6.
[12] Yang OO, Kalams SA, Trocha A, Cao H, Luster A, Johnson RP, etal. Suppression of human immunodeficiency virus type 1 replicationby CD8+ cells: evidence for HLA class I-restricted triggering ofcytolytic and noncytolytic mechanisms. J Virol 1997;71:3120–8.
[13] Wagner E, Yang OO, Garcia-Zepeda EA, Ge Y, Kalams SA, WalkerBD, et al. Beta-chemokines are release from HIN-1-specific T cellsgranules complexes to proteoglycans. Nature 1998;391:908–11.
[14] Price DA, Sewell AK, Dong T, Tan R, Goulder PJ, Rowland-JonesSL, et al. Antigen-specific release of beta-chemokines by anti-HIV-1cytotoxic T lymphocytes. Curr Biol 1998;8:355–8.
[15] Daniliuk NK, Serpinskii OI, Siniakov AN. Design of recombinant-stable plasmids of the pFH series. Bioorg Khim 1991;17:81–7 (inRussian).
[16] Mamaeva OA, Pliasunova OA, Proniaeva TR, Pokrovskii AG,Tkachev VK, Karamov EV, et al. The characteristics of the long-term
continuous cultivation of a cell line chronically infected with thehuman immunodeficiency virus type-1. Vopr Virusol 1991;36:447–50.
[17] Sambrook J, Fritsch EF, Maniatis T. In: Molecular cloning. 2nd ed.Cold Spring Harbor Laboratory Press; 1989.
[18] Sanger F, Coulson AR. A rapid method for determining sequencesin DNA by primed synthesis with DNA polymerase. J Mol Biol1975;94:441–8.
[19] Rychlik W, Rhoads RE. A computer program for choosing optimaloligonucleotides for filter hybridization, sequencing and in vitroamplification of DNA. Nucl Acids Res 1989;17:8543–51.
[20] Laemmli E. Cleavage of structural proteins during the assembly ofthe head of bacteriophage T4. Nature 1970;227:680–5.
[21] Dittmer U, Luke W, Stahl-Hennig C, Coulibaly C, Petry H, BodemerW, et al. Early helper T-cell disfunction in SIV- but not HIV-2-infected macaques. J Med Primatol 1994;23:21–6.
[22] Oldstone MB. Viral persistence. Cell 1989;56:517–20.[23] Goldberg AL, Rock KL. Proteolysis, proteasomes and antigen
presentation. Nature 1992;357:375–9.[24] York IA, Rock KL. Antigen processing and presentation by the class
I major histocompatibility complex. Ann Rev Immunol 1996;14:369–96.
[25] Rammensee HG, Falk K, Roetzschke O. MHC molecules as peptidereceptors. Curr Opin Immunol 1993;5:35–44.
[26] Lalvani A, Aidoo M, Allsopp CE, Plebanski M, Whittle HC, Hill AV.An HLA-based approach to the design of a CTL-inducing vaccineagainst Plasmodium falciparum. Res Immunol 1994;145:461–8.
[27] Sidney J, Grey HM, Kubo RT, Sette A. Practical, biochemical andevolutionary implication of the discovery of HLA class I supermotifs.Immunol Today 1996;17:261–6.
[28] Maksiutov AZ, Bachinskii AG, Bazhan SI. Searching for localsimilarities between HIV-1 and human proteins. Application tovaccines. Mol Biol (Mosk) 2002;36:447–59 (in Russian).
[29] Woodberry T, Gardner J, Mateo L, Eisen D, Medveczky J, RamshawIA, et al. Immunogenicity of a human immunodeficiency virus(HIV) polytope vaccine containing multiple HLA A2 HIV CD8(+)cytotoxic T-cell epitopes. J Virol 1999;73:5320–5.
[30] Livingston BD, Newman M, Crimi C, McKinney D, Chesnut R, SetteA. Optimization of epitope processing enhances immunogenicity ofmultiepitope DNA vaccines. Vaccine 2001;19:4652–60.
[31] Perkus ME, Tartaglia J, Paoletti E. Poxvirus-based vaccine candidatesfor cancer, AIDS, and other infectious diseases. J Leukoc Biol1995;58:1–13.
[32] Shen H, Slifka MK, Matloubian M, Jensen ER, Ahmed R, Miller JF.Recombinant Listeria monocytogenes as a live vaccine vehicle forthe induction of protective anti-viral cell-mediated immunity. ProcNatl Acad Sci USA 1995;92:3987–91.
[33] Waine GJ, McManus DP. Nucleic acids: vaccines of the future.Parastiol Today 1995;11:113.
[34] Rodriguez F, An LL, Harkins S, Zhang J, Yokoyama M, WideraG, et al. DNA immunization with minigenes: low frequency ofmemory cytotoxic T lymphocytes and inefficient antiviral protectionare rectified by ubiquitination. J Virol 1998;72:5174–81.
[35] MacGregor RR, Boyer JD, Ugen KE, Lacy KE, Gluckman SJ,Bagarazzi ML, et al. First human trial of a DNA-based vaccine fortreatment of human immunodeficiency virus type 1 infection: safetyand host response. J Infect Dis 1998;178:92–100.