enhancement in inulinase production by …nopr.niscair.res.in/bitstream/123456789/19874/1/ijbt...
TRANSCRIPT

Indi an Journal of Biotechnology Vol I , July 2002, pp 270-274
Enhancement in Inulinase Production by Mutagenesis in Penicillium purpurogenum
Arun Dev Sharma" , Jagpreet Singh Nanda', Prabhjot Kaur Gill' , Sukdev Singh Bhullar' , Prabhjeet Singh' and Dhiraj Vyas2
I Department of Biotechno logy, Guru Nanak Dev University , Amritsar 143005, India 2 Biotechnology Division, Institute of Himalayan Bioresource and Technology, Palampur 17606 1. In ia
Received 5 September 2001 ; accepted 13 March 2002
Extracellular inulinase producing strain of Penicillium pIlrpuroge1lllm was isolated from rhizosphere soil of chicory. Conidia of this selected strain were subjected to mutagenesis with UV and NTG (3-nitro, S-methyl guanidine). After mutagenesis, of various colonies screened, few were selected to carry out the inulinase study. The study revealed 2.S-fold higher inulinase activity with 8 to 21-fold liS ratio in all the selected colonies, indicating the enhancement in inulinase production after both mutagenic treatments in all the selected mutants.
Keywords: Pe1licillium pllrp"rogen"m, liS ratio, inulil1lase, invertase, mutants
Introduction Fructose, a sweetest natural sugar having beneficial
medicinal effects, is emerging as an alternative sweetener to sucrose in the pharmaceutical industry. It can be produced from starch by enzymatic methods involving a-amylase, amyloglucosidase and glucose isomerase resulting in production of an invert sugar mixture (oligosaccharides, 5; fructose, 45; glucose, 50%). This method is uneconomical. Fructose can also be produced by acid hydrolysi s of inulin, but this method results in production of difructose anhydrides , which have no sweetening capacity (Vandamme & Derycke, 1983). Another method may include microbi al sources, which secrete higher amount of inulinase having tendency to cleave fructose from inulin (a polymer of fructose with terminal glucose), by single-step catalyzed reaction and yields up to 95 % of fructose. In this study, attempt has been made to enhance the inulinase production by two different mutagenic methods in fungal isolate (Sharma et al, 1998). Although there are reports on enhancement in inulinase production by mutagenesis on Aspergillus JumigatLis (Fiedurek et ai, 2000) and A. niger (Vi shwanthan & Ku lkarni , 1995), present communication is the first report on Penicillium purpurogenum where UV and NTG (3-nitro, 5-methyl
"Author fo r correspondence: Fax: 0 183-258272 E-mail : [email protected] arundevsharma@ indi atimes.com
guanidine) mutagenesis was carried out to enhance the inulinase production.
Materials and Methods Microorganism
The strain of P. purpurogenum, used in the present study, was isolated from rhizosphere soil of chicory (Sharma et ai, 1998) and deposited with Microbial Type Culture Collection (MTCC No. 3009) at Institute of Microbial Technology, Chandigarh (India). The stock was maintained on Czapek 's Dox Agar (CDA) at 4 DC and, whenever required, fresh culturing was done on CDA slants. The composition of CDA medium is as follows: inulin/sucrose, 10.0; NaNO), 3; K2HP04, 1.0; MgS04. 7H20 , 0.5; KCI , 0.5; FeS04. 7H20 , 0.01 and agar, 20.0 gr l
.
Preparation of Inoculum A spore suspensi on was prepared by adding sterile
Phys iological saline solution [0.9 % w/v NaCI containing 1 % (v/v) Tween-80], to 4-day old slant culture.
Mutagenizatiol'l by UV Light Inoculum having 104/ml spores of P. purpura
genum (wild strain) was prepared in phys iological sal ine solution. 10 ,ul of spore suspension was poured at the centre of 9-cm wide agar pl ates and spread with the help of a sterile spreader following ex posure to UV light (60 w) for 0- 10 min (in 5 di ffe rent replicates) at a di stance of 15cm from the UV source.

SHARMA et at: ENHANCED INULINASE PRODUCTION BY MUTAGENESIS 271
After UV treatment, each petri plate was wrapped in a black paper and incubated at 34°C. After 4 days of incubation, number of colonies was counted and petri plates having 1.0% survival rate were used for inulinase study. After selecting well-separated colonies, each colony was picked up with the help of a steri le loop, and transferred to fresh CDA slants and incubated at 34°C for 4 days. After 4 days of growth, spore suspension was prepared as described earlier and inoculum-containing 106/ml spores were inoculated in 250 ml Erlenmeyer flasks containing 50 ml CDA supplemented with 1 % inulin (w/v). The flasks were incubated in rotatory incubator shaker (New Brunswick Scientific Co. , USA) at 200 rpm at 34°C for 9 days. The culture broth was centrifuged at 8000 rpm for 20 min at 4°C and supernatants were stored at -20°C for enzyme assays.
Mutagenization by NTG In chemical mutagenesis (Nakamura et ai, 1994),
inoculum containing 104/ml spore suspension of P. purpurogenum was prepared. Different amounts of NTG (0-2 mg) were added to the spore suspension (NTG solution was prepared in 0.05 M phosphate buffer, pH 6.0, each treatment was given in 5 replicates) followed by incubation at 34°C for 30 and 60 min, respectively. Further, it was diluted 100 times with 0.056 M phosphate buffer (PH 8.0) and 1 ml of the spore suspension was spread on the agar plates. Plates were incubated at 34°C for 7 days and treatments giving lowest number of colonies were further studied for inulinase production.
The well-separated colonies were transferred to fresh CDA slants and incubated at 34°C for 4 days . After 4 days of growth, mutagenization was carried out as per the same parameters described under mutagenization by UV.
Enzyme Assay Inulinase and invertase activities were assayed
(Nelson, 1944) by measuring the amount of reducing sugars released from inulin and sucrose, respectively. The assay mixture for inulinase contained 0.1 ml of enzyme extract and 0.9 ml of 1.1 % inulin in 0.1 M sodium acetate buffer (PH 5.5). The mixture was incubated at 60°C for 1 hr. Invertase assay mixture contained 0.1 ml of enzyme preparation and 0.9 ml of sucrose (55.5 mM) in 0.1 M sodium acetate buffer (PH 5.5). After incubating for 30 min at 60°C, the reaction was stopped by adding 1 ml of Nelson' s
reagent and amount of reducing sugars thus released was measured. The reducing sugars already present in the enzyme samples were also estimated and appropriate correction made while calculating the actual amount of sugar released.
One unit of inulinase was defined as the amount of enzyme that released ImM of fructose per min. One unit of invertase activity was defined as the amount of enzyme that hydrolyzed ImM of sucrose per min. Calibration curve was drawn with fructose (10-100 mg). Protein content in crude extract was estimated (Lowry et ai, 1951) using bovine serum albumin as a standard.
Results and Discussion 13 UV mutants were screened after 10 min UV
treatment (Fig. 1) and inulinase profile was studied in these mutants for 9 days after culturing. Maximum inulinase activity was observed at 7 lh day of culturing (Table 1). Among these mutants, 6 were selected as the representative samples for inulinase study and further tested on 250 ml culturing medium on a rotatory shaker. The inulinase activity of the investigated mutants was substantially higher in comparison to the parent strain but a significant enhancement was observed in mutant UVmlO having 2.5-fold higher inulinase activity with 21 times higher liS (inulin/sucrose) ratio (Table 2). The increased liS ratio and decisively lower invertase activity in all the selected mutants further confirm the enhancement in inulinase production after UV treatment. It is generally accepted that the liS ratio is higher than 10-2
for inulinase, while lower than 10-4 for invertase (Guiraud et ai, 1981). The results of the present study support several findings (Fiedurek et ai, 2000; Viswanathan et ai, 1994), which have observed 2-fold increase in inulinase activity after UV mutagenesis.
I- 150-r-------------~ z Wu..
~ 0 Cf.) 100 w-.J w (l.~Z w_O ~~5 50 I-::lU :)Cf.) W ~ O~~--.l~_~~__.l~ _ _L~_~
C 2 4 6 8 10
W EXPOSURE T1tvE ( rrin)
Fig. I-Per cent survival of colonies after UV treatment for different time intervals. C---control. Each value is the ± SE of the mean from 5 different replicates.
272 INDIAN J BIOTECHNOL, JULY 2002
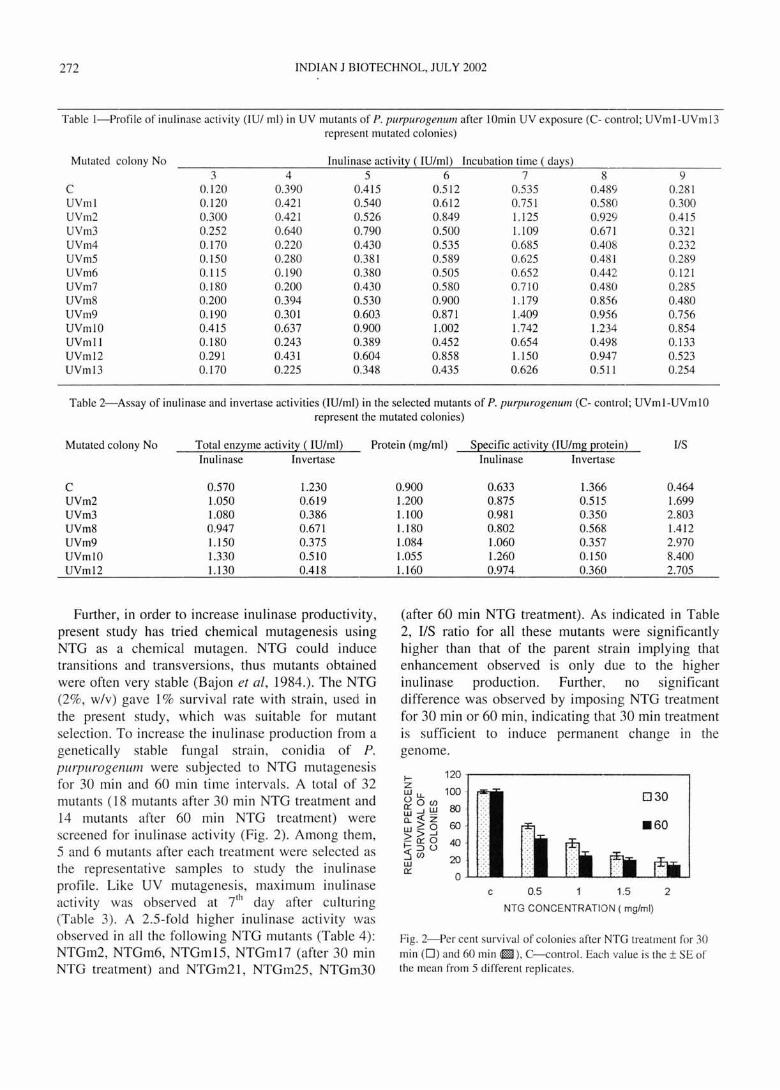
Table I-Profile of inulinase activity (lUI ml) in UV mutants of P. purpurogenu111 after 10min UV exposure (C- control ; UVm I-UVm 13 represent mutated colonies)
Mutated colony No Inulinase activit:t ( IUlml) Incubation time ( da:ts) 3 4 5 6 7 8 9
C 0.120 0.390 0.415 0.512 0.535 0.489 0.28 1 UVml 0.120 0.42 1 0.540 0.6 12 0.75 1 0.580 0.300 UVm2 0.300 0.421 0.526 0.849 1. 125 0.929 0.4 15 UVm3 0.252 0.640 0.790 0.500 1.109 0.671 0.321 UVm4 0.170 0.220 0.430 0.535 0.685 0.408 0.232 UVm5 0.150 0.280 0.381 0.589 0.625 0.481 0.289 UVm6 0.11 5 0.190 0.380 0.505 0.652 0.442 0.121 UVm7 0.180 0.200 0.430 0.580 0.710 0.480 0.285 UVm8 0.200 0.394 0.530 0.900 1.179 0.856 0.480 UVm9 0.190 0.301 0.603 0.871 1.409 0.956 0.756 UVmlO 0.415 0.637 0.900 1.002 1.742 1.234 0.854 UVml1 0. 180 0.243 0.389 0.452 0.654 0.498 0.133 UVml2 0.291 0.431 0.604 0.858 1.150 0.947 0.523 UVm l3 0.170 0.225 0.348 0.435 0.626 0.511 0.254
Table 2-Assay of inulinase and invertase act ivities (IUlml) in the selected mutants of P. purpurogenu111 (C- control; UVml-UVm 10 represent the mutated colonies)
Mutated colony No Total enz:tme activit:t ( IU/ml ) Protein (mg/ml ) SQecific activit:t (lUlmg Qrotein) IIS Inulinase Invertase
C 0.570 1.230 UVm2 1.050 0.619 UVm3 1.080 0.386 UVm8 0.947 0.671 UVm9 1.150 0.375 UVmlO 1.330 0.510 UVml2 1.130 0.418
Further, in order to increase inulinase productivity, present study has tried chemical mutagenesis using NTG as a chemical mutagen. NTG could induce transitions and trans versions, thus mutants obtained were often very stable (Bajon et ai, 1984.). The NTG (2%, w/v) gave 1 % survival rate with strain, used in the present study, which was suitable for mutant select ion . To increase the inulinase production from a genetically stable fungal strain, conidia of P. purpurogenum were subjected to NTG mutagenesis for 30 min and 60 min time intervals. A total of 32 mutants ( 18 mutants after 30 min NTG treatment and 14 mutants after 60 min NTG treatment) were screened for inulinase activity (Fig. 2). Among them, 5 and 6 mutants after each treatment were selected as the representative samples to study the inulinase profile. Like UV mutagenesis, maximum inulinase activity was observed at 7th day after culturing (Table 3). A 2.5-fold higher inulinase act ivity was observed in all the following NTG mutants (Table 4): NTGm2, NTGm6, NTGmI5 , NTGm17 (after 30 min NTG treatment) and NTGm21, NTGm25, NTGm30
Inulinase Invertase
0.900 0.633 1.366 0.464 1.200 0.875 0.515 1.699 1.l00 0.981 0.350 2.803 1.180 0.802 0.568 1.412 1.084 1.060 0.357 2.970 1.055 1.260 0.150 8.400 1.160 0.974 0.360 2.705
(after 60 min NTG treatment). As indicated in Table 2, liS ratio for all these mutants were significantly higher than that of the parent strain implying that enhancement observed is only due to the higher inulinase production . Further, no significant difference was observed by imposing NTG treatment for 30 min or 60 min , indicating that 30 min treatment is suffici ent to induce permanent change in the genome.
I- 120 Z
100 w LL (00) IY-,w 80 w« -Cl.>z 60 w- O »-' -IYO 40 I-::Jo ::SO)
20 w IY
0
c 0.5 1.5 2
NTG CONCENTRATION ( mg/ml)
Fig. 2- Per cent survival of colon ies after NTG treatment for 30 min (0 ) and 60 min iIE.I ). C-{;ontrol. Each value is the ± SE of the mean from 5 different replicates.

SHARMA et al: ENHANCED INULINASE PRODUCTION BY MUTAGENESIS 273
Table 3--Profi le of inulinase activity (lUlml) in NTG mutants of P. purpurogenulIl (C- control; NTGm I-NTGm32 represent the mutated colonies)
Mutated Treatment time Inulinase activity ( IUlml) colony No Incubation time (da;ts)
3 4 5 6 7 8 9
C 0.003 0.123 0.399 0.455 0.515 0.433 0.202 NTGml 30 min 0.011 0.173 0.357 0.562 0.752 0.438 0.238 NTGm2 0.020 0.357 0.410 0.732 1.1 32 0.627 0.412
NTGm3 0.01 8 0.2 10 0.385 0.498 0.816 0.517 0.380
NTGm4 0.008 0.215 0.131 0.135 0.236 0.162 0.123
NTGm5 0.01 3 0.311 0.43 1 0.512 0.832 0.713 0.312
NTGm6 0.027 0.Dl8 0.521 0.821 1.120 0.812 0.412
NTGm7 0.100 0.230 0.336 0.530 0.688 0.632 0.438
NTGm8 0.008 0.111 0.246 0.488 0.751 0.586 0.321
NTGm9 0.009 0.11 8 0.258 0.438 0.647 0.32 1 0.121
NTGm10 0.180 0.230 0.489 0.558 0.920 0.741 0.381
NTGml1 0.011 0.489 0.655 0.889 1.000 0.851 0.453 NTGm12 0.004 0.121 0.257 0.321 0.611 0.42 1 0.290
NTGml3 0.007 0. 141 0.23 1 0.431 0.658 0.351 0.255 NTGml4 0.008 0.151 0.3 15 0.512 0.781 0.58 1 0.357 NTGml5 0.243 0.489 0.730 0.988 1.230 0.845 0.458 NTGm l6 0.013 0.139 0.322 0.538 0.890 0.601 0.341 NTGml7 0.132 0.34 1 0.479 0.887 1.131 0.921 0.432 NTGml8 0.048 0.223 0.387 0.599 0.821 0.633 0.225 NTGml9 60 min 0.015 0.125 0.258 0.398 0.579 0.45 1 0.354 NTGm20 0.0 18 0.1310 0.278 0.458 0.784 0.548 0.325 NTGm2 1 0.023 0.275 0.310 0.668 0.827 0.589 0.430 NTGm22 0.208 0.437 0.789 0.989 1.125 0.851 0.451 NTGm23 0.117 0.200 0.375 0.675 0.860 0.500 0.3 12 NTGm24 0.012 0.210 0.431 0.5 11 0.789 0.454 0.322 NTGm25 0.141 0.3 18 0.632 0.832 1.025 0.852 0.455 NTGm26 0.03 1 0.188 0.339 0.521 0.632 0.420 0.200 NTGm27 0.0 19 0.192 0.360 0.532 0.679 0.499 0.289 NTG m28 0.11 0 0.232 0.522 0.689 0.723 0.458 0.352 NTGm29 0.018 0.201 0.377 0.522 0.68 1 0.53 1 0.302 NTGm30 0.282 0.47 1 0.732 0.954 1.125 0.895 0.546 NTGm31 0.018 0.35 1 0.5 12 0.552 0.633 0.521 0.324 NTGm32 0.111 0.225 0.325 0.552 0.712 0.451 0.352
Table 4 -Assay of inulinase and invertase ac ti vities (lUlml ) in the selected mutants of P. pUlpurogel/ulII (C--control; NTGm2-NTGm30 represent the mutated colonies)
Mutated Treatment Total enzyme act ivity Protein (mg/ml) Specific activity liS ratio colony No time (lUI ml ) (lU/mg Qrotei n)
Inulinase Invertase Inulinase In vertase
C 0.525 1.100 0.868 0.605 1.267 · 0.477 NTGm2 30 min 1.122 0.58 1 1.058 1.060 0.553 1.917 NTGm6 1.122 0.69 1 1.099 1.021 0.628 1.623 NTGml1 0.826 0.432 1.123 0.735 0.384 1.914 NTGml5 1.133 0.5 12 1.1 2 1 1.010 0.457 2.210 NTGml7 1.222 0.732 1.133 1.078 0.884 1.220 NTGm2 1 60 min 1.113 0.432 1.043 1.067 0.414 2.577 NTGm22 0.773 0.3 13 0.912 0.847 0.343 2.470 NTGm23 0.727 0.423 0.813 0.894 0.520 1.720 NTGm24 0.660 0.232 0.933 0.707 0.248 2.850 NTGm25 1.130 0.430 1.0 II 1.11 8 0.425 2.630 NTGm30 1.210 0.510 1.133 1.068 0.45 1 2.340

274 INDIAN J BIOTECHNOL, JULY 2002
According to authors, there are no other reports on mutagenic enhancement in inulinase production in P. purpurogenum. The results of present communication can be compared with studies (Fiedurek et aI, 2000; Bajon et aI, 1984) for A. fumigatus and Pichia polymorpha, where 2-fold increase in inulinase production after NTG mutagenesis have been observed. Furthermore, studies on extracellularly produced inulinase in P. purpurogenum (Sharma et at, 1998) show that this enzyme possesses higher thermostabilty by maintaining 95% residual activity after Ihr incubation at 60°C and it shows maximum inulinase activity at 50°C. Hence improved strain of this study, which has maintained thermostability , must be a significant one for industry where inulin hydrolysis is carried out at higher temperature (about 60°C) to prevent microbial contamination, and also because of higher solubility of inulin at elevated temperature.
These hyperinulinase mutants were retested twice for their stability and, after UV and NTG mutagenesis, were found stable. The stability of all these mutants was also verified after 3-months subculturing. No significant change in inulinase activity was observed in UV and NTG mutants. Therefore, using these mutants would be a logical step for large-scale production of fructooligosaccharides. Further, studies are in progress on enhancing the production and thermostability of P. purpurogenum inulinase by site directed mutagenesis. Purification
and immobilization of this enzyme is also underway for exploiting its commercial potential.
Acknowledgement The authors are grateful to Department of
Biotechnology, Government of India, New Delhi , for providing the financial assistance for research work.
References Bajon A M et ai, 1984. Isolation of an inul inase depressed mutant
of Pichia polymorpha for the production of fructose. Biotechnol Bioeng, 26, 128- 133.
Fiedurek J et ai, 2000. Direct methods for selecting mutants with increased production of in vertase from mutagenised culture of Aspergillusjumigafus. J Basic Microbiol , 40, 111 -118.
Guiraud J P, 1981 . Utilization des lev ures pour la valorisation industrielle de polyfructosanes de type inliline. These de Doctorat d ' etat. Universite Science et Techniques du Languedoc, France.
Lowry 0 H et ai, 1951. Protein estimation with Folin phenol reagent. J Bioi Chem , 193,265-275.
Nakamura T et ai, 1994. Occurrence of two fo rms of extracellular endoinulinmase from Aspergillus niger mutant 8 17. J Ferment Bioeng, 78, 134-139.
Nelson N, 1944. A photometric adaptation of Somogyi method for the determination of glucose. J Bioi Chern, 153,375-380.
Sharma D et ai, 1998. Themostabil ity of extracellular inlilinases from Aspergillus and Penicillium . Indian J Microbiol , 38, 235-236.
Vandamme E J & Derycke D G. 1983. Microbial inulinases; fermentation process, properties and applications. Adv Appl Microbiol, 29, 139-176.
Vishwanthan P & Kulkarni P R, 1995. Enhancement of inulinase production by Aspergillus niger Van Teighem. J Appl Bacteriol, 78, 384-386.