distribution of preprovasopressin mrna in the rat central nervous system
TRANSCRIPT

Distribution of Preprovasopressin mRNAin the Rat Central Nervous System
MARTIN HALLBECK,* OLA HERMANSON, AND ANDERS BLOMQVIST
Division of Cell Biology, Department of Biomedicine and Surgery, Faculty of Health Sciences,University of Linkoping, S-581 85 Linkoping, Sweden
ABSTRACTVasopressin released in the central nervous system has been shown to be involved both in
homeostatic mechanisms (e.g., water balance, thermoregulation, cardiovascular regulation,metabolism, and antinociception) and in higher brain functions (e.g., social recognition andcommunication, and learning and memory). Many nuclear groups have been proposed tosynthesize vasopressin, but available data are conflicting. We have used a sensitive in situhybridization technique to identify the distribution of the neurons that may be the origin ofthe vasopressin in the central nervous system of the male Sprague-Dawley rat. VasopressinmRNA-expressing neurons were most abundant in the hypothalamus (e.g., the paraventricu-lar, supraoptic, and suprachiasmatic nuclei) but were also seen in the medial amygdaloidnucleus, the bed nucleus of stria terminalis, and the nucleus of the horizontal diagonal band.Previously unreported vasopressinergic neurons were seen in the entorhinal and piriformcortices, the ventral lateral portion of the parabrachial nucleus, the pedunculopontinenucleus, and the rostral part of the ventral periaqueductal gray matter and the adjacentportion of the mesencephalic reticular nucleus. Vasopressin mRNA expression suggestive ofneuronal labeling was seen in the pyramidal layer of the CA1–3 fields and the dentate gyrus ofthe hippocampus. In addition, vasopressin mRNA expression, probably representing axonalmRNA, was detected over the hypothalamopituitary tract. No or insignificant preprovasopres-sin mRNA expression was present in the cerebellum, locus coeruleus, subcoeruleus, or thespinal cord. These findings provide novel information on the distribution of vasopressinneurons that are important for our understanding of how vasopressin acts in the brain. J.Comp. Neurol. 411:181–200, 1999. r 1999 Wiley-Liss, Inc.
Indexing terms: paraventricular hypothalamus; entorhinal cortex; hippocampus; parabrachial
nucleus; periaqueductal gray matter; axonal mRNA
In its classic role as a neuroendocrine hormone, vasopres-sin is antidiuretic and vasoconstrictive. However, vasopres-sin has also been shown to be a neurotransmitter in thecentral nervous system (for a review, see de Wied et al.,1993). Some of its effects as a neurotransmitter are closelycoordinated with its effects as a hormone, such as centralmaintenance of water balance (Buijs, 1978), thermoregula-tion (Cooper et al., 1979; Banet and Wieland, 1985),cardiovascular regulation (Stebbins et al., 1992; Malpasand Coote, 1994), and release of adrenocorticotropic hor-mone (ACTH) from the anterior pituitary (Wolfson et al.,1985). Other central effects involve metabolic regulation(Wideman and Murphy, 1993), alcohol tolerance (Hoffmanet al., 1990), sexual behavior (Sodersten et al., 1983;Smock et al., 1992), and anti-nociception (Berntson andBerson, 1980; Bodnar et al., 1980; Berson et al., 1983;Kordower and Bodnar, 1984; Thurston et al., 1988). Vaso-pressin is also involved in higher brain functions such associal recognition (Bluthe et al., 1990), mating (Young et
al., 1998), and learning and memory (Bohus, 1977; Lesh-ner and Roche, 1977; Bohus et al., 1978; de Wied andVersteeg, 1979; Metzger et al., 1993). In addition, vasopres-sin has been proposed to be involved in brain development(Boer et al., 1982).
Immunoreactivity to vasopressin has been demon-strated in many areas of the rat brain, but the reports onthe distribution of vasopressinergic cell bodies are conflict-ing. Thus, immunohistochemical studies have shown vaso-
Grant sponsor: Swedish Medical Research Council; Grant number: 7879;Grant sponsor: Swedish Society for Medical Research.
Dr. Hermanson’s current address is: Rosenfeld Lab, CMM West, Depart-ment and School of Medicine, UCSD, La Jolla, CA 92093.
*Correspondence to: Martin Hallbeck, Division of Cell Biology, Depart-ment of Biomedicine and Surgery, Faculty of Health Sciences, University ofLinkoping, S-581 85 Linkoping, Sweden. E-mail: [email protected]
Received 12 November 1996; Revised 23 February 1999; Accepted 16March 1999
THE JOURNAL OF COMPARATIVE NEUROLOGY 411:181–200 (1999)
r 1999 WILEY-LISS, INC.

pressinergic neurons in a variety of structures, such as thebed nucleus of stria terminalis (De Vries and Buijs, 1983;van Leeuwen and Caffe, 1983; Sofroniew, 1985), the verti-cal limb of the nucleus of the horizontal diagonal band(Sofroniew, 1985), the septum (Sofroniew, 1985), the para-ventricular hypothalamic nucleus (PVH; Buijs, 1978; DeVries et al., 1985), the supraoptic nucleus (De Vries et al.,1985), the suprachiasmatic nucleus (Buijs, 1978; So-froniew and Weindl, 1980), the dorsomedial hypothalamus(Caffe and van Leeuwen, 1983; De Vries et al., 1985), themedial amygdaloid nucleus (MEA; Caffe and van Leeu-wen, 1983; De Vries et al., 1985; Sofroniew, 1985), the locuscoeruleus (Caffe and van Leeuwen, 1983; Caffe et al., 1985;De Vries et al., 1985; Sofroniew, 1985), the nucleus subcoer-uleus (Caffe et al., 1985; Sofroniew, 1985), and the spinalcord (Kai-Kai et al., 1986). Because neuropeptides areoften produced in small amounts and are often rapidlytransported from the cell body, many of these studies usedthe axonal transport inhibitor colchicine to detect cell bodystaining. However, it is now known that colchicine hasprofound effects on the ultrastructural features of neurons(Alonso, 1988) and that it can affect peptide production(Ceccatelli et al., 1991; Rethelyi et al., 1991). This problemis overcome by using in situ hybridization histochemistrythat permits detection of peptidergic cell bodies in naiveanimals, but the in situ hybridization studies on vasopres-sin expression performed so far have mainly been focussedon the hypothalamus and basal forebrain (Uhl and Rep-pert, 1986; Urban et al., 1990; Lepetit et al., 1993; Miller etal., 1993; Planas et al., 1995), and no comprehensiveanalysis of the distribution of vasopressin transcripts inthe central nervous system has been carried out. Becausethe correct localization of vasopressinergic neurons isimportant for the understanding of the physiologic func-tions of vasopressin, we have, in the present study, exam-ined the central nervous system of rats for preprovasopres-sin mRNA (VP mRNA), by using a sensitive in situhybridization technique that uses long radiolabeled ribo-probes (Simmons et al., 1989). Part of these data has beenpresented in abstract form (Hallbeck et al., 1996a).
MATERIALS AND METHODS
Tissue preparation
Ten adult, naive, male Sprague-Dawley rats, each weigh-ing 200–500 g, (B&K Universal, Sollentuna, Sweden) wereused. The animals were maintained on a 12:12 light:darkcycle and were given food and water ad libitum. Thestudies were approved by the Animal Care and UseCommittee at the University of Linkoping.
The rats were anesthetized with sodium pentobarbital(100 mg/kg; Apoteksbolaget, Umea, Sweden) and perfused
transcardially with 100 ml of 0.9% NaCl (room tempera-ture), followed by 500 ml of 4% paraformaldehyde (PFA) in0.1 M phosphate-buffered saline (PBS; pH 7.4, 4°C). Thebrain and spinal cord were removed and placed in 20%sucrose and 4% PFA in 0.1 M PBS overnight (4°C), andthen cut transversely at 20µm on a freezing microtome.The sections were collected in 4% PFA in 0.1 M PBS. Everyeighth (brain) or every tenth (spinal cord) section was usedfor in situ hybridization against VP mRNA, and an adja-cent section was used for Nissl staining. In some controlexperiments (see below), adjacent sections were processedfor preprooxytocin (OT) or preproenkephalin mRNA. Otheradjacent sections were treated with a sense VP mRNAprobe or pretreated with RNase before vasopressin hybrid-ization. The sections for in situ hybridization were mountedon poly-L-lysine coated slides, and stored at 270°C untiluse.
Probes
Vasopressin. The probe against VP mRNA was pre-pared from a 0.6-kb preprovasopressin cDNA (Rehbein etal., 1986), subcloned into a Bluescript KS1 vector. Anti-sense probe synthesis and labeling were performed byusing T3 polymerase (Promega, Madison, WI) and 35S-labeled UTP (Amersham, Amersham, UK; for in situhybridization) or 32P-labeled UTP (Amersham; for dot blothybridization) after linearization with BamHI (Promega).A sense probe was synthesized with T7 polymerase (Pro-mega) after linearization with HindIII (Promega).
Oxytocin. An antisense OT mRNA probe was pre-pared from a 0.6-kb preprooxytocin cDNA (Rehbein et al.,1986), subcloned into a pSP65 vector. Probe synthesis wasdone with SP6 polymerase (Promega) after linearizationwith HindIII. For sense OT mRNA synthesis, the 0.6-kbpreprooxytocin cDNA was subcloned into a BluescriptKS1 vector. Linearization and synthesis was done withEcoRI (Promega) and T3 polymerase, respectively.
Enkephalin. A preproenkephalin probe was preparedfrom a 0.9-kb preproenkephalin cDNA, part of the pYSEAIplasmid (Yoshikawa et al., 1984).Antisense preproenkepha-lin probe synthesis was done with SP6 polymerase afterlinearization with SacI (Promega).
In situ hybridization
In situ hybridization was performed as described bySimmons et al. (1989). The slides were treated with 4%PFA in 0.1 M phosphate buffer for 30 minutes, prehybrid-ized with 0.001% proteinase K (Boehringer Mannheim,Mannheim, Germany) in a solution of 0.1 M Tris and 0.05M ethylenediaminetetraacetic acid (EDTA; Sigma, St.Louis, MO) for 30 minutes at 37°C, dehydrated withgraded ethanols, and vacuum dried.
Respective probe was placed in hybridization buffer(50% formamide, 10% dextran sulfate, 13 Denhardt’ssolution, 0.3 M NaCl, 10 mM Tris, and 1 mM EDTA)containing tRNA (0.5 mg ml-1, Boehringer Mannheim) anddithiothreitol (0.1 M, Sigma), and the specific activity wasadjusted to 5 3 106 cpm ml-1. The hybridization wasperformed at 58 6 1°C for 16–18 hours. The slides werethen rinsed in 43 standard saline citrate (SSC), beforedigestion with 0.002% RNase A (Boehringer Mannheim) in0.5 M NaCl, 10 mM Tris, and 1 mM EDTA (30 minutes at37°C). The slides were then rinsed in falling concentra-tions of SSC buffers before being heated to 65–70°C in0.13 SSC for 30 minutes. Dithiothreitol (1 mM, Sigma)
Abbreviations
OT mRNA (prepro)oxytocin mRNAPVH paraventricular hypothalamic nucleusap anterior parvocellular subdivisiondp dorsal parvocellular subdivisionlp lateral parvocellular subdivisionmp medial parvocellular subdivisionmpdd dorsal part the dorsal mpmpdv ventral part of the dorsal mpmpv ventral mppm posterior magnocellular subdivisionVP mRNA (prepro)vasopressin mRNA
182 M. HALLBECK ET AL.

was added to all buffers. After dehydration and delipidiza-tion, the slides were dipped in photographic emulsion(Kodak NTB2 [Kodak, Rochester, NY], diluted 1:1 indistilled water), stored at 220°C for 10–21 days, developedin Kodak D-19 (Kodak, Paris, France), fixed, and cover-slipped. All chemicals were from Merck (Darmstadt, Ger-many) if not otherwise stated.
Controls
To ensure that the in situ hybridization and autoradiog-raphy did not produce artifactual labeling, the followingcontrols were performed: (1) hybridization with probesdirected toward other mRNAs (preproenkephalin and pre-prooxytocin); (2) hybridization with 35S-labeled sense-strand cRNA; and (3) RNase treatment (0.002% RNase A,37°C, 60 minutes) of sections before hybridization. Toavoid digestion of the probe the RNase treatment was donebefore the PFA and the proteinase K steps.
In addition to these controls, the specificity of the VPmRNA probe was assessed by dot blot hybridization. Twoidentical dot blot membranes (Hybond N, Amersham) wascreated by using a dot blot apparatus (Bio-Rad, Hercules,CA) and VP or OT mRNA (obtained through synthesis ofcold sense probe from the respective cDNAs; see above)was subsequently fixed to the membrane in an ultravioletcross-linker (Stratagene, La Jolla, CA). The membraneswere prehybridized for two hours (42°C) in plastics bags.Prehybridization buffer contained 50% (vol/vol) formamide,53 SSC, 13 Denhart’s solution, 100 mg of heat-denaturedsheared salmon sperm DNA, 5 mM NaHPO4 (pH 6.5), 5mM EDTA, and 0.5% sodium dodecyl sulfate (SDS). Therespective probe was added to fresh buffer at a concentra-tion of 2 3 106 cpm ml-1, and hybridization was thenperformed at 60 6 3°C for 18 hours during gentle shaking.
After the hybridization, the membranes were washed in23 SSC and 0.1% SDS (2 3 5 minutes, room temperature)followed by digestion with 0.002% RNase A in 0.5 M NaCl,10 mM Tris, and 1 mM EDTA (30 minutes at 37°C).Subsequently, the membranes were heated to 70 6 3°C in0.13 SSC and 1% SDS for 30 minutes and finally themembranes were rinsed in 23 SSC. The dot blots werevisualized after phosphoscreen exposure (30 minutes) byusing a computer-assisted Phosphoimager (BAS 1000;Fujix, Japan).
Analysis
Sections processed for in situ hybridization were ana-lyzed under darkfield illumination. The localization oflabeled neurons was determined with the help of theadjacent Nissl-stained sections. Labeling density was de-termined by microscopic examination at a magnification of2503, with the assistance of an image analysis system(MCID 2; Imaging Research, Canada). The naming anddelineation of brain structures followed the atlas by Swan-son (1992).
RESULTS
Neurons expressing VP mRNA were identified by thepresence of an accumulation of silver grains in the emul-sion layer over the cell body. The autoradiographic labelingwas not uniform among the labeled neurons (e.g., compareFig. 3B,C,F). Instead, some neurons were very denselylabeled, whereas others showed weaker labeling. However,
due to a low background, VP mRNA-expressing neuronswere generally easily distinguishable.
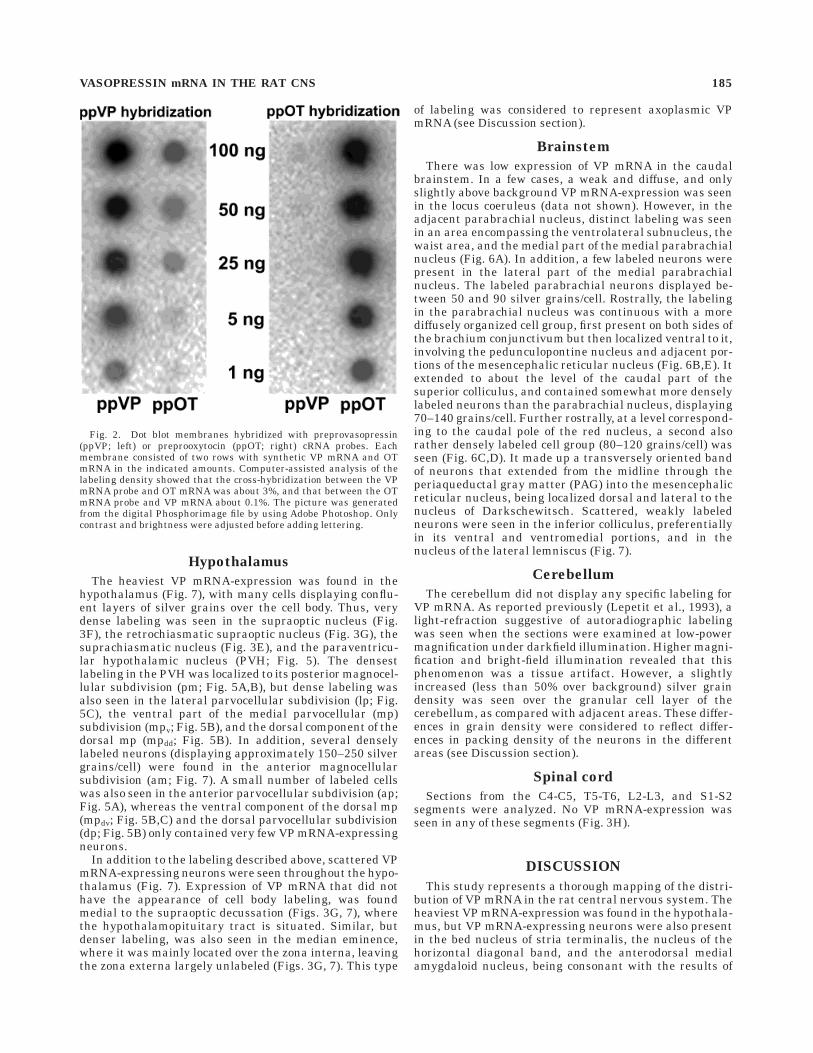
Several controls were made to assess the possibility ofartifactual labeling. Hybridization with a sense-strand VPmRNA probe showed no labeling (Fig. 1A). Pretreatmentwith RNase abolished or greatly attenuated all specificlabeling (Fig. 4C). Hybridization with an unrelated probe(against preproenkephalin mRNA) showed a completelydifferent pattern (data not shown). Because the 600-bp VPprobe used in the present study share sequence homologywith OT mRNA, the possibility of cross-reactivity betweenthe VP mRNA probe and OT mRNA was also examined.Hence, consecutive sections were hybridized with therespective probe. Although VP and OT mRNA-labelinggenerally were seen in the same hypothalamic regions, thelabeling patterns were different. Thus, some areas thatdisplayed strong labeling after hybridization for VP mRNA,contained few or no labeled cell after hybridization for OTmRNA, and vice versa (Fig. 1B,C). These observationswere consistent with the results of the dot blot hybridiza-tion, performed under stringency conditions similar tothose used in the in situ hybridizations (Fig. 2). Thus,quantitative analysis of the dot blots showed little cross-reactivity with OT mRNA; the binding of the vasopressinantisense probe to synthetic OT mRNA being about 3% ofthe binding to VP mRNA.
Forebrain
In the forebrain, VP mRNA-expressing neurons wereseen in the bed nucleus of the stria terminalis (Figs. 3A, 7),the nucleus of the horizontal diagonal band (Figs. 3B, 7),and the anterodorsal medial amygdaloid nucleus (Figs.3C, 7). Whereas the VP mRNA-positive neurons in theanterodorsal medial amygdaloid nucleus and the bednucleus of stria terminalis generally were heavily labeled(Fig. 3A,C), displaying about 150–300 and 200–400 silvergrains/cell,1 respectively, those in the nucleus of the hori-zontal diagonal band were more weakly labeled (Fig. 3B),displaying about 40–50 grains/cell. The background in allthese regions was ,3 grains over an adjacent cell body-sized area. Furthermore, VP mRNA-expressing neurons,displaying varying degrees of labeling density (60–150grains/cell), were seen in the entorhinal cortex (Figs. 3D,7). Single cells accompanied by weak diffuse labeling couldalso be followed ventrally into the piriform cortex.
VP mRNA-expression was also seen in the pyramidallayer of CA1–3 fields and the dentate gyrus throughout therostrocaudal extent of the hippocampus (Fig. 4). It waspresent both on X-ray film (Fig. 4D) and on emulsioncoated sections and was seen both under darkfield (Fig.4A,B) and brightfield (Fig. 4E) illumination, but was notpresent on adjacent RNase-treated sections, hybridizedsimultaneously (compare Fig. 4B with C). Grain countsover the labeled areas showed the labeling to be threetimes or more above background (as measured in adjacentunlabeled areas). Although diffuse, the labeling tended toappear as cell-sized aggregations of silver grains (Fig.4E8), suggestive of cell body labeling.
1 In heavily labeled cells, silver grains were confluent; an approximationof grain density was then obtained by dividing the labeled area by the grainsize. However, the figures obtained are likely to be underestimationsbecause grains were confluent also throughout the depth of the emulsion.
VASOPRESSIN mRNA IN THE RAT CNS 183

Fig. 1. Darkfield photomicrographs showing control hybridiza-tions. Accumulations of silver grains represent labeled neurons.A,A8: Labeling in the anterior paraventricular hypothalamic nucleus(PVH) after hybridization with a sense (A) and an antisense (A8)preprovasopressin cRNA probe. Note the absence of labeling aftersense probe hybridization. pm, posterior magnocellular subdivision;3v, third ventricle. B,B8: Adjacent sections through the middle part ofthe PVH hybridized for preprooxytocin mRNA (B) and preprovasopres-sin mRNA (B8). Note the differences, as highlighted by the arrow-heads, in expression in (i) the dorsolateral part of the posteriormagnocellular subdivision (pm), which was labeled with the preprova-sopressin (B8), but not with the preprooxytocin (B) cRNA probe; and (ii)
the dorsal parvocellular subdivision (dp), which expressed a muchlarger number of labeled neurons after hybridization with the pre-prooxytocin cRNA probe (B) than after hybridization with the prepro-vasopressin cRNA probe (B8). The same differences in the expressionpatterns of vasopressin and oxytocin have previously been shown inimmunohistochemical studies (e.g., Rhodes et al., 1981). 3v, thirdventricle. C,C8: Adjacent sections through the bed nucleus of striaterminalis hybridized with preprooxytocin (C) and preprovasopressin(C8) cRNA, respectively. Note the different labeling patterns obtainedwith the two probes. Capillaries are indicated (asterisks) for orienta-tion. Scale bar 5 70 µm in A,A8; 110 µm in B,B8; 140 µm in C,C8.
184 M. HALLBECK ET AL.
Hypothalamus
The heaviest VP mRNA-expression was found in thehypothalamus (Fig. 7), with many cells displaying conflu-ent layers of silver grains over the cell body. Thus, verydense labeling was seen in the supraoptic nucleus (Fig.3F), the retrochiasmatic supraoptic nucleus (Fig. 3G), thesuprachiasmatic nucleus (Fig. 3E), and the paraventricu-lar hypothalamic nucleus (PVH; Fig. 5). The densestlabeling in the PVH was localized to its posterior magnocel-lular subdivision (pm; Fig. 5A,B), but dense labeling wasalso seen in the lateral parvocellular subdivision (lp; Fig.5C), the ventral part of the medial parvocellular (mp)subdivision (mpv; Fig. 5B), and the dorsal component of thedorsal mp (mpdd; Fig. 5B). In addition, several denselylabeled neurons (displaying approximately 150–250 silvergrains/cell) were found in the anterior magnocellularsubdivision (am; Fig. 7). A small number of labeled cellswas also seen in the anterior parvocellular subdivision (ap;Fig. 5A), whereas the ventral component of the dorsal mp(mpdv; Fig. 5B,C) and the dorsal parvocellular subdivision(dp; Fig. 5B) only contained very few VP mRNA-expressingneurons.
In addition to the labeling described above, scattered VPmRNA-expressing neurons were seen throughout the hypo-thalamus (Fig. 7). Expression of VP mRNA that did nothave the appearance of cell body labeling, was foundmedial to the supraoptic decussation (Figs. 3G, 7), wherethe hypothalamopituitary tract is situated. Similar, butdenser labeling, was also seen in the median eminence,where it was mainly located over the zona interna, leavingthe zona externa largely unlabeled (Figs. 3G, 7). This type
of labeling was considered to represent axoplasmic VPmRNA (see Discussion section).
Brainstem
There was low expression of VP mRNA in the caudalbrainstem. In a few cases, a weak and diffuse, and onlyslightly above background VP mRNA-expression was seenin the locus coeruleus (data not shown). However, in theadjacent parabrachial nucleus, distinct labeling was seenin an area encompassing the ventrolateral subnucleus, thewaist area, and the medial part of the medial parabrachialnucleus (Fig. 6A). In addition, a few labeled neurons werepresent in the lateral part of the medial parabrachialnucleus. The labeled parabrachial neurons displayed be-tween 50 and 90 silver grains/cell. Rostrally, the labelingin the parabrachial nucleus was continuous with a morediffusely organized cell group, first present on both sides ofthe brachium conjunctivum but then localized ventral to it,involving the pedunculopontine nucleus and adjacent por-tions of the mesencephalic reticular nucleus (Fig. 6B,E). Itextended to about the level of the caudal part of thesuperior colliculus, and contained somewhat more denselylabeled neurons than the parabrachial nucleus, displaying70–140 grains/cell. Further rostrally, at a level correspond-ing to the caudal pole of the red nucleus, a second alsorather densely labeled cell group (80–120 grains/cell) wasseen (Fig. 6C,D). It made up a transversely oriented bandof neurons that extended from the midline through theperiaqueductal gray matter (PAG) into the mesencephalicreticular nucleus, being localized dorsal and lateral to thenucleus of Darkschewitsch. Scattered, weakly labeledneurons were seen in the inferior colliculus, preferentiallyin its ventral and ventromedial portions, and in thenucleus of the lateral lemniscus (Fig. 7).
Cerebellum
The cerebellum did not display any specific labeling forVP mRNA. As reported previously (Lepetit et al., 1993), alight-refraction suggestive of autoradiographic labelingwas seen when the sections were examined at low-powermagnification under darkfield illumination. Higher magni-fication and bright-field illumination revealed that thisphenomenon was a tissue artifact. However, a slightlyincreased (less than 50% over background) silver graindensity was seen over the granular cell layer of thecerebellum, as compared with adjacent areas. These differ-ences in grain density were considered to reflect differ-ences in packing density of the neurons in the differentareas (see Discussion section).
Spinal cord
Sections from the C4-C5, T5-T6, L2-L3, and S1-S2segments were analyzed. No VP mRNA-expression wasseen in any of these segments (Fig. 3H).
DISCUSSION
This study represents a thorough mapping of the distri-bution of VP mRNA in the rat central nervous system. Theheaviest VP mRNA-expression was found in the hypothala-mus, but VP mRNA-expressing neurons were also presentin the bed nucleus of stria terminalis, the nucleus of thehorizontal diagonal band, and the anterodorsal medialamygdaloid nucleus, being consonant with the results of
Fig. 2. Dot blot membranes hybridized with preprovasopressin(ppVP; left) or preprooxytocin (ppOT; right) cRNA probes. Eachmembrane consisted of two rows with synthetic VP mRNA and OTmRNA in the indicated amounts. Computer-assisted analysis of thelabeling density showed that the cross-hybridization between the VPmRNA probe and OT mRNA was about 3%, and that between the OTmRNA probe and VP mRNA about 0.1%. The picture was generatedfrom the digital Phosphorimage file by using Adobe Photoshop. Onlycontrast and brightness were adjusted before adding lettering.
VASOPRESSIN mRNA IN THE RAT CNS 185
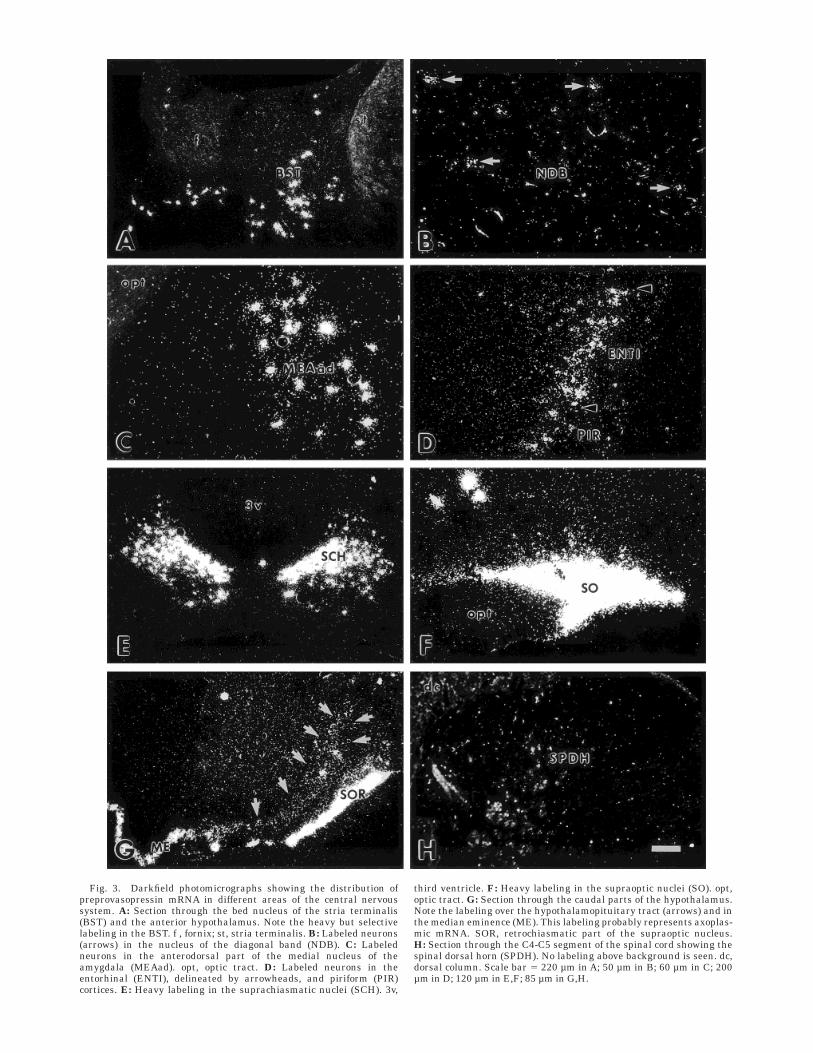
Fig. 3. Darkfield photomicrographs showing the distribution ofpreprovasopressin mRNA in different areas of the central nervoussystem. A: Section through the bed nucleus of the stria terminalis(BST) and the anterior hypothalamus. Note the heavy but selectivelabeling in the BST. f , fornix; st, stria terminalis. B: Labeled neurons(arrows) in the nucleus of the diagonal band (NDB). C: Labeledneurons in the anterodorsal part of the medial nucleus of theamygdala (MEAad). opt, optic tract. D: Labeled neurons in theentorhinal (ENTI), delineated by arrowheads, and piriform (PIR)cortices. E: Heavy labeling in the suprachiasmatic nuclei (SCH). 3v,
third ventricle. F: Heavy labeling in the supraoptic nuclei (SO). opt,optic tract. G: Section through the caudal parts of the hypothalamus.Note the labeling over the hypothalamopituitary tract (arrows) and inthe median eminence (ME). This labeling probably represents axoplas-mic mRNA. SOR, retrochiasmatic part of the supraoptic nucleus.H: Section through the C4-C5 segment of the spinal cord showing thespinal dorsal horn (SPDH). No labeling above background is seen. dc,dorsal column. Scale bar 5 220 µm in A; 50 µm in B; 60 µm in C; 200µm in D; 120 µm in E,F; 85 µm in G,H.
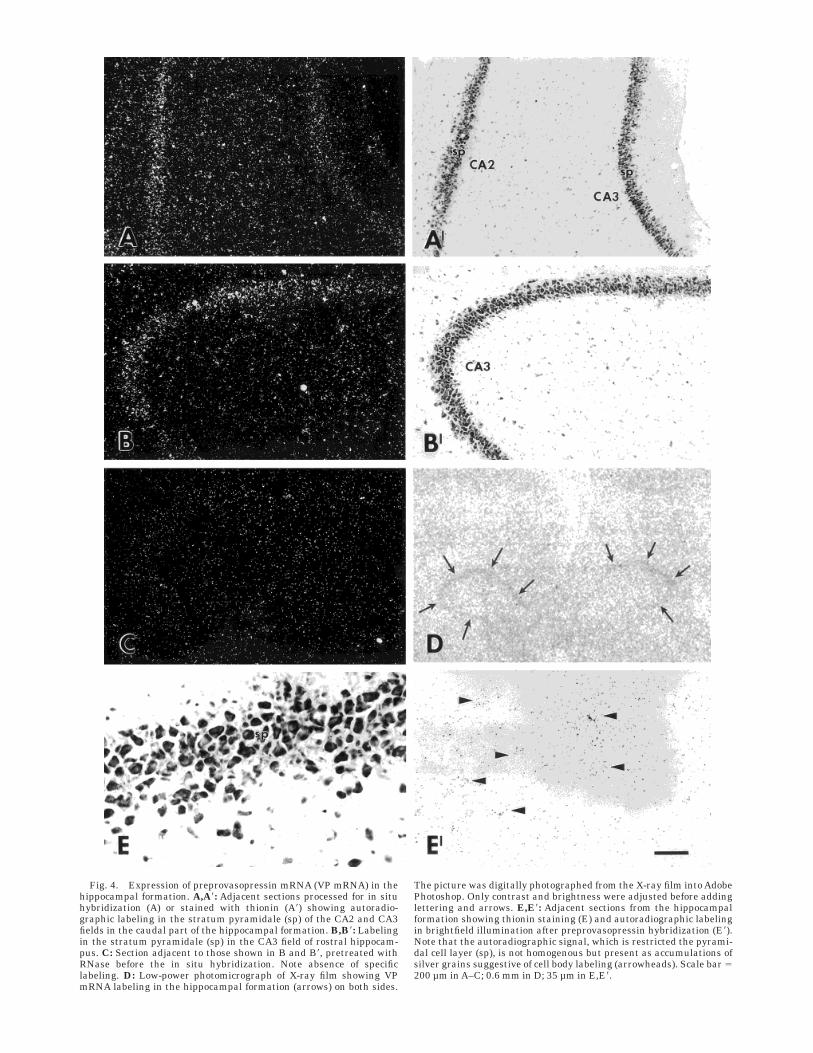
Fig. 4. Expression of preprovasopressin mRNA (VP mRNA) in thehippocampal formation. A,A8: Adjacent sections processed for in situhybridization (A) or stained with thionin (A8) showing autoradio-graphic labeling in the stratum pyramidale (sp) of the CA2 and CA3fields in the caudal part of the hippocampal formation. B,B8: Labelingin the stratum pyramidale (sp) in the CA3 field of rostral hippocam-pus. C: Section adjacent to those shown in B and B8, pretreated withRNase before the in situ hybridization. Note absence of specificlabeling. D: Low-power photomicrograph of X-ray film showing VPmRNA labeling in the hippocampal formation (arrows) on both sides.
The picture was digitally photographed from the X-ray film into AdobePhotoshop. Only contrast and brightness were adjusted before addinglettering and arrows. E,E8: Adjacent sections from the hippocampalformation showing thionin staining (E) and autoradiographic labelingin brightfield illumination after preprovasopressin hybridization (E8).Note that the autoradiographic signal, which is restricted the pyrami-dal cell layer (sp), is not homogenous but present as accumulations ofsilver grains suggestive of cell body labeling (arrowheads). Scale bar 5200 µm in A–C; 0.6 mm in D; 35 µm in E,E8.

previous studies (see Introductory remarks for references).Novel populations of VP mRNA-expressing neurons weredetected in the hippocampus, the piriform cortex, theparabrachial nucleus, the dorsal pontine and midbraintegmentum, and the PAG, and scattered neurons wereseen in the nucleus of the lateral lemniscus and the
inferior colliculus. Expression of VP mRNA was not de-tected in the spinal cord, the cerebellum or the nucleussubcoeruleus, and was very sparse in the locus coeruleus,regions that in previous studies have been reported tocontain large numbers of cell bodies immunoreactive forvasopressin (Caffe and van Leeuwen, 1983; Caffe et al.,
Fig. 5. The distribution of preprovasopressin mRNA-positive neu-rons in the paraventricular hypothalamic nucleus (PVH). A–C: Dark-field photomicrographs of frontal sections throughout the hypothala-mus, separated by 240 µm. A is rostral. A8–C8: Adjacent Nissl-stainedsections, showing the different subdivisions of PVH, as described bySwanson and Sawchenko (1983). To assist orientation, capillaries areindicated by arrowheads. Note the difference in preprovasopressinmRNA labeling between the various parvocellular subdivisions of
PVH. ap, anterior parvocellular subdivision; dp, dorsal parvocellularsubdivision; lp, lateral parvocellular subdivision; mpdd, dorsal part thedorsal medial parvocellular subdivision; mpdv, ventral part of thedorsal medial parvocellular subdivision; mpv, ventral medial parvocel-lular subdivision; pm, posterior magnocellular subdivision. Bordersbetween the subdivisions are indicated by dashed lines. Scale bar 5100 µm in C8 (applies to A–C8).
188 M. HALLBECK ET AL.
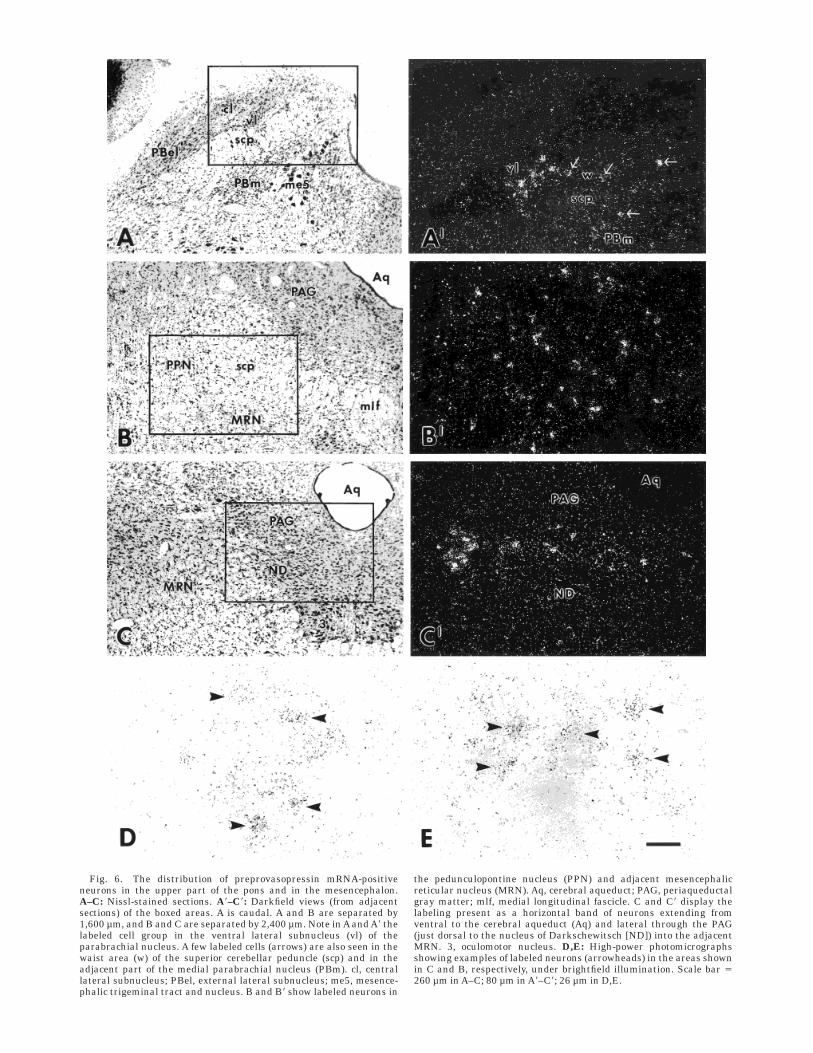
Fig. 6. The distribution of preprovasopressin mRNA-positiveneurons in the upper part of the pons and in the mesencephalon.A–C: Nissl-stained sections. A8–C8: Darkfield views (from adjacentsections) of the boxed areas. A is caudal. A and B are separated by1,600 µm, and B and C are separated by 2,400 µm. Note in A and A8 thelabeled cell group in the ventral lateral subnucleus (vl) of theparabrachial nucleus. A few labeled cells (arrows) are also seen in thewaist area (w) of the superior cerebellar peduncle (scp) and in theadjacent part of the medial parabrachial nucleus (PBm). cl, centrallateral subnucleus; PBel, external lateral subnucleus; me5, mesence-phalic trigeminal tract and nucleus. B and B8 show labeled neurons in
the pedunculopontine nucleus (PPN) and adjacent mesencephalicreticular nucleus (MRN). Aq, cerebral aqueduct; PAG, periaqueductalgray matter; mlf, medial longitudinal fascicle. C and C8 display thelabeling present as a horizontal band of neurons extending fromventral to the cerebral aqueduct (Aq) and lateral through the PAG(just dorsal to the nucleus of Darkschewitsch [ND]) into the adjacentMRN. 3, oculomotor nucleus. D,E: High-power photomicrographsshowing examples of labeled neurons (arrowheads) in the areas shownin C and B, respectively, under brightfield illumination. Scale bar 5260 µm in A–C; 80 µm in A8–C8; 26 µm in D,E.

1985; De Vries et al., 1985; Kai-Kai et al., 1986; Lepetit etal., 1993).
Technical considerations
It should be pointed out that the distribution of VPmRNA demonstrated in this study is that of naive, maleSprague-Dawley rats. Several reports have shown that thevasopressin synthesis is influenced by humoral and otherfactors. For example, it has been demonstrated that thevasopressinergic projections from the bed nucleus of striaterminalis and the anterodorsal medial amygdaloid nucleusare dependent on gonadal steroids for their normal appear-ance (De Vries and Buijs, 1983; De Vries et al., 1985) andthat stress increases the number of vasopressin mRNA-
expressing neurons in the parvocellular PVH (Sawchenkoet al., 1984a; Bartanusz et al., 1994). Accordingly, it ispossible that the very low expression seen in the locuscoeruleus in the present study represents a basal transcrip-tion level that could increase significantly during stressfulconditions. These considerations are also of interest whencomparing the findings of the present study with those ofearlier immunohistochemical studies on the distributionof vasopressinergic neurons in the central nervous system.Such studies in animals treated with the neurotoxincolchicine to inhibit axonal transport for the enhancementof cell body labeling have reported the presence of largenumbers of vasopressin-immunoreactive neurons in thelocus coeruleus, subcoeruleus, and spinal cord (Caffe and
Fig. 7. Drawings of frontal sections through the upper brainstemand basal forebrain, based on the atlas by Swanson (1992), showingthe distribution of preprovasopressin mRNA labeling. Numbers referto the position of the levels, as shown in the figurine of the rat brain.Large dots represent distinctly labeled single neurons; black areasrepresent areas with densely packed, heavily labeled neurons; areaswith small dots represent weak, probably cellular, labeling in areaswith densely packed neurons; and areas with hatching representsaxoplasmic preprovasopressin mRNA. Abbreviations follow Swanson(1992). aco, anterior commissure, olfactory limb; act, anterior commis-sure, temporal limb; ADP, anterodorsal preoptic nucleus; AHA, ante-rior hypothalamic area; AHN, anterior hypothalamic nucleus; AHNa,AHN, anterior part; AMd, anteromedial nucleus thalamus, dorsalpart; APN, anterior pretectal nucleus; AQ, cerebral aqueduct; ARH,arcuate nucleus hypothalamus; AV, anteroventral nucleus thalamus;AVP, anteroventral preoptic nucleus; BLAa, basolateral nucleus amyg-dala, anterior part; BMAa, basomedial nucleus amygdala, anteriorpart; BST, bed nucleus of the stria terminalis; BSTad, BST, anteriordivision, anterodorsal area; BSTal, BST, anterior division, anterolat-eral area; BSTdl, BST, anterior division, dorsolateral nucleus; BSTif,BST, posterior division, interfascicular nucleus; BSTpr, BST, posteriordivision, principal nucleus; BSTrh, BST, anterior division, rhomboidnucleus; BSTtr, BST, posterior division, transverse nucleus; BSTv,BST, anterior division, ventral nucleus; CA1, Ammon’s horn, fieldCA1; CA1slm, CA1, stratum lacunosum-moleculare; CA1so, CA1,stratum oriens; CA1sp, CA1, pyramidal layer; CA1spd, CA1, pyrami-dal layer, deep; CA1sps, CA1, pyramidal layer, superficial; CA1sr,CA1, stratum radiatum; CA2, Ammon’s horn, field CA2; CA2sp, CA2,pyramidal layer; CA3, Ammon’s horn, field CA3; CA3slu, CA3, stra-tum lucidum; CA3so, CA3, stratum oriens; CA3sp, CA3, pyramidallayer; CA3sr, CA3, stratum radiatum; cc, corpus callosum; CEAc,central nucleus amygdala, capsular part; CEAm, central nucleusamygdala, medial part; CENT2, central lobule, lobule II; CENT3,central lobule, lobule III; cic, commissure of the inferior colliculus;cing, cingulum bundle; CL, central lateral nucleus thalamus; CLI,central linear nucleus raphe; CM, central medial nucleus thalamus;CP, caudoputamen; cpd, cerebral peduncle; CSl, superior centralnucleus raphe, lateral part; CSm, superior central nucleus raphe,medial part; cst, corticospinal tract; CUN, cuneiform nucleus; DGlb,dentate gyrus, lateral blade; DGlb-mo, DGlb, molecular layer; DGlb-sg, DG-lb, granule cell layer; DGmb, dentate gyrus, medial blade;DGmb-sg, DGmb, granule cell layer; DMH, dorsomedial nucleushypothalamus; DR, dorsal nucleus raphe; dscp, decussation of thesuperior cerebellar peduncle; ENTl, entorhinal area, lateral part; EPd,endopiriform nucleus, dorsal part; fi, fimbria; FL, flocculus; fx, col-umns of the fornix; GPl, globus pallidus, lateral segment; GPm, globuspallidus, medial segment; IA, intercalated nuclei amygdala; ICc,inferior colliculus, central nucleus; ICd, inferior colliculus, dorsalnucleus; ICe, inferior colliculus, external nucleus; III, oculomotornucleus; INC, interstitial nucleus of Cajal; INFin, infundibulum,internal lamina; int, internal capsule; IPNi, interpeduncular nucleus,intermediate subnucleus; isl, islands of Calleja; LA, lateral nucleusamygdala; LC, locus coeruleus; LD, lateral dorsal nucleus thalamus;LDT, laterodorsal tegmental nucleus; LGd, lateral geniculate complex,dorsal part; LH, lateral habenula; LHA, lateral hypothalamic area; ll,lateral lemniscus; LP, lateral posterior nucleus thalamus; LPO, lateralpreoptic nucleus; LSd, lateral septal nucleus, dorsal part; LSi, lateral
septal nucleus, intermediate part; LSv, lateral septal nucleus, ventralpart; LT, lateral terminal nucleus accessory optic tract; MA, magnocel-lular preoptic nucleus; mcp, middle cerebellar peduncle; MD, mediodor-sal nucleus thalamus; MDc, MD, central part; MEAad, medial nucleusof the amygdala, anterodorsal part; MEPO, median preoptic nucleus;MGd, medial geniculate complex, dorsal part; MGv, medial geniculatecomplex, ventral part; MH, medial habenula; ml, medial lemniscus;mlf, medial longitudinal fasciculus; mp, mammillary peduncle; MPO,medial preoptic area; MPT, medial pretectal area; MRN, mesence-phalic reticular nucleus; MS, medial septal nucleus; NB, nucleusbrachium inferior colliculus; ND, nucleus of Darkschewitsch; NDB,nucleus of the diagonal band; NLL, nucleus of the lateral lemniscus;NOT, nucleus of the optic tract; NTB, nucleus of the trapezoid body;och, optic chiasm; OP, olivary pretectal nucleus; opt, optic tract; PAG,periaqueductal gray matter; PB, parabrachial nucleus; PBG, parabi-geminal nucleus; PBlc, PB, central lateral part; PBle, PB, externallateral part; PBlv, PB, ventral lateral part; PBme, PB, external medialpart; PBmm, PB, medial medial part; PCG, pontine central gray; PG,pontine gray; PH, posterior hypothalamic nucleus; PIR, piriform area;PO, posterior complex thalamus; POR, periolivary nucleus; PPN,pedunculopontine nucleus; PRNc, pontine reticular nucleus, caudalpart; PRNr, pontine reticular nucleus, rostral part; PS, parastrialnucleus; PSV, principal sensory nucleus of the trigeminal nerve; PT,parataenial nucleus; PV, periventricular nucleus; PVH, paraventricu-lar hypothalamic nucleus; PVHam, PVH, anterior magnocellular part;PVHap, PVH, anterior parvicellular part; PVHdp, dorsal parvicellularpart; PVHf, PVH, forniceal part; PVHlp, PVH, lateral parvicellularpart; PVHmpd, PVH, medial parvicellular part, dorsal zone; PVH-pmm, PVH, posterior magnocellular part, medial zone; PVp, posteriorperiventricular nucleus hypothalamus; PVpo, preoptic periventricularnucleus hypothalamus; PVT, paraventricular nucleus thalamus; py,pyramidal tract; RE, nucleus reuniens; RH, rhomboid nucleus; RL,rostral linear nucleus raphe; RN, red nucleus; RR, mesencephalicreticular nucleus, retrorubal area; RT, reticular nucleus thalamus;rust, rubrospinal tract; SAG, nucleus sagulum; SC, superior colliculus;SCdg, SC, deep gray layer; SCH, suprachiasmatic nucleus; SCig, SC,intermediate gray layer; scp, superior cerebellar peduncle; SCsg, SC,superficial gray layer; SF, septofimbrial nucleus; SFO, subfornicalorgan; SH, septohippocampal nucleus; SI, substantia innominata;SIM, simple lobule; SLD, sublaterodorsal nucleus; sm, stria medul-laris; SNc, substantia nigra, compact part; SNr, substantia nigra,reticular part; SO, supraoptic nucleus; SOCl, superior olivary com-plex, lateral part; SOr, SO, retrochiasmatic part; SPFp, subparafascicu-lar nucleus thalamus, parvicellular part; sptV, spinal tract of thetrigeminal nerve; STN, subthalamic nucleus; SUBd, subiculum, dor-sal part; SUT, supratrigeminal nucleus; TRN, tegmental reticularnucleus, pontine grey; TRS, triangular nucleus septum; V, motornucleus of the trigeminal nerve; V3, third ventricle; V4, fourthventricle; VAL, ventral anterior-lateral complex thalamus; VCOa,ventral cochlear nucleus, anterior part; VIIn, facial nerve; VIP, veluminterpositum; VL, lateral ventricle; VM, ventral medial nucleusthalamus; VMH, ventromedial nucleus hypothalamus; Vn, trigeminalnerve; VPL, ventral posterolateral nucleus thalamus; VPM, ventralposteromedial nucleus thalamus; VTA, ventral tegmental area; vtd,ventral tegmental decussation; VTN, ventral tegmental nucleus; ZI,zona incerta.
190 M. HALLBECK ET AL.

Figure 7

Figure 7 (Continued)

Figure 7 (Continued)
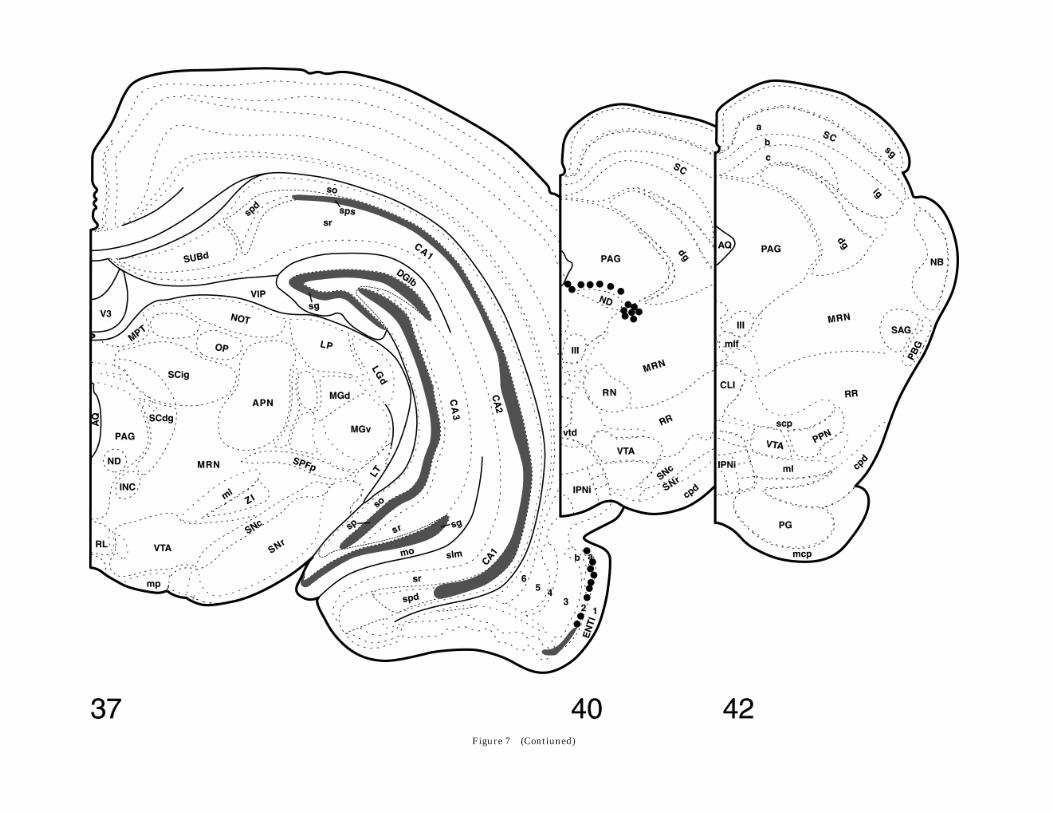
Figure 7 (Contiuned)

Figure 7 (Continued)

van Leeuwen, 1983; Caffe et al., 1985; De Vries et al., 1985;Kai-Kai et al., 1986). It is possible that the use of colchicinein those studies may have induced a vasopressin synthesisthat does not occur normally, because colchicine has beenshown to induce gene transcription that does not occur innaive animals (Ceccatelli et al., 1991; Rethelyi et al.,1991).
The possibility that the technique used in the presentstudy did not permit detection of some populations of VPmRNA-expressing neurons should also be considered. How-ever, the in situ hybridization technique used here is verysensitive and will readily detect small amounts of mRNA(Simmons et al., 1989). Accordingly, we have previouslydetected a novel enkephalinergic cell group in the thala-mus by using the same technique (Hermanson et al.,1995), and in the present study, we found previously notdescribed populations of vasopressinergic neurons in thehippocampus, entorhinal and piriform cortices, and brain-stem. These observations suggest that the unlabeled nervecell groups do not synthesize significant amounts of VPmRNA in the normal Sprague-Dawley rat. However, theabsolute sensitivity of the method used here is not known(Angerer et al., 1987), and it is possible that other tech-niques such as in situ polymerase chain reaction willreveal low-expressing VP mRNA-neurons not detected inthe present study.
Although the use of full-length riboprobes for in situhybridization secures high sensitivity, it carries the riskfor cross-hybridization with mRNAs containing homolo-gous sequences. In the present experiments, the possibilityof cross-hybridization between the VP mRNA probe andOT mRNA must be considered. This possibility was exam-ined both by comparing the labeling pattern in consecutivesections processed for VP and OT mRNA, respectively, andby using dot blot hybridization. Although the comparisonof the labeling in consecutive tissue sections was compli-cated by the fact the VP and OT mRNAs largely areexpressed in the same nuclei, this comparison neverthe-less revealed distinct differences in the distribution ofneurons labeled with the two probes. For example, in thedorsal parvocellular subdivision of PVH (Fig. 1B) and inthe bed nucleus of stria terminalis (Fig. 1C), the labelingpattern for VP mRNA was clearly different from that forOT mRNA. Thus, with the exception for a few VP mRNA-expressing neurons that were strongly and distinctlylabeled, the dorsal parvocellular region of the PVH did notdisplay labeling over background (Fig. 1B8), whereas theconsecutive section hybridized for OT mRNA showeddensely packed, heavily labeled OT mRNA-expressingneurons (Fig. 1B). Any significant cross-reactivity betweenthe vasopressin probe and OT mRNA should have resultedin increased background staining in the vasopressin-hybridized section. Similarly, the OT mRNA probe did notappear to cross-react with VP mRNA, because areas thatdisplayed dense and strong VP mRNA-expression (thedorsal part of the magnocellular division of the PVH [Fig.1B8] and the bed nucleus of stria terminalis [Fig. 1C8]) didnot show any significant labeling in the consecutive oxyto-cin-hybridized section (Fig. 1B,C, respectively). The lack ofsignificant cross-reactivity was confirmed by the dot-blothybridization (Fig. 2) that was performed during similarconditions as the in situ hybridization. The very weakbinding of the vasopressin-probe to OT mRNA reveled bythe dot-blot seems to exclude that OT mRNA-expressingcells would display any significant labeling in the vasopres-
sin-hybridized sections, being consonant with the observa-tions from consecutive sections. However, an unexpectedfinding that could be interpreted as evidence of cross-hybridization was the presence of labeled neurons in theanterior magnocellular PVH (am; also named the anteriorcommissural nucleus) after VP mRNA hybridization. Theam is generally considered to be an oxytocinergic cellgroup (Swanson and Sawchenko, 1983; Burbach et al.,1987; Chung et al., 1991), and although some immunohis-tochemical studies have found VP-expressing neurons inam (Sawchenko and Swanson, 1982; Caffe and van Leeu-wen 1983; van Leeuwen and Caffe, 1983; Sawchenko et al.,1984b), these observations previously have not been sup-ported by in situ hybridization histochemistry (Burbach etal., 1987; Chung et al., 1991). The am cells that expressedVP mRNA were heavily labeled. It is unlikely that suchdense labeling would result from cross-hybridization,which, if present, rather would have produced a muchfainter labeling.
Novel vasopressin-expressing regions
Hippocampus. Labeling was seen in the pyramidallayer of the hippocampus throughout the CA1–3 fields andthe dentate gyrus. This labeling was present both on X-rayfilm (Fig. 4D) and on emulsion-coated sections (Fig.4A,B,E8), and it was abolished after pretreatment of thesections with RNase (Fig. 4C). Thus, it was not due toartifactual autoradiographic labeling, nor a result of unspe-cific binding of the VP mRNA probe, but most likelyrepresents specific VP mRNA expression. However, it isimportant to point out that the high packing density of thepyramidal cells will concentrate any signal originatingfrom these cells onto a small area, permitting the detectionof a low mRNA expression that may not have beenvisualized in less cell dense areas. Accordingly, to checkthat the increased expression seen in the pyramidal celllayer of the hippocampus was not a general phenomenonfor areas consisting of tightly packed small cells, we alsoexamined the expression over the granular cell layer of thecerebellum. This cell layer displayed a somewhat higherhybridization signal than adjacent cell sparse areas, butthe difference was clearly far from that seen in thehippocampus. Furthermore, whereas the hybridizationsignal over the granular cell layer in the cerebellum wasevenly dispersed, the labeling seen over the pyramidal celllayer of the hippocampus was characterized by the pres-ence of aggregations suggestive of cell body labeling.
The possible function of vasopressin in the hippocampusis not known, but vasopressin binding in this structure hasbeen demonstrated (Brinton et al., 1984). Vasopressin hasbeen shown to activate hippocampal neurons (e.g., Ver-steeg et al., 1984; Brinton and McEwen, 1989; Giri et al.,1990; Maegawa et al., 1992), an observation that is conso-nant with the involvement of vasopressin in memory andlearning (Bohus, 1977; Leshner and Roche, 1977; Bohus etal., 1978; de Wied and Versteeg, 1979). Thus, it has beenshown that rats bred to perform better in memory testshave increased hippocampal vasopressin levels (Hess etal., 1992) and that changes in the intrahippocampalvasopressin have effects on memory retrieval and learning(Metzger et al., 1993). Another proposed function of vaso-pressin in the hippocampus concerns the inhibition ofcentrally induced pressor responses (Versteeg et al., 1984),which has been suggested to be mediated by a vasopressin-ergic projection from the amygdala to the hippocampus
196 M. HALLBECK ET AL.

(Caffe et al., 1987; Smock et al., 1992). The present findingindicates that many of the vasopressin effects in thehippocampus could be elicited by intrinsic vasopressiner-gic neurons.
Entorhinal and piriform cortices. A novel, ratherdense VP mRNA-expression was seen in the entorhinaland piriform cortices. Although vasopressin neurons havenot previously been described here, vasopressin V1 recep-tors have been demonstrated (Ostrowski et al., 1994), andperipheral vasopressin-administration has been shown toresult in immediate-early gene expression (Wu et al.,1995). Because of its close association with the hippocam-pus, the vasopressin neurons in the entorhinal cortex mayaccount for several of the vasopressin effects ascribed tothe former structure (see above). The piriform cortex isalso a component in the corticohippocampal pathway andhas been shown to be involved in learning and memory(e.g., Granger et al., 1996); thus, the vasopressin neuronsin the piriform cortex could be of importance for suchmechanisms, similar to the vasopressin neurons in theentorhinal cortex and hippocampus. Furthermore, in addi-tion to its role for olfaction, the piriform cortex has beenassociated with sexual behavior, and vasopressin is knownto influence such behavior (Sodersten et al., 1983; Meyer-son et al., 1988), pointing to another possible mechanismfor vasopressin in this part of the brain.
Parabrachial nucleus. The labeling in the parabra-chial nucleus encompassed preferentially the ventral lat-eral subnucleus, but involved also the dorsomedial part ofthe medial parabrachial nucleus and the waist area inter-connecting these two regions (see Moga et al., 1990, fornomenclature). These regions of the parabrachial nucleushave similar efferent projection patterns, providing outputto the insular cortex, the central nucleus of the amygdala,the substantia innominata, and the zona incerta (e.g.,Fulwiler and Saper, 1984). The ascending input originatesprimarily from the nongustatory, general-visceral regionsof the nucleus of the solitary tract (Herbert et al., 1990).Thus, the anatomic data suggest that vasopressin-express-ing parts of the parabrachial nucleus are involved in theprocessing of some aspect(s) of viscerosensory information.Of particular interest is the recent demonstration by Kellyand Watts (1998) that corticotropin-releasing hormone(CRH) mRNA-expressing neurons in the lateral hypotha-lamic area that are activated by cellular dehydrationproject densely to the parts of the parabrachial nucleus,shown in the present study to express VP mRNA. Theseauthors suggest that the CRH pathways to the parabra-chial nucleus may be involved in the generation of dehydra-tion-induced anorexia, as a component of a homeostaticresponse to dehydration aimed at protecting the animal’sfluid compartment (Kelly and Watts, 1998). Many of theeffects of vasopressin are related to the maintenance ofwater balance, and vasopressin is also known to be anorec-tic (Langhans et al., 1991); accordingly, the vasopressinneurons in the parabrachial nucleus could be the target ofthe CRH fibers.
Other brainstem loci. The VP mRNA-expressing cellgroup in the parabrachial nucleus could be followed ros-trally into the peripeduncular nucleus. However, whetherthe vasopressinergic neurons in the latter structure arefunctionally related to those in the parabrachial nucleusremains to be clarified. In addition, a second, topographi-cally distinct VP mRNA-expressing cell group was seen inthe brainstem at midcollicular level. It involved a thin
sector of the PAG immediately dorsal to the nucleus ofDarkschewitsch, and the adjacent portion of the midbrainreticular formation. Despite its homogenous appearance,the PAG encompasses a large number of functionallydistinct cell groups that regulate a variety of complexbehaviors that include behavioral adaptive autonomicregulations. At present, we can only speculate about therole of the vasopressinergic neurons in the rostral mid-brain; for example, they could be involved in some aspectsof vasomotor control (Carrive, 1991).
Scattered VP mRNA-expressing neurons were seen inthe nucleus of the lateral lemniscus and the inferiorcolliculus. Their functional role in the rat is not known;however, in some other species vasopressin has beenshown to be involved in sexual-related auditory mecha-nisms (Dubois-Dauphin et al., 1987; Penna et al., 1992).
Axoplasmic VP mRNA. An interesting observationwas the labeling seen over the hypothalamopituitary tractand the median eminence. This labeling was not localizedto cell bodies but had the appearance of fiber labeling.Thus, it probably represented axoplasmic VP mRNA trans-ported from the magnocellular vasopressin groups to thepituitary. Axoplasmic mRNA for vasopressin was firstdetected in the posterior pituitary lobe. This mRNA wasinitially believed to originate from the pituicytes (Murphyet al., 1989; Lehmann et al., 1990; McCabe et al., 1990),but it was later shown that interruption of the hypothal-amopituitary tract abolished pituitary expression of VPmRNA (Mohr et al., 1990), suggesting an axonal localiza-tion. It has also been show that VP mRNA is present in theneural stalk and in the zona interna of the medianeminence in salt-loaded animals and that the pituitary islacking primary vasopressin transcripts (Mohr et al.,1991; but see also Pu et al., 1995). In addition, OT mRNAhas been shown in the pituitary (Mohr et al., 1990), andelectron microscopic studies have demonstrated the pres-ence of OT and VP mRNA in axons of the hypothalamopitu-itary tract (Jirikowski et al., 1990; Trembleau et al., 1994).However, to our knowledge, the present study is the first todemonstrate axoplasmic VP mRNA outside the posteriorpituitary and median eminence in naive animals.
The function of axoplasmic mRNA is still not known (seevan Minnen, 1994, for a review). It has been proposed thataxoplasmic mRNA could be a rapid way of regulating themRNA content in the perikaryon (Jirikowski et al., 1990),thus controlling peptide production. This idea is supportedby the fact that mRNA injected into the pituitary istransported back to the hypothalamus where it is tran-scribed (Jirikowski et al., 1990; Maciejewski-Lenoir et al.,1993). However, the axon storage hypothesis is contra-dicted by the fact that VP mRNA increases in the axoncompartment during salt loading (Murphy et al., 1989;McCabe et al., 1990; Trembleau et al., 1994, 1996), whenthere is an increased demand for vasopressin production.It has also been demonstrated that squid axons containactive polysomes (Giuditta et al., 1991), and de novosynthesis of neurofilament proteins in the axonal compart-ment of this species has been demonstrated (Crispino etal., 1993). De novo protein synthesis in axons has also beenproposed in other species, including rats (Koenig, 1991).Another function of axonal mRNA is suggested by therecent finding that preprotachykinin-A mRNA is releasedfrom C-fibers upon stimulation (Wang et al., 1996), raisingthe possibility of a role after secretion.
VASOPRESSIN mRNA IN THE RAT CNS 197

Efferent targets of vasopressinergiccell groups
Vasopressinergic fibers constitute an extensive networkin the rat brain. This study shows that a rather smallnumber of cell groups can produce vasopressin, at least inthe naive rat, and, accordingly, these cell groups must havevery widespread projections. However, no complete picturehas yet emerged on the origin of the vasopressin fibers indifferent brain regions, although there are several reportson the organization of the vasopressinergic projections.One important source for vasopressin fibers in the brain isthe bed nucleus of the stria terminalis, which projects to alarge number of regions, including the olfactory tubercle,the nucleus of the horizontal diagonal band, the lateralseptum, the anterior amygdaloid area, the lateral habenu-lar nucleus, the periventricular gray matter, the dorsalraphe nucleus, and the locus coeruleus (De Vries andBuijs, 1983). The suprachiasmatic nucleus is anotherimportant source for vasopressin fibers in the forebrainand has been reported to give rise to vasopressinergicprojections to the organum vasculosum lamina terminalis,the periventricular hypothalamic nucleus, and the dorsalpart of the paraventricular thalamic nucleus (Hoornemanand Buijs, 1982). The medial amygdaloid nucleus has beenshown to have vasopressinergic projections to the lateralseptum and the ventral hippocampus, but not to themedulla oblongata or the spinal cord (Caffe et al., 1987;Smock et al., 1992). The supraoptic nucleus, including itsretrochiasmatic part, has been shown to project mainly tothe neural lobe (e.g., Swanson and Sawchenko, 1983) buthas also been proposed to have collaterals to adjacentareas of the lateral hypothalamus (Mason et al., 1984). ThePVH sends vasopressinergic fibers to the external layer ofthe median eminence (Dierickx et al., 1976), the dorsalvagal complex, and the spinal cord (Swanson and Kuypers,1980; Sawchenko and Swanson, 1982), in addition to itsprojection to the neural lobe. No obvious change in thevasopressin innervation of the forebrain has been foundafter lesions of the PVH (De Vries and Buijs, 1983).
Interestingly, no substantial VP mRNA-expression wasseen in the locus coeruleus and subcoeruleus in thepresent study. This is contradictory to the findings inseveral immunohistochemical investigations (Caffe andvan Leeuwen, 1983; Caffe et al., 1985; De Vries et al., 1985;Sofroniew, 1985) but corroborates the result of a previousin situ hybridization study (Urban et al., 1990). It is alsoconsistent with earlier studies showing that lesions of thelocus coeruleus and subcoeruleus did not affect vasopres-sin innervation of the brain (Caffe et al., 1988), whereaslesions of the PVH abolished almost all vasopressin inbrainstem autonomic nuclei (De Vries and Buijs, 1983) andspinal cord (Lang et al., 1983). We have previously shownthat dorsal root ganglion cells in naive Sprague-Dawleyrats do not express VP mRNA (Hallbeck et al., 1996b).Thus, because neither the spinal gray matter (presentstudy) nor dorsal root ganglia (Hallbeck et al., 1996b)contains VP mRNA, the PVH, which expresses VP mRNAin subdivisions known to contain neurons with spinalprojections (i.e., dp, lp, and mpv; Swanson and Kuypers,1980), is likely to be the major source of the descendingvasopressin projections to the brainstem and spinal cord.The distribution of the spinally projecting VP mRNA-expressing neurons within the PVH is described in theaccompanying paper (Hallbeck and Blomqvist, 1999).
ACKNOWLEDGMENTS
We thank Dr. P.P. Sanna, Dr. S.L. Sabol, and Dr. E. Mohrfor their generous gifts of the preprovasopressin, preproen-kephalin, and preprooxytocin cDNA, respectively. We thankDr. D. Larhammar for preparation of the plasmids.
LITERATURE CITED
Alonso G. 1988. Effects of colchicine on the intraneural transport ofsecretory material prior to the axon: a morphofunctional study inhypothalamic neurosecretory neurons of the rat. Brain Res 453:191–203.
Angerer LM, Stolter MH, Angerer RC. 1987. In situ hybridization with RNAprobes: an annotated recipe. In: Valentino KL, Eberwine JH, BarchasJD, editors. In situ hybridization. Application to neurobiology. Oxford:Oxford University Press. p 42–70.
Banet M, Wieland U-E. 1985. The effect of intraseptally applied vasopres-sin on thermoregulation in the rat. Brain Res Bull 14:113–116.
Bartanusz V, Aubry J-M, Steimer T, Baffi J, Kiss JZ. 1994. Stressor-specificincrease of vasopressin mRNA in paraventricular hypophysiotrophicneurons. Neurosci Lett 170:35–38.
Berntson GG, Berson BS. 1980. Antinociceptive effects of intraventricularor systemic administration of vasopressin in the rat. Life Sci 26:455–459.
Berson BS, Berntson GG, Zipf W, Torello MW, Kirk WT. 1983. Vasopressin-induced antinociception: an investigation into its physiological andhormonal basis. Endocrinology 113:337–343.
Bluthe R-M, Schoenen J, Dantzer R. 1990. Androgene-dependent vaso-pressinergic neurons are involved in social recognition in rats. BrainRes 519:150–157.
Bodnar RJ, Zimmerman EA, Nilaver G, Mansour A, Thomas LW, Kelly DD,Glusman M. 1980. Dissociation of cold-water swim and morphineanalgesia in Brattleboro rats with diabetes insipidus. Life Sci 26:1581–1590.
Boer GJ, Van Rheenen-Verberg CM, Uylings HB. 1982. Impaired braindevelopment of the diabetes insipidus Brattleboro rat. Brain Res255:557–575.
Bohus B. 1977. Effect of desglycinamide-lysine vasopressin (DG-LVP) onsexually motivated T-maze behavior of the male rat. Horm Behav8:52–61.
Bohus B, Kovacs GL, de Wied D. 1978. Oxytocin, vasopressin and memory:opposite effects on consolidation and retrieval processes. Brain Res157:414–417.
Brinton RE, McEwen BS. 1989. Vasopressin neuromodulation in thehippocampus. J Neurosci 9:752–759.
Brinton RE, Gee KW, Wamsley JK, Davis TP, Yamamura HL. 1984.Regional distribution of putative vasopressin receptors in rat brain andpituitary by quantitative autoradiography. Proc Natl Acad Sci USA81:7248–7252.
Buijs RM. 1978. Intra- and extrahypothalamic vasopressin and oxytocinpathways in the rat. Cell Tissue Res 192:423–435.
Burbach JP, Voorhuis TA, van Tol HH, Ivell R. 1987. In situ hybridization ofoxytocin messenger RNA: macroscopic distribution and quantitation inrat hypothalamic cell groups. Biochem Biophys Res Commun 145:10–14.
Caffe AR, van Leeuwen FW. 1983. Vasopressin-immunoreactive cells in thedorsomedial hypothalamic region, medial amygdaloid nucleus andlocus coeruleus of the rat. Cell Tissue Res 233:23–33.
Caffe AR, van Leeuwen FW, Buijs RM, de Vries GJ, Geffard M. 1985.Coexistence of vasopressin, neurophysin and noradrenaline immunore-activity in medium-sized cells of the locus coeruleus and subcoeruleusin the rat. Brain Res 338:160–164.
Caffe AR, van Leeuwen FW, Luiten PGM. 1987. Vasopressin cells in themedial amygdala of the rat project to the lateral septum and ventralhippocampus. J Comp Neurol 261:237–252.
Caffe AR, van Leeuwen FW, Buijs RM, van der Gugten J. 1988. Vasopressinand noradrenaline coexistence in the rat locus coeruleus: differentialdecreases of their levels in distant brain areas after thermal andneurotoxic lesions. Brain Res 459:386–390.
Carrive P. 1991. Functional organization of PAG neurons controllingregional vascular beds. In: Depaulis A, Bandler R, editors. The mid-brain periaqueductal gray matter. New York: Plenum. p 67–100.
198 M. HALLBECK ET AL.

Ceccatelli S, Cortes R, Hokfelt T. 1991. Effect of reserpine and colchicine onneuropeptide mRNA levels in the rat hypothalamic paraventricularnucleus. Mol Brain Res 9:57–69.
Chung SK, McCabe JT, Pfaff DW. 1991. Estrogen influences on oxytocinmRNA expression in preoptic and anterior hypothalamic regions stud-ied by in situ hybridization. J Comp Neurol 307:281–295.
Cooper KE, Kasting NW, Lederis K, Veale WL. 1979. Evidence supporting arole for endogenous vasopressin in natural suppression of fever in thesheep. J Physiol (Lond) 295:33–45.
Crispino M, Perrone Capano C, Kaplan BB, Giuditta A. 1993. Neurofila-ment proteins are synthesized in nerve endings from squid brain. JNeurochem 61:1144–1146.
de Wied D, Versteeg DHG. 1979. Neurohypophyseal principles and memory.Fed Proc 38:2348–2354.
de Wied D, Diamant M, Fodor M. 1993. Central nervous system effects ofthe neurohypophyseal hormones and related peptides. Front Neuroen-docrin 14:251–302.
De Vries GJ, Buijs RM. 1983. The origin of the vasopressinergic andoxytocinergic innervation of the rat brain with special reference to thelateral septum. Brain Res 273:307–317.
De Vries GJ, Buijs RM, Van Leeuwen FW, Caffe AR, Swaab DF. 1985. Thevasopressinergic innervation of the brain in normal and castrated rats.J Comp Neurol 233:236–254.
Dierickx K, Vandesande F, De Mey J. 1976. Identification, in the externalregion of the rat median eminence, of separate neurophysin-vasopres-sin and neurophysin-oxytocin containing fibers. Cell Tissue Res 168:141–151.
Dubois-Dauphin M, Tribollet E, Dreifuss JJ. 1987. A sexually dimorphicvasopressin innervation of auditory pathways in the guinea pig brain.Brain Res 437:151–156.
Fulwiler CE, Saper CB. 1984. Subnuclear organization of the efferentconnections of the parabrachial nucleus in the rat. Brain Res Rev7:229–259.
Giri PR, Dave JR, Tabakoff B, Hoffman PL. 1990. Arginine vasopressininduces the expression of c-fos in the mouse septum and hippocampus.Mol Brain Res 7:131–137.
Giuditta A, Menichini E, Perrone Capano C, Langella M, Martin R, CastigliE, Kaplan BB. 1991. Active polysomes in the axoplasm of the squidgiant axon. J Neurosci Res 28:18–28.
Granger R, Wiebe SP, Taketani M, Lynch G. 1996. Distinct memory circuitscomposing the hippocampal region. Hippocampus 6:567–578.
Hallbeck M, Blomqvist A. 1999. Spinal cord-projecting vasopressinergicneurons in the rat paraventricular hypothalamus. J Comp Neurol,companion paper.
Hallbeck M, Hermanson O, Blomqvist A. 1996a. Paraventricular hypothala-mus, the source of spinal vasopressin fibers in the rat. Soc NeurosciAbstr 22:872.
Hallbeck M, Hermanson O, Blomqvist A. 1996b. Preprovasopressin mRNAis not present in dorsal root ganglia of the rat. Neurosci Lett 209:125–128.
Herbert H, Moga MM, Saper, CB. 1990. Connections of the parabrachialnucleus with the nucleus of the solitary tract and the medullaryreticular formation in the rat. J Comp Neurol 293:540–580.
Hermanson O, Hallbeck M, Blomqvist A. 1995. Preproenkephalin mRNA-expressing neurones in the rat thalamus. Neuroreport 6:833–836.
Hess J, Lesser D, Landgraf R. 1992. Vasopressin and oxytocin in brainareas of rats selectively bred for differences in behavioral performance.Brain Res 569:106–111.
Hoffman PL, Ishizawa H, Giri PR, Dave JR, Grant KA, Liu L-I, Gulya K,Tabakoff B. 1990. The role of arginine vasopressin in alcohol tolerance.Ann Med 22:269–274.
Hoorneman EM, Buijs RM. 1982. Vasopressin fiber pathways in the ratbrain following suprachiasmatic nucleus lesioning. Brain Res 243:235–241.
Jirikowski GF, Sanna PP, Bloom FE. 1990. mRNA coding for oxytocin ispresent in axons of the hypothalamo-neurohypophysial tract. Proc NatlAcad Sci USA 87:7400–7404.
Kai-Kai MA, Anderton BH, Keen P. 1986. A quantitative analysis of theinterrelationships between subpopulations of rat sensory neuronscontaining arginine vasopressin or oxytocin and those containingsubstance P, fluoride-resistant acid phosphatase or neurofilamentprotein. Neuroscience 18:475–486.
Kelly AB, Watts AG. 1998. The region of the pontine parabrachial nucleus isa major target of dehydration-sensitive CRH neurons in the rat lateralhypothalamic area. J Comp Neurol 394:48–63.
Koenig E. 1991. Evaluation of local synthesis of axonal proteins in thegoldfish Mauthner cell axon and axons of dorsal and ventral roots of therat in vitro. Mol Cell Neurosci 2:384–394.
Kordower JH, Bodnar RJ. 1984. Vasopressin analgesia: specificity of actionand non-opioid effects. Peptides 5:747–756.
Lang RE, Heil J, Ganten D, Hermann K, Rascher W, Unger T. 1983. Effectsof lesions in the paraventricular nucleus of the hypothalamus onvasopressin and oxytocin contents in brainstem and spinal cord of rat.Brain Res 260:326–329.
Langhans W, Delprete E, Scharrer E. 1991. Mechanisms of vasopressin’sanorectic effect. Physiol Behav 49:169–176.
Lehmann E, Hanze J, Pauschinger M, Ganten D, Lang RE. 1990. Vasopres-sin mRNA in the neurolobe of the rat pituitary. Neurosci Lett 111:170–175.
Lepetit P, Fevre-Montange M, Gay N, Belin MF, Bobillier P. 1993. Vasopres-sin mRNA in the cerebellum and circumventricular organs: a quantita-tive in situ hybridization study. Neurosci Lett 159:171–174.
Leshner AI, Roche KE. 1977. Comparison of the effects of ACTH and lysinevasopressin on avoidance-of-attack in mice. Physiol Behav 18:879–883.
Maciejewski-Lenoir D, Jirikowski GF, Sanna PP, Bloom FE. 1993. Reduc-tion of exogenous vasopressin RNA poly(A) tail length increases itseffectiveness in transiently correcting diabetes insipidus in the Brat-tleboro rat. Proc Natl Acad Sci USA 90:1435–1439.
Maegawa H, Katsube N, Okegawa T, Aishita H, Kawasaki A. 1992.Arginine-vasopressin fragment 4–9 stimulates the acetylcholine re-lease in hippocampus of freely-moving rats. Life Sci 51:285–293.
Malpas SC, Coote JH. 1994. Role of vasopressin in sympathetic response toparaventricular nucleus stimulation in anesthetized rats. Am J Physiol266:R228–R236.
Mason WT, Ho YW, Hatton GI. 1984. Axon collaterals of supraopticneurones: anatomical and electrophysiological evidence for their exis-tence in the lateral hypothalamus. Neuroscience 11:169–182.
McCabe JT, Lehmann E, Chastrette N, Hanze J, Lang RE, Ganten D, PfaffDW. 1990. Detection of vasopressin mRNA in the neurointermediatelobe of the rat pituitary. Mol Brain Res 8:325–329.
Metzger D, Alescio-Lautier B, Bosler O, Devigne C, Soumireu-Mourat B.1993. Effect of changes in the intrahippocampal vasopressin on memoryretrieval and relearning. Behav Neural Biol 59:29–48.
Meyerson BJ, Hoglund U, Johansson C, Blomqvist A, Ericson H. 1988.Neonatal vasopressin antagonist treatment facilitates adult copulatorybehavior in female rats and increases hypothalamic vasopressin con-tent. Brain Res 473:344–351.
Miller MA, Kolb PE, Raskind MA. 1993. Extra-hypothalamic vasopressinneurons coexpress galanin messenger RNA as shown by double in situhybridization histochemistry. J Comp Neurol 329:378–384.
Moga MM, Herbert H, Hurley KM, Yasui Y, Gray TS, Saper CB. 1990.Organization of cortical, basal forebrain, and hypothalamic afferents tothe parabrachial nucleus in the rat. J Comp Neurol 295:624–661.
Mohr E, Zhou A, Thorn NA, Richter D. 1990. Rats with physicallydisconnected hypothalamo-pituitary tracts no longer contain vasopres-sin-oxytocin gene transcripts in the posterior pituitary lobe. FEBS Lett263:332–336.
Mohr E, Fehr S, Richter D. 1991. Axonal transport of neuropeptideencoding mRNAs within the hypothalamo-hypophyseal tract of rats.EMBO J 10:2419–2424.
Murphy D, Levy A, Lightman S, Carter D. 1989. Vasopressin RNA in theneural lobe of the pituitary: dramatic accumulation in response to saltloading. Proc Natl Acad Sci USA 86:9002–9005.
Ostrowski NL, Lolait SJ, Young WS III. 1994. Cellular localization ofvasopressin V1a receptor messenger ribonucleic acid in adult male ratbrain, pineal, and brain vasculature. Endocrinology 135:1511–1528.
Penna M, Capranica RR, Somers J. 1992. Hormone-induced vocal behaviorand midbrain auditory sensitivity in the green treefrog, Hyla cinerea. JComp Physiol A 170:73–82.
Planas B, Kolb PE, Raskind MA, Miller MA. 1995. Vasopressin and galaninmRNAs coexist in the nucleus of the horizontal diagonal band: a novelsite of vasopressin gene expression. J Comp Neurol 361:48–56.
Pu L-P, van Leeuwen FW, Tracer HL, Sonnemans MAF, Loh YP. 1995.Localization of vasopressin mRNA and immunoreactivity in pituicytesof pituitary stalk-transected rats after osmotic stimulation. Proc NatlAcad Sci USA 92:10653–10657.
Rethelyi M, Mohapatra NK, Metz CB, Petrusz P, Lund PK. 1991. Colchicineenhances mRNAs encoding the precursor of calcitonin gene-relatedpeptide in the brainstem motorneurons. Neuroscience 42:531–539.
Rehbein M, Hillers M, Mohr E, Ivell R, Morley S, Schmale H, Richter D.
VASOPRESSIN mRNA IN THE RAT CNS 199

1986. The neurohypophyseal hormones vasopressin and oxytocin. Pre-cursor structure, synthesis and regulation. Biol Chem 367:695–704.
Rhodes CH, Morrell JI, Pfaff DW. 1981. Immunohistochemical analysis ofmagnocellular elements in rat hypothalamus: distribution and num-bers of cells containing neurophysin, oxytocin, and vasopressin. J CompNeurol 198:45–64.
Sawchenko PE, Swanson LW. 1982. Immunohistochemical identification ofneurons in the paraventricular nucleus of the hypothalamus thatproject to the medulla or to the spinal cord in the rat. J Comp Neurol205:260–272.
Sawchenko PE, Swanson LW, Vale WW. 1984a. Co-expression of corticotro-pin-releasing factor and vasopressin immunoreactivity in parvocellularneurosecretory neurons of the adrenalectomized rat. Proc Natl Acad SciUSA 81:1883–1887.
Sawchenko PE, Swanson LW, Vale WW. 1984b. Corticotropin-releasingfactor: co-expression within distinct subsets of oxytocin-, vasopressin-,and neurotensin-immunoreactive neurons in the hypothalamus of themale rat. J Neurosci 4:1118–112.
Simmons DM, Arizza JL, Swanson LW. 1989. A complete protocol for in situhybridization of messenger RNAs in brain and other tissues withradiolabeled single-stranded RNA probes. J Histotechnol 12:169–181.
Smock T, Arnold S, Albeck D, Emerson P, Girritano J, Burrows K, Derber W,Sanson C, Marrs K, Weatherly H, Kruse K. 1992. A peptidergic circuitfor reproductive behavior. Brain Res 598:138–142.
Sodersten P, Henning M, Melin P, Ludin S. 1983. Vasopressin alters femalesexual behaviour by acting on the brain independently of alterations inblood pressure. Nature 301:608–610.
Sofroniew MV. 1985. Vasopressin- and neurophysin-immunoreactive neu-rons in the septal region, medial amygdala and locus coeruleus incolchicine-treated rats. Neuroscience 15:347–358.
Sofroniew MV, Weindl A. 1980. Identification of parvocellular vasopressinand neurophysin neurons in the suprachiasmatic nucleus of a variety ofmammals including primates. J Comp Neurol 193:659–675.
Stebbins CL, Oritiz-Acevedo A, Hill JM. 1992. Spinal vasopressin modu-lates the reflex cardiovascular response to static contraction. J ApplPhysiol 72:731–738.
Swanson LW. 1992. Brain maps: structure of the rat brain. Amsterdam:Elsevier.
Swanson LW, Kuypers HGJM. 1980. The paraventricular nucleus of thehypothalamus: cytoarchitectonic subdivisions and organization of pro-jections to the pituitary, dorsal vagal complex, and spinal cord asdemonstrated by retrograde fluorescence double-labeling methods. JComp Neurol 194:555–570.
Swanson LW, Sawchenko PE. 1983. Hypothalamic integration: organiza-
tion of the paraventricular and supraoptic nuclei. Annu Rev Neurosci6:269–324.
Thurston CL, Culhane ES, Suberg SN, Carstens E, Watkins LR. 1988.Antinociception vs motor effects of intrathecal vasopressin as measuredby four pain tests. Brain Res 463:1–11.
Trembleau A, Morales M, Bloom FE. 1994. Aggregation of vasopressinmRNA in a subset of axonal swellings of the median eminence andposterior pituitary: light and electron microscopic evidence. J Neurosci14:39–53.
Trembleau A, Morales M, Bloom FE. 1996. Differential compartmentaliza-tion of vasopressin messenger RNA and neuropeptide within the rathypothalamo-neurohypophysial axonal tracts: light and electron micro-scopic evidence. Neuroscience 70:113–125.
Uhl GR, Reppert SM. 1986. Suprachiasmatic nucleus vasopressin messen-ger RNA: circadian variation in normal and Brattleboro rats. Science232:390–393.
Urban JH, Miller MA, Drake CT, Dorsa DM. 1990. Detection of vasopressinmRNA in cells of the medial amygdala but not the locus coeruleus by insitu hybridization. J Chem Neuroanat 3:277–283.
van Leeuwen F, Caffe R. 1983. Vasopressin-immunoreactive cell bodies inthe bed nucleus of the stria terminalis of the rat. Cell Tissue Res228:525–534.
van Minnen I. 1994. RNA in the axonal domain: a new dimension in neuralfunctioning? Histochem J 26:377–391.
Wang Z, Zhou L, Hulihan TJ, Insel TR. 1996. Immunoreactivity of centralvasopressin and oxytocin pathways in microtine rodents: a quantitativecomparative study. J Comp Neurol 366:726–737.
Versteeg CAM, de Jong W, Bohus B. 1984. Arginine-vasopressin inhibitscentrally induced pressor responses by involving hippocampal mecha-nisms. Brain Res 292:317–326.
Wideman CH, Murphy HM. 1993. Modulatory effects of vasopressin onglucose and protein metabolism during food-restriction stress. Peptides14:259–261.
Wolfson B, Manning RW, Davis LG, Arentzen R, Baldino F Jr. 1985.Co-localization of corticotropin releasing factor and vasopressin mRNAin neurones after adrenalectomy. Nature 315:59–61.
Wu PH, Lanca AJ, Liu JF, Man CF, Kalant H. 1995. Peripheral injection ofarginine8-vasopressin increases fos in specific brain areas. Eur JPharmacol 281:263–269.
Yoshikawa K, Williams C, Sabol SL. 1984. Rat brain preproenkephalinmRNA cDNA cloning, primary structure, and distribution in the centralnervous system. J Biol Chem 259:14301–14308.
Young LJ, Wang Z, Insel TR. 1998. Neuroendocrine bases of monogamy.Trends Neurosci 21:71–75.
200 M. HALLBECK ET AL.