developmental expression of the lipocalin lazarillo and its role in
TRANSCRIPT

INTRO
Amonbiologdiversihow tneuronneurogat the systeminsectscalled neurobposition. After their differentiation, NBs undergo severaloriented asymmetric divisions giving rise to smaller ganglionmother cells (GMCs) that will divide symmetrically to generatetwo postmitotic neurons. The stereotypy and invariancedisplayed by the progeny (Doe et al., 1985; Huff et al., 1989)suggest that GMC and neuron identity is mainly determined byNB identity and by lineage relationships. ‘NB-identity’molecules are expressed in unique subsets of NBs; while ‘sub-lineage-identity’ molecules are expressed in NBs and theirprogeny only during a certain fraction of the lineage andgenerate differences between sister neurons (Cui and Doe,1992; Doe, 1992). Finding new molecules with this cellulardistribution is of special interest to uncover the mechanismsgenerating cellular diversity in the CNS.
Once postmitotic neurons start axonogenesis then epigeneticinteractions with the surroundings are essential to direct theirgrowth cones (Doe et al., 1985). In this context, moleculeshave been described that are expressed on the surface of
Then of cor asawaer 88;f atric axgit
ty mothe
which predicts that axon fascicles differentially labeled bysurface molecules are used for growth cone guidance. This rolehas been assayed in invertebrates for several surface proteinsby antibody perturbation (Harrelson and Goodman, 1988;Seaver et al., 1991; Kolodkin et al., 1992; Wang et al., 1992)or genetic mutation (Elkins et al., 1990; Hedgecock et al.,1990; Grenningloh et al., 1991; McIntire et al., 1992).Moreover, such a restricted molecular code has also beeninvoked in the developing vertebrate nervous system (Doddand Jessell, 1988; Bixby and Harris, 1991; Kuwada, 1992) sup-porting the idea that common mechanisms of guidance havebeen conserved during evolution.
Lazarillo is a highly glycosylated protein anchored by aglycosyl-phosphatidylinositol (GPI) tail to the plasmamembrane of a subset of neurons in the nervous system of thegrasshopper embryo. The deduced Lazarillo sequence revealsthat it belongs to the lipocalin family, a group of proteins func-tioning as extracellular carriers of small hydrophobic
DevelopPrinted i
This aanalysexpresand a by a neuroneuroof neuin a f
ocloentneer a anc
cali
SUMM
Dev r
path
Diego
Biolog
*M. D. G†Author
y could be part of a molecular the multiple axonal pathways.
uld be functioning as receptors the signal for other axons thaty from them. The fasciclins,and fly (Bastiani et al., 1987; Patel et al., 1987; Kolodkin etxonal-identity molecules thattion; they are expressed region-ons defining certain pathwaysudinal). The existence of these
olecules is a prerequisite forsis’ (Goodman et al., 1982),
135
nal antibody raised against and direction of growth ofr neurons. We propose thatmidline morphogen involvede of these neurons.
n, grasshopper, lipid binding
ole in axonal
ment 121, 135-147 (1995)n Great Britain © The Company of Biologists Limited 1995
ARY
elopmental expression of the lipocalin Lazarillo and its
finding in the grasshopper embryo
Sánchez*,†, María D. Ganfornina* and Michael J. Bastiani
y Department, University of Utah, Salt Lake City, UT 84112, USA
. and D. S. contributed equally to this workfor correspondence
DUCTION
g the more intriguing issues that developmental neuro-ists have faced is the generation of the enormous cellularty in the nervous system and the mechanisms that definehese diverse cells establish the intricate network ofal connections. A general strategy for understandingenesis and axon pathfinding has been to examine, bothcellular and molecular level, the less complex nervouss of invertebrates. The central nervous system (CNS) in develops from a monolayer of ventral ectodermal cellsthe neurogenic region where a set of enlarged cells, thelasts (NBs), differentiate in a stereotyped number and
restricted subsets of axons. code governing the formatio‘Axonal-identity’ moleculesfor a specific guidance cue ofasciculate with, or grow described in both grasshoppHarrelson and Goodman, 19al., 1992), are examples opossess a second level of resally only in portions of the(e.g. commissural versus lonaxonal- and pathway-identithe ‘labeled pathways hyp
rticle describes the expression pattern and functionalis of Lazarillo, a novel cell surface glycoproteinsed in the embryonic grasshopper nervous system,
member of the lipocalin family. Lazarillo is expressedsubset of neuroblasts, ganglion mother cells andns of the central nervous system, by all sensoryns of the peripheral nervous system, and by a subsetrons of the enteric nervous system. It is also presentew non neuronal cells associated mainly with the
excretory system. A monLazarillo perturbs the extidentified commissural pioLazarillo is the receptor foin the outgrowth and guid
Key words: axon guidance, lipoproteins

136
moleculework wetional ansuggests moleculenon-neuragreemenits role antibodygrasshopa subset ocentered of commmechanismay be g
MATER
Grasshopp
a colony mpercentag(1979). T(1990) agnization m
ImmunoThe antiboby Denbuand disseNaCl, 3 mglycine, fipH 7.2 anmM NaCl50 mM Tserum albmAb 10Enatant diluthe primaTBS, blocIgG (Jack2 hours atlast changEmbryos oxidase (3
M TBS amicroscopa secondapreparatiostudy the developmstandard pbodies (3-antibody)solutions.fixed tissuin 10% fo
In situSánchez a
AntibodyThe mAbaffinity pSepharoseprepared
D
s (Ganfornina, Sánchez and Bastiani, 1995). In this report the developmental expression pattern and func-alysis of this new lipocalin. Its unique distributionthat Lazarillo is a sublineage- and axonal-identity in the developing nervous system. Its presence inonal tissues suggests other functions more int with the lipocalins found in invertebrates. To testin axon guidance we have used the monoclonal (mAb) 10E6 to perturb the protein in culturedper embryos. Binding of the mAb to its epitope causesf growth cones to be delayed and misrouted. We haveour study on the altered behavior of an identified pairissural pioneer neurons. The cellular and molecularms by which a GPI-anchored membrane lipocalinuiding these commissural neurons are discussed.
(Pierce). The concentration of antigen-binding sites was estimatedtaking into account absorbance at 280 nm, relative molecular mass,and number of antigen binding sites in each molecule.
Grasshopper embryos were dissected in medium consisting of: 50%Schneider’s Drosophila Medium (Gibco-BRL), 49% MinimumEssential Medium (α medium, Gibco-BRL), 1% antibiotic-antimy-cotic solution (Sigma), 0.5 ng/ml juvenile hormone I and 0.3 ng/mlβ-ecdysterone (Sigma). Each embryo was transferred to a HLA platewell (Nunc) containing 20 µl of prewarmed medium and the experi-mental or control antibodies. Embryos were incubated at 33°C for 24hours. Embryos used for each perturbation experiment were chosenfrom a single clutch and 4 embryos were fixed at the initiation ofculture period (‘t=0’ embryos). After the culture period, embryos werewashed in Ringer’s solution and then fixed and labeled as describedfor immunocytochemistry. Embryos that were cultured with mAb orFab 10E6 were subsequently exposed to a secondary antibody (HRP-conjugated goat anti-mouse IgG, from Jackson Immunoresearch).Control and t=0 embryos were labeled with mAb 10E6 and secondaryantibody as described above. The mAb 10H11 (obtained in the same
. Sánchez, M. D. Ganfornina and M. J. Bastiani
I
eha
c
rc
d
r
6
r
s
r
pe
u
AL AND METHODS
er (Schistocerca americana) embryos were obtained fromaintained at the University of Utah. They were staged by of embryonic development according to Bentley et al.e mAb 10E6 was generated by Carpenter and Bastianiinst embryonic nervous tissue using a subtractive immu-ethod (Hockfield, 1987).
ytochemistry and in situ hybridizationdy labeling protocol is a modified version of the proceduresg et al. (1989). Embryos were removed from the egg caseted in Ringer’s solution (4.6 mM Tes, pH 7.4, 150 mMM KCl, 2 mM CaCl2, 1 mM MgSO4) with 6 mg/ml of
xed in 4% paraformaldehyde in 0.1 M sodium phosphate, washed in PBS (20 mM potassium phosphate, pH 7.2, 150
). Embryos were incubated for 1 hour in blocking solution:is, pH 7.2, 350 mM NaCl (50 mM TBS); 30 mg/ml bovineumin (BSA fraction V, Sigma); 2% normal goat serum. was used as ascites fluid diluted 1:200 or hybridoma super-ted 1:1 in blocking solution. Embryos were incubated withy antibody at 4°C for 8-12 hours, then washed with 50 mMked again and exposed to HRP-conjugated goat anti-mouseon Immunoresearch) diluted 1:500 in blocking solution for
room temperature, washed again with 50 mM TBS with ae to 0.1 M TBS (0.1 M Tris, pH 7.2, 350 mM NaCl).were reacted with diaminobenzidine (1 mg/ml), glucose
screening as mAb 10E6) and anti-HRP antibodies (JacksonImmunoresearch) were used in control experiments because both arenervous system specific and label the AcP cells.
To analyze the effects of the immunoperturbants used we chose theAcP cells of the S2 segment (see Results). A percentage of develop-ment was assigned to each cell according to Fig. 5F. Total growthachieved was estimated by subtracting the value of each control orexperimental cell from the average of t=0 embryos. The mean growthand s.e.m. were calculated for the control and experimental groups ofeach experiment. To graphically represent the results of all experi-ments, they were grouped by type (A or B, see Results), antibody used(none, mAb 10H11, anti-HRP antibodies, mAb or Fab 10E6), andantigen binding site concentration. In each group of experiments thedifference between experimental growth values and the mean valueof control growth (growth cone delay) were calculated and its meanand s.e.m. were represented normalized to the mean growth obtainedby control embryos in culture. The nonparametric test of Mann-Whitney was used to assess statistical significance of differences ingrowth. Altered AcP cells were scored when a major deviation fromthe normal pathway (Fig. 5) was observed. Cases were not scoredwhen only filopodia extended along aberrant pathways. A χ2 test wasused to assess differences in frequency of abnormalities.
RESULTS
A general view of the Lazarillo expression pattern revealed by
mU/ml, type V) and β-D-glucose (2 mg/ml), washed in 0.1nd cleared in glycerol. Differential interference contrasty (DIC) was used to analyze the preparations. Occasionallyy antibody conjugated to FITC or rhodamine was used andns were analyzed by standard fluorescence microscopy. Toattern of expression in grasshoppers ranging from 60% ofnt to 3rd instar adults, nerve cords were dissected and therotocol was followed using longer incubation times for anti-6 days for the primary and 18-24 hours for the secondaryand adding 1% saponin to the blocking and antibodyAdult nerve cord sections were obtained by embeddinge in OCT medium (Miles), sectioning at −18°C and fixingrmaldehyde. Antibody labeling proceeded as above.
hybridization procedures are detailed in Ganfornina,nd Bastiani (1995).
perturbation experiments10E6 used as a perturbing agent on cultured embryos wasrified from hybridoma supernatants using a Protein G
column (Pharmacia). The monovalent Fab fragments wereas described by the Immunopure Fab Preparation Kit
labeling with mAb 10E6 is outlined in Fig. 1. At 14% of devel-opment the embryos consist of an anterior enlarged portion, thepresumptive head, and a smaller posterior primordium. As thisprimordium enlarges, gastrulation and segmentation occursequentially in an anteroposterior direction. At 24% of devel-opment (Fig. 1A) segments are morphologically defined up tothe second abdominal segment. The first pair of neuronslabeled by mAb 10E6 appear in the head at 22% of develop-ment. Their axons will cross the midline by the primary com-missural tract of the brain. Slightly later, a more lateral groupof neurons is also labeled in the head. At 30% of developmentsegmentation is complete and the embryo consists of a head,three subesophageal segments (S1-S3), three thoracicsegments (T1-T3) and eleven abdominal segments (A1-A11).The ventromedial region of the ectodermal layer has beendefined as the neurogenic region throughout the embryo and astereotyped set of neuroblasts (NBs) differentiate in eachsegment. The neurons they produce will set up an orthogonalscaffold of axon tracts consisting of two longitudinal connec-

137Expression pattern and function of Lazarillo
tives ruhemigaanterio10E6 latudinalin somsegmenearly stnot expCNS sclabel walso difof devesegmenCNS wwhich motor athe intelabeleddevelopexpress
Fig. 1. D10E6. Wgroup oprimaryof the manterioranteriorauditoryforegut fascicle(ISN, inat 80% panel).
3-A3 (fused together), and A5 (lower panel) of an embryonicerve cord at 80% of development. Lazarillo expression is
bryo. Lazarillo is visualized by immunocytochemistry with mAb Anterior is up. (A) At 24% of development a medial and a lateraleir surface. (B) At 32% of development, labeling appears on thedian neuroblast (MNB) and its progeny appear labeled at the midlinements are intensely labeled with mAb 10E6. The pioneers of theof development the first commissural fascicles appear in the mosty neuronal clusters start to label in the lateral body wall. AO,ystem is also labelled (arrowhead) on the anterior margin of the
ression pattern of Lazarillo is seen. Commissural and longitudinalgment in C and D), and well developed sensory organs and nervesa of T2-A2 segments (upper panel) and A5 (lower panel) are shownd as well as the peripheral nerves exiting the ganglia (arrow in lowers: A-D 300 µm, E 150 µm.
nning between segments, two commissures linking bothnglia and a dorsal median fiber tract (MFT) that extends
Tn
evelopmental expression pattern of Lazarillo in the grasshopper emhole embryos are viewed from the dorsal surface with DIC optics.
f cells in the developing brain (arrowheads), express Lazarillo on th commissure and longitudinal connectives (L) of the brain. The meore anterior segmental ganglia. Four cells in the subesophageal seg commissure (AcP neurons) are indicated by an arrow. (C) At 38% segments (example shown with long arrow in T2 segment). Sensor organ in A1 segment. The frontal ganglion of the enteric nervous sinvagination. (D) At 45% of development, the complete neural exps are evident in the brain and ventral nerve cord (compare the T2 setersegmental nerve) are present at the lateral body wall. (E) Gangliof development. Commissural and longitudinal pathways are labeleThe MNB is labeled at this age (arrowhead in the T3 ganglion). Bar
rly along the midline. At 32% of development (Fig. 1B)beling appears in the brain commissure, in the longi-
tracts connecting the brain with the segmental ganglia,e brain NBs, in commissural neurons in S1 and S2ts, and in MFT neurons up to the A2 segment. At theseages other neurons that have extended their axons doress Lazarillo. At 38% of development (Fig. 1C) theaffold has acquired more axonal tracts, some of whichith the mAb. Sensory neurons expressing Lazarillo areferentiating in the body wall and appendages. At 45%lopment (Fig. 1D) the axonal scaffold of the brain andtal ganglia is completely outlined. Completion of theill be carried out mostly by addition of new neuronswill follow preformed axonal fascicles. Sensory andxons travel along either the segmental nerve (SN) orrsegmental nerve (ISN). These peripheral nerves are by mAb 10E6. The enteric nervous system (ENS) thats on the surface of the foregut invagination alsoes Lazarillo (Fig. 1C,D). Fig. 1E shows the ganglia T2,
maintained in the neuropilar region as well as in the longitu-dinal connectives and peripheral nerves up to hatching. Thelabeling in adult nerve cords appears restricted to certaingroups of neurons and a ventral medial tract in the longitudi-nal connectives (Ganfornina, Sánchez and Bastiani, 1995, Fig.1).
Identification of Lazarillo-expressing neurons in thegrasshopper CNSLazarillo is localized on the membrane surface of the cell body,axon, growth cone and filopodia of a subset (5-10%) of theCNS neurons. At 32% of development (Fig. 2A) only a pair ofcells that pioneer a fascicle in the anterior commissure and theMP4 and MP6 cells whose axons form the MFT are labeled.The median neuroblast (MNB) is also labeled. However, othercells that participate in forming the axonal scaffold do notexpress Lazarillo (open arrows). At 37% more neurons areadded to the labeling pattern (Fig. 2B), such as a pair of lon-gitudinal axons, a fascicle in the posterior commissure, and

138
neuronsFig. 2CSome of1984), athe Q5
D D. Ganfornina and M. J. Bastiani
Fig. 2. The expression ofLazarillo in the CNS isrestricted to a specific subset ofneurons, NBs and GMCs in aprecise temporal window.(A) The labeling at 32% ofdevelopment in the metathoracicganglion is seen on the neuronsthat pioneer the anteriorcommissure (AcP cells) and theMFT (MP4 and MP6 cells), aswell as on the MNB. Refer to Cfor identity of neurons. Openarrows indicate unlabeled axons.(B) Later on (37%), moreneurons express Lazarillo asthey differentiate: for examplethe Q5 and Q6 cells (arrowhead)that cross the midline by the
of labelinshown asevidenceof a metamidline. ventral suappears s1%. At eII: NB 7-
. Sánchez, M.
sa
posterior commissure, or theVm neurons that extends alonga medial longitudinal fascicle(arrow). (C) Map of the 10E6-labeled neurons and NBs at 45%of embryonic development. Thepattern of NBs labeled by mAb10E6 is shown on the righthemisegment with dashed lines.Newly identified neurons are:anterior commissure pioneers(AcP), anterior segmentalmotoneurons (AsM), lateralmotoneurons (lM), ventromedialtract neurons (Vm) that followthe vMP2 fascicle, the Zneurons that contribute to theposterior bundle of the anteriorcommissure, and the lL (laterallongitudinal neurons) that runalong the A/P fascicle.Previously identified neurons (initalics) are: the corner cells(aCC and pCC), Q5 and Q6,MP4, MP6 and DUM (dorsalunpaired motoneurons). NBsand neurons noted with asterisks
d
T
a4
that exit the CNS along the anterior edge of the SN.shows a map of the neurons labeled by mAb 10E6. them have been identified previously (Goodman et al., the MP4 and MP6, which contribute to the MFT, ornd Q6 neurons, which emerge from the NB 7-4 and
send their axons by the posterior commissure (see Fig. 2B,arrowhead). Examples of newly identified cells are the twolateral neurons we have named Z cells, which cross the midlineby a posterior fascicle of the anterior commissure, and the Vmcells, which send their growth cones slightly dorsally and then
have segment-specificcharacteristics. NB 1-1 islabeled only in the S2 segmentand the aCC and pCC cells onlyin the S1 segment. The intensity
g of the AcP cells is much higher in S2 than in other segments. (D) A single focal plane of an A2 segment at 34% of development is an example of NBs expressing Lazarillo. (E,F) The NBs themselves and not the surrounding sheath cells express Lazarillo as by the correlation between in situ hybridization with a Lazarillo RNA probe (E) and mAb labeling (F) of the MNB. (G) Caudal view
thoracic ganglion at 37% of development (dorsal is up). Families of cells derived from NBs 6-1 and 6-2 are seen on both sides of thehe arrowhead points to a NB 6-1 GMC. The expression of Lazarillo in NB 6-1 has started to fade. No neuroepithelial cells at the
rface of the ganglion show labeling. The MNB family is seen darkly labeled at the dorsal midline. (H) The mAb 10E6 labelingequentially in NBs. The diagrams represent a thoracic ganglion from 26% (I) to 30% (IV) of development at approximate intervals ofch stage, newly appearing NBs are depicted in black, while previous ones are in gray. The order of appearance is as follows. I: MNB;; III: NBs 2-1, 2-5, 5-6, 6-1, 6-4, and 7-1; IV: NBs 4-3, 4-4, 5-4, and 6-2. Scale bars: A-D and G, 50 µm; E and F, 20 µm.

anteriorlnewly idfirst fasclateral mof the Studinal caxons naThe sibsegmentits anter1v descneurons and vMP6 cells alabeled
LazarillsubsetmAb 10of NBs.1, 7-4 aclosely HoweveexpresscytoplasneuroblNB-idenobserve
The Nticular oically indevelopto start T3). Ththe ordeplace onlium. Nstarts fathe MN(Fig. 1Eof mostthey ceaexpresssibility tdynamicactively thereforeidentity’
In thecorrespoenitor Nand NB ticular cthe progNBs is aa detaile
LazarillnervouIn the Pcells, of
139Expression pattern and function of Lazarillo
y (Fig. 2B, arrow) following the vMP2 fascicle. Otherentified neurons are the AcP neurons, pioneers of theicle of the anterior commissure (Fig. 2A). A group ofotoneurons (lM) exit the CNS by the anterior border
N. Finally the lL cells are located lateral to the longi-onnectives by the anterior branch of the ISN and theirvigate posteriorly fasciculating with the A/P fascicle.
lings of the AcP neurons, which we call anterioral motoneurons (AsM) pioneer the SN (running alongior border) and could correspond to the motoneuronsribed by Whitington (1989). There are many knownthat do not label with mAb 10E6 such as the Q1, Q22 cells, that pioneer the fascicles followed by the Q5-nd the Vm cells respectively. The aCC and pCC are
only in the S1 segment.
o is a NB-identity molecule expressed by a of NBs and GMCs in the CNS
restricted than in the CNS; all described sensory neurons(Bentley and Keshishian, 1982; Meier et al., 1991) expressLazarillo. The labeled sensory groups that travel to the CNSthrough the ISN are shown in Fig. 3A. Sensory neurons of theSN are also labeled (not shown). The only unlabeled axonfascicle is the anterior branch of the ISN, or U fascicle,composed at this age of motoneuron axons exiting the CNS.The group termed lGp (lateral guidepost) cells is uniquelylabeled by mAb 10E6. Meier et al. (1991) studied the grasshop-per PNS in the body wall using anti-HRP antibodies as ageneral neuronal marker and they reported this particular groupof cells (they called it lSO2) as being weakly or rarely labeledand only in T1-A1. A double labeling of segment A2 with mAb10E6 detected with a rhodamine-conjugated secondaryantibody and anti-HRP antibodies conjugated to FITC showsthat anti-HRP antibodies do not label the lGp cells while mAb10E6 does (Fig. 3B,C). The labeling of lGp cells is very intensein comparison to other sensory groups, and is present from T1
E6 reveals the presence of Lazarillo on a particular set Several examples are shown in Fig. 2D (NBs 2-1, 6-nd 7-1) and the whole map is shown in Fig. 2C. GMCsassociated with labeled NBs are also labeled (Fig. 2G).r, the neuroepithelial cells surrounding the NBs never
Lazarillo. The Lazarillo mRNA is present in them (Fig. 2E) and the protein is on the surface of the
ast (Fig. 2F). Lazarillo can therefore be considered atity molecule. Segment-specific differences are also
d, such as NB 1-1 only labeled in the S2 segment.B expression of Lazarillo is not synchronous. The par-rder of appearance of the labeling is depicted schemat- Fig. 2H. The MNB is the first labeled NB (at 26%ment in the metathoracic segment, T3) and the last oneexpressing Lazarillo is NB 6-2 (at 30% development ine onset of Lazarillo expression is not correlated withr of NB birth described by Doe et al. (1985) and takesce all NBs have differentiated from the neuroepithe-
B expression of Lazarillo is also transitory. Labelingding sequentially, starting with NB 7-4 and ending withB, whose labeling is observed until 80% developmnet; arrowhead). Because of the weak immunoreactivity
NBs after 40% stage, it is difficult to establish whense the expression of Lazarillo. Moreover, some of them
Lazarillo intermittently, so we cannot rule out the pos-
to A10 segments. lGp cells send their neurites toward thelateral dorsal body wall (dBw) cells before these begin axono-genesis (Fig. 3D). Later on, the dBw axons contact the lGpaxons, fasciculate with them and navigate to the CNS. The lGpcells appear labeled at least until 60% of embryonic develop-ment, but we do not know the fate of this putative sensoryorgan.
The appendages of grasshopper embryos have been a modelfor studies on guidance of pioneer axons (reviewed by Bentleyand O’Connor, 1992). The first sensory cells that send theiraxons to the CNS, the pioneer Ti1 neurons, follow a highlystereotyped pathway in which they encounter neuronalguidepost cells (Fe1, Tr1, and Cx1). Lazarillo is expressed bythe sensory neurons of the developing legs. Fig. 3E shows ametathoracic leg at 32% of embryogenesis. Both the Ti1 andthe guidepost cells are 10E6 positive, but with a darker labelingin the latter. All subsequent differentiating groups of sensoryneurons are also labeled (see Figs 3, 6 in Ganfornina, Sánchezand Bastiani, 1995). Every sensory neuron is labeled in theantennae (Fig. 3F), labrum, mouth appendages and cerci.
Unlike in the CNS, Lazarillo is absent in the sensory organmother cells (SOCs) in the PNS. Neurogenesis of sensory cellstakes place in the periphery, close to the final location of thesensory organ. A SOC differentiates within the epithelium anddivides symmetrically giving rise to a pair of neurons that
hat NB labeling reappears later in development. These changes in expression are taking place while NBs aredividing to produce a portion of their progeny and Lazarillo can also be considered a ‘sublineage- molecule (Cui and Doe, 1992). cases where the lineage has been traced there is a clearndence between 10E6-positive neurons and the prog-B also being labeled. Examples are the Q5 and Q6 cells7-4, or the DUM neurons and the MNB, and the par-ase of aCC and pCC cells in the S1 segment that areeny of NB 1-1 of segment S2. A subset of cerebrallso labeled by mAb 10E6, but we have not attemptedd analysis of them.
o is present in the peripheral and enterics systemNS only neurons, and not supportive non-neuronalsensory organs label with the mAb. The pattern is less
delaminate into the lumen and start extending their axons alongthe inner surface of the epithelium. Fig. 3F shows two daughterneurons in an antenna before delamination from the epi-thelium.
A subpopulation of cells expressing Lazarillo are alsoobserved in the outer layer of the foregut invagination wherethe ENS is developing (Fig. 1C,D). These cells will eventuallyform part of the frontal ganglion (not shown).
Expression of Lazarillo outside the nervous systemLazarillo is not strictly specific of the nervous system. In theMalpighian tubules, a part of the insect excretory system,Lazarillo is present in the six cellular groups that initially dif-ferentiate from the proctodaeum epithelium at 45% of devel-opment (Fig. 4A). While the tubes are elongating theexpression is restricted to a few cells at the tips (Fig. 4B). By70% a new round of Malpighian tubule differentiation takesplace; again Lazarillo appears at the tip of newly growing

140
tubules (related tsystem (revieweolder emdistendethat Malactive trthe haemtubules.
Othertributed wall (Fiorigin ofgroup ofof the in4C,D). Twith it presencecells (Fproducetaking iThree mdescribecharactepositive nephrocFuture ecells and
The labels wis an arcof loosarising fthe S1 involvedhemolym1985).
The mAoutgrowAfter a ctifiable we chossegmentmAb oncells ptoward axons lathe arealabelingfilopodiapreciselyexamineplace duwas chois imporlabel muthan in immunohybridiz
D
not shown). It is intriguing that other genes have beeno selection of progenitor cells in both the nervousand the developing tip cells of Malpighian tubulesd by Campos-Ortega, 1994). At later stages (65% andbryos) Lazarillo expression always correlates with a
d shape of the tubule tips. Chapman (1971) reportedpighian tubule tips appear swollen during periods ofansport of constituents fromolymph to the lumen of the
10E6-labeled cells are dis-along the sides of the bodyg. 4E). We could trace the these polymorphic cells to a mesodermal cells at the tipvaginating proctodaeum (Fig.hey eventually loose contact
The axons of the AcP cells follow the stereotyped pathwayshown in Fig. 5. At 29%, the AcP cell bodies are locatedslightly ventral to the dorsal basal lamina and extend randomlydirected filopodia (Fig. 5A). The AcP growth cones extendtoward the embryonic midline at 32% of development (Fig.5B). They encounter the contralateral AcP growth cone andboth axons fasciculate together at 34% of embryogenesis (Fig.
. Sánchez, M. D. Ganfornina and M. J. Bastiani
and migrate anteriorly. The of Lazarillo mRNA in theseig. 4C) suggest that they Lazarillo and are not justt up from the hemolymph.ain cellular types have beend in insects that could fit theristics of these Lazarillo-
cells: sessile haemocytes,ytes, or the fat body cells.xperiments will identify these define their function.subesophageal body also
ith mAb 10E6 (Fig. 4F). Thished structure composed largeely arranged nephrocytesrom the lateral mesoderm ofsegment and functionally
in excretion and synthesis ofph components (Crossley,
b 10E6 perturbs axonth and guidance
lose examination of the iden-cells that express Lazarillo,e the AcP cells in the S2 to analyze the effects of the axonal pathfinding. These
roject their growth conesthe midline when no otherbeled by 10E6 are present in, and show a conspicuous
on the growth cone and that allows us to assess their behavior. We alsod other processes takingring the culture period that
sen to study the AcP cells. Ittant to note that the AcP cellsch more intensely in the S2other segments with both
cytochemistry and in situation.
Fig. 3. Lazarillo is present on all sensory neurons in the PNS of the body wall and appendages.In A-D, CNS is on the right, and anterior is up. (A) Sensory neurons entering the CNS via theintersegmental nerve (ISN) are shown labeled by mAb 10E6. The anterior branch of the ISN, orU fascicle (Uf), is never labeled, while axons travelling by the posterior branch are labeled(arrowhead). dBw, dorsal body wall group; ‘SR’, wing hinge stretch receptor homologue inabdominal segments; lGp, lateral guidepost cells; AO, auditory organ. (B,C) A new neuronalgroup in the ISN (lGp cells) has been identified with mAb 10E6 to be present in all segmentsfrom T1 to A10. Anti-HRP antibodies, commonly used as a general neuronal marker, do notrecognize these cells. A double labeling of the A2 segment is shown with mAb 10E6 evidencedwith rhodamine-conjugated secondary antibodies (B) and anti-HRP antibodies directly taggedwith FITC (C). (D) lGp cells send their neurites towards the more distal dBw cells before thelatter start axonogenesis. The dBw axons follow the former as guideposts on their way to theCNS. (E) At the 32% stage the pioneer sensory neurons of the legs (Ti1, arrow) and theguidepost neurons (Fe1, Tr1, and Cx1, arrowheads) are labeled by mAb 10E6. Guidepost cellsalways label more intensely. Other neurons that are differentiating from the leg epithelium areseen out of focus. (F) Example of a sensory neuron in the antenna (arrow). The arrowheadpoints to two sibling cells that will become sensory neurons, which have not yet delaminatedfrom the epithelium. Scale bars: A-C, 50 µm; D, 25 µm; E, 75 µm; F, 40 µm.

5C). Dfollow(not scell bfollowwhichsiderecomm5E). Adefineon culto anaat 29-started
Twor Fadelayeexampthe ty(Con)A1 seat 33experiAcP csiderain all oembryA1 wbecaustage 1989)shown33% experiat a while anterialent 100%growtMethoof deAcP gfoundcell boThe edelay experiallow stop igrowtfollow
In embryof 8.8delay compain con6). It abseniment
141Expression pattern and function of Lazarillo
uring this period other commissural neurons start to the AcP axons, but they are not labeled by mAb 10E6hown). The AcP growth cone reaches the contralateralody and then turns anteriorly at 36% of development,ing the longitudinal vMP2 axon (open arrow, Fig. 5D), is not labeled by mAb 10E6. The AcP axons can be con-d followers along this pathway. They reach the posteriorissure of the S1 segment at 39% of development (Fig.ll the stages are schematically drawn in Fig. 5F. We
d two types of antibody perturbation experiment basedture periods that give us the greatest temporal resolutionlyze the AcP pathway. Type A experiments were started32% of development, while type B experiments were at 33-35%.
o effects were observed in embryos cultured with mAbb 10E6. (1) The position of AcP growth cones wasd with respect to control embryos. Representativeles are shown in Fig. 6. In
propose could be explained by a partial blocking of Lazarillofunction by mAb 10E6. No differences were observed in AcPcells in other segments. The delay in both types of experimentincreases with mAb concentration and reaches a saturationlevel as predicted for a specific interaction. Statistically sig-nificant differences in growth are observed at 0.88 µM andhigher. Experiments with 8.8 µM of Fab antigen binding sitesresult in similar delays in growth in both types of experiment,ruling out an artifactual cross linking of Lazarillo moleculesby the bivalent mAb. Other antibodies that label the AcP cells(see Materials and Methods) were used to assess whether otherIgGs can cause the observed delays. Neither the mAb 10H11,nor the anti-HRP antibodies produced a significant delay. Datagathered from 52 independent experiments, including 369control and 261 experimental embryos, are shown in Table 1.
(2) The AcP growth cones are misrouted during the cultureperiod. In type A experiments (Fig. 7A-D), a significant propor-
pe A control experiment the AcP cells of the S2 andgments reach the midline% of development. Inmental embryos (Exp) theells of S2 showed a con-ble delay while AcP cellsther segments of the sameo grow normally. Segmentas chosen for comparisonse it has a developmentalsimilar to S2 (Patel et al.,. The type B experiment in Fig. 6 was started atof development. Thesemental AcP cells remainedstage equivalent to 33%,control cells reach the nextor segment, a stage equiv-to 38%. This represents a delay compared to controlh (see Materials andds). Although the degreelay varies, experimentalrowth cones are frequently stalled at the contralateraldy in type B experiments.ffects are described as ain position because our
mental assay does notus to distinguish among an growth, a lower rate ofh or normal growthed by axon retraction.
both types of experiment,os cultured in the presence µM mAb 10E6 showed a
of approximately 35%red to the growth attainedtrols (see Table 1 and Fig.is important to note the
ce of delay in some exper-al AcP cells, which we
Fig. 4. Lazarillo is also expressed outside the nervous system. Embryos are viewed from the dorsalsurface with DIC optics. Anterior is up.(A) The Malpighian tubules express Lazarillo as soon as theystart invaginating from the proctodaeum into the body cavity (45% of development). Arrowheadpoints to one of the six groups of cells. (B) Labeled cells (arrowhead) are restricted to the slightlyswollen tips of the growing tubules (55% embryo). (C,D) Amoebic-shaped cells expressing Lazarillooriginate at the tip of the proctodaeum. (C) RNA in situ hybridization of a 29% embryo, (D) mAblabeling of a 32% embryo. (E) Lazarillo expression is maintained while these mesodermal cells aredifferentiating and migrating anteriorly along the sides of the abdominal body wall (40% embryolabeled with mAb). Arrow points the anterior edge of the proctodaeum in C-E. (F) The cellscomposing the subaesophageal body (arrowhead) are darkly stained with mAb 10E6 (45% embryo).Scale bars: A-E, 100 µm; F, 200 µm.

142
tion of Ashowed anext S1 s470 cellsposterior in type Babnormalof the axowith mA190 cells267 cells
Defascthe comobservedProfuse experimethey wesome canomaloimental never exareas, sugthe anternot selec
Althousensory the postedelayed atations wHoweverby 10E6other seneurons Ti1 neuronormal. Wall otherSeveral fascicles appear chosen general dThe effemAb 10Escatteredgesting thwere mo
Thus, by mAb cones tochange tresults suin the specific system o
DISCUS
Lazarilloembryogment and
D
cP cells cultured in the presence of mAb or Fab 10E6n anomalous axon growth directed anteriorly toward theegment (53 of 332 cells in experimental embryos; 8 of in controls; Fig. 7I). Axon extension in lateral ordirections was never observed. The AcP growth cones experiments follow their normal pathway. However, anterior branches arising from the commissural portionn were seen in embryos cultured
b or Fab 10E6 (Fig. 7E,I; 16 of in experimental embryos; 4 of in controls).iculation of AcP axons alongmissural pathway was never in either type of experiment.filopodia were observed inntal embryos (Fig. 7F,G) butre not scored. Interestingly,
on its physiological role during embryogenesis. Although noother molecule has been described with the same tissue distri-bution as Lazarillo, a close resemblance exists with DSS-8, acell surface protein in the cockroach embryo (Denburg et al.,1989). However, its localization in other tissues, as well as bio-chemical properties (Wang et al., 1992), weaken the possibil-ity of homology between these two proteins.
. Sánchez, M. D. Ganfornina and M. J. Bastiani
ontrol filopodia explore theus pathways followed in exper-conditions (Fig. 7H), but theytend to posterior and lateralgesting that in control embryos
omedial region is surveyed butted for growth.gh not extensively analyzed,
axons normally growing alongrior branch of the ISN are oftennd have altered anterior orien-hen cultured with mAb 10E6., other axonal pathways labeled, such as the AcP neurons ingments (Fig. 6), their sibling(AsM) that pioneer the SN, thens of the leg, and the MFT weree have not, however, assayed
labeled neurons at this time.longitudinal and commissuralunlabeled by mAb 10E6 also
correctly formed during theculture periods, ruling out aisruption of the axonal scaffold.cts caused by the presence of6 or Fab fragments were evenly
throughout experiments, sug-at no particular clutches of eggs
re susceptible to perturbation.blocking the epitope recognized10E6 causes the S2 AcP growth delay, and often dramaticallyheir direction of growth. Theseggest that Lazarillo is involvedoutgrowth and guidance ofgrowth cones in the nervousf the grasshopper embryo.
SION
is expressed throughoutenesis, postembryonic develop- adulthood. We have focused
Fig. 5. The growing axons of the anterior commissure pioneer (AcP) cells follow astereotyped pathway. (A) At 29% of development the AcP cells start producing filopodialextensions without any preferential direction. (B) At 32% of development the growth coneextends toward the midline. (C) At 34% of development the two AcP axons have crossed themidline and fasciculated with each other. (D) At 36% of development, after encountering thecontralateral cell body, they extend anteriorly following the unlabeled vMP2 axon (openarrow). (E) At 39% of development the AcP axons have reached the posterior commissure ofthe next anterior segment (S1). Scale bar, 25 µm. (F) Staging of AcP cells by 1% intervals.Two types of antibody perturbation experiment were conducted. Type A experiments startedat 29-32% of development. Type B experiments started at 33-35% of development.

143Expression pattern and function of Lazarillo
Table 1. Average delay values and total number of observations in each group of experiments Antigen binding site 0.176 0.88 1.76 8.88 17.6concentration (µM) n delay(%) n delay(%) n delay(%) n delay(%) n delay(%)
TYPE A:Control 10 0±3.1 96 0±3.5 48 0±6.6 200 0±4.7 54 0±7.1mAb 10E6 12 2.1±11.3 94 25.3±4.4* 50 33.3±5.1* 38 35.1±5.9* 54 38.9±5.8*Fab 10E6 96 43.1±4.6*mAb 10H11 38 2.8±2.3αHRP serum 24 −3.1±2.7
TYPE B:Control 12 0±8.8 36 0±5.5 48 0±4.1 102 0±4.1 26 0±6.3mAb 10E6 12 −3.8±8.5 40 24.5±7.7** 48 37.3±6.1* 46 34.9±6.0* 26 25.7±7.3**Fab 10E6 30 37.9±6.6**mAb 10H11 43 7.8±3.3
Data expressed in mean±s.e.m. Asterisks mean that the difference from the control was statistically significant (Mann-Whitney U test, *P<0.005, **P<0.05).
Fig. 6. The AcP axons are delayed in adose dependent way when embryos arecultured in the presence of mAb 10E6 orits Fab fragments. In a Type Aexperiment, started at the 29% stage, theAcP cells are shown in the S2 and A1segments of an embryo cultured for 24hours in control conditions (Con) andanother cultured with 8.8 µM of mAb10E6 (Exp). A segment-specific greaterintensity of labeling is observed in S2where a clear arrest is seen in theexperimental embryo. The AcP cells incontrol as well as in the experimental A1segment have reached the midline(arrowhead). The type B experimentshown, started at 33% of development.In the control embryo the AcP growthcones have completed their navigation tothe S1 segment (arrowhead in Con). Inthe experimental embryo, cultured with8.8 µM of mAb 10E6, cells are still atthe midline and their growth cones sendabundant filopodia (arrowhead in Exp).Growth cone delay is plotted versusantigen binding site concentration inboth types of experiment. Each cell wasstaged according to Fig. 5F, and thegrowth in culture estimated bysubtracting the average stage of the t=0embryos. Delay was defined as thedifference between each experimentalgrowth value and the average growth ofthe control embryos in a givenexperiment. The average delay in eachgroup of experiments with the sameconcentration of immunoperturbant isnormalized to the average growthattained in the control cultures. Errorbars represent s.e.m. Asterisks indicate astatistically significant difference fromthe control (Mann-Whitney U test,*=P<0.005, **=P<0.05). Scale bars:Type A, 30 µm; Type B, 25 µm.
mAb
mAb
144
LazariMalpighiare involbe capturbe internor activeMalpighifunctionsand retin
D ina and M. J. Bastiani
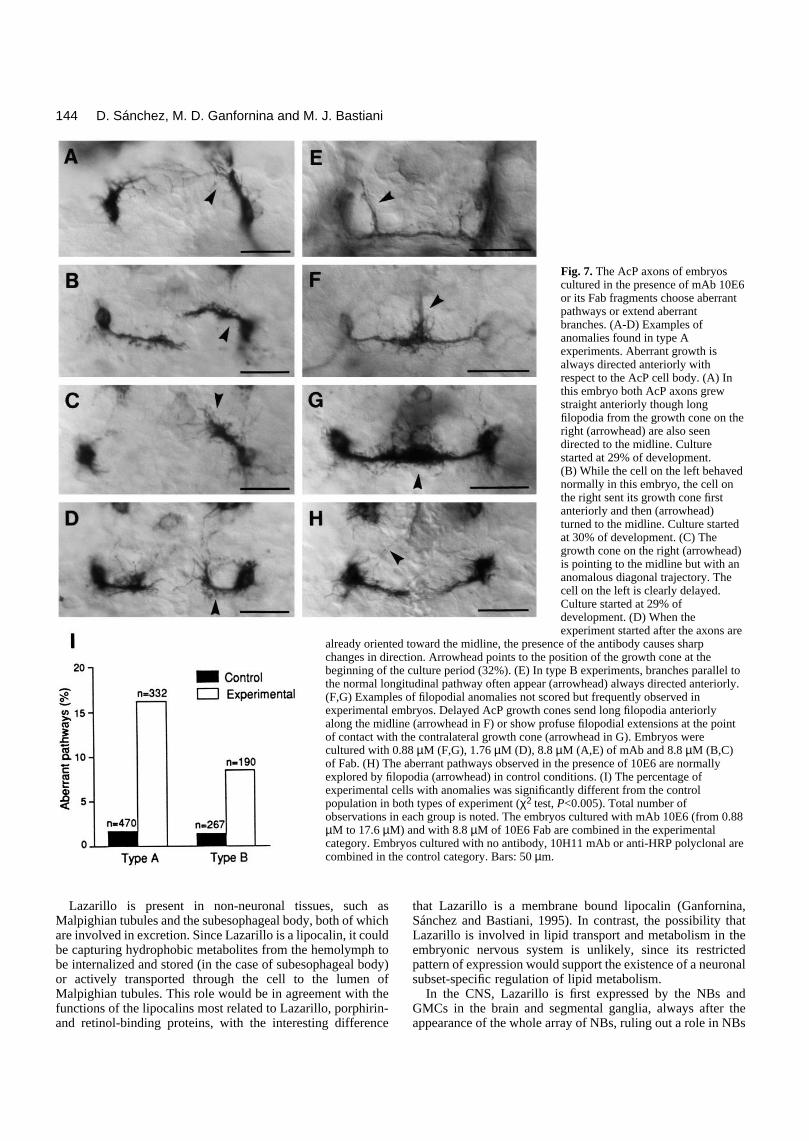
Fig. 7. The AcP axons of embryoscultured in the presence of mAb 10E6or its Fab fragments choose aberrantpathways or extend aberrantbranches. (A-D) Examples ofanomalies found in type Aexperiments. Aberrant growth isalways directed anteriorly withrespect to the AcP cell body. (A) In
. Sánchez, M. D. Ganforn
this embryo both AcP axons grewstraight anteriorly though longfilopodia from the growth cone on theright (arrowhead) are also seendirected to the midline. Culturestarted at 29% of development.(B) While the cell on the left behavednormally in this embryo, the cell onthe right sent its growth cone firstanteriorly and then (arrowhead)turned to the midline. Culture startedat 30% of development. (C) Thegrowth cone on the right (arrowhead)is pointing to the midline but with ananomalous diagonal trajectory. Thecell on the left is clearly delayed.Culture started at 29% ofdevelopment. (D) When theexperiment started after the axons are
already oriented toward the midline, the presence of the antibody causes sharpchanges in direction. Arrowhead points to the position of the growth cone at thebeginning of the culture period (32%). (E) In type B experiments, branches parallel tothe normal longitudinal pathway often appear (arrowhead) always directed anteriorly.(F,G) Examples of filopodial anomalies not scored but frequently observed inexperimental embryos. Delayed AcP growth cones send long filopodia anteriorlyalong the midline (arrowhead in F) or show profuse filopodial extensions at the pointof contact with the contralateral growth cone (arrowhead in G). Embryos were
llo is present in non-neuronal tissues, such asan tubules and the subesophageal body, both of whichved in excretion. Since Lazarillo is a lipocalin, it coulding hydrophobic metabolites from the hemolymph toalized and stored (in the case of subesophageal body)ly transported through the cell to the lumen ofan tubules. This role would be in agreement with the of the lipocalins most related to Lazarillo, porphirin-ol-binding proteins, with the interesting difference
that Lazarillo is a membrane bound lipocalin (Ganfornina,Sánchez and Bastiani, 1995). In contrast, the possibility thatLazarillo is involved in lipid transport and metabolism in theembryonic nervous system is unlikely, since its restrictedpattern of expression would support the existence of a neuronalsubset-specific regulation of lipid metabolism.
In the CNS, Lazarillo is first expressed by the NBs andGMCs in the brain and segmental ganglia, always after theappearance of the whole array of NBs, ruling out a role in NBs
cultured with 0.88 µM (F,G), 1.76 µM (D), 8.8 µM (A,E) of mAb and 8.8 µM (B,C)of Fab. (H) The aberrant pathways observed in the presence of 10E6 are normallyexplored by filopodia (arrowhead) in control conditions. (I) The percentage ofexperimental cells with anomalies was significantly different from the controlpopulation in both types of experiment (χ2 test, P<0.005). Total number ofobservations in each group is noted. The embryos cultured with mAb 10E6 (from 0.88µM to 17.6 µM) and with 8.8 µM of 10E6 Fab are combined in the experimentalcategory. Embryos cultured with no antibody, 10H11 mAb or anti-HRP polyclonal arecombined in the control category. Bars: 50 µm.

differenof the Nidentitysubset reportefied NBwindowThese NB-idelineageand poentiatioTechnacomparexpressfunctioLazarilfor motiation lineagenew cemechansystem
An ifrom tdivisiobe formeage-idonce a shows why Laneuronwith thcalled antibod(Snow betweebohydrneuriteISN, wthe proISN wial., 199
The ing Laneuronhas bee(motonor withment (fgrammless, wneurotr10E6. with thmotoneISN, dsegmensophagthose m
The
145Expression pattern and function of Lazarillo
tiation. However, the restriction to approximately 40%Bs in a segmental ganglion suggests Lazarillo is a NB- molecule. No exact correlation is found between the
of NBs labeled by 10E6 and the NB-identity moleculesd so far (Doe, 1992; Doe and Technau, 1993). Identi-s and GMCs express Lazarillo during a particular time and there are also examples of intermittent expression.
features suggest that Lazarillo belongs to the group ofntity molecules that specify the GMC identity in a sub--specific manner. The different cell types (NBs, GMCsstmitotic neurons) that are involved in neuronal differ-n express a number of proteins (Doe, 1992; Doe andu, 1993) that appear distributed in different cellulartments: nuclear proteins, which regulate geneion, extracellularly secreted proteins, which mightn as diffusible signals, and membrane bound molecules.lo, a membrane protein, could be acting as a receptorrphogenetic signals participating in neuronal differen-
and its restriction to particular axon fascicles during theprocesses of axonogenesis, fascicle formation and pathwayselection, suggest a role in pathfinding. We tested this hypoth-esis by perturbing the molecular interactions of Lazarillo withmAb 10E6 in cultured grasshopper embryos. A pair of cellsthat pioneer the first fascicle in the anterior commissure (AcPcells) were chosen for this study and two kinds of perturbationwere observed: a delay of the AcP growth cones and an alteredtrajectory of growth. Nevertheless, a remarkable absence ofcommissural defects occurs in the AcP cells of body segmentsother than S2, which correlates with segmental quantitative dif-ferences in the level of Lazarillo expression as revealed byimmunocytochemistry and in situ hybridization. This could beexplained either by proposing the existence of redundantguidance cues and a segment-specific role for Lazarillo inguiding commissural axons, or an only partial perturbationeffect by mAb 10E6.
The AcP growth cone emerges from the cell body and
n
l
e
T
and determination. mAb 10E6 can be considered a sub--specific marker and will be a useful tool to ascertainll lineages, an essential requirement to understand theisms generating cellular diversity in the nervous
.ntriguing finding in the PNS is that Lazarillo is absenthe SOCs. As every neuron arises by symmetrical from its own SOC, neuronal sublineages need not toed during neurogenesis in the PNS. Therefore, sublin-entity molecules might not be necessary. Likewise,cluster of sensory neurons is formed the emerging nerveno distinctive fascicle subdivisions. This could explainzarillo expression is not restricted to subsets of sensorys. A particularly interesting sensory cluster associatede ISN is labeled by mAb 10E6. This group we haveGp, has been missed in other studies where anti-HRPies, which recognizes a complex carbohydrate epitopeet al., 1987), were used. The lack of correspondencen anti-HRP and 10E6 labeling suggests a different car-ate moiety in Lazarillo. Since the lGp cells send theirs toward the periphery before any other axon travels thee propose a guidepost role for them. This contradictsposal that dBw cells pioneer the peripheral part of thethout help from any neuronal guidepost cell (Meier et1).
invariably grows toward the midline. It is currently believedthat guidance of commissural neurons relies on specializedmidline cells, the floor plate cells in vertebrates (Tessier-Lavigne et al., 1988; Hatta, 1992) and the midline glial cellsin invertebrates (Menne and Klämbt, 1994). There is evidencefor both attractive and repulsive guidance cues provided by themidline, possibly in the form of soluble morphogen gradients,that determine commissural versus ipsilateral growth cone tra-jectories (reviewed by Goodman and Shatz, 1993; Goodman,1994; Baier and Bonhoeffer, 1994). In those cases where theAcP growth cones are perturbed by mAb 10E6 they often stopgrowing toward the midline, and in a significant number ofcases they turn and always grow in an anterior direction. Asmentioned above, one could propose that Lazarillo participatesin the lipid metabolism of the cell and, in this case, is necessaryfor the rapid expansion of membrane associated with axono-genesis. Nevertheless, many AcP axons were able to extendcomparable distances along aberrant pathways, and we proposethat the delay is a consequence of altering a specific signalingmechanism for directional guidance. The invariant anteriorextension of abnormal growth cones is a strong indication forthe existence of an anterior guidance signal.
The AcP growth cone normally encounters and fasciculateswith its contralateral homologue at the midline. This contactwith its homologue may allow the AcP growth cone to extend
sublineage constraint observed in the NB array express-zarillo is in agreement with the restricted pattern ofs and axonal bundles labeled by 10E6. No correlationn detected though with a particular mature cellular typeeurons, interneurons and sensory neurons are labeled) cells bearing functional significance during develop-ascicle pioneers and followers, neurons undergoing pro-ed cell death or not, all express Lazarillo). Neverthe- have not assayed all other possibilities e.g., a common
ansmitter expression in the sublineages labeled byhere is, however, a striking example of a correlation
e pathway a certain neuron takes. The aCC, segmentalurons that exit the CNS by the anterior branch of theo not express Lazarillo in thoracic or abdominalts, but they appear darkly labeled by 10E6 in the sube-eal segment S1 (Fig. 5E), where the ISN is absent andotoneurons follow a different pathway.presence of Lazarillo on growth cones and filopodia,
away from the peak of a midline morphogen. A similarmechanism in which cell-cell interactions are dominant tomorphogen gradients has been proposed to account for thebehavior of other pioneer neurons in the grasshopper embryo,such as the Q1 neurons in the CNS (Myers and Bastiani, 1993and unpublished observations), and the Ti1 neurons in the limbbud (Bentley and Caudy, 1983; Caudy and Bentley, 1986).Defasciculation between the two AcP axons was neverobserved in the mAb blocking experiments, indicating thatLazarillo is not involved in this selective axon-axon recogni-tion. However, although fasciculation with its homologue isnormal there are many anomalous branches that extend anteri-orly towards the segment border. There is strong experimentalsupport for a role of boundary regions, such as the segmentborder, in axon guidance (Wilson et al., 1993).
Next, the AcP axon leaves its homologue to turn and growanteriorly along the vMP2 fascicle. Commissural neurons mustoften initially ignore an ipsilateral target, but later specifically

146
recognizeversing ththat axonof cell su1988) a(RothbergImmunopfound starecognitio
Althouthose derLazarillo for commthe behavrespond morphogesegment receptor fwith mAbprevent gto responspecific cgradientsthe midliWhen theside it is nthe recepwould prcone fromresponse midline sabnormal
In this morphogecell surfaThe earlywould acThe vast of substrpathway to define involved rectly in choices. ligand thunknownsuggests (Ganfornwill also bproteins product (membersaxon guid1994). Fmodel.
The redirectly itheir corralso inferbe involvas well a
D
that same target on the contralateral side after tra-e midline. Experimental observations indeed suggests traversing the midline can change their expressionrface proteins (Bastiani et al, 1987; Dodd and Jessell,nd therefore acquire new pathfinding abilities et al., 1990; Campbell and Peterson, 1993).erturbed AcP growth cones are often delayed andlled at the contralateral cell body, suggesting thatn of the vMP2 axon is impaired.
gh other hypotheses are formally possible, such asived from considering mAb 10E6 as an agonist offunction, we propose the following model to accountissure formation in the grasshopper embryo based onior of the perturbed AcP growth cones. The AcP cellsto segment wide mediolateral and anteroposteriorn gradients, possibly originating at the midline and
border respectively. We propose that Lazarillo is the
system of insects. A better understanding of its functional roleswill be of interest to formulate new hypotheses about thecellular and molecular mechanisms governing the processes ofneuronal identity determination and axon guidance.
We are grateful to the members of our laboratory and of the BiologyDepartment for support and critical reading of the manuscript; C.Goodman for advice, and E. Carpenter for generating mAb 10E6. Thiswork was supported by NIH grant to M. J. B. (NS25387). M. D. G.holds a fellowship from the Fulbright/MEC of Spain, and D. S. is therecipient of a NIH postdoctoral fellowship (1F05TW04686-01).
REFERENCES
Baier, H. and Bonhoeffer, F. (1994). Attractive axon guidance molecules.Science 265, 1541-1542.
Bastiani, M. J., Harrelson, A. L., Snow, P. M. and Goodman, C. S. (1987).
. Sánchez, M. D. Ganfornina and M. J. Bastiani
or a midline morphogen. Blocking Lazarillo function Expression of Fasciclin I and II glycoproteins on subsets of axon pathways
10E6 before the AcP cells reach the midline wouldrowth toward the midline and allow the growth coned to an anterior morphogen. Once at the midline,ell-cell interactions would be dominant to the global, allowing the AcP growth cone to extend away fromne by fasciculating on its contralateral homologue. perturbed AcP growth cone reaches the contralateralot able to recognize vMP2. We propose that alteringtion of the midline signal by perturbing Lazarilloevent the switch in the specificity of the AcP growth its homologue to the vMP2 fascicle. In contrast, the
to the anterior signal seems to be independent of theignal, since the immunoperturbed AcP cell extends processes both before and after crossing the midline.model the interplay between the anterior and midlinens determines the polarity of growth, and localized
ce cues determine the precise position of pathways. mediolateral and anteroposterior segment gradientscount for the behavior of pioneering growth cones.majority of later developing neurons would make useate bound cues on individual fascicles for theirchoices, but may still rely on these global gradientstheir polarity of growth. Lazarillo would be directlyin the reception of the midline morphogen and indi-
during neuronal development in the grasshopper. Cell 48, 745-755.Bentley, D., Keshishian, H., Shankland, M. and Toroian-Raymond, A.
(1979). Quantitative staging of embryonic development of the grasshopper,Schistocerca nitens. J. Embryol. Exp. Morph. 54, 47-74.
Bentley, D. and Keshishian, H. (1982). Pathfinding by peripheral pioneerneurons in grasshoppers. Science 218, 1082-1088.
Bentley, D. and Caudy, M. (1983). Pioneer axons lose directed growth afterselective killing of guidepost cells. Nature 304, 62-65.
Bentley, D. and O’Connor, T. P. (1992). Guidance and steering of peripheralpioneer growth cones in grasshopper embryos. In The Nerve Growth Cone,(ed. P. C. Letourneau, S. B. Kater and E. R. Macagno), pp. 265-282. NewYork: Raven Press.
Bixby, J. L. and Harris, W. A. (1991). Molecular mechanisms of axon growthand guidance. Annu. Rev. Cell Biol. 7, 117-159.
Campbell, R. M. and Peterson, A. C. (1993). Expression of a lacZ transgenereveals floor plate cell morphology and macromolecular transfer tocommissural axons. Development 119, 1217-1228.
Campos-Ortega, J. A. (1994). Cellular interactions in the developing nervoussystem of Drosophila. Cell 77, 969-975.
Carpenter, E. M. and Bastiani, M. J. (1990) Expression and characterizationof nervous system antigens in grasshopper embryos. Soc. Neurosci. Abstr.16, 303.
Caudy, M. and Bentley, D. (1986). Pioneer growth cone steering along a seriesof neuronal and non-neuronal cues of different affinities. J. Neurosci. 6,1781-1795.
Chapman, R. F. (1971). Nitrogenous excretion and salt and water regulation.In The insects. Structre and Function (ed. W. S. Bullough), pp. 490-512. NewYork: Elsevier.
Crossley, A. C. (1985). Nephrocytes and pericardial cells. In Comprehensive
the switch in specificity for subsequent pathwayThe identity of the proposed morphogens and theat the lipocalin Lazarillo can bind is currently, but the phylogenetic analysis of Lazarillo sequenceheme metabolites and retinoids as strong candidatesina, Sanchez and Bastiani, 1995). In this context, ite important to test the binding of Lazarillo to soluble
secreted by midline structures such as the slit geneRothberg et al., 1990) or the recently discovered of the netrin family of outgrowth-promoting andance proteins (Serafini et al., 1994; Kennedy et al.,
uture experiments will test the predictions of this
sults presented in this paper suggest that Lazarillo isnvolved in the guidance of developing axons alongect pathways. A multifunctional role of Lazarillo isred from its complex expression pattern; it could alsoed in the specification of NBs and neuronal identitiess in the development and function of the excretory
Insect Physiology, Biochemistry and Pharmacology (eds. G. A. Kerkut andL. I. Gilbert), Vol 3, pp 487-515. Oxford: Pergamon Press.
Cui, X. and Doe, C. Q. (1992). ming is expressed in neuroblast sublineages andregulates gene expression in the Drosophila central nervous system.Development 116, 943-952.
Denburg, J. L., Norbeck, B. A., Caldwell, R. T. and Marner, J. A. M.(1989). Developmental stage-specific antigens in the nervous system of thecockroach. Dev. Biol. 132, 1-13.
Dodd, J. and Jessell, T. M. (1988). Axon guidance and the patterning ofneuronal projections in vertebrates. Science 242, 692-699.
Doe, C. Q., Kuwada, J. Y. and Goodman, C. S. (1985). From epithelium toneuroblasts to neurons: the role of cell interactions and cell lineage duringinsect neurogenesis. Phil. Trans. R. Soc. Lond. 312, 67-81.
Doe, C. Q. (1992). Molecular markers for identified neuroblasts and ganglionmother cells in the Drosophila central nervous system. Development, 116,855-863.
Doe, C. Q. and Technau, G. M. (1993). Identification and cell lineage ofindividual neuronal precursors in the Drosophila CNS. Trends Neurosci. 16,510-514.
Elkins, T., Zinn, K., McAllister, L., Hoffmann, F. M. and Goodman, C. S.(1990). Genetic analysis of a Drosophila neural cell adhesion molecule:Interaction of Fasciclin I and Abelson tyrosine kinase mutations. Cell 60,565-575.

Ex
Ganfornina, M. D., Sánchez, D. and Bastiani, M. J. (1995). Lazarillo, a GPI-linked surface glycoprotein restricted to a subset of neurons in thegrasshopper embryo, is a new member of the lipocalin family. Development121, 123-134..
Goodman, C. S., Raper, J. A., Ho, R. K. and Chang, S. (1982). Pathfinding ofneuronal growth cones in grasshopper embryos. In Developmental Order: ItsOrigin and Regulation (ed. S. Subtelny and P. B. Green), pp. 275-316. NewYork: Alan R. Liss.
Goodman, C. S., Bastiani, M. J., Doe, C. Q., du Lac, S., Helfand, J.,Kuwada, J. Y. and Thomas, J. B. (1984). Cell recognition during neuronaldevelopment. Science 225, 1271-1279.
Goodman, C. S. and Shatz, C. J. (1993). Developmental mechanisms thatgenerate precise patterns of neuronal connectivity. Cell 72/Neuron 10(Suppl.), 77-98.
Goodman, C. S. (1994). The likeness of being: Phylogenetically conservedmolecular mechanisms of growth cone guidance. Cell 78, 353-356.
Grenningloh, G., Rehm, E. J. and Goodman, C. S. (1991). Genetic analysisof growth cone guidance in Drosophila: Fasciclin II functions as a neuronalrecognition molecule. Cell 67, 45-57.
Harrelson, A. L. and Goodman, C. S. (1988). Growth cone guidance ininsects: Fasciclin II is a member of the immunoglobulin superfamily. Science242, 700-708.
Hatta, K. (1992). Role of the floor plate in axonal patterning in the zebrafishCNS. Neuron 9, 629-642.
Hedgecock, E. M., Culotti, J. G. and Hall, D. H. (1990). The unc-5, unc-6,and unc-40 genes guide circumferential migrations of pioneer axons andmesodermal cells in the epidermis in C. elegans. Neuron 4, 61-85.
Hockfield, S. (1987). Mab to a unique cerebellar neuron generated byimmunosuppression and rapid immunization. Science 237, 67-70.
Huff, R., Furst, A. and Mahowald, A. P. (1989). Drosophila embryonicneuroblasts in culture: autonomous differentiation of specificneurotransmitters. Dev. Biol. 134, 146-157.
Kennedy, T. E., Serafini, T., de la Torre, J. R. and Tessier-Lavigne, M.(1994). Netrins are diffusible chemotropic factors for commissural axons inthe embryonic spinal cord. Cell 78, 425-435.
Kolodkin, A. L., Matthes, D. J., O’Connor, T. P., Patel, N. H., Admon, A.,Bentley, D. and Goodman, C. S. (1992). Fasciclin IV: sequence,expression, and function during growth cone guidance in the grasshopperembryo. Neuron 9, 831-845.
Kuwada, J. Y. (1992). Growth cone guidance in the zebrafish central nervoussystem. Curr. Opin. Neurobiol. 2, 31-35.
McIntire, S. L., Garriga, G., White, J., Jacobson, D. and Horvitz, H. R.(1992). Genes necessary for directed axonal elongation or fasciculation in C.elegans. Neuron 8, 307-322.
Meier, T., CembryonSchistoce112, 241-
Menne, T. DrosophiDevelopm
Myers, P. Zmigration143.
Patel, N. Hcloning oaxon path
Patel, N. Hengrailed107, 201-
Rothberg, J(1990). Sglia and cGenes De
Seaver, E., and tempaxon out881-893
Serafini, TTessier-promotin
Snow, P., specific Drosoph
Tessier-LaT. M. (1central n
Wang, L., developmin pathfin
Whitingtothe locusganglia.
Wilson, S.boundari323.
147pression pattern and function of Lazarillo
habaud, F. and Reichert, H. (1991). Homologous pattern in theic development of the peripheral nervous system in the grasshopperrca gregaria and the fly Drosophila melanogaster. Development253.V. and Klämbt, C. (1994). The formation of commissures in thela CNS depends on the midline cells and on the Notch gene.ent 120, 123-133.. and Bastiani, M. J. (1993). Growth cone dynamics during the of an identified commissural growth cone. J. Neurosci. 13, 127-
., Snow, P. M. and Goodman, C. S. (1987). Characterization andf Fasciclin III: a glycoprotein expressed on a subset of neurons andways in Drosophila. Cell 48, 975-988.., Kornberg, T. B. and Goodman, C. S. (1989). Expression ofduring segmentation in grasshopper and crayfish. Development
212.. M., Jacobs, J. R., Goodman, C. S. and Artavanis-Sakonas, S.
lit: an extracellular protein necessary for development of midlineommissural axon pathways contains both EGF and LRR domains.v. 4, 2169-2187.
Karlstrom, R. O. and Bastiani, M. J. (1991). The restricted spatialoral expression of a nervous-system-specific antigen involved in
growth during development of the grasshopper. Development 111,.., Kennedy, T. E., Galko, M. J., Mirzayan, C., Jessell, T. M. andLavigne, M. (1994). The netrins define a family of axon outgrowth-g proteins homologous to C. elegans UNC-6. Cell 78, 409-424.Patel, N., Harrelson, A. and Goodman, C. S. (1987). Neural-carbohydrate moiety shared by many surface glycoproteins inila and grasshopper embryos. J. Neurosci. 7, 4137-4144.vigne, M., Placzek, M., Lumsden, A. G. S., Dodd, J. and Jessell,988). Chemotropic guidance of developing axons in the mammalianervous system. Nature 336, 775-778.Feng, Y. and Denburg, J. L. (1992). A multifunctional cell surface
ental stage-specific antigen in the cockroach embryo: involvementding by CNS pioneer axons. J. Cell. Biol. 118, 163-176.
n, P. M. (1989). The early development of motor axon pathways int embryo: the establishment of the segmental nerves in the thoracicDevelopment 105, 715-721. W., Placzek, M. and Furley, A. J. (1993). Border disputes: Does play a role in growth-cone guidance? Trends Neurosci. 16, 316-
(Accepted 30 September 1994)