determination of the luciferin contents in luminous and non-luminous beetles
TRANSCRIPT

Determination of the Luciferin Contents in Luminous
and Non-Luminous Beetles
Yuichi OBA,1;y Takeru SHINTANI,1 Takanori NAKAMURA,1
Makoto OJIKA,1 and Satoshi INOUYE2
1Graduate School of Bioagricultural Sciences, Nagoya University, Nagoya 464-8601, Japan2Yokohama Research Center, Chisso Co., 5-1 Okawa, Kanazawa, Yokohama 236-8605, Japan
Received January 9, 2008; Accepted January 12, 2008; Online Publication, May 7, 2008
[doi:10.1271/bbb.80017]
The contents of firefly luciferin in luminous and non-
luminous beetles were determined by the methods of
HPLC with fluorescence detection and the luminescence
reaction of luciferin and firefly luciferase. Luminous
cantharoids and elaterids contained various amounts
of luciferin in the range of pmol to hundreds of nmol,
but no luciferin was detected in the non-luminous
cantharoids and elaterids.
Key words: bioluminescence; Cantharidae; Elateridae;
Lampyridae; Lycidae
A remarkable feature of beetles (Coleoptera) isbioluminescence that is found in many cantharoidbeetles and some click beetles (Elateridae). The cantha-roid beetles, formerly the superfamily Cantharoidea,1)
consist of Lampyridae (firefly), Phengodidae (railroadworm), Lycidae (net-winged beetle), Cantharidae (sol-dier beetle), and six other small families.2) All knownspecies of Lampyridae and Phengodidae are luminous,3)
but the members of the other cantharoid families arenon-luminous.1) The family Elateridae consists ofluminous and non-luminous species: all luminouselaterids belong to the subfamily of Pyrophorinae.4)
The fireflies, railroad worms, and luminous clickbeetles use the same luciferin (firefly luciferin) in theirluciferin-luciferase reaction (L-L reaction).5) The luci-ferase genes were cloned from various species ofLampyridae, Phengodidae and Elateridae,6) and theprimary sequences show high similarities (>48%) amongthem.7) Thus, the bioluminescence systems of luminousbeetles are essentially the same (Eqs. 1 and 2).5,7)
Luciferinþ ATP � Luciferyl-AMPþ PPi ð1Þ
Luciferyl-AMPþ O2
! Oxyluciferinþ CO2 þ Light ð2Þ
The amount of luciferin has been measured in severalspecies of Lampyridae by the different methods.
Approximately 9.0mg of luciferin was crystallized from15,000 adult lanterns of the North American firefly,Photinus pyralis (2.1 nmol/lantern).8) Thereafter, ap-proximately 5.5mg of luciferin was crystallized from12,000 adult abdomens of the Japanese firefly, Luciolacruciata (1.6 nmol/abdomen).9) Strause et al.10) deter-mined the amount of luciferin in Photuris pennsylvanicaby the L-L reaction and reported that a specimen of larvaand adult contains 90 nmol and 100 nmol of luciferin,respectively. The amount of luciferin in a larva of theIranian firefly, Lampyris turkestanicus, was measured byHPLC with a UV detector,11) and was approximately0.5 nmol/mg of dried lantern. The amount of luciferinin L. lateralis was recently determined by HPLC witha fluorescence detector, and the result showed that alarva and an adult have approximately 0.1 and 1 nmolof luciferin, respectively.12)
The aim of this study is to determine the amountof luciferin in luminous and non-luminous beetles. Weapplied two different methods to analyze the luciferincontent in a specimen: the L-L reaction and an HPLCanalysis with fluorescence detection. The analyticalmethods have an advantage and a disadvantage indetermining the luciferin contents in beetles. Themethod using the L-L reaction is highly sensitive, butthe amount of luciferin might be underestimated due tothe presence of inhibitory factors for the L-L reaction,such as long-chain fatty acids.13) On the other hand, themethod using HPLC with a fluorescence detector is alsohighly sensitive, but the result might be overestimatedwhen the signal of luciferin overlaps with the peak ofother components. Thus, it would be preferable to applytwo independent methods for accurately estimating theamount of luciferin.The mealworm beetle, Tenebrio molitor, was pur-
chased from Jupiter Co. (Tochigi, Japan). The larvae ofL. cruciata and L. lateralis were purchased from ChikyuCo. (Hyogo, Japan). A frozen adult of Pyrophorus sp.collected at Santa-Fe in Panama was kindly supplied by
y To whom correspondence should be addressed. Tel/Fax: +81-52-789-4280; E-mail: [email protected]
Abbreviation: L-L reaction, luciferin-luciferase reaction
Biosci. Biotechnol. Biochem., 72 (5), 1384–1387, 2008
Note

Tama Zoological Gardens (Tokyo, Japan). All the otherbeetles were collected in Aichi, Gifu, and Nagano,Japan. The specimens were frozen by liquid nitrogen,lyophilized with an FDU-506 freeze dryer (TokyoRikakikai, Tokyo, Japan), and then homogenized withten times the weight per volume of hot water at 95 �C.After centrifugation, the supernatant was washed withan equal volume of n-hexane, and the aqueous layer wassubjected to centrifugal filtration with Ultrafree-MC(PVDF, 0.45 mm; Millipore, Bedford, MA, USA). Toestimate the efficiency of luciferin recovery, the adult ofT. molitor or A. binodulus was extracted in the presenceof 100 pmol of authentic firefly luciferin (potassium salt;Wako Pure Chemicals, Osaka, Japan). The recovery ofluciferin was calculated to be 80–90%. For the L-Lreaction assay, the reaction mixture (50 ml) in a 96-wellplate contained the insect extract (38 ml, but 16 ml forthe small species of Plateros coracinus), 5mM ATP,0.25mM CoA and 5mM MgCl2 in 500mM Tris–HCl(pH 7.8). The L-L reaction was initiated by adding 50 mlof recombinant P. pyralis luciferase (1.49 mg in 500mM
Tris–HCl; Promega, Madison, WI, USA). After tenseconds, the light intensity was measured for 60 s byusing a JNR AB-2100 luminometer (Atto, Tokyo,Japan). After the measurement, the pH value of thereaction solution was confirmed to be unchanged. Forthe HPLC analyses, an extract (22–33 ml) was subjectedto reversed-phase HPLC (Jasco, Tokyo, Japan), using aDevelosil ODS UG-5 column (4:6� 250mm; NomuraChemical, Seto, Japan) eluted with a linear gradientof 10–70% MeOH in 80mM Tris–HCl (pH 9.0; thefluorescence intensity of firefly luciferin under the basiccondition is higher than that under the acidic condition)5)
for 30min at a flow rate of 0.6ml/min. Fluorescencewas detected (excitation at 383 nm; emission at 528 nm)with an FP-1520 fluorescence detector (Jasco). Theretention time and peak area were analysed by Bowinsoftware (Jasco), using authentic firefly luciferin. Underthe foregoing conditions for HPLC analysis, luciferinwas eluted at 21min. When a peak was detected at asimilar retention time to that of luciferin, authenticluciferin was mixed with the extract and reanalyzed todistinguish true luciferin from false.
The calibration curve and detection limit of luciferinin the L-L reaction (Fig. 1A) and HPLC (n ¼ 5,Fig. 1B) analyses were determined by using authenticfirefly luciferin. The concentration of authentic fireflyluciferin was determined spectrophotometrically, usingan absorption coefficient of 18,450 M�1 cm�1 at 328 nmin water.14) Quantitative linearity (correlation efficientR2 > 0:999) was observed from 10 fmol to 20 pmol perwell by the L-L reaction (Fig. 1A) and from 15 fmol to20 pmol/injection by HPLC (Fig. 1B). The detectionlimit of luciferin (the mean value� SE is significantlydifferent (Student’s t-test, P < 0:05) from that withoutluciferin) was 5 fmol/well by the L-L reaction. Thedetection limit by the HPLC (signal to noise ratio, S/N>3) was 15 fmol/injection.
The results showed that adult fireflies (Lampyridae) ofL. cruciata, L. lateralis, and L. parvula contain approx-imately 3.5, 0.5, and 1.5 nmol of luciferin per specimen,respectively (Table 1). These amounts of luciferin areapproximately equal to the values previously reported:L. lateralis by HPLC with a fluorescence detector;12)
P. pyralis and L. cruciata by purification.8,9) Theamount of luciferin in an adult of L. parvula isapproximately three times higher than that in L. later-alis, although they have roughly same body size. Theamount of luciferin in larval L. cruciata determined bythe L-L reaction was higher than that by HPLC(Student’s t-test, P < 0:05). The luciferin in larvalL. lateralis was approximately 20 pmol/insect, whichis nearly the same as the result by Niwa et al.12)
L-Luciferin is not a substrate for the bioluminescencereaction,15) and Niwa et al.12) suggested that the larvaof L. lateralis possesses approximately 1:1 mixture ofD- and L-luciferin by an HPLC analysis using a chiralcolumn. In our analysis, the content of luciferin in thelarva of L. lateralis measured by HPLC analysis waslarger than that by the L-L reaction analysis (althoughstatistically not significant; Table 1). This may beexplained by the presence of L-luciferin as suggestedby Niwa et al.12) or by the presence of compounds thatinhibit the L-L reaction in the extract. The adult ofLucidina biplagiata is a diurnal and seems to commu-nicate between male and female by pheromones.16) Theadult of L. biplagiata is usually considered to be non-
Rel
ativ
e lig
ht u
nits
106
108
104
102
A
R2=0.9994
Luciferin (fmol)10-1 101 103 1050
//
B
102
104
106R2=0.9998
Pea
k ar
ea0 -1 101 103 105
Luciferin (fmol)
//10
Fig. 1. Calibration Curves of Luciferin Determined by the Luciferin-
Luciferase Reaction (A) and by HPLC Analysis with a Fluorescence
Detector (B).
Luciferin Contents in Luminous and Non-Luminous Beetles 1385
luminous or almost non-luminous: the bioluminescenceis undetectable by the human eye, although very weakluminescence was detected by a photomultiplier.16) Inthis study, the amount of luciferin in an adult ofL. biplagiata was determined to be approximately3 pmol/insect (Table 1). Our result suggests that thevery weak luminescence of L. biplagiata is caused bythe L-L reaction, as same as the other luminous fireflies.
The beetles in Cantharidae and Lycidae are diurnaland non-luminous,1,3) and the families of Cantharidaeand Lycidae are supposed to be closely related toLampyridae.3,17) Our results indicated that the species ofCantharidae have no luciferin (Table 1). In Lycidae(Lycostomus modestus and P. coracinus), luciferin couldnot be detected by the L-L reaction, but a small peakcorresponding to luciferin on an HPLC chromatogramwas detected in the extract. If this peak was luciferin, thecontent of luciferin was approximately 50 fmol/insectwhich can be readily detected by the L-L reaction. Thus,the peak detected in the extract of lycids by the HPLCanalysis would not be luciferin.
Click beetles (Elateridae) are widespread throughoutthe world, but the luminous species are only found intropical and subtropical America and Melanesia.4) Theluciferin in luminous click beetles was determined to beidentical to firefly luciferin,5) but the amount of luciferinhas not been estimated. In the present study, the amountof luciferin in an adult of the luminous elaterid,Pyrophorus sp., was determined to be approximately250 nmol, which is the highest value in our present
analysis (Table 1). It is known that the luminescenceintensity of luminous elaterids, especially of the genusPyrophorus, is much brighter than that in lampyrids.18)
No luciferin in four non-luminous elaterid species wasdetected, suggesting that non-luminous elaterids haveno luciferin (Table 1). Among the four species, A. bi-nodulus and Cryptalaus berus belong to the subfamilyPyrophorinae, the same as luminous Pyrophorus.19) Wehave recently cloned and characterized the orthologousgene of the beetle luciferase of A. binodulus (AbLL).20)
The gene product of AbLL is fatty acyl-CoA synthetase,and not luciferase. Thus, the non-luminous click beetleA. binodulus lacks both luciferin and the luminescenceactivity of the beetle luciferase-like gene, in spite of itsclose relationship to luminous click beetles.The mealworm beetle T. molitor (Tenebrionidae) is
an outgroup taxa of cantharoids and elaterids.17,21) Ourresults showed that luciferin was not detected in larvaand adult of T. molitor (Table 1). We also found that thehomologous genes of beetle luciferase in T. molitorhave fatty acyl-CoA synthetic activity but not luciferaseactivity with luciferin.21) Thus, T. molitor lacks both aluciferin and a luciferase.On the basis of the morphological analysis, Crowson1)
suggested that the luminosity of Pyrophorus (clickbeetle) and of various cantharoids (firefly and railroadworm) derived from a common ancestor is unlikely. Ourmolecular phylogenetic analysis also suggested that theancestral state of the Elateridae was non-luminous.19) Itmay be hypothesized that the larvae of luminous click
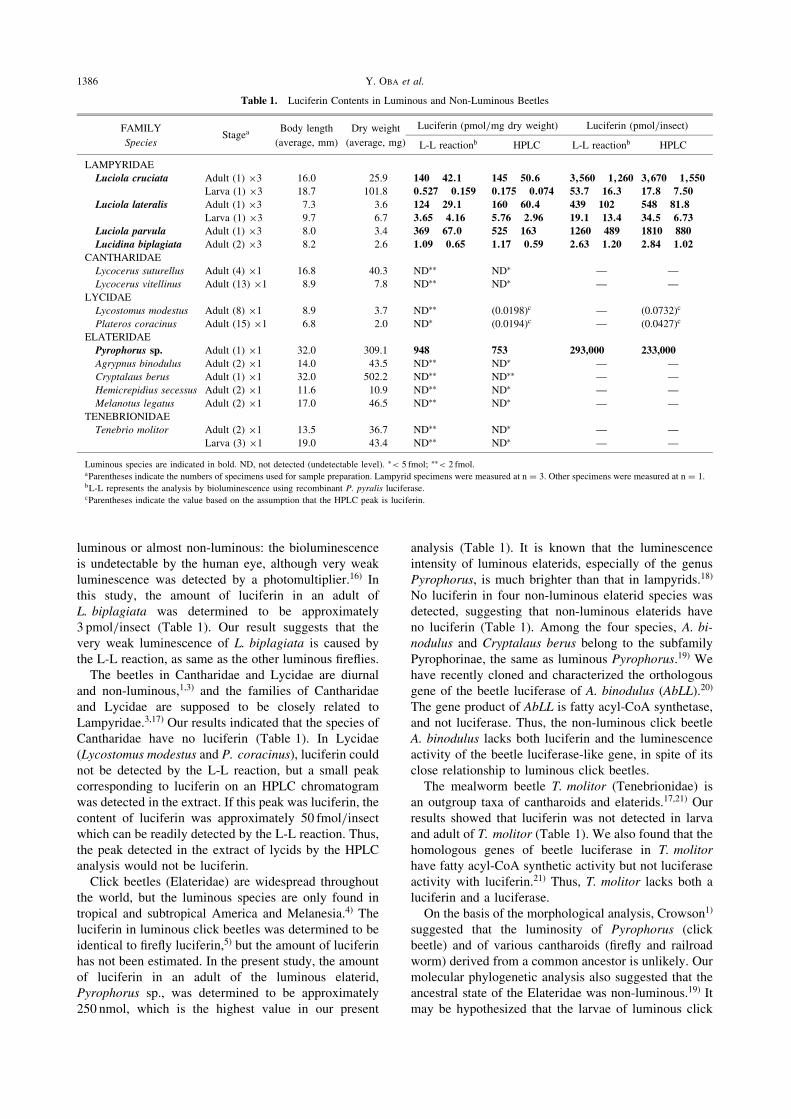
Table 1. Luciferin Contents in Luminous and Non-Luminous Beetles
FAMILYStagea
Body length Dry weight Luciferin (pmol/mg dry weight) Luciferin (pmol/insect)
Species (average, mm) (average, mg) L-L reactionb HPLC L-L reactionb HPLC
LAMPYRIDAE
Luciola cruciata Adult (1) �3 16.0 25.9 140� 42:1 145� 50:6 3;560� 1;260 3;670� 1;550
Larva (1) �3 18.7 101.8 0:527� 0:159 0:175� 0:074 53:7� 16:3 17:8� 7:50
Luciola lateralis Adult (1) �3 7.3 3.6 124� 29:1 160� 60:4 439� 102 548� 81:8
Larva (1) �3 9.7 6.7 3:65� 4:16 5:76� 2:96 19:1� 13:4 34:5� 6:73
Luciola parvula Adult (1) �3 8.0 3.4 369� 67:0 525� 163 1260� 489 1810� 880
Lucidina biplagiata Adult (2) �3 8.2 2.6 1:09� 0:65 1:17� 0:59 2:63� 1:20 2:84� 1:02
CANTHARIDAE
Lycocerus suturellus Adult (4) �1 16.8 40.3 ND�� ND� — —
Lycocerus vitellinus Adult (13) �1 8.9 7.8 ND�� ND� — —
LYCIDAE
Lycostomus modestus Adult (8) �1 8.9 3.7 ND�� (0.0198)c — (0.0732)c
Plateros coracinus Adult (15) �1 6.8 2.0 ND� (0.0194)c — (0.0427)c
ELATERIDAE
Pyrophorus sp. Adult (1) �1 32.0 309.1 948 753 293,000 233,000
Agrypnus binodulus Adult (2) �1 14.0 43.5 ND�� ND� — —
Cryptalaus berus Adult (1) �1 32.0 502.2 ND�� ND�� — —
Hemicrepidius secessus Adult (2) �1 11.6 10.9 ND�� ND� — —
Melanotus legatus Adult (2) �1 17.0 46.5 ND�� ND� — —
TENEBRIONIDAE
Tenebrio molitor Adult (2) �1 13.5 36.7 ND�� ND� — —
Larva (3) �1 19.0 43.4 ND�� ND� — —
Luminous species are indicated in bold. ND, not detected (undetectable level). �< 5 fmol; ��< 2 fmol.aParentheses indicate the numbers of specimens used for sample preparation. Lampyrid specimens were measured at n ¼ 3. Other specimens were measured at n ¼ 1.bL-L represents the analysis by bioluminescence using recombinant P. pyralis luciferase.cParentheses indicate the value based on the assumption that the HPLC peak is luciferin.
1386 Y. OBA et al.

beetles, which are carnivorous in the larval stage, alsoobtain luciferin from luminous cantharoids such asLampyridae and Phengodidae sympatrically occurring inCentral and South America. However, this possibility isinvalid, because adults of Pyrophorus were luminouseven when the specimens were reared from egg to adultby feeding only T. molitor as bait (K. Ogura, personalcommunication). These results suggested that the lumi-nous click beetle and luminous cantharoids independ-ently developed the ability of luciferin biosynthesis.
Acknowledgments
The authors thank Kanjirou Ogura (Tama ZoologicalGardens, Tokyo) for information about the laboratoryrearing of luminous click beetles, Dr. Hitoo Ohira forthe specimen identification, and Tooru Ojika forsupplying specimens.
References
1) Crowson, R. A., A review of the classification ofCantharoidea (Coleoptera), with the definition of twonew families, Cneoglossidae and Omethidae. Rev. Univ.Madrid, 21, 35–77 (1972).
2) Lawrence, J. F., Coleoptera. In ‘‘Synopsis and Classi-fication of Living Organisms,’’ ed. Parker, S. P.,McGraw-Hill, New York, pp. 482–553 (1982).
3) Branham, M. A., and Wenzel, J. W., The origin of photicbehavior and the evolution of sexual communication infireflies (Coleoptera: Lampyridae). Cladistics, 19, 1–22(2003).
4) Stibick, J. N. L., Classification of the Elateridae(Coleoptera): relationships and classification of thesubfamilies and tribes. Pacific Insects, 20, 145–186(1979).
5) Shimomura, O., ‘‘Bioluminescence: Chemical Principlesand Methods,’’ World Scientific, Singapore, pp. 1–29(2006).
6) Viviani, V. R., The origin, diversity, and structurefunction relationships of insect luciferases. Cell. Mol.Life Sci., 59, 1833–1850 (2002).
7) Wood, K. V., The chemical mechanism and evolutionarydevelopment of beetle bioluminescence. Photochem.Photobiol., 62, 662–673 (1995).
8) Bitler, B., and McElroy, W. D., The preparation andproperties of crystalline firefly luciferin. Arch. Biochem.Biophys., 72, 358–368 (1957).
9) Kishi, Y., Matsuura, S., Inoue, S., Shimomura, O., andGoto, T., Luciferin and luciopterin isolated from the
Japanese firefly, Luciola cruciata. Tetrahedron Lett., 9,2847–2850 (1968).
10) Strause, L. G., DeLuca, M., and Case, J. F., Biochemicaland morphological changes accompanying light organdevelopment in the firefly, Photuris pennsylvanica. J.Insect Physiol., 25, 339–347 (1979).
11) Hadj-Mohammadi, M. R., and Chaichi, M. J., Separa-tion, identification and determination of luciferin in theIranian firefly, Lampyris turkestanicus by HPLC andspectroscopic methods. Photochem. Photobiol., 64,821–822 (1996).
12) Niwa, K., Nakamura, M., and Ohmiya, Y., Stereo-isomeric bio-inversion key to biosynthesis of fireflyD-luciferin. FEBS Lett., 580, 5283–5287 (2006).
13) Ueda, I., and Suzuki, A., Is there a specific receptorfor anesthetics? Contrary effects of alcohols and fattyacids on phase transition and bioluminescence of fireflyluciferase. Biophys. J., 75, 1052–1057 (1998).
14) Morton, R. A., Hopkins, T. A., and Seliger, H. H., Thespectroscopic properties of firefly luciferin and relatedcompounds. An approach to product emission. Biochem-istry, 8, 1598–1607 (1969).
15) Seliger, H. H., McElroy, W. D., White, E. H., and Field,G. F., Stereospecificity and firefly bioluminescence, acomparison of natural and synthetic luciferins. Proc.Natl. Acad. Sci. USA, 47, 1129–1134 (1961).
16) Ohba, N., Studies on the communication system ofJapanese fireflies. Sci. Rept. Yokosuka City Mus., 30,1-62 + 6 pls (1983).
17) Hunt, T., Bergsten, J., Levkanicova, Z., Papadopoulou,A., St. John, O., Wild, R., Hammond, P. M., Ahrens, D.,Balke, M., Caterino, M. S., Gomez-Zurita, J., Ribera, I.,Barraclough, T. G., Bocakova, M., Bocak, L., andVogler, A. P., A comprehensive phylogeny of beetlesreveals the evolutionary origins of a superradiation.Science, 318, 1913–1916 (2007).
18) Levy, H. C., ‘‘University of Florida Book of InsectRecords’’ Chapter 29: Greatest bioluminescence, ed.Walker, J., Univ. Florida, pp. 72–73 (1998).
19) Sagegami-Oba, R., Oba, Y., and Ohira, H., Phylogeneticrelationships of click beetles (Coleoptera: Elateridae)inferred from 28S ribosomal DNA: insight into theevolution of bioluminescence in Elateridae. Mol. Phylo-genet. Evol., 42, 410–421 (2007).
20) Oba, Y., Iida, K., Ojika, M., and Inouye, S., Orthologousgene of beetle luciferase in non-luminous click beetle,Agrypnus binodulus (Elateridae), encodes a fatty acyl-CoA synthetase. Gene, 407, 169–175 (2008).
21) Oba, Y., Sato, M., and Inouye, S., Cloning andcharacterization of the homologous genes of fireflyluciferase in the mealworm beetle, Tenebrio molitor.Insect Mol. Biol., 15, 293–299 (2006).
Luciferin Contents in Luminous and Non-Luminous Beetles 1387