dna la scoperta - lumsa.it dna la... · chirurgiche con il pus (formato di cellule) e osservò una...
TRANSCRIPT

DNA ...la scoperta

1665 il matematico, fisico, astronomo e naturalista inglese Robert Hooke (1635-1702), usando un microscopio di sua invenzione, notò che il sughero e altri tessuti vegetali erano formati da piccole cavità separate da pareti; egli chiamò queste cavità «celle», cioè «piccole stanze». Il termine «cellula» ha assunto il suo attuale significato, cioè «unità di base della materia vivente», soltanto 150 anni dopo la scoperta di Hooke.

1838, Matthias J. Schleiden (1804-1881), un botanico tedesco, giunse alla conclusione che tutti i tessuti vegetali sono costituiti da insiemi organizzati di cellule.
1839, lo zoologo tedesco Theodor Schwann (1810-1882) estese le osservazioni di Schleiden ai tessuti animali e propose una base cellulare comune a tutti gli organismi viventi.
1838-1839
Che noia……
La mancanza di figure rende la lezione molto noiosa, proviamo ad aggiungere figure

1869 La scoperta del DNA è attribuita al ricercatore svizzero Friedrich Miescher già nel 1869. La storia racconta che Miescher lavorava in un laboratorio dove si cercava di comprendere la struttura delle cellule. Per avere a disposizione tanto materiale, raccolse le bende chirurgiche con il pus (formato di cellule) e osservò una sostanza microscopica a cui diede il nome di nucleina. Questa sostanza era associata con proteine e derivava dal nucleo delle cellule. Miescher scoprì che la nucleina era composta di idrogeno, ossigeno azoto e fosforo ed è stato il primo ad identificare il DNA come una molecola distinta.
http://www.dnaftb.org/15/animation.html
Ancora non ci siamo… i bambini devono poter immaginare ciò di cui stiamo parlando, quindi servono più figure…

E allora…..
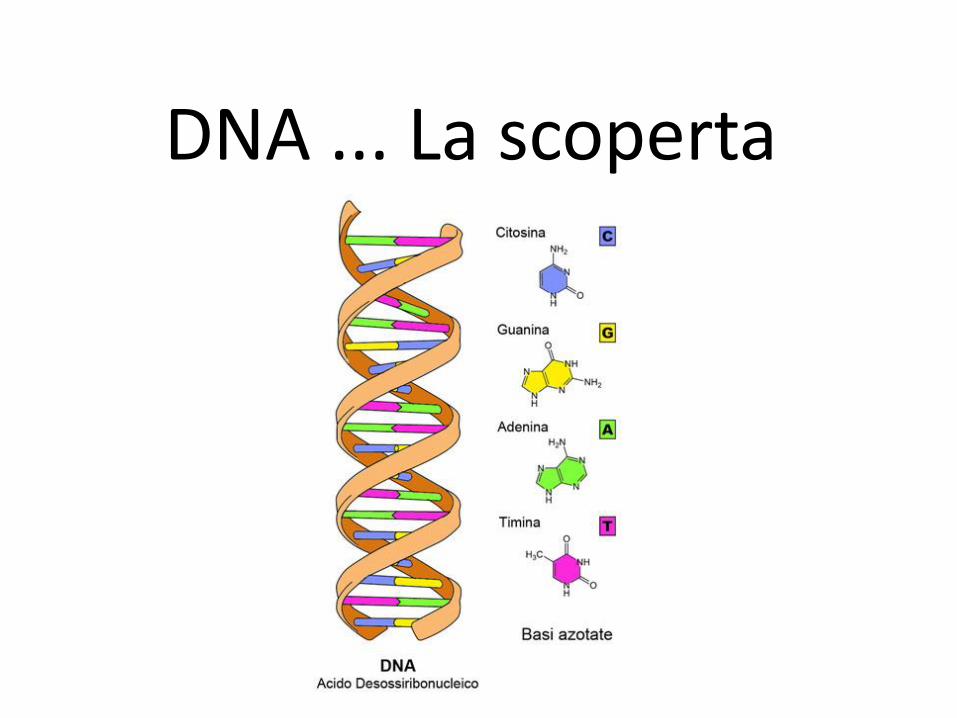
DNA ... La scoperta

1665 il matematico, fisico, astronomo e naturalista inglese Robert Hooke (1635-1702), usando un microscopio di sua invenzione, notò che il sughero e altri tessuti vegetali erano formati da piccole cavità separate da pareti; egli chiamò queste cavità «celle», cioè «piccole stanze». Il termine «cellula» ha assunto il suo attuale significato, cioè «unità di base della materia vivente», soltanto 150 anni dopo la scoperta di Hooke.

1838, Matthias J. Schleiden (1804-1881), un botanico tedesco, giunse alla conclusione che tutti i tessuti vegetali sono costituiti da insiemi organizzati di cellule.
1839, lo zoologo tedesco Theodor Schwann (1810-1882) estese le osservazioni di Schleiden ai tessuti animali e propose una base cellulare comune a tutti gli organismi viventi.
1838-1839

1858 l’idea che tutti gli organismi fossero formati da una o più cellule assunse un significato ancora più ampio quando l’anatomopatologo tedesco Rudolf Virchow (1821-1902) affermò che le cellule possono essere originate solo da altre cellule preesistenti: «Quando una cellula esiste, ci dev’essere stata una cellula preesistente, proprio come un animale si origina solo da un animale e una pianta si origina solo da una pianta».
Il suo studio

1869 La scoperta del DNA è attribuita al ricercatore svizzero Friedrich Miescher già nel 1869. La storia racconta che Miescher lavorava in un laboratorio dove si cercava di comprendere la struttura delle cellule. Per avere a disposizione tanto materiale, raccolse le bende chirurgiche con il pus (formato di cellule) e osservò una sostanza microscopica a cui diede il nome di nucleina. Questa sostanza era associata con proteine e derivava dal nucleo delle cellule. Miescher scoprì che la nucleina era composta di idrogeno, ossigeno azoto e fosforo ed è stato il primo ad identificare il DNA come una molecola distinta.
http://www.dnaftb.org/15/animation.html

1919 Phoebus Levene nel 1919 scoprì la struttura dei nucleotidi, formati da base azotata, fosfato e zuccheri. Levene sostenne per la prima volta che il DNA è un filamento di nucleotidi legati tra loro tramite fosfati la cui ripetitività era costante. Levene maturò anche la convinzione che tale filamento fosse corto e che le proteine con la loro struttura più complessa fossero le giuste candidate ad essere le molecole dell’eredità.
160,000 combinazioni

1928 L'esperimento di Frederick Griffith del 1928
?
Questo fu il primo esperimento a suggerire che i batteri possono trasferire informazioni genetiche attraverso un processo noto come trasformazione. Il materiale genetico di un batterio si traferisce ad un altro. L’esperimento aprì la strada alla determinazione di quale fosse la natura del materiale genetico.

1937 Trascorrono quasi venti anni prima che William Astbury, nel 1937, possa presentare alla comunità scientifica i primi risultati dei suoi studi sulla diffrazione a raggi X, i quali dimostrano che il DNA ha una struttura molto regolare.
Questa è una delle prime cristallografie pubblicate da Astbury, è un capello di Mozart!

1943 L'esperimento di Oswald Theodore Avery e dei suoi colleghi Colin MacLeod e Maclyn McCarty, rappresenta una delle esperienze fondamentali per l'avanzamento delle conoscenze nel campo della genetica e della biologia molecolare.
Tramite l'esperimento riuscirono a
dimostrare che il cosiddetto principio
trasformante (ovvero il portatore di
informazioni geniche scoperto nel 1928 da
Griffith in seguito al suo famoso
esperimento) era il DNA.

1952 L'esperimento di Alfred D. Hershey e Martha Chase prova definitivamente nel 1952 che il materiale genetico è costituito da DNA e non da proteine.
In seguito a questi risultati incontrovertibili anche gli scienziati che avevano criticato esperimenti precedenti, si convincono dell'importantissimo ruolo biologico del DNA.

1951-52
Erwin Chargaff formula due regole: 1) Esiste un rapporto 1:1 tra le basi puriniche (A+G) e le basi pirimidiniche (T+C) contenute nel DNA di una cellula. Il rapporto è costante in tutte le specie.

2) in una molecola di DNA a doppio filamento la concentrazione di adenina eguaglia quella di timina e la concentrazione di citosina quella di guanina (%A = %T; %C = %G).
Questa ultima semplice regola è stata uno degli elementi essenziali che hanno permesso la formulazione del modello di DNA da parte di James Watson e Francis Crick. Grazie anche a questa regola si sono dedotte le corrette forme di appaiamento delle basi tra i due filamenti del DNA.

Watson e Crick costruirono diversi modelli di DNA, con cartone e fil di ferro, cercando di far combaciare tutte le informazioni che avevano a disposizione. Costruirono il modello basandosi su un bozzetto di Odile Speed, moglie di Crick, che lo aveva disegnato seguendo le istruzioni del marito: il bozzetto compare sulla rivista scientifica Nature il 25 aprile del 1953 quando la struttura a doppia elica viene presentata per la prima volta.


1953 Nel 1953 i due scienziati ipotizzano che il DNA si componga di due catene di nucleotidi disposte a formare una doppia elica. Ciascuna purina, composta da due anelli, si trova di fronte ad una pirimidina, composta da un singolo anello. L'adenina (A) si lega alla timina (T) mentre la citosina (C) si lega alla guanina (G), in accordo con i risultati di Chargaff. I legami tra le basi puriniche e pirimidiniche avvengono tramite ponti idrogeno.

Avery e Chargaff • Avery stabilì che il DNA (e non le
proteine) era il materiale genetico
della cellula.
• Chargaff scoprì che la propozione
delle quattro basi azotate nel DNA è
la stessa in tutte le cellule degli
individui della stessa specie.
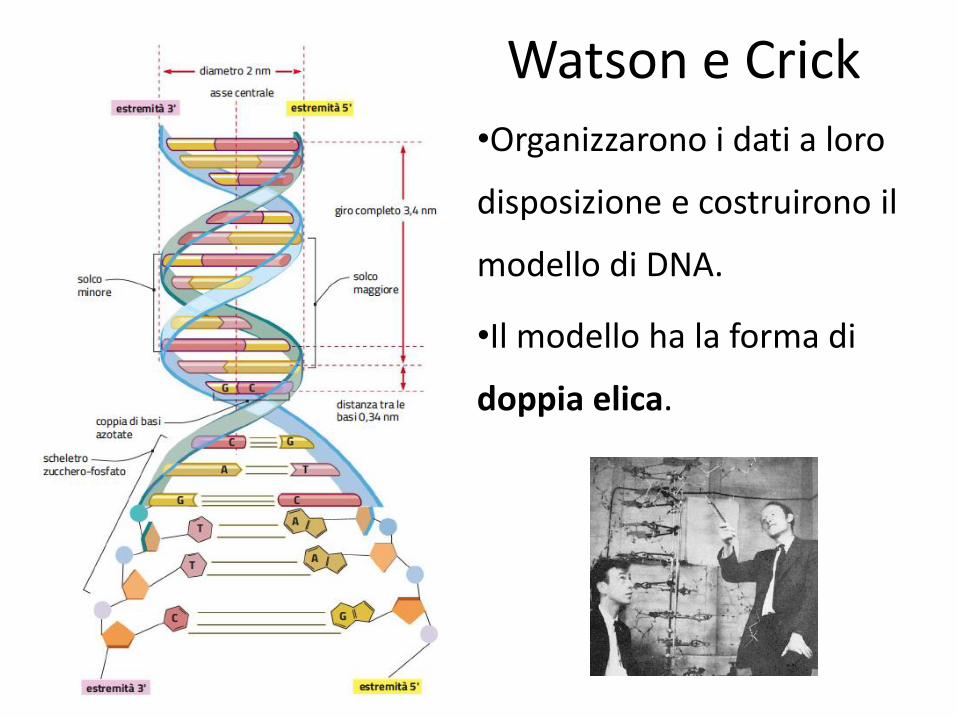
Watson e Crick •Organizzarono i dati a loro
disposizione e costruirono il
modello di DNA.
•Il modello ha la forma di
doppia elica.

Il DNA
• I cromosomi sono costituiti
sia da acido deossiribonucleico,
o DNA, sia da proteine.
• Gli elementi di base del DNA
sono i nucleotidi.

I nucleotidi
• Ogni nucleotide è composto
da una base azotata, uno
zucchero (deossiribosio) e un
gruppo fosfato.
• Le basi azotate sono di due
tipi: le purine, adenina (A) e
guanina (G); le pirimidine,
citosina (C) e timina (T).



L’appaiamento tra basi
• Ogni base forma un legame covalente
con lo zucchero adiacente.
• Le basi appaiate si incontrano
sull’asse centrale dell’elica con legami a
idrogeno.
• Le basi appaiate sono complementari
dove (A=T) e (G≡C).

La struttura
• Contiene due catene
• Le percentuali G=C e A=T
• Legami fosfodiesterici tra nucleotidi




Lewin, IL GENE VIII, Zanichelli editore S.p.A. Copyright © 2006

N
N N
N
N N
O
H
H
O
N
s u g a r
H
s u g a r
C H 3 timina
adenina
N
N N
N
O
N
H
N N
N
H
O
s u g a r
H
H
H
s u g a r
citosina
guanina
L’appaiamento delle basi
implica la formazione di legami idrogeno

Lewin, IL GENE VIII, Zanichelli editore S.p.A. Copyright © 2006

Figura riportata nella
pubblicazione originale
di Watson e Crick
Nature, Aprile 1953

Lewin, IL GENE VIII, Zanichelli editore S.p.A. Copyright © 2006
20

La struttura del DNA
• Il DNA ha una forma ad elica regolare con legami idrogeno tra le basi
• L’impilamento delle basi è determinato da
interazioni idrofobiche
• Ogni coppia è ruotata di 36°
• Solchi maggiore (22Å) e minore (12Å)
• Avvolgimento in senso orario (elica destrorsa)

Nature, Vol. 171, p.737, April 25, 1953
MOLECULAR STRUCTURE OF
NUCLEIC ACIDS
A Structure for Deoxyri bose Nucleic Acid
We wish to s uggest a structure for the salt of
deoxyribose nucleic acid (D.N.A.). This structure has
novel features which are of considerable biological
interest.
A structure for nucleic acid has already been
proposed by Pauling and Corey (1). They kind ly made
their manuscript available to us in advance of
publication. Their model consists of three intertwined
chains, with the phosphates near the fibre axis, and the
bases on the outside. In our opinion, this structure is
unsatisfactory for two reasons: (1) We believe that the
material which gives the X-ray diagrams is the salt, not
the free acid. Without the acidic hydrogen atoms it is
not clear what forces would hold the structure together,
especially as the negatively charged phosphates near
the axis will repel each other. (2) Some of the van der
Waals distances appear to be too small.
Another three-chain structure has also been
suggested by F raser (in the press). In his model the
phosphates are on the outside and the bases on the
inside, linked together by hyd rogen bonds. This
structure as described is rather ill- defined, and for this
reason we shall not comment on it.
We wish to put forward a ra dically different
structure for the salt of deoxyribose nucleic acid. This
structure has two helical chains each coiled round the
same axis (see diagram). We have made the usual
chemical assumptions, namely, that each chain consists
of phosphate diester groups joining §-D-
deoxyribofuranose residues with 3',5' linkages. The two
chains (but not their bases) are related by a dyad
perpendicular to the fibre axis. Both chains follow
right- handed helices, but owing to the dyad the
sequences of the atoms in t he two chains run in
opposite directions. Each chain loosely resembles
Furberg's2 model No. 1; that is, the bases are on the
inside of the helix and the phosphates on the outside.
The configuration of the sugar and the atoms near it is
close to Furberg's 'standard configu ration', the sugar
being roughly perpendicular to the attached base. There
is a residue on each every 3.4 A. in the z-direction. We
have assumed an ang le of 36¡ between adjacent
residues in the same chain, so that the structure repeats
after 10 residues on each chain, that is, after 34 A. The
distance of a phosphorus atom from the fibre axis is 10
A. A s the phosphates are on the outside, cations have
easy access to them.
The structure is an open one, and its water content
is rather high. At lower water contents we would expect
the bases to tilt so that the structure could become more
compact.
The novel feature of the structure is the manner in
which the two chains are held together by the purine
and pyrimidine bases. The planes of the bases are
perpendicular to the fibre axis. The are joined together
in pairs, a single base from the other chain, so that the
two lie side by side with identical z-co-ordinates. One
of the pair must be a purine and the other a pyrimidine
for bonding to occur. The hydrogen bonds are made as
follows : purine position 1 t o pyrimidine position 1 ;
purine position 6 to pyrimidine position 6.
If it is assumed that the bases only occur in the
structure in the most plausible tautomeric forms (that is,
with the keto rather than the enol configu rations) it is
found that only specific pairs of bases can bond
together. These pairs are : adenin e (purine) with
thymine (pyrimidine), and guanine (purine) with
cytosine (pyrimidine).
In other words, if an adenine forms one member of
a pair, on either chain, then on th ese assumptions the
other member must be thymine ; similarly for guanine
and cytosine. The sequence of bases on a s ing le chain
does not appear to be restricted in any way. However, if
only specific pairs of bases can be formed, it follows
that if the sequence of bases on one chain is given, then
the sequence on t he other chain is automatically
determined.
It has been found experimentally (3,4) that the ratio
of the amounts of adenine to thymine, and the ration of
guanin e to cytosine, are always bery close to unity for
deoxyribose nucleic acid.
It is probably impossible to build this structure
with a ribose sugar in place of the deoxyribose, as the
extra oxygen atom would make too close a van der
Waals contact. The previously published X-ray data
(5,6) on deoxyribose nucleic acid are in sufficient for a
rigorous test of our structure. So far as we can tell, it is
roughly compatible with the experimental data, but it
must be regarded as unproved until it has been checked
against more exact results. Some of these are giv en in
the following communications. We were not aware of
the details of the results presented there when we
devised our structure, which rests mainly though not
entirely on published experimental data and
stereochemical arguments.
It has not escaped our notice that the specific
pairing we have postulated immediately suggests a
possible copying mechanism for the genetic material.
Full details of the structure, includ ing the
conditions assumed in building it, together with a set of
co-ordinates for the atoms, will be published elsewhere.
We are much ind ebted to Dr. Jerry Donohue for
constant advice and criticism, especially on interatomic
distances. We have also been stimulated by a
knowledge of the general nature of the unpublished
experimental results and ideas of Dr. M. H. F. Wilkins,
Dr. R. E . Franklin and their co-workers at King's
College, London. One of us (J. D. W.) has been aided
by a fellowship from the National Foundation for
Infantile Paralysis.
J. D. WATSON
F. H. C. CRICK
Medical Research Council Unit for the Study of
Molecular Structure of Biological Systems, Cavendish
Laboratory, Cambridge.
1. Pauling, L., and Corey, R. B., Nature, 171, 346
(1953); Proc. U.S. Nat. Acad. Sci., 39, 84 (1953).
2. Furberg, S., Acta Chem. Scand., 6, 634 (1952).
3. Ch argaff, E., for references see Zamenhof, S.,
Brawerman, G., and Chargaff, E., Bio chim. et Biophy s.
Acta, 9, 402 (1952).
4. Wyatt, G. R., J. Gen. Physiol., 36, 201 (1952).
5. A stbury, W. T., Symp. Soc. Exp. Biol. 1, Nucleic
Acid, 66 (Camb. Univ. Press, 1947 ).
6. W ilkins, M. H . F., and Randall, J. T., Biochim. et
Biophys. Acta, 10, 192 (1953).

«It has not escaped our notice that the specific pairing we have postulated
immediately suggests a possible copying mechanism for the genetic material»

Il modello a doppia elica di Crick e Watson
suggerisce un meccanismo di replicazione
OLD NEW OLD NEW
NEW NEW
OLD OLD
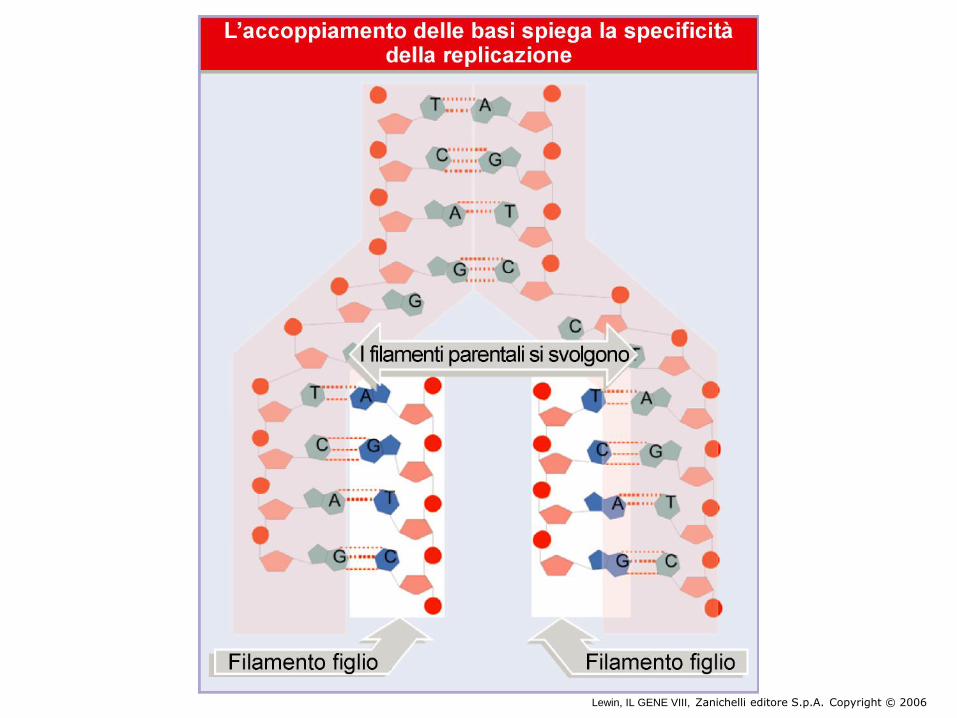
Lewin, IL GENE VIII, Zanichelli editore S.p.A. Copyright © 2006

La molecola di DNA forma
copie identiche di se stessa

Lewin, IL GENE VIII, Zanichelli editore S.p.A. Copyright ©
2006

Lewin, IL GENE VIII, Zanichelli editore S.p.A. Copyright © 2006

Lewin, IL GENE VIII, Zanichelli editore S.p.A. Copyright © 2006

Enzimi per gli acidi nucleici
•Polimerasi: DNA polimerasi RNA polimerasi
•Nucleasi: esonucleasi endonucleasi

Lewin, IL GENE VIII, Zanichelli editore S.p.A. Copyright © 2006

Lewin, IL GENE VIII, Zanichelli editore S.p.A. Copyright © 2006
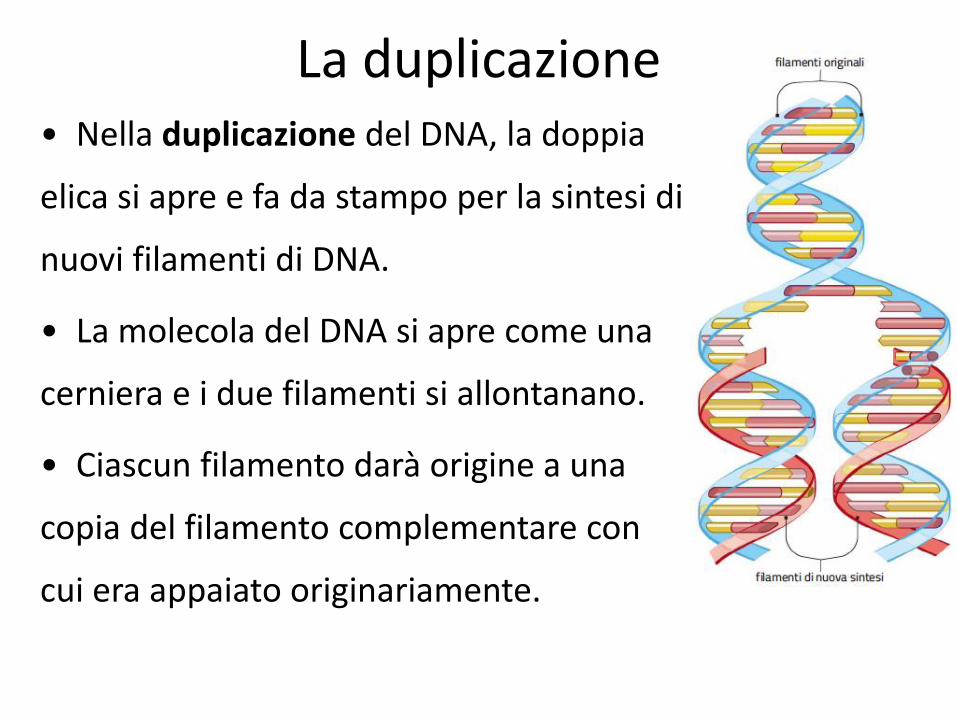
La duplicazione • Nella duplicazione del DNA, la doppia
elica si apre e fa da stampo per la sintesi di
nuovi filamenti di DNA.
• La molecola del DNA si apre come una
cerniera e i due filamenti si allontanano.
• Ciascun filamento darà origine a una
copia del filamento complementare con
cui era appaiato originariamente.

La duplicazione è semiconservativa
• Le basi si appaiano in modo
complementare.
• Ogni nuova doppia elica è formata
per metà da un filamento già
esistente e per metà da uno appena
sintetizzato: perciò è detta
semiconservativa.

Gli enzimi e il proofreading
• Gli enzimi controllano con precisione il processo di
duplicazione.
• Le DNA-polimerasi hanno il compito di aggiungere
nucleotidi al filamento in costruzione.
• Se si verifica un errore, lo correggono.
• La capacità di leggere le sequenze e di rimuovere i
nucleotidi che non sono appaiati in modo corretto è
detta proofreading.

La Sintesi Proteica Dal DNA alla proteina:
dai nucleotidi agli amminoacidi
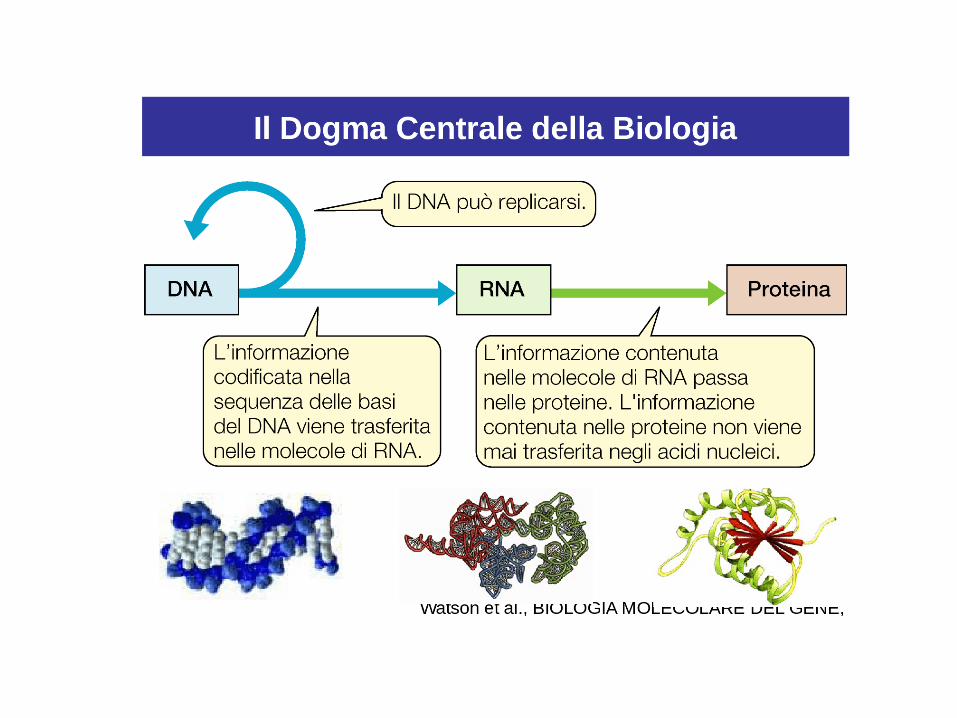

La traduzione e la trascrizione
• Il passaggio di informazioni dal DNA alla proteina è
detto traduzione.
• La trascrizione ha lo scopo di copiare o «trascrivere» il
messaggio contenuto nel DNA in una molecola
complementare.
• L’RNA è indispensabile nel processo di trascrizione.


La trascrizione
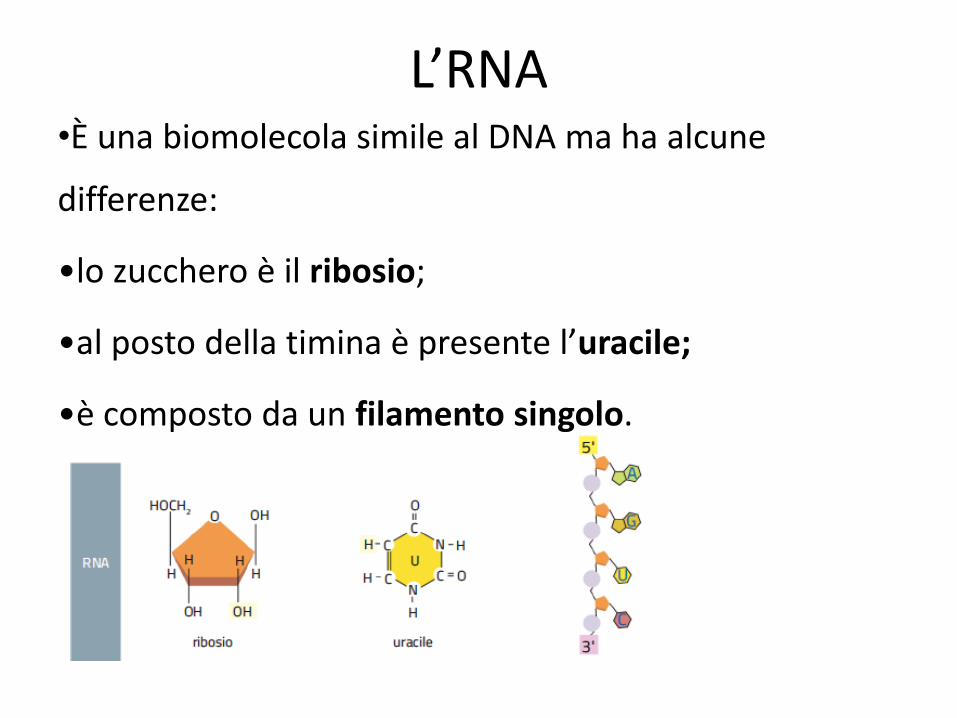
L’RNA •È una biomolecola simile al DNA ma ha alcune
differenze:
•lo zucchero è il ribosio;
•al posto della timina è presente l’uracile;
•è composto da un filamento singolo.

La trascrizione
• Le molecole di RNA messaggero sono copie (o trascritti)
di sequenze di nucleotidi presenti nel DNA.
• La trascrizione è catalizzata dall’enzima RNA polimerasi;
• L’RNA polimerasi e il DNA scorrono uno sull’altra
permettendo così la sintesi dell’mRNA a partire dal
filamento stampo.

La traduzione

I codoni
• I codoni sono segmenti di RNA contenenti tre
nucleotidi.
• Il codice genetico a triplette codifica le diverse
proteine.
• La struttura primaria di ogni tipo di proteina è
formata da una specifica disposizione lineare dei 20
differenti amminoacidi.

RNA ribosomale
• I ribosomi sono i siti dove avviene la
sintesi proteica. Sono costituiti in
parte da proteine e in parte da RNA,
detto RNA ribosomiale (rRNA).
• Ogni ribosoma è formato da due
subunità (maggiore e minore).

RNA di trasporto
• Ogni molecola di tRNA ha la
funzione di agganciare uno
specifico amminoacido per
trascinarlo fino all’mRNA.
• È composto da un sito di
attacco e un anticodone.

La sintesi proteica
•È il
trasferimento di
informazioni
dagli acidi
nucleici agli
amminoacidi.

La sintesi proteica nelle cellule eucariote
• Le proteine sono
sintetizzate sui ribosomi del
reticolo endoplasmatico e
poi inviate all’apparato di
Golgi.
• Da qui sono smistate ai
diversi comparti cellulari.