enzyme. halo enzyme= apoenzyme+ cofactor coenzyme( non covalane) organic prostatic(covalan) cofactor...
TRANSCRIPT

Enzyme

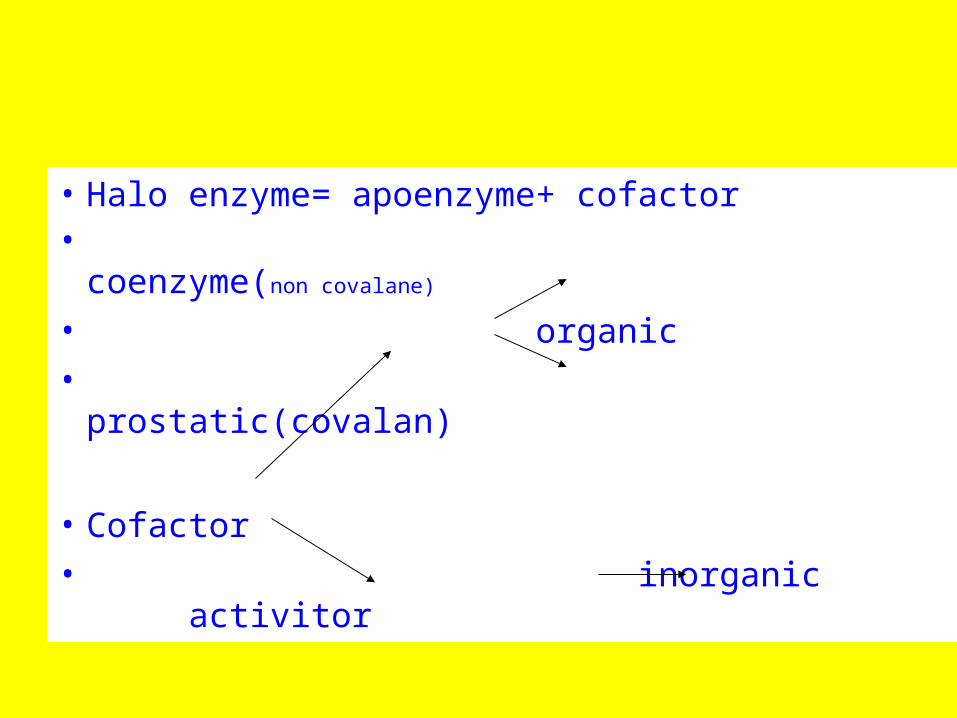
• Halo enzyme= apoenzyme+ cofactor• coenzyme(non covalane)
• organic
• prostatic(covalan)
• Cofactor
• inorganic activitor

• Artificial enzymes, termed abzymes, have been synthesized by making antibodies against chemicals that are transition-state analogs.
• Abzymes have been designed to catalyze over 100 different chemical reactions

• Metalloenzymes contain a tightly bound transition metal, such as Zn2+ or Fez+,
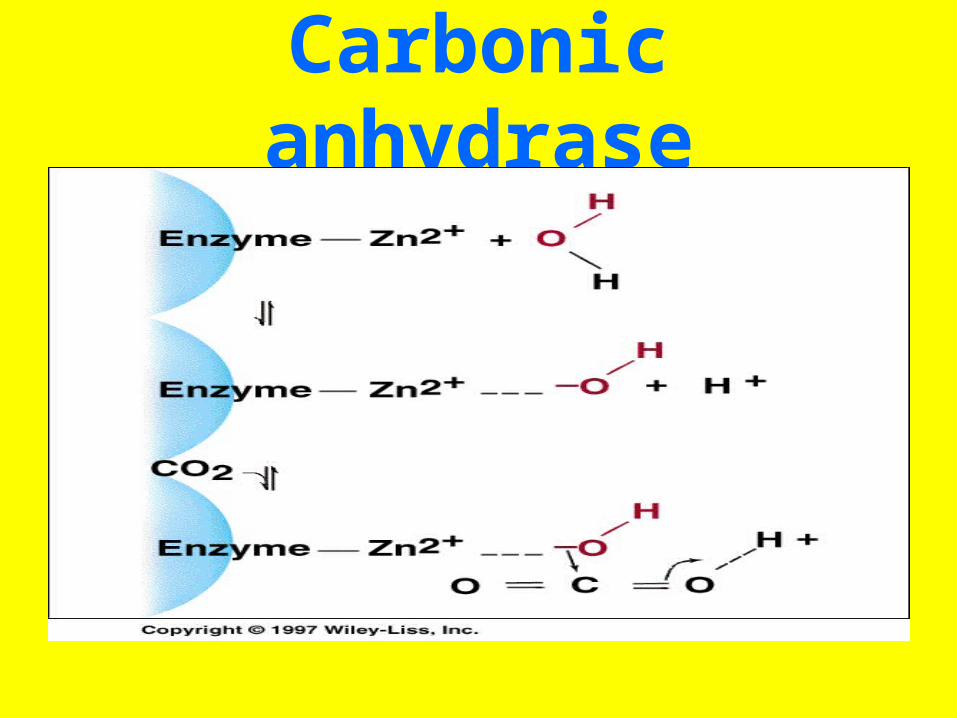
Carbonic anhydrase


• In mid-2004, information was available for over 83,000 different enzymes from 9800 different organisms;
• International Union of Biochemistry and Molecular Biology (IUBMB) to characterize each enzyme

Enzyme nomenclature
• 1- Ordinary • Substrate name+ enzyme action+ase• Urea + H2O 2NH3+H2CO3• Urea hydrolase = Urease
• 2- Classic IUB• EC=1.2.3.4• Butyrylcholinesterase BChE (EC.3.1.1.8 • acetylcholinesterase (ACHE, EC.3.1.1.7)
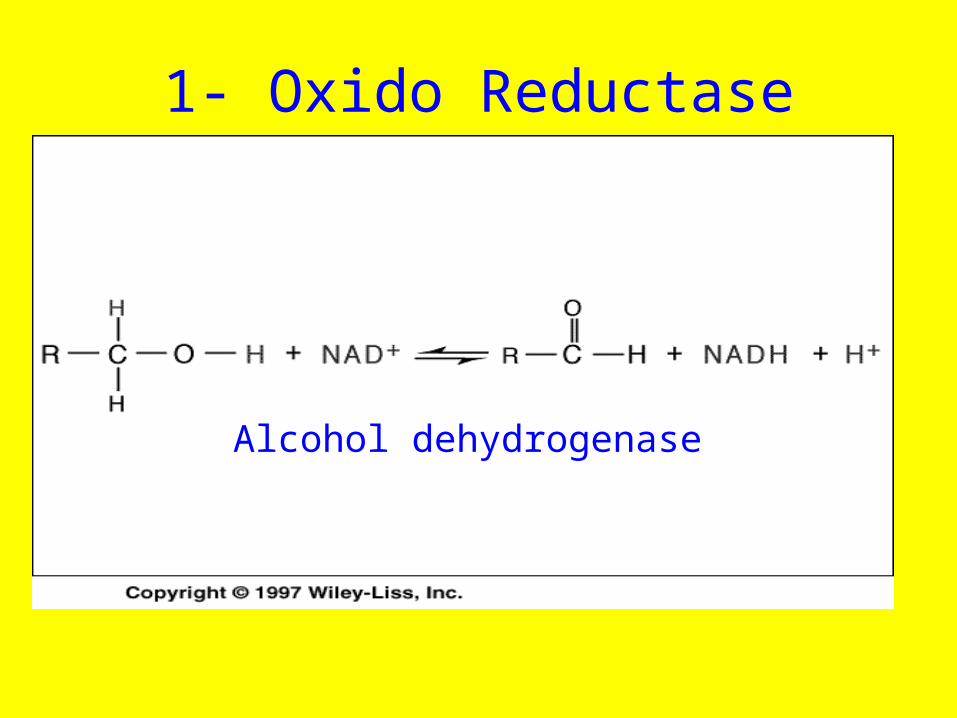
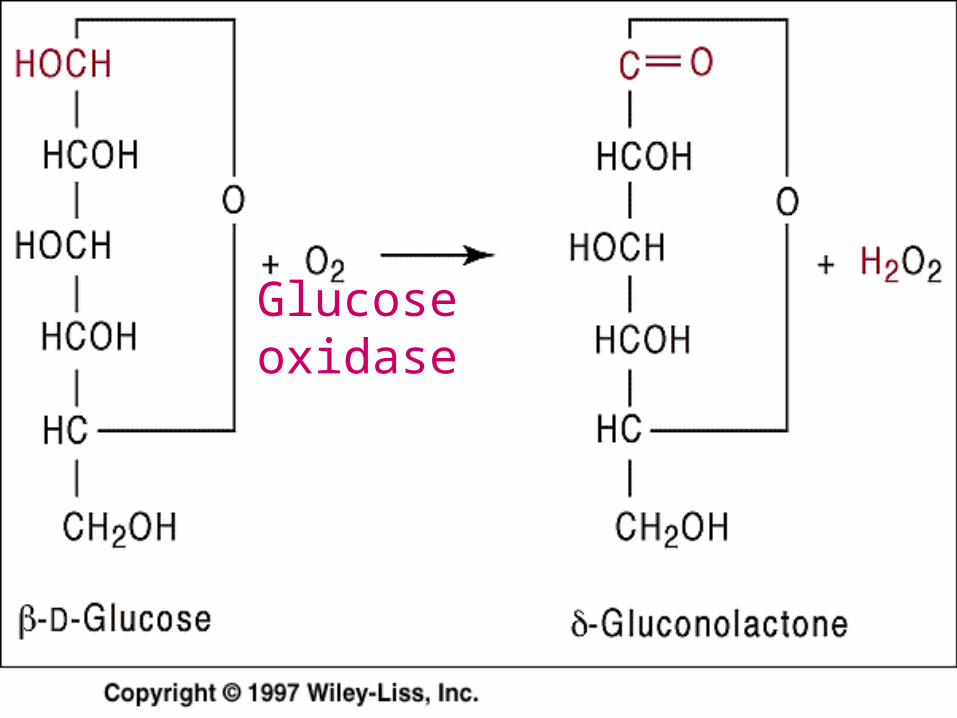
1- Oxido Reductase
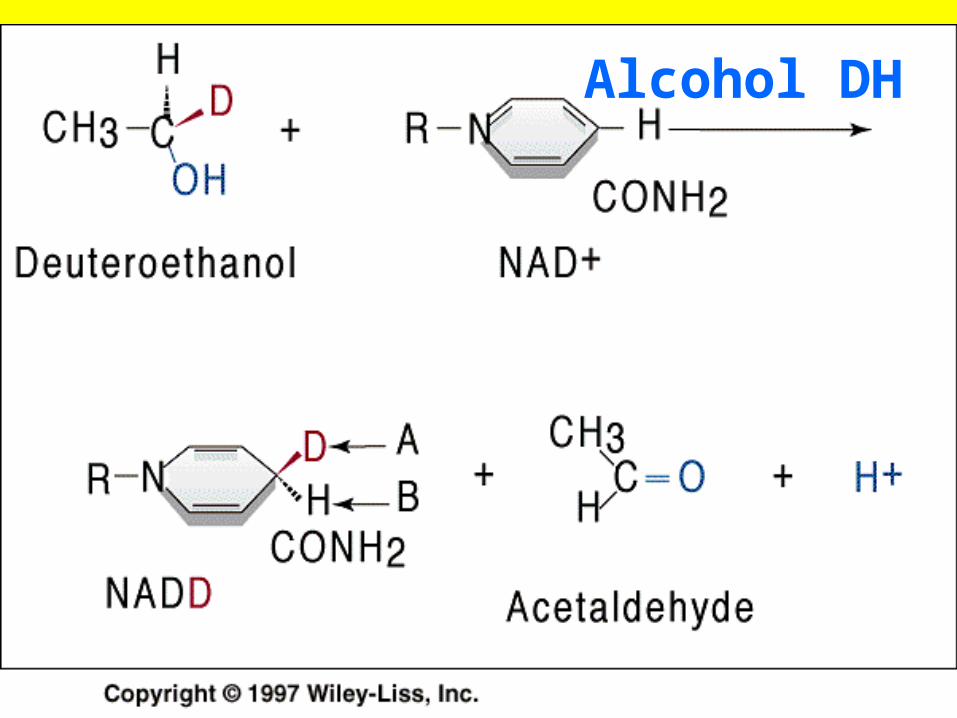
Alcohol dehydrogenase

• Alcohol dehydrogenase an enzyme that oxidizes an alcohol to an aldehyde has the IUBMB number 1.1.1.1.T his indicates
• ( 1) that the enzyme is involved in an oxidation-reduction reaction (first number),
• (2) it removes hydrogen as a hydride ion with NAD+ as the electron accepror (second n umber),
• and (3) substrates for the enzyme can be most primary alcohols (third number).
• (4 ) The last number is reserved to different substrat each enzyme that catalyzes the same overall reaction but with different substrates. Lactate dehydrogenas e (l.l .1.27
Alcohol DH
Glucose oxidase
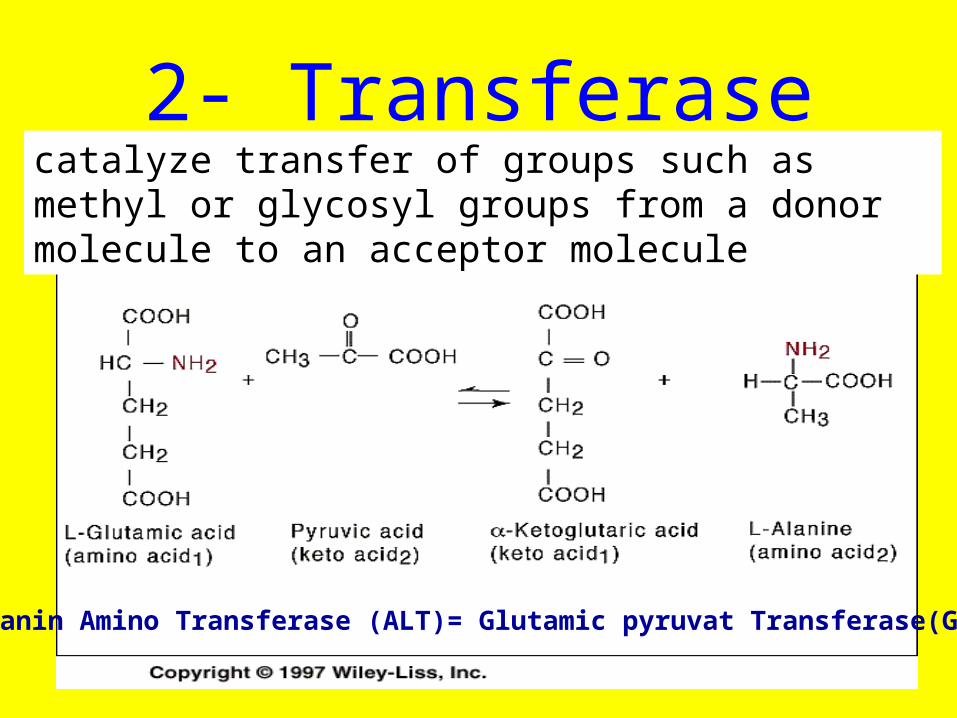
2- Transferase
Alanin Amino Transferase (ALT)= Glutamic pyruvat Transferase(GPT)
catalyze transfer of groups such as methyl or glycosyl groups from a donor molecule to an acceptor molecule
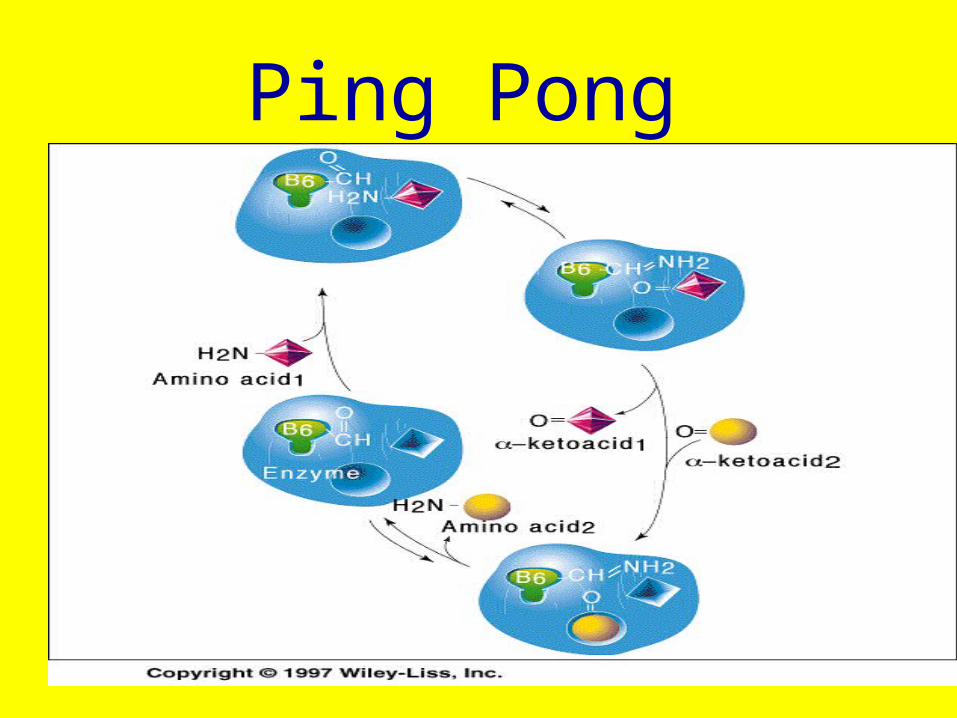
Ping Pong
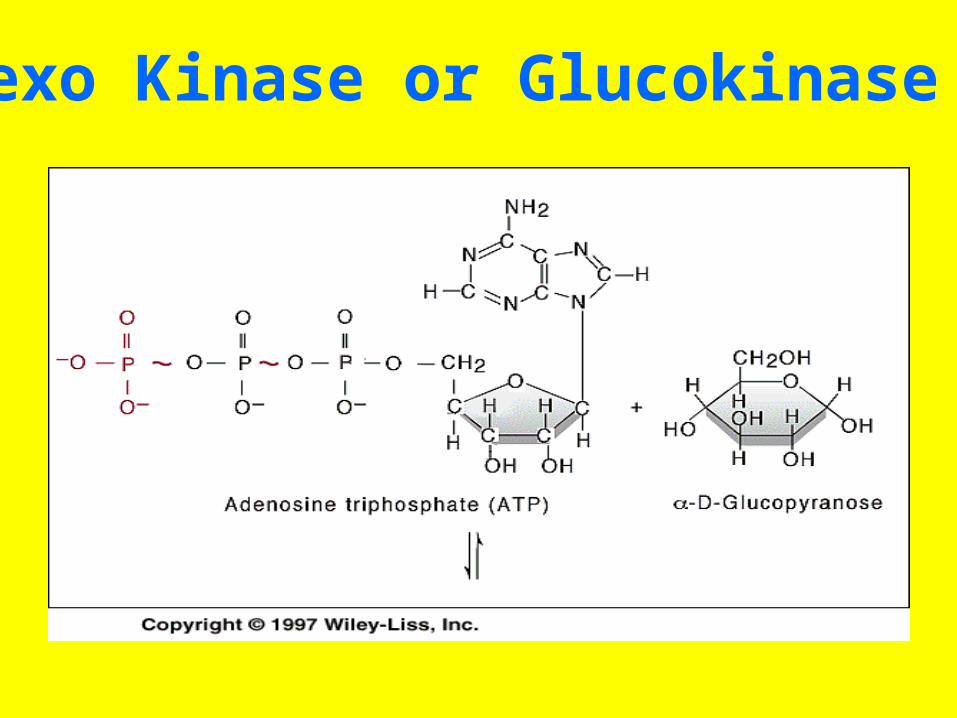
Hexo Kinase or Glucokinase
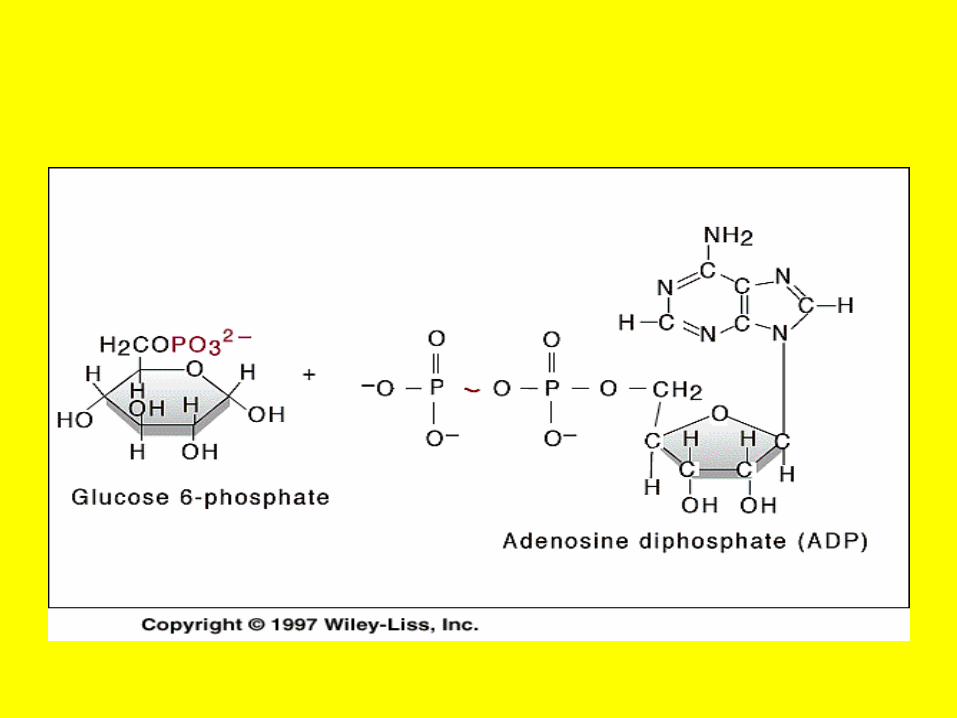

• For example, the enzyme commonly called “hexokinase” is designated “ATP:D-hexose-6-phosphotransferase
• E.C. 2.7.1.1.” This identifies hexokinase as a member of class 2 (transferases), subclass 7 (transfer of a
• phosphoryl group), • sub-subclass 1 (alcohol is the phosphoryl
acceptor). • Finally, the term “hexose-6” indicates that
the alcohol phosphorylated is that of carbon six of a hexose.

3- Hydrolase• catalyze the hydrolytic cleavage of C-C, C-O, C-N, P-
O, and certain other bonds, including acid anhydride bonds.
• ButyrylCholine +H2O Choline+Butyric Acid
• Butyrylcholinesterase BChE (EC.3.1.1.8)
• acetylcholinesterase (ACHE, EC.3.1.1.7)
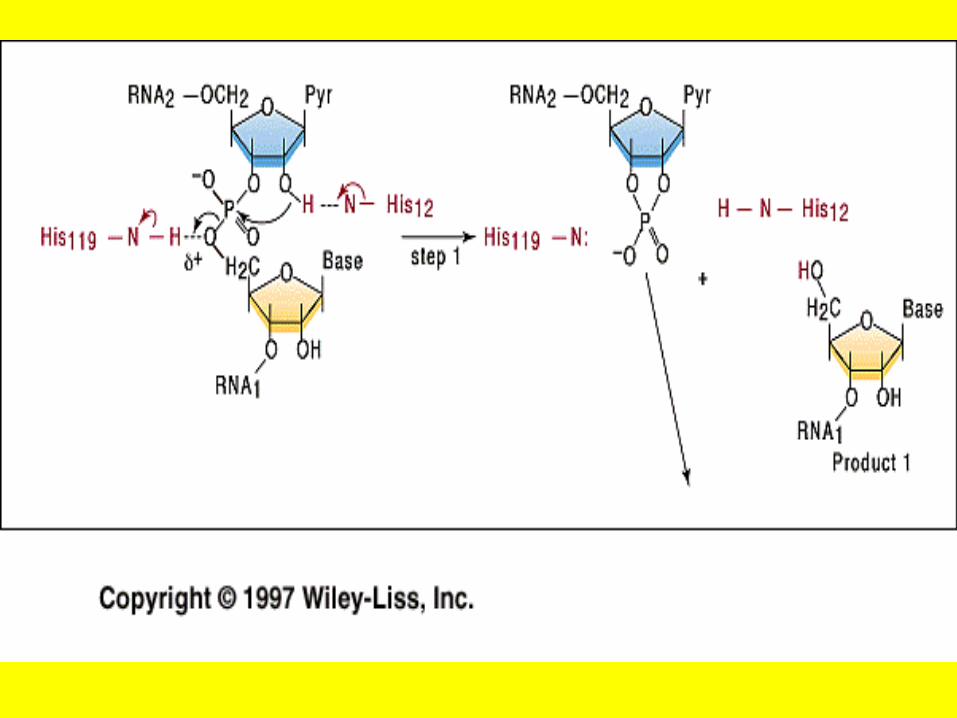
• RNases and DNases are enzymes that hydrolyze the phospho—ester bond
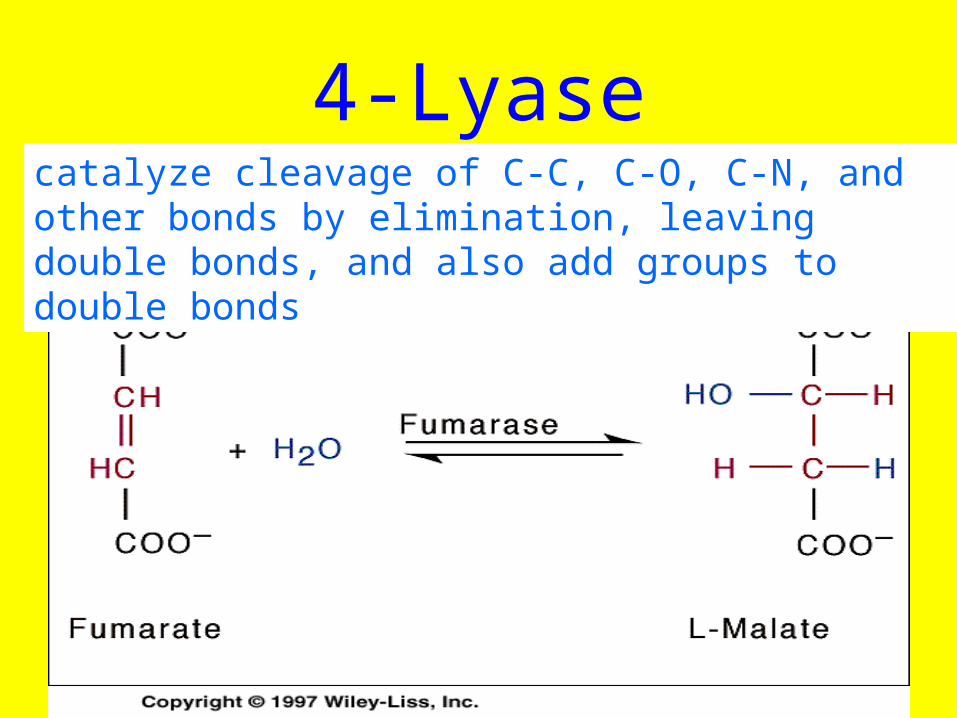
4-Lyasecatalyze cleavage of C-C, C-O, C-N, and other bonds by elimination, leaving double bonds, and also add groups to double bonds

Hydrolysis of Asparaginaseand Leukemia
• Some forms of juvenile leukemia require the nonessential amino acid asparagine
• hydrolase enzyme that converts asparagine into aspartic acid
• This form of (the enzyme was encoded in polyethlenglycol)
• the enzyme is commercially available and is approved for human treatment.

5- Isomerasecatalyze geometric or structural changes within a single molecule (moving a group or a double bond within the same molecule)

• Glucose6Phosphate Fructose6Phosphate
• Hexose6Phosphate isomerase
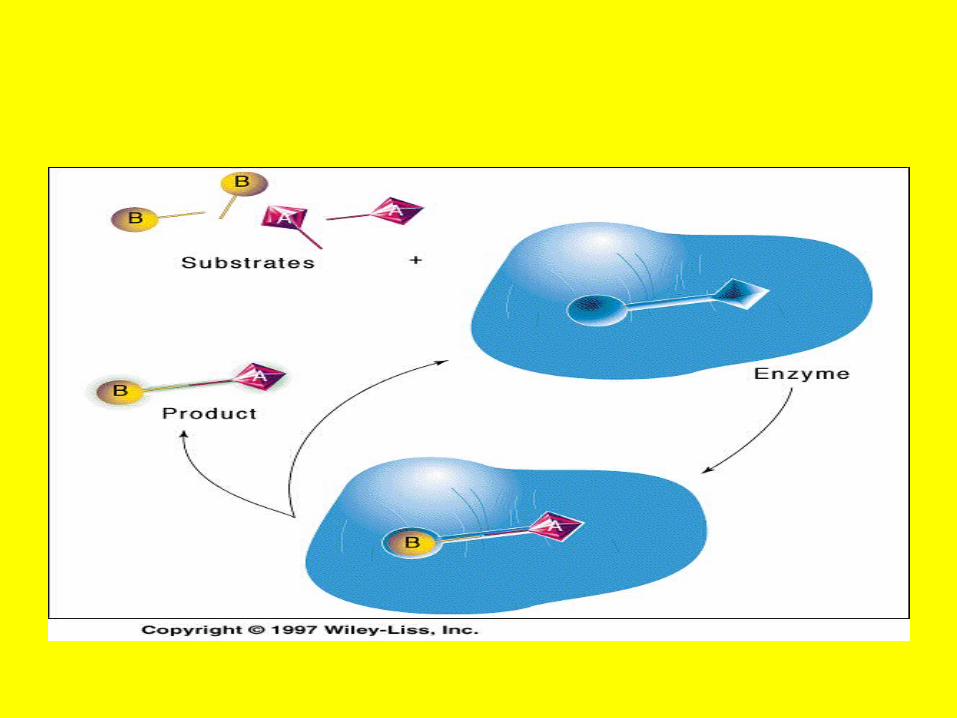

6- ligase or synthetase• Urea+NH3+ATP Carbomyl phoshate +
• ADP
• Cabomylphosphate synthetase
• Oxaloacetate+AcetylCoA Citrate+CoA
• Citrate syntase


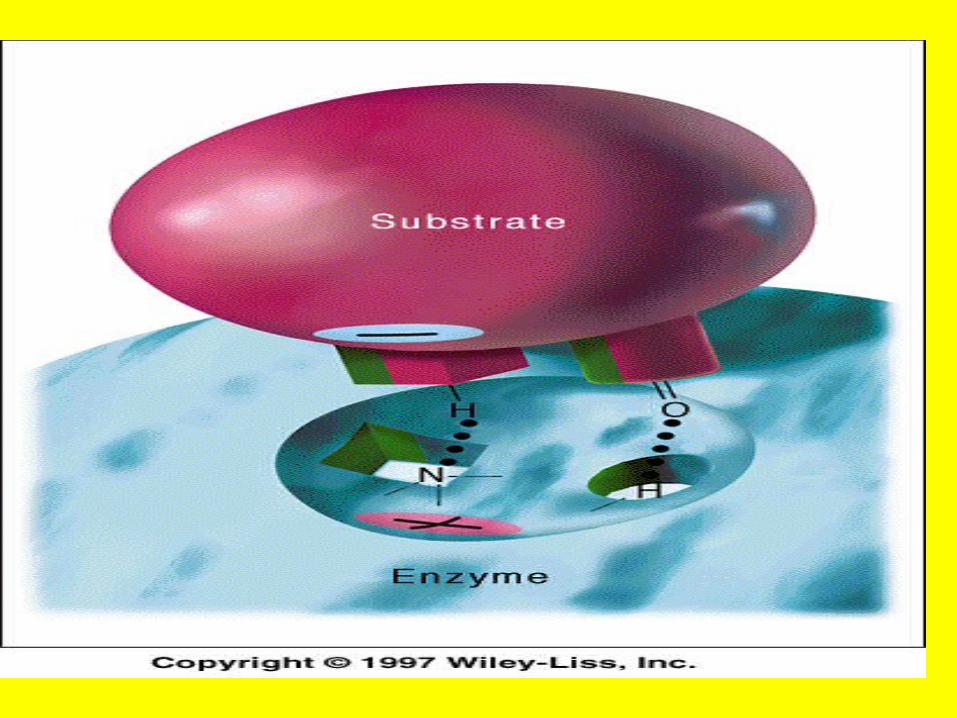
Induced-fit model
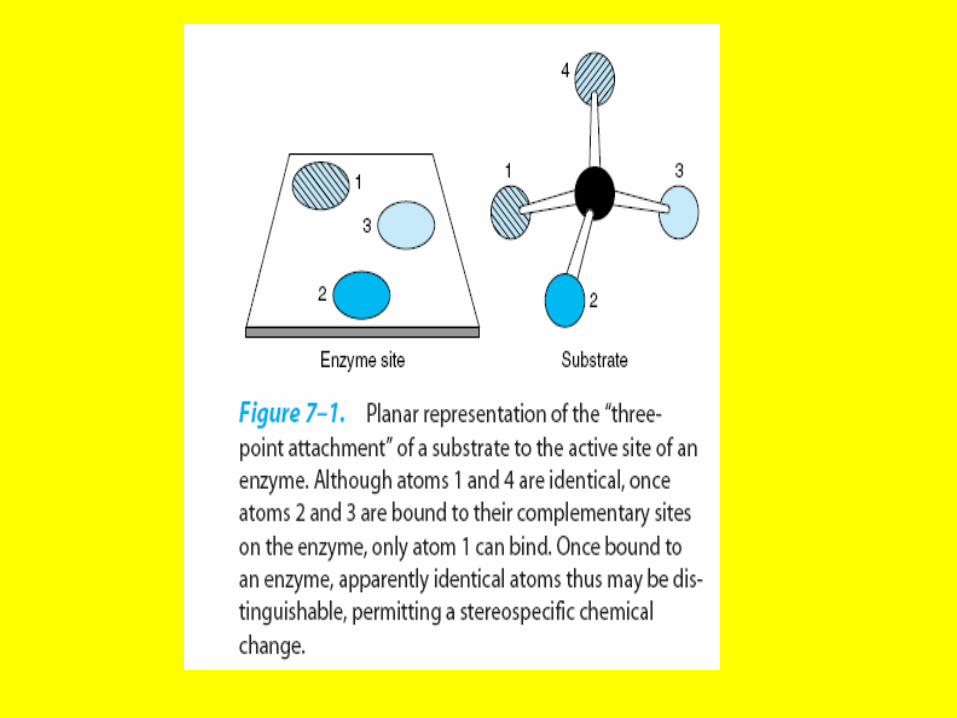
Lack-and-key model
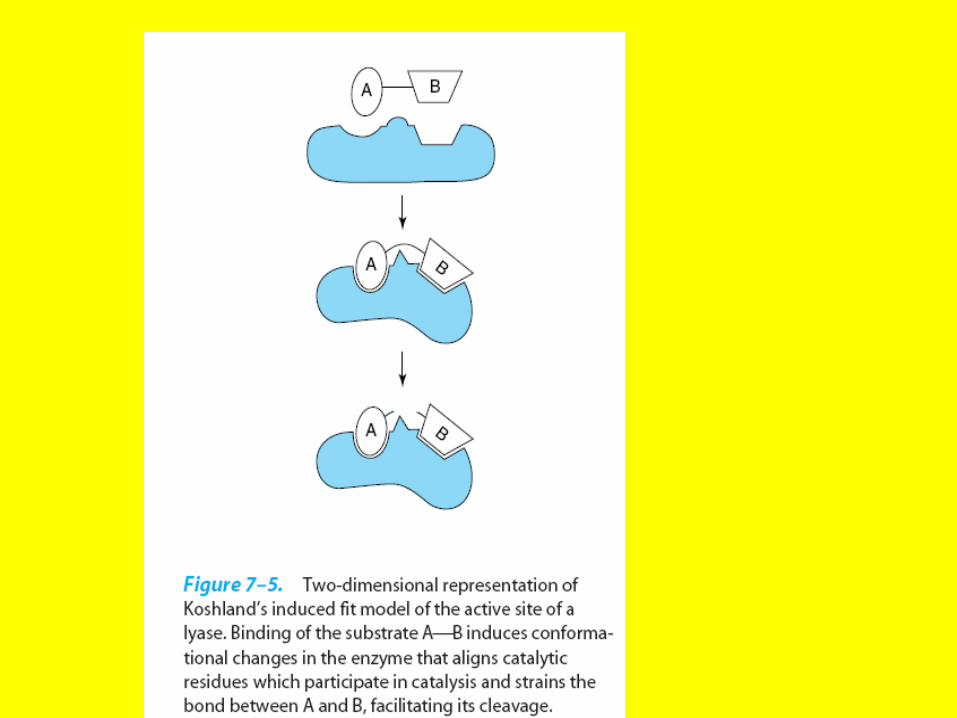

• Hydrophobic portions of the substrate bind such that they are in a hydrophobic portion of the protein, referred to as a
hydrophobic pocket

• ISOZYMES ARE DISTINCT ENZYME• FORMS THAT CATALYZE THE• SAME REACTION• Higher organisms often elaborate several
physically distinct versions of a given enzyme, each of which catalyzes the same reaction.
• Like the members of other protein families, these protein catalysts or isozymes arise through gene duplication.
• Isozymes may exhibit subtle differences in properties such as sensitivity to

Enzyme kinetic
•E+S ES E+P
Active site
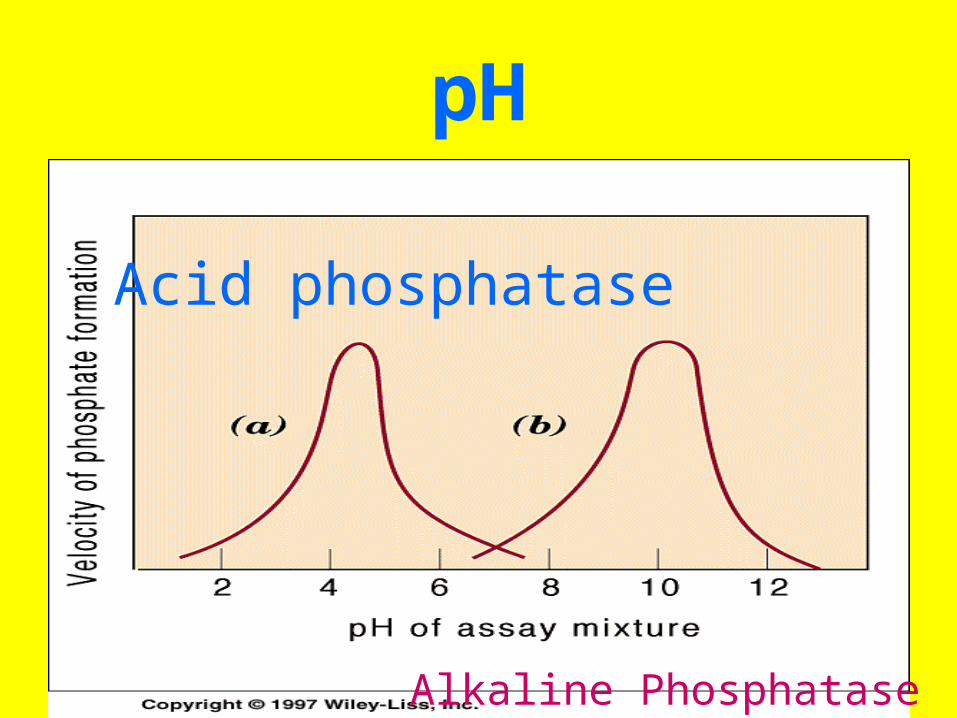
pH
Acid phosphatase
Alkaline Phosphatase
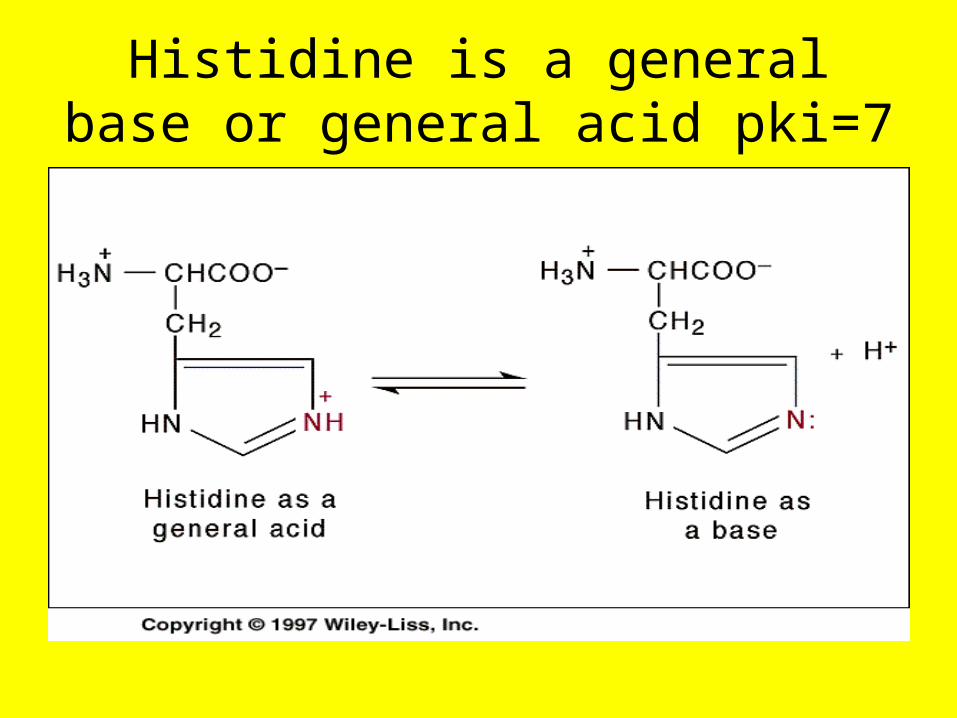
Histidine is a general base or general acid pki=7
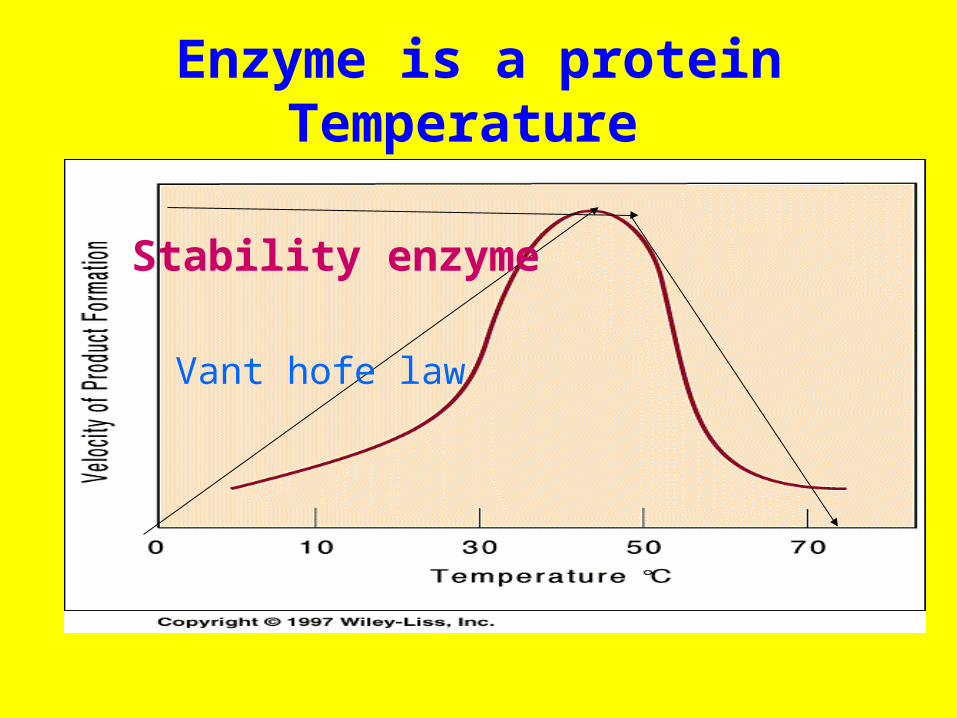
Enzyme is a proteinTemperature
Stability enzyme
Vant hofe law

[E]

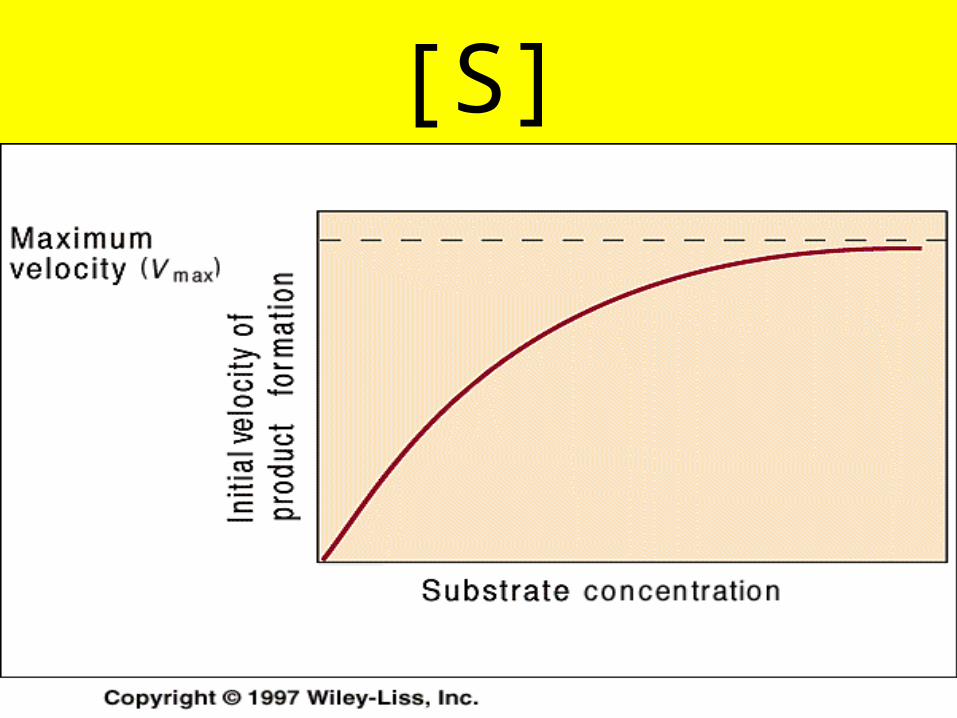
[S]
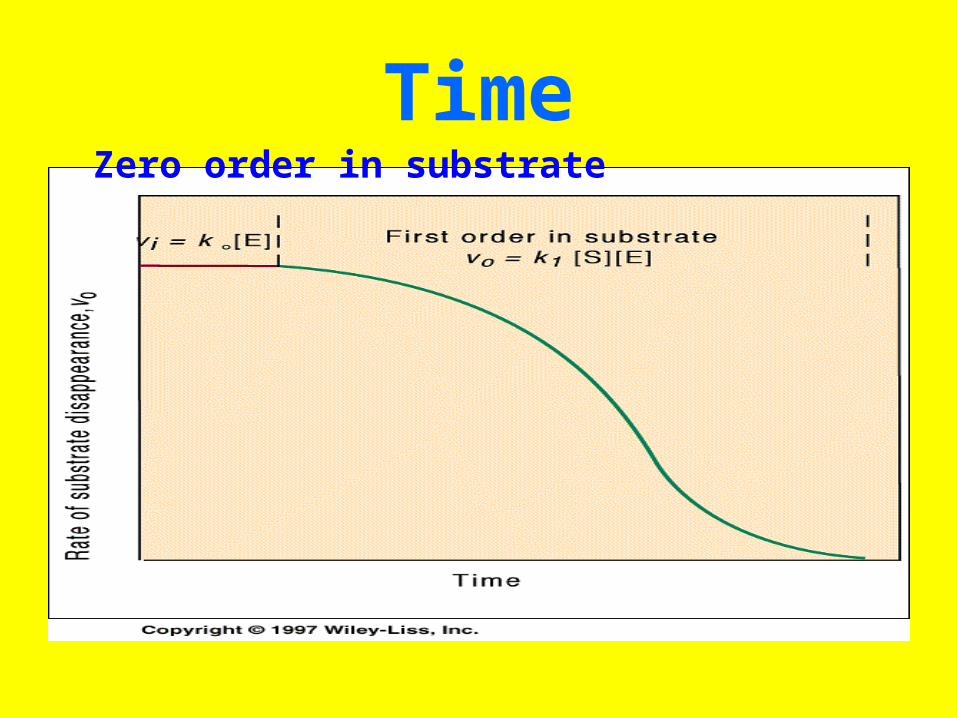
TimeZero order in substrate


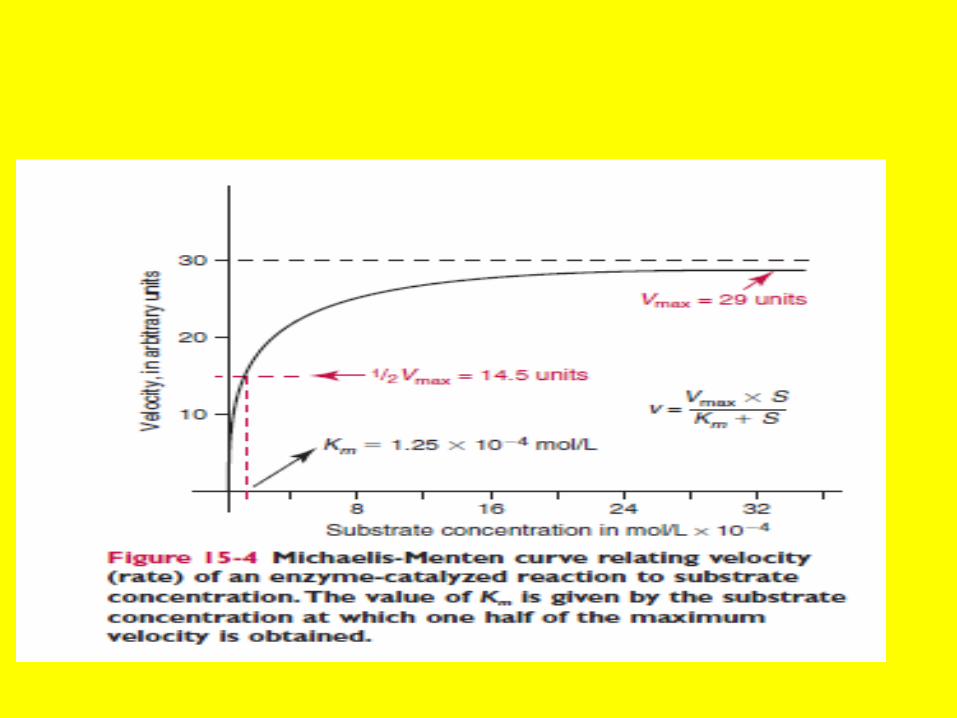

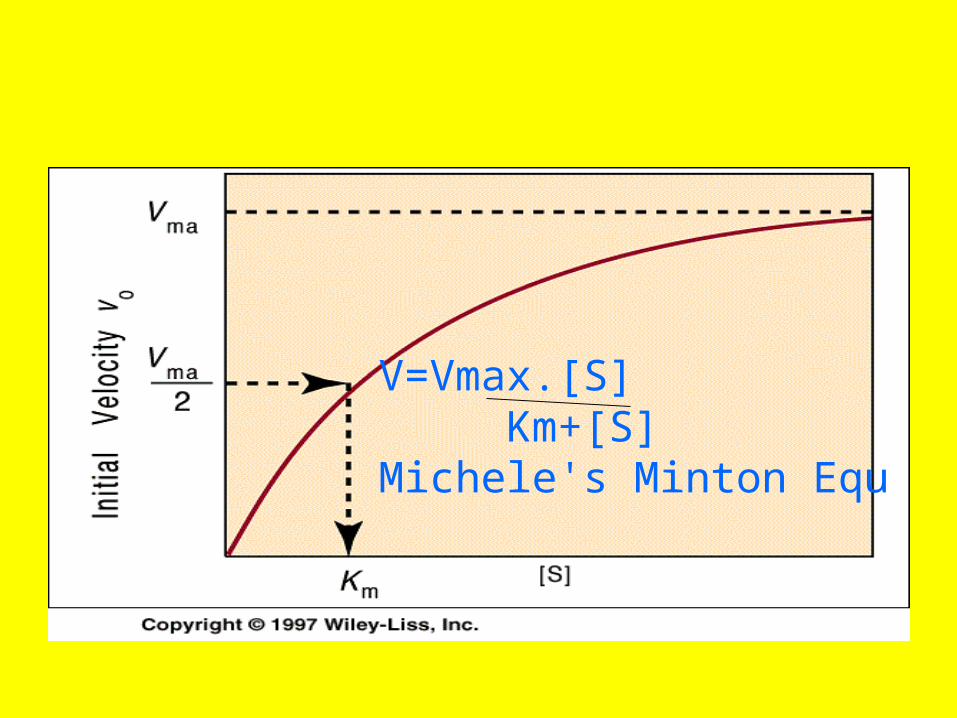
V=Vmax.[S] Km+[S]Michele's Minton Equ
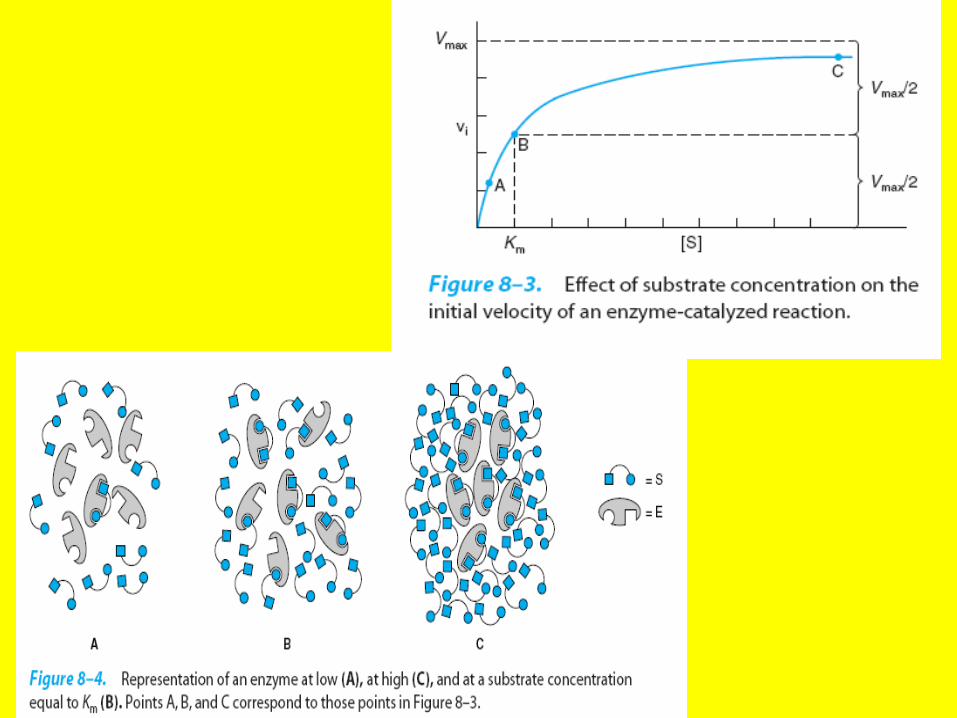


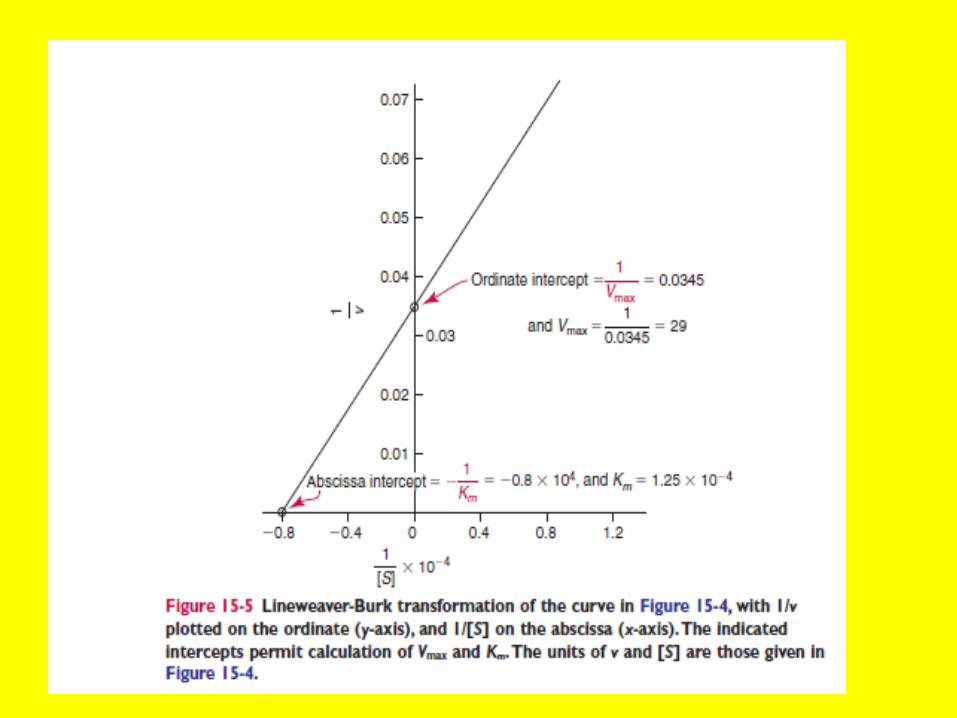
Line Weaver Burk Equ

• An example of the importance of Km is the physiological utilization of glucose. Glucose can be phosphorylated by two different kinases to form glucose6 –phosphate Liver contains
• both hexokinase and glucokinase that catalyze the identical reaction of
• glucose+ ATP glucose6 -phosphate+ ADP.
• For hexokinase the Km-for glucose is 0. mM whereas for glucokinase it is 5 mM. lf then the concentration of blood sugar is low, as occurs in the fasted state, hexokinase is used to phosphorylated glucose, but when blood glucose increases
• after feeding, the high K-enzyme also function



The Hill Equation Describes the Behaviorof Enzymes That Exhibit Cooperative
Binding of Substrate
• While most enzymes display the simple saturation kinetics depicted in Figure 8–3 and are adequately described by the Michaelis-Menten expression,
• some enzymes bind their substrates in a cooperative fashion analogous to the binding of oxygen by hemoglobin

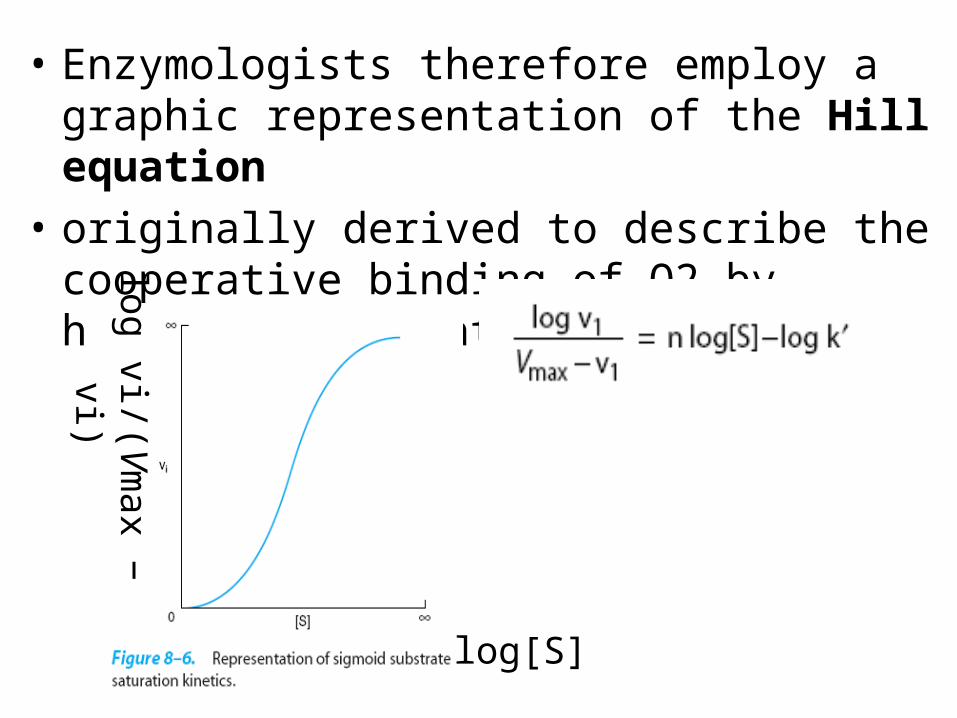
• Enzymologists therefore employ a graphic representation of the Hill equation
• originally derived to describe the cooperative binding of O2 by hemoglobin. Equation (43
log vi/(Vm
ax − vi)
log[S]

• A graph of log vi/(Vmax − vi) versus log[S] gives a straight line (Figure 8–7), where the slope of the line n is the Hill coefficient,
• an empirical parameter whose value is a function of the number, kind, and strength of the interactions of the multiple substrate-binding sites on the enzyme.
• When n = 1, all binding sites behave independently, and simple Michaelis-Menten kinetic behavior is observed.
• If n is greater than 1, the enzyme is said to exhibit positive cooperatively

• Binding of the first substrate molecule then enhances the affinity of the enzyme for binding additional substrate.
• The greater the value for n, the higher the degree of cooperativity and the more sigmoidal will be the plot of vi versus [S].
• A perpendicular dropped from the point where the y term log vi/(Vmax − vi) is zero intersects the x axis at a substrate concentration termed S50,
• the substrate concentration that results in half-maximal velocity.
• S50 thus is analogous to the P50 for oxygen binding to hemoglobin


• Inhibitors and activators

Inhibitors•Reversible
•Irreversible

Competitive Inhibitors TypicallyResemble Substrates
• The effects of competitive inhibitors can be overcome by raising the concentration of the substrate.
• Most frequently, in competitive inhibition the inhibitor, I, binds to the substrate-binding portion of the active site and blocks access by the substrate.
• The structures of most classic competitive inhibitors therefore tend to resemble the structures of a substrate and thus are termed substrate analogs.
• Inhibition of the enzyme succinate dehydrogenase by malonate illustrates competitive inhibition by a substrate analog
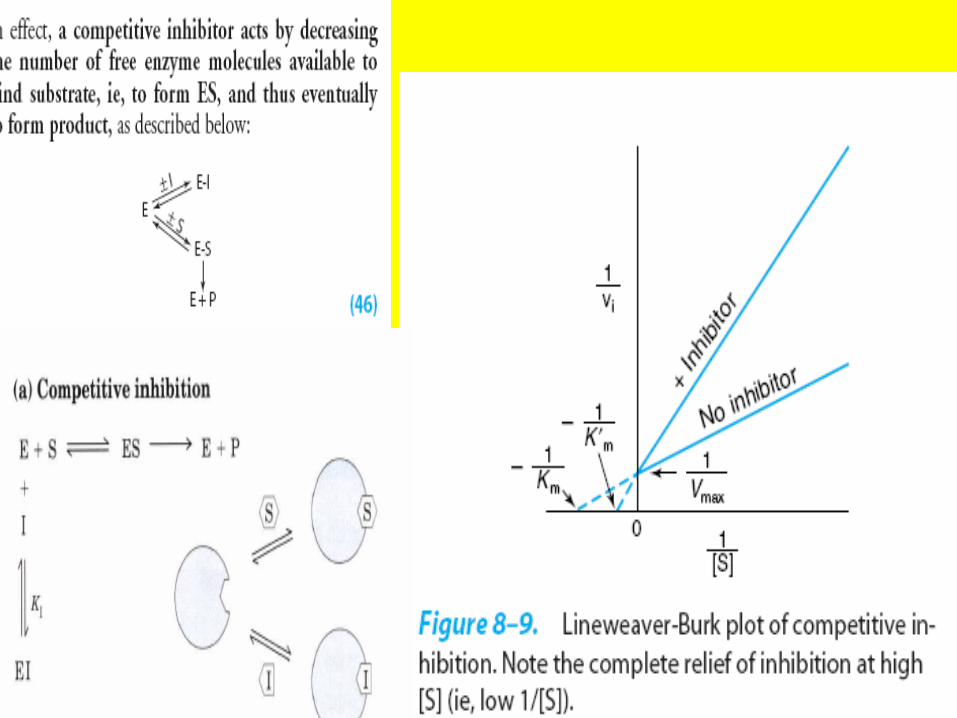
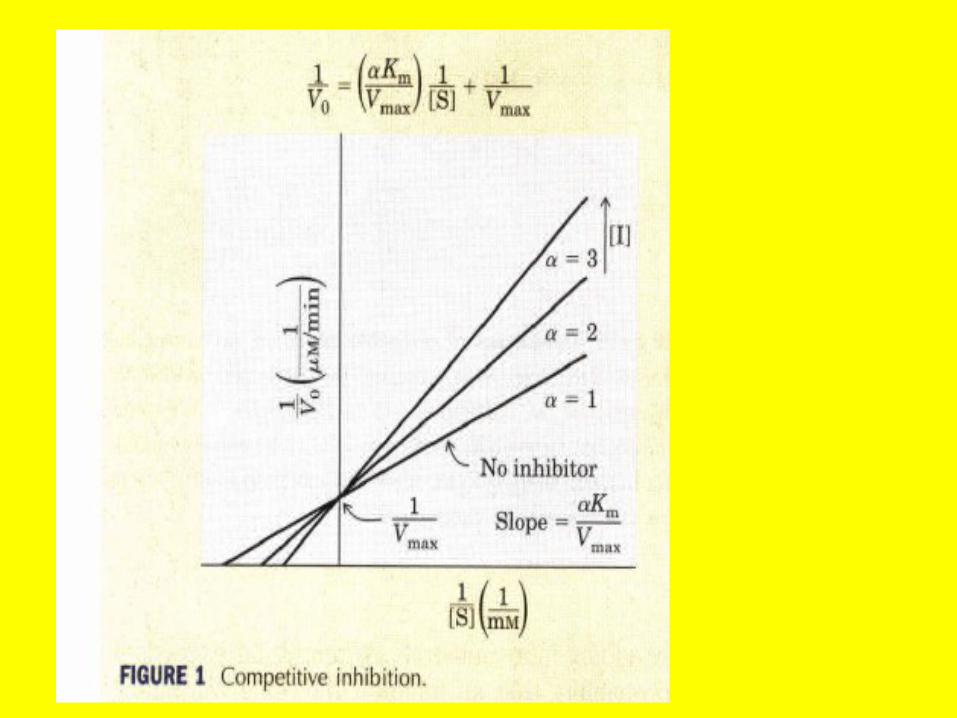

Competitive

Simple Noncompetitive Inhibitors LowerVmax but Do Not Affect Km
• In noncompetitive inhibition, binding of the inhibitor does not affect binding of substrate. Formation of both
• EI and EIS complexes is therefore possible. However,• while the enzyme-inhibitor complex can still bind
substrate,• its efficiency at transforming substrate to product,• reflected by Vmax, is decreased. Noncompetitive• inhibitors bind enzymes at sites distinct from the
substrate-• binding site and generally bear little or no structural• resemblance to the substrate


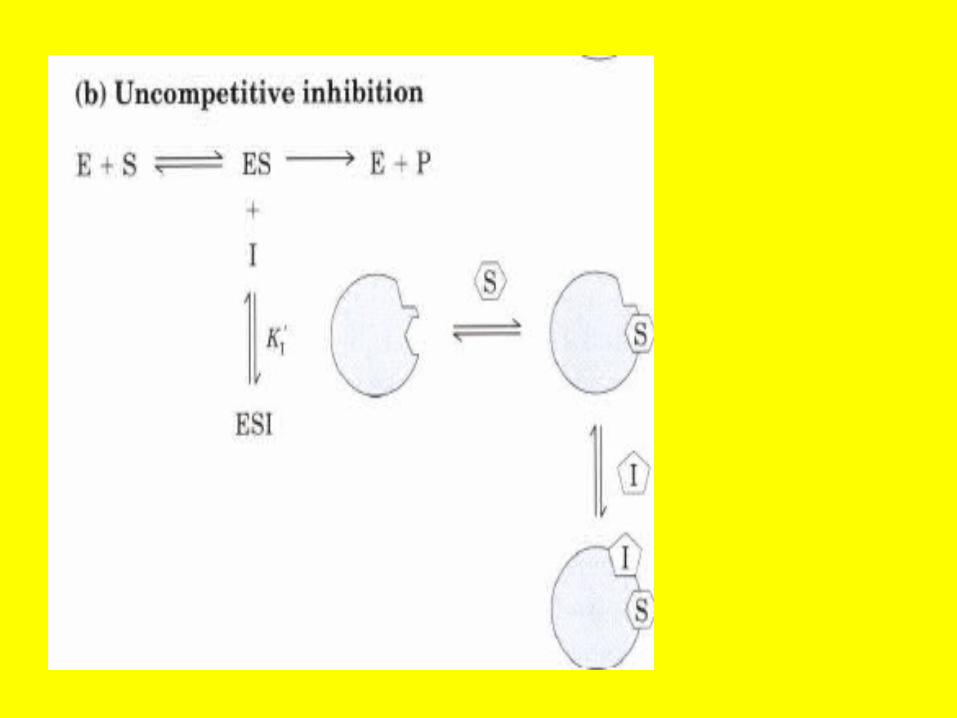
Uncompetitive ES+ I ESI

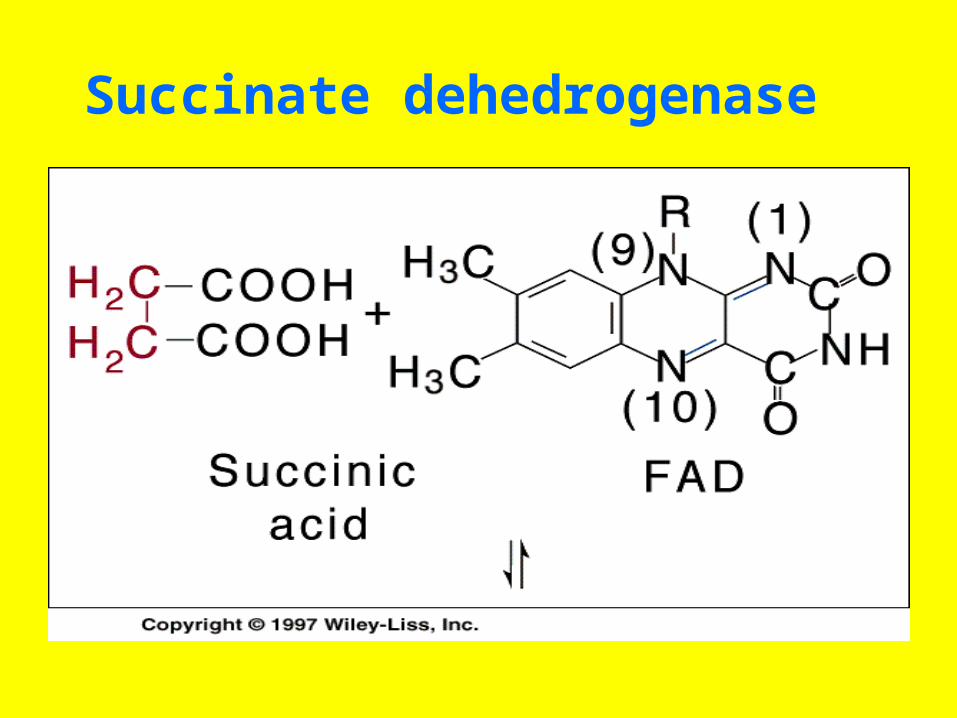
Succinate dehedrogenase
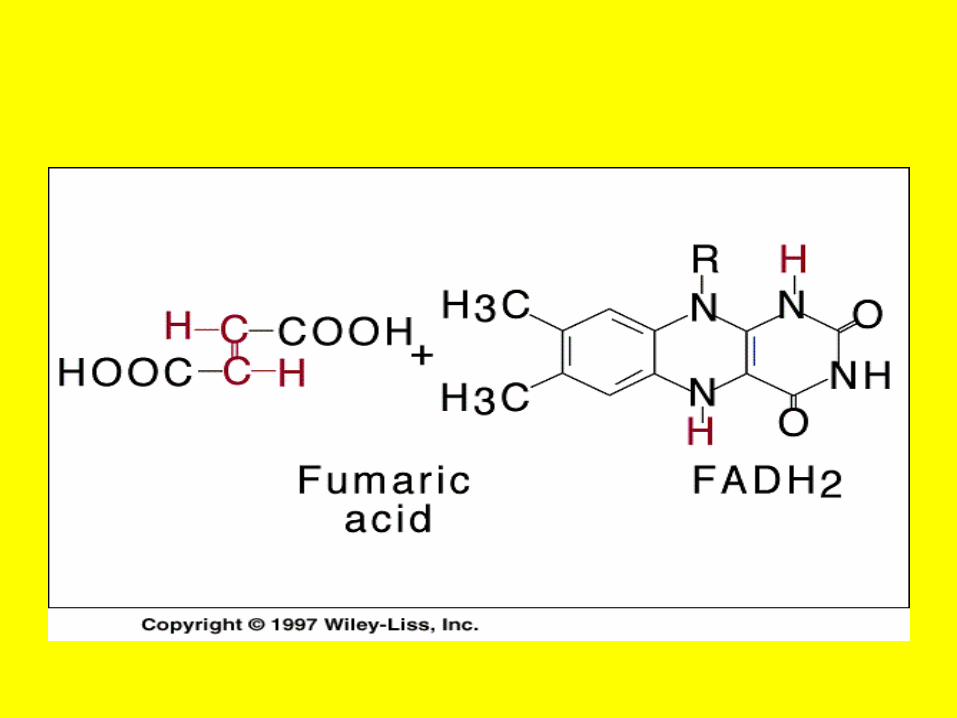


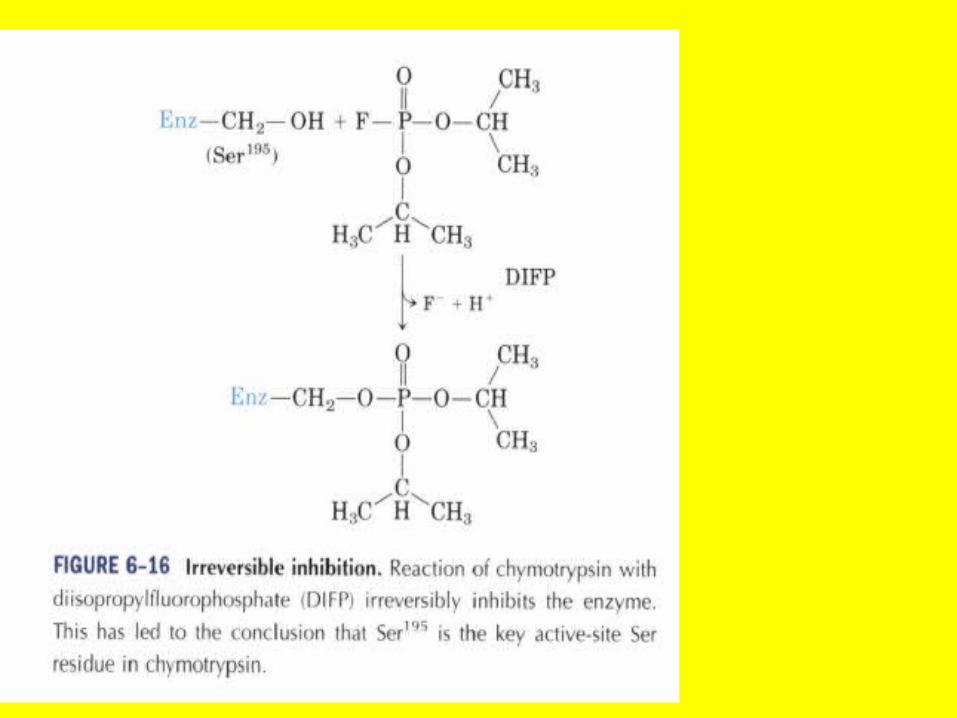
Irreversible
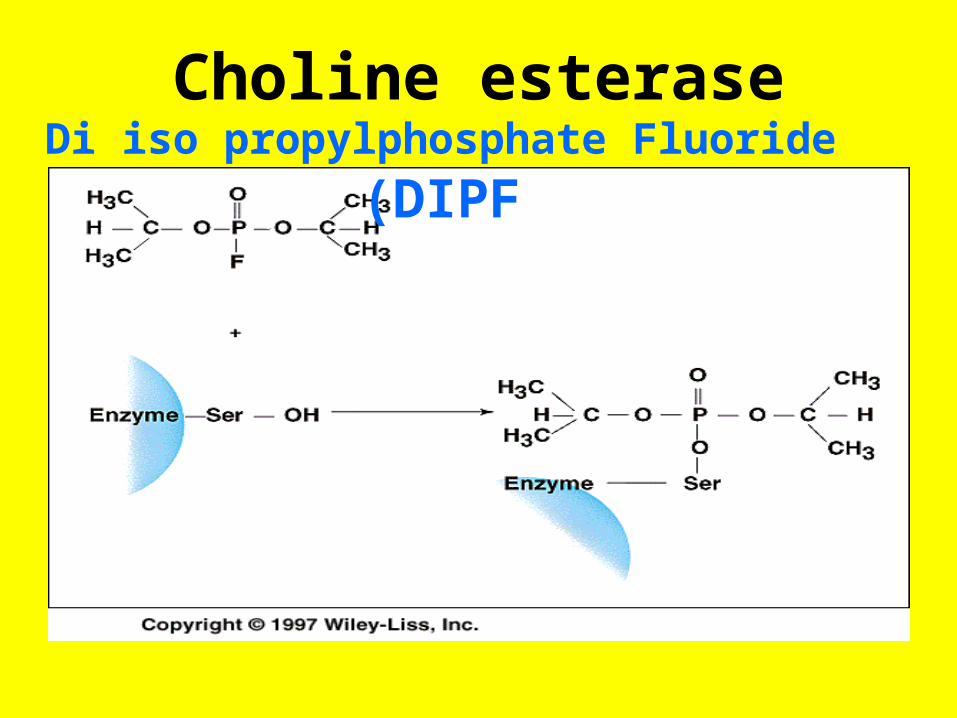
Choline esteraseDi iso propylphosphate Fluoride (DIPF
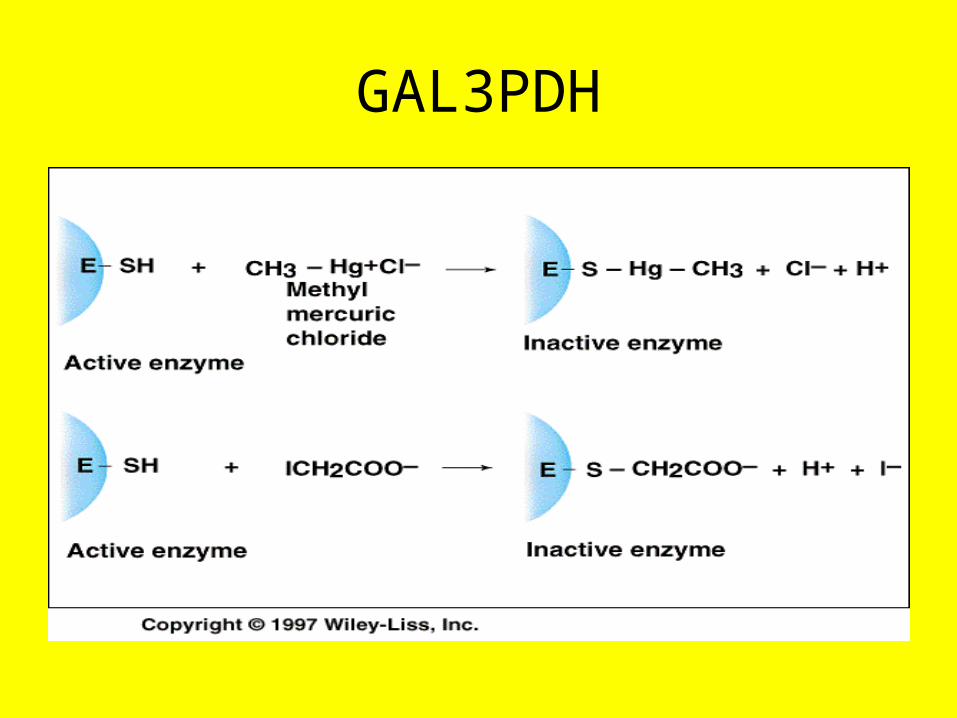
GAL3PDH
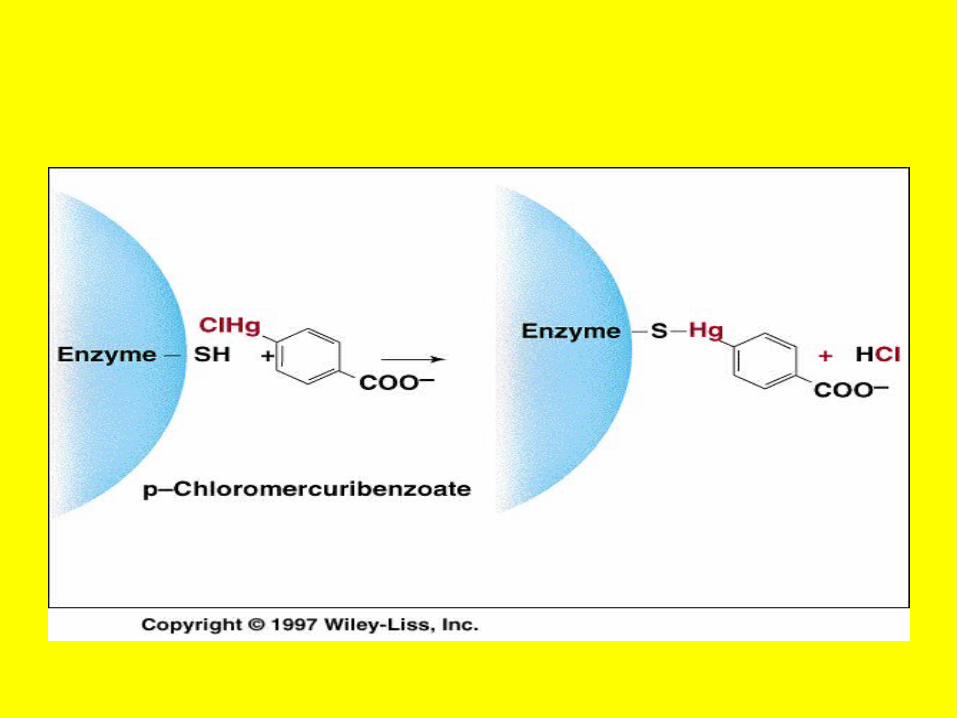
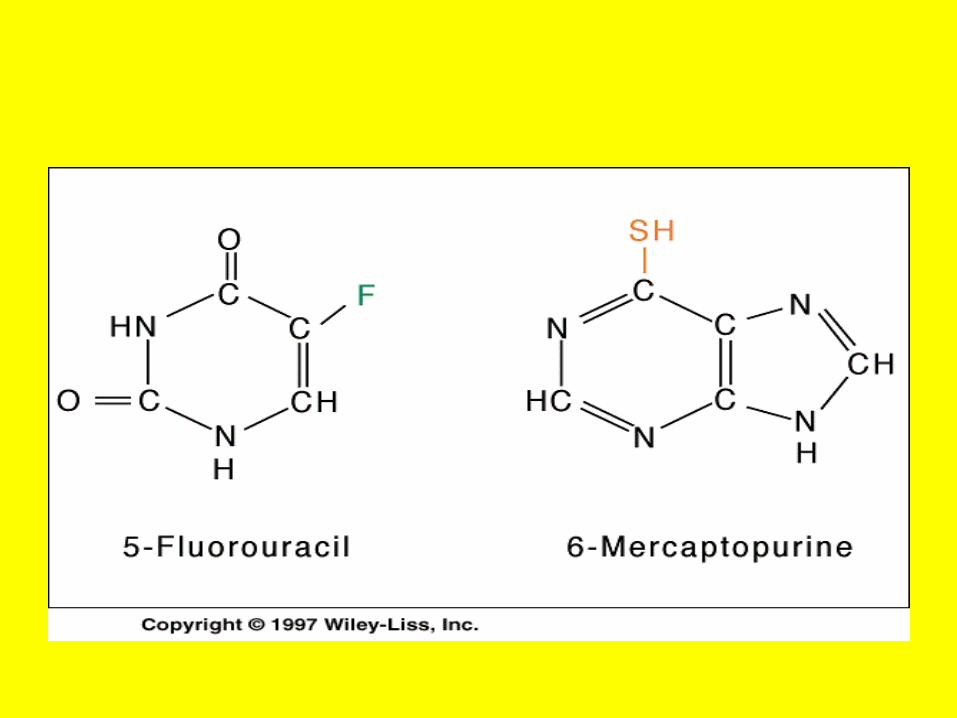
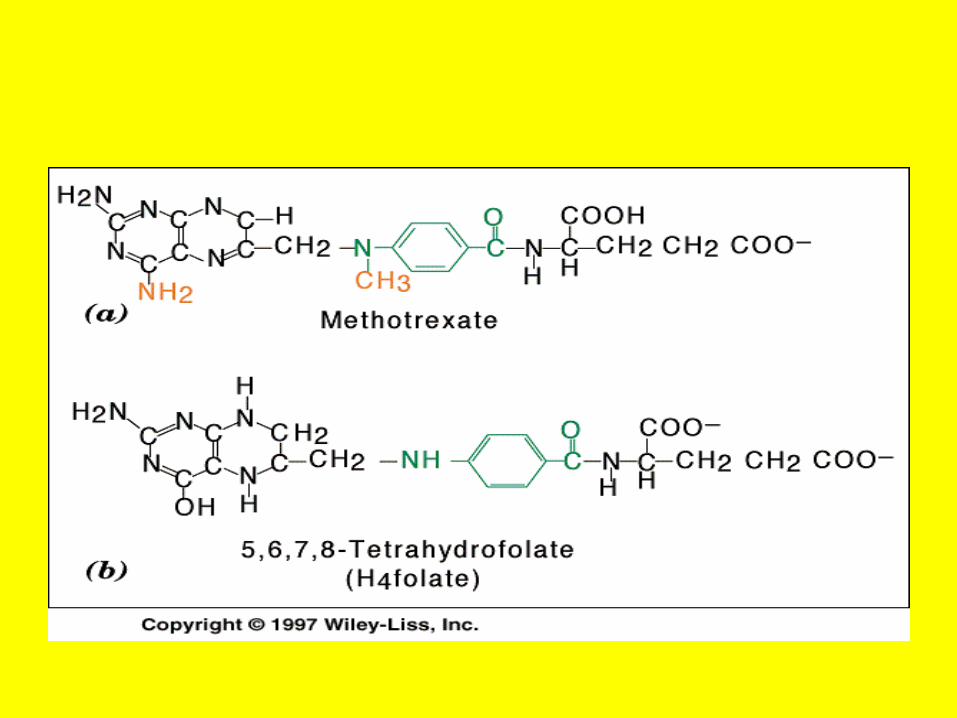
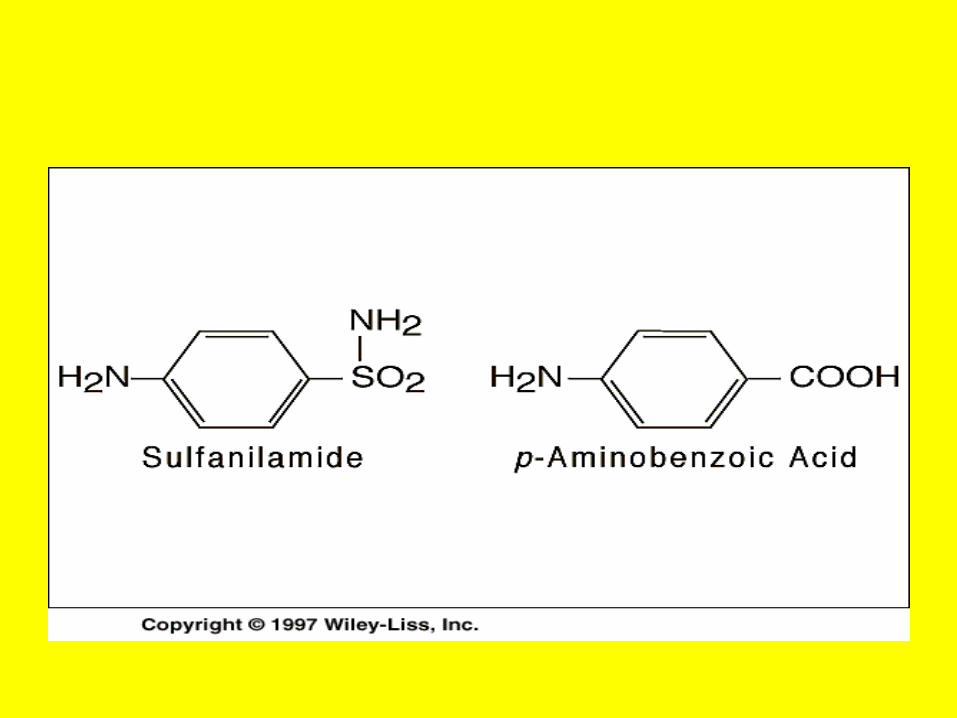

• A Case of Poisoning Emergency room personnel encounter many instances of pesticide poisoning and must be equipped to recognize and treat these cases.
• Many of the common insecticides are organophosphate compounds that irreversibly inhibit the action of acetylcholine esterase(AChE) in the postsynaptic fibers of the cholinergic neurons (p. 948) by
• Forming stable phosphate esters with a specific serine in the active site of the esterase Inhibition of AChE prevents the hydrolysis of acetylcholine in the synapse resulting in constant stimulation of the end organs of these neurons.
• The most prominent effects of pesticide poisoning in humans are paralysis of the respiratory muscles and pulmonary
• edema.
• If given early enough, a drug like pralidoxime can displace the alkyl phosphate from the p€sticide bound to the active site serine and regenerate an active AchE

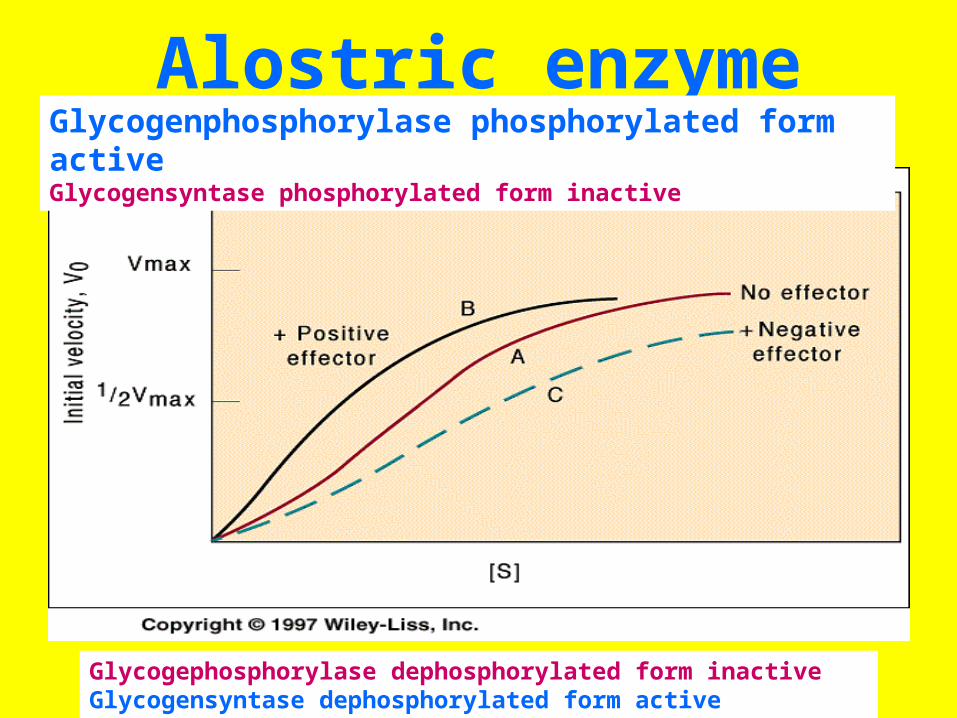
Alostric enzymeGlycogenphosphorylase phosphorylated form activeGlycogensyntase phosphorylated form inactive
Glycogephosphorylase dephosphorylated form inactiveGlycogensyntase dephosphorylated form active