epidémiologie de la résistance aux antibiotiques des
TRANSCRIPT

République Algérienne Démocratique et Populaire
Ministère de l’enseignement supérieur et de la recherche scientifique
Université Abou Bekr Belkaid – Tlemcen
Faculté des Sciences de la Nature et de la Vie
et Sciences de la Terre et de l’Univers
Département de Biologie
Laboratoire Antibiotiques, Antifongiques : Physico-chimie, Synthèse et Activité Biologiques
Mémoire de Magister en Biologie
Option : Biochimie appliquée
THEME
Epidémiologie de la résistance aux antibiotiques des
entérobactéries au niveau du C.H.U de Sidi Bel Abbes
Présenté par
Melle SOUNA Djahida
Devant le jury composé de
Président Mme BOUCHERIT Z. Maître de conférences, Université de Tlemcen
Promoteur Mr DRISSI M. Maître de conférences, Université de Tlemcen
Examinateur Mr ABDELOUAHID D.E. Professeur, Université de Tlemcen
Examinatrice Mme HASSAINE H. Maître de conférences, Université de Tlemcen
Année Universitaire : 2010-2011

Remerciements
Remerciements
Merci à
Dieu
Le tout puissant qui m’a doté de volonté
De patience pour ce travail

Remerciements
Ce travail a été réalisé au niveau du « Laboratoire Antibiotiques,
Antifongiques : Physico-chimie, Synthèse et Activités Biologiques » de la
faculté SNV-STU de l’université Abou Bekr Belkaid.
Je tiens tout d'abord à remercier Mr DRISSI M., maître de conférences
à l'université Abou Bekr Belkaid de Tlemcen pour avoir accepté de diriger ce
travail, pour ses conseils, son aide, ses suggestions sur la rédaction de ce
mémoire ainsi que la confiance qu'il m’a témoigné tout au long de cette
étude.
Je tiens également à remercier :
Mm BOUCHERIT Z., maître de conférences à l'université Abou Bekr
Belkaid de Tlemcen, pour avoir accepté de présider le jury.
Mr ABDELOUAHID D.E., Professeur à l'université Abou Bekr Belkaid
de Tlemcen, pour avoir accepté d'examiner ce travail.
Mm HASSAINE H., maître de conférences à l'université Abou Bekr
Belkaid de Tlemcen, pour avoir accepté de faire partie du jury et d’examiner
ce travail.
J'aimerais remercier touts le personnel du C.H.U de Sidi Bel Abbes
pour m'avoir facilité l’accès, ainsi pour son aide de près ou de loin, par leur
expertise et leur bonne humeur intarissable et plus particulièrement:
Mm BADSI AMIR S., Médecin chef de service de Réanimation au C.H.U
de Sidi Bel Abbes, ainsi que l'ensemble du Personnel Aide Soignant.

Remerciements
Mr BELHANDOUZ., Médecin chef de service de chirurgie au C.H.U de
Sidi Bel Abbes, ainsi que l’ensemble des Infirmiers surtout Mr Maacho.
Mm KADI K., Médecin chef de service de Neurochirurgie au C.H.U de Sidi
Bel Abbes ainsi que le Personnel Aide Soignant.
L’équipe du laboratoire de microbiologie au C.H.U de Sidi Bel Abbes,
j’adresse mes vifs remerciements surtout à Mr Hamza et Melle Karima.
Je tiens à remercier aussi Mr RAHMOUN N., Ingénieurs au
laboratoire « Antibiotiques, Antifongiques » et la Technicienne du
Laboratoire.
Un grand merci à tous les membres de mon équipe: Mm BABA
AHMED Zahira Zakia, Melle AYAD Amel, Melle MESLI Asma et Melle
BERRAZEG Meryem. Leurs conseils judicieux m'auront grandement aidé à
bien réaliser mon travail.
Un merci tout spécial à mes collègues et amis, les étudiants de magister
« Biochimie Appliquée », pour leur aide et leurs encouragements lors de la
préparation de ce mémoire et au cours de l’année théorique: Imane, Sara,
Wafaa, Wassila, Habib, Zinedine et Samir. Les bons moments que nous
avons passés ensemble seront toujours inoubliables.
Merci…..!

Dédicace
Dédicace
J'exprime ma profonde affection :
À mes parents Mostefa et Nacira pour leur soutien constant, leur
amour et leurs mots d'encouragement qui m'ont permis de me rendre ici
aujourd'hui ;
À mes sœurs Amel, Ibtissem et Chaimaa ;
À mon frère Radouane et mon beau frère Mohamed ;
À toute la famille SOUNA et la famille BOUKHARI et surtout mes
tantes Rachida et Sabrina ;
À mon amie Imane avec laquelle j'ai partagé de très bons moments au
laboratoire et l’hôpital ainsi à Hadj Abdelkader pour m’avoir soutenu,
encouragé et conseillé. Je lui dédie cette thèse.
Djahida

Abréviations
Abréviations
spp. : espèce
subsp. : sous-espèce
S.E. : Escherichia coli (S : Sidi Bel Abbes)
S.Kp. : Klebsiella pneumoniae
S.Ec. : Enterobacter cloacae
S.Pm. : Proteus mirabilis
S.Pv. : Proteus vulgaris
S.Pp. : Proteus penneri
S.Ps. : Providencia stuartii
S.Mm.1 : Morganella morganii
S.Cf.1 : Citrobacter freundii
S.Ss.1 : Shigella sonnei
S.Sel.spp. : Salmonella spp.
TC. : transconjugant
ARN : acide ribonucléique
°C : dégrée celcus
DNAse : Désoxyribonucléase
ONPG : Orthonitrophényl-β-D-galactopyrannoside
ADN : Acide désoxyribonucléique
D-ala : D-alanine
C1G : Céphalosporine de 1ere
génération
C2G : Céphalosporine de 2eme
génération
C3G : Céphalosporine de 3eme
génération
C4G : Céphalosporine de 4eme
génération
CTX-M : Céfotaximase-Munich
TEM : TEMoneira- nom du patient
SHV : SulfHydryl Variable
PER : Pseudomonas Extended Resistance
VEB : Vietnam Extended-spectrum -lactamase
GES : Guyana Extended-Spectrum -lactamase
TLA : Tlahuicas - tribu indienne
BES : Brazilian extended-spectrum -lactamase

Abréviations
SFO : Serratia fonticola
FEC : Fecal Escherichia coli
OXA : Oxacillinase
MIR-1 : Miriam hospital
BIL-1 : Belgium extended-spectrum -lactamase
CMY-2 : Cephamycinase-2
ACT-1 : AmpC Type
DHA : Dharhan hospital
ACC-1 : Ambler Class C
SME1 : Serratia marcescens
PASE: pénicillinase
CASE: céphalosporinase hyperproduite
ARNr : ARN ribosomal
ARNt : ARN de transfert
ARNm : ARN messager
LPS : lipo-polysaccharide
CHU : Centre hospitalo-universitaire
ATCC : American Type Culture Collection
EDTA : Ethylenediamintetraacetic acid
TE : Tris-EDTA
TBE : Tris-Borate-EDTA
BET : Bromure d’éthidium
H2S : sulfure d'hydrogène
BHIB: brain-heart infusion broth
UFC : unité formant cellule
H : heure
m : mettre
nm : nano-mettre
mn : minutes
mm : milli-mettre
M : molaire
g : gramme
CA-SFM : Comité d’antibiogramme- société française de microbiologie
SDS : Sodium dodécyl sulfate
LB : Luria Broth
rpm : rotation par minute
D.O : Densité optique
Kb: Kilo-bases

Sommaire
Sommaire
Introduction 1
Synthèse Bibliographique 3
1. Généralités sur les entérobactéries… …....................................................... 3
1.1. Escherichia coli……………………………………………………...………………. 5
1.2. Shigella ……………………………………………………………………… 5
1.3. Salmonella…………………………………………………………………… 6
1.4. Les entérobactéries opportunistes……………………………………………. 7
1.4.1. Groupe des K.E.S………………………………………………………….. 7
1.4.1.1. Klebsiella……..………………………………………………….............. 7
1.4.1.2. Enterobacter………………………………………… ................... … 8
1.4.1.3. Serratia………………………………………………………………......... 8
I.4.2. Groupe des Proteae……………………………………………........................… 9
1.4.2.1. Proteus.................................................................................................. 9
1.4.2.2. Providencia............................................................................................ 10
1.4.2.3. Morganella............................................................................................. 10
1.5. Yersinia……………………………………………………………………… 10
1.6. Les entérobactéries rares ou récemment décrites………………………..….. 11
2. La résistance aux antibiotiques……….....………………………………… 11
2.1. Support génétique de la résistance…………………………………….…….. 13
2.2. Mécanismes biochimiques de la résistance aux antibiotiques……..……….. 14
3. La résistance des entérobactéries aux -lactamines……………………… 16
3.1. Définition des -lactamines…………………………….………………….. 16
3.2. Mécanisme d’action………………………………….…………………….. 18
3.3. Mécanismes de résistance……………………..…………….……………... 18
3.3.1. La résistance non enzymatique aux -lactamines……………………….. 18
3.3.1.1. Imperméabilité....................................................................................... 18

Sommaire
3.3.1.2. Modification de la cible PLP................................................................. 19
3.3.1.3. Pompe à efflux ...................................................................................... 19
3.3.2. La résistance enzymatique aux -lactamines (-lactamases)……..……… 19
3.3.2.1. Définition………………………………………………………..……… 19
3.3.2.2. Mécanisme d’action…………………………………………..………… 20
3.3.2.3. Classification des -Lactamases………………………………………... 21
3.4. Phénotypes de résistance aux -lactamines…………………...……….…… 22
3.4.1. Résistance naturelle ou phénotypes « sauvages »………….….…………. 22
3.4.2. Résistance acquise ou phénotypes « résistants »……..………..………… 24
3.4.3. Résistance aux carbapénèmes ……………………………………………. 27
4. La résistance aux aminosides………..........………………………………. 28
5. La résistance aux quinolones…………………….……………………….. 29
6. La résistance aux autres antibiotiques…………...……………………….. 31
6.1. Triméthoprime-Sulfaméthoxazole………………………….………………. 31
6.2. Polymyxines ………………………………………….……………………. 32
Matériel et Méthodes
34
1. Matériel……………………………………………………….……………….. 34
1.1. Matériel biologique ………………………………………….…………….. 34
1.2. Milieux de culture ……………………………………………….………… 34
1.3. Tests biochimiques ………………………………………….……………... 34
1.4. Solutions et Tampons ……………………..……………………………….. 35
1.5. Antibiotiques ……………………………………………………………..... 35
2. Méthodes ……………………………………………………………………… 36
2.1. Prélèvements………………………………………………………………... 36
2.2. Isolement et purification …………………………………………………… 36
2.3. Identification ……………………………………....………………………. 36
2.4. Antibiogramme………………………………………………...…………… 38
2.5. Détermination de la CMI en milieu solide ………………………...………. 40
2.6. Détermination de la CMI pour les Proteus………………………………..……. 41
2.7. Test de synergie ……………………………………...…………………….. 43

Sommaire
2.8. Test à la cloxacilline ………………………………………………...……... 44
2.9. Test de Hodge ……………………………………………………...……… 45
2.10. Test de l’EDTA …………………………………………………………..... 46
2.11. Transfert des plasmides par conjugaison …………………………………... 47
2.12. Extraction de l’ADN plasmidique …………………………...…………….. 48
2.13. Electrophorèse sur gel d’agarose…………………………………...………. 49
Résultats et Interprétation 51
1. Prélèvements ………………………………………..……..…………..…. 51
2. Identification de souches ………………………………………..……..…. 52
2.1. Répartition des entérobactéries en fonction des services ……...………….... 54
2.2. Répartition des entérobactéries en fonction des sites de prélèvement…….… 56
3. Résistance des entérobactéries aux antibiotiques ………...…………...….. 58
3.1. Résistance d’Escherichia coli ……………………………………………..……… 61
3.2. Résistance de Klebsiella pneumoniae………………………………………….… 62
3.3. Résistance de Proteus spp. …………………………………………….…… 62
3.4. Résistance d’Enterobacter cloacae…………………………………………. 63
4. Phénotypes de résistance …………...………………….…………………. 65
5. Résistances associées des entérobactéries ………………….……….......... 70
6. Conjugaison bactérienne ………………………………..………………... 71
7. Extraction et visualisation des plasmides …………………………..…….. 74
Discussion 75
Conclusion 81
Références bibliographiques 83
Annexes 105

Liste des figures
Liste des figures
Figure 1. Cibles des principaux antibiotiques……………………………………. 12
Figure 2. Cycle -lactame........................................................................................ 16
Figure 3. Classification des -lactamines………………………………………… 17
Figure 4. Mécanisme d’hydrolyse d’une -lactamine par une -lactamase……… 21
Figure 5. Disposition des antibiotiques sur les boîtes d’antibiogrammes
selon le schéma de Vedel…………………………………………………….
39
Figure 6. Envahissement (swarming) de Proteus spp. sur gélose Mueller Hinton.. 43
Figure 7. Lecture du test de Hodge……………………………………………….. 45
Figure 8. Répartition de l’ensemble des espèces d’entérobactéries isolées……… 52
Figure 9. Identification des principales espèces par galerie API 20E et ID 32E…. 53
Figure 10. Répartition des entérobactéries au niveau du service de réanimation.... 54
Figure 11. Répartition des entérobactéries au niveau du service de chirurgie...….. 55
Figure 12. Répartition des entérobactéries au niveau des autres services………… 55
Figure 13. Répartition des entérobactéries au niveau des sondes urinaires……… 56
Figure 14. Répartition des entérobactéries au niveau des plaies chirurgicales …. 56
Figure 15. Répartition des entérobactéries au niveau des prélèvements trachéaux. 57
Figure 16. Répartition des entérobactéries au niveau des prélèvements de
l’environnement…………………………………………………………………….
57
Figure 17. Pourcentage de résistance des entérobactéries aux -lactamines…….. 58
Figure 18. Pourcentage de résistance des entérobactéries aux autres classes
d’antibiotique………………………………………………………………………
59

Liste des figures
Figure 19. Pourcentage de résistance des entérobactéries aux -lactamines au
niveau de la réanimation, de la chirurgie et des autres services………...…………
60
Figure 20. Pourcentage de résistance des entérobactéries aux autres antibiotiques
au niveau de la réanimation, de la chirurgie et des autres services……...…………
60
Figure 21. Pourcentage de résistance d’Escherichia coli aux antibiotiques……... 61
Figure 22. Pourcentage de résistance de Klebsiella pneumoniae (A), Proteus
spp. (B) et Enterobacter cloacae (C) aux -lactamines…………………………...
64
Figure 23. Pourcentage de résistance de Klebsiella pneumoniae (A), Proteus
spp. (B) et Enterobacter cloacae (C) aux autres antibiotiques…………………...
64
Figure 24. Phénotypes de résistance des entérobactéries aux -lactamines……. 65
Figure 25. La répartition des phénotypes de résistance en fonction des services... 66
Figure 26. Phénotype BLSE……………………………………………………… 67
Figure 27. Phénotype BLSE+CASE……………………………………………... 67
Figure 28. Phénotype CASE……………………………………………………… 68
Figure 29. Phénotype PASE………………………………………………………. 68
Figure 30. Test de Hodge…………………………………………………………. 69
Figure 31. Test de l’EDTA pour la VIM (A) et la souche S.Ec.14 (B)…………... 69
Figure 32. Transfert de phénotype BLSE ……………………………….............. 71
Figure 33. Transfert de phénotype BLSE+CASE ……………………………….. 72
Figure 34. Transfert de phénotype BLSE chez les souches présentant le
phénotype BLSE+CASE…………………………………………………...............
73
Figure 35. Profils plasmidiques des 4 espèces d’entérobactéries et de leurs
transconjugants……………………………………………………………………..
74

Liste des tableaux
Liste des tableaux
Tableau 1. Entérobactéries rares ou récemment décrites………………………... 11
Tableau 2. Les mécanismes de résistance bactérienne aux antibiotiques………. 15
Tableau 3. Répartition des prélèvements selon les services et les sites de
prélèvement………………………………………………………………………..
51
Tableau 4. Intervalles de CMI (g/ml) observés en fonction des phénotypes de
résistance aux -lactamines……………………………………………………….
66
Tableau 5. Résistances associées des entérobactéries en fonction des phénotypes
de résistance aux -lactamines……………………………………………………
70
Tableau 6. Intervalles de CMI (g/ml) observés en fonction des phénotypes de
résistance…………………………………………………………………………..
70
Tableau 7. Intervalles de CMI (g/ml) observés en fonction des phénotypes de
résistance aux -lactamines des transconjugants………………………………….
73

Introduction

Introduction
1
La découverte de la pénicilline suivie par celle des autres antibactériens au cours du
siècle dernier a permis d’accroître l’espérance de vie de plus de dix ans. Cependant, ce
progrès essentiel de la médecine moderne comporte des inconvénients, c’est pourquoi
l’on parle de « paradoxe des antibiotiques ». Ce paradoxe est la conséquence du
développement des résistances liées à une large utilisation (et parfois abusive) des
antibiotiques, conjuguée à la pénurie de nouvelles molécules, limitant l’arsenal
thérapeutique contre les infections à bactéries résistantes. La multirésistance aux
antibiotiques concerne principalement les bactéries à Gram négatif (Frasca et al.,
2008). L’apparition de bactéries résistantes aux antibiotiques et leur propagation à
l’hôpital comme en ville sont devenues une préoccupation sanitaire majeure depuis les
années 1980 (Stahl, 2005). Malheureusement, nous sommes confrontés à la monté
croissante de cette résistance, atteignant à l’heure actuelle des niveaux alarmants (Van
Bambeke et al., 2002).
Les entérobactéries forment une vaste famille de bactéries à Gram-négatif, qui sont
à l’origine de maladies de gravité très variable, en raison de mécanismes
pathogéniques distincts (Mirabaud, 2003). Les espèces de cette famille ont été depuis
une vingtaine d’années largement exposées à une utilisation extensive des
antibiotiques. Revers de la médaille, elles n’ont pas été épargnées par la résistance
croissante aux molécules les plus fréquemment utilisées (Péan et al., 2001).
L’apparition de la résistance chez les Enterobacteriaceae est un problème important
qui nécessite une attention immédiate. La résistance liée à la production de -
lactamase à spectre étendu (BLSE) est un problème particulier dans le traitement des
infections à entérobactéries, mais d’autres mécanismes ont également émergé,
conduisant à la multirésistance et menaçant de créer des espèces pan-résistants
(Paterson, 2006).

Introduction
2
En milieu hospitalier, l'utilisation des antibiotiques est souvent associée à une
augmentation de la résistance. Bien que de bonnes politiques d'utilisation des
antibiotiques permettent souvent de contrôler les épidémies causées par les bactéries
multirésistantes, le problème de la résistance demeure imposant et coûteux pour la
société. Il est donc impératif d'étudier le phénomène afin de mieux comprendre
l'origine, le fonctionnement et les mécanismes de la résistance aux antibiotiques
(Tremblay, 2007).
C'est dans cet ordre d'idée que nous nous sommes proposés de réaliser une
étude à l’hôpital de Sidi Bel Abbes selon les étapes suivantes:
Isoler et identifier les entérobactéries;
Étudier la résistance aux antibiotiques de ces souches;
Détecter les phénotypes de résistances associées;
Extraire et visualiser les plasmides.

Synthèse
Bibliographique

Synthèse Bibliographique
3
1. Généralités sur les entérobactéries:
Les entérobactéries constituent un grand groupe de bactéries ayant une forte
similitude. La création de ce groupe a été proposée par Rahn en 1937 qu’il dénomma
Enterobacteriaceae. Si le nom de famille est toujours maintenu, en revanche le
classement des bactéries dans la famille a beaucoup évolué (Joly et Reynaud, 2007).
Actuellement, les entérobactéries sont classées sur la base du séquençage des ARN
5S et 16S dans :
Domaine : Eubacteria ;
Phylum XII : Proteobacteria ;
Classe : Cammaproteobacteria ;
Ordre des Enterobacteriales ;
Famille des Enterobacteriaceae (Meyer et al., 2004).
44 genres sont regroupés en cinq tribus, d’après leurs propriétés fermentatives:
Escherichiae, Klebsielleae, Proteae, Yersiniae et Erwiniae (Larpent, 2000). Les
genres les plus communément isolés en bactériologie clinique sont: Citrobacter,
Enterobacter, Escherichia, Hafnia, Klebsiella, Morganella, Proteus, Providencia,
Salmonella, Serratia, Shigella et Yersinia (Morice, 2003).
Le nom « entérobactérie » fait référence à la localisation de cette famille de
microorganismes dans le tube digestif et principalement le côlon de l’homme et des
animaux (Scheftel, 2008). La famille des entérobactéries se définit par un ensemble
de caractères généraux communs. Ce sont des bacilles à Gram négatif, non sporulés,
le plus souvent mobiles grâce a une ciliature péritriche mais immobiles dans le cas
des bactéries des genres Klebsiella, Shigella et Yersinia pestis. Elles fermentent le D-
glucose avec ou sans production de gaz et réduisent les nitrates en nitrites. Elles n’ont
pas d’oxydases et possèdent une catalase (Joly et Reynaud, 2007). Elles sont aérobies
-anaérobies facultatives et elles cultivent sur les milieux ordinaires. La température
optimale de croissance est 37°C mais la culture est possible entre 20°C et 40°C. Leur
temps de division varie de 20 à 40 minutes. Sur gélose, les colonies sont lisses et

Synthèse Bibliographique
4
régulières et atteignent 2 millimètres de large sauf celles des Yersinia, qui sont plus
petites. Les Proteus ont tendance à envahir la gélose et à y former un tapis uniforme.
En milieu liquide, les entérobactéries occasionnent un trouble uniforme de bouillon
(Decoster, 2005).
Les entérobactéries sont une famille très hétérogène pour ce qui est de leur
pathogénie et de leur écologie. 130 espèces sont en effet soit parasites (Shigella,
Yersinia pestis), soit commensales (Escherichia coli, Proteus mirabilis, Klebsiella
spp.), soit encore saprophytes (Serratia spp., Enterobacter spp.) (Decoster, 2005;
Morice, 2003). Elles semblent plus spécifiquement adaptées à l'homme ou l'animal;
certaines sont responsables d'infections humaines parfois sévères (fièvre typhoïde,
dysenterie bacillaire, peste). D'autres groupes pourtant prolifèrent en abondance dans
l'environnement (sol-eaux). Elles participent aux grands cycles de dégradation des
matières organiques ou sont étroitement associées aux plantes chez lesquelles elles
peuvent déterminer des altérations nuisibles dans le domaine agro-alimentaire
(nécroses, dégénérescence ou ramollissement tissulaire, pourriture molle) (Philippon,
2001).
Les entérobactéries fréquemment rencontrées en bactériologie clinique, alimentaire,
des eaux… sont succinctement présentées en considérant leur habitat naturel ou leur
habitat lié aux activités anthropiques, leurs caractères particuliers, leurs pouvoirs
pathogènes ou leur intérêt (Delarras, 2007). Seuls les genres et les espèces qui ont un
intérêt médical reconnu seront envisagés par la suite. Une centaine d’espèces
d’Enterobacteriaceae sont individualisées, mais 23 d’entre elles représentent 99% des
souches isolées en clinique (Avril et al., 2000).

Synthèse Bibliographique
5
1.1. Escherichia coli:
Isolée pour la première fois par Escherich en 1885, Escherichia coli est l'espèce
bactérienne qui a été la plus étudiée par les fondamentalistes pour des travaux de
physiologie et de génétique (Avril et al., 2000). Elle représente l’espèce type de genre
Escherichia. Appelée communément « colibacille » cette espèce possède des
caractères biochimiques particuliers permettant de la différencier des espèces voisines.
La production d’indole à partir de tryptophane, l’absence d’utilisation du citrate
comme source de carbone et l’absence de production d’acétoïne (réaction de Voges-
Proskauer négative) (Joly et Reynaud, 2007).
Escherichia coli se retrouve en abondance dans la flore commensale humaine, en
particulier dans le tube digestif de l’homme qu’elle colonise dès les premières heures
de la naissance. Elle constitue l’espèce dominante de la flore aérobie anaéro-tolérante
(Ahoyo et al., 2007; Bonacorsi et al., 2001).
Les colibacilles, hôtes normaux de l'intestin, ne provoquent normalement pas de
maladie. Cependant ils possèdent un potentiel pathogène qu'ils expriment dans
certaines circonstances (pathogènes opportunistes) (CHU-PS, 2003).C’est une des
espèces bactériennes les plus souvent rencontrées en pathologie humaine. La plupart
sont commensales, mais certaines souches possèdent des facteurs de virulence qui leur
permettent de déclencher des diarrhées. Escherichia coli est une cause majeure de
diarrhée aigue dans le monde (Berche, 2003). A côté des infections intestinales, elle
est responsable d’infections extra-intestinales diverses: urinaires, abdominale,
méningées et les bactériémies (Fauchère et Avril, 2002).
1.2. Shigella:
Nommé Shigella en l'honneur du bactériologiste japonais Kiyoshi SHIGA qui a
découvert le bacille de la dysenterie en 1897 (Dufour, 2005). Les Shigella sont des
entérobactéries immobiles extrêmement proches d’Escherichia coli mais qui ne
fermentent pas le lactose. Elles n’ont pas d’uréase et ne produisent pas de gaz. (CHU-
PS, 2003). Ce sont des parasites intestinaux rencontrés seulement chez l'homme, qui

Synthèse Bibliographique
6
les élimine par ses selles et les disperse dans le milieu extérieur (sol, eau) où elles ne
survivent pas longtemps (Peiffer, 2000).
Il existe 4 espèces, Shigella dysenteriae responsable de la dysenterie bacillaire,
Shigella flexneri, Shigella boydi et Shigella sonnei. Ces bactéries envahissent la
muqueuse colique, déclenchant des entérites inflammatoires fébriles dans le monde
entier. La gravité de cette infection est liée à la déshydratation qu’elle provoque,
nécessitant une antibiothérapie et une réhydratation d’urgence (Berche, 2003).
1.3. Salmonella:
Kauffmann, le pionnier de l’analyse du genre Salmonella, avait individualisé sur la
base de tests phénotypiques et de tests sérotypiques plusieurs sous-genres et de très
nombreuses espèces de Salmonella (par exemple, Salmonella typhi, espèce
responsable de la fièvre typhoïde) (Weill, 2009). Depuis 2005, une nouvelle
nomenclature est en vigueur sur le plan international (Tindall et al., 2005). Elle fait
suite aux études moléculaires (hybridations ADN-ADN) qui ont révélé la présence de
seulement deux espèces dans le genre Salmonella (S. enterica, espèce majoritaire et S.
bongori, espèce rare). S. enterica est elle-même subdivisée en six sous-espèces.
L’espèce bongori et les différentes sous espèces d’enterica sont ensuite subdivisées
sur la base du sérotypage en de très nombreux sérotypes. À ce jour, plus de 2500
sérotypes (ou sérovars) sont décrits (Popoff, 2001).
Le réservoir des bactéries du genre Salmonella est principalement le tube digestif
des vertébrés. De très nombreuses espèces animales hébergent ces agents pathogènes
(volailles, bovins, porcs, poissons, reptiles,….). La sous-espèce enterica est plutôt
adaptée aux animaux à sang chaud et à l’homme (Weill, 2009). Les salmonelles sont
responsables de fièvres typhoïdes et de salmonelloses non typhiques qui représentent
une cause majeure de diarrhées dans le monde avec un taux important de mortalité
infantile (Dagnra et al., 2007 ).

Synthèse Bibliographique
7
1.4. Les entérobactéries opportunistes:
1.4.1. Groupe des K.E.S:
Dans le groupe Klebsiella - Enterobacter - Serratia, dit K.E.S., sont rassemblées
des Enterobacteriaceae qui ont en commun les caractères suivants:
La réaction de Voges-Proskauer (VP) est généralement positive;
Bactéries pathogènes opportunistes;
Multirésistance aux antibiotiques (Avril et al., 2000).
1.4.1.1. Klebsiella:
Les Klebsiella sont des bactéries immobiles, en diplobacilles, généralement
capsulées (Delarras, 2007) et fermentent de nombreux sucres avec production de gaz,
mais elles ne sont pas protéolytiques (Fauchère et Avril, 2002). Sur milieu gélosé, les
colonies sont caractéristiques: elles sont volumineuses, bombées, brillantes et très
visqueuses à cause de la capsule. (Fauchère et Avril, 2002).
Le genre Klebsiella comporte actuellement cinq espèces K. pneumoniae comportant
3 sous espèces: K. pneumoniae subsp. pneumoniae, K. pneumoniae subsp. ozanae et
K.pneumoniae subsp. Rhinoscleromatis ; Klebsiella oxytoca ; Klebsiella
ornithinolytica ; Klebsiella terrigena et Klebsiella planticola (Farah et
Boutefnouchet, 2007).
L’espèce type est Klebsiella pneumoniae, germe très répandu dans la nature (sol et
eau), saprophyte des voies respiratoires supérieures et il est l’agent des surinfections
respiratoires (Lamnaouer, 2002). Klebsiella pneumoniae est un pathogène à fort
potentiel épidémique fréquemment impliqué dans des infections sévères. De
nombreuses épidémies nosocomiales causées par cette bactérie ont été décrites,
notamment chez des patients hospitalisés dans des unités de soins intensifs adultes ou
pédiatriques (Carrër et Nordmann, 2009; Boukadida et al., 2002).

Synthèse Bibliographique
8
1.4.1.2. Enterobacter:
Les Enterobacter sont des bacilles à Gram négatif généralement mobiles,
fermentent ou non le lactose et ils ont une -galactosidase (Fauchère et Avril, 2002).
Différentes espèces constituent ce genre. Certains n’ont jamais été associés à des
infections humaines. Les espèces les plus souvent isolés incluent Enterobacter cloacae
et Enterobacter aerogenes, suivie par Enterobacter sakazakii. Les espèces du genre
Enterobacter, en particulier Enterobacter cloacae et Enterobacter aerogenes, sont des
pathogènes responsables d'infections nosocomiales diverses, y compris la bactériémie,
les infections des voies respiratoires et urinaires, l'endocardite, les infections intra-
abdominales et ophtalmiques, l'arthrite septique et les ostéomyélites (Fraser et al.,
2010). Enterobacter sakazakii est l’agent d’infections rares mais sévères touchant
particulièrement les très jeunes enfants, les personnes âgées et les sujets
immunodéprimés. Cette espèce se différencie des autres Enterobacter par son pigment
jaune (Leclercq, 2006).
1.4.1.3. Serratia:
Toutes les Serratia possèdent une gélatinase et une DNAse sauf (S. fonticola)
(Denis et Ploy, 2007). D’une manière générale, les espèces de ce genre sont isolées
des plantes (légumes, champignons, mousses), du tube digestif des rongeurs (40% des
petits mammifères sauvages sont porteurs de Serratia spp.), des insectes, de l’eau et du
sol (Euzéby, 2003). Le genre Serratia comprend maintenant dix espèces: Serratia
marcescens, Serratia liquefaciens, Serratia proteomaculans, Serratia grimesii,
Serratia polmuthica, Serratia rubidaea, Serratia odorifera, Serratia ficaria, Serratia
fonticola, et Serratia entomophila (Sekhsokh et al., 2007). La principale espèce
pathogène du genre est Serratia marcescens qui provoque habituellement des
infections nosocomiales. Toutefois, des souches de S. plymuthica, S. liquefaciens, S.
rubidaea et S. odorifera ont causé des maladies à travers des infections (Basilio et
Anía, 2009).

Synthèse Bibliographique
9
1.4.2. Groupe des Proteae:
La tribu des Proteae ou groupe Proteus- Morganella- Providencia, appartient à la
famille des Enterobacteriaceae.
Ce groupe se caractérise par la désamination d’acides aminés en acides cétoniques
qui, additionnés d’ions ferriques, donnent des réactions colorées grâce à des enzymes
comme:
Tryptophane désaminase (TDA);
Phénylalanine désaminase (ph.al. DA) qui catalysent la désamination du
tryptophane en acide indolpyruvique (coloration rouge brun avec Fe) et de la L
phénylalanine en acide phenylpyruvique (coloration vert foncé avec Fe).
Les Proteus et Morganella hydrolysent rapidement l’urée contrairement aux
Providencia qui ne possèdent pas d’uréase (Amhis, 2004).
1.4.2.1. Proteus:
Les Proteus spp. sont des bacilles à Gram négatif, généralement très mobiles,
polymorphes, mesurant de 0,4 à 0,8 μm de diamètre sur 1,0 μm à 80 μm de longueur.
Les espèces du genre Proteus sont largement répandues dans la nature et elles sont
isolées du sol, de l’eau, de l’intestin de l’homme et de nombreuses espèces animales.
Actuellement, le genre Proteus rassemble cinq espèces (Lamnaouer, 2002) dont 3
espèces importantes pour l’homme, Proteus mirabilis, Proteus vulgaris et Proteus
penneri (Goubau et Pellegrims, 2000). Certaines espèces comme Proteus mirabilis
et Proteus vulgaris ont une propriété très connue de s’étaler très rapidement sur boîte
de gélose: c’est le phénomène de swarming (Pelmont, 2005).
Proteus mirabilis est l’espèce la plus fréquemment isolée de prélèvements
cliniques. (Lamnaouer, 2002). Après Escherichia coli, elle est la bactérie la plus
souvent isolée des urines et elle est à l’origine aussi d’infections nosocomiales
(Mahrouki et al., 2009).

Synthèse Bibliographique
10
1.4.2.2. Providencia:
Les espèces du genre Providencia sont habituellement considérées comme
commensaux dans le tube digestif, mais certaines espèces (Providencia stuartii et
Providencia alcalifaciens) ont été associées à des infections nosocomiales et sont
considérées comme des pathogènes opportunistes (Tribe et Rood, 2002; O’Hara et
al., 2000). Tous les isolats rapportés d’espèces de Providencia ont été isolés à partir de
cas cliniques chez les humains et les animaux et on en sait peu sur la source de ces
infections. Chez l'homme, ces bactéries sont la cause des infections urinaires
(Chander et al., 2006).
1.4.2.3. Morganella:
Le genre Morganella se compose actuellement d’une seule espèce avec deux sous-
espèces, morganii et sibonii (O'Hara et al., 2000). Morganella morganii est un
organisme facultatif, anaérobique et ne fermente pas le lactose (Sinan Bilgin et al.,
2003). Il se trouve normalement dans le sol, l'eau, les eaux usées, et il fait également
partie de flore fécale de l’homme (Chou et al., 2009). Ce bacille est reconnu comme
étant un pathogène commun responsable d’infections opportunistes dans les voies
respiratoires, urinaires et aussi les infections des plaies (Kim et al., 2007).
1.5. Yersinia:
Ces bactéries ont une croissance plus lente que celle des autres entérobactéries.
Elles ont toutes un tropisme pour les tissus lymphoïdes. On les rencontre dans diverses
espèces animales et plus particulièrement chez les rongeurs (Nauciel, 2000).
Ce genre bactérien comprend 3 espèces pathogènes pour l’homme : Yersinia pestis,
Y. enterocolitica et Y. pseudotuberculosis. Yersinia pestis est l’agent causatif de la
peste. Y. enterocolitica et Y. pseudotuberculosis sont des agents pathogènes d’origine
alimentaire, répandus dans l’environnement et fréquemment isolés à partir d’animaux
infectés (Branger et al., 2009).

Synthèse Bibliographique
11
1.6. Les entérobactéries rares ou récemment décrites:
En plus des entérobactéries courantes, 18 autres genres d’entérobactéries rares ou
récemment décrits (tableau 1) sont connus, mais ils sont moins fréquemment isolés
que les premiers. Ils ne sont généralement pas spontanément pathogènes pour
l’homme, les animaux ou les végétaux; ils peuvent se comporter dans quelques cas
comme des bactéries pathogènes opportunistes (Delarras, 2007). Monnet et Freney en
2000 ont retenu un regroupement fondé sur deux caractères biochimiques essentiels: la
dégradation du lactose détectée par le test ONPG et la production d’acétoine par le test
VP (Monnet et Freney, 2000).
Tableau 1. Entérobactéries rares ou récemment décrites (Delarras, 2007).
Test
ONPG Test VP Entérobactéries (genres)
+ + Cedesea, Ewingella, Pantoea et Rahnella.
+ - Budvicia, Buttiauxella, Kluyvera, Leclercia, Moellerella,
Trabulsiella et Yokenella.
- Aucune
indication
Edwardsiella, Leminorella, Obesumbacterium, Pragia,
Photorhabdus, Taumella et Xenorhabdus.
2. La résistance aux antibiotiques :
Un antibiotique est une substance antibactérienne d'origine biologique, c'est à dire
produit par des micro-organismes (champignons microscopiques et bactéries) ou de
synthèse chimique et qui est capable d'inhiber la multiplication ou de détruire les
micro-organismes (Yala et al., 2001). C'est en 1941 que fut utilisé pour la première
fois un antibiotique, la pénicilline, découverte en 1928 par le bactériologiste anglais
Alexander Fleming pour traiter un patient atteint de septicémie à staphylocoques
(Besassier et al., 2005).

Synthèse Bibliographique
12
Les antibiotiques constituent un important groupe de médicaments pour la
médecine. À coté de leurs propriétés de lutter contre les infections humaines dues aux
bactéries pathogènes, ils sont également utilisés en médecine vétérinaire (Emmanuel,
2003). Les antibiotiques agissent à un niveau précis dans les structures bactériennes et
chaque famille possède son site d’action propre (figure 1):
Action sur la paroi bactérienne;
Action sur la structure de la membrane;
Action sur la synthèse protéique;
Action sur la synthèse de l’ADN;
Action sur le métabolisme de l’acide folique (Tissot, 2005).
Figure 1. Cibles des principaux antibiotiques (Benlmouden et Hakkou, 2007).
a. Paroi bactérienne ; b. Espace périplasmique ; 1. -lactamine (PLP) ; 2. Glycopeptides (D-
ala) ; 3. Dihydroptéorate synthétase (sulfamides) ; 4. Fixation à la sous-unité 50 S du
ribosome (macrolides, synergistines, lincosamides, phénicolés) ; 5. Fixation à la sous-unité 30
S du ribosome (aminosides, tétracyclines) ; 6. Acides nucléiques (quinolones, rifamycines,
nitro-imidazolés) ; 7. Membranes cytoplasmiques (polymyxines).

Synthèse Bibliographique
13
De la fin des années 1940 jusqu’aux années 1970, de nombreuses molécules
antibiotiques d’origine naturelle ou synthétique ont été découvertes. Le succès
fulgurant des premiers traitements anti-infectieux a fait considérer un peu hâtivement
le problème des maladies infectieuses comme définitivement réglé. Mais, rapidement,
l’enthousiasme a décliné avec l’apparition des premières résistances des bactéries aux
antibiotiques. À chaque nouvel antibiotique introduit en thérapeutique, les bactéries
ont su s’adapter et résister plus ou moins vite (Ploy et al., 2005). Ces résistances
peuvent avoir un spectre étroit, limité à un ou quelques antibiotiques de structure
voisine, mais on observe depuis plusieurs années l’émergence de mécanismes de
résistances croisées à des drogues de structures et de modes d’actions différents
(Walsh, 2000). Apparaissent aujourd’hui de véritables « monstres » bactériens
résistants à tous les antibiotiques potentiellement actifs (Ploy et al., 2000).
2.1. Support génétique de la résistance:
Il existe deux grands types de résistance aux antibiotiques: La résistance intrinsèque
et la résistance acquise.
La résistance intrinsèque (ou naturelle ou insensibilité): c’est-à-dire bactéries
dont tous les représentants au sein de l’espèce sont résistants dans la nature à tel
ou tel antibiotique, indépendamment de la présence de celui-ci (Skurnik et
Andremont, 2006);
A l’inverse, la résistance acquise n’est présente que chez certaines souches de
la même espèce ou du même genre; dans certains cas, elle peut concerner la
grande majorité de ces souches comme, par exemple, la production de
pénicillinase chez le staphylocoque qui intéresse plus de 90 % des souches
(Courvalin, 2008).
Sur le plan génétique, la résistance peut être acquise par deux voies totalement
distinctes, soit des mutations affectant des gènes présents sur le chromosome, soit
l’acquisition de gènes étrangers. Ces gènes peuvent provenir du chromosome
d’espèces différentes ou être véhiculés par des éléments génétiques mobiles pouvant

Synthèse Bibliographique
14
être transférés d’une bactérie à l’autre, on parle alors de transmission horizontale
(Galimand et al., 2005). L’ADN exogène peut provenir de cellules bactériennes
appartenant à une autre souche de la même espèce ou même d’une espèce, voire d’un
genre différent (Fauchère et Avril, 2002). Le transfert horizontal est le principal
mécanisme responsable de la dissémination des gènes de résistance au sein du monde
bactérien et concerne 80 % des cas de résistances aux antibiotiques rencontrés en
médecine humaine. Le plus souvent, lors de ce transfert, les gènes sont véhiculés par
des éléments génétiques mobiles, plasmides ou transposons (Ploy et al., 2005).
Au cours des années 1980, de nouveaux éléments génétiques susceptibles
d’acquérir ou de perdre des gènes de résistance aux antibiotiques ont été décrits par
Stokes et Hall et désignés sous le nom d’intégrons (Miriagou et al., 2006; Ploy et al.,
2005). Les intégrons constituent un système de capture et d’expression de gènes
contenus dans des cassettes. Les cassettes sont des éléments mobiles capables d’être
intégrés ou excisés par un système de recombinaison spécifique de site médié par une
intégrase. Les intégrons ne sont pas mobiles par eux-mêmes; ils sont incapables
d’autoréplication et sont généralement portés par des plasmides ou des transposons
(Fluit et Schmitz, 2004; Faure, 2009b).
2.2. Mécanismes biochimiques de la résistance aux antibiotiques :
Une souche bactérienne est dite résistante à un antibiotique quand elle est capable
de se développer en présence d’une concentration élevée de cet antibiotique (Lavigne,
2007). Alors ces micro-organismes ont développé trois types de mécanismes de
résistance (tableau 2).
Le premier est la production d’enzymes inactivatrices des antibiotiques (ex.-
lactamases);
Le deuxième est la modification des cibles des antibiotiques empêchant l’action
de ces derniers, comme par exemple la résistance aux fluoroquinolones par
modification des topoisomérases de classe II;

Synthèse Bibliographique
15
Enfin, le troisième mécanisme est la réduction des concentrations
intracellulaires d’antibiotique. Ce phénomène peut être dû à un transport actif
vers l’extérieur de la cellule via des transporteurs membranaires appelés
pompes d’efflux et/ou à une imperméabilité (Cattoir, 2004) (ex. résistance à
l’imipénème causée par la surproduction de AmpC pouvant être associée à des
perte de porines de la membrane externe et/ou la surexpression des pompes à
efflux) (Penteado et al., 2009).
Tableau 2. Les mécanismes de résistance bactérienne aux antibiotiques
(Li et Nikaido, 2004).
Antibiotique Cible
bactérienne
Mécanismes de résistance
Inactivation
enzymatique
Modification
de cible
Imperméabilité
cellulaire
Efflux
actif
Inhibition de la synthèse de la paroi (peptidoglycane)
-lactamines PLP
+++ ++ ++ ++
Glycopeptides Précurseurs
D-Ala-D-Ala +++
Inhibition de la synthèse protéique
Aminosides ARN ribosomal
30S +++ ++ + +
MLS ARN ribosomal
50S + +++ ++
Tétracyclines ARN ribosomal
30S ++ +++
Phénicolés ARN ribosomal
50S ++ ++
Oxazolidinones ARN ribosomal
50S ++
Inhibition de la synthèse ou de fonctionnement de l’ADN
Fluoroquinolones Topoisomérases +++ ++
Rifamycines ARN
polymérase +++ +
Sulfamides DHFS ++ +
Triméthoprimes DHFR ++ +

Synthèse Bibliographique
16
3. La résistance des entérobactéries aux -lactamines:
3.1. Définition des -lactamines:
Les β-lactamines sont les antibiotiques les plus utilisés dans la pratique clinique
courante (Rodriguez-Villalobos et Struelens, 2006) surtout dans le traitement des
infections dues aux entérobactéries (Zogheib et Dupont, 2005). Ce nom est du au fait
que tous les membres de cette classe portent une fonction lactame en position (figure
2) (Bessard, 2005).
Figure 2. Cycle -lactame (Bessard, 2004).
La famille des -lactamines comprend un grand nombre de molécules, toutes
caractérisées par la présence d’un cycle -lactame indispensable à l’activité
antibiotique, une faible toxicité, associées à un mode d’action fort complexe sur des
protéines de la membrane cytoplasmique, dénommées protéines liant la pénicilline
(PLP) ou penicillin binding proteins (Cavallo et al., 2004).
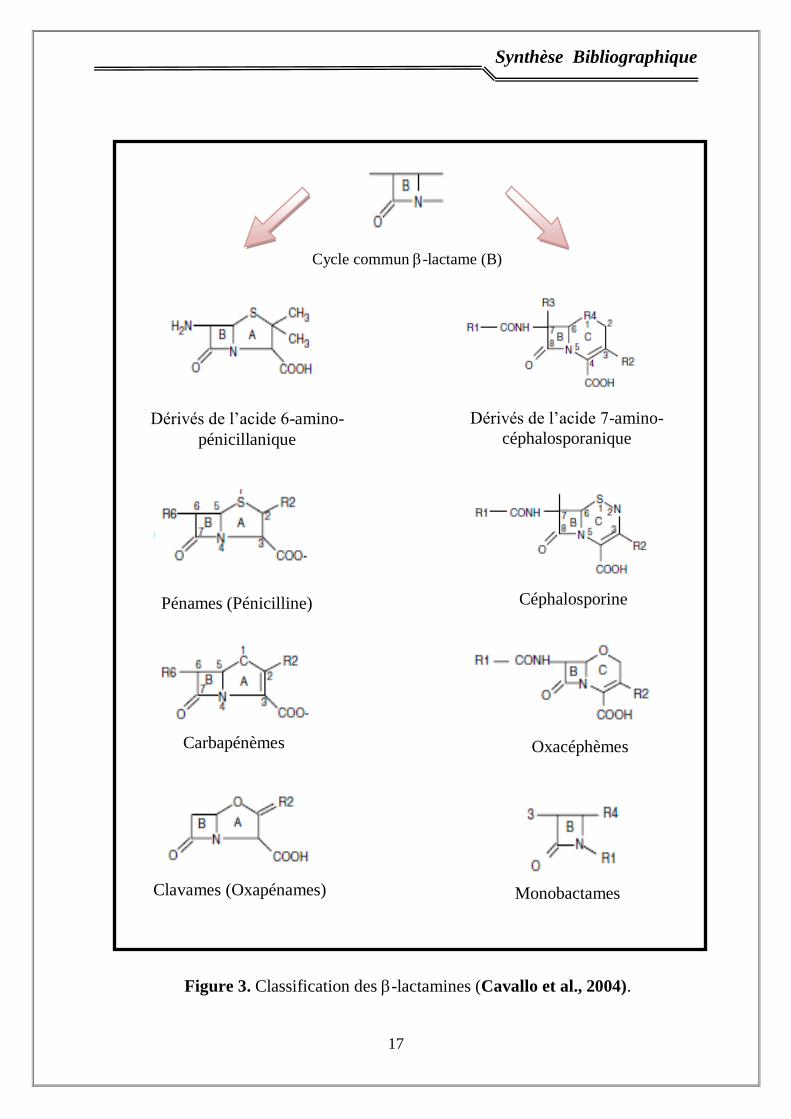
Synthèse Bibliographique
17
Figure 3. Classification des -lactamines (Cavallo et al., 2004).
Cycle commun -lactame (B)
Dérivés de l’acide 7-amino-
céphalosporanique
Dérivés de l’acide 6-amino-
pénicillanique
Pénames (Pénicilline) h Céphalosporine
Carbapénèmes Oxacéphèmes
Clavames (Oxapénames) Monobactames

Synthèse Bibliographique
18
3.2. Mécanisme d’action:
Les -lactamines sont des antibiotiques bactéricides principalement temps-
dépendant (Aubert et Carricajo, 2004) qui inhibent la synthèse de la paroi
bactérienne. Les transpeptidases et carboxypepytidases, enzymes associées à la
membrane cytoplasmique, fixent de façon covalente ces antibiotiques. Cette liaison est
due à une analogie structurale entre le substrat naturel de ces enzymes, l’acyl-D -
alanyl- D- alanine et le cycle -lactame. Ces enzymes qui lient les pénicillines et les
céphalosporines, sont également dénommées protéines de liaison aux pénicillines
(PLP). La nature de ces PLP est relativement spécifique d’espèce et leur nombre varie
d’une espèce bactérienne à une autre. Chacune a une fonction bien définie, mais
certaines jouent un rôle prépondérant dans la synthèse du peptidoglycane (Abdou
Souley Lie, 2002), c’est-à-dire l’étape de polymérisation à partir de sous unités faites
d’un disaccharide peptide. L’activité enzymatique des PLP est inhibée par leur liaison
avec les -lactamines (Nauciel, 2000).
3.3. Mécanisme de résistance:
Les bactéries ont développé différents mécanismes pour contrecarrer l’action des -
lactamines (Gangoue Pieboji, 2007): enzymatique ou non enzymatique.
3.3.1. La résistance non enzymatique aux -lactamines:
3.3.1.1. Imperméabilité:
Pour atteindre leurs cibles situées à la surface de la membrane cytoplasmique, les -
lactamines doivent diffuser aux travers de canaux spécialisés appelés porines. La diffusion
est en fonction de la charge, de la masse moléculaire et de la polarité des molécules. Les -
lactamines touchées diffèrent selon la porine absente. La disparition de porine provoque
l’augmentation des concentrations minimales inhibitrices de certaines -lactamines comme
cela a été mis en évidence chez certaines entérobactéries (Enterobacter aerogenes,
Klebsiella pneumoniae, Serratia marcescens, Enterobacter cloacae, Salmonella
typhimurium et Escherichia coli) (Nicaido, 2000).

Synthèse Bibliographique
19
3.3.1.2. Modification de la cible PLP:
Plusieurs facteurs peuvent concourir à la résistance par modification de la cible :
perte d’affinité des PLP pour les -lactamines par mutation, acquisition de gène ou
fragments de gènes codant pour les PLP d’affinité diminuée ou hyperproduction de
PLP normales. Des souches de Proteus mirabilis résistantes à l’imipénème et au
mécillinam ont été observées suite à une perte d’affinité de la PLP 2 et à une
diminution de la quantité de PLP 1A. Cependant, ce type de mécanisme de résistance
reste très rare chez les entérobactéries (Bonnet, 2006).
3.3.1.3. Pompe à efflux:
Acquisition ou surproduction des pompes efflux peuvent expulser l’antibiotique
hors de la cellule même contre le gradient de concentration (Gangoue Pieboji, 2007).
Chez les bactéries à Gram-négatif, l’imperméabilité de la membrane externe et les
pompes à efflux jouent en synergie un rôle important dans la résistance intrinsèque de
ces bactéries. Les mutations ou/et les modifications inductibles de ces mécanisme peut
conduire à une diminution de l’afflux et l’augmentation de l’efflux des antibiotiques,
ainsi d’empêcher l’accès des antibiotiques à leur cibles. Toutefois, les niveaux de
résistance conférés par ces mécanismes devraient être faibles, mais ils sont renforcés
par l’interaction entre les mécanismes non enzymatiques et enzymatiques (par
production de -lactamases). Les expressions élevées des pompes à efflux ont été
observés chez Escherichia coli et Salmonella spp. dans l’alimentation des animaux (Li
et al., 2007).
3.3.2. La résistance enzymatique aux -lactamines (-lactamases):
3.3.2.1. Définition:
La résistance bactérienne aux β-lactamines est due principalement à la production
d’enzymes (β-lactamases) capables d’hydrolyser l’anneau β-lactame commun à cette
classe d’antibiotiques (Rodriguez-Villalobos et Struelens, 2006). Les -lactamases

Synthèse Bibliographique
20
sont des enzymes d'inactivation (Philippon, 2005), appartenant à la grande famille des
hydrolases d'amides cycliques (De Wals, 2007).
Sur la base du mécanisme catalytique, on distingue:
les enzymes à sérine active réparties en classes A, C et D ont en commun un
résidu sérine dans le site actif;
Les métallo-enzymes: les enzymes de classes B ont besoins d'un ion métallique
Zn2+
catalysant leurs activités (Materon et al., 2004).
3.3.2.2. Mécanisme d’action:
Les -lactamases sont des enzymes d’inactivation hydrolysant le pont amide du
cycle -lactame pour donner un acyl-enzyme qui sera ensuite dégradé en acide inactif.
Ainsi, les pénicillines sont dégradées en acide pénicilloïque et les céphalosporines en
acide céphalosporoïque. Ces enzymes sont localisées au niveau de l’espace
périplasmique chez les bactéries à Gram négatif (Barrial et Scotet, 2005).
Les -lactamases ont une structure proche des enzymes impliquées dans la synthèse
du peptidoglycane de la bactérie. Le mode d’action des -lactamines sur une bactérie
sensible consiste à entraîner une erreur des peptidases aboutissant à un défaut de
synthèse du peptidoglycane, ce qui provoque la mort bactérienne. Pour éviter que les
peptidases ne se « trompent », la bactérie synthétise une -lactamase qui va hydrolyser
le cycle -lactame. Son ouverture va empêcher sa reconnaissance par la peptidase et
donc la synthèse du peptidoglycane est possible: la multiplication bactérienne n’est
alors pas affectée (Lavigne et al., 2002). La différence majeure entre les 2 types
d'enzymes (les -lactamases et les PLP) consiste dans la vitesse avec laquelle l'acyl-
enzyme est hydrolysé. En effet, si les PLP ne sont capables d'hydrolyser qu'une -
lactame par heure de tel sorte que l'acyl-enzyme apparaît dans ce cas comme un
intermédiaire stable, les -lactamases les plus efficaces en hydrolysent 1000 par
seconde, rendant l'antibiotique totalement inactif et régénérant l'enzyme pour une
nouvelle réaction d'hydrolyse (Tulkens et Spinewine, 2002).

Synthèse Bibliographique
21
Figure 4. Mécanisme d’hydrolyse d’une -lactamine par une -lactamase
(Ruppé, 2010).
3.3.2.3. Classification des -Lactamases:
Les -lactamases sont des enzymes d’une extrême diversité (Gautier, 2007).
Actuellement, plus de 400 β-lactamases, dont plus de 200 BLSE, sont décrites et
aucune β-lactamine, seule ou en association avec des inhibiteurs de β-lactamases, n’est
invulnérable à leur action potentielle (Li et al., 2007; Rodriguez-Villalobos et
Struelens, 2006). De nombreuses classifications existent, qui prennent comme
critères: le phénotype, la nature du site actif, la séquence d’acide aminé, les
caractéristiques physiques et l’origine (Vidon et Bourdin, 2005).
Les deux schémas les plus utilisés pour la classification des -Lactamases sont
celui d’Ambler qui est fondé sur l’homologie de séquence des acides aminés et celui
de Bush-Jacoby-Medeiros fondé sur les propriétés fonctionnelles des enzymes
(Nordmann et Poirel, 2002).
La classification d’Ambler divise les β-lactamases en quatre groupes (de A à
D). Cette classification moléculaire, impliquant la connaissance de la structure
primaire des enzymes, a été suggérée par Ambler en 1980 (Bryskier, 1999);

Synthèse Bibliographique
22
En 1995, Bush-Jacoby-Medeiros a présenté le dernier système de classification
qui repose sur quatre groupes (1-4) et sous-groupes (A à F) (Shah et al., 2004).
Elle est fondée sur les caractéristiques physicochimiques des enzymes comme
leur point isoélectrique, poids moléculaire, substrat d’activité et profil
d’inhibition (Nordmann et Poirel, 2002).
3.4. Phénotypes de résistance aux -lactamines:
3.4.1. Résistance naturelle ou phénotypes « sauvages »:
Les entérobactéries ont une résistance naturelle à certaines familles d’antibiotiques
hydrophobes comme les pénicillines G et M, les macrolides et apparentés
(lincosamides, synergistines) et les glycopeptides. De nombreuses classes
d’antibiotiques restent cependant actives comme la plupart des -lactamines, les
aminoglycosides, les tétracyclines, les quinolones et les sulfamides. Classiquement, on
classe les entérobactéries en groupes en fonction de leur sensibilité naturelle aux -
lactamines (Zogheib et Dupont, 2005).
Groupe 0: Phénotype « sensible » d’espèces dépourvues de gènes de -lactamases
Salmonella spp. et Proteus mirabilis sont dépourvus de -lactamase à l’état
« sauvage » et sont naturellement sensibles aux amino-pénicillines, carboxy-
pénicillines, uréido-pénicillines, à l’aztréonam, aux céphalosporines et aux
carbapénèmes (Bonnet, 2006).
Groupe 1: Phénotype « sensible » d’espèces produisant naturellement une
céphalosporinase de classe C
Une β-lactamase de classe C constitutive à bas niveau est présente chez les souches
sauvages d’Escherichia coli et Shigella spp. et ne confère de résistance qu’aux -
lactamines qui pénètrent faiblement dans l’espace périplasmique (benzylpénicilline,
cefsulodine) (Gautier, 2007).

Synthèse Bibliographique
23
Groupe 2 : Phénotype « pénicillinase de bas niveau »
Ce phénotype, observé chez Klebsiella (K.pneumoniae, K.oxytoca), Citrobacter
koseri (C.diversus), Citrobacter amalonaticus et Escherichia hermannii, se caractérise
par la résistance à bas niveau de ces espèces à l’amoxicilline et à la ticarcilline, par la
sensibilité à la céfalotine, au céfotaxime et aux autres -lactamines (Vedel, 1998).
Groupe 3 : Phénotype « céphalosporinase de bas niveau »
Des espèces comme Enterobacter cloacae, Enterobacter aerogenes, Citrobacter
freundii, Morganella morganii, Serratia marcescens, Hafnia alvei, Yersinia
enterocolitica, Providencia stuartii et Providencia rettgeri possèdent une
céphalosporinase inductible, leur conférant une résistance aux amino-pénicillines, aux
céphalosporines de 1ere
génération et à l’action de l’acide clavulanique. Ce caractère
inductible est détecté dans l’antibiogramme en milieu gélosé par un antagonisme
("écrasement" du diamètre d’inhibition) entre une β-lactamine inductrice (imipénème,
céfoxitine) et une céphalosporine de 3eme
génération. Ce type d’expression est le fait
d’un mécanisme complexe faisant intervenir plusieurs gènes régulateurs (ampR,
ampD, et ampG) (Philippon et Arlet, 2006).
Groupe 4 : Yersinia enterocolitica et Serratia fonticola
Elles sont résistantes aux amino-pénicillines, uréido-pénicillines, carboxy-
pénicillines et aux céphalosporines de 1ere
génération par l’action associée d’une
pénicillinase et d’une céphalosporinase naturelle produite à bas niveau (Zomahoun,
2004).
Groupe 5 : Phénotype « céfuroximase »
Proteus vulgaris et Proteus penneri possèdent une -lactamase particulière parfois
dénommée "céfuroximase" qui rend ces espèces résistantes aux pénicillines du groupe
A et aux céphalosporines de 1ere
et 2eme
générations (à l’exception des céphamycines
comme la céfoxitine) mais dont l’activité enzymatiques et inhibe par l’action d’acide
clavulanique (Denis et Ploy, 2007).

Synthèse Bibliographique
24
Groupe 6 : Phénotype « -lactamase à spectre étendu chromosomique »
Les entérobactéries d'isolement très rare tell que Kluyvera montrant un phénotype
de résistance inhabituel "pénicillinase de bas niveau". Ce phénotype est nettement à
distinguer de celui "pénicillinase de bas niveau" (Philippon, 2005). Il s’agit une BLSE
naturelle exprimé à bas niveau (Denis et Ploy, 2007). Après clonage du gène
correspondant (KLUA-1) dans une souche réceptrice d’E.coli, a été identifié, par la
suite, un des progénitures de BLSE plasmidiques de type CTX-M (Philippon, 2005).
3.4.2. Résistance acquise ou phénotypes « résistants »:
Toutes les entérobactéries, quel que soit leur groupe, sont capables d’intégrer des
gènes de résistance codant pour une -lactamase (Zogheib et Dupont, 2005).
Phénotype « pénicillinase de haut niveau » ou « pénicillinase acquise »
Les pénicillinases des bacilles à Gram négatif sont nombreuses (TEM-1, TEM-2,
SHV-1…) et non inductibles. Environ 75% de -lactamases isolées des entérobactéries
sont des TEM-1. Ces enzymes sont codées par des plasmides et donc facilement
transférables. Elles confèrent aux bactéries, qui les produisent à haut niveau, une
résistance aux amino-pénicillines, carboxy-pénicillines, uréido-pénicillines, amidino-
pénicillines, C1G et de C2G. Cependant elles conservent leur sensibilité aux C3G,
céfamycines, monobactames et carbapénèmes. Elles sont inhibées plus ou moins par
les inhibiteurs enzymatiques (Collignon et al., 2007).
Phénotype « pénicillinase résistante aux inhibiteurs »
Plus récemment, des -lactamases dérivées de pénicillinases plasmidiques
entraînant une résistance aux inhibiteurs suicides de -lactamases ont été décrites.
Elles sont produites par Escherichia coli, Proteus et Klebsiella. Elles confèrent une
résistance à l’amoxicilline et à la ticarcilline, seules ou en association avec l’acide
clavulanique et un bas niveau de résistance aux C1G. Pour la pipéracilline, le niveau
de résistance est plus faible (Lavigne et al., 2002).

Synthèse Bibliographique
25
Phénotype « -lactamase à spectre étendu »
Classiquement, les BLSE sont définies comme des enzymes, appartenant à la classe
A ou D de la classification d'Ambler (Rodriguez-Villalobos et Struelens, 2006),
capables d’hydrolyser et causer une résistance aux oxymino-céphalosporines
(céfotaxime, céftazidime, céftriaxone, céfuroxime et céfépime) et les monobactames
(aztréonam), mais pas aux céphamycines (céfoxitine et céfotetan) ou carbapénèmes
(imipénème, méropénème, doripénème et ertapénème). Ces enzymes sont inhibés par
les inhibiteurs de -lactamases telle que l’acide clavulanique, le sulbactam et le
tazobactam (Peirano et Pitout, 2010).
La majorité des BLSE sont dérivées de mutations ponctuelles dans la séquence
génétique codant pour le site actif des 1ere
-lactamases connues (TEM-1, TEM-2 et
SHV-1) (Paterson et Bonomo, 2005).
Plus de 200 BLSE naturelles ont été décrites; elles ont été classées en 11 familles
différentes sur la base de leur séquence d’acides aminés: TEM, SHV, CTX-M, PER,
VEB, GES, TLA, BES, SFO, FEC et OXA ((Vidon et Bourdin, 2005). Les 4 familles
majeures sont représentées par les enzymes de type TEM, SHV, OXA et CTX-M
(Gniadkowski, 2001).
Récemment, les BLSE de type CTX-M ont émergé comme le type le plus courant
de BLSE, avec une répartition mondiale (Seonghan et al., 2007). Découvertes pour la
première fois au milieu des années 1980, elles montrent un niveau de dissémination
dans les bactéries et partout dans le monde qui augmente dramatiquement depuis 1995
(Bonnet, 2004). Les enzymes de la famille CTX-M comprennent aujourd’hui plus de
90 enzymes (Ruppé, 2010) isolées de nombreuses espèces d’entérobactéries et qui, sur
la base de leurs séquences protéiques, sont divisés en 5 groupes (Bonnet, 2004). Elles
sont similaires aux autres -Lactamases à spectre étendu dans la mesure où elles
hydrolysent à large spectre les céphalosporines et l’aztréonam mais agissant comme
des cefotaximases (Brenwald et al., 2008).

Synthèse Bibliographique
26
Phénotype « Hyper OXY »
Chez Klebsiella oxytoca, des mutations ponctuelles dans la région du promoteur de
transcription des enzymes chromosomiques K1 (type OXY-1 ou OXY-2), conduisent à
augmentation du niveau d’expression et se traduisent in vivo par une résistance de
haut niveau aux pénicillines, aux céphalosporines de 1ere
et 2eme
génération, ainsi qu’à
l’aztréonam. Dans ce cas, une synergie peut être détectée entre les céphalosporines de
3eme
génération ou l’aztréonam, et le clavulanate (Sougakoff et Trystram, 2003).
Phénotype « céphalosporinase de haut niveau »
Le phénotype de résistance céphalosporinase de haut niveau se traduit par une
résistance à l’ensemble des -lactamines excepté les carbapénèmes (Eyquem et
Montagnier, 2000). Il peut subsister une activité des céphalosporines à large spectre
(céfépime, cefpirome). Il s’agit d’un phénotype retrouvé principalement chez les
bactéries possédant naturellement une céphalosporinase ampC qui peut être alors
surexprimée (Enterobacter cloacae, Citrobacter freundii, Escherichia coli,
Morganella morganii et autres entérobactéries du même groupe) (Gueudet et al.,
2010).
Depuis 1990, des céphalosporinases codées par des gènes plasmidiques ont été
décrites. Parmi celles-ci, il y a MIR-1, BIL-1 et CMY-2 (Seck, 2005).
Cette résistance a été retrouvée chez Klebsiella pneumoniae mais également chez
d’autres entérobactéries. Ces enzymes à médiation plasmidique dérivent des
céphalosporinases chromosomiques d’Enterobacter (ACT-1 et MIR-1), de Citrobacter
(CMY), de Morganella (DHA), d’Hafnia (ACC-1) et d’autres entérobactéries (MOX,
FOX. . .) (Philippon et Arlet, 2006).

Synthèse Bibliographique
27
3.4.3. Résistance aux carbapénèmes:
Les carbapénèmes sont aujourd’hui parmi les traitements de choix des infections
sévères dues aux entérobactéries productrices de -lactamases à spectre étendu
(BLSE). Mais leur utilisation pourrait être compromise par l’émergence de souches de
bactéries résistantes également aux carbapénèmes (Cuzon et al., 2009).
Chez les entérobactéries, cette résistance est principalement due à trois mécanismes
(Livermore Woodford, 2006):
i. L’association entre la diminution de la perméabilité membranaire avec la
production d’une céphalosporinase (chromosomique ou plasmidique) ou d’une
BLSE (Patel et al., 2009): chez les entérobactéries, une surproduction de
céphalosporinase ou d’une BLSE associée à une perte de porine de la membrane
externe et/ou une surexpression de pompes à efflux (Castanheira et al., 2008) est
la cause la plus fréquente de résistance à l’imipénème (Bonnet, 2006);
ii. Acquisition des métallo--lactamases (VIM, IMP): ces enzymes, appartenant au
classe B d’ambler, hydrolysent toutes les -lactamases à l’exception de
l’aztréonam (Trigoso, 2010);
iii. Acquisition de non métallo--lactamases: ces enzymes sont soit les oxacillinases
(la famille des OXA), soit les carbapénèmases, inhibées par l’acide clavulanique,
qui peuvent être codées par des gènes chromosomiques, décrits pour des souches
de Serratia marcescens (SME1) et Enterobacter cloacae (IMTI NMC-A), ou
plasmidiques comme certains variants de GES et Klebsiella pneumoniae
carbapénèmase (KPC) (Nordmann et al., 2009; Pourriat et Martin, 2005 ).
Actuellement, les carbapénèmases qui sont les plus répandues chez les
entérobactéries sont les enzymes de type KPC. Ces enzymes ont principalement été
décrites chez Klebsiella pneumoniae avec un support plasmidique. Elles possèdent un
pouvoir de dissémination important et sont émergentes dans de nombreux pays
(Lautenbach et al., 2010).

Synthèse Bibliographique
28
4. La résistance aux aminosides:
Les aminosides, également appelés aminoglycosides, oligosaccharides ou amino-
cyclitols sont constitués de sucres aminés dérivés du noyau 2 désoxystreptamine et
élaborés par des Actinomycètes (Amikacine, Tobramycine) ou des Micromonospora
(Gentamicine) (Ezaitouni et al., 1999). Après fixation à des sites chargés
négativement sur la paroi bactérienne (Faure, 2009a), en raison de leur caractère
polycationique, ils diffusent dans les bactéries à travers les canaux porines jusqu'à
l'espace périplasmique. Leur pénétration à travers la membrane cytoplasmique est
active et nécessite un apport d'énergie et la présence d'oxygène. Les aminosides
perturbent la synthèse protéique bactérienne (Benlmouden et Hakkou, 2007) en se
liant au site aminoacyl de l'ARN ribosomal 16S (ARNr), dans la sous-unité 30S du
ribosome (Durante-Mangoni et al., 2009 ). Cette liaison de l’antibiotique permet:
l’inhibition de l’étape d’élongation (en empêchant le transfert du peptidyl-
ARNt depuis le site A vers le site P) conduisant à un arrêt de la synthèse
protéique;
l’introduction d’erreurs dans la lecture des codons de l’ARNm, engendrant la
production de protéines aberrantes.
C’est l’accumulation des protéines erronées synthétisées qui est responsable de la
létalité induite par les aminosides (Faure, 2009a).
Ce sont des antibiotiques bactéricides concentration-dépendante (Aubert et
Carricajo, 2004). Ils sont réservés au traitement des infections sévères à bactéries à
Gram négatif et à staphylocoques notamment dans leurs manifestations pulmonaires,
endocardites, rénales et bactériémiques. Ils sont souvent employés dans le traitement
probabiliste de ces infections en association avec une -lactamine ou une
fluoroquinolone (Martin, 2008).
Ces dernières années, l’utilisation des aminosides comme des antibiotiques à large
spectre importante en clinique a diminué considérablement en raison de l’émergence
des souches bactériennes résistantes (Thuresson et al., 2007). La résistance à ces

Synthèse Bibliographique
29
médicaments est devenue un grave problème de santé publique. Le mécanisme de
résistance principal aux aminoglycosides est l’acquisition d’enzymes bactériennes qui
modifient les antibiotiques par acétyltransférase (AAC), adényltransférase (ANT) ou
phosphotransférase (APH). Chaque classe de ces enzymes effectue une réaction régio-
spécifique soit sur le groupe amine (AAC) ou un groupe hydroxyle (ANT et APH) et
les produits de ces réactions sont dépourvus d'activité antibactérienne (Chen et al.,
2008).
La diminution de l'accumulation de l’antibiotique est un autre mécanisme majeur de
résistance. Les bactéries peuvent exprimer des systèmes d’efflux qui résultent d’une
accumulation réduite des aminosides à l’intérieur de la cellule (Durante-Mangoni et
al., 2009).
La méthylation de l'ARNr 16S au sein de la sous-unité 30S des espèces comme
Escherichia coli et Klebsiella altère la fixation des aminosides, ce qui bloque leurs
activités antimicrobiennes (Doi et Arakawa, 2007).
5. La résistance aux quinolones:
Les quinolones, découvertes en 1962 par Lesher, sont des antibiotiques
synthétiques. Du fait de leur bonne diffusion tissulaire, ces antibiotiques sont
largement utilisés en médecine humaine et vétérinaire, notamment dans le cas
d’infections urinaires et respiratoires (Meradi et al., 2009).
Les quinolones agissent en se fixant au niveau de l’hélice 4 topoisomérase II, la
topoisomérase IV et de l’ADN gyrase. Le complexe ternaire ainsi formé, molécule
d’antibiotique, enzyme et ADN, empêche la progression de la fourche de réplication
(Guillard et al., 2008).
Les quinolones de 1ere
génération, dont le chef de file est l’acide nalidixique,
n’agissent que sur les bacilles à gram négatif et ne sont utilisées que dans le traitement
des infections urinaires. Les quinolones de 2eme
génération ou fluoroquinolones,
utilisées depuis les années 80, comprennent principalement la pefloxacilline,

Synthèse Bibliographique
30
l’ofloxacine, la ciprofloxacilline. Elles sont beaucoup plus actives que les quinolones
de 1ere
génération (Nseir et al., 2005; Nauciel, 2000).
La résistance acquise à ces antibiotiques est le résultat de la combinaison de
plusieurs mécanismes. Pendant plus de 30 ans, les seuls mécanismes de résistance aux
quinolones connus avaient un support chromosomique (Skurnik et Andremont,
2006).
Les mutations responsables sont localisées dans des gènes qui conduisent soit à:
La perte d’affinité de l’antibiotique pour sa cible (mutation de la topoisomérase
dans la région quinolone resistance determining region [QRDR];
Une augmentation de son excrétion hors du cytoplasme (par surexpression des
systèmes d’efflux);
Une diminution de sa pénétration transmembranaire (par déficit quantitatif ou
qualitatif de la synthèse des porines) (Nordmann et Mammeri, 2007).
Le support de la résistance aux quinolones était supposé être uniquement
chromosomique jusqu’en 1998 où, Martinez- Martinez et al. ont décrit la première
souche (Klebsiella pneumoniae UAB1) dont le support de la résistance est un plasmide
transférable (pMG252). Le déterminant génétique de cette résistance est le gène qnr
dont la caractéristique est d’être porté par différents types d’intégrons. L’importance
de ce support est sa transférabilité et sa capacité à accélérer la diffusion de la
résistance aux quinolones. Les gènes qnr ont été identifiés dans différentes espèces
d’entérobactéries et souvent associés à la production de -lactamases à spectre élargi
(Guessennd et al., 2008).
À ce jour, trois qnr ont été mises en évidence, qnrA, qnrB et qnrS (Guillard et al.,
2009). Toutes ces protéines sont capables de se fixer sur les topoisomérases II et IV en
compétition avec l’ADN. La réduction du nombre des complexes binaires
topoisomérases-ADN diminue la fixation ultérieure des quinolones sur les
topoisomérases (Tran et al., 2005).

Synthèse Bibliographique
31
Les souches porteuses du gène qnr appartenaient le plus souvent aux espèces
Escherichia coli, Klebsiella pneumoniae et récemment Enterobacter cloacae, et sont
toutes multirésistantes, en particulier résistantes aux céphalosporines de 3eme
génération par production de -lactamase à spectre étendu (BLSE) (ex. SHV-7) ou de
céphalosporinase plasmidique (ex. FOX-5) (Honoré et al., 2006).
En 2005, un second mécanisme de résistance plasmidique a été identifié. Il s’agit
d’un variant de l’acétylase en 6-isoforme Ib dénommé AAC (6_)-Ib-cr (pour
ciprofloxacine resistance) et qui confère une résistance de bas niveau à la
ciprofloxacine et à la norfloxacine par acétylation enzymatique (Robicsek et al.,
2006).
6. La résistance aux autres antibiotiques:
6.1. Triméthoprime-Sulfaméthoxazole:
Le cotrimoxazole est une combinaison (synergique) de triméthoprime et de
sulfaméthoxazole, un sulfamide, deux drogues qui bloquent des réactions différentes
dans la même voie métabolique (Singleton, 2005). Le triméthoprime est un inhibiteur
de la déhydrofolate réductase qui peut potentialiser l’activité des sulfamides par
inhibition séquentielle de la synthèse de l’acide folique. Triméthoprimes, sulfamides et
leurs associations ont une activité thérapeutique bien établie en pathologie respiratoire,
digestive, urinaire, notamment en traitements courts et en prophylaxie (Bryskier,
1999).
De toutes les familles d’antibiotiques, l’association sulfamide-triméthoprime
possède indiscutablement la plus grande diversité de mécanismes de résistance acquise
et de leur support génétique: modification de la perméabilité, activation de pompes
d’efflux, modification quantitative ou qualitative des cibles, contournement
métabolique, hyperproduction de précurseurs, absence de certaines enzymes et toute
une variété de gènes exogènes acquis par la bactérie (Golsdein, 2006).

Synthèse Bibliographique
32
6.2. Polymyxines:
Les polymyxines ont été découvertes en 1947. Bien que 5 molécules de polymyxine
soient connus de manière séquentielle et nommées polymyxine A à E, seules 2 sont
disponibles pour un usage thérapeutique: polymyxine B et la polymyxine E (colistine)
(Hermsen et al., 2003). L’antibiotique le plus utilisé est la colistine. Elle n’agit que
sur les bacilles à Gram négatif et elle reste cependant inactive sur les Proteus,
Providencia et Serratia (Nauciel et Vildé, 2005).
La colistine est un polypeptide de la famille des polymyxines du groupe E, produit
par Bacillus polymyxa subsp colistinus. Il s’agit d’un mélange complexe d’au moins
30 composés, dont les deux majeurs sont la colistine A (polymyxine E1) et la colistine
B (polymyxine E2) (Frasca et al., 2008).
Les polymyxines agissent sur la paroi cellulaire des bactéries à Gram négatif par
l'intermédiaire de trois mécanismes connus.
Les polymyxines sont des molécules cationiques, qui perturbent
électrostatiquement les membranes de surface bactérienne par le déplacement
des ions Ca2+
et Mg2+
qui stabilisent les molécules de lipo-polysaccharide
(LPS);
Les polymyxines sont des agents tensio-actifs amphipathiques contenant à la
fois des groupements lipophiles et lipophobes. Ils pénètrent dans les membranes
cellulaires et interagissent avec des phospholipides, conduisant à des
changements de perméabilité qui perturbent rapidement les membranes et
résultent la mort cellulaire;
Les polymyxines peuvent également se lier au lipide A de l'endotoxine ou
molécule LPS et, dans des études sur les animaux, ils bloquent plusieurs effets
biologiques des endotoxines (Chen et Kaye, 2009; Cohen, 2010).

Synthèse Bibliographique
33
Malheureusement, la colistine n'échappe pas au développement de résistance. Il
existe plusieurs mécanismes suggérés de résistance à la colistine pour les bactéries à
Gram négatif, dont la plupart impliquent des changements dans la membrane externe
(Li et al., 2006).
La résistance naturelle semble due à l’architecture de la paroi qui empêche la
polymyxine d’interagir avec les groupements phosphates cibles du LPS. Quant à la
résistance acquise, l’apparition de mutants au cours de traitement est très rare et il n’y
a pas de plasmides de résistance décrits (Li et al., 2005). À noter qu’il n’existe sans
doute pas de phénomènes d’imperméabilité membranaire ou d’efflux actif à cause de
leur haut poids moléculaire. Par contre, la résistance est totalement croisée entre la
colistine et la plymyxine B (Gattoir, 2006).

Matériel
Méthodes

Matériel et Méthodes
34
1. Matériel:
1.1. Matériel biologique:
1.1.1. Souches étudiées:
Le présent travail a pour but d’étudier 140 souches entérobactéries isolées à partir de
différents services du CHU de Sidi Bel Abbes.
1.1.2. Souches de références:
ATCC 27853: Pseudomonas aeruginosa de phénotype sauvage;
ATCC 25922: Escherichia coli;
VIM: Pseudomonas aeruginosa productrice de métallo--lactamase VIM;
K 12: Escherichia coli résistante à la rifampicine;
VP 517: Escherichia coli.
1.2. Milieux de culture:
1.2.1. Milieux de culture liquides:
BN : Bouillon Nutritif (Institut Pasteur d’Algérie);
BHIB : Bouillon Cœur Cerveau (Fluka);
MHL : Mueller Hinton Liquide (Fluka);
LB : Bouillon Luria (Sigma).
1.2.2. Milieux de culture solides:
Gélose nutritive (Fluka);
Mac Conkey (Fluka);
Mueller Hinton (Fluka);
TSI : Triple Sucre à Identifié.
1.3. Tests biochimiques:
Galerie API 20 E (Bio Mérieux);
Galerie API 32 E (Bio Mérieux).

Matériel et Méthodes
35
1.4. Solutions et Tampons:
Solution de l’EDTA 0,5 M pH 8;
Tampon TE;
Solution de lyse;
Phénol-Chloroform-Isoamylalcool (24 :25 :1) (Sigma);
Tampon de charge;
Tampon TBE 1X;
Solution de BET (Sigma).
1.5. Antibiotiques:
1.5.1. En disque:
-lactamines: Ampicilline (10 g), Amoxicilline (25 μg),
Amoxicilline/acide clavulanique (20/10 μg), Ticarcilline (75 μg),
Ticarcilline/acide clavulanique (75/10 g), Pipéracilline (75μg),
Pipéracilline/tazobactam (75/10μg), Imipénème (10 μg), Aztréonam (30
g), Céfalotine (30 g), Céfuroxime (30 g), Céfoxitine (30 g), Latamoxef
(30 g), Céfotaxime (30 g), Ceftriaxone (30 g), Ceftazidime (30 g),
Céfépime (30 g) et Cefpirome (30 g).
Aminosides: Kanamycine (30 UI), Tobramycine (30 μg), Amikacine (30
μg) et Gentamicine (15 μg).
Quinolones: Acide nalidixique (30 μg), Ofloxacine (5 μg) et Ciprofloxacine
(5 μg).
Autres: Colistine (50 g), Triméthoprime/Sulfaméthoxazole (75 μg) et
Rifampicine (30 μg).
1.5.2. En poudre:
-lactamines: Pipéracilline (DakotaR Pharm.), Pipéracilline/tazobactam
(Wyeth Pharmaceuticals), Imipénème (MSD), Céfoxitine (Panpharma),
Aztréonam (Sanofi- synthelabo), Céfotaxime (Aventis), Céftazidime

Matériel et Méthodes
36
(Glaxo Smith Kline), Céfépime (Bristol-Myers Squibb) et Orbénine
(Astellas).
Aminosides : Tobramycine (Merck), Amikacine (Bristol-Myers Squibb) et
Gentamicine (Panpharma).
Quinolones : Ciprofloxacine (Bayer Pharma).
Autre : Rifampicine (Saidal).
2. Méthodes :
2.1. Prélèvements:
Les prélèvements ont été réalisés par écouvillonnage sur des plaies suppurées,
d’escarres de malade et de l’environnement hospitalier ainsi que par la
récupération directe des sondes trachéales et vésicales, puis acheminés au
laboratoire (l’écouvillon est introduit dans un tube contenant du bouillon
nutritif afin d’assurer la survie des souches) pour être incubés dans l’étuve à
37°± 1 pendant 18 à 24H.
2.2. Isolement et purification:
On procède directement à une recherche de germes d’entérobactéries et pour
cela on réalise un ensemencement en strie sur toute la surface d’une gélose
Mac Conkey pour l’isolement des bactéries à Gram négatif grâce à l’action de
deux inhibiteurs présent dans le milieu. Le Cristal Violet pour l’inhibition de la
flore Gram positive, et les sels biliaires pour la sélection des Entérobactéries.
Ce milieu ensemencé, est incubé à 37°C ± 1°C pendant 24 H. Après une
lecture morphologique, on fait une purification des colonies par passage
successif sur bouillon nutritif puis sur gélose Mac Conkey.
2.3. Identification:
L’identification est l’étape qui précède toujours l’antibiogramme et elle est
prise à partir des cultures pures représentées par des colonies que l’on peut
différencier par leur aspect, forme, taille, bord, surface et chromogénèse.

Matériel et Méthodes
37
L’identification a été réalisée par le test TSI pour différencier entre les espèces
dites fermentants ou non-fermentants puis par les galeries d’identification.
2.3.1. Le test TSI (milieu triple sucres):
i. Principe:
Ce milieu permet d'étudier la fermentation de trois sucres (glucose, lactose,
saccharose), d'apprécier la production ou non de H2S et de noter la production
ou non de gaz à partir du glucose.
ii. Technique:
Elle consiste à ensemencer en stries serrées la pente de la gélose puis par
piqûre centrale le culot, la lecture se fait après 18H d’incubation à 37°C.
iii. lecture:
a)Lecture de glucose et du gaz au niveau du culot:
La fermentation du glucose se traduit par le virage au jaune du culot;
La production de gaz se traduit par la formation ou non de bulles de gaz
dans la masse du culot.
b) lecture de la pente:
la fermentation du glucose et /ou du saccharose se traduit par le virage au
jaune de la masse du culot.
c) production de H2S: se traduit par noircissement du milieu.
2.3.2. Galeries d’identification:
Le système API® Bio Mérieux (Appareillage et Procédé d’Identification) est
une version miniaturisée et standardisée des techniques biochimiques
conventionnelles pour l’identification des bactéries.

Matériel et Méthodes
38
Lorsqu’une suspension bactérienne de densité convenable est répartie dans les
différentes alvéoles qui composent la micro-galerie (contenant de substrats
déshydratés), les métabolites produits durant la période d’incubation se
traduisent par des changements de couleur spontanés ou révélés par addition de
réactifs.
Ces plaques permettent l’identification des entérobactéries et autres bacille à Gram
négatif ; l’API 20 E comprend 20 tests biochimiques alors que l’ ID 32 E est basé
sur 32 tests biochimiques miniaturisés.
La lecture de ces réactions est réalisée à l’aide du tableau de lecture (Annexe 1
et 2) et l’identification obtenue à l’aide du tableau d’identificat ion du
catalogue analytique (Annexe 3 et 4).
2.4. Antibiogramme (CASFM, 2010; Vedel, 1998):
2.4.1. Principe:
Antibiogramme standard ou les méthodes par diffusion sont plus utilisées par
les laboratoires cliniques. Elles consistent à disposer des disques de papier
buvard imprégnés de concentration déterminée d’antibiotiques à la surface
d’un milieu gélosé. Dès l’application des disques, l’antibiotique diffuse à partir
du disque de manière uniforme dans la gélose. Après incubation, les disques
s'entourent de zones d'inhibition circulaires correspondant à une absence de
culture (Soude, 2005).
2.4.2. Technique:
Préparation de l’inoculum:
Ensemencer 5 ml de bouillon BHIB avec 3 à 4 colonies de la souche à
tester;
Incuber 3 à 5 H sous agitation à 37± 1°C. La culture doit être de 108
UFC/ml soit de densité optique 0,08-0,1 à une longueur d’onde de 625nm.

Matériel et Méthodes
39
Ensemencement:
Diluer la suspension inoculum au 1/100 ( 106 UFC/ml) dans de l’eau
physiologique;
Ensemencer par inondation les boites de Pétri coulées par 20 ml de milieu
Muller-Hinton;
Après l'aspiration de surplus, faire sécher les boites de Pétri 20 à 30 mn à
l’étuve.
Application des disques:
Déposer les disques d’antibiotiques à l’aide d’une pince stérile;
Pour ce qui concerne les -lactamines, placer les disques dans un ordre qui
obéit au schéma de Vedel (Figure 5).
Figure 5. Disposition des antibiotiques sur les boîtes d’antibiogrammes selon
le schéma de Vedel (1998).
Marqueurs principaux Marqueurs secondaires
Amoxicilline: AMX
Ticarcilline: TIC
Céfalotine: CF
Céfotaxime: CTX
Amoxicilline+ Ac.clavulanique: AMC
Céfoxitine: FOX
Céfuroxime: CXM
PIP
MEC
CF
TIC
AMX
IMP
AMC
CTX
CXM
CAZ
MOX
ATM
FOX
AMC

Matériel et Méthodes
40
On laisse les boites 20 mn à la température ambiante pour permettre une pré -
diffusion de l’antibiotique, puis on incube pendant 18-24 H à 37°C ±1°C.
2.4.3. Lecture:
Mesurer avec précision les diamètres des zones d’inhibition, et comparer
ces résultats aux valeurs critiques figurant dans l’annexe ;
Classer les bactéries dans l’une des catégories: Sensib le, Intermédiaire ou
Résistance;
Interpréter les phénotypes de résistance aux -lactamines.
2.5. Détermination de la CMI en milieu solide (CASFM, 2010):
2.5.1. Principe :
La méthode de dilution successive en milieu solide est la méthode de référence pour
déterminer la sensibilité bactérienne aux antibiotiques. Elle consiste à mettre un
inoculum bactrien standardisé au contact de concentrations croissantes d’antibiotiques.
2.5.2. Technique:
Préparation des solutions d’antibiotiques:
Pour chacun d’antibiotiques, préparer une solution mère à 5120 mg/l puis réaliser des
dilutions séries de progression géométrique de raison 1/2 (Annexe 6).
Préparation des boîtes:
Distribuer 2 ml de chaque dilution d’antibiotique dans une série de boîte de Pétri
ainsi que 2 ml d’eau distillée stérile dans une boîte de Pétri témoin;
Ajouter 18 ml de milieu Mueller Hinton glosé maintenu en surfusion;
Bien mélanger et laisser solidifier;
Sécher les boîtes 30 mn à l’étuve à 37°C.

Matériel et Méthodes
41
Préparation de l’inoculum:
Préparer des suspensions de 108 UFC/ml pour chacune des souches à tester.
Ensemencement:
Diluer la suspension d’inoculum au 1/10;
Ensemencer par spot 1 à 2 l de la suspension bactérienne, soit un inoculum de
104 UFC/spot.
Incubation:
Incuber 18 à 24 H à 37°C.
2.5.3. Lecture:
S’assurer de la croissance des souches au niveau de la boîte témoin;
La concentration minimale inhibitrice est définie comme étant la plus faible
concentration ou il n’y a pas de croissance visible. La présence d’une ou de deux
colonies ou d’un fin film n’est pas prise en considération.
2.6. Détermination de la CMI pour les Proteus (CASFM, 2010; Schwalbe et al.,
2007):
En milieu gélose, Proteus mirabilis et Proteus vulgaris peuvent envahir la surface du
milieu en formant des ondes concentriques. Cet essaimage, ou « swarming », est dû à
la grande mobilité de la bactérie (figure 6). Cependant il est peu marqué pour Proteus
penneri (Amhis, 2004).

Matériel et Méthodes
42
Figure 6. Envahissement (swarming) de Proteus spp. sur gélose Mueller Hinton.
L'envahissement des cultures par les Proteus peut être réduit par:
L’augmentation de la concentration en gélose (>3%);
La diminution de la concentration en Nacl;
T° à 43°C;
Adjonction de diverses substances alcool éthylique (5p100), chloral (1 à
2p1000), azide de sodium (1p5000);
Les métaux lourds affecteraient la production de super oxyde dismutase
indispensable au métabolisme de la bactérie;
Les chélateurs ferreux (le fer est nécessaire à la croissance bactérienne);
Les agents osmotiques spécifiques;
Le salycilate de Sodium diminuerait le nombre de flagelles;
l’ajout de 50 mg/l de p-nitrophenyl glycérol (PNPG) ou 350 mg/l Matexil.
Cependant le PNPG, Matexil ou l’augmentation de concentration en gélose pouvons
tous altérer les résultats significatif des CMI avec certains agents (Amhis, 2004;
Andrews, 2006).
Pour cela, on procède à déterminer les concentrations minimales inhibitrices en milieu
solide avec la technique décrite précédemment mais en augmentant la température
d’incubation à 43°C au lieu de 37°C ± 1°C.
P.m P.v
P.p

Matériel et Méthodes
43
2.7. Test de synergie (Rahal, 2005):
2.7.1. Principe:
Cette technique, sur gélose dite des « bouchons de champagne » recommandée
par le CASFM, reste la plus utilisée. Elle consiste à rechercher une synergie
sur gélose de Mueller-Hinton entre un disque d’antibiotique contenant un
inhibiteur de -lactamases (d’amoxicilline + acide clavulanique) et des disques
(placés à 2 ou 3 cm du disque central) de C3G (ceftazidime, de céfotaxime et
de céfépime) et/ou d’aztréonam. Elle permet de différencier une bactérie
hyperproductrice de céphalosporinases d'une bactérie produisant une BLSE
dans la famille des entérobactéries.
2.7.2. Technique:
La recherche de la -lactamase à spectre étendu se fait dans les conditions standard
de l’antibiogramme en disposant un disque d’amoxicilline/acide clavulanique
AMC à 30 mm (centre à centre) d’un disque de C3G.
La détection des BLSE chez les souches également hyperproductrices de
céphalosporinase (Enterobacter….) est facilitée par la recherche d’une
synergie entre un disque d’amoxicilline/acide clavulanique AMC et le
céfépime ou cefpirome (C4G).
Chez Proteus mirabilis, P vulgaris, P pinneri, Morganella morganii,
Providencia stuartii, les BLSE s’expriment à bas niveau dans ce cas le test
de synergie est optimisé en disposant les disques à une distance de 40 à 45
mm au lieu de 30 mm.
2.7.3. Lecture:
Il y a présence d’une BLSE ou un test de synergie positif quand il y a une
image caractéristique, en « bouchon de Champagne ». Un résultat positif est
basé sur l'inhibition des BLSE par l 'acide clavulanique, et par conséquent
l'augmentation de l'activité des céphalosporines de troisième et quatrième
génération en présence d'acide clavulanique.

Matériel et Méthodes
44
2.8. Test à la cloxacilline (De Champs, 2002; Naas, 2003):
2.8.1. Principe:
Pour certaines souches de bacille à Gram négatif, il est parfois difficile de
distinguer sur l’antibiogramme habituel les hyperproductions de
céphalosporinases des -lactamases à spectre élargi (BLSE). En cas
d’hyperproduction, les diamètres autour des disques de C3G très diminués peuvent
masquer les images de synergies évocatrices de BLSE entre des disques de C3G et
d’acide clavulanique. La cloxacilline (Orbénine®), ajoutée au milieu pour
l’antibiogramme (Mueller-Hinton), inhibe in vitro les céphalosporinases de la
classe C d’Ambler chromosomiques ou plasmidiques et reste inefficace sur les
pénicillinases des bacilles à Gram négatif.
2.8.2. Technique:
Réaliser un antibiogramme par diffusion sur gélose Mueller Hinton contenant de la
cloxacilline à une concentration finale de 250 μg/ml;
Dans ce cas il faut dissoudre 0,25 g de Cloxacilline dans 25 ml d’eau
distillée pour préparer une concentration de 10000 μg/ml puis procéder à une
dilution à 2500 μg/ml;
Pour une boite ronde (90mm), mettre 2 ml de cette concentration et 18 ml
de Mueller-Hinton.
2.8.3. Lecture:
L’interprétation des résultats peut se traduire, en comparant les boites de Pétri
contenant une gélose Mueller-Hinton simple et celles contenant les géloses
supplémentées de la cloxacilline.
Un résultat positif est noté lorsque une augmentation des diamètres
d'inhibition, d'au moins 8 mm autour des disques de céfotaxime CTX et
céftazidime CAZ, est observé.

Matériel et Méthodes
45
2.9. Test de Hodge (Cuzon et al., 2009; Anderson et al., 2007):
2.9.1. Principe:
Un disque d’imipénème est appliqué au centre d’une boîte de Mueller Hinton (MH)
préalablement ensemencée avec une souche d’Escherichia coli sauvage (ATCC
25922) sensible aux carbapénèmes afin d’obtenir une culture confluente et un diamètre
dans la zone de sensibilité autour de l’imipénème.
2.9.2. Technique:
Préparer une suspension d’Escherichia coli ATCC 25922 de 108 UFC/ml;
Ensemencer par écouvillonnage une gélose Mac Conkey;
Faire une lourde strie de la souche à tester à partir d’un prélèvement de quelques
colonies d’une culture sur milieu Mueller Hinton gélosé du centre de la boite vers
la périphérique;
Sécher les biotes 15 mn;
Déposer un disque d’imipénème au centre (à l’extrémité de la strie);
Incuber 18 H à 37°C ± 1°C.
2.9.3. Lecture:
L’hydrolyse de l’imipénème par la souche à tester se traduit par l’échancrure de la
zone d’inhibition de la souche d’Escherichia coli ATCC 25922 (Figure 7).
Figure 7. Lecture du test de Hodge (Anderson et al., 2007).
IMP
Test positif
Test négatif
E. coli
Strie de la souche
à tester

Matériel et Méthodes
46
2.10. Test de l’EDTA (Aggoune-Khinache et al., 2009; Galani et al., 2008 ):
2.10.1. Principe:
La recherche de la métallo--lactamases a été effectuée sur les souches en mettant à
profit la faculté de l’ethylenediamintetraacetic acid (EDTA), chélateur bivalent à
restaurer l’activité de l’imipénème en chélatant les ions ZN2+
indispensables à
l’activité de la métallo--lactamases en la rendant inactive. Il permet de différencier
entre les métallo--lactamases et les autres carbapénèmases de classe A ou D.
2.10.2. Technique de DDST (Double-disc synergy tests):
Préparer des suspensions de la souche à tester a une densité équivalente à 0.5 Mc
Farland et inoculer une boite de Mueller Hinton;
Placer un disque d’imipénème (10 g) ou un disque de céftazidime (30 g) et un
disque blanc de papier filtre a distance de 10, 15 et 20 mm (centre à centre);
Ajouter 10 l de solution d’EDTA 0.5 M au disque blanc;
Incuber 18 H à 35°C ± 1°C.
2.10.3. Lecture:
Après l’incubation, la présence d’une zone d’inhibition synergétique est interprétée
positive.
2.10.4. Technique de CDT (The combination disc test):
Placer 2 disques d’imipénème (10 g) et 2 disques de céftazidime (30 g) sur une
boite de Mueller Hinton inoculée par la souche à tester (0.5 Mc Farland);
Ajouter 4 l de solution d’EDTA (0,5 M) a un disque de chaque antibiotique.
2.10.5. Lecture:
Après une incubation pendant 18 H à 35°C ± 1°C, une différence de diamètre des
zones entre les disques d’imipénème / imipénème + l’EDTA ou de céftazidime /
céftazidime + l’EDTA de plus 7 mm est interprétée comme résultat positive.

Matériel et Méthodes
47
2.11. Transfert des plasmides par conjugaison (Touati, 2006):
2.11.1. Principe:
La conjugaison représente le phénomène de contact physique par lequel les plasmides
peuvent passer d’une cellule donatrice à une autre réceptrice (K 12). La sélection des
transconjugants s’effectue en présence de 2 antibiotiques: l’un correspond à l’une des
résistances transférées, l’autre à la résistance non transférable de la souche réceptrice.
2.11.2. Technique:
Ensemencer la souche réceptrice la K 12 et les souches donatrices en bouillon
BHIB et incuber 18 à 24 H à 37°C ± 1°C;
Réaliser un mélange donatrice réceptrice dans un rapport 1 : 10;
Mélanger par inversion;
Incuber 18 à 24 H à 37°C ± 1°C;
Ensemencer par strie le mélange sur un milieu de sélection;
Vérifier la sélectivité des milieux de sélection en ensemençant la souche donatrice
et la souche réceptrice sur la même boite;
Incuber 24 H à 37°C ± 1°C.
2.11.3. Préparation de boites de sélections:
La sélection a été faite sur Mac Conkey (18 ml) additionné de 1 ml de céfotaxime et 1
ml de rifampicine à des concentrations finales de 16 g/ml et de 256 g/ml
respectivement.
2.11.4. Lecture:
Analyser les transconjugants en réalisant des antibiogrammes et/ou des CMI.

Matériel et Méthodes
48
2.12. Extraction de l’ADN plasmidique (Kado et Liu, 1981- modifiée):
2.12.1. Principe:
L’extraction a pour but de séparer l’ADN plasmidique de l’ADN de la cellule hôte. La
technique la plus répondue comprend une étape de lyse alcaline en présence de
détergent (SDS), au cours de laquelle les deux types d’acide nucléique sont dénaturés.
Le pH est ensuite neutralisé rapidement, ce qui aboutit à une forme soluble pour les
plasmides et à un agrégat insoluble pour l’ADN chromosomique, ce dernier incluant
également les protéines dénaturées et le SDS précipité. Après centrifugation, l’ADN
plasmidique contenu dans le surnageant est précipité par l’éthanol et le culot après
centrifugation repris par le TE ou de l’eau stérile.
2.12.2. Technique:
Réaliser une culture bactérienne dans 3 ml de LB et incuber 24 H à 37° C ± 1°C;
Centrifuger la culture à 14000 rpm pendant 2 mn à 4° C;
Eliminer le surnageant et sécher le culot;
Re-suspendre le culot dans 100 μl de TE;
Bien vortexer;
Ajouter 200 μl de solution de lyse;
Mélanger par inversion 5 à10 fois;
Incuber à 55° C pendant 1 H en agitant de temps à l’autre;
Ajouter 600 μl de phénol- chloroforme- isoamylalcool (25/24/1);
Mélanger par inversion 5 à 10 fois;
Centrifuger à 14000 rpm pendant 15 mn à 4° C;
Récupérer délicatement la phase aqueuse dans un nouveau microtube;
Ajouter 300 μl d’acétate de sodium 3 M pH 4,8;
Mélanger par inversion 5 à 10 fois;
Centrifuger à 14000 rpm pendant 10 mn à 4° C;
Récupérer le surnageant dans un nouveau microtube;
Ajouter 1 ml d’éthanol absolue à -20° C;

Matériel et Méthodes
49
Précipiter 30 mn à -20° C;
Centrifuger à 14000 rpm pendant 10 mn à 4° C;
Laver le culot avec 500 μl d’éthanol 70% à température ambiante;
Centrifuger à 14000 rpm pendant 5 mn à 4° C;
Eliminer le surnageant et bien sécher le culot;
Re-suspendre le culot dans 40 μl de TE.
2.13. Electrophorèse sur gel d’agarose (Sambrook et Russel, 2001):
2.13.1. Principe:
L'électrophorèse sur gel d'agarose est la méthode de choix pour la séparation, la
purification et l'identification des fragments d'ADN et d'ARN. Sous l’influence d’un
courant électrique, les acides nucléiques chargés négativement se déplacent à travers le
gel d’agarose vers la borne positive (l'anode). Les molécules seront séparées dans le
gel dû au fait qu’elles ne migrent pas toutes à la même vitesse.
2.13.2. Technique:
Préparation du gel d’agarose:
Pour un gel de 0,8%, dissoudre par chauffage jusqu’à ébullition, 1,6 g d’agarose
(Sigma) dans 200 ml de tampon TBE 1 X;
Refroidir la solution à 55°C dans un bain marie, puis couler le gel dans un moule
dont les 2 extrémités ont été préalablement fermées par du ruban adhésif;
Déposer un peigne dans le gel afin de réaliser des puits;
Laisser solidifier;
Retirer le peigne du gel et le ruban adhésif du moule;
Placer le moule avec le gel dans la cuve d’électrophorèse;
Ajouter un volume de tampon TBE 1X dans la cuve préalablement remplie jusqu’à
ce que le gel soit immergé (environ 1 mm au dessus).

Matériel et Méthodes
50
Ensemencement:
Répartir dans des tubes eppendorfs 40 l de chaque solution d’ADN;
Ajouter 10 l de tampon de charge;
Mélanger à l’aide d’une micropipette puis transférer les 25 l des mélanges dans
les puits du gel.
Migration:
Brancher le générateur après avoir raccordé la cuve;
Appliquer un voltage de 90 volts pendant 4 H.
Révélation:
Colorer le gel en l’immergeant dans une solution de TBE 1X contenant du BET à
une concentration de 0,5 g/ml pendant 30 à 45 mn à température ambiante;
Placer le gel dans un transilluminateur.

Résultats
Interprétation
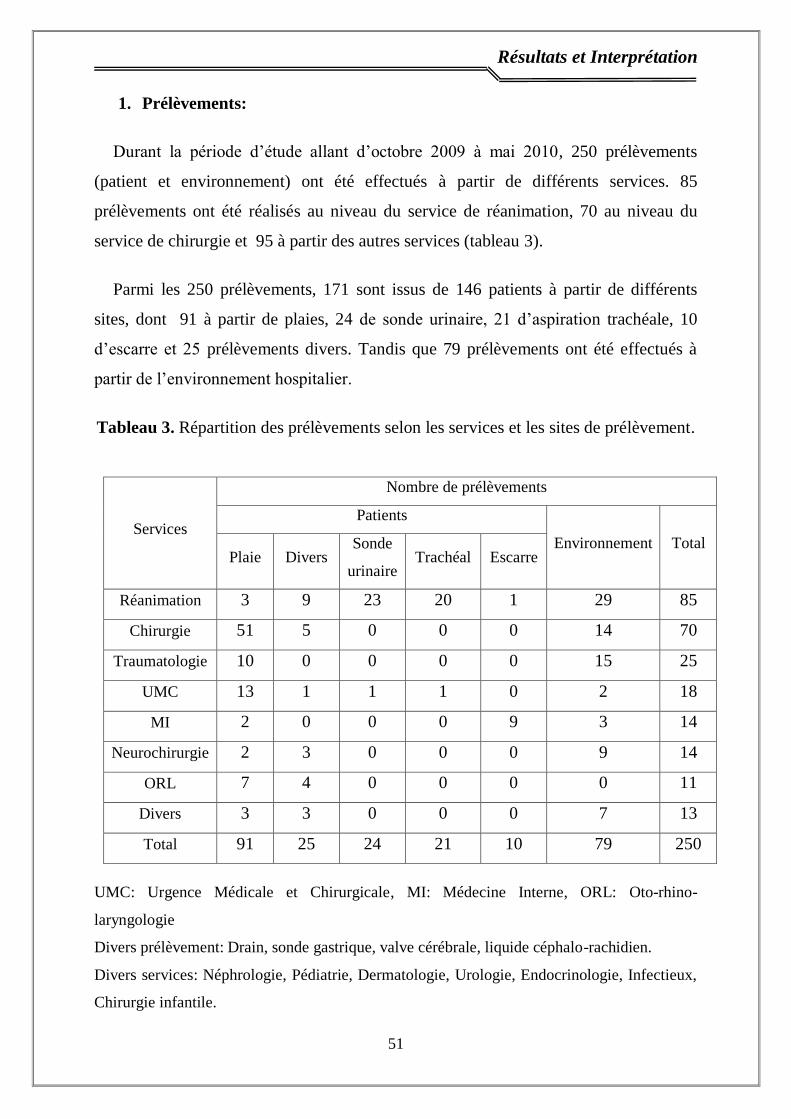
Résultats et Interprétation
51
1. Prélèvements:
Durant la période d’étude allant d’octobre 2009 à mai 2010, 250 prélèvements
(patient et environnement) ont été effectués à partir de différents services. 85
prélèvements ont été réalisés au niveau du service de réanimation, 70 au niveau du
service de chirurgie et 95 à partir des autres services (tableau 3).
Parmi les 250 prélèvements, 171 sont issus de 146 patients à partir de différents
sites, dont 91 à partir de plaies, 24 de sonde urinaire, 21 d’aspiration trachéale, 10
d’escarre et 25 prélèvements divers. Tandis que 79 prélèvements ont été effectués à
partir de l’environnement hospitalier.
Tableau 3. Répartition des prélèvements selon les services et les sites de prélèvement.
Services
Nombre de prélèvements
Patients
Environnement Total Plaie Divers
Sonde
urinaire Trachéal Escarre
Réanimation 3 9 23 20 1 29 85
Chirurgie 51 5 0 0 0 14 70
Traumatologie 10 0 0 0 0 15 25
UMC 13 1 1 1 0 2 18
MI 2 0 0 0 9 3 14
Neurochirurgie 2 3 0 0 0 9 14
ORL 7 4 0 0 0 0 11
Divers 3 3 0 0 0 7 13
Total 91 25 24 21 10 79 250
UMC: Urgence Médicale et Chirurgicale, MI: Médecine Interne, ORL: Oto-rhino-
laryngologie
Divers prélèvement: Drain, sonde gastrique, valve cérébrale, liquide céphalo-rachidien.
Divers services: Néphrologie, Pédiatrie, Dermatologie, Urologie, Endocrinologie, Infectieux,
Chirurgie infantile.

Résultats et Interprétation
52
2. Identification de souches:
140 entérobactéries ont été identifiées, parmi un total de 240 bacilles à Gram
négatif, avec une fréquence d’isolement de 56% (140/250). Ainsi 120 souches ont été
isolées à partir de patients avec 70.2% (120/171) et 20 souches à partir de
l’environnement soit 25.3% (20/79).
L’analyse des espèces identifiées par rapport à l’ensemble des souches
d’entérobactéries isolées, a montré une prédominance d’Escherichia coli avec 52
souches soit 37.1%, suivie de Klebsiella pneumoniae avec 30 souches (21.4%), de
Proteus spp. avec 27 souches (19.3%) et d’Enterobacter cloacae avec 26 souches
(18.6%). Les différentes espèces de Proteus sont représentées par Proteus mirabilis
avec 19 souches, Proteus vulgaris (7 souches) et Proteus pinneri (une souche).
Les autres entérobactéries viennent en dernière position avec 5 isolats soit 3.6% et
qui correspond à Citrobacter freundii, Morganella morganii, Providencia stuartii,
Shigella sonnei et Salmonella spp.
Figure 8. Répartition de l’ensemble des espèces d’entérobactéries isolées.
37,1%
21,4%
19,3%
18,6%
3,6%
Escherichia coli
Klebsiella pneumoniae
Proteus spp.
Enterobacter cloacae
Divers
Résultats et Interprétation
53
Figure 9. Identification des principales espèces par galerie API 20E et ID 32E.
Pro
teu
s m
irabil
is
Esc
heri
ch
ia c
oli
K
lebsi
ell
a p
neu
mon
iae
En
tero
bacte
r clo
acae

Résultats et Interprétation
54
2.1. Répartition des entérobactéries en fonction des services:
Au niveau du service de réanimation, 39 souches d’entérobactéries (27.8%) ont été
isolées, dont 41% proviennent de prélèvements trachéaux et de sondes urinaires. 37
souches (26.4%) ont été collectées au niveau du service de chirurgie et 64 souches
(45.7%) ont été isolées à partir de divers services, essentiellement de plaies
chirurgicales.
L’étude de la distribution des souches d’entérobactéries identifiées est variable
selon les services. En effet, au niveau de la réanimation, Escherichia coli et Klebsiella
pneumoniae occupent la première place avec 33.3%, suivie d’Enterobacter cloacae
(20.5%) et de Proteus spp. (10.3%).
Figure 10. Répartition des entérobactéries au niveau du service de réanimation.
33,3%
33,3%
10,3%
20,5%
2,6%
Escherichia coli
Klebsiella pneumoniae
Proteus spp.
Enterobacter cloacae
Divers

Résultats et Interprétation
55
Dans le service de chirurgie, Escherichia coli est l’espèce majoritaire (32.4%)
suivie de Proteus spp. avec un taux de 29.7%, celui-ci étant plus élevé que celui
obtenu au niveau des autres services, suivie enfin de Klebsiella pneumoniae (21.6%) et
d’Enterobacter cloacae (13.5%).
Figure 11. Répartition des entérobactéries au niveau du service de chirurgie.
Pour le reste des services, Escherichia coli est encore l’espèce dominante avec un
taux de 42.2%, suivie cette fois d’Enterobacter cloacae avec 20.3%, de Proteus spp.
(18.7%) et de Klebsiella pneumoniae (14%).
Figure 12. Répartition des entérobactéries au niveau des autres services.
42,2%
14%
18,7%
20,3%
4,7%
Escherichia coli
Klebsiella pneumoniae
Proteus spp.
Enterobacter cloacae
Divers
32,4%
21,6%
29,7%
13,5%2,7%
Escherichia coli
Klebsiella pneumoniae
Proteus spp.
Enterobacter cloacae
Divers

Résultats et Interprétation
56
2.2. Répartition des entérobactéries en fonction des sites de prélèvement:
La distribution des espèces isolées en fonction des sites de prélèvement révèle
qu’Escherichia coli occupe la 1ere
place parmi les entérobactéries pour les
prélèvements de sondes urinaires, d’escarres et de plaies chirurgicales avec
respectivement des taux de 61.1% (11/18), 54.5% (6/11) et 34.8% (23/66).
À noter que les souches de Proteus spp. prédominent dans les prélèvements de plaie
chirurgicale par rapport aux autres sites, soit 70.4% (19/27).
Figure 13. Répartition des entérobactéries au niveau des sondes urinaires.
Figure 14. Répartition des entérobactéries au niveau des plaies chirurgicales.
34,8%
19,7%
28,8%
13,6%3%
Escherichia coli
Klebsiella pneumoniae
Proteus spp.
Enterobacter cloacae
Divers
61,1%16,7%
11,1%
11,1% 0%
Escherichia coli
Klebsiella pneumoniae
Proteus spp.
Enterobacter cloacae
Divers

Résultats et Interprétation
57
A l’inverse, les sondes trachéales sont beaucoup plus colonisées par les souches de
Klebsiella pneumoniae avec un taux d’isolement de 50 % (8/16).
Figure 15. Répartition des entérobactéries au niveau des prélèvements trachéaux.
Enfin, pour l’environnement de l’hôpital de Sidi Bel Abbes, c’est Enterobacter
cloacae qui prédomine avec un taux de 45% (9/20).
Figure 16. Répartition des entérobactéries au niveau des prélèvements de
l’environnement.
12,5%
50%6,2%
31,2%
0%
Escherichia coli
Klebsiella pneumoniae
Proteus spp.
Enterobacter cloacae
Divers
30%
10%
10%
45%
5%
Escherichia coli
Klebsiella pneumoniae
Proteus spp.
Enterobacter cloacae
Divers
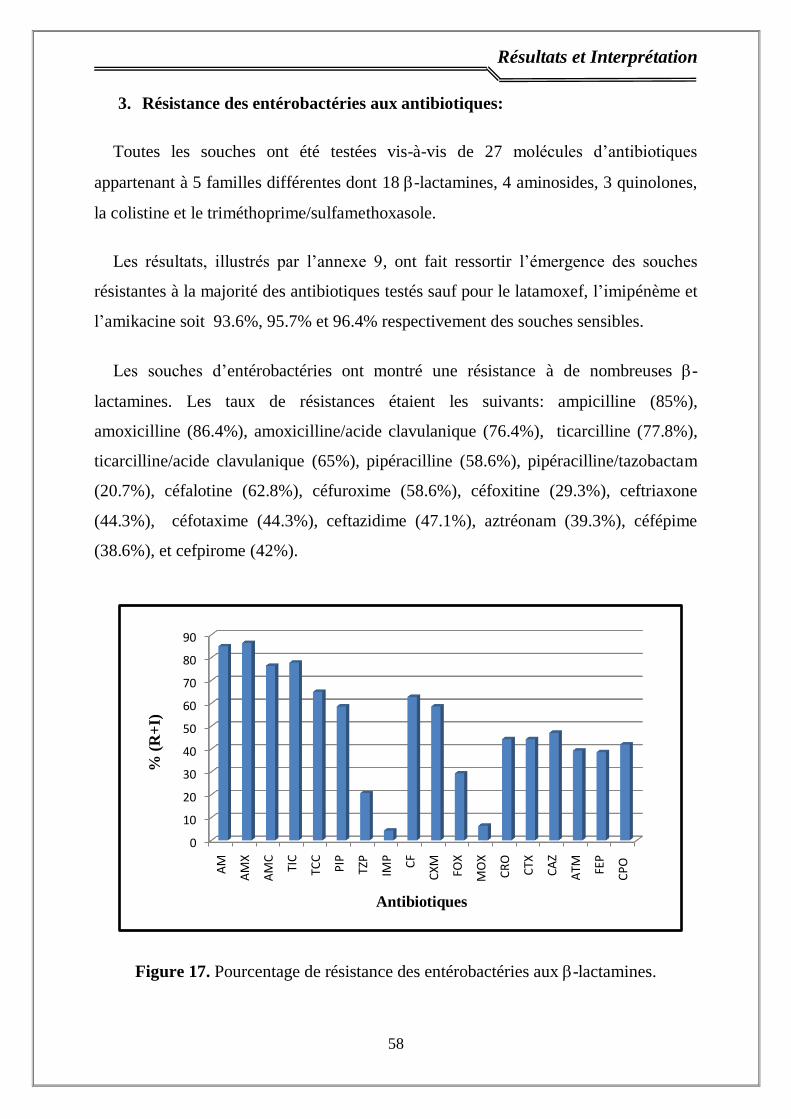
Résultats et Interprétation
58
3. Résistance des entérobactéries aux antibiotiques:
Toutes les souches ont été testées vis-à-vis de 27 molécules d’antibiotiques
appartenant à 5 familles différentes dont 18 -lactamines, 4 aminosides, 3 quinolones,
la colistine et le triméthoprime/sulfamethoxasole.
Les résultats, illustrés par l’annexe 9, ont fait ressortir l’émergence des souches
résistantes à la majorité des antibiotiques testés sauf pour le latamoxef, l’imipénème et
l’amikacine soit 93.6%, 95.7% et 96.4% respectivement des souches sensibles.
Les souches d’entérobactéries ont montré une résistance à de nombreuses -
lactamines. Les taux de résistances étaient les suivants: ampicilline (85%),
amoxicilline (86.4%), amoxicilline/acide clavulanique (76.4%), ticarcilline (77.8%),
ticarcilline/acide clavulanique (65%), pipéracilline (58.6%), pipéracilline/tazobactam
(20.7%), céfalotine (62.8%), céfuroxime (58.6%), céfoxitine (29.3%), ceftriaxone
(44.3%), céfotaxime (44.3%), ceftazidime (47.1%), aztréonam (39.3%), céfépime
(38.6%), et cefpirome (42%).
Figure 17. Pourcentage de résistance des entérobactéries aux -lactamines.
0
10
20
30
40
50
60
70
80
90
AM
AM
X
AM
C
TIC
TCC
PIP
TZP
IMP CF
CX
M
FOX
MO
X
CR
O
CTX
CA
Z
ATM FE
P
CP
O
% (
R+
I)
Antibiotiques

Résultats et Interprétation
59
Pour les aminosides, le taux le plus important a été observé pour la tobramycine
(42.1%) suivi de la gentamicine (38.6%) et de la kanamycine (35%).
En ce qui concerne les fluoroquinolones (quinolone de 2eme
génération), les taux de
résistance étaient de 41.4 % pour l’ofloxacine et 33.6% pour la ciprofloxacine. Par
contre l’acide nalidixique (quinolone de 1ere
génération) est toutefois moins efficace
avec 46.4% de souches résistantes.
Une activité importante a été remarquée vis-à-vis de la colistine, soit 24.3%, en
tenant compte que cette molécule est totalement inactive sur les espèces de Proteus
(Cattoir, 2006). Alors qu’une activité faible a été observée vis-à-vis du
triméthoprime/sulfaméthoxazole avec un taux de 60%.
Figure 18. Pourcentage de résistance des entérobactéries aux autres classes
d’antibiotique.
Au sein des services, l’analyse globale des résultats d’antibiogramme a montré une
différence de taux de résistance. En effet pour la majorité des antibiotiques, les taux de
résistance sont plus importants au niveau de la réanimation par rapport aux autres
services. Cependant, on note que les taux de résistance pour la céfuroxime et la
colistine sont plus élevés au niveau de la chirurgie, cela est en rapport avec le nombre
de souches Proteus spp. isolés au niveau de ce service. Ainsi pour la céfoxitine et
0
10
20
30
40
50
60
TM GM AN K NA OFX CIP CS SXT
% (
R+
I)
Antibiotiques

Résultats et Interprétation
60
l’amikacine où on trouve une augmentation des taux de résistance au niveau des autres
services par rapport à la réanimation et à la chirurgie (figure 19 et 20).
Figure 19. Pourcentage de résistance des entérobactéries aux -lactamines au niveau
de la réanimation, de la chirurgie et des autres services.
Figure 20. Pourcentage de résistance des entérobactéries aux autres antibiotiques au
niveau de la réanimation, de la chirurgie et des autres services.
0
10
20
30
40
50
60
70
80
90
100
AM
AM
X
AM
C
TIC
TCC
PIP
TZP
IMP CF
CX
M
FOX
MO
X
CR
O
CTX
CA
Z
ATM FE
P
CP
O
%(R
+I)
Antibiotiques
Réanimation
Chirurgie
Autres
0
10
20
30
40
50
60
70
TM GM AN K NA OFX CIP CS SXT
% (
R+
I)
Antibiotiques
Réanimation
Chirurgie
Autres
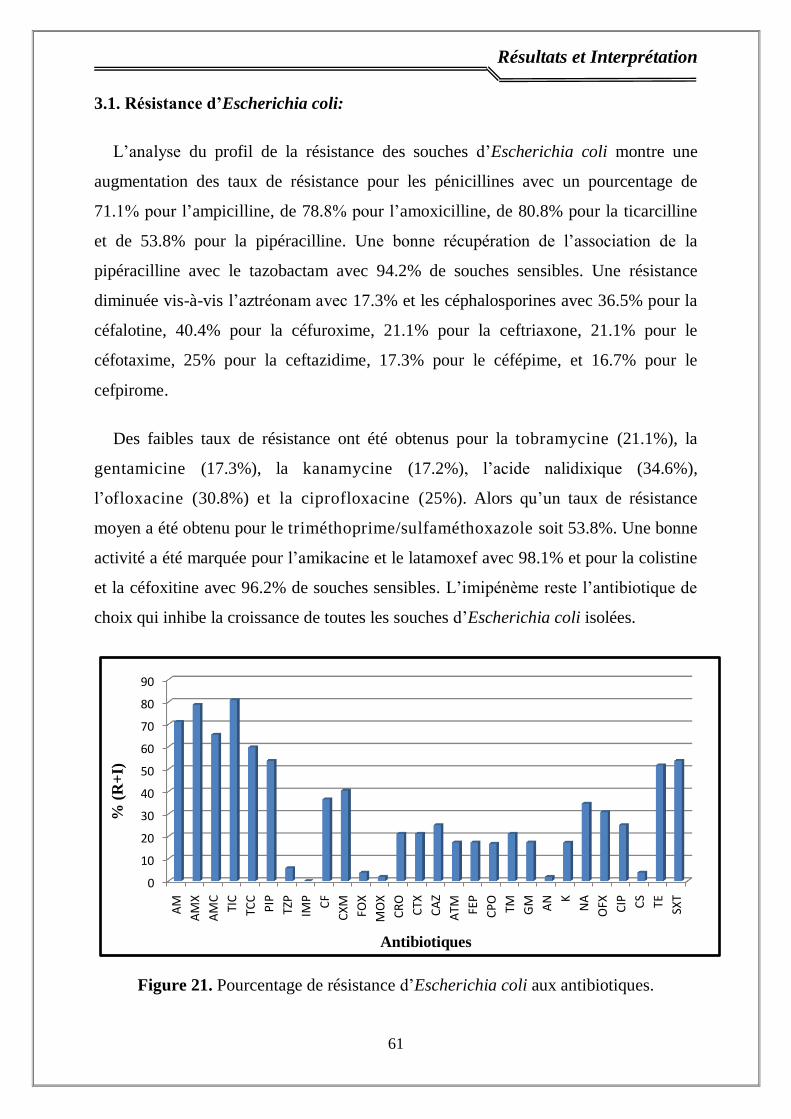
Résultats et Interprétation
61
3.1. Résistance d’Escherichia coli:
L’analyse du profil de la résistance des souches d’Escherichia coli montre une
augmentation des taux de résistance pour les pénicillines avec un pourcentage de
71.1% pour l’ampicilline, de 78.8% pour l’amoxicilline, de 80.8% pour la ticarcilline
et de 53.8% pour la pipéracilline. Une bonne récupération de l’association de la
pipéracilline avec le tazobactam avec 94.2% de souches sensibles. Une résistance
diminuée vis-à-vis l’aztréonam avec 17.3% et les céphalosporines avec 36.5% pour la
céfalotine, 40.4% pour la céfuroxime, 21.1% pour la ceftriaxone, 21.1% pour le
céfotaxime, 25% pour la ceftazidime, 17.3% pour le céfépime, et 16.7% pour le
cefpirome.
Des faibles taux de résistance ont été obtenus pour la tobramycine (21.1%), la
gentamicine (17.3%), la kanamycine (17.2%), l’acide nalidixique (34.6%),
l’ofloxacine (30.8%) et la ciprofloxacine (25%). Alors qu’un taux de résistance
moyen a été obtenu pour le triméthoprime/sulfaméthoxazole soit 53.8%. Une bonne
activité a été marquée pour l’amikacine et le latamoxef avec 98.1% et pour la colistine
et la céfoxitine avec 96.2% de souches sensibles. L’imipénème reste l’antibiotique de
choix qui inhibe la croissance de toutes les souches d’Escherichia coli isolées.
Figure 21. Pourcentage de résistance d’Escherichia coli aux antibiotiques.
0
10
20
30
40
50
60
70
80
90
AM
AM
X
AM
C
TIC
TCC
PIP
TZP
IMP CF
CX
M
FOX
MO
X
CR
O
CTX
CA
Z
ATM FE
P
CP
O
TM GM AN K
NA
OFX CIP CS
TE SXT
% (
R+
I)
Antibiotiques

Résultats et Interprétation
62
3.2. Résistance de Klebsiella pneumoniae:
L’étude de la résistance des souches de Klebsiella pneumoniae révèle des taux de
résistance très élevés pour l’ampicilline (100%), l’amoxicilline (100%),
l’amoxicilline/acide clavulanique (90%), la ticarcilline (100%), la ticarcilline/acide
clavulanique (93.3%), la pipéracilline (90%), la pipéracilline/tazobactam (46.7%), la
céfalotine (83.3%), la céfuroxime (83.3%), la ceftriaxone (86.7%), le céfotaxime
(80%), la ceftazidime (80%), l’aztréonam (73.3%), le céfépime (80%) et le cefpirome
(80%).
Pour les aminosides, le taux le plus important a été observé pour la gentamicine
(86.7%), suivi de la tobramycine (83.3%) et de la kanamycine (65%). En ce qui
concerne les quinolones, les taux de résistance étaient les suivants: 66.7% pour l’acide
nalidixique et 63.3% pour L’ofloxacine et la ciprofloxacine. Notons également un
taux de résistance important pour le triméthoprime/sulfaméthoxazole avec 83.3%.
En revanche une bonne activité du latamoxef, du céfoxitine et de l’amikacine a été
remarquée avec respectivement 96.7%, 90% et 86.7% de souches sensibles. Aucune
résistance n’a été observée pour l’imipénème et la colistine avec 100% de souches
sensibles.
3.3. Résistance de Proteus spp.:
Selon les figures 22 et 23, chez Proteus spp., on rapporte généralement une
diminution des taux de résistance aux antibiotiques par rapport aux souches de
Klebsiella pneumoniae: ampicilline (77.8%), amoxicilline (70.4%), amoxicilline/acide
clavulanique (55.5%), ticarcilline (63%), ticarcilline/acide clavulanique (37%),
pipéracilline (37%), pipéracilline/tazobactam (7.4%), céfalotine (51.8%), céfuroxime
(63%), céfoxitine (29.6%), ceftriaxone (25.9%), céfotaxime (33.3%), ceftazidime
(37%), aztréonam (22.2%), céfépime (18.5%), cefpirome (18.5%), tobramycine
(22.2%), gentamicine (18.5%), kanamycine (33.3%), acide nalidixique (33.3%),
ofloxacine (29.6%), ciprofloxacine (22.2%) et triméthoprime/sulfaméthoxazole
(51.8%).

Résultats et Interprétation
63
L'amikacine et le latamoxef présentent une bonne activité sur les souches de
Proteus avec une sensibilité totale, contrairement à la colistine qui reste inactive avec
une résistance totale (100%) due à une résistance naturelle des souches à cet
antibiotique.
Notons également une légère augmentation du taux de résistance à l imipénème
jusqu’à 11.1%.
3.4. Résistance d’Enterobacter cloacae:
Les souches identifiée d’Enterobacter cloacae présentent une résistance importante
vis-à-vis de l’ampicilline (100%), l’amoxicilline (100%), l’amoxicilline/acide
clavulanique (100%), la ticarcilline (65.4%), la ticarcilline/acide clavulanique (73%),
la pipéracilline (57.7%), la céfalotine (100%), la céfuroxime (69.2%), la céfoxitine
(100%), la ceftriaxone (65.4%), le céfotaxime (65.4%), la ceftazidime (65.4%),
l’aztréonam (65.4%), le céfépime (57.7%), le cefpirome (50%), la tobramycine
(57.7%), la gentamicine (53.8%), l’acide nalidixique (61.5%), l’ofloxacine (46.1%)
et le triméthoprime/sulfaméthoxazole (57.7%).
Les taux de résistance les plus faibles pour cette espèce sont observés avec 37.5%
pour la kanamycine, 34.6% pour la pipéracilline/tazobactam et la ciprofloxacine,
26.9% pour le latamoxef et 11.5% pour la colistine et l’imipénème. Pour l imipénème,
on note aussi une légère augmentation du taux de résistance semblable a celle observée
pour les souches de Proteus
L’amikacine reste l’antibiotique le plus actif sur les souches d’Enterobacter cloacae
avec une sensibilité totale.
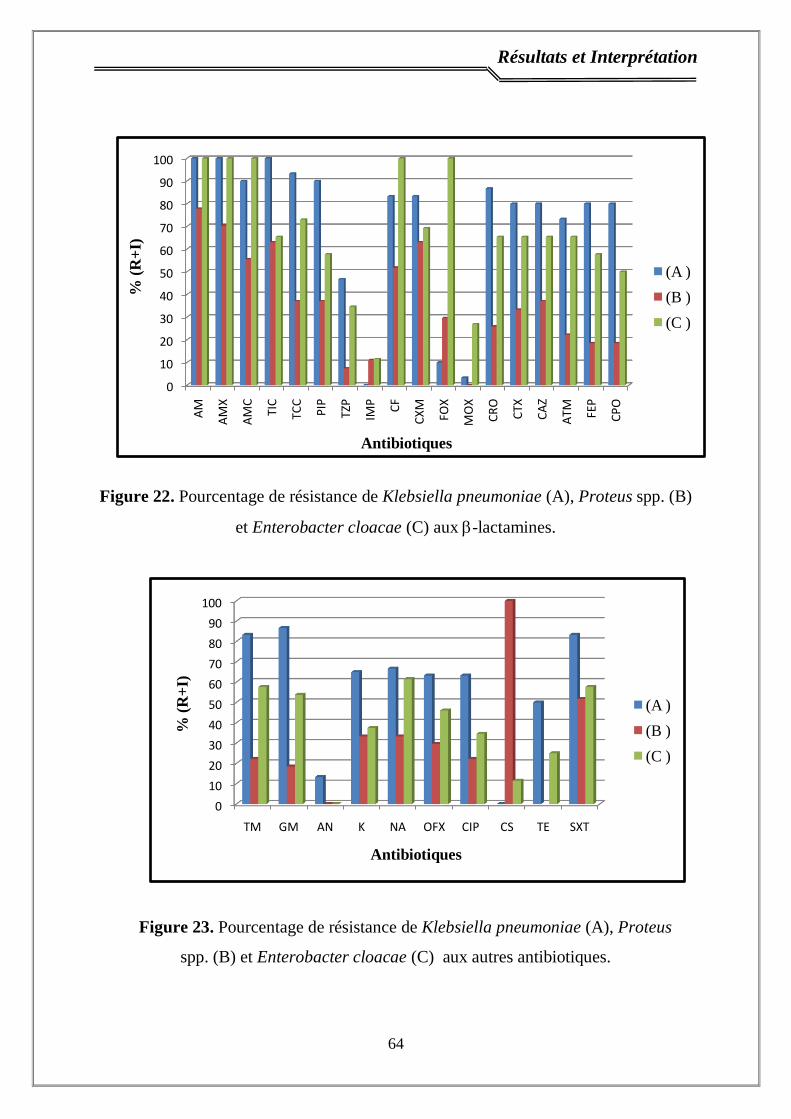
Résultats et Interprétation
64
Figure 22. Pourcentage de résistance de Klebsiella pneumoniae (A), Proteus spp. (B)
et Enterobacter cloacae (C) aux -lactamines.
Figure 23. Pourcentage de résistance de Klebsiella pneumoniae (A), Proteus
spp. (B) et Enterobacter cloacae (C) aux autres antibiotiques.
0
10
20
30
40
50
60
70
80
90
100
AM
AM
X
AM
C
TIC
TCC
PIP
TZP
IMP CF
CX
M
FOX
MO
X
CR
O
CTX
CA
Z
ATM FE
P
CP
O
% (
R+
I)
Antibiotiques
(A )
(B )
(C )
0
10
20
30
40
50
60
70
80
90
100
TM GM AN K NA OFX CIP CS TE SXT
% (
R+
I)
Antibiotiques
(A )
(B )
(C )

Résultats et Interprétation
65
4. Phénotypes de résistance:
Le comportement des entérobactéries vis-à-vis des -lactamines nous a permis de
classer, selon Vedel (1998), les souches en fonction de leur phénotype de résistance.
L’analyse phénotypique des espèces d’entérobactéries est en faveur d’une
production de -lactamase à spectre étendu (BLSE) soit (37.1%), dont 24 souches de
Klebsiella pneumoniae, 15 souches d’Enterobacter cloacae, 8 souches d’Escherichia
coli et 5 souches de Proteus spp. Parmi ces souches, 20 souches (14.3%), dont 9
Klebsiella pneumoniae, 5 Escherichia coli et Enterobacter cloacae et 1 Proteus spp.,
produisent également une céphalosporinase associée.
Le phénotype céphalosporinase hyperproduite seul est manifesté chez 6 souches soit
4.3%. La production de pénicillinase a été trouvée chez 35 souches avec 25%,
majoritairement observée chez Escherichia coli avec 23 souches. Le reste des souches
(33.6%) dont 20 Escherichia coli, 13 Proteus spp., 9 Enterobacter cloacae et 3
Klebsiella pneumoniae ont présenté le phénotype sauvage.
Figure 24. Phénotypes de résistance des entérobactéries aux -lactamines.
22,8%
14,3%
4,3%
25%
33,6%
BLSE
BLSE+CASE
CASE
PASE
Souches sauvages
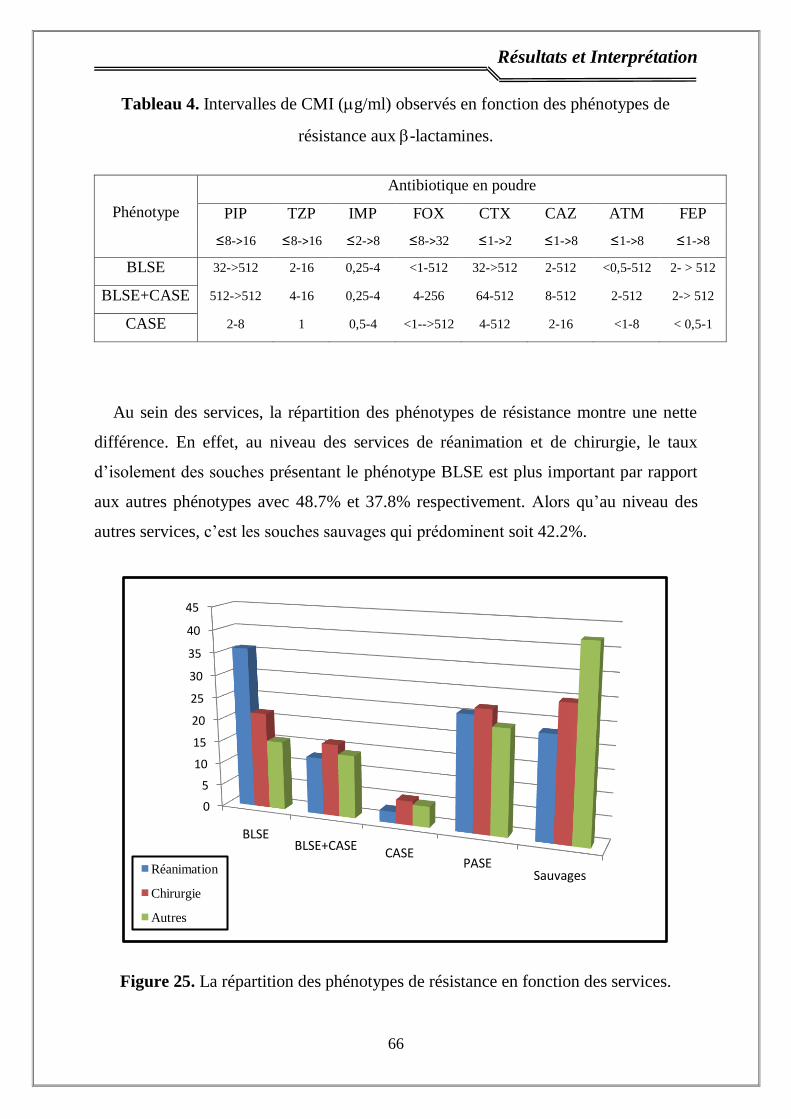
Résultats et Interprétation
66
Tableau 4. Intervalles de CMI (g/ml) observés en fonction des phénotypes de
résistance aux -lactamines.
Phénotype
Antibiotique en poudre
PIP TZP IMP FOX CTX CAZ ATM FEP
≤8->16 ≤8->16 ≤2->8 ≤8->32 ≤1->2 ≤1->8 ≤1->8 ≤1->8
BLSE 32->512 2-16 0,25-4 <1-512 32->512 2-512 <0,5-512 2- > 512
BLSE+CASE 512->512 4-16 0,25-4 4-256 64-512 8-512 2-512 2-> 512
CASE 2-8 1 0,5-4 <1-->512 4-512 2-16 <1-8 < 0,5-1
Au sein des services, la répartition des phénotypes de résistance montre une nette
différence. En effet, au niveau des services de réanimation et de chirurgie, le taux
d’isolement des souches présentant le phénotype BLSE est plus important par rapport
aux autres phénotypes avec 48.7% et 37.8% respectivement. Alors qu’au niveau des
autres services, c’est les souches sauvages qui prédominent soit 42.2%.
Figure 25. La répartition des phénotypes de résistance en fonction des services.
0
5
10
15
20
25
30
35
40
45
BLSEBLSE+CASE
CASEPASE
SauvagesRéanimation
Chirurgie
Autres

Résultats et Interprétation
67
La présence de phénotype BLSE a été confirmée par des images de synergie
caractéristiques entre l’acide clavulanique et les céphalosporines de 3eme
génération
et/ou l’aztréonam (figure 26).
Figure 26. Phénotype BLSE.
Dans certains cas, la forte résistance aux -lactamines, due à l’hyperproduction de
céphalosporinase, masque les images de synergie indicatrices de phénotype BLSE. Un
antibiogramme en présence d’une concentration définie de cloxacilline est nécessaire
pour vérifier l’association des 2 mécanismes (BLSE avec céphalosporinase) (figure
27).
Figure 27. Phénotype BLSE+CASE.
A: antibiogramme sans cloxacilline; B: antibiogramme avec cloxacilline à 250g/ml
Synergie
A B

Résultats et Interprétation
68
Par contre, l’absence des images de synergie et la restauration de l’activité des
céphalosporines de 3ème
génération, en présence de cloxacilline, est en faveur d’une
hyperproduction de céphalosporinase seule (figure 28)
Figure 28. Phénotype CASE.
A: antibiogramme sans cloxacilline; B: antibiogramme avec cloxacilline à 250g/ml
Pour les souches présentant le phénotype pénicillinase, l’acide clavulanique ne
restaure que partiellement l’activité de l’amoxicilline et de la ticarcilline. Alors que
l’association du tazobactam avec la pipéracilline permet une bonne récupération de
l’activité de l’antibiotique soit 88.6% de bactéries sensibles (figure 29).
Figure 29. Phénotype PASE.
Pipéracilline
Synergie
Ticarcilline
Pipéracilline/tazobactam
Ticarcilline/ac.clavulanique
A B

Résultats et Interprétation
69
La majorité des entérobactéries étaient sensibles à l’imipénème, indiquant sa place
en premier choix dans le traitement des infections sévères à bactéries multirésistantes.
Quoique, dans cette étude, certaines souches (6 souches), dont 3 Enterobacter cloacae
et 3 Proteus spp. ont présenté une résistance intermédiaire avec des valeurs de CMI de
4 g/ml vis-à-vis de l’imipénème.
Pour cela des tests de Hodge et de l’EDTA ont été réalisés afin de détecter
d’éventuelle production de carbapénèmases. Pour le test de Hodge cette méthode, bien
que facile à réaliser, ne préjuge en rien de l’identification de la -lactamase en cause.
En outre, le test est parfois difficile à interpréter (Cuzon et al., 2009). Alors que pour
le test de l’EDTA, aucune image de synergie n’a été détectée pour les souches testées.
Figure 30. Test de Hodge.
Figure 31. Test de l’EDTA pour la VIM (A) et la souche S.Ec.14 (B).
VIM
S.Ec.14 S.Ec.26
IMP IMP
+
EDTA
IMP EDTA
A B
Résultats et Interprétation
70
5. Résistances associées des entérobactéries:
Pour les souches produisant une BLSE ou une BLSE+CASE, en dehors de
l’amikacine la molécule la plus active, les autres aminosides présentent un fort risque
d’échec thérapeutique. L’association triméthoprime/sulfaméthoxazole et les
quinolones présentent également un taux de résistance considérable.
En revanche pour les souches qui produisent une céphalosporinase ou une
pénicillinase, les aminosides sont modérément affectés alors que les quinolones
présentent une efficacité limitée avec prés d’une souche résistante sur 2. L’amikacine
reste la molécule de référence avec une sensibilité totale.
Tableau 5. Résistances associées des entérobactéries en fonction des phénotypes de
résistance aux -lactamines.
Phénotype Nombre
(%)
Nombre de souches résistantes (%)
TM GM AN K NA OFX CIP CS SXT
BLSE 32
(22,8)
30
(93,7)
31
(96,8)
2
(6,2)
15
(65,2)
23
(71,8)
21
(65,6)
17
(53,1)
6
(18,7)
30
(93,7)
BLSE+CASE 20
(14,3)
19
(95)
18
(90) 3 (15)
7
(63,6)
13
(65)
12
(60)
12
(60) 2 (10)
18
(90)
CASE 6 (4,3) 1
(16,7)
1
(16,7) 0 (0) 0 (0) 3 (50) 3 (50) 3 (50) 3 (50)
2
(33,3)
PASE 35 (25) 6
(17,1)
4
(11,4) 0 (0)
11
(42,3)
19
(54,3)
17
(48,6)
11
(31,4)
9
(25,7)
29
(82,8)
Sauvage 47
(33,6) 3 (6,4) 0 (0) 0 (0) 0 (0)
7
(14,9)
5
(10,6)
4
(8,5)
14
(29,8)
5
(10,6)
Tableau 6. Intervalles de CMI (g/ml) observés en fonction des phénotypes de
résistance.
Phénotype
Antibiotique en poudre
TM GN AN CIP RIF
≤2- >4 ≤2- >4 ≤8- >16 ≤0,5- >1 ≤4- >16
BLSE 2- > 512 2- > 512 <0,5- > 512 <0,125- 64 2- 512
BLSE+CASE 2- >512 <0,5- > 512 <0,5- > 512 <0,125->128 2- 512
CASE <0,25- 2 <0,5- 4 <0,5- 8 <0,125- 2 4- 32

Résultats et Interprétation
71
6. Conjugaison bactérienne:
A fin de déterminer le support génétique de la résistance aux antibiotiques, des
expériences de conjugaison ont été réalisées entre la souche de référence Escherichia
coli résistante à la rifampicine (K12 Rif R) et 29 souches d’entérobactéries dont 13
BLSE+CASE, 10 BLSE et 6 CASE. 18 souches sur les 29 souches soumises au test
de conjugaison ont transféré une partie de leurs caractères de résistance à la souche
réceptrice, soit 62.1%. Les transconjugants ont été sélectionnés sur gélose Mac
Conkey contenant du céfotaxime et de la rifampicine à des concentrations finales de
16 g/ml et de 256 g/ml respectivement.
L’analyse des profils d’antibiorésistance des transconjugants par la méthode de
diffusion en milieu solide a montré le transfert de la résistance aux -lactamines, aux
aminosides et au triméthoprime/sulfaméthoxazole pour la majorité des souches.
Par contre pour les quinolones, le transfert de la résistance est constaté chez seulement
quelques souches.
Pour toutes les souches présentant le phénotype BLSE, des images de synergie,
indicatrice de ce phénotype, ont été observées chez leurs transconjugants qui
confirment le support plasmidique de cette résistance (figure 32).
Figure 32. Transfert de phénotype BLSE: (A) la souche réceptrice K12 Rif R, (B) la
souche donatrice et (C) son transconjugant.
A B C

Résultats et Interprétation
72
Parmi les souches ayant présenté le phénotype BLSE+CASE, certaines ont transféré
aussi bien le phénotype BLSE que le phénotype céphalosporinase. Le transfert de
gènes AmpC aux plasmides a donné lieu à leur diffusion parmi les entérobactéries, et
par conséquence que les -lactamases codés par les AmpC sont maintenant présentes
dans les souches de Klebsiella spp., Escherichia coli, Proteus mirabilis et Salmonella
spp., dont la sensibilité réduite à la céfoxitine peut être un indicateur de l’activité de
l’AmpC (Tan et al., 2009). La nature plasmidique transférée chez les transconjugants
a été confirmée par une augmentation de la concentration minimale inhibitrice de la
céfoxitine jusqu’au 32g/ml (figure 33).
Figure 33. Transfert de phénotype BLSE+CASE: (A) la souche réceptrice K12 Rif R,
(B) la souche donatrice et (C) son transconjugant.
Par contre certaines souches présentant le phénotype BLSE+CASE, seul le
phénotype BLSE a été transféré (figure 34), le plus souvent, ce cas caractérise les
espèces qui produisent une -lactamases de type AmpC chromosomique (Choi et al.,
2008).
A C B
Fox

Résultats et Interprétation
73
Figure 34. Transfert de phénotype BLSE chez les souches présentant le phénotype
BLSE+CASE: (A) la souche réceptrice K12 Rif R, (B) la souche donatrice et (C) son
transconjugant.
Ainsi aucun transfert n’a été observé chez les souches S.E.15, S.Pm.9, S.Pm.12,
S.Pv.4, S.Ec.4 et S.Ec.11 qui présentent le phénotype céphalosporinase seul.
Tableau 7. Intervalles de CMI (g/ml) observés en fonction des phénotypes de
résistance aux -lactamines des transconjugants.
Phénotype
Antibiotique en poudre
FOX CTX CAZ ATM FEP TM GN CIP RIF
≤8->32 ≤1->2 ≤1->8 ≤1->8 ≤1->8 ≤2- >4 ≤2- >4 ≤0,5- >1 ≤4->16
BLSE 2-4 16- >512 0.5-512 16-256 4-16 2-256 4->512 <0,125-2 >512
BLSE+CASE 2-32 16-512 0.5-32 8-16 1-8 <0,5-16 <0,5-32 <0,125-2 >512
B
A
C A

Résultats et Interprétation
74
7. Extraction et visualisation des plasmides:
L’analyse du contenu plasmidique de 8 souches d’entérobactéries (2 souches de
chaque espèce) ainsi que de leurs transconjugants respectifs a permis de visualiser la
taille et le nombre de plasmides hébergés par ces souches. La taille des plasmides a été
estimée par comparaison avec des plasmides de taille connue de la souche Escherichia
coli V 517. La plupart des souches contiennent 1 à 2 plasmides de taille supérieure à
55 Kb. Les 2 souches de Klebsiella pneumoniae comportent 3 et 5 plasmides et les 2
souches d’Enterobacter cloacae comportent 3 et 4 plasmides de taille inférieure.
Figure 35. Profils plasmidiques des 4 espèces d’entérobactéries et de leurs
transconjugants.
(1: V.517); (2: S.E.2 ); (3: Tc.E.2 ); (4: S.E.47); (5: Tc.E.47); (6: S.Kp.18); (7: Tc.Kp.18);
(8: S.Kp.21); (9: Tc.Kp.21); (10: S.Ec.12); (11: Tc.Ec.12); (12: S.Ec.14 ); (13: Tc.Ec.14);
(14: S.Pm.2); (15: Tc.Pm.2); (16: S.Pm.3); (17: Tc.Pm.3 ); (18: V.517).
Enterobacter cloacae Escherichia coli
Klebsiella pneumoniae Proteus spp.
55 kb
7,56 Kb
5,8 Kb
5,35 Kb
4,1 Kb
3,15 Kb
2,8 Kb
2,2 Kb
1 2 3 4 5 6 7 8 9 10 11 12 13 14 15 16 17 18

Discussion

Discussion
75
L’hôpital où a eu lieu notre étude est le centre hospitalo-universitaire Hassani
Abdelkader de Sidi Bel Abbes, contenant 639 lits, accueillant des enfants et des
adultes, avec 23 services d’hospitalisation et 7 services d’exploration. Cette étude a
permis d’évaluer l’activité antibactérienne des antibiotiques sur les entérobactéries.
Entre octobre 2009 et mai 2010, 240 bacilles à Gram négatif ont été isolés à partir de
différents services. Les entérobactéries dominent dans notre étude avec une fréquence
d’isolement de 56% (140 souches) par rapport à 40% des bacilles à Gram négatif
(BGN) non fermentants (Sefraoui, 2010). Ces résultats concordent bien avec ceux
obtenus au Maroc (Elouennass, 2008), en France (Zogheib et Dupont, 2005) et au
Cameroun (Gangoué Piéboji et al., 2004). Dans notre étude, les entérobactéries ont
été retrouvées dans tous les types de services. Cependant, certaines spécialités sont
apparues plus concernées dont la réanimation et la chirurgie avec 27.8% et 26.4%
respectivement.
Dans l’ensemble des entérobactéries isolées, Escherichia coli reste l’espèce la plus
fréquente (37.1%), suivie par Klebsiella pneumoniae (21.4%), comme cela été
rapporté par plusieurs études (Messai et al., 2007; Nadmia et al., 2010). Alors que,
Proteus spp. et Enterobacter cloacae occupent la dernière
place avec des taux
d’isolement pratiquement identiques soit 19.3% et 18.6% respectivement. Le même
classement a été rapporté par Nijssen et al. (2004) mais avec des pourcentages de
2.9% pour Enterobacter cloacae et de 2.2% pour Proteus mirabilis.
Aussi, ces espèces sont réparties différemment selon les services et les sites de
prélèvement. L’infection urinaire est une pathologie fréquente en pratique quotidienne.
L’espèce bactérienne principale impliquée dans cette infection est Escherichia coli
puisqu’elle représente 50 à 80% des agents en cause (Farrell et al., 2003; Matute et
al., 2004; Zhanel et al., 2006; Bouzenoune, 2009). Ceci correspond aux résultats
obtenus dans notre étude avec un taux de 61.1%. Cela est en rapport avec la
physiopathologie de l’infection urinaire qui est en général ascendante, et il existe une
forte colonisation du périnée par les entérobactéries d’origine digestive, et en
particulier Escherichia coli. À cela s’ajoutent des facteurs spécifiques

Discussion
76
d’uropathogénicité. Ainsi, Escherichia coli possède des adhésines, capables de lier la
bactérie à l’épithélium urinaire et d’empêcher son élimination. Klebsiella et Proteus
sécrètent une uréase qui alcalinise l’urine dont le pH, naturellement acide, empêche la
prolifération des germes (Sekhsokh, 2008). En effet, Klebsiella pneumoniae est
beaucoup plus isolée à partir des voies respiratoires (50 %) et elle est l’agent
responsable de pneumonies (Paterson, 2006). À noter également que les souches de
Proteus spp. sont essentiellement isolées des prélèvements de plaies chirurgicales soit
70.4%, ces espèces se comportent souvent comme des pathogènes opportunistes. Elles
sont alors impliquées dans des pathologies infectieuses, notamment chez les individus
hospitalisés (unités de gériatrie, de psychiatrie, de soins intensifs), immunodéprimés,
sondés, paraplégiques ou ayant des anomalies des voies urinaires. Ces infections sont
associées aux trois espèces principales: Proteus vulgaris, Proteus mirabilis et Proteus
penneri. Toutefois, Proteus mirabilis est l’espèce la plus communément isolée des
prélèvements cliniques. Cela peut s’expliquer par son taux élevé de portage (25%)
dans le tube digestif de l’homme (Mahamat et al., 2006). Enfin, on observe une
prédominance des Enterobacter cloacae au niveau de l’environnement du CHU de
Sidi Bel Abbes, avec un taux de 45%.
Les niveaux des résistances bactériennes varient d’un pays à l’autre et d’une année à
l’autre. Aussi, la connaissance de la situation locale et de son évolution sont
nécessaires pour le choix de l’antibiothérapie de première intention (El Bakkouri et
al., 2009). En effet, nos résultats montrent un taux de résistance considérable à la
majorité des antibiotiques testés. Les entérobactéries résistantes aux céphalosporines
de 3eme
génération (C3G) occupent une place importante dans les infections
nosocomiales, notamment en milieu de réanimation. Ces bactéries deviennent de plus
en plus résistantes aux antibiotiques et commencent à franchir les limites de l’hôpital
pour émerger dans la communauté. La dissémination de ces bactéries présente une
menace grave qui met en cause la validité de l’arsenal antibiotique actuellement
disponible, d’autant plus qu’aucune classe nouvelle d’antibiotique n’est attendue dans
les prochaines années (Mkaouar, 2008). L’étude de la sensibilité des entérobactéries
aux C3G, montrait des taux de résistances acquises moyenne à l’hôpital. Les niveaux

Discussion
77
de résistance étant considérés inférieurs à ceux observés au CHU de Tlemcen (Ayad,
2010), sont en revanche plus élevés que ceux rapportés en Algérie (Amazian et al.,
2006), en Tunisie (Boutiba et al., 2004), en Australie, en France, en Argentine
(Navon-Venezia et al., 2003; Masterton et Turner, 2006) et dans certains pays
asiatiques (Inde et Chine) (Hirakata et al., 2005).
Depuis plus de 20 ans, la résistance des entérobactéries aux céphalosporines de 3eme
génération (C3G) ne cesse de se renforcer notamment par l’acquisition de -
lactamases à spectre élargi (BLSE) (Belmonte et al., 2010). Ainsi 37.1% des souches
étudiées sont productrices de -lactamases à spectre étendu (BLSE). Cette valeur est
nettement inférieure à celle retrouvée à Tlemcen (Ayad, 2010) (67%), se rapproche de
celle obtenue en Tunisie (30.8 %) (Messai et al., 2007) et en Egypte (38.5%) mais
supérieure à celle rapportée en France (5.2%), en Allemagne (2.6%) et en Hollande
(2%) (Ho et al., 2000; Bouchillon et al., 2004). Aussi, La coexistence des différentes
-lactamases dans une même bactérie peut poser un problème de diagnostic et de
thérapeutique. Ces associations conduisent à une co-sélection des gènes de résistance
et au scénario de pandémie à l’hôpital et en communauté (Chen et al., 2007; Meradi
et al., 2009). Cependant, on note que le nombre des souches productrices de BLSE est
plus important chez Klebsiella pneumoniae avec 80% qui correspond aux résultats
obtenus par Sekhsokh (2008) et par Edelstein et al. (2003). Enterobacter cloacae
arrive en 2eme
place avec 57.7%. Ce taux est supérieur à celui rapporté en France par
Belmonte et al. (2010) (39%) et par Doit et al. (2010) (18%). Notons également un
taux de production de BLSE de 18.5% pour les souches de Proteus spp. plus élevé par
rapport aux résultats obtenus aux États-Unis (9.5 %) (Saurina et al., 2000), en Italie
(8.8 à 10 %) (Pagani et al., 2002) et en France (10.2%) (Mahamat et al., 2006). Les
résultats révèlent une faible fréquence du phénotype BLSE chez Escherichia coli
(15.4%) contrairement à plusieurs études réalisées par Leotard et Negrin (2010) et
Holstein et al. (2010). Le taux de production de pénicillinase acquise de haut niveau
est de 25% dont prés de 11.4 % n’étaient pas récupérées par le tazobactam et le
phénotype sauvage a concerné 33.6% des isolats.

Discussion
78
Le taux d’isolement des souches présentant le phénotype BLSE est plus important
en réanimation (48.7%) qu’en chirurgie, ou autres services signalés, ce qui est
certainement lié à l’utilisation abusive voire massive des antibiotiques en soins
intensifs (Carlet, 2002). C'est ainsi que l'utilisation large des céphalosporines de 3eme
génération (C3G) a eu comme conséquence le développement d'entérobactéries
résistantes par production de -lactamases à spectre étendu (BLSE), la production
d’une telle enzyme permet à la bactérie de résister à toutes les -lactamines excepté les
céphamycines et l'imipénème. Elles sont souvent responsables d'épidémies notamment
dans les unités de soins intensifs (Ben Redjeb et al., 2000).
En outre, le problème lié aux BLSE est surtout la présence fréquente de co-
résistances rendant les souches multirésistantes (Leotard et Negrin, 2010). En effet,
les BLSE sont généralement portées par de grands plasmides qui portent aussi des
gènes de résistance aux classes d’antibiotiques non -lactamines, tels que les
aminoglycosides, les quinolones et le triméthoprime/sulfaméthoxazole. Aussi,
l’utilisation de ces antibiotiques contribue à la sélection de souches productrice
de BLSE (Paterson et Bonomo, 2005) . L’incidence d’apparition des résistances
aux aminosides a augmenté ces derniers années et particulièrement chez les souches
productrice de BLSE (Spanu et al., 2002). Les niveaux de résistance de ces souches
dans notre étude est plus important. L’amikacine reste la molécule la plus efficace
avec 96.4% de souches sensibles comme cela été rapporté dans plusieurs études
(Gangoue Piebouji, 2007; Wu et al., 2007). Les quinolones montrent une activité
moyenne, la sensibilité globale des souches chutant considérablement dans le cas des
souches productrices de BLSE. Pour la majorité des souches, la résistance acquise est
la conséquence d’une mutation, ce qui limite sa diffusion (Larabi et al., 2003). Une
activité faible est détectée pour le triméthoprime/sulfaméthoxazole avec 60%. Ce
taux reste supérieur à celui décrit dans la littérature par plusieurs études (Ammari et
al., 2001; Kahlmeter, 2003; Guneysel et al., 2009) dont l’évolution de la résistance à
cette molécule a été marquée par une décroissance significative. Cela est
vraisemblablement lié à une moindre utilisation de cet antibiotique en pratique
courante (Mahamat et al., 2006). En ce qui concerne la colistine, c’est l’une des

Discussion
79
molécules gardant les taux de sensibilité les plus élevés sur les espèces naturellement
sensibles (en dehors des Proteus) avec un taux de résistance plus élevé au service de
chirurgie (Hamze et al., 2003).
Des gènes de résistances à différentes familles d’antibiotiques sont décrits comme
présents sur un même plasmide, représentant ainsi un mode de diffusion efficace de
plusieurs mécanismes associés (Leotard et Negrin, 2010). Cette dissémination de la
multirésistance et cette diffusion des gènes de résistance sont liées à l’existence
d’éléments génétiques mobiles entre bactéries d’une même espèce ou d’espèces
différentes, ainsi qu’à l’existence de structures génétiques permettant de cumuler de
nombreux gènes de résistance au sein d’une même souche (Skurnik et Andremont,
2006). Les résultats de la visualisation des plasmides sur gel d’agarose semblent
révéler que les gènes codant pour cette résistance soient situés sur des plasmides de
taille supérieure à 55 Kb chez les souches ayant transféré le phénotype BLSE et qui
souvent, portent des déterminants de la résistance à d'autres familles d'antibiotiques, ce
qui peut expliquer que les bactéries qui les hébergent deviennent multi-résistantes
(Ben Redjeb et al., 2000).
Les carbapénèmes sont les traitements de choix des infections sévères causées par
les entérobactéries productrices de BLSE (Gülmez et al., 2008). La résistance des
entérobactéries à ces molécules reste encore un phénomène marginal comme en
témoignent les données épidémiologiques obtenues pour un grand nombre de souches,
avec des pourcentages de sensibilité de 99-100 % (Wolff et al., 2008). Dans notre
étude, 4.3% des souches présentent une résistance intermédiaire à l’imipénème avec
des CMI de 4 g/ml. Cette baisse de la sensibilité pourrait être le résultat d’une
modification des porines, associée à la production de β-lactamases de type BLSE ou
AmpC, diminuant ainsi la perméabilité de la membrane externe (Bennett et al., 2010).
Le plus inquiétant serait la production de carbapénèmases puisque l’activité des
carbapénèmes peut être compromise par l’émergence de ces enzymes (Pasteran et al.,
2008; Sekhri et al., 2010). Dans l’édition de janvier 2008 du document CLSI M100
(Clinical and Laboratory Standards Institute, 2008), un article a inclu une alerte

Discussion
80
aux laboratoires pour contrôler et chercher toute production de carbapénèmase chez les
entérobactéries avec des CMI de 2 ou 4 g/ml (Patel et al., 2009). Les méthodes
phénotypiques, qui peuvent être combinées entre elles, ont l’avantage d’être faciles à
réaliser. Elles permettent de mettre en évidence une résistance aux carbapénèmes sans
en identifier précisément le mécanisme. La confirmation de la présence de ces gènes
ne peut se faire actuellement que par l’utilisation d’outils moléculaires (Cuzon et al.,
2009). Le traitement des infections causées par les entérobactéries résistantes aux
carbapénèmes est difficile en effet, les choix thérapeutiques contre ce type de germes
sont limités. Pour éviter ces épidémies, on doit procéder à des surveillances strictes de
la résistance et de la consommation des antibiotiques à l’hôpital (Wu et al., 2010).
Les entérobactéries produisant une BLSE (E-BLSE+) sont toujours d’actualité dans
les structures de soins, malgré les mesures de prévention de leur dissémination
aujourd’hui bien codifiées. On observe plutôt une augmentation de leur nombre, en
particulier, avec des BLSE de type CTX-M alors que les BLSE plus « anciennes » de
la famille des TEM ou des SHV semblent diminuer (Giraud-Morin et Fosse, 2008).
Les niveaux de CMI pour le céfotaxime chez la plupart des souches présentant le
phénotype BLSE sont plus importants par rapport à la céftazidime, ce qui oriente vers
des BLSE de type CTX-M. Les enzymes de ce type ont eu un fort succès
épidémiologique et ont diffusé dans le monde entier (Doit et al., 2010), parmi elles,
les CTX-M15 semblant avoir une capacité particulière de dissémination (Nicolas-
Chanoine et al., 2008). Au cours de cette étude, une souche de Proteus mirabilis
(S.Pm.3) a été envoyée au laboratoire de bactériologie de l’hôpital Tenon à Paris pour
la recherche des gènes codant pour la production de BLSE. L’analyse moléculaire est
réalisée par l’équipe de Mr Guillaume Arlet, le résultat indique que cette souche est
productrice d’une BLSE de type CTX-M du groupe 1. La détermination et la
compréhension des mécanismes moléculaires du pouvoir pathogène et en particulier de
l’émergence des caractères de résistance aux antibiotiques représentent un des buts
essentiels de la bactériologie médicale pouvant faire évoluer les stratégies
thérapeutiques (Boukerzaza, 2005).

Conclusion

Conclusion
81
Les résultats de cette étude révèlent d’abord une certaine diversité pour les espèces
d’entérobactéries identifiées. Les quatre espèces représentées sont Escherichia coli,
Klebsiella pneumoniae, Proteus spp. et Enterobacter cloacae. Au total, 140 souches
d’entérobactéries ont été isolées à partir de différents services aux CHU de Sidi Bel
Abbes entre octobre 2009 et mai 2010. De plus, un grand nombre de ces souches ont
présenté des phénotypes de résistance à une ou plusieurs familles d’antibiotiques en
particulier les -lactamines, les aminosides, les quinolones, la colistine et le
triméthoprime/sulfamethoxasole.
Les niveaux de résistance atteints vis-à-vis les -lactamines sont associés à de
nombreux mécanismes de résistance, avec prédominance de la production de -
lactamases à spectre étendu (BLSE) soit (37.1%), mais aussi la production de
pénicillinase avec 25% et une minorité la céphalosporinase hyperproduite avec 4.3%.
Par ailleurs, les aminosides, les quinolones et la colistine ont gardé globalement
leur activité dans le traitement des infections à entérobactéries. Notons toutefois que
l’imipénème et l’amikacine restent les antibiotiques les plus actifs.
Le problème des BLSE est surtout lié à la présence fréquente de co-résistances
comme par exemple, celles aux aminosides et quinolones. Des gènes de résistances à
différentes familles d’antibiotiques sont décrits comme présents sur un même
plasmide, représentant ainsi un mode de diffusion efficace de plusieurs mécanismes
simultanément. La diffusion de ces résistances pourrait aussi être le fait d’intégrons
souvent décrits chez les CTX-M (Leotard et Negrin, 2010).
Une meilleure maîtrise en termes d’hygiène hospitalière (renforcement de la
formation du personnel aux règles préventives, précocité du dépistage. . .) et un
meilleur contrôle de la consommation d’antibiotiques au sein de l’établissement
seraient toutefois des facteurs favorisant une meilleure maîtrise des risques (Belmonte,
2010).

Conclusion
82
L’analyse moléculaire des clones détectés localement paraît indispensable à terme
pour une compréhension et une réaction adaptée à l’émergence des entérobactéries. La
détection ou non de la circulation à grande échelle de certains types de BLSE et
notamment des CTX-M permettrait de rattacher les évènements locaux à ceux
observés au sein de l’Algérie et plus globalement à l’échelle mondiale. Cela
permettrait également d’envisager de façon plus précise le risque encouru à l’échelle
communautaire.

Références
Bibliographiques

Références Bibliographiques
83
A:
1 Abdou Souley Lie. 2002. Sensibilité et évolution de la résistance de
Pseudomonas aeruginosa aux antibiotiques à l'hôpital du point G. La faculté de
Médecine, de Pharmacie et d'Odonto-Stomatologie. Docteur en Pharmacie
(Diplôme d'état).
2 Aggoune-Khinache N., Bensersa D., Henniche F.Z., Daoudi M., Abdouni M.A.,
Chabani A., Tiouit D. et Naim M. 2009. Pseudomonas aeruginosa producteurs
de métallo--lactamases en Algérie. Médecine et maladies infectieuses; 39:
413-414.
3 Ahoyo A.T., Baba-Moussa L., Anago A.E., Avogbe P., Missihoun T.D., Loko
F., Prévost G., Sanni A. et Dramane K. 2007. Incidence d’infections liées à
Escherichia coli producteur de -lactamase à spectre élargi au Centre
hospitalier départemental du Zou et Collines au Bénin. Médecine et maladies
infectieuses; 37: 746-752.
4 Amazian K., Fendri C., Missoum M.F.K., Bouzouaia N., Rahal K., Savey A., et
al. 2006. Multicenter pilot survey of resistant bacteria in the Mediterranean
area. Eur J Clin Microbiol Infect Dis; 25: 340-3.
5 Amhis W. 2004. « Le genre Proteus, épidémiologie, bactériologie, résistance
aux antibiotiques et biologie moléculaire». Thèse de DESM.
6 Ammari H., Ramdani-Bouguessa N. et Bellouni R. 2001. Antibiothérapie dans
les infections ORL. Médecine du Maghreb; n°91.
7 Anderson K., Lonsway D.R., Rasheed J.K., Biddle J., Jensen B., McDougal
L.K., et al. 2007. Evaluation of Methods to Identify the Klebsiella pneumoniae
Carbapenemase in Enterobacteriaceae. J. Clin. Microbiol; 45(8): 2723-2725.
8 Andreuws J.M. 2006. Determination of Minimum Inhibitory Concentrations.
Department of microbiology, City Hospital NHS Trust, Birmingham B187 QH.
9 Aubert G. et Carricajo A. 2004. Place du laboratoire dans le choix et le suivi
pharmacodynamique de l’antibiothérapie des infections sévères. Annales
Françaises d’Anesthésie et de Réanimation; 23: 704–713.
10 Avril J.L., Dabernat H., Denis F. et Monteil H. 2000. Bactériologie clinique.
3ème
édition. Ellipses édition marketing S.A. Paris. p 171-229.

Références Bibliographiques
84
11 Ayad A. 2010. Etude de la résistance aux antibiotiques des entérobactéries au
niveau du CHU de Tlemcen. Thèse de magister.
B:
12 Barrial. K. et Scotet. J. 2005. Classification raisonnée des -lactamases chez les
bacilles à Gram négatif, Perspectives d’évolution. Encadrement : Dr Sylvestre
TIGAUD. DES BACTERIOLOGIE, Semestre Hiver; 05(06): 1-18.
13 Basilio J.A. 2009. Serratia. eMedicine Specialties. Infectious Diseases.
Bacterial Infections. Sur le lien :
http://emedicine.medscape.com/article/228495-overview.
14 Belmonte O., Drouet D., Alba J. Moiton M.-P., Kuli B., Lugagne-Delpon N.,
Mourlan C. et Jaffar-Bandjee M.-C. 2010. Evolution de la résistance des
entérobactéries aux antibiotiques sur l’île de la Réunion : émergence des -
lactamases à spectre élargi. Pathologie Biologie; 58: 18–24.
15 Benlmouden A. et Hakkou F. 2007. Antibiotiques : Mécanismes d’action et de
résistance. Service de Pharmacologie, Faculté de Médecine et de Pharmacie,
Casablanca, Maroc. La Chronique Ibn Rochd; 1: 46-54.
16 Bennett J.W., Mende K., Monica L., Herrera, Yu X., Lewis J. S., Wickes B. L.,
Jorgensen J. H. et Murray C.K. 2010. Mechanisms of carbapenem resistance
among a collection of Enterobacteriaceae clinical isolates in a Texas City.
Diagnostic Microbiology and Infectious Disease; 66: 445–448.
17 Ben Redjeb S., Ben Hassen A., Hammami A. et Kechrid A. 2000.
Epidémiologie des résistances bactériennes en Tunisie. Laboratoire “Résistance
aux antibiotiques” - Faculté de Médecine - Tunis.
18 Berche P. 2003. Bactériologie systématique D.C.E.M. 1. Faculté de médecine
Necker-Enfant malade. Sur le lien : www.medix.free.fr/.../bacterie-diarrhee-
aigue.php
19 Besassier R., Califano J. Carrette M. et Lombardo M. 2005. La lutte
antibactérienne. Université Nice Sophia Antipolis. Polytech’ Nice-Sophia.
Département Génie Biologique. Sur le lien :
http://bioinfo.unice.fr/enseignements/EPU_2005/2005_UET1/Rapport%20lutte
%20antibact%E9rienne%20et%20r%E9sum%E9.pdf.

Références Bibliographiques
85
20 Bessard G. 2005. Pharmacologie des antibiotiques. Faculté de médecine de
Grenoble. Université Joseph Fourier. DCEM I. Sur le lien: www-sante.ujf-
grenoble.fr/SANTE/pharma/site.fac/diaporama/antibio_oct2005.ppt.
21 Bessard G. 2004. Pharmacologie des antibiotiques. Faculté de médecine de
Grenoble. Université Joseph Fourier. DCEM I. Sur le lien: www-sante.ujf-
grenoble.fr/SANTE/pharma/site.fac/diaporama/antibio2.ppt.
22 Bonacorsi S., Houdoin V. et Bingen E. 2001. Facteurs de virulence associés à
E. coli responsable de méningite néonatale. Elsevier SAS. Service de
microbiologie, hôpital Robert-Debré, 48, boulevard Sérurier, 75019 Paris,
France. Arch Pédiatr; 8 Suppl; 4: 726-31.
23 Bonnet R. 2006. -lactamines et entérobactéries. Chapitre 15. Livre
antibiogramme. Paris. ESKA: 2éme édition. p 143.
24 Bonnet R. 2004. Growing group of extended-spectrum -lactamases. The CTX-
M enzymes. Antimicrob. Agents Chemother; 48: 1-14.
25 Bouchillon, S.K., Johnson B.M.et Hoban D.J. 2004. Determining incidence of
extended-spectrum β-lactamase producing Enterobacteriaceae, Vancomycin-
resistant Enterococcus faecium and methicillin-resistant Staphylococcus aureus
in 38 centres from 19 countries: The PEARLS study 2001-2002. Int. J.
Antimicrob. Agents; 24: 119-124.
26 Boukadida J., Salem N., Hannachi N., Monastiri K.et Snoussi N. 2002.
Exploration génotypique d’une bouffée épidémique nosocomiale néonatale à
Klebsiella pneumoniae productrice de -lactamase à spectre étendu. Éditions
scientifiques et médicales Elsevier SAS. Arch Pédiatr; 9: 463-8.
27 Boukerzaza S. 2005. Etude génétique et moléculaire du phénotype de résistance
des Salmonella enterica sérotypes Senftenberg. Thèse de Magistère en
Génétique Moléculaire. Université Mentouri de Constantine. Faculté des
Sciences de la Nature et de la vie. Département des Sciences de la Nature et de
la vie.
28 Boutiba I, Ghozzi R, Ben Abdallah H, Mamlouk K, Kamoun A. et Ben Redjeb
S. 2004. Evolution of acquired resistance to third generation Cephalosporins in

Références Bibliographiques
86
Enterobacteriaceae in a Tunisian Hospital 1993–2001. Clin Microbiol Infect;
10 (7): 657-78.
29 Bouzenoune F., Boudersa F., Bensaad A., Harkat F. et Siad N. 2009. Les
infections urinaires à Ain M’lila (Algérie). Résistance aux antibiotiques des 239
souches isolées entre 2006 et 2007. Médecine et maladies infectieuses; 39: 142-
143.
30 Branger C. G., Torres-Escobar A., Sun W., Perry R., Fetherston J., Roland K.
L. et Curtiss R. 2009. Oral vaccination with LcrV from Yersinia pestis KIM
delivered by live attenuated Salmonella enterica serovar Typhimurium elicits a
protective immune response against challenge with Yersinia pseudotuberculosis
and Yersinia enterocolitica. Vaccine; 27(39): 5363-5370.
31 Brenwald N. P., Jevons G., Andrews J. M., Xiong J.-H., Hawkey P. M. et Wise
R. 2008. Un foyer d'un CTX-M-type ß-lactamase-production Klebsiella
pneumoniae: l'importance de l'utilisation de détecter cefpodoxime extended-
spectrum ß-lactamases. Department of Microbiology, city hospital, Birmingham
B18 7QH; Division of immunity and infection, medical school, university of
Birmingham, Birmingham B15 2TT, UK.
32 Bryskier A. 1999. Antibiotiques, agents antibactériens et antifongiques.
Ellipses; Paris. p 436-445.
C:
33 Carlet J. 2002. Risque infectieux en réanimation: gestion et prévention. Elsevier
Masson. p 99-100.
34 Carrër A. et Nordmann P. 2009. Klebsiella pneumoniae CTX-M-15 : vers une
modification de l’épidémiologie des ß-lactamases à spectre étendu. Pathologie
Biologie.
35 CA-SFM. 2010. Comité de l’antibiogramme de la société française de
microbiologie. Sur le lien : http://www.sfm.asso.fr/.
36 Castanheira M., Sader H. S., Deshpande L.M., Fritsche T. R. et Jones R. N.
2008. Antimicrobial Activities of Tigecycline and Other Broad-Spectrum
Antimicrobials Tested against Serine Carbapenemase and Metallo--
Lactamase-Producing Enterobacteriaceae: Report from the SENTRY

Références Bibliographiques
87
Antimicrobial. Surveillance Program. Antimicrobial Agents and Chemotherapy;
52: 570-3.
37 Cattoir V. 2006. Chloramphenicol, fosfomycine, acide fusidique et
polymyxines. Chapitre 28. In Antibiogramme. Paris. ESKA : 2éme édition. p
359-363.
38 Cattoir V. 2004. Pompes d’efflux et résistance aux antibiotiques chez les
bactéries. Pathologie Biologie; 52: 607-616.
39 Cavallo J.-D., Fabre R., Jehl F., Rapp C. et Garrabé E. 2004. -lactamines.
EMC-Maladies Infectieuses ; 1 : 129-202.
40 Chander Y., Goyal S.M. et Gupta S.C. 2006. Antimicrobial resistance of
Providencia spp. isolated from animal manure. The Veterinary Journal; 172:
188-191.
41 Chen L., Hainrichson D., Bourdetsky M., Mor A., Yaron S. et Baasov T. 2008.
Structure–toxicity relationship of aminoglycosides: Correlation of 2’-amine
basicity with acute toxicity in pseudo-disaccharide scaffolds. Bioorganic &
Medicinal Chemistry; 16: 8940-8951.
42 Chen L.F. et Kaye D. 2009. Current Use for Old Antibacterial Agents:
Polymyxins, Rifamycins, and Aminoglycosides. Antibacterial Therapy and
Newer Agents. Infectious Disease Clinics of North America. Antibacterial
Therapy and Newer Agents; 23(4): 1053-1075.
43 Chen Y.-T., Lauderdale T.-L., Liao T.-L., Shiau Y.-R., Shu H.-Y., Wu K.-M.,
Yan J.-J., I.-J. Su, et Tsai1 S.-F. 2007. Sequencing and Comparative Genomic
Analysis of pK29, a 269-Kilobase Conjugative Plasmid Encoding CMY-8 and
CTX-M-3 -Lactamases in Klebsiella pneumoniae. Antimicrobial Agents and
Chemotherapy; 51(8): 3004-3007.
44 Choi, S. H., J. E. Lee, S. J. Park, S. O. Lee, J. Y. Jeong, M. N. Kim, J. H. Woo,
et Kim Y. S. 2008. Emergence of antibiotic resistance during therapy for
infections caused by Enterobacteriaceae producing AmpC -lactamase:
implications for antibiotic use. Antimicrob. Agents Chemother; 52: 995-1000.

Références Bibliographiques
88
45 Chou Y.Y., Chiu S-K., Lai H-C. et Chang F-Y. 2009. Tubo-ovarian abscess
with Morganella Morganii bacteremia. J Microbiol Immunol Infect; 42: 357-
359.
46 CHU-PS Pitié-Salpêtrière. 2003. Bactériologie DCEM1. Université PARIS-VI
Pierre et Marie Curie. Faculté de Médecine Pitié-Salpêtrière. Service de
Bactériologie.
47 Clinical and Laboratory Standards Institute. 2008. Performance standards for
antimicrobial susceptibility testing; 18th informational supplement. CLSI
document M100-S18. CLSI,Wayne, PA.
48 Cohen R. 2010. Colimycine: un vieil antibiotique qu’il faut apprendre à
connaître. Archives de Pédiatrie;17:S171-S176.
49 Collignon A., Beljean-Leymarie M., Farinotti R. et Doutremepuich C. 2007.
Infectiologie. Volume 3 de Collection Le Moniteur internat. Le Moniteur des
Pharmacies N° 3. p: 354-355.
50 Courvalin P. 2008. La résistance des bactéries aux antibiotiques: combinaisons
de mécanismes biochimiques et génétiques. Bacterial antibiotic resistance:
combinations of biochemical and genetic mechanisms. Bull. Acad. Vét. France-
Tome 161 - N°1: 7-12.
51 Courvalin P. Leclerck R. et Bingen E. 2006. Antibiogramme. Paris. ESKA :
2éme édition. p 141-162.
52 Courvalin P., Drugeon N., Flandrois J.P. et Goldstein F. 1991. Aspects
théoriques et thérapeutiques. Bactéricidie. Ed Maloine. p113-26.
53 Cuzon G., Naas T. et Nordmann P. 2009. Carbapénèmases de type KPC: quel
enjeu en microbiologie clinique ? Pathologie Biologie.
D:
54 Dagnra A.-Y., Akolly K., Gbadoe A., Aho K. et David M. 2007. Émergence
des souches de salmonelles multirésistantes aux antibiotiques à Lomé (Togo).
Médecine et maladies infectieuses; 37 : 266-269.
55 De Champs C. 2002. Perspective survey of -lactamase produced ceftazidim.
Agent chemother; 46(9):30-34.

Références Bibliographiques
89
56 Decoster A. 2005. Entérobactéries. FLM. p 1-16. Sur le lien
http://anne.decoster.free.fr/btelechar/bpoly/enteroba05.pdf.
57 Delarras C. 2007. Microbiologie pratique pour le laboratoire d’analyses ou de
contrôle sanitaire. Edition Techniques et Documentation. Paris. p 248-296.
58 Denis F. et Ploy M.-C. 2007. Bactériologie médicale: techniques usuelles.
Elsevier Masson. p 316-318.
59 De Wals P.-Y. 2007. Analyses mutationnelles et cinétiques de la β-lactamase
TEM-1 d’Escherichia coli vers une meilleure compréhension du phénomène de
résistance aux antibiotiques. Département de biochimie. Faculté de médecine.
Université de Montréal.
60 Doi Y. et Arakawa Y. 2007. 16S ribosomal RNA methylation: emerging
resistance mechanism against aminoglycosides. Clin Infect Dis; 45: 88–94.
61 Doit C., Mariani-Kurkdjian P. et Bingen E. 2010. Entérobactéries productrices
de -lactamases à spectre étendu. Archives de Pédiatrie;17: S140 -S144.
62 Dufour J-P. 2005. Les diarrhées du macaque cynomolgus (Macaca
fascicularis): essai de prophylaxie dans un élevage de l’île Maurice.
L’université Paul-Sabatier De Toulouse. Thèse pour obtenir le grade de docteur
vétérinaire.
63 Durante-Mangoni E., Grammatikos A., Utili R. et Falagas M.E. 2009. Do we
still need the aminoglycosides? International Journal of Antimicrobial Agents;
33: 201-205.
E:
64 Edelstein M., Pimkin M., Palagin I., Edelstein I. et Stratchounski L. 2003.
Prevalence and molecular epidemiology of CTX-M extended-spectrum -
lactamase-producing Escherichia coli and Klebsiella pneumoniae in Russian
hospitals. Antimicrob Agents Chemother; 47: 3724-3732.
65 El Bakkouri J., Belabbes H., Zerouali K., Belaiche A., Messaouidi D., Gros
Claude J. D. P. et El Mdaghri N. 2009. Résistance aux Antibiotiques
d’Escherichia coli Uropathogène Communautaire et Consommation
d’Antibiotiques à Casablanca (Maroc). European Journal of Scientific Research
ISSN 1450-216X; 36(1): 49-5.

Références Bibliographiques
90
66 Elouennass M., Sahnoun I., Zrara A., Bajjou T. et Elhamzaoui S. 2008.
Épidémiologie et profil de sensibilité des isolats d’hémoculture dans un service
de réanimation (2002–2005). Médecine et maladies infectieuses; 38 : 18-24.
67 Emmanuel E. 2003. Evaluation des risques sanitaires et ecotoxicologiques lies
aux effluents hospitaliers. L’institut national des sciences appliquées de LYON
pour l'obtenir de grade du docteur. Formation doctorale: science et technique
du déchet école doctoral de chimie de Lyon. p 59-60.
68 Euzéby J.P. 2003. Dictionnaire de Bactériologie Vétérinaire. SERRATIA. Sur
le liens : http://www.bacterio.cict.fr/bacdico/.
69 Eyquem A. et Montagnier L. 2000. Traité de microbiologie clinique:
deuxièmes mises à jour et compléments. PICCIN. p 33-34.
70 Ezaitouni F., Rhou H., Benamar L., Ouzeddoun N., Bayahya R. et Balafrej L.
1999. Rein et aminosides. Médecine du Maghreb n°77.
F:
71 Farah A., Boutefnouchet N., Dekhil M. et Bouzerna N. Klebsiella pneumoniae
productrices de –lactamases a spectre élargi (BLSE) isolées dans les hôpitaux
de la ville de Annaba, Algérie. Scientific Study & Research ; VIII (2) (1582-
540X): 199-214.
72 Farrell DJ, Morissey I, De Rubeis D et al. 2003. A UK multicentre study and
the antimicrobial susceptibility of bacterial pathogens causing urinary tract
infection. J Infect; 46(2): 94-100.
73 Fauchère J.L. et Avril J.L. 2002. Bactériologie générale et médicale. Ellipses
Edition Marketing. Paris. p 250-260.
74 Faure S. 2009.a. Les aminosides, Pharmacothérapeutique pratique. Actualité
pharmaceutique n° 482 : 51-55.
75 Faure S. 2009.b. Transfert d’un gène de résistance aux β-lactamines bla CTX-
M-9 entre Salmonella et les entérobactéries de la flore intestinale humaine :
impact d’une antibiothérapie. Thèse De Doctorat de L’université de Rennes.
76 Fluit, A. et Schmitz F. 2004. Resistance integrons and super-integrons. Clin
Microbiol Infect; 10: 272-288.

Références Bibliographiques
91
77 Frasca D., Dahyot-Fizelier C. et Mimoz O. 2008. La colistine en réanimation.
Réanimation; 17: 251-258.
78 Fraser S. L., Arnett M. et Sinave C.P. 2010. Enterobacter Infections. eMedicine
Specialties. Infectious Diseases. Bacterial Infections. Contributor Information
and Disclosures.
G:
79 Galani I., Rekatsina P. D. Hatzaki D., Plachouras D., Souli M. et Giamarellou
H. 2008. Evaluation of different laboratory tests for the detection of Metallo--
lactamase production in Enterobacteriaceae. Journal of Antimicrobial
Chemotherapy; 61: 548-553.
80 Galimand M., Sabtcheva, S., Courvalin, P. et Lambert, T. 2005. Worldwide
disseminated armA aminoglycoside resistance methylase gene is borne by
composite transposon Tn1548. Antimicrob Agents Chemother; 49: 2949-2953.
81 Gangoue Pieboji J. 2007. Caractérisation des -lactamases et leur inhibition
par les extraits des plantes médicinales. Centre d'ingénierie des protéines.
Université de Liége. Thèse de doctorat. p 1-27.
82 Gangoué Piéboji J., Koulla-Shiro S., Ngassam P., Adiogo D., Njine T. et
Ndumbe P. 2004. Antimicrobial resistance of Gram-negative bacilli isolates
from inpatients and outpatients at Yaounde Central Hospital, Cameroon.
International Journal of Infectious Diseases; 8: 147-154.
83 Gattoir V. 2006. Chloramphénicol, Fosfomycine, Acide fusidique et
Polymyxines. Chapitre 28. Livre antibiogramme. ESKA : 2éme édition. p 359-
364.
84 Gautier V. 2007. Caractérisation et expression des gènes codant pour les -
lactamases chromosomiques au sein des entérobactéries de l’environnement. Ecole
pratique des hautes études Sciences de la Vie et de la Terre. Mémoire Pour
l’obtention du diplôme de l’Ecole Pratique des Hautes Etudes.
85 Giraud-Morin C. et Fosse T. 2008. Évolution récente et caractérisation des
entérobactéries productrices de BLSE au CHU de Nice (2005–2007).
Pathologie Biologie; 56 :417-423.

Références Bibliographiques
92
86 Gniadkowski M. 2001. Evolution and epidemiology of extended-spectrum -
lactamases (ESBLs) and ESBL-producing microorganisms. Clin Microbiol
Infect 7. p 597-608.
87 Golsdein F.W. 2006. Sulfamides et Triméthoprime. Chapitre 27. Livre
antibiogramme. ESKA : 2éme édition. p 341-348.
88 Goubau P. et Pellegrims E. 2000. Repères en microbiologie. Garant, Édition3. p
116-117.
89 Guessennd N, Bremont S., Gbonon V., Kacou-NDouba A., Ekaza E., Lambert
T., Dosso M. et Courvalin P. 2008. Résistance aux quinolones de type qnr chez
les entérobactéries productrices de -lactamases à spectre élargi à Abidjan en
Côte d’Ivoire. Pathologie Biologie; 56 : 439-446.
90 Gueudet T., Richter S., Szulc M. et Jehl F. 2010. Les nouvelles formes de
résistance des bactéries aux antibiotiques : deux cas de Klebsiella pneumoniae
produisant une céphalosporinase plasmidique. Médecine et maladies
infectieuses ; 40: 177-179.
91 Guillard T., Cavallo J.-D., Cambau E., Duval V., Bajolet O. Brasme L., de
Champs C. et Vernet-Garnier V. 2009. Mise au point d’une technique de PCR
en temps réel pour la détection rapide des gènes qnr chez des entérobactéries
productrices de -lactamases à spectre étendu. Pathologie Biologie (Paris) p 1-
4.
92 Guillard T., Heckel E. et Galimand M. 2008. Résistance plasmidique aux
quinolones chez les bactéries à Gram négatif. p 1-8.
93 Gulmez D., Woodford N., Palepou M.-F. I., Mushtaq S., Metan G.,
Yakupogullari Y., Kocagoz S., Uzung O., Hascelik G. et Livermore D. M.
2008. Carbapenem-resistant Escherichia coli and Klebsiella pneumoniae
isolates from Turkey with OXA-48-like carbapénèmases and outer membrane
protein loss. International Journal of Antimicrobial Agents.
94 Guneysel O., Onur O., Erdede M., et Denizbasi A. 2009.
Trimethoprim/sulfamethoxazole resistance in urinary tract infections. The
Journal of Emergency Medicine; 36(4): 338-341.

Références Bibliographiques
93
H:
95 Hamze M., Dabboussi F. et Izard D. 2003. Enterobacterial susceptibility to
antibiotics in northern Lebanon (1998-2001). Cahiers d'études et de recherches
francophones / Santé; 13(2): 107-12.
96 Hermsen E.D., Sullivan C.J. et Rotschafer J.C. 2003. Polymyxins:
pharmacology, pharmacokinetics, pharmacodynamics, and clinical applications.
Infect Dis Clin North Am;17(3): 545-62.
97 Hirakata Y, Matsuda J, Miyazaki Y, Kamihira S, Kawakami S, Miyazawa Y, et
al. 2005. Regional variation in the prevalence of extended spectrum -
lactamase producing clinical isolates in the Asia–Pacific region (SENTRY
1998–2002). Diagn Microbiol Infect Dis; 52: 323-9.
98 Ho P.L., Tsang D.N.C., Que T.L., Ho M. et Yuen K.Y. 2000. Comparison of
screening methods for detection of extended–spectrum β–lactamases and their
prevalence among Esherichia coli and Klebsiella spp. in Hong Kong. APMIS,
108: 237-240.
99 Holstein A., Grillon A., Yzon L., Morange V., Baty G., Lartigue M.-F.,
Mereghetti L., Goudeau A. et Lanotte P. 2010. Prévalence des souches de
Escherichia coli et de Klebsiella pneumoniae productrices de -lactamases à
spectre étendu de type CTX-M à l’hôpital Bretonneau (CHRU de Tours)
Pathologie Biologie; 58 : 67-69.
100 Honoré S., Lascols C., Malin D., Targaouchi R., Cattoir V., Legrand P., Soussy
C.-J. et Cambau E. 2006. Émergence et diffusion chez les entérobactéries du
nouveau mécanisme de résistance plasmidique aux quinolones Qnr (résultats
hôpital Henri-Mondor 2002–2005). Pathologie Biologie; 54: 270-279.
I:
101 Iroha, I.R., Adikwu M.U., Amadi E.S., Aibinu I. et Esimone C.O. 2008.
Characterization of extended spectrum β-lactamase producing E. coli from
secondary and tertiary hospital in South Eastern Nigeria. Res. J. Microb; 3:
514-519.
J:

Références Bibliographiques
94
102 Andrews J.M. 2006. Determination of Minimum Inhibitory Concentrations.
Department of Microbiology, City Hospital NHS Trust, Birmingham B18 7QH.
p 1-19.
103 Joly B. et Reynaud A. 2007. Entérobactéries : systématique et méthodes de
diagnostic. Edition Techniques et Documentation. Paris. p 3-182.
K:
104 Kado C.I. et Liu S.T. 1981. Rapid procedure for detection and isolation
of large and small plasmids. J. Bacteriology; 145(3): 1365-1373.
105 Kahlmeter G. 2003. An international survey of the antimicrobial susceptibility
of pathogens from uncomplicated urinary tract infections: the ECOSENS
Project. J Antimicrob Chemother; 51: 69-76.
106 Kim J. H., Cho C. R., Um T. H. , Rhu J. Y., Kim E.S., Jeong J. W. et Lee H.R.
2007. Morganella Morganii Sepsis with Massive Hemolysis. J Korean Med
Sci; 22: 1082-1084.
L:
107 Lamnaouer D. 2002. Détermination des propriétés biologiques (activités
pharmacologiques et toxicologiques) des plantes médicinales et aromatiques du
PNT. Programme de l’UICN en Afrique du Nord: Phase III. Etat d’avancement.
p 3-7.
108 Larabi K., Masmoudi A. et Fendri C. 2003. Étude bactériologique et
phénotypes de résistance des germes responsables d’infections urinaires dans
un CHU de Tunis : à propos de 1930 cas. Médecine et maladies infectieuses;
33: 348-352.
109 Larpent J.P. 2000. Introduction à la nouvelle classification bactérienne. Les
principaux groupes bactériens. Ed. TEC & DOC. Paris p 280.
110 Lautenbach E., Weoltje K. et Malani P. 2010. Practical Healthcare
Epidemiology. University of Chicago Press. Third Edition. p 212-213.
111 Lavigne J-P. 2007. Effets des antibiotiques et mécanismes de résistance. MB7 :
Bactériologie B6 –Antibiotiques et résistance. Faculté de Médecine Montpellier
– Nîmes. p 1-3.

Références Bibliographiques
95
112 Lavigne J-P., Sotto A., Merle C., Jourdan J., Soussy C-J. et Sirot D. 2002.
Résistance enzymatique d’Escherichia coli aux -lactamines et prévalence en
clinique. Pathol Biol; 50: 388-93.
113 Leclercq M. 2006. Enterobacter sakazakii. Agence française de sécurité
sanitaire des aliments AFSSA. p 1-6.
114 Leotard S. et Negrin N. 2010. Epidémiologie des entérobactéries sécrétrices de
-lactamases à spectre étendu (E-BLSE) au centre hospitalier de Grasse (2005–
2008). Pathologie Biologie; 58 : 35-38.
115 Li J., Nation R.L., Milne R.W., Turnidge J. D. et Coulthard K. 2005.
Evaluation of colistin as an agent against multi-resistant Gram-negative
bacteria. Int. J. Antimicrob. Agents; 25: 11-25.
116 Li J., Nation R. L., Turnidge J. D., Milne R. W., Coulthard K., Rayner C. R. et
Paterson D. L. 2006. Colistin: the re-emerging antibiotic for multidrug-resistant
Gram-negative bacterial infections. Lancet Infect Dis; 6: 589-601.
117 Li X-Z., Mehrotra M., Ghimire S. et Adewoye L. 2007. -Lactam resistance
and -lactamases in bacteria of animal origin. Veterinary Microbiology; 121:
197-214.
118 Li X-Z. et Nikaido H. 2004. Efflux-mediated drug resistance in bacteria.
Drugs; 64:159-204.
119 Livermore D.M. et Woodford N. 2006. The -lactamase threat in
Enterobacteriaceae, Pseudomonas and Acinetobacter. Trends in Microbiology;
14:413-20.
M:
120 Mahamat A., Lavigne J.-P., Bouziges N., Daurès J.-P. et Sottob A. 2006. Profils
de résistance des souches urinaires de Proteus mirabilis de 1999 à 2005 au
CHU de Nîmes. Pathologie Biologie; 54 : 456-461.
121 Mahrouki S., Ben–Achour N., Chouchani C., Ben–Moussa M. et Belhadj O.
2009. Identification of plasmid-encoded extended spectrum -lactamases
produced by a clinical strain of Proteus mirabilis. Pathologie Biologie; 57: 55-
59.

Références Bibliographiques
96
122 Martin C. 2008. Urgences et infections: Guide du bon usage des antibiotiques,
antifongiques, antiviraux, antiseptiques. Editions Arnette. p : 41-42.
123 Martinez-Martinez L, Pascual A et Jacoby GA. 1998. Quinolone resistance
from a transferable plasmid. Lancet; 351: 797-9.
124 Masterton R.G. et Turner P.J. 2006. Trends in antimicrobial Susceptibility in
UK centers: the MYSTIC program (1997–2002). Int J Antimicrob Agents; 27:
69-72.
125 Materon I. C., Beharry Z., Huang W., Perez C. et Palzkill T. 2004. Analysis of
the context dependent sequence requirements of active site residues in the
metallo--lactamase IMP-1. J. Mol. Biol; 344: 653-663.
126 Matute A.J, Hak E., Schurink C. et al. 2004. Resistance of uropathogens in
symptomatic urinary tract infection in Leon, Nicaragua. Int J Antimicrob
Agents; 23: 506-9.
127 Meradi L., Djahoudi A., Abdi A., Bouchakour M., Perrier Gros Claude J.-D. et
Timinouni M. 2009. Résistance aux quinolones de types qnr, aac (60)-Ib-cr
chez les entérobactéries isolées à Annaba en Algérie. Pathologie Biologie
(Paris). p 1-6.
128 Messai L., Achour W. et Ben Hassen A. 2007. Profil épidémiologique des
entérobactéries isolées chez des patients neutropéniques. Pathologie Biologie;
55: 230-234.
129 Meyer A., Deiana J. et Bernard A. 2004. Cours de microbiologie générale:
avec problèmes et exercices corrigés. Editions 2 Doin. p 430.
130 Mirabaud M. I. 2003. Entérobactéries a -lactamases à spectre elargi en
pédiatrie en 1996. La Faculté de Médecine de l’Université de Genève. Thèse
de Doctorat en médecine.
131 Miriagou V., Carattoli A. et Fanning S. 2006. Antimicrobial resistance islands:
resistance gene clusters in Salmonella chromosome and plasmids. Microbes and
Infection; 8: 1923-1930.
132 Mkaouar D., Mahjoubi F., Mezghani S., Znazen A., Ktari S. et Hammami A.
2008. Étude de la résistance des entérobactéries aux céphalosporines de

Références Bibliographiques
97
troisième génération dans les hôpitaux de Sfax, Tunisie (1999–2005). Médecine
et maladies infectieuses; 38 :293-298.
133 Monnet D. et Freney J. 2000. Autres Entérobactéries. In : Freney J., Renaud F.,
Hansen W. et Bollet C. Précis de Bactériologie Clinique. ESKA, Paris, p 1215-
1237.
134 Morice V. 2003. Chapitre 7 - Entérobactéries et autres bacilles à Gram négatif
non exigeants. Sur le lien :
http://www.chups.jussieu.fr/polys/bacterio/bacterio/POLY.Chp.7.html.
N:
135 Naas T. 2003. Phénotype de résistance aux antibiotiques de la souche
Acinetobacter baumannii productrice de la -lactamase VEB-1. Service de
Bactériologie-Virologie-Hygiène (P. Nordmann), CHU de Bicêtre. Journal of
Clinical Microbiology; 41: 3542-7.
136 Nadmia H., Elotmani F., Talmi M., Zerouali K., Perrier-Gros-Claude J.D. et
Timinouni M. 2010. Profil de résistance aux antibiotiques des entérobactéries
uropathogènes communautaires à El Jadida (Maroc). Médecine et maladies
infectieuses; 40: 303-305.
137 Nauciel C. et Vildé J.-L. 2005. Bactériologie médicale, connaissance et pratique.
2ème
édition Masson. p
138 Nauciel C. 2000. Bactériologie médicale: connaissance et pratique. Edition
Masson. Paris. p 125-146.
139 Navon-Venezia S, Hammer-Munz O, Schwartz D, Turner D, Kuzmenko B. et
Carmeli Y. 2003. Occurrence and Phenotypic characteristics of Extended
Spectrum -lactamases among Members of the family Enterobacteriaceae at
the Tel-Aviv Medical center (Israel) and Evaluation of Diagnostic Tests. J Clin
Microbiol; 41 (1):155-8.
140 Nicaido H. 2000. "Crossing the envelope: how cephaslosporin sreach their targets."
Clin Microbiol Infect; 6: 22-26.
141 Nicolas-Chanoine MH, Blanco J, Leflon-Guibout V, Demarty R, Alonso MP,
Caniça MM, et al. 2008. Intercontinental emergence of Escherichia coli clone
O25: H4-ST131 producing CTX-M-15. J Antimicrob Chemother; 61(2):273-81.

Références Bibliographiques
98
142 Nijssen S., Florijnb A., Bontena M.J.M., Schmitz F.J., Verhoef J. et Fluit A.C.
2004. -lactam susceptibilities and prevalence of ESBL-producing isolates
among more than 5000 European Enterobacteriaceae isolates. International
Journal of Antimicrobial Agents; 24: 585-591.
143 Nordmann P. et Poirel L. 2002. Emerging carbapenemases in Gram-negative
aerobes. Clin Microbiol Infect; 8: 321-31.
144 Nordmann P. et Mammeri H. 2007. Résistance plasmidique aux quinolones.
Antibiotiques; 9: 246-53.
145 Nordmann P. Cuzon G. et Naas T. 2009. The real threat of Klebsiella
pneumoniae carbapenemase-producing bacteria. Lancet Infect Dis; 9: 28-36.
146 Nseir S., Ader F., Marquette C.-H. et Durocher A. 2005. Impact de l’utilisation
des fluoroquinolones sur l’émergence des bactéries multirésistantes. Pathologie
Biologie; 53: 470-475.
O:
147 O’Hara, C.M., Brenner, F.W. et Miller, M., 2000. Classification and
identification, and clinical significance of Proteus, Providencia, and
Morganella. Clinical Microbiology Reviews; 13: 534-546.
P:
148 Pagani L, Migliavacca R, Pallecchi L, Matti C, Giacobone E, Amicosante G, et
al. 2002. Emerging extended-spectrum -lactamases in Proteus mirabilis. J Clin
Microbiol; 40: 1549-52.
149 Patel J. B., Kamile Rasheed J. et Kitchel B. 2009. Carbapenemases in
Enterobacteriaceae: Activity, Epidemiology, and Laboratory Detection.
Clinical Microbiology Newsletter; 31: 8.
150 Pasteran Fernaldo G., Otaegui L., Guerriero L., Radice G., Maggiora R.,
Rapoport M., Faccone D., Di Matino A. et Galas M. 2008. Klebsiella
pneumonia Carbapenemase-2, Buenos Aires, Aegentina. EID Journal Home.
14.
151 Paterson D. L. 2006. Resistance in Gram-negative bacteria:
Enterobacteriaceae. Pittsburgh, Pennsylvania; 34(5): S20-S28.

Références Bibliographiques
99
152 Paterson D.L. 2006. Resistance in Gram-Negative Bacteria:
Enterobacteriaceae. The American Journal of Medicine; 119 (6A): S20-S28.
153 Paterson D.L.et Bonomo R.A. 2005. Extended-spectrum -lactamases: a
clinical update. Clin Microbiol Rev; 18: 657-86.
154 Peiffer B. 2000. Dysenterie bacillaire ou shigellose. Sur le lien :
http://www.liste-hygiene.org/SHIGELLA.html.
155 Péan Y., Goldstein F.W. et De Bels F. 2001. Évolution de la sensibilité et
épidémiologie de la résistance des entérobactéries de ville au cours des
enquêtes Vigil’Roc. Méd Mal Infect; 31: 609-21.
156 Peirano G. et Pitout J.D.D. 2010. Molecular epidemiology of Escherichia coli
producing CTX-M -lactamases: the worldwide emergence of clone ST131
O25: H4. International Journal of Antimicrobial Agents; 35: 316-321.
157 Pelmont J. 2005. Biodégradations et métabolismes: les bactéries pour les
technologies de l'environnement. Collection Grenoble sciences. L'Editeur :
EDP Sciences. p 124-125.
158 Penteado A.P., Castanheira M., Pignatari A.C.C., Guimarães T., Mamizuka
E.M. et Gales A.C. 2009. Dissemination of blaIMP-1-carrying integron In86
among Klebsiella pneumoniae isolates harboring a new trimethoprim resistance
gene dfr27. Diagnostic Microbiology and Infectious Disease; 63: 87-91.
159 Philippon A. 2001. Entérobactéries. Cours de Bactériologie Médicale Faculté
de Médecine COCHIN-PORT-ROYAL, Université PARIS V. Sur le lien :
www.microbe-edu.org/etudiant/entero.html.
160 Philippon A. 2005. ß-lactamases: généralités. Université Paris V - Faculté de
Médecine Descartes, Service de Bactériologie - 75014 PARIS. Sur le lien :
www.microbesedu.org/mecanisme/bla/generalites.html - 63k.
161 Philippon A., et Arlet G. 2006. -lactamases de bacilles à Gram négative : le
mouvement perpétuel ! Ann Biol Clin; 64 (1): 37-51.
162 Ploy M.-C., Gassama A., Chainier D. et Denis F. 2005. Les intégrons en tant
que support génétique de résistance aux antibiotiques. Integrons: an antibiotic
resistance gene capture system. Immuno-analyse & Biologie spécialisée; 20:
343-352.

Références Bibliographiques
100
163 Ploy M.-C., Lambert T., Gassama A. et Denis F. 2000. Place des intégrons
dans la dissémination de la résistance aux antibiotiques. Annales de Biologie
Clinique; 58(4), 439-44.
164 Popoff M.Y. 2001. Antigenic formulas of the Salmonella serovars, 8th ed.
WHO Collaborating center for reference and research on Salmonella, Institut
Pasteur, Paris, France.
165 Pourriat J.-L., Martin C. 2005. Principes de réanimation chirurgicale. Editions
Arnette 2. p 306-308.
Q:
R:
166 Rahal K. 2005. Standardisation de l’antibiogramme en médicine humaine à
l’échelle nationale. République Algérienne démocratique et populaire ministère
de la santé, de la population et de la reforme hospitalière. 4 eme
édition. p 40-42.
167 Robicsek A, Strahilevitz J et Jacoby GA. 2006. Fluoroquinolone modifying
enzyme: a new adaptation of a common aminoglycoside acetyltransferase. Nat
Med; 12: 83-8.
168 Rodriguez-Villalobos H. et Struelens M.-J. 2006. Résistance bactérienne par β-
lactamases à spectre étendu : implications pour le réanimateur. Réanimation;
15: 205-213.
169 Ruppé E. 2010. Épidémiologie des -lactamases à spectre élargi : l’avènement
des CTX-M. Antibiotiques;12: 3-16.
S:
170 Safraoui I. 2010. Epidémiologie de la résistance aux antibiotiques des bacilles à
Gram négatif non fermentants au CHU de Sidi Bel Abbes. Université de
Tlemcen. Thèse de magister.
171 Sambrook J. et Russel D.W. 2001. Molecular cloning. A Laboratory Manual.
Ed CSHL PRESS. VOL 1.
172 Saurina G., Quale J.M., Manikal V.M., Oydna E. et Landman D. 2000.
Antimicrobial resistance in Enterobacteriaceae in Brooklyn, NY: epidemiology
and relation to antibiotic usage patterns. J Antimicrob Chemother; 45: 895-8.

Références Bibliographiques
101
173 Scheftel J.-M. 2008. Entérobactéries. Sur le lien :
irespsdgs.free.fr/.../Escherichia%20coli%20cours%20%202008.ppt.
174 Schwalbe R., Steele-Moore L. et Goodwin A. C. 2007. Antimicrobial
Susceptibility Testing Protocols. Taylor & Francis Group. p 75-78.
175 Sekhri-Arafa N., Smati F., Scheftel J.M. et Meunier O. 2010. Marqueurs
épidémiologiques de souches de Klebsiella pneumoniae subsp pneumoniae
isolées au CHU de Constantine (ALGERIE). Rev. Microbiol. Ind. San et
Environn; 4(1): 82-98
176 Sekhsokh Y., Chadli M. et El Hamzaoui S.A. 2008. Fréquence et sensibilité aux
antibiotiques des bactéries isolées dans les urines. Médecine et maladies
infectieuses; 38 : 324-327.
177 Sekhsokh Y., Arsalane L., El Ouenass M., Doublali T., Bajjou T. et Lahlou
Amine I. 2007. Bactériémie à Serratia rubidaea. Médecine et maladies
infectieuses; 37 : 287-289.
178 Seck R. 2005. Résistance des souches d’Escherichia coli et de Klebseilla
pneumoniae isolées d’infections urinaires. Thèse de doctorat. Université Cheikh
Anta Diop de Dakar. Faculté de médecine, de pharmacie et d’Odonto-
Stomatologie. Département de pharmacie.
179 Seonghan K., Jinxin H., Romesh G., Junyoung K., Bokkwon L., et Boyle D. S.
2007. CTX-M Extended-spectrum β-Lactamases. Washington State.
Washington Department of Health, Shoreline, Washington, USA; and †Korea
Centers for Disease Control and Prevention, Seoul, Republic of Korea. EID
Journal Home; 13(3): 513-514.
180 Shah A.A., Hasan F., Ahmed S. et Abdul H. 2004. Characteristics,
epidemiology and clinical importance of emerging strains of Gram-negative
bacilli producing extended-spectrum β-lactamases. Microbiology Research
Laboratory, Department of Biological Sciences, Quaid-i-Azam University,
Islamabad, Pakistan. Research in Microbiology; 155: 409–421.
181 Sinan Bilgin S., Eren Olcay S. et Mehmet Demirtaş A. 2003. Complication of
felon caused by Morganella morgagni; case report. JOURNAL oF ANKARA
MEDICAL SCHOOL; 25(4): 199-204.

Références Bibliographiques
102
182 Singeton P. 2005. Bactériologie. Pour la médecine, la biologie et les
biotechnologies. 6eme
édition. Dunod. p 463.
183 Skurnik D. et Andremont A. 2006. Antibiothérapie sélectionnante: de la théorie
à la pratique. Réanimation; 15: 198–204.
184 Soude S.G.A.A. 2005. Bactéries isolées des hémocultures au laboratoire du
centre national hospitalier et universitaire Hubert Koutoukou Maga De
Cotonou. Thèse pour l'obtention du grade de Docteur en pharmacie diplôme
d'état. République du Mali, université de Bamako, faculté de Médecine, de
Pharmacie et d'Odonto-Stomatologie.
185 Sougakoff W. et Trystram D. 2003. Résistances aux β-lactamines. Service de
Bactériologie-Hygiène du CHU Pitié-Salpêtrière. Université Paris-VI.
186 Spanu T., Luzzaro F., Perilli M., Amicosante G., Toniolo A. et Fadda G. 2002.
Occurrence of extended-spectrum -lactamases in members of the family
Enterobacteriaceae in Italy: implications for resistance to -lactams and other
microbial drugs. Antimicrob Agents Chemother; 46: 196-202.
187 Stahl J.-P. 2005. La tigécycline, un nouvel antibiotique à l’étude chez l’homme.
Médecine et maladies infectieuses; 35: 62–67.
T:
188 Tan T. Y.,* Yong L. S., He J., Koh T. H. et Hsu L. Y. 2009. Evaluation of
Screening Methods To Detect Plasmid-Mediated AmpC in Escherichia coli,
Klebsiella pneumoniae, and Proteus mirabilis. Antimicrobial Agents And
Chemotherapy; 53(1): 146–149.
189 Thuresson A.-C., Kirsebom L.A. et Virtanen A. 2007. Inhibition of poly (A)
polymerase by aminoglycosides. Biochimie; 89: 1221-1227.
190 Tindall B.J., Grimont P.A.D., Garrity G.M., Euzéby J. 2005, Nomenclature and
taxonomy of the genus Salmonella, Int. J. Syst. Evol. Microbiol; 55: 521-524.
191 Tissot P. 2005. Antibiothérapie 1 & 2. I F S I Année 2004 – 2005. Pharmacien-
Hyères U/. sur le lien :
ifsi.chhyeres.fr/IMG/pdf/Antibiotherapie.pdf.
192 Touati A., benallaoua S., forte D., Madoux J., Brasme L. et De Champs C.
2006. First report of CTX-M-15 CTX-M-3 -lactamase among clinical isolates

Références Bibliographiques
103
of Enterobacteriacae in Béjaia, Algeria. International Journal of Antimicrobial
Agents; 27: 397-402.
193 Tran J.H., Jacoby G.A. et Hooper D.C. 2005. Interaction of the plasmid
encoded quinolone resistance protein QnrA with Escherichia coli DNA gyrase.
Antimicrob Agents Chemother; 49: 118-25.
194 Tremblay S. 2007. Etude moléculaire du recrutement des gènes de résistance
aux antibiotiques. Département de biochimie et microbiologie, Faculté des
sciences et de génie, Université LAVAL, Québec. Mémoire présenté pour
l'obtention du grade de maître en sciences.
195 Tribe, G.W. et Rood M.J., 2002. Providencia alcalifaciens in diarrhoeic dogs
and cats. The Veterinary Record; 150: 386-387.
196 Trigoso C. 2010. Carbapenemases: A Brief Review for Pediatric Infectious
Disease Specialists. Pediatr Infect Dis J; 29(1):68-70.
197 Tulkens P. et Spinewine A. 2002. Pharmacologie spéciale: les beta-lactames.
Université catholique de Louvain. Pharmacologie et pharmacothérapie des anti-
infectieux.
U:
V:
198 Van Bambeke F., Spiniwine A., Verbist L. et Tulkens P.M. 2002. Les
antibiotiques moins souvent et mieux: le rôle du pharmacien. Société
scientifiques des pharmaciens francophones (SSPF). p 1-7.
199 Vedel G. 1998. Lecture interprétative de l'antibiogramme (interprétation
phénotypique). In outil d'investigation microbiologique en réanimation. Edition
Arnette. p 183-188.
200 Vidon O. et Bourdin C. 2005. -lactamases à spectre étendu (BLSE). Intérêt
porté aux CTX-M. sur le lien : umr5558-mq1.univ-
lyon1.fr/.../BétaLactamasesSpectreEtendu?...BétaLactamases...
W:
201 Walsh C. 2000. Molecular mechanisms that confer antibacterial drug resistance.
Nature; 406: 775-81.

Références Bibliographiques
104
202 Weill F.-X. 2009. Salmonella: épidémiologie, typage et résistance aux
antibiotiques. Centre national de référence des Salmonella Laboratoire des
bactéries pathogènes émergentes. Institut Pasteur Elsevier Masson SAS. Revue
Francophone Des Laboratoires. Supplément Au N°409 : 25.
203 Wolff M., Joly-Guillou M.-L. et Pajot O. 2008. Le point sur les carabapénèmes.
Réanimation; 17: 242-250
204 Wu Q., Liu Q., Han L., Sun J. et Ni Y. 2010. Plasmid-mediated carbapenem-
hydrolyzing enzyme KPC-2 and ArmA 16S rRNA methylase conferring high-
level aminoglycoside resistance in carbapenem-resistant Enterobacter cloacae
in China. Diagnostic Microbiology and Infectious Disease; 66: 326-328.
205 Wu J.J., Ko W.C., Tsai S.H. et Yan J.J. 2007. Prevalence of plasmid-mediated
quinolones resistance determinants QnrA, QnrB, and QnrS among clinical
isolates of Enterobacter cloacae in a Taiwanese hospital; 51(4): 1223-1227.
X:
Y:
206 Yala D., Merad A.S., Mohamedi D. et Ouar Korich M.N. 2001. Classification
et mode d'action des antibiotiques. Médecine du Maghreb n°91.
Z:
207 Zhanel G., Hisanaga T., Laing N. et al. 2006. Antibiotic resistance in
Escherichia coli out patient urinary isolates: final results from the North
American Urinary Tract Infection Collaborative Alliance (NAUTICA). Int J
Antimicrob Agents; 27: 468–475.
208 Zogheib E. et Dupont H. 2005. Entérobactéries multirésistantes. Elsevier SAS.
P : 153-165. Sur le lien :
http://www.sfar.org/sfar_actu/ca05/html/ca05_13/ca05_13.htm#94992.
209 Zomahoun C. I. N. P. 2004. Evaluation de la sensibilité aux antibiotiques des
bactéries isolées des infections urinaires au laboratoire de bactériologie du
centre national hospitalier universitaire – HUBERT KOUTOUKOU MAGA
(C.N.H.U.-H.K.M.) DE COTONOU. Faculté de médecine de pharmacie et
d’odontostomatologie.

Annexes
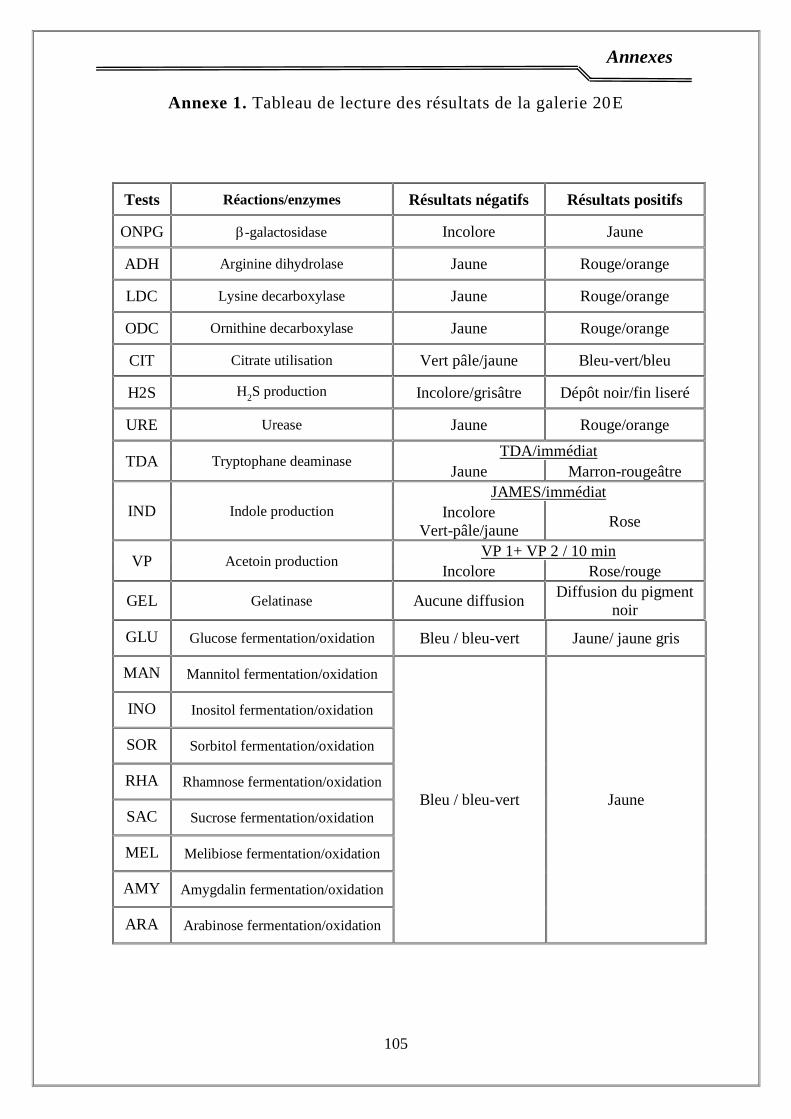
Annexes
105
Annexe 1. Tableau de lecture des résultats de la galerie 20E
Tests Réactions/enzymes Résultats négatifs Résultats positifs
ONPG -galactosidase Incolore Jaune
ADH Arginine dihydrolase Jaune Rouge/orange
LDC Lysine decarboxylase Jaune Rouge/orange
ODC Ornithine decarboxylase Jaune Rouge/orange
CIT Citrate utilisation Vert pâle/jaune Bleu-vert/bleu
H2S H2S production Incolore/grisâtre Dépôt noir/fin liseré
URE Urease Jaune Rouge/orange
TDA Tryptophane deaminase TDA/immédiat
Jaune Marron-rougeâtre
IND Indole production
JAMES/immédiat
Incolore
Vert-pâle/jaune Rose
VP Acetoin production VP 1+ VP 2 / 10 min
Incolore Rose/rouge
GEL Gelatinase Aucune diffusion Diffusion du pigment
noir
GLU Glucose fermentation/oxidation Bleu / bleu-vert Jaune/ jaune gris
MAN Mannitol fermentation/oxidation
Bleu / bleu-vert Jaune
INO Inositol fermentation/oxidation
SOR Sorbitol fermentation/oxidation
RHA Rhamnose fermentation/oxidation
SAC Sucrose fermentation/oxidation
MEL Melibiose fermentation/oxidation
AMY Amygdalin fermentation/oxidation
ARA Arabinose fermentation/oxidation

Annexes
106
Annexe 2. Tableau de lecture des résultats de la galerie 32E
No Test Réaction Résultats
Négative Positive
1.0 ODC Ornithine decarboxylase Jaune / jaune-orange Rouge / orange
1.1 ADH Arginine dihydrolase
1.2 LDC Lysine decarboxylase Jaune-vert Bleu-violet
1.3 URE Urease Jaune / jaune-orange Rose-violet
1.4 LARL L-Arabitol (Acidification)
Bleu / bleu-vert Jaune /
vert-jaune 1.5 GAT Galacturonate (Acidification)
1.6 5KG 5 Ketoglutarate (Acidifacation
1.7 LIP Lipase Incolore Bleu
1.8 RP Phenol red (Acidification) Rouge / orange Jaune
1.9 βGLU β-Glucosidase Incolore Jaune
1.A MAN Mannitol (Acidification)
Bleu /bleu-vert Jaune /
vert-jaune
1.B MAL Maltose (Acidification)
1.C ADO Adonitol (Acidification)
1.D PLE Palatinose (Acidification)
1.E βGUR β-Glucuronidase Incolore Jaune
1.F MNT Malonate Jaune / vert pâle Bleu-vert / bleu
0.0 IND 1
Indole production
JAMES / immédiat
Incolore /
jaune / beige Rose / rouge
0.1 βNAG N-acetyl-β-glucosaminidase Incolore Bleu
0.2 βGAL β-Galactosidase Incolore Jaune
0.3 GLU Glucose (Acidification)
Bleu / bleu-vert Jaune / vert-
jaune
0.4 SAC Saccharose/sucrose
(Acidification)
0.5 LARA L-Arabinose (Acidification)
0.6 DARL D-Arabitol (Acidification)
0.7 αGLU α-Glucosidase Incolore Jaune
0.8 αGAL α-Galactosidase
0.9 TRE Trehalose (Acidification)
Bleu / bleu-vert Jaune /
vert-jaune
0.A RHA Rhamnose (Acidification)
0.B INO Inositol (Acidification)
0.C CEL Cellobiose (Acidification)
0.D SOR Sorbitol (Acidification)
0.E αMAL α-Maltosidase Incolore /
Jaune très pâle Jaune
0.F AspA L-Aspartic acid arylamidase Incolore /
Jaune très pâle Jaune

Annexes
107
Annexe 3. Tableau d’identification du catalogue analytique API 20 E

Annexes
108
Annexe 3. Tableau d’identification du catalogue analytique API 20E

Annexes
109
Annexe 4. Tableau d’identification du catalogue analytique ID 32E

Annexes
110
Annexe 4. Tableau d’identification du catalogue analytique ID 32E

Annexes
111
Annexe 4. Tableau d’identification du catalogue analytique ID 32E

Annexes
112
Annexe 5. Tableau des Valeurs critiques des diamètres des zones d’ inhibition
et des CMI pour les entérobactéries (CA-SFM, 2010).
Familles
d’antibiotiques Antibiotiques testés Signe
Charge des
disques
Concentrations
critiques (mg/L)
Diamètres
critiques (mm)
S R S R
Ampicilline AM 10 g ≤4 > 8
≥19
<16
-lactamines
Amoxicilline AMX 25 μg ≤4 > 8
≥21
<16
Amoxicilline+
Ac.clavulanique AMC 20/10 μg ≤ 4/2 > 8/2 ≥21 <16
Ticarcilline TIC 75 μg
≤ 8 16 ≥24 <22
Ticarcilline+
Ac.clavulanique TCC 75/10 g ≤ 8/2 >16/2 ≥24 <22
Pipéracilline PIP 75μg ≤ 8 > 16 ≥20 <16
Pipéracilline+
tazobactum TZP 75/10μg ≤ 8/4 > 16/4 ≥ 21 < 17
Imipénème IPM 10 μg ≤ 2 > 8 ≥24 <17
Aztréonam ATM 30 g ≤ 1 > 8 ≥27 <21
Céfalotine CF 30 μg ≤ 8 > 32 ≥18 <12
Céfuroxime CXM 30 g ≤ 8 > 8 ≥22 <22
Céfoxitine FOX 30 μg ≤ 8 > 32 ≥22 <15
Latamoxef MOX 30 g ≤ 4 > 32 ≥23 <17
Céfotaxime CTX 30 μg ≤ 1 > 2 ≥26 <23
Céftriaxone CRO 30 g ≤ 1 > 2 ≥26 <23
Ceftazidime CAZ 30 μg ≤ 1 > 8 ≥26 <19
Céfépime FEP 30 μg ≤ 1 > 8 ≥24 <17
Cefpirome CPO 10 g ≤ 1 > 8 ≥24 <17
Aminosides
Kanamycine K 30UI ≤ 8 > 16 ≥17 <15
Tobramycine TM 30 μg ≤ 2 > 4 ≥ 18 < 16
Amikacine AN 30 μg ≤ 8 > 16 ≥17 <15
Gentamicine GM 15 μg (10UI) ≤ 2 > 4 ≥18 <16
Quinolones
Ac.nalidixique NA 30 μg
≤ 8 > 16 ≥20 <15
Ofloxacine OFX 5 μg ≤ 0.5 > 1 ≥25 <22
Ciprofloxacine CIP 5 μg
≤ 0.5 > 1 ≥25 <22
Divers
Colistine CS 50 g ≤ 2 > 2 ≥15 <15
Triméthoprime+
Sulfaméthoxazole SXT 1,25/23,75μg ≤ 2/38 >4/76 ≥16 <13
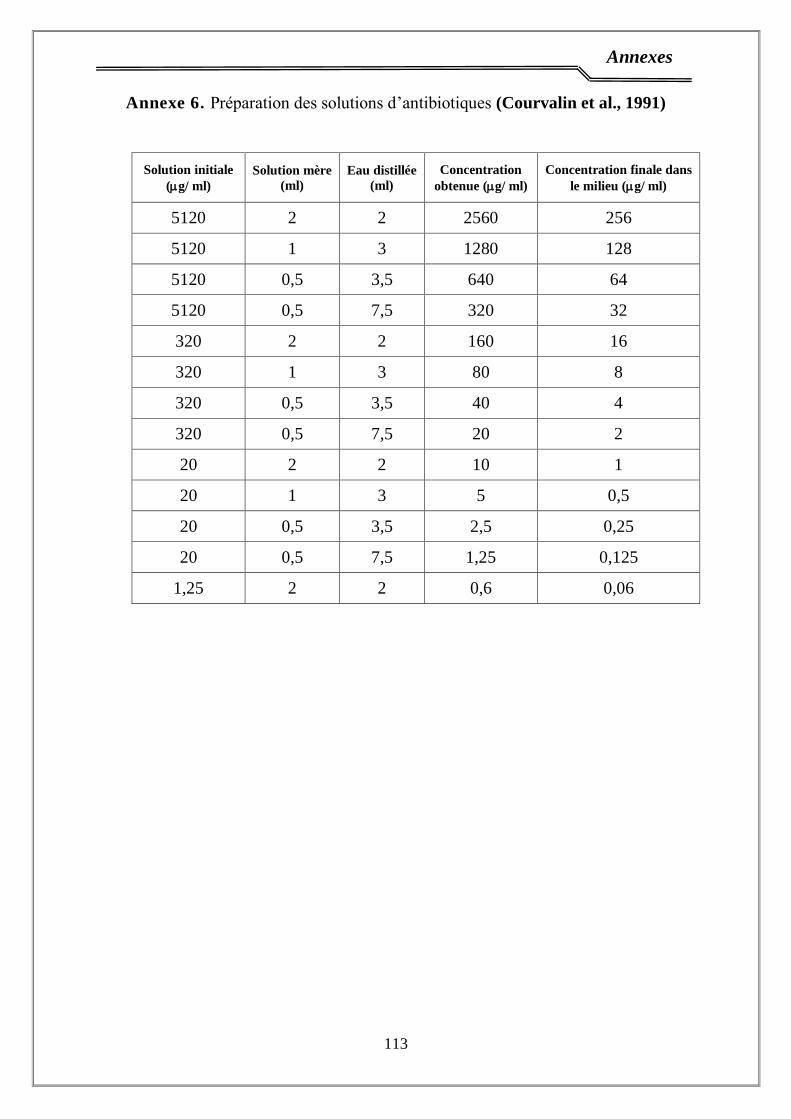
Annexes
113
Annexe 6. Préparation des solutions d’antibiotiques (Courvalin et al., 1991)
Solution initiale
(g/ ml)
Solution mère
(ml)
Eau distillée
(ml)
Concentration
obtenue (g/ ml)
Concentration finale dans
le milieu (g/ ml)
5120 2 2 2560 256
5120 1 3 1280 128
5120 0,5 3,5 640 64
5120 0,5 7,5 320 32
320 2 2 160 16
320 1 3 80 8
320 0,5 3,5 40 4
320 0,5 7,5 20 2
20 2 2 10 1
20 1 3 5 0,5
20 0,5 3,5 2,5 0,25
20 0,5 7,5 1,25 0,125
1,25 2 2 0,6 0,06

Annexes
114
Annexe 7. Solution et tampon pour l’extraction de l’ADN plasmidique
(Kado et Liu., 1981).
Solutions et tampons Composition
Solution de lyse
Tampon TE
Solution d’acétate de sodium
Tris (Sigma) 50 mM pH 12,6
SDS (Sigma) 3%
Tris (Sigma) 50 mM
EDTA (Sigma) 10 mM
pH 8
Acétate de sodium 3 M
pH 4,8

Annexes
115
Annexe 8. Solution et tampon pour l’électrophorèse de l’ADN plasmidique
(Sambrook et Russel, 2001).
Solution et tampon Composition
Tampon TBE 5 X
Tampon de charge
Tris base 54g
Acide borique 27,5g
EDTA 0,5 M 2 ml
Eau distillée 1 litre
Glycérol 3 ml
Bleu de bromophénol 75 mg
Eau distillée 7 ml

Annexes
116
Annexe 9. Résultats d’antibiogramme .
-lactamines :
Souche AM AMX AMC TIC TCC PIP TZP IMP CF CXM FOX MOX CRO CTX CAZ ATM FEP CPO Phénotype
S.E.1 7
R 7 R 20 I 7 R
20
R
22
S
28
S
35
S
22
S 26 S
27
S 30 S 34 S
37
S
29
S 32 S S
PASE
S.E.2 7
R 7 R 21 S 7 R 22 I
8
R
27
S
32
S 7 R 7 R
27
S 32 S
12
R
10
R
18
R 19 R R
BLSE
S.E.3 7
R 7 R 19 I 7 R
18
R
11
R
23
S
30
S
16
I 19 I
25
S 27 S 28 S
28
S
26
S 28 S S
PASE
S.E.4 21
S 20 I 23 S
29
S
27
S
30
S
25
S
33
S
23
S 30 S
25
S 30 S 30 S
34
S
29
S 33 S S
Sauvage
S.E.5 7
R 7 R 20 I 7 R
20
R
22
S
25
S
30
S
19
S 26 S
26
S 30 S 26 S
31
S
26
S 30 S S
PASE
S.E.6 7
R 7 R 16 I 7 R
15
R
7
R
22
S
25
S
13
I 7 R
25
S 28 S 22 I
28
S 25 I 28 S S
PASE
S.E.7 19
S 19 I 23 S
27
S
24
S
25
S
25
S
29
S
22
S 14 R
25
S 30 S 26 S
30
S
30
S 28 S S
Sauvage
S.E.8 23
S 19 I 23 S
25
S
28
S
28
S
26
S
30
S
22
S 10 R
27
S 30 S 30 S
30
S
34
S 32 S S
Sauvage
S.E.9 24
S 20 I 24 S
29
S
29
S
30
S
28
S
32
S
20
S 10 R
34
S 35 S 30 S
34
S
32
S 34 S S
Sauvage
S.E.10 7
R 7 R 20 I 7 R
19
R
13
R
25
S S
15
I 7 R
26
S 29 S 25 S
30
S
26
S 31 S S
PASE
S.E.11 7
R 7 R 18 I 7 R
15
R
9
R
23
S S
11
R 7 R
25
S 29 S 25 S
32
S
26
S 32 S S
PASE
S.E.12 7
R 7 R 17 I 7 R
13
R
10
R 17 I S
12
I 7 R
24
S 23 S 25 S
26
S
26
S 27 S S
PASE
S.E.13 7
R 7 R 19 I 7 R
17
R
11
R
23
S S
14
I 9 R
29
S 29 S 26 S
30
S
30
S 35 S S
PASE
S.E.14 7
R 7 R 21 S
11
R 9 R
12
R
26
S
31
S
19
S 28 S S 34 S 32 S
35
S
30
S S
36
S PASE
S.E.15 7
R 7 R 9 R
18
R
16
R
20
S
21
S
28
S 7 R 17 R R 22 I
21
R
22
R
18
R I
27
S CASE
S.E.16 7
R 7 R 18 I 7 R
10
R
10
R
22
S
30
S 7 R 16 R S 25 S
19
R
17
R
10
R R 23 I
BLSE
S.E.17 21
S 25 S 24 S
28
S
26
S
29
S
28
S
31
S
21
S 25 S S 30 S 31 S
31
S
29
S S
33
S Sauvage
S.E.18 20
S 21 S 21 S
25
S
24
S
25
S
24
S
28
S
21
S 26 S S 29 S 26 S
33
S
28
S S
31
S Sauvage
S.E.19 7
R 7 R 18 I 7 R 22 I
12
R
25
S
24
S
14
I 21 R S 24 S 31 S 23 I 22 I S
29
S PASE
S.E.20 7
R 7 R 17 I 7 R
24
S
18
I
25
S
30
S
19
S 23 S
26
S 29 S 29 S
30
S
26
S S
31
S PASE
S.E.21 7
R 7 R 18 I 7 R
17
R
16
I 18 I
24
S
17
I 18 R S 23 S
21
R
22
R 20 I I 23 I
PASE
S.E.22 7
R 7 R 7 R
30
S
32
S
28
S
34
S
35
S
24
S 30 S S 34 S 34 S
34
S
35
S S
34
S Sauvage
S.E.23 7
R 7 R 16 I 7 R
10
R
7
R
27
S
26
S 7 R 7 R S 27 S 7 R 7 R
11
R R
15
R
BLSE+
CASE
S.E.24 20
S 19 I 18 I
25
S
25
S
23
S
23
S
28
S
20
S 22 S S 24 S 26 S
26
S
26
S S
30
S Sauvage
S.E.25 7
R 7 R 15 R 7 R
21
R
12
R
23
S
28
S
20
S 24 S S 29 S 26 S
30
S
26
S S
26
S PASE
S.E.26 7
R 7 R 11 R 7 R
12
R
7
R 19 I
28
S 7 R 7 R S 24 S 9 R 8 R
12
R R
16
R
BLSE+
CASE
S.E.27 22
S 25 S 22 S
28
S
28
S
27
S
25
S
30
S
22
S 25 S S 30 S 26 S
30
S
26
S S
32
S Sauvage
S.E.28 7
R 7 R 15 R 9 R
21
R
15
R
23
S
28
S
17
I 20 R S 24 S 26 S
26
S
26
S S
27
S PASE
S.E.29 7
R 7 R 16 I 7 R 23 I
13
R
25
S
28
S
19
S 24 S S 31 S 34 S
32
S
30
S S S
32
S PASE
S.E.30 7
R 7 R 15 R 7 R
21
R
18
I
25
S
28
S
19
S 25 S S 30 S 30 S
31
S
29
S S S
29
S PASE
S.E.31 20
S 23 S 22 S
25
S
25
S
25
S
23
S
29
S
20
S 22 S S 28 S 26 S
28
S 25 I S S
30
S Sauvage
S.E.32 20
S 22 S 24 S
28
S
27
S
26
S
24
S
27
S
21
S 26 S S 30 S 29 S
31
S
28
S S S
30
S Sauvage

Annexes
117
S.E33 7
R 7 R 15 R 7 R
21
R
18
I
25
S
28
S
19
S 24 S S 29 S 28 S
29
S
28
S S S
29
S PASE
S.E.34 7
R 7 R 20 I 7 R 23 I
18
I
25
S
30
S
24
S 28 S S 31 S 30 S
34
S
30
S S S
33
S PASE
S.E.35 7
R 7 R 15 R 7 R
21
R
15
R
25
S
29
S
20
S 25 S S 30 S 28 S
30
S
27
S S S
30
S PASE
S.E.36 18
I 20 I 24 S
28
S
24
S
26
S
23
S
30
S
20
S 23 S S 29 S 26 S
30
S
26
S S S
26
S Sauvage
S.E.37 7
R 7 R 17 I 7 R 23 I
18
I
28
S
28
S
22
S 24 S S 30 S 31 S
30
S
28
S S S
32
S PASE
S.E.38 7
R 7 R 18 I 7 R
18
R
7
R
26
S
32
S 7 R 7 R S 30 S 8 R 8 R
18
R R R
11
R BLSE
S.E.39 7
R 7 R 15 R 7 R
15
R
12
R
23
S
32
S
16
I 26 S S 30 S 30 S
30
S
30
S S S
30
S PASE
S.E.40 7
R 7 R 16 I 7 R
21
R
18
I
27
S
29
S
22
S 32 S S 31 S 32 S
30
S
30
S S S
32
S PASE
S.E.41 17
I 22 S S
28
S
26
S S
27
S
32
S
18
S 32 S S 34S 30 S
30
S
33
S S S
30
S Sauvage
S.E.42 25
S 28 S S
30
S
32
S S 29S
34
S
24
S 28 S S 34 S 31 S
30
S
32
S S S
30
S Sauvage
S.E.43 18
I 19 I S
29
S
31
S S
28
S
28
S
18
S 28 S S 34 S 30 S
30
S
30
S S S
30
S Sauvage
S.E.44 18
I 20 I 16 I
30
S
31
S S
29
S
31
S
18
S 26 S S 34 S 29 S
30
S
30
S S S
30
S Sauvage
S.E.45 22
S 21 S 17 I
29
S
28
S S
29
S
30
S
19
S 27 S S 31 S 28 S
30
S
28
S S S
30
S Sauvage
S.E.46 19
S 23 S S
28
S
28
S S
28
S
29
S
18
S 26 S S 30 S 28 S
30
S
28
S S S
30
S Sauvage
S.E.47 7
R 7 R 7 R 7 R 9 R
7
R
25
S
32
S 7 R 7 R S 34 S 9 R
12
R 25 I R R
16
R
BLSE+
CASE
S.E.48 32
S 21 S 26 S
24
S
30
S
30
S
30
S
30
S
30
S 30 S S 34 S 30 S
34
S
30
S S S
34
S Sauvage
S.E.49 7
R 7 R 7 R 7 R 9 R
7
R
25
S
31
S 7 R 7 R S 30 S 9 R
11
R 22 I R R
15
R
BLSE+
CASE
S.E.50 7
R 7 R 16 I 7 R
18
R S
29
S
31
S
21
S 28 S S 32 S 30 S
30
S
31
S S S
30
S PASE
S.E.51 7
R 7 R 7 R 7 R 9 R
7
R
28
S
32
S 7 R 7 R I 29 S 7 R 7 R
16
R R R 7 R
BLSE+
CASE
S.E.52 19
S 24 S 22 S
30
S
31
S S
29
S
31
S
18
S 26 S S 34 S 29 S
30
S
30
S S S
30
S Sauvage
S.Kp.1 7
R 7 R 10 R 7 R 7 R
7
R 20 I
33
S 7 R 7 R
26
S 27 S 7 R 7 R 7 R 7 R R R BLSE
S.Kp.2 7
R 7 R 17 I 7 R
18
R
7
R 20 I
24
S 7 R 7 R 21 I 24 S 8 R 7 R
15
R 15 R R R
BLSE+
CASE
S.Kp.3 7
R 7 R 13 R 7 R
16
R
7
R
21
S
26
S 7 R 7 R
25
S 26 S
11
R
12
R 21 I 20 R R R BLSE
S.Kp.4 7
R 7 R 19 I 7 R
19
R
7
R
25
S
32
S 7 R 7 R
26
S 31 S
15
R
15
R 23 I 27 S R R BLSE
S.Kp.5 7
R 7 R 14 R 7 R
15
R
15
R 18 I
29
S
17
I 28 S
25
S 29 S 28 I
30
S
27
S 30 S S S PASE
S.Kp.6 7
R 7 R 13 R 7 R
12
R
7
R 18 I
25
S 7 R 7 R
22
S 25 S 7 R 7 R
15
R 15 R R R
BLSE+
CASE
S.Kp.7 7
R 7 R 14 R 7 R
14
R
7
R
21
S
27
S 7 R 7 R
24
S 26 S
12
R
10
R
16
R 19 R R R BLSE
S.Kp.8 7
R 7 R 13 R 7 R
13
R
7
R 19 I
30
S 7 R 7 R
23
S 25 S
10
R 9 R
14
R 16 R R R BLSE
S.Kp.9 7
R 7 R 15 R 7 R
13
R
7
R
21
S
28
S 7 R 7 R
24
S 29 S
11
R
10
R
15
R 7 R R R BLSE
S.Kp.10 7
R 7 R 15 R 7 R
14
R
15
R
21
S
29
S
21
S 23 S
23
S 26 S 30 S
27
S
26
S 30 S S S PASE
S.Kp.11 7
R 7 R 16 I 7 R
14
R
7
R
22
S
26
S 7 R 7 R
24
S 29 S
18
R
12
R 19 I 27 S R R BLSE
S.Kp.12 10
R 9 R 23 S
14
R
24
S
22
S
24
S
25
S
23
S 25 S
24
S 28 S 30 S
30
S
26
S 30 S S S Sauvage
S.Kp.13 7
R 7 R 19 I
11
R
19
R
20
S
21
S
24
S
18
S 10 R
24
S 25 S
16
R
26
S
26
S 32 S S S Sauvage
S.Kp.14 7
R 7 R 14 R 7 R
14
R
7
R 19 I S 7 R 7 R 21 I 25 S 7 R 7 R 7 R 8 R
12
R R
BLSE+
CASE
S.Kp.15 7
R 7 R 14 R 7 R 9 R
7
R 20 I S 7 R 11 R
26
S 30 S
10
R 7 R
15
R 15 R R R BLSE
S.Kp.16 7
R 7 R 15 R 7 R
10
R
7
R 19 I S 7 R 7 R 21 I 25 S 7 R 7 R 7 R 7 R R R
BLSE+
CASE
S.Kp.17 7
R 7 R 14 R 7 R
12
R
7
R 19 I S 7 R 7 R
22
S 27 S
15
R 8 R
15
R 19 R R R BLSE

Annexes
118
S.Kp.18 7
R 7 R 20 I 7 R
21
R
7
R
24
S
27
S 7 R 7 R S 30 S
13
R
16
R 25 I R
16
R R BLSE
S.Kp.19 15
R 15 R 24 S
21
R
29
S
28
S
29
S
29
S
28
S 32 S S 34 S 30 S
38
S
32
S S
35
S S Sauvage
S.Kp.20 7
R 7 R 7 R 7 R
16
R
7
R 18 I
27
S 7 R 7 R S 26 S 9 R
10
R
17
R R
16
R R BLSE
S.Kp.21 7
R 7 R 21 S 7 R
20
R
7
R
25
S
40
S 7 R 7 R S 35 S 7 R 7 R 7 R R
15
R R BLSE
S.Kp.22 7
R 7 R 14 R 7 R
11
R
8
R 20 I
24
S 7 R 7 R S 25 S 8 R
10
R
17
R R
16
R R BLSE
S.Kp.23 7
R 7 R 7 R 7 R
14
R
7
R 19 I
26
S 7 R 7 R S 26 S 7 R 7 R
11
R R 9 R R
BLSE+
CASE
S.Kp.24 7
R 7 R R 7 R
20
R
13
R
24
S
28
S
20
S 26 S S 30 S 29 S
32
S
28
S S
30
S S PASE
S.Kp.25 7
R 7 R R 7 R
15
R
7
R
24
S
29
S 7 R 7 R S 26 S 8 R 8 R
16
R R R
10
R
BLSE+
CASE
S.Kp.26 7
R 7 R R 7 R
13
R
7
R 20 I
28
S 7 R 7 R S 28 S 9 R 9 R
13
R R R
10
R
BLSE+
CASE
S.Kp.27 7
R 7 R R 7 R
16
R
7
R 17 I
26
S 7 R 7 R S 20 I 8 R
11
R
15
R R R
12
R BLSE
S.Kp.28 7
R 7 R 7 R 7 R 9 R R
21
S
29
S 7 R 7 R S S 8 R 8 R
18
R R R R BLSE
S.Kp.29 7
R 7 R 7 R 7 R
10
R R
24
S
27
S 7 R 7 R S 28 S 7 R 8 R
18
R R R 8 R
BLSE+
CASE
S.Kp.30 7
R 7 R 7 R 7 R 8 R R
21
S
30
S 7 R 7 R S 26 S 7 R 7 R 9 R R R 7 R
BLSE+
CASE
S.Pm.1 7
R 7 R 23 S
18
R
28
S
30
S
35
S
31
S
25
S 30 S
30
S 30 S 43 S
30
S
30
S 30 S S
PASE
S.Pm.2 7
R 7 R 20 I 7 R 22 I
13
R
24
S
24
S 8 R 9 R
24
S 29 S
20
R
15
R 8 R 22 I I
BLSE
S.Pm.3 7
R 7 R 9 R 7 R
19
R
12
R
24
S 23 I 7 R 7 R 20 I 28 S
10
R
10
R
16
R 25 I I
BLSE+
CASE
S.Pm.4 7
R 7 R 22 S 7 R
21
R
14
R
32
S
31
S 9 R 7 R
30
S 34 S 25 S
16
R
10
R 34 S R
BLSE
S.Pm.5 7
R 7 R 36 S
18
R
28
S
30
S
40
S
30
S
36
S 9 R
30
S 30 S 50 S
30
S
34
S 34 S S
PASE
S.Pm.6 7
R 8 R 22 S
25
S
27
S
27
S
24
S
24
S
24
S 15 R
26
S 29 S 37 S
35
S
29
S 26 I S
PASE
S.Pm.7 20
S 21 S 22 S S
25
S
25
S
24
S S
19
S 13 R
23
S 25 S 33 S
32
S
26
S 31 S S
Sauvage
S.Pm.8 11
R 7 R 20 I
20
R
25
S
25
S
25
S S
19
S 10 R
24
S 25 S 26 S
30
S
26
S 30 S S
PASE
S.Pm.9 7
R 7 R 7 R
21
R
20
R
11
R 20 I 21 I 7 R 10 R S 26 S
21
R
18
R
14
R S
27
S CASE
S.Pm.10 23
S 25 S 24 S
25
S
25
S
24
S
24
S
24
S
24
S 26 S S 26 S 29 S
31
S
26
S S
28
S Sauvage
S.Pm.11 7
R 7 R 23 S 7 R
20
R
15
R
28
S
24
S
20
S 28 S S 28 S 26 S
26
S
29
S S
31
S PASE
S.Pm.12 7
R 7 R 10R 7 R
20
R
12
R
25
S
24
S 9 R 13 R S 29 S
22
R
20
R 23 I S
28
S CASE
S.Pm.13 7
R 8 R 12 R
22
I
25
S
18
I
23
S 22 I
22
S 26 S S 28 S 34 S
30
S
30
S S
26
S PASE
S.Pm.14 3O
S 30 S 30 S
32
S
32
S
31
S
30
S
34
S
30
S 28 S S 30 S 34 S
30
S
30
S S
34
S Sauvage
S.Pm.15 26
S 29 S S
30
S
29
S
30
S S
29
S
28
S 28 S S 30 S 34 S S
30
S S
30
S Sauvage
S.Pm.16 26
S 35 S S
30
S
30
S S
32
S
40
S
28
S 30 S S 32 S 30 S
32
S
31
S S
30
S Sauvage
S.Pm.17 26
S 31 S S
29
S
34
S S
30
S
30
S
30
S 31 S S 34 S 30 S
32
S
30
S S
32
S Sauvage
S.Pm.18 28
S 33 S S
35
S
34
S S
30
S
29
S 30S 31 S S 32S 31 S
32
S
30
S S
32
S Sauvage
S.Pm.19 7
R 7 R 19 I
11
R
27
S
19
I
23
S
29
S 7 R 8 R I 29 S
11
R
11
R 9 R R 9 R
BLSE
S.Pv.1 7
R 7 R 20 I
15
R
24
S
15
R
24
S
27
S 7 R 12 R
24
S 34 S
16
R
18
R
12
R 22 I
14
R BLSE
S.Pv.2 7
R 7 R 19 I S
21
R
25
S
25
S S 7 R 7 R 15 I 23 S 28 S
22
R 23 I 27 S
28
S Sauvage
S.Pv.3 7
R 7 R 18 I S 23 I
24
S
27
S S 7 R 7 R 20 I 26 S 30 S
30
S
28
S 35 S
28
S Sauvage
S.Pv.4 7
R 7 R 7 R 7 R
16
R
12
R 20 I
24
S 7 R 12 R S 26 S
17
R
17
R
16
R S
25
S CASE
S.Pv.5 7
R 7 R 7 R
29
S
30
S
31
S
34
S
32
S 7 R 16 R 15 I 36 S 34 S
35
S
33
S 28 S
34
S Sauvage
Annexes
119
S.Pv.6 7
R 7 R 20 I
24
S
30
S
24
S
27
S
23
S 7 R 7 R 20 I 27 S 30 S
31
S
29
S 30 S
28
S Sauvage
S.Pv.7 16
I 16 I 19 I
32
S
28
S
24
S
30
S
32
S
14
I 30 S 19 I 34 S 30 S
30
S
32
S 31 S
30
S
30
S Sauvage
S.Pp.1 7
R 7 R 19 I
15
R
20
R
23
S
23
S S 7 R 7 R 16 I 23 S 27 S
21
R 20 I 23 I
28
S Sauvage
S.Ec.1 7
R 7 R 7 R
30
S
27
S
28
S
28
S
28
S 7 R 26 S 7 R 30 S 30 S
36
S
28
S 30 S S S Sauvage
S.Ec.2 7
R 7 R 8 R 7 R
16
R
7
R 18 I
24
S 7 R 7 R 7 R 25 S 7 R 7 R
12
R 12 R R R BLSE
S.Ec.3 7
R 7 R 7 R 7 R
13
R
7
R 20 I
25
S 7 R 7 R 7 R 29 S
13
R 9 R
12
R 16 R R R BLSE
S.Ec.4 7
R 7 R 7 R 9 R 7 R
20
S
23
S 20 I 7 R 7 R 7 R 18 I 26 S
22
R 20 I 26 I S S CASE
S.Ec.5 7
R 7 R 7 R 7 R
10
R
9
R
21
S
25
S 7 R 7 R 9 R 20 I
11
R
17
R 9 R 12 R I I BLSE
S.Ec.6 7
R 7 R 7 R 7 R 9 R
7
R 17 I
24
S 7 R 7 R 7 R 14 R 7 R 7 R
14
R 17 R R R BLSE
S.Ec.7 7
R 7 R 7 R
24
S
21
R
21
S
30
S
24
S 7 R 7 R 7 R 24 S
20
R
29
S
32
S 28 S S S Sauvage
S.Ec.8 7
R 7 R 7 R 7 R 7 R
7
R 20 I
24
S 7 R 7 R 8 R 16 R 7 R 7 R
11
R 10 R R R
BLSE+
CASE
S.Ec.9 7
R 7 R 7 R 7 R 9 R
10
R
25
S
27
S 7 R 7 R 8 R 25 S
15
R
20
R
11
R 12 R I I BLSE
S.Ec.10 7
R 7 R 7 R 7 R
11
R
7
R 20 I
24
S 7 R 7 R 9 R 27 S
15
R
10
R
16
R 20 R R R BLSE
S.Ec.11 7
R 7 R 7 R
22
I
20
R
25
S
23
S S 7 R 7 R 7 R 26 S
21
R 7 R 25 I 26 I S S CASE
S.Ec.12 7
R 7 R 10 R 7 R
15
R
9
R 19 I 20 I 7 R 7 R 18 I 14 R
19
R
10
R
16
R 21 I R R
BLSE+
CASE
S.Ec.13 9
R 9 R 7 R
25
S
27
S
24
S
26
S
25
S 7 R 24 S R 30 S 26 S
27
S
26
S S
30
S S Sauvage
S.Ec.14 7
R 7 R 7 R 7 R
14
R
7
R
16
R 22 I 7 R 7 R R 27 S 7 R 7 R
13
R R
12
R R CASE
S.Ec.15 7
R 7 R 10 R 7 R
16
R
7
R 19 I
25
S 7 R 7 R I 23 S 7 R 7 R
11
R R
16
R R
BLSE+
CASE
S.Ec.16 7
R 7 R 7 R
24
S
24
S
22
S
22
S
24
S 7 R 22 S R 24 S 26 S
26
S
26
S S
28
S S Sauvage
S.Ec.17 7
R 7 R R 7 R
12
R
7
R
21
S
28
S 7 R 7 R R 20 I
14
R
15
R 9 R R I 22 I BLSE
S.Ec.18 7
R 7 R R
26
S
25
S
23
S
24
S
28
S 7 R 23 S R 29 S 26 S
29
S
26
S S S
28
S Sauvage
S.Ec.19 7
R 7 R R
29
S
30
S
22
S
25
S
30
S 7 R 22 S R 34 S 30 S
34
S
34
S S S
34
S Sauvage
S.Ec.20 7
R 7 R R
27
S
30
S
25
S
25
S
25
S 7 R 27 S R 34 S 30 S
34
S
34
S S S
30
S Sauvage
S.Ec.21 7
R 7 R R
25
S 22 I
24
S
23
S
24
S 7 R 22 S R 26 S 26 S
26
S
26
S S S
25
S Sauvage
S.Ec.22 7
R 7 R R 7 R
13
R
7
R
25
S
26
S 7 R 12 R R 25 S
13
R
14
R
13
R R I
24
S
BLSE+
CASE
S.Ec.23 7
R 7 R R 7 R
12
R
7
R
24
S
29
S 7 R 13 R R 28 S
14
R
15
R
12
R R I
24
S BLSE
S.Ec.24 7
R 7 R R 7 R 9 R
7
R
22
S
27
S 7 R 7 R R 21 I
10
R
11
R 7 R R I 21 I BLSE
S.Ec.25 7
R 7 R 7 R
24
S
24
S
24
S
31
S
34
S 7 R 28 S R 30 S 30 S
32
S
30
S S S
34
S Sauvage
S.Ec.26 7
R 7 R R 7 R
14
R
7
R 20 I
24
S 7 R 7 R R 26 S 7 R 7 R
13
R R R 7 R BLSE
S.Ps.1 I R R S S S S S R S S S S S I S S
Sauvage
S.Mm.1 R R R R S S S S R R I S S S S S S
PASE
S.CF.1 R R R S R S I S R S R S R I I I I
Sauvage
S.SS.1 R R R R R R S S I S S S S S S S S
PASE
S.Sel.spp. R R R R R R S S S S S S S S S S S
PASE

Annexes
120
Autres classe d’antibiotiques:
Souche Aminosides Quinolones
CS SXT TM GM AN K NA OFX CIP
S.E.1 22 S 26 S 25 S 26 S 26 S 31 S 35 S 17 S 30 S
S.E.2 16 I 10 R 22 S 22 S 23 S 33 S 34 S 16 S 29 S
S.E.3 20 S 21 S 20 S 20 S 21 S 30 S 30 S 16 S 7 R
S.E.4 20 S 23 S 21 S 21 S 24 S 32 S 36 S 16 S 26 S
S.E.5 11 R 11 R 23 S 19 S 7 R 7 R 7 R 17 S 7 R
S.E.6 17 I 20 S 20 S 20 S 20 S 30 S 33 S 15 S 24 S
S.E.7 19 S 20 S 20 S 20 S 20 S 27 S 30 S 15 S 22 S
S.E.8 20 S 25 S 21 S 20 S 23 S 30 S 30 S 16 S 28 S
S.E.9 20 S 25 S 23 S 20 S 27 S 34 S 34 S 17 S 28 S
S.E.10 21 S 24 S 21 S 21 S 7 R 9 R 12 R 15 S 7 R
S.E.11 19 S 22 S 21 S 19 S 7 R 7 R 7 R 15 S 7 R
S.E.12 16 I 17 I 18 S 16 I 21 S 22 I 27 S 15 S 7 R
S.E.13 20 S 22 S 20 S 19 S 23 S 28 S 33 S 15 S 25 S
S.E.14 21 S 24 S 23 S 24 S 7 R 21 R 25 S 16 S 7 R
S.E.15 20 S 21 S 19 S 24 S 7 R 22 I 24 I 16 S 7 R
S.E.16 11 R 10 R 22 S 34 S 25 S 32 S 35 S 15 S 7 R
S.E.17 22 S 26 S 22 S 30 S 24 S 35 S 35 S 16 S 28 S
S.E.18 19 S 23 S 20 S 27 S 22 S 26 S 32 S 15 S 21 S
S.E.19 22 S 24 S 20 S 8 R 7 R 9 R 10 R 13 R 7 R
S.E.20 19 S 22 S 21 S 29 S 24 S 28 S 26 S 15 S 24 S
S.E.21 14 R 19 S 19 S 26 S 19 I 27 S 31 S 13 R 15 I
S.E.22 23 S 32 S 30 S 34 S 30 S 34 S 30 S 20 S 25 S
S.E.23 7 R 7 R 7 R 27 S 26 S 27 S 35 S 15 S 7 R
S.E.24 19 S 22 S 19 S 28 S 18 I 26 S 30 S S 20 S
S.E.25 20 S 23 S 21 S 25 S 21 S 25 S 28 S S 7 R
S.E.26 9 R 11 R 19 S 7 R 7 R 7 R 7 R S 7 R
S.E.27 22 S 25 S 22 S S 21 S 27 S 30 S S 23 S
S.E.28 20 S 21 S S R 7 R 15 R 21 R S 7 R
S.E.29 24 S 27 S 24 S R 7 R 8 R 10 R S 7 R
S.E.30 21 S 23 S S
22 S 29 S 34 S S 7 R
S.E.31 20 S 23 S S
21 S 28 S 33 S S 23 S
S.E.32 21 S 24 S S
22 S 31 S 36 S S 27 S
S.E33 20 S 23 S S
7 R 10 R 14 R S 7 R
S.E.34 22 S 25 S S
7 R 12 R 15 R S 7 R
S.E.35 20 S 23 S S
7 R 20 R 25 S S 7 R
S.E.36 20 S 24 S S
20 S 26 S 30 S S 22 S
S.E.37 23 S 25 S S
23 S 30 S 36 S S 7 R
S.E.38 16 I 12 R S
25 S 30 S 36 S S 7 R
S.E.39 19 S 22 S S
24 S 33 S 34 S S 7 R
S.E.40 20 S 21 S S
11 R 26 S 32 S S 7 R
S.E.41 18 S 20 S S
24 S 31 S 33 S S 24 S
S.E.42 18S 22 S S
27 S 35 S 38 S S 27 S
S.E.43 18 S 21 S S
26 S 31 S 36 S S 25 S
S.E.44 19 S 22 S S
25 S 32 S 36 S S 25 S
S.E.45 20 S 24 S S
7 R 9 R 14 R S 7 R
S.E.46 18 S 22 S S
25 S 30 S 35 S S 25 S
S.E.47 21 S 22 S S
28 S 36 S 40 S S 7 R

Annexes
121
S.E.48 21 S 23 S S
33 S 34 S 34 S S 26 S
S.E.49 16 I 12 R S
7 R 7 R 7 R S 7 R
S.E.50 19 S 23 S S
25 S 31 S 38 S S 7 R
S.E.51 13 R 12 R S
7 R 7 R 7 R S 7 R
S.E.52 19 S 22 S S
25 S 32 S 36 S S 25 S
S.Kp.1 7 R 9 R 23 S 7 R 7 R 11 R 7 R 17 S 7 R
S.Kp.2 16 I 10 R 22 S 19 S 21 S 25 S 29 S 15 S 18 S
S.Kp.3 16 I 10 R 23 S 20 S 21 S 26 S 28 S 15 S 10 R
S.Kp.4 18 S 12 R 25 S 22 S 25 S 30 S 33 S 16 S 13 I
S.Kp.5 7 R 7 R 18 S 8 R 15 I 18 R 16 R 16 S 7 R
S.Kp.6 7 R 7 R 18 S 9 R 7 R 11 R 10 R 15 S 7 R
S.Kp.7 9 R 9 R 20 S 11 R 16 I 18 R 16 R 15 S 7 R
S.Kp.8 8 R 9 R 17 S 10 R 15 R 17 R 15 R 15 S 7 R
S.Kp.9 11 R 11 R 21 S 13 R 7 R 7 R 7 R 17 S 7 R
S.Kp.10 8 R 8 R 18 S 9 R 16 I 19 R 18 R 18 S 7 R
S.Kp.11 9 R 9 R 21 S 12 R 16 I 19 R 19 R 16 S 7 R
S.Kp.12 22 S 25 S 24 S 23 S 21 S 28 S 30 S 16 S 7 R
S.Kp.13 18 S 20 S 20 S 20 S 18 I 24 I 23 I 15 S 19 S
S.Kp.14 7 R 7 R 14 R 7 R 7 R 10 R 7 R 15 S 12 R
S.Kp.15 8 R 10 R 14 R 11 R 15 I 19 R 19 R 15 S 7 R
S.Kp.16 7 R 7 R 14 R 7 R 7 R 10 R 8 R 15 S 7 R
S.Kp.17 8 R 10 R 19 S 9 R 14 R 16 R 16 R 15 S 7 R
S.Kp.18 16 I 8 R 25 S 30 S 25 S 31 S 33 S 15 S 23 S
S.Kp.19 23 S 25 S 25 S 31 S 26 S 30 S 36 S 17 S 27 S
S.Kp.20 9 R 10 R 19 S 15 R 14 R 17 R 16 R 15 S 7 R
S.Kp.21 7 R 7 R 7 R
22 I 26 S 26 S 15 S 7 R
S.Kp.22 7 R 7 R 19 S
15 I 18 R 17 R 15 S 7 R
S.Kp.23 9 R 8 R S
7 R 12 R 7 R 15 S 7 R
S.Kp.24 23 S 24 S S
21 S 27 S 32 S 15 S 7 R
S.Kp.25 12 R 9 R S
20 S 26 S 28 S 15 S 7 R
S.Kp.26 7 R 7 R S
20 S 26 S 26 S 15 S 7 R
S.Kp.27 8 R 9 R S
14 R 18 R 16 R 15 S 7 R
S.Kp.28 10 R 12 R S
9 R 21 R 17 R 15 S 7 R
S.Kp.29 12 R 11 R S
22 S 29 S 30 S 15 S 23 S
S.Kp.30 9 R 10 R S
14 R 20 R 17 R 15 S 7 R
S.Pm.1 26 S 30 S 28 S 7 R 20 S 34 S 32 S 7 R 7 R
S.Pm.2 16 I 15 R 22 S 16 I 17 I 29 S 30 S 7 R 7 R
S.Pm.3 12 R 12 R 20 S 7 R 7 R 7 R 7 R 7 R 7 R
S.Pm.4 22 S 22 S 28 S 20 S 25 S 35 S 44 S 7 R 7 R
S.Pm.5 29 S 31 S 30 S 30 S 7 R 20 R 27 S 7 R 7 R
S.Pm.6 22 S 25 S 23 S 7 R 20 S 30 S 35 S 7 R 7 R
S.Pm.7 17 I 19 S 17 S 19 S 22 S 27 S 30 S 8 R 21 S
S.Pm.8 18 S 21 S 21 S 7 R 21 S 31 S 34 S 7 R 7 R
S.Pm.9 19 S 21 S 19 S 30 S 25 S 30 S 36 S 7 R 20 S
S.Pm.10 18 S 19 S 22 S 26 S 22 S 25 S 28 S 7 R 19 S
S.Pm.11 21 S 23 S 21 S 7 R 7 R 7 R 21 R 7 R 7 R
S.Pm.12 21 S 23 S 21 S 18 S 7 R 11 R 21 R 8 R 7 R
S.Pm.13 21 S 23 S 22 S S 16 I 28 S 28 S 7 R 7 R
S.Pm.14 21 S 23 S S S 23 S 33 S 38 S 7 R 20 S
S.Pm.15 19 S 22 S S S 20 S 30 S 35 S 7 R 19 S
S.Pm.16 23 S 26 S S S 26 S 32 S 34 S 7 R 22 S
S.Pm.17 24 S 27 S S S 27 S 33 S 35 S 7 R 21 S
S.Pm.18 22 S 25 S S S 24 S 32 S 34S 7R 20 S
Annexes
122
S.Pm.19 12 R 13 R S R 7 R 12 R 19 R 7 R 7 R
S.Pv.1 12 R 14 R 23 S 7 R 20 S 22 I 28 S 7 R 7 R
S.Pv.2 20 S 23 S 21 S 20 S 22 S 26 S 30 S 7 R 15 I
S.Pv.3 21 S 26 S 23 S 21 S 21 S 29 S 30 S 7 R 23 S
S.Pv.4 16 I 13 R 19 S 12 R 7 R 7 R 16 R 7 R 19 S
S.Pv.5 24 S 29 S 28 S 20 S 14 R 16 R 24 I 13 R 7 R
S.Pv.6 22 S 26 S 24 S 32 S 20 S 32 S 40 S 7 R 23 S
S.Pv.7 20 S 24 S 23 S 30 S 27 S 35 S 35 S 7 R 27 S
S.Pp.1 20 S 21 S 20 S 20 S 20 S 27 S 31 S 7 R 19 S
S.Ec.1 24 S 26 S 28 S 25 S 25 S 30 S 39 S 19 S 29 S
S.Ec.2 8 R 9 R 18 S 10 R 11 R 17 R 17 R 15 S 7 R
S.Ec.3 9 R 10 R 19 S 12 R 9 R 18 R 17 R 16 S 7 R
S.Ec.4 20 S 23 S 22 S 22 S 21 S 27 S 32 S 19 S 25 S
S.Ec.5 12 R 9 R 23 S 18 S 16 I 22 I 29 S 15 S 7 R
S.Ec.6 9 R 10 R 20 S 10 R 12 R 17 R 18 R 14 R 7 R
S.Ec.7 20 S 22 S 22 S 20 S 15 I 23 I 17 R 16 S 18 S
S.Ec.8 16 I 11 R 26 S 20 S 22 S 28 S 36 S 17 S 7 R
S.Ec.9 16 I 11 R 27 S 21 S 19 I 25 S 33 S 18 S 7 R
S.Ec.10 9 R 10 R 20 S 14 R 15 I 20 R 18 R 14 R 7 R
S.Ec.11 20 S 25 S 20 S 21 S 23 S 25 S 28 S 16 S 25 S
S.Ec.12 16 I 15 R 23 S 20 S 7 R 13 R 7 R 13 R 7 R
S.Ec.13 20 S 23 S 21 S 26 S 23 S 26 S 30 S 19 S 19 S
S.Ec.14 7 R 7 R 18 S 7 R 7 R 7 R 7 R 17 S 7 R
S.Ec.15 11 R 23 S S 7 R 7 R 7 R 7 R S 7 R
S.Ec.16 20 S 22 S 22 S 26 S 19 I 26 S 30 S S 23 S
S.Ec.17 12 R 9 R S
16 I 23 I 28 S S 7 R
S.Ec.18 20 S 22 S 22 S
25 S 26 S 33 S S 23 S
S.Ec.19 19 S 21 S 22 S
27 S 30 S 36 S S 25 S
S.Ec.20 18 S 20 S 21 S
22 S 28 S 33 S S 19 S
S.Ec.21 18 S 20 S 22 S
20 S 29 S 31 S S 21 S
S.Ec.22 12 R 12 R S
16 I 27 S 34 S S 7 R
S.Ec.23 12 R 13 R S
16 I 26 S 32 S S 7 R
S.Ec.24 13 R 11 R S
16 I 24 I 32 S S 7 R
S.Ec.25 20 S 23 S 22 S
25 S 35 S 40 S S 23 S
S.Ec.26 9 R 10 R S
10 R 17 R 17 R S 7 R
S.Ps.1 R S S S S S S R R
S.Mm.1 S S S R R R S R R
S.CF.1 I S S S I I S S S
S.SS.1 S S S S S S S S S
S.Sel.spp. S S S S R R S S S

Annexes
123
Annexe 10. Résultats des concentrations minimales ihnibitrice des souches .
Souches Antibiotiques
Phénotypes PIP TZP IMP FOX CTX CAZ ATM FEP TM GM AN CIP RIF
S.E.2 512
R 2 S
0,25
S <1 S
>512
R 16 R 32 R 64 R 4 I 16 R 2 S 0,5 S 16 I BLSE
S.E.15 8 S / 0,5
S 64 R 8 R 16 R 8 I
< 0,5
S 2 S 0,5 S 1 S 0,25 S 4 S CASE
S.E.16 >512
R /
0,25
S 2 S 32 R
128
R
256
R 4 I 8 R 32 R 2 S
<0,125
S 4 S BLSE
S.E.23 >512
R /
0,25
S 4 S
>512
R
128
R
128
R
256
R
>512
R
>512
R
>512
R 1 I 8 I BLSE+CASE
S.E.26 >512
R /
0,25
S 4 S
>512
R 64 R 64 R
128
R 16 R 16 R 2 S 32 R 2 S BLSE+CASE
S.E.38 >512
R /
0,25
S 4 S
>512
R 64 R 64 R
256
R 4 I 8 R 0,5 S
<0,125
S 2 S BLSE
S.E.47 >512
R /
0,5
S 4 S
>512
R 8 I 16 R 64 R 2 S 0,5 S 1 S
<0,125
S 4 S BLSE+CASE
S.E.49 >512
R / 1 S 4 S
>512
R 8 I 16 R 32 R 4 I 32 R 1 S 16 R 4 S BLSE+CASE
S.E.51 >512
R /
0,25
S 32 I
>512
R
128
R
256
R
256
R 8 R 32 R 2 S 64 R 4 S BLSE+CASE
S.Kp.1 512
R 16 I
0,5
S 8 S
>512
R
128
R
256
R
512
R 64 R
128
R 8 S 16 R
512
R BLSE
S.Kp.2 512
R 16 I 1 S 32 I
>512
R 64 R
128
R
512
R 4 I 32 R
<0,5
S
<0,125
S
32
R BLSE+CASE
S.Kp.3 512
R 4 S
0,25
S 4 S
128
R 4 I 16 R 32 R 4 I 16 R
<0,5
S
<0,125
S
64
R BLSE
S.Kp.4 512
R 4 S
0,5
S
< 1
S
256
R 2 I
<0,5
S 64 R 2 S 16 R
<0,5
S
<0,125
S
64
R BLSE
S.Kp.6 512
R 16 I 1 S 8 S
512
R 32 R 64 R 64 R 32 R 64 R 4 S 8 R
32
R BLSE+CASE
S.Kp.7 512
R 8 S
0,5
S 4 S
>512
R 16 R 64 R 32 R 32 R 32 R 8 S 2 R
32
R BLSE
S.Kp.8 512
R 16 I
0,5
S 4 S
512
R 16 R 64 R 64 R 32 R 64 R 8 S 4 R
32
R BLSE
S.Kp.9 512
R 8 S
0,5
S 8 S
>512
R 32 R
512
R
128
R 16 R 32 R
<0,5
S 64 R
32
R BLSE
S.Kp.11 512
R 8 S
0,5
S
< 1
S
>512
R 8 I
<0,5
S 32 R 32 R 32 R
<0,5
S 2 R
32
R BLSE
S.Kp.14 >512
R 16 I
0,5
S 16 I
>512
R
512
R
256
R
512
R 64 R
256
R 32 R 32 R
64
R BLSE+CASE
S.Kp.15 >512
R 16 I 1 S 4 S
>512
R
128
R
128
R
512
R 16 R 64 R 32 R 2 R
64
R BLSE
S.Kp.16 >512
R 16 I
0,5
S 32 I
512
R
512
R
512
R
256
R 64 R
256
R 32 R 64 R
512
R BLSE+CASE
S.Kp.17 >512
R 16 I
0,5
S 4 S
512
R 32 R 32 R 64 R 16 R 64 R 2 S 4 R
64
R BLSE
S.Kp.18 >512
R /
0,25
S 4 S
256
R 4 I 16 R 64 R 4 I 32 R 0,5 S
<0,125
S 4 S BLSE
S.Kp.20 >512
R / 1 S 4 S
>512
R 32 R 32 R 64 R 16 R 16 R 2 S 2 R
32
R BLSE
S,Kp.21 >512
R / 1 S 8 S
>512
R
512
R
512
R
>512
R
> 512
R
>512
R
>512
R
<0,125
S 16 I BLSE
S,Kp.22 >512
R / 1 S 4 S
>512
R 32 R 64 R
128
R 32 R 32 R 2 S 2 R
32
R BLSE
S,Kp.23 >512
R / 1 S 8 S
>512
R
256
R
512
R
>512
R 32 R 32 R 2 S 32 R
32
R BLSE+CASE
S.Kp.25 >512
R / 1 S 4 S
>512
R
256
R
256
R
>512
R 8 R 32 R 1 S 2 R
32
R BLSE+CASE
S.Kp.26 >512
R / 1 S 4 S
>512
R
128
R
128
R
512
R 32 R 32 R 8 S
<0,125
S
32
R BLSE+CASE
S.Kp.27 >512
R /
0,5
S 4 S
>512
R 64 R 64 R
128
R 16 R 16 R 2 S 2 R
32
R BLSE

Annexes
124
S.Kp.28 >512
R /
0,5
S 4 S
512
R 32 R 64 R 32 R 16 R 32 R 2 S 2 R
32
R BLSE
S.Kp.29 >512
R /
0,5
S 4 S
>512
R 16 R 64 R
512
R 8 R 16 R 1 S
<0,125
S
32
R BLSE+CASE
S.Kp.30 >512
R /
0,5
S 4 S
>512
R
256
R
256
R
512
R 16 R 16 R 2 S 2 R
32
R BLSE+CASE
S.Pm.2 512
R 2 S >2 <1 S 32 R
256
R 2 I 8 I 4 I 8 R 8 S
<0,125
S
256
R BLSE
S.Pm.3 512
R 4 S >2 8 S
>512
R 16 R 2 I 8 I 16 R 16 R 8 S
>128
R 8 I Case+BLSE
S.Pm.4 256
R 4 S 1 S 4 S
128
R
512
R 1 S
128
R 2 S 2 S 8 S
<0,125
S
256
R BLSE
S.Pm.9 / / 2 S <1 4 R 16 R <1 S <1 S <0,25
S
<0,5
S <1 S
<0,125
S 8 I CASE
S.Pm.12 / / 1 S <1 S 4 R 2 I <1 S <1 S 2 S 1 S 8 S 2 R <16 CASE
S.Pm.19 / / 2 S 16 I 512
R
512
R 64 R 16 R 32 R 32 R 8 S 2 R 32 BLSE
S.Pv.1 32 R 4 S 1 S 2 S >512
R
512
R 4 I 64 R 8 R 8 R <1 S 0,25 S 8 I BLSE
S.Pv.4
1 S 2 S 4 R 16 R <1 S <1 S 2 S 4 I <1 S 2 R
CASE
S.Ec.2 512
R 16 I 1 S
512
R
>512
R 16 R 64 R
256
R 16 R 16 R 4 S 2 R 16 I BLSE
S.Ec.3 512
R 16 I 1 S
512
R
>512
R 64 R
128
R
256
R 16 R 32 R 4 S 2 R 16 I BLSE
S.Ec.4 4 S 1 S 4 I >512
R 16 R 2 I 8 I
<0,5
S
<0,25
S 0,5 S
<0,5
S
<0,125
S
32
R CASE
S.Ec.5 >512
R 8 S 1 S
256
R 64 R
256
R
256
R 4 I 8 R 32 R
<0,5
S 0,5 S
32
R BLSE
S.Ec.6 >512
R 16 I 1 S
512
R
>512
R 64 R 64 R
128
R 16 R 64 R 8 S 4 R
32
R BLSE
S.Ec.8 >512
R 16 I 1 S
256
R
>512
R
128
R
128
R
512
R 4 I 64 R
<0,5
S
0,125
S
32
R BLSE+CASE
S.Ec.9 >512
R 8 S 1 S
256
R 64 R
256
R
256
R 8 I 4 I 32 R
<0,5
S 0,5 S
32
R BLSE
S.Ec.10 512
R 16 I 1 S
512
R
>512
R 64 R 64 R
256
R 16 R 32 R 4 S 4 R
64
R BLSE
S.Ec.11 2 S 1 S 0,5
S
512
R
512
R 8 I 2 I 1 S 0,5 S 0,5 S
<0,5
S
<0,125
S
32
R CASE
S.Ec.12 512
R 16 I 4 I 32 I
>512
R 16 R 2 I 32 R 4 I 8 R
<0,5
S 64 R
32
R
BLSE+
CASE
S.Ec.14 >512
R / 2 S
512
R
>512
R 64 R
256
R
>512
R 32 R 32 R 2 S 32 R
32
R BLSE+CASE
S.Ec.15 >512
R /
0,5
S 16 I
>512
R
256
R
256
R
>512
R 8 R
<0,5
S 2 S 128 R
32
R BLSE+CASE
S.Ec.17 >512
R / 1 S
256
R
128
R
256
R
256
R 4 I 8 R 32 R 1 S
<0,125
S 16 I BLSE
S.Ec.22 >512
R / 1 S 64 R 64 R
128
R
128
R 2 I 8 R 16 R 1 S 0,25 S
32
R BLSE+CASE
S.Ec.23 >512
R /
0,5
S 64 R 64 R
128
R
128
R 2 I 8 R 16 R 1 S
<0,125
S
32
R BLSE
S.Ec.24 >512
R / 1 S
256
R
128
R
256
R
256
R 8 I 8 R 32 R
<0,5
S
<0,125
S 16 I BLSE
S.Ec.26 >512
R / 2 S
256
R
>512
R
128
R
128
R
>512
R 32 R 16 R 2 S 4 R
32
R BLSE
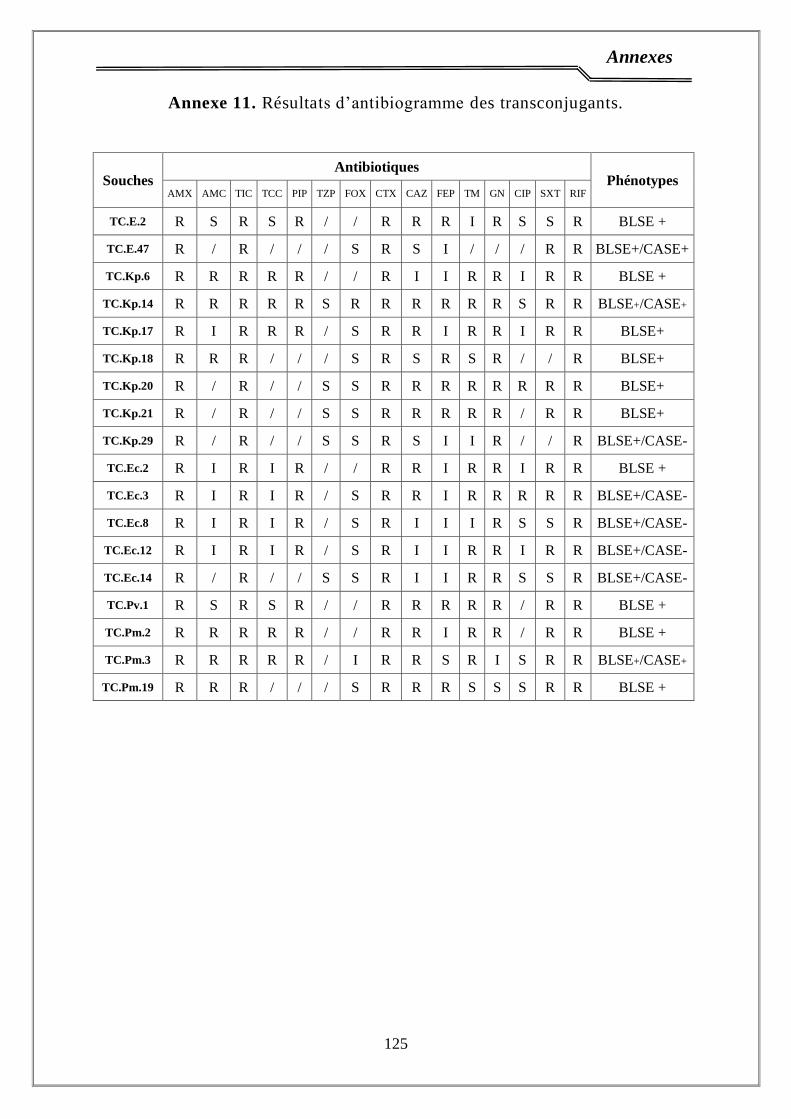
Annexes
125
Annexe 11. Résultats d’antibiogramme des transconjugants.
Souches Antibiotiques
Phénotypes AMX AMC TIC TCC PIP TZP FOX CTX CAZ FEP TM GN CIP SXT RIF
TC.E.2 R S R S R / / R R R I R S S R BLSE +
TC.E.47 R / R / / / S R S I / / / R R BLSE+/CASE+
TC.Kp.6 R R R R R / / R I I R R I R R BLSE +
TC.Kp.14 R R R R R S R R R R R R S R R BLSE+/CASE+
TC.Kp.17 R I R R R / S R R I R R I R R BLSE+
TC.Kp.18 R R R / / / S R S R S R / / R BLSE+
TC.Kp.20 R / R / / S S R R R R R R R R BLSE+
TC.Kp.21 R / R / / S S R R R R R / R R BLSE+
TC.Kp.29 R / R / / S S R S I I R / / R BLSE+/CASE-
TC.Ec.2 R I R I R / / R R I R R I R R BLSE +
TC.Ec.3 R I R I R / S R R I R R R R R BLSE+/CASE-
TC.Ec.8 R I R I R / S R I I I R S S R BLSE+/CASE-
TC.Ec.12 R I R I R / S R I I R R I R R BLSE+/CASE-
TC.Ec.14 R / R / / S S R I I R R S S R BLSE+/CASE-
TC.Pv.1 R S R S R / / R R R R R / R R BLSE +
TC.Pm.2 R R R R R / / R R I R R / R R BLSE +
TC.Pm.3 R R R R R / I R R S R I S R R BLSE+/CASE+
TC.Pm.19 R R R / / / S R R R S S S R R BLSE +
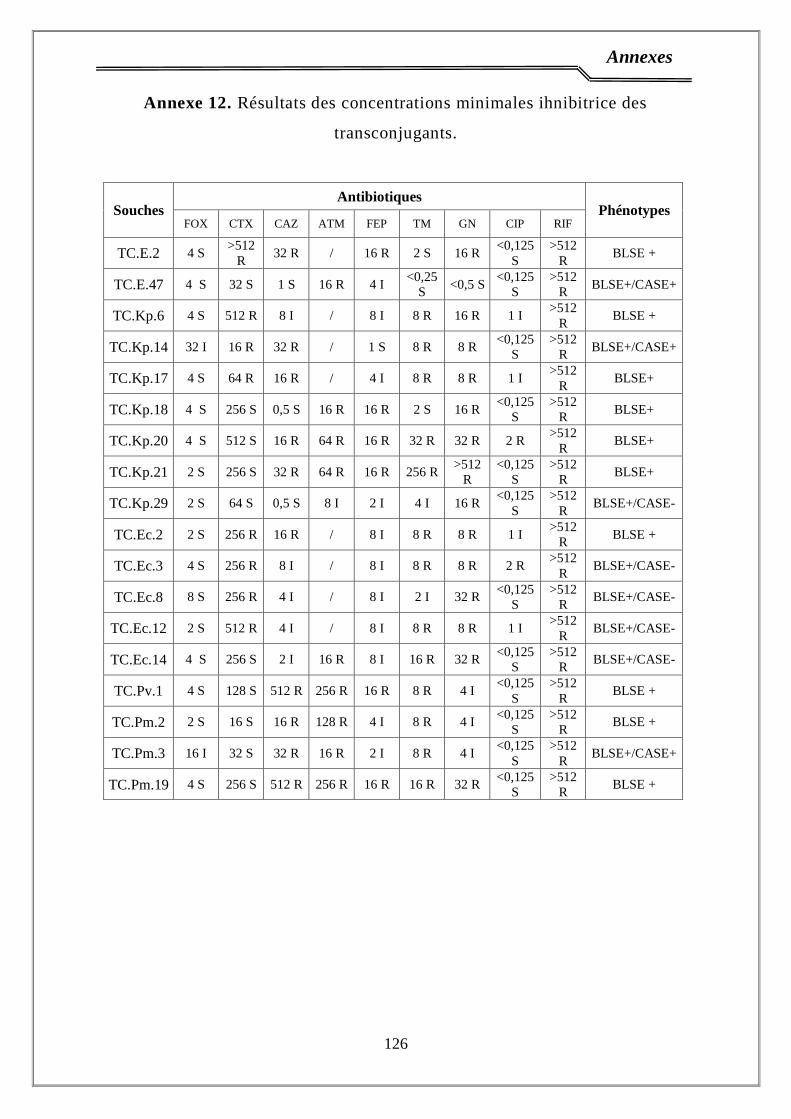
Annexes
126
Annexe 12. Résultats des concentrations minimales ihnibitrice des
transconjugants.
Souches Antibiotiques
Phénotypes FOX CTX CAZ ATM FEP TM GN CIP RIF
TC.E.2 4 S >512
R 32 R / 16 R 2 S 16 R
<0,125
S
>512
R BLSE +
TC.E.47 4 S 32 S 1 S 16 R 4 I <0,25
S <0,5 S
<0,125
S
>512
R BLSE+/CASE+
TC.Kp.6 4 S 512 R 8 I / 8 I 8 R 16 R 1 I >512
R BLSE +
TC.Kp.14 32 I 16 R 32 R / 1 S 8 R 8 R <0,125
S
>512
R BLSE+/CASE+
TC.Kp.17 4 S 64 R 16 R / 4 I 8 R 8 R 1 I >512
R BLSE+
TC.Kp.18 4 S 256 S 0,5 S 16 R 16 R 2 S 16 R <0,125
S
>512
R BLSE+
TC.Kp.20 4 S 512 S 16 R 64 R 16 R 32 R 32 R 2 R >512
R BLSE+
TC.Kp.21 2 S 256 S 32 R 64 R 16 R 256 R >512
R
<0,125
S
>512
R BLSE+
TC.Kp.29 2 S 64 S 0,5 S 8 I 2 I 4 I 16 R <0,125
S
>512
R BLSE+/CASE-
TC.Ec.2 2 S 256 R 16 R / 8 I 8 R 8 R 1 I >512
R BLSE +
TC.Ec.3 4 S 256 R 8 I / 8 I 8 R 8 R 2 R >512
R BLSE+/CASE-
TC.Ec.8 8 S 256 R 4 I / 8 I 2 I 32 R <0,125
S
>512
R BLSE+/CASE-
TC.Ec.12 2 S 512 R 4 I / 8 I 8 R 8 R 1 I >512
R BLSE+/CASE-
TC.Ec.14 4 S 256 S 2 I 16 R 8 I 16 R 32 R <0,125
S
>512
R BLSE+/CASE-
TC.Pv.1 4 S 128 S 512 R 256 R 16 R 8 R 4 I <0,125
S
>512
R BLSE +
TC.Pm.2 2 S 16 S 16 R 128 R 4 I 8 R 4 I <0,125
S
>512
R BLSE +
TC.Pm.3 16 I 32 S 32 R 16 R 2 I 8 R 4 I <0,125
S
>512
R BLSE+/CASE+
TC.Pm.19 4 S 256 S 512 R 256 R 16 R 16 R 32 R <0,125
S
>512
R BLSE +

ملخصف اىسخشف شاقبت حسبست اىضبداث اىحت. بنخشب اىؼت حثو احذة اىؼبئالث اىؼصبث سيبت اىغشا اىسؤىت ػ إصبببث خغشةاى
اىبنخشب اىؼت سالىت 041 دساست .اىحت خغة ضشست ألب حج اخخبس اىؼالج اىخجشب حقيو اىضغط اىخ حبسسب اىضبداث حثو
ح ،37.1 % :ـححخو اىشحبت االى ب lico Escherichiaحب ا ، 9101 ب 9112 بيؼببط ب أمخبش خخيفت حذاث سخشف سذاىؼضىت
ضبد ح 79إى اىحسبست lla pneumoniaeKlebsie %)4.(21 . spp Proteus 3%)(19. e cloacaEnterobacter . (18.6%) حيب
18 -- ب ىؼبشــظ ال فقـػ عشق اخشبس األقشاص ف س سىفبثمضاصه -مى ، مىسخ حشثبش 3 ، اصذ 4، ىنخب
SFM -CA بت ىو ـ، حب اخشبس سالالث ق-رىل بــ بب اإل نبسألابء ـت ببسخثـاىذسس األخشش اىضبداث اىحت ـاس بئشــىنخب س :
نشف ػي حع ظش غ ت بىسحج ب ىنخب -ىقبت ىوااىظبشت األبط ححيو . اىحسبستاىخاى اىسالالث ػي 95.7% 26.4 %
خخببس ححو اىقبت إح BLSE ىز اىسالالث راث اىظ اىظبش. 37.0 % :ـ رىل ب BLSE ىذ اىاسغار ىنخببص -ىو اىخجت اىسالالث
راىخحيو مشف خخيف األبط جد اىؼذذ اىبيسذاث غ احذ شخشك . قبسصىنخشفساص ػي ال األإلع بشبذة اىض اىبالسذ بب
.Kb 55 مبش أجضئ ص
كلمات مفتاحية:
.قبسصألىنخشفساص ػي ال ااإل-ححو اىقبت -BLSE-قبتاى-اىضبداث اىحت -اىؼصبث اىؼت
Résumé
Les entérobactéries représentent une des principales familles de bacilles à Gram négatif responsables d’infections
humaines graves. La surveillance de la sensibilité aux antibiotiques à l’hôpital est une étape essentielle; elle oriente le
choix des traitements empiriques et permet de réduire la pression de sélection exercée par les antibiotiques. L’étude de
140 souches d’entérobactérie isolées en milieu hospitalier au niveau de différents services du CHU de Sidi Bel Abbes
entre octobre 2009 et mai 2010, montre qu’Escherichia coli occupe la première place soit 37.1% suivie de Klebsiella
pneumoniae (21.4%), de Proteus spp. (19.3%) et d’Enterobacter cloacae (18.6%). La résistance vis-à-vis de 27
antibiotiques (18 -lactamines, 4 aminosides, 3 quinolones, colistine et triméthoprime/sulfamethoxasole) par la méthode
de diffusion des disques en milieu gélosé, selon les normes du CA-SFM, a révélé l’émergence des souches résistantes
aux -lactamines et à la majorité des autres classes d’antibiotiques sauf pour l’amikacine et l’imipénème soit 96.4% et
95.7% respectivement des souches sensibles. L’analyse des phénotypes de résistance aux -lactamines a permis de
détecter une diversité phénotypique avec dominance des souches productrices de -lactamase à spectre étendu (BLSE)
soit 37.1%. Ces souches présentant le phénotype BLSE ont fait l’objet d’un test de conjugaison suivi d’une visualisation
du contenu plasmidique par électrophorèse sur gel d’agarose. L’analyse révèle différents profils et la présence de
plusieurs plasmides dont un commun de poids moléculaire supérieur à 55 kb.
Mots clés:
Entérobactéries-antibiotiques-résistance-BLSE-conjugaison-électrophorèse sur gel d’agarose.
Summary
Enterobacteria represent major families of Gram-negative bacteria responsible of serious human infections.
Surveillance of antibiotic susceptibility in the hospital is an essential step; it guides the choice of empirical treatment
and reduces the selection pressure exerted by antibiotics. The study of 140 strains of Enterobacteriaceae isolated in
various units in the hospital of Sidi Bel Abbes between october 2009 and may 2010, shows that Escherichia coli is
ranked first (37.1%) followed by Klebsiella pneumoniae (21.4%), Proteus spp. (19.3%) and Enterobacter cloacae
(18.6%). The susceptibility to 27 antibiotics (18 -lactams, 4 aminoglycosides, 3 quinolones, colistin and trimethoprim /
sulfamethoxazole) by the disk diffusion method in agar medium, according to CA-SFM, revealed the emergence of
strains resistant to -lactam antibiotics and most other classes of antibiotics except the amikacin and imipenem with
96.4% and 95.7% respectively of susceptible strains. The analysis of resistance phenotypes to -lactam has detected
diversity phenotypic with dominance of strain producing extended spectrum -lactamase (ESBL) or 37.1%. These
strains with ESBL phenotype were tested conjugation followed by a visualization of plasmidic content by agarose gel
electrophoresis. The analysis reveals differents profiles and the presence of severals plasmids including a common with
molecular weight greater than 55 kb.
Key words:
Enterobacteria- antibiotics-resistance- ESBL- conjugation- agarose gel electrophoresis.