enhanced mutability associated with a temperature- - journal of
TRANSCRIPT

JOURNAL OF VIROLOGY, Aug. 1981, p. 377-3890022-538X/81/080377-13$02.00/0
Vol. 39, No. 2
Enhanced Mutability Associated with a Temperature-Sensitive Mutant of Vesicular Stomatitis Virus
C. R. PRINGLE,* V. DEVINE, M. WILKIE, C. M. PRESTON, A. DOLAN, AND D. J. McGEOCHMedical Research Council Virology Unit, Institute of Virology, Glasgow Gll 5JR, Scotland
Received 3 March 1981/Accepted 28 April 1981
Temperature-sensitive (ts) mutant tsDl of vesicular stomatitis virus, NewJersey serotype, is the sole representative of complementation group D. Clonesderived from this mutant exhibited three different phenotypes with respect toelectrophoretic mobility of the G and N polypeptides of the virion in sodiumdodecyl sulfate-polyacrylamide gel. Analysis of non-ts pseudorevertants showedthat none of the three phenotypes was associated with the temperature sensitivityof mutant tsDl. Additional phenotypes, some also involving the NS polypeptide,appeared during sequential cloning, indicating that mutations were generated athigh frequency during replication of tsDl. Furthermore, mutations altering theelectrophoretic mobility of the G, N, NS, and M polypeptides were induced inheterologous viruses multiplying in the same cells as tsDl. These heterologousviruses included another complementing ts mutant of vesicular stomatitis virusNew Jersey and ts mutants of vesicular stomatitis virus Indiana and Chandipuravirus. Complete or incomplete virions of tsDl appeared to be equally efficientinducers of mutations in heterologous viruses. Analysis of the progeny of a mixedinfection of two complementing ts mutants of vesicular stomatitis virus NewJersey with electrophoretically distinguishable G, N, NS, and M proteins yieldedno recombinants and excluded recombination as a factor in the generation of theelectrophoretic mobility variants. In vitro translation of total cytoplasmic RNAfrom BHK cells indicated that post-translational processing was not responsiblefor the aberrant electrophoretic mobility of the N, NS, and M protein mutants.Aberrant glycosylation could account for three of four G protein mutants, how-ever. Some clones of tsDl had an N polypeptide which migrated faster in sodiumdodecyl sulfate-polyacrylamide gel than did the wild type, suggesting that thepolypeptide might be shorter by about 10 amino acids. Determination of thenucleotide sequence to about 200 residues from each terminus of the N gene ofone of these clones, a revertant, and the wild-type parent revealed no changescompatible with synthesis of a shorter polypeptide by premature termination orlate initiation of translation. The sequence data indicated, however, that the N-protein mutant and its revertant differed from the parental wild type in two ofthe 399 nucleotides determined. These sequencing results and the phenomenonof enhanced mutability associated with mutant tsDl reveal that rapid andextensive evolution of the viral genome can occur during the course of normalcytolytic infection of cultured cells.
Rapid and progressive mutation of a viralgenome, which occurred during the course of anoncytopathic persistent infection of BHK-21cells, has been described by Holland et al. (9).This persistent infection was initiated by a tem-perature-sensitive (ts) mutant [tsG31(III)] oftheIndiana serotype of vesicular stomatitis virus(VSV) in the presence of its homologous defec-tive interfering (DI) particle. Numerous oligon-ucleotide map changes in viral RNA accumu-lated in the genome of the virus released fromthese cells during continuous propagation for 5years, in marked contrast to the apparent ge-
netic stability of virus propagated by cycles oflytic infection (2). The virus released after 5years exhibited a number of peptide changes inthe G (glycosylated spike), N (nucleocapsid),and M (non-glycosylated membrane) proteins,indicating that mutational change occurredthroughout the coding regions of the genome.Sequencing of the extremities of the genomerevealed that there were numerous base substi-tutions (7 of the first 46 nucleotides) in thenoncoding 5'-terminal region (25), whereas atthe 3'-terminus there was one base substitutionin the noncoding leader region and one in the
377
Dow
nloa
ded
from
http
s://j
ourn
als.
asm
.org
/jour
nal/j
vi o
n 28
Dec
embe
r 20
21 b
y 11
2.16
5.86
.196
.

378 PRINGLE ET AL.
three-nucleotide junction between the leader se-quence and the N gene (24). Thus, the mutationsgenerated during persistent infection were notrestricted to any specific region of the genome.These observations suggested that persistent in-fection could be an important source of biologi-cal variation in nature.High mutability of VSV Indiana has been
observed also during propagation of the virus ininsect (Drosophila melanogaster) cells. In thisinstance, enhanced mutability was indicated bythe accumulation of variants with multiple phe-notypic changes during sequential passage (14).The absence of cytopathogenicity in persistentlyinfected mammalian cells and in insect cells mayhave been a relevant factor in the enhancedmutability of VSV Indiana. However, the lowtemperature of incubation and the differentcomposition of the culture medium may havecontributed to the apparent genetic instabilityof VSV Indiana when the virus was propagatedin insect cells.We now describe an instance of high mutabil-
ity associated with a ts mutant ofthe New Jerseyserotype of VSV, which illustrates that highmutability can exist under conditions of lyticinfection and that it is not a phenomenonuniquely associated with noncytopathic infec-tion.Mutant tsDl is the sole representative ofcom-
plementation group D of VSV New Jersey (20).This mutant exhibited an RNA-positive pheno-type, and intracellular viral polypeptide synthe-sis was not inhibited at restrictive temperature(26). However, the virions of tsDl had two elec-trophoretically atypical proteins: both the G andthe N polypeptides of mutant tsDl migratedfaster in sodium dodecyl sulfate (SDS)-poly-acrylamide gel than did the corresponding poly-peptides of wild-type virus. Reversion of the tsphenotype was accompanied by restoration ofnormal N mobility, and none of 10 non-ts re-vertants showed any change in G mobility (27).Consequently, it was concluded that comple-mentation group D represented the N-proteingene, an assignment at variance with expectation(21).
Reinvestigation of the phenotype of mutanttsDl has established that the tsDl stock is het-erogeneous and that temperature sensitivity isnot associated with either the G- or the N-mo-bility difference. A characteristic ofmutant tsDlwas the frequent appearance, during sequentialcloning, of electrophoretically atypical polypep-tides. Furthermore, comparable mutations couldbe induced in other viruses during coinfectionwith mutant tsDl. Thus, enhanced mutabilityappeared to be a property of mutant tsDl.
(A partial account of these findings was pre-sented at the Fourth International NegativeStrand Virus Meeting, St. Thomas, V.I., October28-November 1 1980.)
MATERIALS AND METHODSCells. BHK-21 cells were used for growth and ra-
diolabeling of virus stocks. Cloning and assay of infec-tivity were carried out with BHK-21 cells and, occa-sionally, BS-C-1 cells.
Virus. The origin of the wild-type and ts mutantsof VSV New Jersey (Missouri strain) has been de-scribed previously (20). Mutant tsDl is an RNA-posi-tive mutant classified in complementation group D; itis the only member of the group. Mutant tsE3 is anRNA-negative mutant with an NS protein defect (5),classified in complementation group E.Mutant tsG114(I) is a thermolabile transcriptase
mutant of VSV Indiana (19, 21), and tsCh851(ChV) isan RNA-positive M-protein mutant of Chandipuravirus (6, 7).
Isolation of clones. Clones were established fromsingle, well-separated plaques on BHK-21 cell mono-layers under agar overlay. An agar plug and underlyingcells were removed by means of a Pasteur pipette andinoculated into a BHK-21 cell monolayer in a 30-mlbottle to establish a stock. The ts and polypeptidemobility phenotypes of each clone were determinedfrom this material.
Purification of virion and DI particles of tsDl.Approximately 5 x 108 BHK-21 cells as monolayers inrotating bottle cultures were infected at a multiplicityof 1 PFU per cell with virus from the original stock ofmutant tsD1. The culture medium was harvested after24 h of incubation at 31°C, and cell debris was removedby low-speed centrifugation. The virus was concen-trated by centrifugation at 20,000 rpm for 1 h in anMSE Superspeed centrifuge (MSE Ltd., London, Eng-land). The pelleted virus was resuspended in a smallvolume of 0.02 M Tris-hydrochloride (pH 8.0) bufferand layered onto a 15 to 45% sucrose gradient. Avisible separation of virion and DI particles was ob-tained by centrifugation for 1 h in a Beckman SW50rotor (Beckman Instruments, Glenrothes, Fife, Scot-land) at 20,000 rpm for 1 h. The virion and DI particlezones were collected and concentrated by centrifuga-tion. Both were resuspended in Tris-hydrochloride(pH 8.0) buffer and subjected to a second cycle ofgradient centrifugation and concentration. Sampleswere negative stained with ammonium molybdate andexamined by electron microscopy. The final DI parti-cle preparation contained 98% homogeneous truncatedparticles, about one-quarter virion length, and <2%intact full-length virions. Coinfection of tsDl-infectedcells with 100 electron microscope particles of this DIparticle preparation per cell reduced the yield of infec-tious virus by 99%.
Neutralization with specific antiserum. Guineapig hyperimmune convalescent antiserum was gener-ously provided by F. Brown, Animal Virus ResearchInstitute, Pirbright, Surrey, England, and used at afinal dilution of 1/150. Equal volumes of DI particlepreparation and antiserum were held at +4°C over-night. Residual infectivity was reduced by more than
J. VIROL.
Dow
nloa
ded
from
http
s://j
ourn
als.
asm
.org
/jour
nal/j
vi o
n 28
Dec
embe
r 20
21 b
y 11
2.16
5.86
.196
.

ENHANCED MUTABILITY 379
4 logio units, and interfering activity was abolished.This neutralized DI virus was used in the experimentsdescribed in Table 5.
Radiolabeling of viral polypeptides. Mono-layers of 5 x 106 BHK-21 cells in 30-ml bottles wereinfected with approximately 5 PFU of wild-type, mu-tant, or revertant virus per cell. The inoculum wasallowed to absorb for 30 min at 31°C before additionof 5 ml of incubation medium containing 5 yCi of [3S]-methionine (Radiochemical Centre, Amersham, Eng-land) per ml. The culture medium was harvested after16 to 20 h of incubation at 31°C, and cell debris wasremoved by centrifugation at 3,000 rpm for 20 min inan MSE Mistral centrifuge (MSE Ltd.). The radiola-beled virus was concentrated from the clarified culturefluid by centrifugation at 60,000 x g for 1 h in aBeckman Ti5O rotor. The centrifuge tubes weredrained carefully, and the pelleted virus was solubi-lized in 75 pl of electrophoresis lysis buffer (0.15 MTris-hydrochloride [pH 7.6], 6% SDS, 15% ,8-mercap-toethanol, 30% glycerol, 0.015% bromophenol blue)and stored at -20°C.
SDS-polyacrylamide gel electrophoresis. Thesolubilized virus samples were heated to 100°C for 5min before electrophoresis. The 3S-labeled polypep-tides were analyzed by discontinuous slab gel electro-phoresis in a 7.5% polyacrylamide-SDS gel as de-scribed by Marsden et al. (10). The gels were stainedwith 2% Coomassie brilliant blue and dried undervacuum. The radioactive bands were located by au-toradiography.
In vitro translation. Total cytoplasmic RNA wasextracted from BHK or BS-C-1 cell monolayers at 5 hafter infection, as described by Preston (17). RNA wastranslated in vitro, using a micrococcal nuclease-treated, fractionated rabbit reticulocyte cell-free sys-tem (16, 18).Sequencing methods. The 3'-terminal sequences
of the genome RNAs of tsDl and of a revertant weredetermined by adding a polyadenylated tail to the 3'-terminus of the purified RNA and then copying theRNA with reverse transcriptase from a p(dT)11-rAprimer in the presence, one at a time, of each dideox-ynucleoside triphosphate, as previously described forVSV Indiana (21). The sequences adjacent to thepolyadenylated tracts of the N mRNA's of tsDl andthe revertant were analyzed in the same way, usingp(dT)8-dC as primer, as described for VSV Indiana(13). It was verified by in vitro translation beforesequencing that the tsDl N mRNA specified the fast-migrating N polypeptide and that the revertant NmRNA specified an N polypeptide with normal mo-bility.
RESULTSHeterogeneity of mutant tsDl of VSV
New Jersey. Electrophoretic analysis of thevirion proteins of clones isolated from the tsDlstock revealed that the mutant stock was het-erogeneous, with three distinct phenotypes.These three phenotypes are illustrated in Fig. 1,and Table 1 gives the frequency of isolation ofclones with these phenotypes. All 48 clones iso-
2 3
NS D
N t
FIG. 1. SDS-polyacrylamide gel electrophoresis ofthe virion proteins of clones of VSV New Jerseyisolated from the tsDl stock. Track 1, Clone with thewild-type phenotype; track 2, clone with fast-migrat-ing G and N polypeptides; track 3, clone with fast-migrating G polypeptide.
TABLE 1. Clonal analysis of wild-type VSV NewJersey and mutants tsDl and tsE3
Virus stock Phenotype" No. of % ofts L G N NS M clones total
Wild type + + + + + + 50 100
tsDl
tsE3
M + M + +M + M M +M + + + +
+ + + + +
+ 34131
71272
+ 47 96+ 2 2
a +, Wild-type electrophoretic mobility in 7.5% poly-acrylamide-SDS gel or the ts+ phenotype; M, mutant,atypical electrophoretic mobility or the ts phenotype.
VOL. 39, 1981
Dow
nloa
ded
from
http
s://j
ourn
als.
asm
.org
/jour
nal/j
vi o
n 28
Dec
embe
r 20
21 b
y 11
2.16
5.86
.196
.

380 PRINGLE ET AL.
lated from the tsDl stock were temperaturesensitive, but only 13 exhibited the atypical mo-bility of G and N (Fig. 1, track 2) originallyassociated with tsDl. The majority (71%) of theclones had a normal N polypeptide and an aber-rant G polypeptide (Fig. 1, track 3), whereas oneclone was indistinguishable from wild-type virus(Fig. 1, track 1).In contrast, stocks of wild-type VSV New
Jersey and mutant tsE3 were homogeneous.Fifty clones of wild-type virus exhibited no de-tectable difference in polypeptide electropho-retic mobility. Forty-seven clones ofmutant tsE3exhibited the NS polypeptide mobility differ-ence associated with this mutant (5), whereasthe remaining two clones had the non-ts phe-notype and wild-type NS mobility characteristicof revertants of tsE3.
Wild-type (i.e., non-ts) revertants of the threecomponents of the tsDl stock were obtained byisolation of virus from plaques appearing onmonolayers at 390C to determine whether anyof the observed polypeptide changes were cor-related with temperature sensitivity. The resultsof these analyses are given in Table 2, whichshows that there was no association betweenchange in polypeptide mobility and reversion tothe non-ts phenotype. Indeed, several cloneswere isolated that had an additional mobilitychange affecting the NS polypeptide (Fig. 2,tracks 2 and 3), which had not been observedearlier. These results indicated that mutant tsDlwas genetically unstable and that the originalobservation of coreversion of the ts and N-mo-bility phenotypes was fortuitous.
Clones of mutant tsE3, on the other hand,isolated at 390C in the same experiment, showed
TABLE 2. Pattern of reversion ofphenotype ofmutants tsDl and tsE3 at 39°C
Mutant Phenotypea of Phenotypesa ofstock (VSV parental clone progeny clones No. of % of to-New Jer- - - clones tal
sey) tsGN NS ts G N NS
tsD1 MMM + +MM + 5 56+M+ + 3 33+MM M 1 11
M + + + + ++ + 22 52+ + +M 12 29+M+ + 8 19
M M + + +M + + 22 92+ ++ + 1 4+ ++ M 1 4
tsE3 M + + M +++ + 16 100a + Wild-type electrophoretic mobility in 7.5% poly-
acrylamide gel or the ts+ phenotype; M, mutant, atyp-ical electrophoretic mobility or the ts phenotype.
1 2 3 4
:i: .0 .:0 X - . .i.l .. :; t:- - 0 :; .. 0 X -.; .-W0.! 70 t:00: :::.. iN ;. Wi. ,77 .......... j R -
.,^.. .,s ..:L D _-t-f -t-;
::
::
::::
C0.:_
::$:
00 ff .t '-?, fi.*. _
WEt 7. C; fffa::> olE ,,E,.,, 4s,C,.D V,,: . - 00. f ;S . :f j . 0 E .- :<S'
M q r:.t: 9{S " /<Z00- -- tS
sS-_N w_F__
FIG. 2. SDS-polyacrylamide gel electrophoresis ofthe virion proteins of clones of tsDl of VSV NewJersey with altered NS mobility. Tracks 1 and 4,Clones with fast-migrating G polypeptide only; track2, clone with fast-migrating G, NS, and N polypep-tides; track 3, clone with fast-migrating NS polypep-tide only.
complete concordance of reversion of the ts andNS mobility phenotypes (Table 2), reconfirmingthe unique association of complementationgroup E with the NS polypeptide. Table 3 sum-marizes the transitions in phenotype observedduring sequential cloning of mutant tsDl, on theassumption that the G-and-N variant was theoriginal phenotype. Seven of the eight possiblesingle-step transitions were observed, and onlyone two-step change was observed. These obser-vations are consistent with change of phenotypeby sequential single-step mutations.Isolation of an M-polypeptide variant of
tsE3. Monolayers of BHK-21 cells were coin-fected with equal amounts (approximately 5PFU per cell) of mutants tsE3 and tsDl at 390C,and 21 clones were isolated from the progenyobtained by genetic complementation betweenthese two ts mutants (Table 4). Four of these tsprogeny clones resembled the tsDl parent and17 resembled the tsE3 parent. One of the tsE3clones had an atypicalM polypeptide in additionto the NS mobility difference invariably associ-ated with this mutant (Fig. 3, tracks 2 and 4).The band marked MR in Fig. 3 appeared to be
J. VIROL.
Dow
nloa
ded
from
http
s://j
ourn
als.
asm
.org
/jour
nal/j
vi o
n 28
Dec
embe
r 20
21 b
y 11
2.16
5.86
.196
.

ENHANCED MUTABILITY 381
TABLE 3. Transition in phenotype duringsequential cloning ofmutant tsDla
Polypeptide
G NS N
MM MT,Imm + Ml
+ + +4111T+ +M++411II,
+ + M ++ M MM M + 4
a , Observed; --+, not observed.
TABLE 4. Progeny from coinfection with mutantstsDl and tsE3 at 39°C
Phenotypea of progeny clonesisolated at 39°C No. Type
ts G N NS M
M M + + + 1 tsDlM M M + + 3 tsDlM + + M + 16 tsE3M + + M M 1 tsE3
a+, Wild-type electrophoretic mobility in 7.5% poly-acrylarnide-SDS gel or the ts+ phenotype; M, mutant,atypical electrophoretic mobility or the ts phenotype.
a virus-specific polypeptide and related to poly-peptide M, since it was only observed to migratefaster in extracts of clones of mutant tsE3 withthe fast M.Other experiments involving coinfection of
this tsE3-derived clone and tsD1 yielded addi-tional clones of tsE3, with mutational changeaffecting other polypeptides (Table 5, experi-ment 1). In these experiments BHK-21 cellmonolayers were infected with tsE3 alone, tsE3plus tsD1 virions, tsE3 plus tsDI DI particles,and tsE3 plus tsD1 DI particles preexposed toneutralizing antibody and then incubated for 24h at 310C. A total of 354 clones were isolatedfrom cultures infected with mutant tsE3 alone,and none differed from the parental phenotype(i.e., they had the variant NS and M of the tsE3parent). Therefore, the frequency (f) of spon-taneous mutants with lesions affecting polypep-tide mobility was -0.28%. By contrast, one of 62clones (f = 1.6%) isolated from the progenyreleased from cells infected with tsE3 and tsD1virions had a mutated M polypeptide with amobility intermediate between that of the tsE3parent and that of the wild type (Fig. 4). Threeof 191 clones (f = 1.6%) isolated from the prog-eny released from cells infected with tsE3 andtsD1 DI particles exhibited mutations affecting
either the N (Fig. 8, track 3) or the G polypeptide(Fig. 5, track 2). In these experiments, in whichtsDl was the inducer and tsE3 was the target,DI particles were used to reduce the probabilityof recovery of infectious parental tsDl and toenhance the proportion of tsE3 clones in theprogeny. It was evident from these results thatcoinfection with mutant tsDl enabled recoveryof polypeptide mobility variants of tsE3 and thatDI particles were as efficient inducers as thetsDl virion.Clones were also isolated from a mixed infec-
tion of tsE3 and DI particles of tsDl pretreatedwith neutralizing antiserum, as described in Ma-terials and Methods, to determine whether theapparent mutation-inducing ability of the tsD1stock was a specific effect or due to an adventi-tious factor. One of 156 clones (f= 0.6%) differedfrom the tsE3 parent. TheM polypeptide of thismutant migrated to a position intermediate be-tween that of the tsE3 M and that of the wild-type M (not shown; similar to difference shownin Fig. 4). The result of this experiment was
1 2 3 4 5
MR
FIG. 3. SDS-polyacrylamide gel electrophoresis ofthe virion proteins of clones of tsDl and tsE3 of VSVNew Jersey. Tracks 1, 3, and 5, Clones of tsDl withfast-migrating G and Npolypeptides; tracks 2 and 4,clones of tsE3 with fast-migrating NS and M poly-peptides. The band marked MR appeared to be re-lated to M, since a faster mobility was only observedin clones of VSV New Jersey with a fast-migratingM.
VOL. 39, 1981
Dow
nloa
ded
from
http
s://j
ourn
als.
asm
.org
/jour
nal/j
vi o
n 28
Dec
embe
r 20
21 b
y 11
2.16
5.86
.196
.

TABLE 5. Effect of coinfection with tsDl on (experiment 1) mutant tsE3 of VSV New Jersey, (experiment 2)mutant tsGl14(I) of VSV Indiana, and (experiment 3) mutant tsCh851(ChV) of Chandipura virus
Electrophoretic mobility variants in
No. of clones progenyExpt Inocuexami*edEtnocuumexamined Polypeptide af- No. of % of total
fected clones
1 tsE3 alone 354 Nil 0 <0.28tsE3 + tsD1/B 62 M 1 1.60tsE3 + tsD1/DI 191 N 2 1.60
GtsE3 + neutralized tsD1/DI 156 M 1 0.60
2 tsG114(I) alone 94 Nil 0 <1.10tsG114(I) + tsD1/DI 130 G (slow) 21 47
G (fast) 2JtsG114(I) + neutralized tsD1/DI 131 Nil 0 -0.8
3 tsCh851(ChV) alone 67 Nil 0 <1.5tsCh851(ChV) + tsDl/B 40 N 1 2.4
a BHK-21 cells were infected simultaneously with 5 PFU of the target virus [tsE3 or tsG114(I)] per cell and5 PFU of complete (B) or 100 elecctron microscope particles of incomplete (DI) virions of tsDl per cell.
1 2 3 4
FIG. 4. SDS-polyacrvlamide ael electrox- - __*_.1--..Iw---C VV ---vv1the virion proteins of a clone of tsE3 of VSV NewJersey with partial reversion of the fast M-mobilityphenotype. Tracks 2 and 3, clones of tsDl with Mpolypeptides of wild-type mobility; track 1, coelectro-phoresis of tsE3 with the fast M and the partial
ambiguous because the reduction in frequencyof mobility variants of tsE3 from 1.6 to 0.6%after neutralization of tsDl DI particles did notexclude the possible role of an adventitious fac-
'1 L tor. However, a second experiment, in whichmutant tsG114(I) of VSV Indiana was used asthe target virus, provided additional support forthe specific involvement of tsDl (see below).
Effect of tsDl on other vesiculovirusesduring coinfection. Monolayers of BHK-21cells were infected with the tsG114(I) mutant ofVSV Indiana either alone or in combination withnormal or preneutralized DI particles of tsDl.The analysis of the progeny is given in Table 5
X G (experiment 2). No variants (i.e., f - 1.1%) wereisolated from the tsG114(I) self-infection,
a N whereas four variants of tsG114(I), all involvingthe G polypeptide, were present among 191clones (f = 4.7%) isolated from the coinfectionwith tsDl DI particles. Figure 6 (track 2) showsa tsG114(I) mutant with a slow G. Preneutrali-zation of the DI particles reduced the frequencyof tsG114(I) mobility variants to <0.8%.
In another experiment (Table 5, experiment3), a mutant of Chandipura virus, a serologicallydistinct member of the vesiculovirus genus, witha variant N was isolated from the progeny of a
X M coinfection with tsDl (Fig. 7, track 2). The fre-quency of isolation was 2.4%, compared with afrequency of s1.5% in the absence of tsDl.These results indicated that coinfection with
Mhoresis of mutant tsDl (or a DI particle derived from it)
revertant, showing separation of theM bands; track4, coelectrophoresis of the partial revertants of tsE3and tsDl, showing separation of the normal M bandand the partially revertedM band.
382 PRINGLE ET AL. J. VIROL.
Dow
nloa
ded
from
http
s://j
ourn
als.
asm
.org
/jour
nal/j
vi o
n 28
Dec
embe
r 20
21 b
y 11
2.16
5.86
.196
.

ENHANCED MUTABILITY 383
1 2 3
G t
FIG. 5. SDS-polyacrylamide gel electrophoresis ofthe virion proteins of a clone of tsE3 of VSV NewJersey with altered mobility of the G polypeptide.Tracks 1 and 3, tsE3 clones of normal phenotype;track 2, clone with altered mobility of the G polypep-tide.
could induce mutational change in heterologousvesiculoviruses as well as in the homologousVSV New Jersey.In vitro translation. Total cytoplasmic
RNA was extracted from infected BHK cellsand translated in vitro in a rabbit reticulocytecell-free system to determine whether post-translational processing contributed to the aber-rant mobility phenotypes. The expected differ-ences in N, NS, and M polypeptides of fourrepresentative mutants were found in transla-tion products (Fig. 8), suggesting that the mo-bility changes resulted from alterations in thenascent polypeptides.The polyacrylamide gel system used in these
experiments resolved the G precursor into two
bands, possibly due to partial glycosylation invitro (23). When RNA from VSV Indiana var-iants with altered G polypeptide was translated(Fig. 9), one mutant (tracks 2 and 6) showedincreased mobility whereas another (track 4)had the wild-type pattern (tracks 3 and 5). Thesame results were obtained with RNA extractedfrom both BHK and BS-C-1 cells. It is con-cluded, therefore, that an aberration of glycosyl-ation could account for the atypical mobility ofclone 80/3/27 (tracks 3 and 5) but not of clone80/21/9 (tracks 2 and 6).The results of these and other in vitro trans-
lation experiments are summarized in Table 6,emphasizing that the observed mobility pheno-types are not due to post-translational process-ing except for certain G-protein mutants.Absence of recombination. The possibility
that these variants originated by a recombina-tional rather than a mutational process was ex-amined by coinfection of BHK-21 cells with twoclones of VSV New Jersey which had been de-rived in these experiments. These clones weretemperature sensitive and had electrophoreti-
1 2 3
N D
FIG. 6. SDS-polyacrylamide gel electrophoresis ofthe virion proteins of tsGl14(I) of VSV Indiana withaltered mobility of the Gpolypeptide. Tracks 1 and 3,tsGl14(I) clones of normal phenotype; track 2, clonewith altered mobility of the G polypeptide.
VOL. 39, 1981
Dow
nloa
ded
from
http
s://j
ourn
als.
asm
.org
/jour
nal/j
vi o
n 28
Dec
embe
r 20
21 b
y 11
2.16
5.86
.196
.

384 PRINGLE ET AL.
2 3
c D
FIG. 7. SDS-polyacrylamide gel electrophoresis ofthe virion proteins of a clone of tsCh851(ChV) ofChandipura virus with altered mobility of the Npolypeptide. Tracks I and 3, Clones of tsCh851(ChV)of normal phenotype; track 2, clone with alteredmobility of the Npolypeptide.
cally distinct G, N, NS, and M proteins (Fig. 10,tracks 1 and 2). Potential non-ts recombinantswere selected by cloning from plaque-formingvirus at 39°C, and 22 isolates were scored for thefour unselected polypeptide mobility markers.All exhibited polypeptides characteristic of bothparental viruses (Fig. 10, tracks 3 to 8, shows sixof these isolates). In each case segregation of thetwo parental types was obtained by cloning at31°C (e.g., Fig. 11, tracks 3 to 8). All of theseclones were temperature sensitive, indicatingthat the plaques obtained on monolayers at 39°Cwere the product of complementation, since theparental clones were derived from the comple-menting mutants tsE3 and tsDl. There was no
evidence for the occurrence of recombination inthis experiment, which involved four independ-ent markers; therefore, it is unlikely that thevariants described could have arisen by a recom-binational event.Comparison of the nucleotide sequences
of the N-gene termini of clones ofVSV NewJersey with distinctive N proteins. As al-
ready described, some clones of tsDl had an Npolypeptide which ran faster than wild-type Non SDS-polyacrylamide gel, suggesting that thepolypeptide was shorter by about 1,000 daltons(or 10 amino acids). Since revertants could beobtained, the phenotype of the mutant was dueeither to a real size change resulting from earlytermination of translation (a nonsense mutation)or late initiation of translation or to an aminoacid substitution in a polypeptide of unalteredchain length. To examine the first possibility,the nucleotide sequences of the terminal regionsof the N gene of a clone of tsD1, with mutant N,and a revertant of tsDl, with N of wild-typemobility, were determined and compared withthe corresponding sequences of wild-type VSVNew Jersey (12).
FIG. 8. SDS-polyacrylamide gel electrophoresis ofthe products of in vitro translation of total cytoplas-mic RNA fr-om mutant-infected cells in a rabbit retic-ulocyte cell-fr-ee system. Track 1, Escherichia coliRNA track 2 uninfected BHK-21 cells RNA; track3 clone (80/15/100) of tsE3 with a slow-migrating Nand fast-migrating NS and Mpolypeptides; track 4,wild- type VSVNew Jersey; track 5, clone oftsE3 withfast-migrating NS and M; track 6, clone of tsDl withcomigrating fast NS and N polypeptides; track 7,clone of tsDl with fast-migrating Npolypeptide. TheG polypeptides were not uniformly resolved in thisgel
J. VIROL.
Dow
nloa
ded
from
http
s://j
ourn
als.
asm
.org
/jour
nal/j
vi o
n 28
Dec
embe
r 20
21 b
y 11
2.16
5.86
.196
.

ENHANCED MUTABILITY 385
FIG. 9. SDS-polyacrylamide gel electrophoresis of
the products of in vitro translation of total cytoplas-
mic RNA fr-om mutant-infected cells in a rabbit retic-
ulocyte cell-free system. Track 1, Uninfected BHK-21
cells RNA; track 2, clone (80/21/9) of tsG114aI) with
fast-migrating pre-G polypeptides translated fr-ominfected BHK-21 cell RNA; track 3, VSV Indiana
wild-typepolypeptides translated from infectedBHK-
21 cell RNA; track 4, clone (80/3/27) of tsGIl14(I) with
a G-polypeptide mobility difference not resolved by in
vitro translation of infected BHK-21 cell RNA; track
5, VSV Indiana wild-type polypeptides translated
from infected ES-C-I cell RNA; track 6, as track 2
but translated from infected ES-C-i1 cell RNA, show-
ing that the fast mobility ofthepre-Gpolypeptides is
independent of host factors; track 7, uninfected ES-
C-i cell RNA.
The sequences for the three viruses are com-
pared diagrammatically in Fig. 12. The number-
ing of nucleotides follows the convention
adopted for the 3'-terminal region of the genome
RNA of VSV Indiana (11) and the 3'-terminal
region of the N mRNA of VSV Indiana (13).
The sequence of the genome RNA of VSV New
Jersey has been determined from the 3'-end to
residue 201, with the exception of residues 15and 106 which remain ambiguous (12). The 3'-terminal sequences of the virion RNA of mutanttsDl and its revertant have been determined forresidues 16 to 212. However, in tsDI residues147 and 148, and in the revertant residues 146 to148, were unresolved. Residue 106 was unre-solved in all three clones. Only one base changewas seen; at residue 46 a C in wild type wasreplaced by a U in both tsDl and its revertant.Since this change occurs in the leader region andis present in both mutant and revertant, it can-not be involved in the N-mobility phenotype.The sequence complementary to the 5'-terminalregion ofN mRNA begins at residue 51, and thetranslation initiation codon is at residues 64 to66. There are no base changes in either virus toresidue 212, a region encoding 50 amino acids.The unresolved regions (106, 146 to 148) do n6tappear to offer any possibility for generation 6fthe short polypeptide phenotype by late imnii-tion of translation.The 3'-terminal sequence of the N mRNA of
wild-type VSV New Jersey has been determinedup to residue 205, numbered from the polyaden-ylated 3 tail (13). Sequences were also obtainedfor the 3'-terminus of the mRNA of mutant tsDland its revertant to residue 205. Residues 97 to99 were not determined for tsDl and its revert-ant due to strong compression of the bands here,using the standard method. The wild-type se-quence was determined by using more severedenaturing conditions. The presence of the samecompression phenomenon in all three RNAs,however, suggests that the sequences are iden-tical. One change in sequence was found at res-idue 53, where an A residue in wild type wasreplaced by G in tsDl and its revertant. Thetranslation termination codon is thought to beresidues 45 to 47. Therefore, the above basechange would correspond to replacement of as-paragine in the wild type by aspartic acid intsDl and its revertant. This is unlikely to beresponsible for the N-polypeptide mobility phe'.notype, since it is found in both mutant andrevertant. (Interestingly, the sequence in thecorresponding position in VSV Indiana is thesame as that of tsDl and its revertant.) All threesequences then predict the same amino acidsequence for a further 16 amino acids up to theincompletely sequenced region at residues 97 to99. This covers the area where any new trans-lation termination codon should be located, i.e.,about 10 amino acids or 30 nucleotides from thewild-type translation terminator. Residues 97 to99, undetermined in tsDI and its revertant, com-prise the codon UCA in the wild type. A singlebase change could convert this to a nonsensecodon; consequently, there is a possibility that
VOL. 39, 1981
Dow
nloa
ded
from
http
s://j
ourn
als.
asm
.org
/jour
nal/j
vi o
n 28
Dec
embe
r 20
21 b
y 11
2.16
5.86
.196
.

TABLE 6. Summary of the in vivo and in vitro phenotypes of the electrophoretic mobility mutants of tsDl orinduced by tsDl
Polypep- Phenotypetide Origina Lesion
ts In vivo In vitro
L
G VSV-NJ tsDl ts/ts+ Fast G Nonmutant (pre-G) GlycosylationVSV-NJ tsE3 ts/ts+ Slow G Nonmutant (pre-G) GlycosylationVSV-IND tsG114(I) ts Slow G Nonmutant (pre-G) Glycosylation
ts Fast G Mutant ?
N VSV-NJ tsDl ts/ts+ Fast N Mutant ?VSV-NJ tsE3 ts/ts+ Slow N Mutant ?CHV tsCh851(ChV) ts Slow N Mutant ?
NS VSV-NJ tsDl ts/ts+ Fast NS Mutant ?VSV-NJ tsE3 ts Fast NS Mutant ?
M VSV-NJ tsE3 ts/ts+ Fast M Mutant ?ts Intermediate M ND ?
VSV-NJ, VSV New Jersey serotype; VSV-IND, VSV Indiana serotype; CHV, Chandipura virus.bND, no data.
1 2 3 4 5 6 7 8
L~~~~~~~~~~~t0f-0s0 t0tmi i- .I busESit bush-
G~ ~ ~___',-' _La mmiWR...._...
NS
_
FIG. 10. SDS-polyacrylamide gel electrophoresisof the virion proteins of39WCplaque-forming virus in
the progeny of a tsDl x tsE3 coinfection. Track 1,
tsDl parent; track 2, tsE3 parent; tracks 3-8, poly-peptide profiles of virus isolated from six separate
plaques observed at 39WC (each exhibited 0, N, NS,and M polypeptides characteristics of both parents).
Figure 11 shows that the biparental Phenotype ofthese viruses is a consequence of complementation.
the mutation resides here, although we considerthis unlikely. The remainder of the three se-quences are identical up to residue 205.
DISCUSSIONThe results presented here establish that mu-
tations were generated at high frequency in astock of mutant tsDl of VSV New Jersey duringlytic cycles of infection. The mutations wererecognized as changes in electrophoretic mobil-ity of viral polypeptides. The molecular basis forthese changes has not been established, apartfrom the three G-protein mutants listed in Table6, whose aberrant mobility was related to gly-cosylation. Sequencing of the ends of the N geneexcluded premature termination, or delayed ini-tiation, in determination of the fast N-mobilityphenotype of mutant tsDl. Presumably theaberrant electrophoretic mobility of the mutantN polypeptide was the consequence of an inter-nal missense mutation. Why this should affectthe mobility of the polypeptide in SDS-poly-acrylamide gel has not been established. It isknown, however, that single amino acid substi-tution in QB coat protein (8), alpha-crystallinprotein (3), and a histidine transport protein ofSalmonella typhimurium (15) can result in amobility difference in SDS-polyacrylamide gelcorresponding to 1 x 103 to 2 x 103 molecularweight. The frequency of ts mutation was alsoenhanced in the presence of tsDl, but, being aquantitative property, was less amenable to thistype of analysis than discrete changes in electro-phoretic mobility.Determination of the sequence of the nucleo-
tides at the termini of the N gene revealed two
"W
386 PRINGLE ET AL. J. VIROL.
p- 19- 17PT,719. 7"77W
Dow
nloa
ded
from
http
s://j
ourn
als.
asm
.org
/jour
nal/j
vi o
n 28
Dec
embe
r 20
21 b
y 11
2.16
5.86
.196
.
ENHANCED MUTABILITY 387
1 2 3 4 5 6 7 8
G t
NS
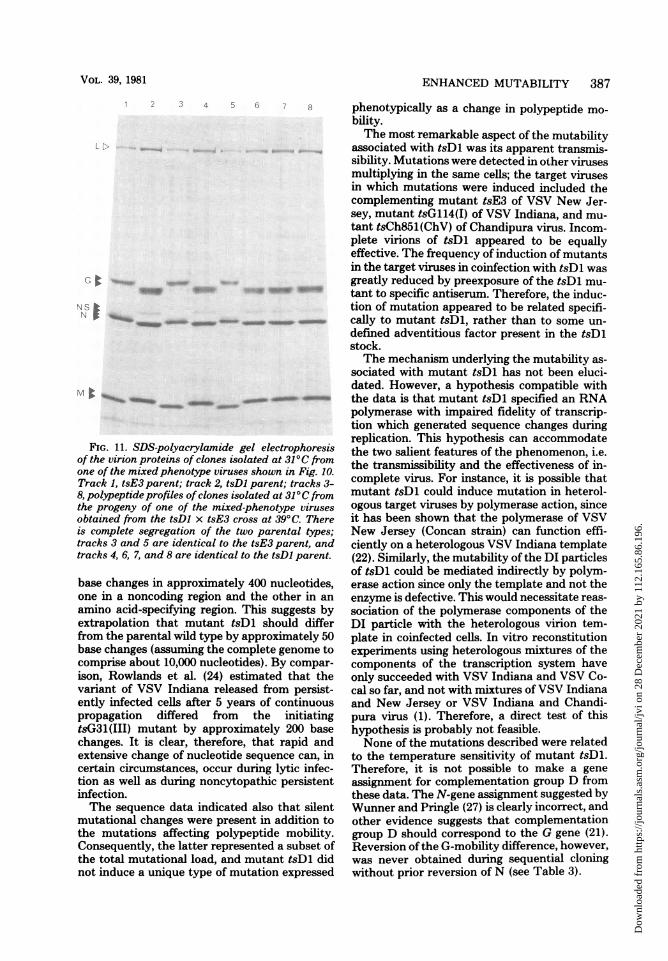
FIG. 11. SDS-polyacrylamide gel electrophoresisof the virion proteins of clones isolated at 31°C fromone of the mixed phenotype viruses shown in Fig. 10.Track 1, tsE3 parent; track 2, tsDl parent; tracks 3-8, polypeptide profiles ofclones isolated at 31°C fromthe progeny of one of the mixed-phenotype virusesobtained from the tsDl x tsE3 cross at 39°C. Thereis complete segregation of the two parental types;tracks 3 and 5 are identical to the tsE3 parent, andtracks 4, 6, 7, and 8 are identical to the tsDl parent.
base changes in approximately 400 nucleotides,one in a noncoding region and the other in an
amino acid-specifying region. This suggests byextrapolation that mutant tsDl should differfrom the parental wild type by approximately 50base changes (assuming the complete genome tocomprise about 10,000 nucleotides). By compar-
ison, Rowlands et al. (24) estimated that thevariant of VSV Indiana released from persist-ently infected cells after 5 years of continuouspropagation differed from the initiatingtsG31(III) mutant by approximately 200 basechanges. It is clear, therefore, that rapid andextensive change of nucleotide sequence can, incertain circumstances, occur during lytic infec-tion as well as during noncytopathic persistentinfection.The sequence data indicated also that silent
mutational changes were present in addition tothe mutations affecting polypeptide mobility.Consequently, the latter represented a subset ofthe total mutational load, and mutant tsDl didnot induce a unique type of mutation expressed
phenotypically as a change in polypeptide mo-bility.The most remarkable aspect of the mutability
associated with tsDl was its apparent transmis-sibility. Mutations were detected in other virusesmultiplying in the same cells; the target virusesin which mutations were induced included thecomplementing mutant tsE3 of VSV New Jer-sey, mutant tsG114(I) of VSV Indiana, and mu-tant tsCh851(ChV) of Chandipura virus. Incom-plete virions of tsDl appeared to be equallyeffective. The frequency of induction of mutantsin the target viruses in coinfection with tsDl wasgreatly reduced by preexposure of the tsDl mu-tant to specific antiserum. Therefore, the induc-tion of mutation appeared to be related specifi-cally to mutant tsDl, rather than to some un-defined adventitious factor present in the tsDlstock.The mechanism underlying the mutability as-
sociated with mutant tsDl has not been eluci-dated. However, a hypothesis compatible withthe data is that mutant tsDl specified an RNApolymerase with impaired fidelity of transcrip-tion which generated sequence changes duringreplication. This hypothesis can accommodatethe two salient features of the phenomenon, i.e.the transmissibility and the effectiveness of in-complete virus. For instance, it is possible thatmutant tsDl could induce mutation in heterol-ogous target viruses by polymerase action, sinceit has been shown that the polymerase of VSVNew Jersey (Concan strain) can function effi-ciently on a heterologous VSV Indiana template(22). Similarly, the mutability of the DI particlesof tsDl could be mediated indirectly by polym-erase action since only the template and not theenzyme is defective. This would necessitate reas-sociation of the polymerase components of theDI particle with the heterologous virion tem-plate in coinfected cells. In vitro reconstitutionexperiments using heterologous mixtures of thecomponents of the transcription system haveonly succeeded with VSV Indiana and VSV Co-cal so far, and not with mixtures of VSV Indianaand New Jersey or VSV Indiana and Chandi-pura virus (1). Therefore, a direct test of thishypothesis is probably not feasible.None of the mutations described were related
to the temperature sensitivity of mutant tsDl.Therefore, it is not possible to make a geneassignment for complementation group D fromthese data. The N-gene assignment suggested byWunner and Pringle (27) is clearly incorrect, andother evidence suggests that complementationgroup D should correspond to the G gene (21).Reversion ofthe G-mobility difference, however,was never obtained during sequential cloningwithout prior reversion of N (see Table 3).
VOL. 39, 1981
Dow
nloa
ded
from
http
s://j
ourn
als.
asm
.org
/jour
nal/j
vi o
n 28
Dec
embe
r 20
21 b
y 11
2.16
5.86
.196
.

Ul)
soR o_i-~~X_ gt
S~~~~~~~~~~~~~b
_ 04 i
tx ~ ~ -X V<_
388
111~~~~t
Dow
nloa
ded
from
http
s://j
ourn
als.
asm
.org
/jour
nal/j
vi o
n 28
Dec
embe
r 20
21 b
y 11
2.16
5.86
.196
.

ENHANCED MUTABILITY 389
ACKNOWLEDGMENT
We are indebted to Matt Dunlop for assistance in prepa-
ration of the cell-free translation experiments and to JohnParry for electron microscopy.
LITERATURE CITED
1. Bishop, D. H. L., S. U. Emerson, and A. Flamand.1974. Reconstitution of infectivity and transcriptaseactivity of homologous and heterologous viruses: vesic-ular stomatitis (Indiana serotype), Chandipura, vesicu-lar stomatitis (New Jersey serotype), and Cocal viruses.J. Virol. 14:139-144.
2. Clewley, J. P., D. H. L. Bishop, C.-Y. Kang, J. Coffin,W. M. Schnitzlein, M. E. Reichmann, and R. E.Shope. 1977. Oligonucleotide fingerprints of RNA spe-
cies obtained from rhabdoviruses belonging to the ve-
sicular stomatitis virus subgroup. J. Virol. 23:152-166.3. de Jong, W. W., A. Zweers, and L. H. Cohen. 1978.
Influence of a single amino acid substitution on electro-phoretic mobility of sodium dodecyl sulfate-proteincomplexes. Biochem. Biophys. Res. Commun. 82:532-539.
4. Emerson, S. U., and R. R. Wagner. 1972. Dissociationand reconstitution of the transcriptase and templateactivities of vesicular stomatitis B and T virions. J.Virol. 10:297-309.
5. Evans, D., C. R. Pringle, and J. F. Szilagyi. 1979.Temperature-sensitive mutants of complementationgroup E of vesicular stomatitis virus New Jersey sero-
type possess altered NS polypeptides. J. Virol. 31:325-333.
6. Gadkari, D. A., and C. R. Pringle. 1980. Temperature-sensitive mutants of Chandipura virus. I. Inter- andintragroup complementation. J. Virol. 33:100-106.
7. Gadkari, D. A., and C. R. Pringle. 1980. Temperature-sensitive mutants of Chandipura virus. II. Phenotypiccharacteristics of the six complementation groups. J.Virol. 33:107-114.
8. Glowacki-Strauss, E., and P. Kaesberg. 1970. Acryl-amide gel electrophoresis of bacteriophage QB: electro-phoresis of the intact virions and of the viral proteins.Virology 42:437-452.
9. Holland, J. J., E. A. Grabau, C. L. Jones, and B. L.Semler. 1979. Evolution of multiple genome mutationsduring long-term persistent infection by vesicular sto-matitis virus. Cell 16:495-504.
10. Marsden, H. S., I. K. Crombie, and J. H. Subak-Sharpe. 1976. Control of protein synthesis in herpesvirus-infected cells: analysis of the polypeptides inducedby wild type and sixteen temperature-sensitive mutantsof HSV strain 17. J. Gen. Virol. 31:347-372.
11. McGeoch, D. J., and A. Dolan. 1979. Sequence of 200nucleotides at the 3'-terminus of the genome RNA ofvesicular stomatitis virus. Nucleic Acids Res. 6:3199-3211.
12. McGeoch, D. J., A. Dolan, and C. R. Pringle. 1980.Comparison of nucleotide sequences in the genomes ofthe New Jersey and Indiana serotypes of vesicular
stomatitis virus. J. Virol. 33:69-77.13. McGeoch, D. J., and N. T. Turnbull. 1978. Analysis of
the 3'-terminal nucleotide sequence of vesicular sto-matitis virus N protein mRNA. Nucleic Acids Res. 5:4007-4024.
14. Mudd, J. A., R. W. Leavitt, D. T. Kingsbury, and J.J. Holland. 1973. Natural selection of mutants of ve-sicular stomatitis virus by cultured cells of Drosophilamelanogaster. J. Gen Virol. 20:341-351.
15. Noel, K. D., K. Nikaido, and G. Ferro-Luzzi Ames.1979. Single amino acid substitution in a histidine-trans-port protein drastically alters its mobility in sodiumdodecyl sulfate-polyacrylamide gel electrophoresis. Bio-chemistry 18:4159-4165.
16. Pelham, H. R. B., and R. J. Jackson. 1976. An efficientmRNA-dependent translation system from reticulocytelysates. Eur. J. Biochem. 67:247-256.
17. Preston, C. M. 1977. The cell-free synthesis of herpesvi-rus-induced polypeptides. Virology 78:349-353.
18. Preston, C. M. 1979. Control of herpes simplex virus type1 mRNA synthesis in cells infected with wild-type virusor the temperature-sensitive mutant tsK. J. Virol. 29:275-284.
19. Pringle, C. R. 1970. Genetic characteristics of conditionallethal mutants of vesicular stomatitis virus induced by5-fluorouracil, 5-azacytidine, and ethyl methane sulfo-nate. J. Virol. 5:559-567.
20. Pringle, C. R., I. B. Duncan, and M. Stevenson. 1971.Isolation and characterization of temperature-sensitivemutants of vesicular stomatitis virus, New Jersey sero-type. J. Virol. 8:836-841.
21. Pringle, C. R., and J. F. Szilagyi. 1980. Gene assignmentand complementation group, p. 141-161. In D. H. L.Bishop (ed.), Rhabdoviruses, vol. 2. CRC Press, BocaRaton, Fla.
22. Repik, P., A. Flamand, and D. H. L. Bishop. 1976.Synthesis of RNA by mutants of vesicular stomatitisvirus (Indiana serotype) and the ability of wild-typeVSV New Jersey to complement the VSV Indiana ts GI-114 transcription defect. J. Virol. 20:157-169.
23. Rothman, J. E., F. N. Katz, and H. F. Lodish. 1978.Glycosylation of a membrane protein is restricted to thegrowing polypeptide chain but is not necessary forinsertion as a transmembrane protein. Cell 15:1447-1454.
24. Rowlands, D., E. Grabau, K. Spindler, C. Jones, B.Semler, and J. Holland. 1980. Virus protein changesand RNA termini alterations evolving during persistentinfection. Cell 19:871-880.
25. Semler, B. L., and J. J. Holland. 1979. Persistent vesic-ular stomatitis virus infection mediates base substitu-tions in viral RNA termini. J. Virol. 32:420-428.
26. Wunner, W. H., and C. R. Pringle. 1972. Protein syn-thesis in BHK-21 cells infected with vesicular stomatitisvirus. II. ts mutants of the New Jersey serotype. Virol-ogy 50:250-253.
27. Wunner, W. H., and C. R. Pringle. 1974. A temperature-sensitive mutant of vesicular stomatitis virus with twoabnormal virus proteins. J. Gen. Virol 23:97-106.
VOL. 39, 1981
Dow
nloa
ded
from
http
s://j
ourn
als.
asm
.org
/jour
nal/j
vi o
n 28
Dec
embe
r 20
21 b
y 11
2.16
5.86
.196
.