effects ofacidification mercury methylation, demethylation ... · lake water and sediment ph was...
TRANSCRIPT

Vol. 54, No. 8APPLIED AND ENVIRONMENTAL MICROBIOLOGY, Aug. 1988, p. 2003-20090099-2240/88/082003-07$02.00/0Copyright © 1988, American Society for Microbiology
Effects of Acidification on Mercury Methylation, Demethylation,and Volatilization in Sediments from an Acid-Susceptible Lake
ROBERT J. STEFFAN,t ERIC T. KORTHALS,t AND MICHAEL R. WINFREY*
River Studies Center, Department of Biology and Microbiology, University of Wisconsin-La Crosse,La Crosse, Wisconsin 54601
Received 2 November 1987/Accepted 19 May 1988
The effect of experimental acidification on mercury methylation, demethylation, and volatilization was
examined in surficial sediment samples from a weakly buffered northern Wisconsin lake. All mercury
transformations were measured with radioisotopic tracers. Acidification of sediment pH with H2SO4, HCI, or
HNO3 significantly decreased 203Hg(II) methylation. Acidification of pH 6.1 (ambient) sediments to pH 4.5 witheither H2SO4 or HCI inhibited methylation by over 65%. The decreased methylation was due to the increasedhydrogen ion concentration because methylation was not affected by concentrations of Na2SO4 or NaCIequimolar to the amount of acid added. Inhibition of methylation was observed even after prolongedacidification of sediments to pH 5.0 for up to 74 days. Acidification of sediments to pH 5.5, 4.5, and 3.5 withHNO3 resulted in a near complete inhibition of methylation at each pH. Similarly, the addition of equimolaramounts of NaNO3 resulted in a near complete inhibition of methylation, indicating that the inhibition was dueto the nitrate ion rather than to the acidity. Demethylation of methyl mercury was not affected by pHs between8.0 and 4.4, but sharply decreased below pH 4.4. Volatilization of 203Hg(II) from surface sediments was lessthan 2% of methylation activity and was not significantly different from that in killed sediments. This studyindicated that acidification of sediments inhibits mercury methylation and that the observed increase in themercury burdens in fish from low pH lakes is not due to increased production of methylmercury in sediments.
Interest in mercury cycling has increased recently as aresult of to observations of elevated mercury concentrationsin fishes from naturally acidic and acidified lakes that lackdirect anthropogenic sources of mercury (1, 4, 19, 34, 37).Lindqvist et al. (19) estimated that 10,000 remote Swedishlakes contain mercury-contaminated fish. Most (>85%) ofthe total mercury in freshwater fish (14) and algae (30) is inthe methylmercury form, indicating that the formation ofmethylmercury is a key process regulating the mercurycontent of aquatic biota.Data on the effect of acidification on mercury methylation
in sediments are conflicting. Fagerstrom and Jernelov (10)observed optimum monomethylmercury formation in sedi-ments between pH 7 and 5 and decreased formation ofmethylmercury at higher pHs. Dimethylmercury was theprimary methylated species formed at higher pHs. This earlyobservation led many investigators (4-6) to assume thatincreased acidity enhances mercury methylation. However,Baker et al. (2) observed methylation in nutrient-enrichedsediments at pH 6.5 and 5.5, but not at pH 4.5 and 3.5, andno difference in methylation activity was observed in OttawaRiver sediments between pH 5 and 6 (20). More recently,Ramlal et al. (24) observed decreased mercury methylationin sediments with decreased pH in the range pH 7 to 4.5.The amount of methylmercury formed in an aquatic sys-
tem is a result of the concomitant processes of methylationand demethylation (31). Ramlal et al. (25) recently developedrapid isotopic methods to measure mercury demethylationand to compare methylation and demethylation activities.These methods have been used to compare relative rates of
* Corresponding author.t Present address: Department of Biology, University of Louis-
ville, Louisville, KY 40292.t Present address: Center for Lake Superior Environmental Stud-
ies, University of Wisconson-Superior, Superior, WI 54880.
methylation and demethylation in lakes in the CanadianShield (24, 38) and northern Wisconsin (17). These studiesdemonstrate that demethylation is an important factor af-fecting variations in net methylmercury formation in aquatichabitats.
Less is known about other components of the mercurycycle in aquatic environments, such as volatilization. Al-though little studied, volatilization could result in significantdecreases in the amount of mercury available for methyla-tion. Mercury volatilization may occur by the formation ofdimethylmercury or by the reduction of mercuric ion toelemental mercury (Hg0). The formation of dimethylmercuryoccurs largely at pHs greater than 7 (10), and volatile loss ofdimethylmercury would be insignificant at low pHs. Thereduction of mercuric ion to volatile Hg0 is the primarymercury-detoxifying mechanism used by mercury-resistantbacteria (28). These organisms may compose a large portionof the native aquatic bacterial population (21), suggestingthat reductive volatilization may be an important, but over-looked, aspect of mercury cycling in aquatic ecosystems.Mercury volatilization has been observed in freshwatersediments (18, 29), but the effect of acidification on mercuryvolatilization and its importance in acidic lakes has not beeninvestigated.The addition of sulfate to lakes may also affect mercury
transformations. Sulfate and hydrogen ion concentrations inacid precipitation are closely correlated (13); thus, an in-crease in acid precipitation will result in increased sulfateloading in acidified lakes. The production of hydrogen sulfideby sulfate-reducing bacteria in anaerobic sediments mayresult in the formation of highly insoluble HgS (Ksr = 4 x10-53), which is unavailable for methylation (10). In con-trast, sulfate-reducing bacteria have recently been impli-cated as agents of mercury methylation (7; M. R. Winfrey,Abstr. Annu. Meet. Am. Soc. Microbiol. 1985, Q79, p. 271),suggesting that the increased sulfate loadings associated with
2003
on May 3, 2019 by guest
http://aem.asm
.org/D
ownloaded from

2004 STEFFAN ET AL.
acidic precipitation may stimulate mercury methylation insediments.The mechanism responsible for the increased mercury
content of fish in remote acidic lakes is unknown, but likelyinvolves increased production of methylmercury. An in-crease in methylmercury production could result from in-creased methylation, but also from decreased demethyl-ation, or decreased volatile loss of mercury from the lake.However, no study has simultaneously examined these threeprocesses. We report here on the effect of acidification onmercury methylation, demethylation, and volatilization fromsediments from a mildly acidic, poorly buffered northernWisconsin lake. Methylation was decreased by acidificationwith H2SO4, HNO3, or HCl. Acidification to pH 4.5 had littleeffect on demethylation, and significant volatilization wasnot observed at any pH.
MATERIALS AND METHODS
Study area. Lake Clara is a mesotrophic, acid-susceptible,drained lake (9) in Lincoln County, Wis. It is one of severallakes that has a fish advisory from the Wisconsin Depart-ment of Natural Resources for mercury-contaminated fish(36). The lake has a maximum depth of 11.3 m and containshighly organic flocculent sediments. Chemical and physicalcharacteristics of the lake have been described by Korthalsand Winfrey (17) and Rada et al. (22).Sample collection. Flocculent surficial sediment and water
samples were collected with a peristaltic sampling pump(Horizon Ecology Co.) from the area of maximum depth.Sediments were transferred to and stored in N,-gassed glassbottles sealed with butyl rubber stoppers. All collection,manipulation, and incubation of sediments were performedby strict anaerobic technique (15, 35). Samples were trans-ported to the laboratory on ice and processed within 24 h.Chemical analysis. Lake water and sediment pH was
measured in the field with a Fisher model 107 pH meter.Dissolved oxygen and temperature were measured with aYellow Springs Instrument model 54A oxygen meter. In thelaboratory, pH was determined with an Orion model 701 pHmeter with a glass combination electrode (Corning GlassWorks, Corning, N.Y.).pH adjustments of sediments. Surficial sediment samples
(pH 6.1 to 6.4) were pooled in a 4-liter jar and mixed undera stream of O,-free N,. Subsamples (200 to 400 ml) weretransferred to 500-ml glass bottles and acidified to variouspH levels with 0.5 N H2SO4, 0.5 N HNO3, or 0.5 N HCI. ThepH readings were recorded when instrument drift decreasedto <0.01 unit/30 s. After acidification, samples were incu-bated at 22 + 2°C for 2 to 24 h and the pH was readjusted, asneeded, before further analysis. The pH of samples in timecourse experiments was monitored weekly and readjusted asneeded. Ion controls for acidification experiments wereprepared by adding 0.5 N Na,S04, 0.5 N NaNO3, or 0.5 NNaCl to subsamples of the pooled sediments. The sodiumsalts of the anion of each acid were added to give concen-trations equimolar to the amount of ion added in the corre-sponding acidified sediment sample.The pH of sediment subsamples was increased above the
ambient pH with the addition of 6 N NaOH and allowed toequilibrate for 2 h, and the pH was readjusted as necessary.Mercury methylation assay. Mercury methylation activity
was determined by incubating a 15-ml sediment sample with1.0 jig of 203Hg(II) [3.05 to 8.66 ,uCi as 2"3Hg(NO3)2; NewEngland Nuclear Corp., Boston, Mass.] in acid-washed,60-ml biochemical oxygen demand bottles. The experiments
shown in Fig. 2 and 3 were done by adding 2.0 jig of2"'3Hg(II). The additional amount of mercury did not alter theeffect of pH on methylation as demonstrated in repeatexperiments with 1.0 jig of 203Hg(II). In one experiment,203HgC1, was added to sediments to examine the effect of theaddition of nitrate when 203Hg(NO3)2 was added to sedi-ments.Samples were incubated for 24 or 48 h, and the experiment
was terminated with the addition of 1.0 ml of 6 N HCI. Allassays for methylation, demethylation, and volatilizationwere set up and incubated by strict anaerobic technique (15).All assays were incubated at 22 ± 2°C. The optimumtemperature for methylation was determined to be about35°C, but the lower temperature was used to approximatemore closely the in situ temperature of the profundal lakesediments (2 to 12°C).Methylmercury formed during incubation was extracted
by the method developed by Furutani and Rudd (11) asmodified by Korthals and Winfrey (17). The radiolabeledmethylmercury in the final benzene extract was counted in 4ml of 4a20 scintillation cocktail (Research Products Interna-tional Corp.).Mercury demethylation assay. Demethylation activity in
sediments was determined by incubating 30-mI sedimentsamples with 1.0 ,ug of Hg(II) (as "4CH HgI; AmershamCorp., Arlington Heights, Ill.) in rubber-stoppered 125-mlacid-washed glass bottles. Samples were incubated at 22 ±2°C for 24 h, and the experiment was terminated by injecting2 ml of 6 N HCI through the butyl rubber stoppers with asyringe.
Products of 14CH3HgI demethylation (14CO2 and 14CH4)were quantified by the method developed by Ramlal et al.(25) as modified by Korthals and Winfrey (17). The 14CO,produced during incubation was captured in a series oftwo vials containing 8 ml of a 14CO -trapping cocktail(14CO,UNT-SORB; Research Products InternationalCorp.). The "'CH4 produced during incubation passedthrough the first series of traps and was oxidized to 14C02 ina tube furnace (Sargent Welch) containing hot cupric oxide(J. T. Baker Chemical Co., Phillipsburg, N.J.). The resultant"'CO, was then trapped in a second series of two vialscontaining the 14CO2-trapping cocktail. The radioactivity ineach vial was quantified by liquid scintillation counting.Mercury volatilization assay. Volatilization assays were
performed by incubating acidified and nonacidified sedi-ments with 1.0 ,ug of 213Hg(II) [as 203Hg(N03)2] in a 60-mlscrew-cap Erlenmeyer flask with a Teflon stopcock attachedto the side (Fig. 1). Each flask was capped with a TeflonMininert cap (Supelco, Inc., Bellefonte, Pa.) and gassedcontinuously with 02-free N,. The outlet tube of the stop-cock was attached to two scintillation vial traps with Teflontubing. Each trap was filled with 4 ml of trapping solution(1.5% HgBr and 10% KBr), which effectively traps volatileHg compounds (29). This trapped more than 95% of thevolatile Hg evolved from the samples. The volatile 203Hg('formed was quantified by counting the trap solution in 8.0 mlof PCS gelling scintillation cocktail (Amersham Corp.). Thetrapped mercury was assumed to be elemental mercury,because no radioactivity was detected in solvent extractionsof mercury traps (as described above).The radioisotopic techniques used in this study do not
provide in situ rates of methylation, demethylation, orvolatilization because of the addition of exogenous radiola-beled substrate. They do, however, provide useful estimatesof potential rates and may be used to compare relative ratesbetween different sites or conditions.
APPL. ENVIRON. MICROBIOL.
on May 3, 2019 by guest
http://aem.asm
.org/D
ownloaded from

MERCURY TRANSFORMATIONS IN ACIDIFIED SEDIMENTS
GAS SUPPLY
TEFLON MININERT
SEDIMENT OR WATERSAMPLE
FIG. 1. Apparatus used to measure mercury volatilization with 203Hg(II).
Quality assurance. Quality assurance for this study con-sisted of killed controls, procedural blanks, and standardadditions. Killed controls were prepared by autoclavingsamples or by adding 6 N HCI, 6 N NaOH, or Formalin priorto the addition of isotope. Procedural blanks were preparedby the addition of radioisotope [213Hg(NO3), or '4CH3HgIIto sterile deionized water. The procedural blanks wereacidified, and the methylation and demethylation assayswere conducted as described above. Methylation and de-methylation activities were not detected in killed controls orprocedural blanks.
Standard additions for the methylation assay were pre-pared by adding a known quantity (disintegrations perminute) of 14CH3HgI to acidified samples, and the 14CH3HgIwas extracted as described above. The standard additionsfor the demethylation assay were prepared by adding aknown amount (disintegrations per minute) of NaH'4CO0(New England Nuclear Corp.) to samples prior to acidifica-tion. Sample bottles were immediately stoppered and acidi-fied with 2 ml of 6 N HCI. Acidification lowered the pHsufficiently to convert all H14C0- to 14CO . The 14CO, wasgassed from the reaction bottles and captured as describedabove. Extraction efficiencies of the methylation and de-methylation assays were essentially 100% (17).
Reagents were routinely checked for radioactive contam-ination by scintillation counting. All radioisotope countingwas done on a Beckman LS 230 liquid scintillation counter.Counts per minute were corrected for the counter efficiencyfrom previously prepared quench curves by using externalstandardization, and disintegrations per minute were used inall calculations.
Chemicals and radioisotopes. All chemicals used werereagent grade except for benzene, which was certified grade.Stock solutions of 203Hg(NO3) in 0.5 M HNO, and 203HgCl,in 0.5 M HCI (New England Nuclear Corp.) were preparedby dilution in distilled water to yield a final concentration of0.10 [Lg of Hg per Il. The 14CH HgI (Amersham Corp.) was
kept at -60°C, and working stock solutions were preparedby dilution in distilled water. Stock solutions of NaH14CO3(New England Nuclear Corp.) were prepared by dilution indistilled water.
Data analyses. All assays were conducted in triplicate andeach experiment was repeated at least once to ensurereproducible results. Results of the assays were expressed asthe percentage of total added mercury methylated, demethyl-ated, or volatilized. Significant differences between means ofdata were evaluated by the Student t test when two meanswere compared. If more than two means were compared,data were analyzed by one-way analysis of variance andStudent-Newman-Keuls multiple comparison tests.
RESULTS
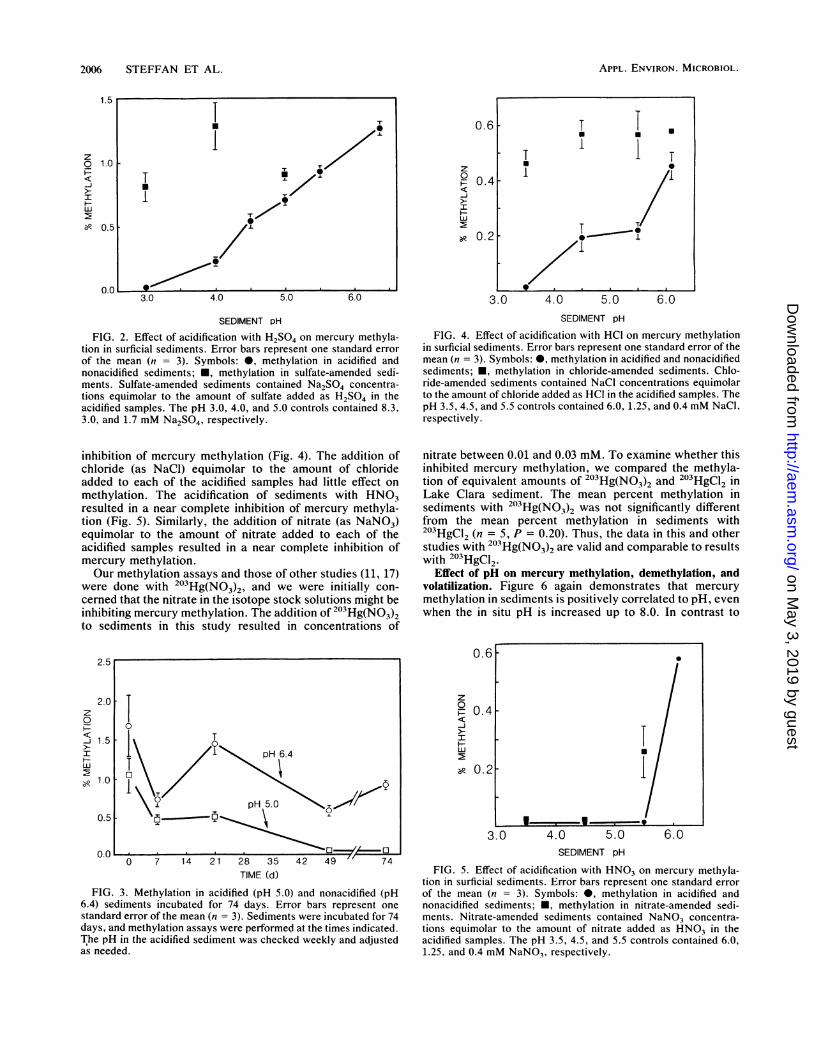
Effect of acid and anion addition on mercury methylation.Acidification of sediments with H2SO4 significantly de-creased mercury methylation activity (Fig. 2). Acidificationof sediments to pH 4.5 with H2S04 decreased methylationactivity by greater than 65%, and the degree of inhibitionwas proportional to the sediment pH between pH 3 and 6.4(ambient). Little effect, however, was observed when addingsulfate (as Na2SO4) equimolar to the amount of sulfate addedto each of the acidified samples. The amount of methylationin samples with 0.4 and 6.0 mM sulfate added (correspondingto pH 5.0 and 3.0, respectively) was significantly less thanthe amount of methylation in sediments without addedsulfate. However, this was not observed in replicate sulfateaddition experiments.
Methylation activity in sediments acidified to pH 5.0 withH.S04, assayed at 7, 21, 49, and 74 days after acidification,did not increase (Fig. 3). In contrast, the amount of methyl-ation in acidified sediments decreased significantly thelonger the sediments had been acidified, while methylationactivity in nonacidified sediments changed little.
Acidification of sediments with HCI resulted in a similar
Vot- 54, 1988 2005
on May 3, 2019 by guest
http://aem.asm
.org/D
ownloaded from
2006 STEFFAN ET AL.
1.5
z
0
7-I
LUill
1O
SEDIMENT pH
FIG. 2. Effect of acidification with H2S04 on mercury methyla-tion in surficial sediments. Error bars represent one standard errorof the mean (n = 3). Symbols: *, methylation in acidified andnonacidified sediments; U, methylation in sulfate-amended sedi-ments. Sulfate-amended sediments contained Na2SO4 concentra-tions equimolar to the amount of sulfate added as H2SO4 in theacidified samples. The pH 3.0, 4.0, and 5.0 controls contained 8.3,3.0, and 1.7 mM Na2SO4, respectively.
inhibition of mercury methylation (Fig. 4). The addition ofchloride (as NaCl) equimolar to the amount of chlorideadded to each of the acidified samples had little effect onmethylation. The acidification of sediments with HNO3resulted in a near complete inhibition of mercury methyla-tion (Fig. 5). Similarly, the addition of nitrate (as NaNO3)equimolar to the amount of nitrate added to each of theacidified samples resulted in a near complete inhibition ofmercury methylation.Our methylation assays and those of other studies (11, 17)
were done with 203Hg(NO3)2, and we were initially con-cerned that the nitrate in the isotope stock solutions might beinhibiting mercury methylation. The addition of 203Hg(NO3)2to sediments in this study resulted in concentrations of
2.5
z
0
7-
LUi
2.0 [
1.51-
1.0
0.51-
0.00 7 14 2 1 28 35
TIME (d)42 49 " 74
FIG. 3. Methylation in acidified (pH 5.0) and nonacidified (pH6.4) sediments incubated for 74 days. Error bars represent onestandard error of the mean (n = 3). Sediments were incubated for 74days, and methylation assays were performed at the times indicated.The pH in the acidified sediment was checked weekly and adjustedas needed.
0.61
z
0
-I
LU
0.41
0.2 [
3.0 4.0 5.0 6.0
SEDIMENT pH
FIG. 4. Effect of acidification with HCI on mercury methylationin surficial sediments. Error bars represent one standard error of themean (n = 3). Symbols: 0, methylation in acidified and nonacidifiedsediments; *, methylation in chloride-amended sediments. Chlo-ride-amended sediments contained NaCI concentrations equimolarto the amount of chloride added as HCI in the acidified samples. ThepH 3.5, 4.5, and 5.5 controls contained 6.0, 1.25, and 0.4 mM NaCl,respectively.
nitrate between 0.01 and 0.03 mM. To examine whether thisinhibited mercury methylation, we compared the methyla-tion of equivalent amounts of 2'3Hg(NO3)2 and 203HgCl2 inLake Clara sediment. The mean percent methylation insediments with 203Hg(NO3)2 was not significantly differentfrom the mean percent methylation in sediments with203HgCl2 (n = 5, P = 0.20). Thus, the data in this and otherstudies with 2033Hg(NO3)2 are valid and comparable to resultswith 203HgC12.
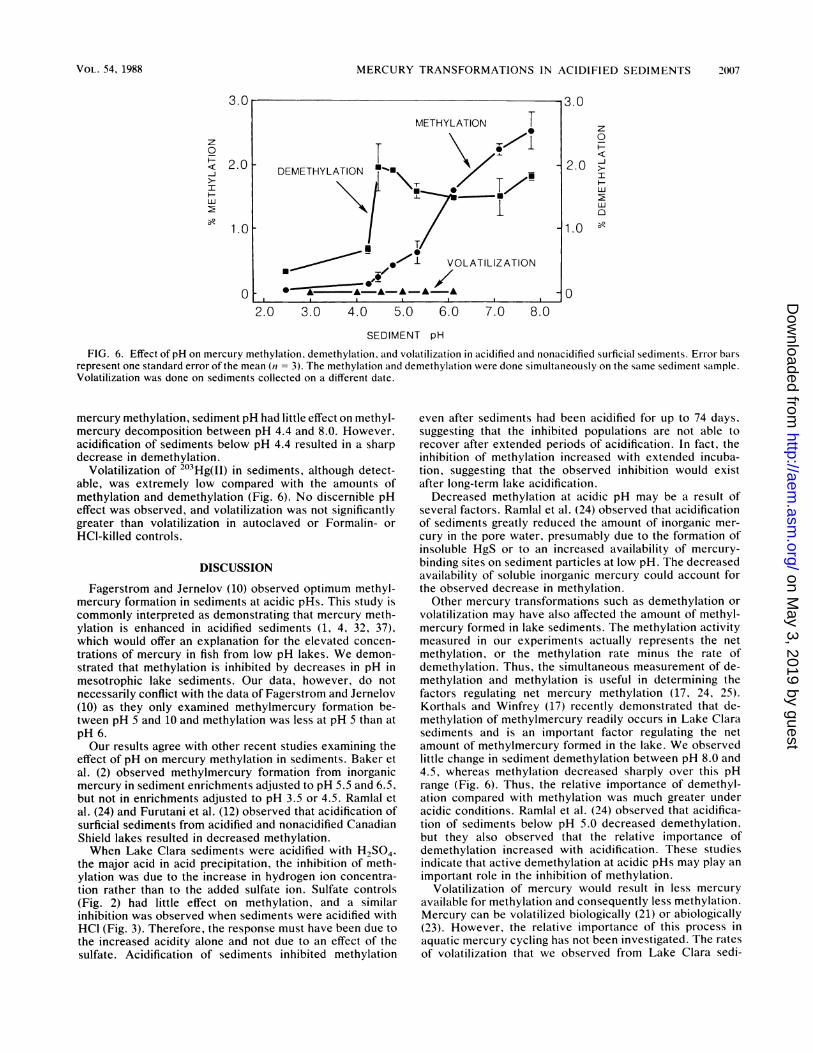
Effect of pH on mercury methylation, demethylation, andvolatilization. Figure 6 again demonstrates that mercurymethylation in sediments is positively correlated to pH, evenwhen the in situ pH is increased up to 8.0. In contrast to
0.6
z
0
7-
ILHo
0.4
0.2
3.0 4.0 5.0 6.0
SEDIMENT pH
FIG. 5. Effect of acidification with HNO3 on mercury methyla-tion in surficial sediments. Error bars represent one standard errorof the mean (n = 3). Symbols: 0, methylation in acidified andnonacidified sediments; *, methylation in nitrate-amended sedi-ments. Nitrate-amended sediments contained NaNO3 concentra-tions equimolar to the amount of nitrate added as HNO3 in theacidified samples. The pH 3.5, 4.5, and 5.5 controls contained 6.0,1.25, and 0.4 mM NaNO,, respectively.
T Ia
TI I
*
T TTII
0
T
APPL. ENVIRON. MICROBIOL.
. o- {t'l o
on May 3, 2019 by guest
http://aem.asm
.org/D
ownloaded from
MERCURY TRANSFORMATIONS IN ACIDIFIED SEDIMENTS
2.0
'Tia
METHYLATION
DEMETHYLATION
1 VOLATILIZATION
AA-A-A-AI a I_ I*
2.0 3.0 4.0 5.0 6.0 7.0 8.0
SEDIMENT pH
FIG. 6. Effect of pH on mercury methylation, demethylation, and volatilization in acidified and nonacidified surficial sediments. Error barsrepresent one standard error of the mean (n = 3). The methylation and demethylation were done simultaneously on the same sediment sample.Volatilization was done on sediments collected on a different date.
mercury methylation, sediment pH had little effect on methyl-mercury decomposition between pH 4.4 and 8.0. However,acidification of sediments below pH 4.4 resulted in a sharpdecrease in demethylation.
Volatilization of 203Hg(II) in sediments, although detect-able, was extremely low compared with the amounts ofmethylation and demethylation (Fig. 6). No discernible pHeffect was observed, and volatilization was not significantlygreater than volatilization in autoclaved or Formalin- or
HCI-killed controls.
DISCUSSION
Fagerstrom and Jernelov (10) observed optimum methyl-mercury formation in sediments at acidic pHs. This study is
commonly interpreted as demonstrating that mercury meth-ylation is enhanced in acidified sediments (1, 4, 32, 37),which would offer an explanation for the elevated concen-
trations of mercury in fish from low pH lakes. We demon-strated that methylation is inhibited by decreases in pH inmesotrophic lake sediments. Our data, however, do notnecessarily conflict with the data of Fagerstrom and Jernelov(10) as they only examined methylmercury formation be-tween pH 5 and 10 and methylation was less at pH 5 than atpH 6.Our results agree with other recent studies examining the
effect of pH on mercury methylation in sediments. Baker etal. (2) observed methylmercury formation from inorganicmercury in sediment enrichments adjusted to pH 5.5 and 6.5,but not in enrichments adjusted to pH 3.5 or 4.5. Ramlal etal. (24) and Furutani et al. (12) observed that acidification ofsurficial sediments from acidified and nonacidified CanadianShield lakes resulted in decreased methylation.When Lake Clara sediments were acidified with H,S04,
the major acid in acid precipitation, the inhibition of meth-ylation was due to the increase in hydrogen ion concentra-tion rather than to the added sulfate ion. Sulfate controls(Fig. 2) had little effect on methylation, and a similarinhibition was observed when sediments were acidified withHCI (Fig. 3). Therefore, the response must have been due tothe increased acidity alone and not due to an effect of thesulfate. Acidification of sediments inhibited methylation
even after sediments had been acidified for up to 74 days,suggesting that the inhibited populations are not able torecover after extended periods of acidification. In fact, theinhibition of methylation increased with extended incuba-tion, suggesting that the observed inhibition would existafter long-term lake acidification.Decreased methylation at acidic pH may be a result of
several factors. Ramlal et al. (24) observed that acidificationof sediments greatly reduced the amount of inorganic mer-
cury in the pore water, presumably due to the formation ofinsoluble HgS or to an increased availability of mercury-binding sites on sediment particles at low pH. The decreasedavailability of soluble inorganic mercury could account forthe observed decrease in methylation.Other mercury transformations such as demethylation or
volatilization may have also affected the amount of methyl-mercury formed in lake sediments. The methylation activitymeasured in our experiments actually represents the netmethylation, or the methylation rate minus the rate ofdemethylation. Thus, the simultaneous measurement of de-methylation and methylation is useful in determining thefactors regulating net mercury methylation (17, 24, 25).Korthals and Winfrey (17) recently demonstrated that de-methylation of methylmercury readily occurs in Lake Clarasediments and is an important factor regulating the netamount of methylmercury formed in the lake. We observedlittle change in sediment demethylation between pH 8.0 and4.5, whereas methylation decreased sharply over this pHrange (Fig. 6). Thus, the relative importance of demethyl-ation compared with methylation was much greater underacidic conditions. Ramlal et al. (24) observed that acidifica-tion of sediments below pH 5.0 decreased demethylation,but they also observed that the relative importance ofdemethylation increased with acidification. These studiesindicate that active demethylation at acidic pHs may play an
important role in the inhibition of methylation.Volatilization of mercury would result in less mercury
available for methylation and consequently less methylation.Mercury can be volatilized biologically (21) or abiologically(23). However, the relative importance of this process inaquatic mercury cycling has not been investigated. The ratesof volatilization that we observed from Lake Clara sedi-
3.0
z
0
-J
I
LUII
0o
1.0
0
z0
2.0 >I
LL
1.0 g
0
VOL. 54, 1988 2007
on May 3, 2019 by guest
http://aem.asm
.org/D
ownloaded from

2008 STEFFAN ET AL.
ments were less than 2% of the rates of mercury methylationand demethylation at the ambient pH, and volatilization wasnot dependent on sediment pH. This implies that volatiliza-tion of mercury from sediments is not a significant processaffecting the availability of mercury.
This was surprising as bacteria capable of volatilizingmercury are abundant in sediment (21; M. R. Winfrey, W. J.Pekarske, and R. J. Steffan, Abstr. Annu. Meet. Am. Soc.Microbiol. 1986, Q124, p. 304). However, the mercury-volatilizing bacteria must not be active in our sedimentssince the small amounts of volatilization that we observedwere primarily abiological. The low rates of volatilizationmay be due to the tendency of mercury to bind readily tosediment particles and organic matter, which may render itunavailable for volatilization.The observation that sulfate had little effect on mercury
methylation in Lake Clara sediment indicates that the sulfideformed from the enhanced sulfate reduction did not precip-itate the mercury and render it unavailable. Inhibition ofmethylation by the formation of insoluble mercuric sulfidehas been suggested by previous workers (3, 16, 33, 39).Recently, sulfate-reducing bacteria have been implicated as
agents of mercury methylation in freshwater (Winfrey,Abstr. Annu. Meet. Am. Soc. Microbiol. 1985) and marine(7) sediments. However, the addition of sulfate did notstimulate methylation, although sulfate reduction wouldhave been markedly increased in the sulfate-depleted sedi-ments. These conflicting data suggest that further researchneeds to be done to clarify the role and relative importanceof sulfate-reducing bacteria in mercury methylation.
Nitric acid is the second most abundant acid in acidprecipitation and is thus part of the acid burden added tolakes receiving acid deposition. The effect of HNO3 additionon mercury methylation in sediments was markedly differentfrom the effect observed with H2SO4 (Fig. 5). The nitratecontrols, however, indicated that the near complete inhibi-tion of methylation by HNO3 was due to an inhibitory effectcaused by the nitrate ion, and not from the acidity. It isunlikely that nitrate would actually affect mercury methyla-tion in situ. The final concentrations of added nitrate thatinhibited methylation in our experiments ranged from 0.4 to6.0 mM. However, the addition of 0.03 mM nitrate inexperiments in which mercury was added as 203Hg(NO3)2did not inhibit methylation. This amount is far above thelevel of nitrate in sediment, which is generally undetectable(<0.001 mM).The results of this study and other recent data (12, 24)
demonstrate that enhanced methylation activity in lakesediments cannot account for the elevated mercury concen-trations observed in fish from low pH lakes. Therefore, othermechanisms such as altered gill permeability in fish (8, 27) oraltered partitioning of methylmercury between sediment andwater at low pH (20) may account for this correlation.Alternatively, Xun et al. (38) have recently demonstratedthat decreased pH in Canadian Shield lakes results in in-creased methylation in the water column. Methylation activ-ity in the water of lakes is low (11, 17, 38), and methylationis generally assumed to occur in the sediments (26). How-ever, water column methylation may play a relatively moreimportant role due to the greater volume of water comparedwith that of sediments. Thus, mercury methylation in thewater column of lakes may be an important, yet oftenoverlooked process regulating the bioavailability of mercuryin lakes. Future studies involving the cycling of mercury inthe water of lakes are needed if we are to understand fully
the factors regulating the biological availability of this toxiccompound.
ACKNOWLEDGMENTS
This study was supported by the Department of Biology andMicrobiology and the River Studies Center at the University ofWisconsin-La Crosse.We are grateful to J. W. M. Rudd of the Freshwater Institute,
Winnipeg, Manitoba, Canada, for providing the radiolabeled methylmercury. Bill Pekarske, Greg Cope, Jim Rogala, Janet Winfrey, PaulRitter, Lori Rislove, and Kristi Honer provided assistance withcollection and processing of samples. We also thank R. G. Rada andT. F. Weeks for their valuable criticism of the manuscript.
LITERATURE CITED1. Akielazek, J. J., and T. A. Haines. 1981. Mercury in the muscle
tissue of fish from three northern Maine lakes. Bull. Environ.Contam. Toxicol. 27:201-208.
2. Baker, M. D., W. E. Inniss, I. Mayfield, P. T. S. Wong, andY. K. Chau. 1983. Effect of pH on the methylation of mercuryand arsenic by sediment microorganisms. Environ. Technol.Lett. 4:89-100.
3. Berman, M., and R. Bartha. 1986. Control of the methylationprocess in a mercury-polluted aquatic environment. Environ.Pollut. Ser. B 11:41-53.
4. Bjorkland, I., H. Borg, and K. Johansson. 1984. Mercury inSwedish lakes-its regional distribution and causes. Ambio 13:119-121.
5. Brouzes, R. J. P., R. A. N. McLean, and G. H. Tomlinson. 1977.The link between pH of natural waters and the mercury contentof fish. Research Report. Dromtar Research Centre, Senneville,Quebec, Canada.
6. Clarkson, T. W., R. Hamaday, and L. Amin-Zaki. 1984. Mer-cury, p. 285-309. In J. 0. Nriagu (ed.), Changing metal cyclesand human health. Springer-Verlag, New York.
7. Compeau, G., and R. Bartha. 1985. Sulfate-reducing bacteria:principle methylators of mercury in anoxic estuarine sediments.Appl. Environ. Microbiol. 48:1203-1207.
8. Drummond, R. A., G. F. Olson, and A. R. Batterman. 1974.Cough response and uptake of mercury by brook trout, Salve-linus fontinalis, exposed to mercuric compounds at differenthydrogen-ion concentrations. Trans. Am. Fish. Soc. 103:224-249.
9. Eilers, J., G. Glass, K. Webster, and J. A. Rogalla. 1983.Hydrologic control of lake susceptibility to acidification. Can. J.Fish. Aquat. Sci. 40:1896-1904.
10. Fagerstrom, T., and A. Jernelov. 1972. Some aspects of thequantitative ecology of mercury. Water Res. 6:1193-1202.
11. Furutani, A., and J. W. M. Rudd. 1980. Measurement ofmercury methylation in lake water and sediment samples. Appl.Environ. Microbiol. 40:770-776.
12. Furutani, A., J. W. M. Rudd, and C. A. Kelly. 1984. A methodfor measuring the response of sediment microbial communitiesto environmental perturbations. Can. J. Microbiol. 30:1408-1414.
13. Gorham, E., F. B. Martin, and J. T. Litzau. 1984. Acid rain:ionic correlations in the eastern United States, 1980-1981.Science 225:407-409.
14. Huckabee, J. W., J. W. Elwood, and S. G. Hildebrand. 1979.Accumulation of mercury in freshwater biota, p. 277-302. InJ. 0. Nriagu (ed.), The biogeochemistry of mercury in theenvironment. Elsevier/North-Holland Biomedical Press, NewYork.
15. Hungate, R. E. 1969. A roll tube method for cultivation of strictanaerobes. Methods Microbiol. 3B:117-137.
16. Jacobs, L. W., and D. R. Keeney. 1974. Methylmercury forma-tion in mercury-treated river sediments during in situ equilibra-tion. J. Environ. Qual. 3:121-126.
17. Korthals, E. T., and M. R. Winfrey. 1987. Seasonal and spatialvariations in mercury methylation and demethylation in anoligotrophic lake. Appl. Environ. Microbiol. 53:2397-2404.
18. Kudo, A., D. R. Miller, H. Akagi, D. C. Mortimer, A. S.
APPL. ENVIRON. MICROBIOL.
on May 3, 2019 by guest
http://aem.asm
.org/D
ownloaded from

MERCURY TRANSFORMATIONS IN ACIDIFIED SEDIMENTS
DeFreitas, H. Nagasse, D. R. Townsend, and R. G. Warnock.1978. The role of sediments on mercury transport (total andmethyl) in a river system. Prog. Water Technol. 10:329-339.
19. Lindqvist, O., A. Jernelov, K. Johansson, and H. Rodhe. 1984.Mercury in the Swedish environment. Global and local sources.Report snv pm 1816. National Swedish Environment ProtectionBoard, Solna, Sweden.
20. Miller, D. R., and H. Akagi. 1979. pH affects mercury distribu-tion, not methylation. Ecotoxicol. Environ. Saf. 3:36-38.
21. Nelson, J. D., and R. R. Colwell. 1975. The ecology of mercury-resistant bacteria in Chesapeake Bay. Microb. Ecol. 1:191-218.
22. Rada, R. G., M. R. Winfrey, J. G. Wiener, and D. E. Powell.1987. A comparison of mercury distribution in sediment coresand mercury volatilization from surface waters of selectednorthern Wisconsin lakes. Completion report to the WisconsinDepartment of Natural Resources, Bureau of Water ResourcesManagement, Surface Water Standards and Monitoring Section,Madison, Wis.
23. Ramamoorthy, S., T. C. Cheng, and D. J. Kushner. 1983.Mercury speciation in water. Can. J. Fish. Aquat. Sci. 40:85-89.
24. Ramlal, P. S., J. W. M. Rudd, A. Furutani, and L. Xun. 1985.The effect of pH on methyl mercury production and decompo-sition in lake sediments. Can. J. Fish. Aquat. Sci. 42:685-692.
25. Ramlal, P. S., J. W. M. Rudd, and R. E. Hecky. 1986. Methodsfor measuring specific rates of mercury methylation and degra-dation and their use in determining factors controlling net ratesof mercury methylation. Appl. Environ. Microbiol. 51:110-114.
26. Robinson, J. B., and 0. H. Tuovinen. 1984. Mechanisms ofmicrobial resistance and detoxification of organomercury com-pounds: physiological, biochemical, and genetic analysis. Mi-crobiol. Rev. 48:95-124.
27. Rodgers, D. W., and F. W. H. Beamish. 1983. Water qualitymodifies uptake of water borne methyl mercury in rainbowtrout, Salmo gairdneri. Can. J. Fish. Aquat. Sci. 40:824-828.
28. Silver, S. 1984. Bacterial transformations of and resistance toheavy metals, p. 199-223. In J. 0. Nriagu (ed.), Changing metalcycles and human health. Springer-Verlag, New York.
29. Spangler, W. J., J. L. Spigarelli, J. M. Rose, and H. M. Miller.
1973. Methylmercury: bacterial degradation in lake sediments.Science 180:192-193.
30. Stokes, P. M., S. I. Dreier, M. 0. Farkas, and R. A. N. McLean.1983. Mercury accumulation by filamentous algae: a promisingbiological monitoring system for methylmercury in acid stressedlakes. Environ. Pollut. Ser. B 5:255-271.
31. Summers, A. O., and S. Silver. 1978. Microbial transformationsof metals. Annu. Rev. Microbiol. 36:637-672.
32. Suns, K., C. Curry, and D. Russel. 1980. The effects of waterquality and morphometric parameters on mercury uptake byyearling yellow perch. Technical report LTS 80-1. OntarioMinistry of the Environment, Rexdale, Ontario, Canada.
33. Tonomura, K., F. Furukawa, and M. Yamada. 1972. Microbialconversion of mercury compounds, p. 115-133. In F. Matsu-mura, G. M. M. Boush, and T. Misato (ed.), Environmentaltoxicology of pesticides. Academic Press, Inc., New York.
34. Wiener, J. E. 1983. Comparative analyses of fish populations innaturally acidic and circumneutral lakes in Northern Wisconsin.FWS/OBS-80/40.16. Eastern Energy and Land Use Team, U.S.Fish and Wildlife Service, Washington, D.C.
35. Winfrey, M. R., D. R. Nelson, S. C. Klevickis, and J. G. Zeikus.1977. Association of hydrogen metabolism with methanogenesisin Lake Mendota sediments. Appl. Environ. Microbiol. 33:312-318.
36. Wisconsin Department of Natural Resources. 1986. Wisconsin'shealth advisory for mercury-contaminated fish. Wisconsin De-partment of Natural Resources, Madison.
37. Wren, C. D., and H. R. MacCrimmon. 1983. Mercury levels inthe sunfish, Lepomis gibbosus, relative to pH and other envi-ronmental variables of Precambrian Shield lakes. Can. J. Fish.Aquat. Sci. 40:1737-1744.
38. Xun, L., N. E. R. Campbell, and J. W. M. Rudd. 1987.Measurement of specific rates of net methyl mercury productionin the water column and surface sediments of acidified andcircumneutral lakes. Can. J. Fish. Aquat. Sci. 44:750-757.
39. Yamada, M., and K. Tonomura. 1972. Microbial methylation ofmercury in hydrogen sulfide evolving environments. J. Fer-ment. Technol. 50:901-909.
VOL. 54, 1988 2009
on May 3, 2019 by guest
http://aem.asm
.org/D
ownloaded from