expression of p2x4r mrna and protein in rats with
TRANSCRIPT

Circulation Journal Vol.75, April 2011
Circulation JournalOfficial Journal of the Japanese Circulation Societyhttp://www.j-circ.or.jp
he pulmonary hypertension that develops in hypobaric hypoxia is characterized by structural remodeling of the heart (eg, right ventricular hypertrophy).1–7 Over
the past 20 years or so, various factors and influences, includ-ing the adrenergic-receptor system, have been shown to be involved in such cardiovascular remodeling processes.8–12
It is well known that hypoxia stimulates endothelial cells to release adenosine triphosphate (ATP). Purinergic P2 nucle-otide receptors on endothelial cells bind this ATP, triggering secretion of nitric oxide and consequent vasodilation. The P2 class of nucleotide receptors includes the P2X receptor, which is a ligand-gated receptor channel, and the G protein-coupled P2Y receptor.13,14 Recently, the P2X4 receptor (P2X4R) has been reported to control vascular tone and vessel remodeling in at least some blood vessels.15–19 Interestingly, P2X4R is an important subunit of the native cardiac myocyte P2X recep-tor,17 and cardiac-restricted overexpression of P2X4R can induce an enhanced contractile state of the intact heart.20 More recently, it was found that cardiac overexpression of
P2X4R increased cardiac contractility and survival in a model of myocardial infarction-induced cardiac failure.21 It has therefore emerged as a key factor in the enhancement of car-diac performance. However, no reports have been published of alterations in P2X4R expression in certain organs, espe-cially the heart, upon exposure to hypobaric hypoxia, and lit-tle is known about any changes in P2X4R expression in hypoxia-induced pulmonary hypertension (in which only RV is exposed to pressure overload).
Even if changes in P2X4R expression occur during expo-sure to hypobaric hypoxia and show an apparent relation to the pulmonary pressure overload, it needs to be firmly estab-lished: (1) whether the changes in P2X4R (at both the protein and mRNA levels) occurring in certain organs, especially the heart, are coupled to the elevation in pulmonary arterial pres-sure seen after exposure to hypobaric hypoxia; (2) whether the changes in P2X4R protein occurring in certain organs as an adaptation to hypobaric hypoxic exposure are mirrored by similar changes at the mRNA level; and (3) whether the dis-
Received August 7, 2009; revised manuscript received November 22, 2010; accepted December 6, 2010; released online March 3, 2011 Time for primary review: 51 days
Department of Pathology and Laboratory Medicine, National Defense Medical College, Tokorozawa (Y.O., S.O., K.N., S.H., S.T., T.K.); Biochemical Section, 2nd Division, Aeromedical Laboratory, Japan Air Self-Defense Force, Tachikawa (F.K.); and Division of Environmental Medicine, National Defense Medical College Research Institute, Tokorozawa (M.U.), Japan
Mailing address: Kuniaki Nakanishi, MD, Department of Pathology and Laboratory Medicine, National Defense Medical College, Tokorozawa 359-8513, Japan. E-mail: [email protected]
ISSN-1346-9843 doi: 10.1253/circj.CJ-09-0575All rights are reserved to the Japanese Circulation Society. For permissions, please e-mail: [email protected]
Expression of P2X4R mRNA and Protein in Rats With Hypobaric Hypoxia-Induced
Pulmonary HypertensionYuichiro Ohata, MD; Sho Ogata, MD; Kuniaki Nakanishi, MD; Fumiko Kanazawa; Maki Uenoyama; Sadayuki Hiroi, PhD; Susumu Tominaga; Toshiaki Kawai, MD
Background: The experimental pulmonary hypertension that develops in hypobaric hypoxia is characterized by structural remodeling of the heart. The P2X4 receptor (P2X4R) controls vascular tone and vessel remodeling in several blood vessels, and it has emerged as a key factor in the enhancement of cardiovascular performance.
Methods and Results: To study the possible effects of hypobaric hypoxia on the P2X4R-synthesis system, 150 male Wistar rats were housed in a chamber at the equivalent of the 5,500 m altitude level for 21 days. After 14 days’ exposure to hypobaric hypoxia, pulmonary arterial pressure (PAP) was significantly increased. In the right ventricle (RV) of the heart, P2X4R expression was significantly increased on days 1 and 14 (mRNA) and on days 7 and 21 (protein) of hypobaric hypoxic exposure. Immunohistochemical staining for P2X4R protein became more intense in RV in the late phase of exposure. These changes in P2X4R synthesis in RV occurred alongside the increase in PAP. In addition, P2X1R and P2Y2R mRNA levels in the RV were significantly increased on days 1, 14, and 21, and day 5, respectively, of exposure. The level of P2X1R protein in the RV was significantly increased on day 21 of exposure.
Conclusions: Conceivably, P2 receptors, including P2X4R and P2X1R, might play roles in modulating the RV hypertrophy that occurs due to pulmonary hypertension in hypobaric hypoxia. (Circ J 2011; 75: 945 – 954)
Key Words: Hypobaric hypoxia; P2X4 receptor; Pulmonary hypertension
T
ORIGINAL ARTICLEPulmonary Circulation

Circulation Journal Vol.75, April 2011
946 OHATA Y et al.
tributions of P2X4R mRNA and protein are altered by expo-sure to hypobaric hypoxia. In the present study, in order to better understand the effects of hypobaric hypoxic exposure on P2X4R, rats were housed in a mechanical chamber at the equivalent of an altitude of 5,500 m for 3 weeks. We mea-sured pulmonary arterial pressure and calculated the levels of right ventricular hypertrophy as well as the expression of P2X4R (mRNA and protein) in a variety of organs to help us examine the possible causes of the morphologic and physio-logic changes occurring in the heart during hypobaric hypoxic exposure. In addition, we made semiquantitative measure-ments of mRNA and proteins for P2X1R and P2Y2R, for the following reasons. It is well known that diadenosine poly-phosphates (formed from 2 adenosine molecules) function as vasoconstrictor agents, probably via actions on the P2X1 receptor (P2X1R) and P2Y2 receptor (P2Y2R),22 and more-over in rats exposed to experimental congestive heart failure, P2X1R and P2Y2R mRNA levels are increased in the myo-cardium of the left ventricle.23
MethodsWe used essentially the same methods as in previous reports.4–7 Briefly, 150 adult male Wistar rats, approximately 8 weeks old and weighing 190 to 210 g, were divided into 7 groups. Groups of 17 or 18 rats each were housed in a 5.6 × 3.0 × 3.0 m mechanical chamber (Hitachi Corp, Tokyo, Japan) and exposed to hypobaric hypoxia equivalent to 5,500 m in alti-
tude (380 mmHg = 50.6 kPa) for 0.5 to 21 days. The chamber comprised a main room to house the animals and an acces-sory room. When investigators (with their own oxygen sup-ply) needed to enter the accessory room, it was decompressed to hypobaric hypoxia equivalent to 5,500 m in altitude. They then took care of the rats exposed to hypobaric hypoxia. Tests on the animals in hypobaric hypoxia were conducted in the accessory room. The oxygen content in the main room was maintained constant at an FiO2 of 0.105 to simulate 5,500 m in altitude. The age of the rats when exposure to hypobaric hypoxia was initiated was such that all were approximately 11 weeks old at the termination of the experiment. The chamber was kept at an ambient temperature of 23±1°C and a relative humidity of 60–70%, with light exposure daily from 06.00 to 18.00 h. All rats were given commercial chow and tap water ad libitum. After 0.5, 1, 3, 5, 7, 14 or 21 days of exposure to hypobaric hypoxia, rats were killed by decapitation. An addi-tional 17 rats kept in the ground-level environment served as controls. This experimental study was carried out in accor-dance with the recommendations in the Guide for the Care and Use of Laboratory Animals of the National Institutes of Health. The protocol was approved by the Committee on the Ethics of Animal Experiments in the National Defense Medi-cal College. Mean pulmonary arterial pressure (PAPm), mean systemic arterial pressure (SAPm), and the ratio of right ven-tricle over left ventricle plus septum (RV/LV + S) were evalu-ated at 0.5, 1, 3, 7, 14 or 21 days of exposure to the hypobaric hypoxic condition described above; the technique used and
Figure 1. Semiquantitative reverse transcription-polymerase chain reaction (RT-PCR) for P2X4 receptor (P2X4R) (A), P2X1 receptor (P2X1R) (B), and P2Y2 receptor (P2Y2R) (C) mRNA in normal rats. To derive the semiquantitative RT-PCR data shown for these tissues, the actual “P2X4R mRNA/18S rRNA” value obtained for “brain” in 1 individual rat was expressed as “1.0”, and all other values were expressed in a relative manner (n=5 for each group).
Circulation Journal Vol.75, April 2011
947P2X4R in Hypobaric Hypoxic Rats
the results obtained have been reported elsewhere.7
Blood Sampling and AutopsyBefore autopsy, the rats were decapitated and blood samples were taken (from rats not subjected to arterial pressure mea-surement) for the determination of hematocrit.
At autopsy, the heart was removed and dissected on ice into bilateral atrial and ventricular tissues. Tissues were fro-zen in liquid nitrogen, and stored at –80°C until use. The car-diac tissues so obtained were subjected to RNA and peptide extractions. All the parts of the various organs isolated from each of 11 rats were frozen in liquid nitrogen, and stored at –80°C until used for reverse transcription-polymerase chain reaction (RT-PCR) and Western blotting analysis. Tissues from 2 to 3 rats were fixed separately in 10% formalin for immunohistochemistry.
Total RNA Extraction and Semiquantitative RT-PCRIn all 6 tissues isolated from 5 control rats in each of the groups exposed to hypobaric hypoxia, total RNA was iso-lated using acid guanidinium isothiocyanate-phenol-chloro-form extraction and ethanol precipitation.5–7 RT-PCR for rat P2X4R and rat 18S rRNA was performed using an amplifica-tion reagent kit (TaqMan EZRT-PCR kit; Applied Biosys-
tems, Alameda, CA, USA). These primers were synthesized by an automated DNA synthesizer. The sense and antisense sequences, TaqMan probe, and predicted product sizes for these primers were as follows: 5’-GGG TGA AGT TTT ATT CCA GC-3’ (sense), 5’-GGG TGA AGT TTT CTG CAG CC-3’ (antisense), 5’-ACC AAC ATG ATC GTC ACC GTG AAC C-3’ (TaqMan probe), giving rise to a 301-bp fragment from position 359 to 659 in the cDNA base sequence of rat P2X4R; 5’-AGT CCC TGC CCT TTG TAC ACA-3’ (sense), 5’-GAT CCG AGG GCC TCA CTA AAC-3’ (antisense), and 5CGC CCG TCG CTA CTA CCG ATT GG-3’ (TaqMan probe), giving rise to a 73-bp PCR product corresponding to bases 1674–1752 in the cDNA base sequence of 18S rRNA. Both TaqMan probes were labeled at the 5’ end with the reporter dye molecule, FAM (6-carboxyfluorescein), and at the 3’ end with the quencher dye, BHQ1α. Furthermore, RT-PCR for rat P2X1R and rat P2Y2R was performed using an amplification reagent kit (Power SYBR RNA-to-CT 1-step kit; Applied Biosystems). The sense and antisense sequences, and predicted product sizes for these primers were as follows: 5’-GCT GAT GGC TTT GAG CCA G-3’ (sense) and 5’-TGG TGT CTG CCT TGC AGG A-3’ (antisense), giving rise to a 62-bp fragment from position 1458 to 1520 in the cDNA of rat P2X1R; 5’- TTC TCT AGA GCG TGG ACC
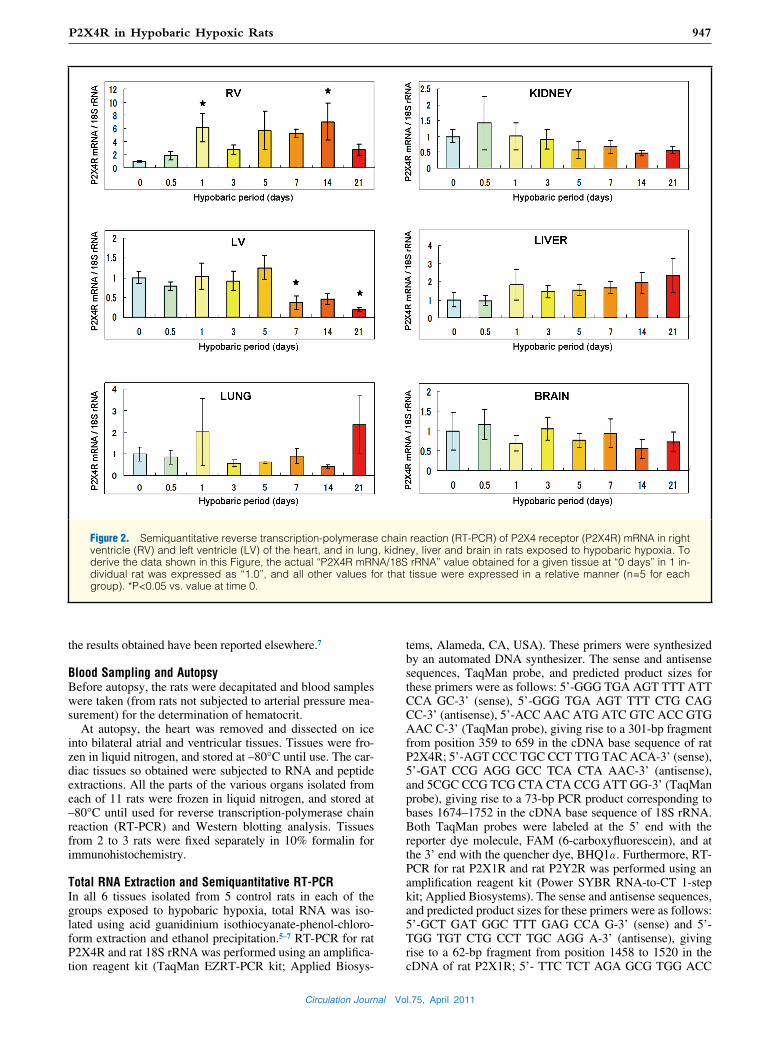
Figure 2. Semiquantitative reverse transcription-polymerase chain reaction (RT-PCR) of P2X4 receptor (P2X4R) mRNA in right ventricle (RV) and left ventricle (LV) of the heart, and in lung, kidney, liver and brain in rats exposed to hypobaric hypoxia. To derive the data shown in this Figure, the actual “P2X4R mRNA/18S rRNA” value obtained for a given tissue at “0 days” in 1 in-dividual rat was expressed as “1.0”, and all other values for that tissue were expressed in a relative manner (n=5 for each group). *P<0.05 vs. value at time 0.

Circulation Journal Vol.75, April 2011
948 OHATA Y et al.
TC-3’ (sense) and 5’-GAC TGA GGC AGG AAA CAG GA-3’ (antisense), giving rise to a 115-bp PCR product corre-sponding to bases 245–359 in the cDNA of P2Y2R. The reaction master mix was prepared according to the manufac-turer’s protocol. To perform PCR for P2X4R and 18S rRNA, the RT reaction was incubated at 60°C for 30 min, followed by incubation at 95°C for 5 min. PCR was performed using 50 and 40 amplification cycles (for P2X4R and 18S rRNA, respectively) at 95°C for 20 s and at 60°C for 1 min using an ABI PRISM 7500 Sequence Detector (Applied Biosystems). To perform PCR for P2X1R and P2Y2R, the RT reaction was incubated at 48°C for 30 min, followed by incubation at 95°C for 10 min. PCR was performed using 50 amplification cycles at 95°C for 20 s and at 60°C for 1 min using an ABI PRISM 7500 Sequence Detector. Total RNA extracts from the brain or liver were used as the control templates for the 4 mRNA. PCR products were separated by electrophoresis in a 3% agarose gel, and stained with ethidium bromide.
Western Blotting AnalysisFor Western blotting analysis, tissue samples were homoge-nized using a sonicated VP-15S Ultras homogenizer (Taitec Co, Tokyo, Japan) for 30 s in 500 μl of an ice-cold homoge-nizing buffer of the following composition: 1% Triton-X,
50 mmol/L Tris/HCL, pH8.0, 150 mmol/L NaCl, and protease inhibitor cocktail (1 tab/10 ml of extraction buffer; Complete Mini; Roche Diagnostics GmbH, Roche Applied Science, Penzberg, Germany). The supernatant was centrifuged at 15,000 g for 30 min at 4°C, and the protein concentration of each supernatant was determined by means of a DC protein assay system (Bio-Rad Laboratories, CA, USA). Then, 5 μg amounts of protein extracts were separated by 7.5% SDS-PAGE, and transferred to polyvinylidene difluoride membranes (Hybond-P; Amersham Biosciences, Buckinghamshire, UK). Membranes were blocked with 5% non-fat dry milk for 90 min at room temperature, then incubated overnight at 4°C with anti-P2X4R antibody (Almone Labs Ltd, Jerusalem, Israel) diluted 1:400, anti-P2X1R (Almone Labs Ltd) diluted 1:250, or anti-P2Y2R (Almone Labs Ltd) diluted 1:250. The same membrane was reprobed with monoclonal anti-β-actin antibody (Sigma, St. Louis, MO, USA) diluted 1:1,000. The densitometry values obtained for the P2X4R blots were stan-dardized to those obtained for the β-actin blots. Finally, after blots had been incubated for 2 h with horseradish peroxidase-conjugated secondary antibodies against rabbit immunoglob-ulins (EnVision + Rabbit/HRP; Dako Cytomation, Denmark) and donkey anti-mouse IgG (HRP conjugated; absorbed for dual labeling) secondary antibody (Millipore Corporation,
Figure 3. Semiquantitative reverse transcription-polymerase chain reaction (RT-PCR) of P2X1 receptor (P2X1R) mRNA in right ventricle (RV) and left ventricle (LV) of the heart, and in lung, kidney, liver and brain in rats exposed to hypobaric hypoxia. To derive the data shown in this Figure, the actual “P2X1R mRNA/18S rRNA” value obtained for a given tissue at “0 days” in 1 individual rat was expressed as “1.0”, and all other values for that tissue were expressed in a relative manner (n=5 for each group). *P<0.05 vs. value at time 0.

Circulation Journal Vol.75, April 2011
949P2X4R in Hypobaric Hypoxic Rats
Billerica, Massachusetts, USA), the blots were visualized by means of an ECL Plus Western Blotting Detection System (GE Healthcare UK Ltd, Buckinghamshire, UK).
ImmunohistochemistryThe indirect immunoperoxidase method was applied to 4 μm-thick formalin-embedded paraffin sections.7 Primary rabbit polyclonal antibodies against P2X4R protein (Almone Labs Ltd) diluted at 1:250 and horseradish peroxidase-labeled sec-ondary antibody against rabbit immunoglobulins (EnVision + Rabbit/HRP; Dako Cytomation) diluted at 1:250 were used. For the negative control, the incubation step with the primary antibody was omitted.
Statistical AnalysisThe results are expressed as mean ± SEM. Fisher’s protected least significant difference (Fisher’s PLSD) test or Scheffé’s test was applied to the data when significant F-ratios were obtained in an analysis of variance. Differences were consid-ered significant at P<0.05.
ResultsGrowth Characterization and Hemodynamic StudyThe body weight of rats exposed to hypobaric hypoxia decreased from 335.5±6.3 g (control rats, n=10) to 279.9±5.2 g (n=10) at 21 days of exposure, while the hematocrit increased from 42.2±0.1% (control rats, n=6) to 60.4±0.5% (n=6) at 21 days of exposure. The PAPm increased from 14.7±1.0 mmHg (control rats, n=6) to 24.5±2.7 mmHg (n=6) at 21 days of hypobaric hypoxic exposure, although SAPm did not change (as previously described in ref. no.7). Among the cardioven-tricular measurements, the ratios of RV over LV + S (RV/LV + S) and RV over body weight (RV/BW) were increased from 0.263±0.004 and 0.055±0.005% (control rats, each n=10–11), respectively, to 0.573±0.018 and 0.120±0.002% (at 21 days of exposure, each n=10–11), respectively, indicat-ing right ventricular hypertrophy (as also previously described in ref. no.7).
Expression of P2X4R mRNASemiquantitative RT-PCR was performed on the heart, lung, brain, kidney and liver. P2X4R mRNA was detected in all 6 tissues obtained from control rats. When we performed semi-quantitative RT-PCR on these tissues, relatively high levels
Figure 4. Semiquantitative reverse transcription-polymerase chain reaction (RT-PCR) of P2Y2 receptor (P2Y2R) mRNA in right ventricle (RV) and left ventricle (LV) of the heart, and in lung, kidney, liver and brain in rats exposed to hypobaric hypoxia. To derive the data shown in this Figure, the actual “P2Y2R mRNA/18S rRNA” value obtained for a given tissue at “0 days” in 1 in-dividual rat was expressed as “1.0”, and all other values for that tissue were expressed in a relative manner (n=5 for each group). *P<0.05 vs. value at time 0.
Circulation Journal Vol.75, April 2011
950 OHATA Y et al.
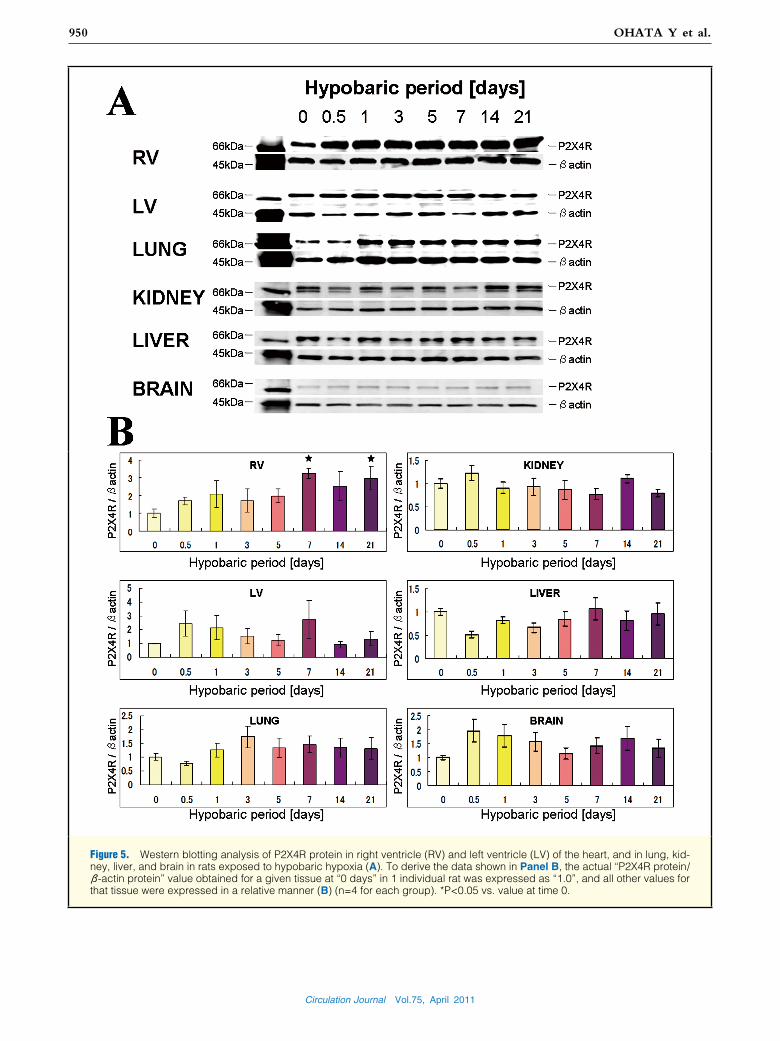
Figure 5. Western blotting analysis of P2X4R protein in right ventricle (RV) and left ventricle (LV) of the heart, and in lung, kid-ney, liver, and brain in rats exposed to hypobaric hypoxia (A). To derive the data shown in Panel B, the actual “P2X4R protein/ β-actin protein” value obtained for a given tissue at “0 days” in 1 individual rat was expressed as “1.0”, and all other values for that tissue were expressed in a relative manner (B) (n=4 for each group). *P<0.05 vs. value at time 0.

Circulation Journal Vol.75, April 2011
951P2X4R in Hypobaric Hypoxic Rats
of mRNA were detected in the kidney and lung, intermediate levels in the brain, LV, and liver, and a low level in the RV (Figure 1A).
In rats exposed to hypobaric hypoxia, the P2X4R mRNA level in the RV was, or tended to be, increased on day 1 and from day 5 to 14 of exposure to hypobaric hypoxia (Figure 2). There was a significant difference between day 1 and day 0 (P<0.05), and between day 14 and day 0 (P<0.05), and bor-derline significance between day 5 and day 0, and between day 7 and day 0 of exposure to hypobaric hypoxia (P=0.06 and P=0.08, respectively). In the LV, the level was signifi-cantly decreased at both 7 days and 21 days of exposure to hypobaric hypoxia (each, P<0.05 vs. control). P2X4R mRNA levels did not change significantly in the lung, kidney, liver or brain.
Expressions of P2X1R and P2Y2R mRNALikewise, semiquantitative RT-PCR for P2X1R and P2Y2R mRNA was performed on the above-mentioned 6 tissues. In the case of P2X1R mRNA, the lung exhibited a high level, the brain, RV, LV and kidney intermediate levels, and the liver a low level (Figure 1B). In the case of the P2Y2R mRNA level, the liver was high, the brain, lung and kidney were intermedi-
ate, and the RV and LV were low (Figure 1C).In rats exposed to hypobaric hypoxia, the P2X1R mRNA
level in the RV was significantly increased at 1, 14 and 21 days’ exposure to hypobaric hypoxia (each, P<0.05 vs. con-trol, Figure 3). In the LV and liver, the level was significantly increased at 7 days or 1 day of exposure to hypobaric hypoxia, respectively (each, P<0.05 vs. control). In contrast, the level was significantly decreased throughout the period of expo-sure to hypobaric hypoxia in the lung (each day, P<0.05 vs. control), and at 21 days of exposure in the kidney (P<0.05 vs. control). In the brain, the level did not change.
In the RV and LV, the P2Y2R mRNA level did not change, while in the lung and liver, the level was significantly increased at 3–7 days and 1–3 days, respectively, of exposure to hypobaric hypoxia (Figure 4; each, P<0.05 vs. control). The level was significantly decreased at 1 day and 7–21 days’ exposure to hypobaric hypoxia in the brain (each, P<0.05 vs. control), and throughout the period of exposure to hypobaric hypoxia in the kidney (each day, P<0.05 vs. control).
Expression of P2X4R ProteinWestern blotting analysis for P2X4R protein was performed on the RV, LV, lung, kidney, liver and brain. P2X4R protein
Figure 6. Western blotting analysis of P2X1R (A) and P2Y2R (B) proteins in right ventricle (RV) and left ventricle (LV) of the heart, and in the lung in rats exposed to hypobaric hypoxia. To derive the data, the actual “P2X1R protein/β-actin protein” value or “P2Y2R protein/β-actin protein” value obtained for a given tissue at “0 days” in 1 individual rat was expressed as “1.0”, and all other values for that tissue were expressed in a relative manner (n=4 for each group). *P<0.05 vs. value at time 0.
Circulation Journal Vol.75, April 2011
952 OHATA Y et al.
was detected in all these organs. In the RV, P2X4R protein was significantly increased at 7 days and 21 days of exposure to hypobaric hypoxia (each, P<0.05 vs. control), while in the other organs its level did not change (Figure 5).
Expressions of P2X1R and P2Y2R ProteinsWestern blotting analyses for P2X1R and P2Y2R were per-formed on the RV, LV and lung. In the RV, P2X1R protein was significantly increased at 21 days of exposure to hypo-baric hypoxia (P<0.05 vs. control), while in the LV it was significantly decreased at 14 days of exposure to hypobaric hypoxia (P<0.05 vs. control). In the lung, its level did not change (Figure 6A).
In the RV and LV, P2Y2R protein did not change, while in the lung its level was significantly increased at 3, 5, 14 and 21 days of exposure to hypobaric hypoxia (each, P<0.05 vs. control) (Figure 6B).
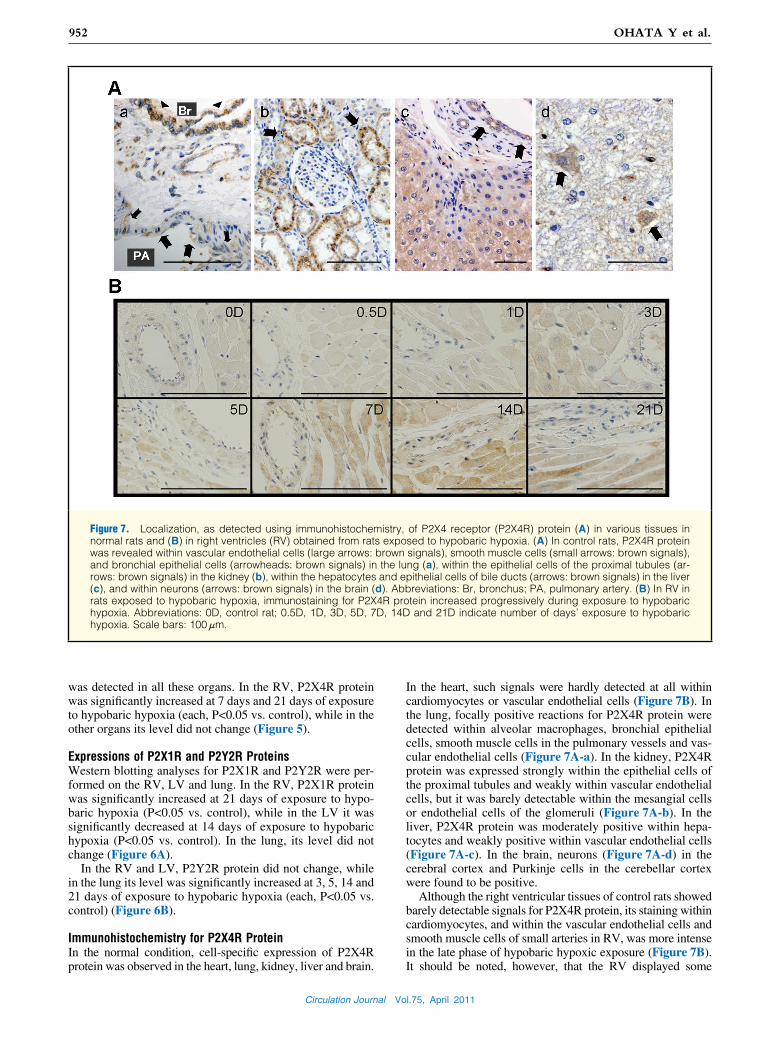
Immunohistochemistry for P2X4R ProteinIn the normal condition, cell-specific expression of P2X4R protein was observed in the heart, lung, kidney, liver and brain.
In the heart, such signals were hardly detected at all within cardiomyocytes or vascular endothelial cells (Figure 7B). In the lung, focally positive reactions for P2X4R protein were detected within alveolar macrophages, bronchial epithelial cells, smooth muscle cells in the pulmonary vessels and vas-cular endothelial cells (Figure 7A-a). In the kidney, P2X4R protein was expressed strongly within the epithelial cells of the proximal tubules and weakly within vascular endothelial cells, but it was barely detectable within the mesangial cells or endothelial cells of the glomeruli (Figure 7A-b). In the liver, P2X4R protein was moderately positive within hepa-tocytes and weakly positive within vascular endothelial cells (Figure 7A-c). In the brain, neurons (Figure 7A-d) in the cerebral cortex and Purkinje cells in the cerebellar cortex were found to be positive.
Although the right ventricular tissues of control rats showed barely detectable signals for P2X4R protein, its staining within cardiomyocytes, and within the vascular endothelial cells and smooth muscle cells of small arteries in RV, was more intense in the late phase of hypobaric hypoxic exposure (Figure 7B). It should be noted, however, that the RV displayed some
Figure 7. Localization, as detected using immunohistochemistry, of P2X4 receptor (P2X4R) protein (A) in various tissues in normal rats and (B) in right ventricles (RV) obtained from rats exposed to hypobaric hypoxia. (A) In control rats, P2X4R protein was revealed within vascular endothelial cells (large arrows: brown signals), smooth muscle cells (small arrows: brown signals), and bronchial epithelial cells (arrowheads: brown signals) in the lung (a), within the epithelial cells of the proximal tubules (ar-rows: brown signals) in the kidney (b), within the hepatocytes and epithelial cells of bile ducts (arrows: brown signals) in the liver (c), and within neurons (arrows: brown signals) in the brain (d). Abbreviations: Br, bronchus; PA, pulmonary artery. (B) In RV in rats exposed to hypobaric hypoxia, immunostaining for P2X4R protein increased progressively during exposure to hypobaric hypoxia. Abbreviations: 0D, control rat; 0.5D, 1D, 3D, 5D, 7D, 14D and 21D indicate number of days’ exposure to hypobaric hypoxia. Scale bars: 100 μm.

Circulation Journal Vol.75, April 2011
953P2X4R in Hypobaric Hypoxic Rats
regional variations in staining intensity. In the other organs, no changes in the staining for P2X4R protein were seen throughout the period of exposure to hypobaric hypoxia.
DiscussionOur purpose in exposing rats to hypobaric hypoxia was to look for possible relationships between changes in the expres-sion of P2X4R (mRNA and/or protein) and hypoxia-induced pulmonary hypertension. We confirmed that an elevation in pulmonary arterial pressure and right ventricular hypertrophy occurred upon exposure to hypobaric hypoxia. In the RV of the heart, we observed overexpression of P2X4R mRNA in the middle period of exposure to hypobaric hypoxia (using semiquantitative RT-PCR analysis), and also an increased expression of P2X4R protein in the middle-to-late period of exposure to hypobaric hypoxia (using Western blotting analy-sis). In addition, we examined the levels of the mRNA (using semiquantitative RT-PCR analysis) and proteins (using West-ern blotting analysis) for P2X1R and P2Y2R. In RV, we observed overexpression of P2X1R mRNA in the early and late periods of exposure to hypobaric hypoxia. Moreover, we detected an increased expression of P2X1R protein (using Western blotting analysis) in RV on day 21 of hypobaric hypoxia. Conceivably, these changes could play important roles in modulating the pulmonary hypertension induced by hypobaric hypoxia, although we have no direct evidence for such a causal link.
In normal rats, we demonstrated that expressions of P2X4R mRNA and protein were present in multiple organs (using semiquantitative RT-PCR and immunohistochemistry). In our semiquantitative RT-PCR, the kidney and lung exhibited high levels of P2X4R mRNA, but the heart, especially RV, a low level. Likewise, our immunohistochemical findings revealed expressions within endothelial cells in some organs, within alveolar macrophages and bronchial epithelial cells in the lung, within the epithelial cells of proximal tubules, and within hepatocytes, neurons, and Purkinje cells, but only a very weak expression within cardiomyocytes. These results are consis-tent with the Western blotting and immunohistochemical results obtained in normal rats by Bo et al.24 In addition, we observed P2X1R and P2Y2R mRNA in several organs, including the brain and liver, in normal rats (using semiquan-titative RT-PCR), and we demonstrated that in the normal condition, P2X1R was abundant in the lung, whereas P2Y2R was abundant in the liver. In contrast, Rice et al25 reported P2Y2R mRNA expression in heart, kidney, lung, spleen and testis, but no expression in rat brain or liver. As the reason for this discrepancy is unclear, further studies will be needed.
In the present examination of the heart, the levels of P2X4R mRNA and protein were quite low in control rats (by our semiquantitative RT-PCR and Western blotting analysis). During exposure to hypobaric hypoxia: (a) the mRNA level in RV displayed a significant increase in the middle period of the exposure, and (b) the protein level showed a significant increase in the late period of the exposure. Furthermore, these changes were quite well paralleled by increases in the inten-sity of immunohistochemical staining within the cytoplasm of cardiomyocytes in RV. In contrast, the levels of P2X4R mRNA and protein did not change significantly in the lung, kidney, liver or brain, although in LV the mRNA level decreased in the middle-to-late period of exposure to hypo-baric hypoxia. Such a functional remodeling occurring only in RV (viz. induction of P2X4R) might be a characteristic of the pulmonary pressure overload resulting from hypoxia.
Very recently, Sonin et al – who examined an infarction model created by ligation of the left anterior descending artery in transgenic mice (Tg) exhibiting a cardiac-specific overex-pression of P2X4R – demonstrated that Tg animals displayed a significantly enhanced cardiac contractile function com-pared with wild-type animals.21 They therefore suggested that an enhanced contractile function in the non-infarcted area of the Tg heart was most likely an important rescue mechanism. Furthermore, Shen et al. demonstrated that the cardiac myo-cyte P2XR was upregulated in an experimental model of hypertrophy and heart failure, and moreover that chronic in vivo administration of a P2XR agonist could rescue the hypertrophic phenotype of the cardiomyopathic animals and prolong their longevity.26 Therefore, they suggested that aug-mentation of the cardiac myocyte P2X current could reverse or attenuate cardiac hypertrophy and failure. We therefore speculate that the induction of P2X4R in RV we observed in our model might have a beneficial effect on the pulmonary hypertension induced by hypobaric hypoxia.
Recently, Yamamoto et al showed that endothelial cells from mouse pulmonary microvessels expressed P2X4R.15 They also demonstrated that P2X4R-knockout mice had higher pulmonary blood pressure, and that the flow-induced release of the potent vasodilator nitric oxide, and the conse-quent vasodilation, were impaired in several arteries and arterioles.16 On the basis of their data, they suggested that endothelial P2X4R might be crucial for the flow-sensitive mechanisms that regulate blood pressure and vascular remod-eling. In the present study, however, we did not detect an induction of P2X4R mRNA or protein in the lung during exposure to hypobaric hypoxia, even though it is well known that pulmonary vascular resistance increases upon exposure to hypobaric hypoxia.1–7 However, the maximal PAPm was 24.5 mmHg at 21 days of hypobaric hypoxic exposure in our study: not an excessively high value. We previously exam-ined BNP induction in RV in rats exposed for 12 weeks to the same hypobaric hypoxia as that employed here, and we found that BNP protein levels were significantly elevated only at 12-weeks’ exposure (vs. those of control rats). Thus, the degree of P2X4R induction we observed might have been limited by near-or-subthreshold levels of pulmonary arterial pressure overload being obtained upon exposure to hypo-baric hypoxia equivalent to 5,500 m in altitude. In other words, P2X4R induction in the lung might require a more severe PAP overload.
In addition, we detected overexpression of P2X1R and P2Y2R mRNA (using semiquantitative RT-PCR) and an increased expression of P2X1R protein (using Western blot-ting analysis) in RV. These receptors might also have a benefi-cial effect on the hypobaric hypoxic pulmonary hypertension. Concerning P2Y2R, Wang and coworkers, who examined the role of shear stress on smooth muscle cells in the human umbilical vein, observed that P2Y2R mRNA and protein were upregulated.27 They suggested that an increased expression of P2Y2R might promote the induction of vascular remodeling by shear stress because P2Y2R stimulate the growth and migration of smooth muscle cells. In the present study, we observed overexpression of both P2Y2R mRNA and protein in the lung. In our previous study on rats exposed for 12 weeks to the same hypobaric hypoxia as that employed here, we observed structural remodeling of all muscular arteries, partially muscular arteries, and non-muscular arteries within the lobules of the lung.4 Such vascular remodeling could be partly due to an increased expression of P2Y2R. Further-more, in the early-to-middle period of exposure to hypobaric

Circulation Journal Vol.75, April 2011
954 OHATA Y et al.
hypoxia we noted overexpression of P2X1R mRNA in the LV and liver, and overexpression of P2Y2R mRNA in the liver. In contrast, P2X1R and P2Y2R mRNA levels were decreased in the lung and kidney, and in the brain and kid-ney, respectively, while P2X1R protein was decreased in the LV. ATP induces both constrictor and dilator responses in vessels. That is, ATP acts on smooth muscle cells to cause contraction and on endothelial cells to cause vasodilation. Consequently, further studies will be needed before we can hope to understand the complex purinergic control of vascu-lar tone and its role in hypobaric hypoxia.
In conclusion, we have provided evidence that during 21 days’ exposure to hypobaric hypoxia (equivalent to the 5,500 m altitude level), increases in the synthesis of P2X4R and P2X1R in the RV occur alongside the increase in PAP and the development of right ventricular hypertrophy. These changes could conceivably play an important role in modu-lating the RV hypertrophy that occurs due to pulmonary hypertension during hypobaric hypoxia. However, we did not try to determine whether the increased P2X4R expression does indeed perform that function during hypobaric hypoxia. Recently, it has been reported that ATP and other nucleotides (adenosine diphosphate, uridine triphosphate, and uridine diphosphate) play important roles in cardiovascular physiol-ogy and pathophysiology, acting via several P2X and P2Y receptors.28 Further studies will be needed to determine which, if any, effects of hypobaric hypoxia are exerted via the purinergic P2 nucleotide receptors located within the car-diovascular system.
DisclosureConflict of interest: There is no conflict of interest for this study.
References 1. Heath D, Williams DR. Man at high altitude. The pathophysiology
of acclimatization and adaptation. Edinburgh: Churchill Livingstone, 1981.
2. Meyrick B, Reid L. The effect of continued hypoxia on rat pulmo-nary arterial circulation: An ultrastructural study. Lab Invest 1978; 38: 188 – 200.
3. Reid LM. Structure and function in pulmonary hypertension; new perceptions. Chest 1986; 89: 279 – 288.
4. Nakanishi K, Tajima F, Osada H, Nakamura A, Yagura S, Kawai T, et al. Pulmonary vascular responses in rats exposed to chronic hypobaric hypoxia at two different altitude levels. Pathol Res Pract 1996; 192: 1 – 12.
5. Nakanishi K, Tajima F, Nakata Y, Osada H, Tachibana S, Kawai T, et al. Expression of endothelin-1 in rats developing hypobaric hypoxia-induced pulmonary hypertension. Lab Invest 1999; 79: 1347 – 1357.
6. Nakanishi K, Tajima F, Itoh H, Nakata Y, Osada H, Hama N, et al. Changes in atrial natriuretic peptide and brain natriuretic peptide associated with hypobaric hypoxia-induced pulmonary hypertension in rats. Virchows Archiv 2001; 439: 808 – 817.
7. Nakanishi K, Osada H, Uenoyama M, Kanazawa F, Ohrui N, Masaki Y, et al. Expressions of adrenomedullin mRNA and protein in rats with hypobaric hypoxia-induced pulmonary hypertension. Am J Physiol Heart Circ Physiol 2004; 286: H2159 – H2168.
8. Richalet JP, Kacimi R, Antezana AM. The control of cardiac chro-
notropic function in hypobaric hypoxia. Int J Sports Med 1992; 13: 22 – 24.
9. Höpfl G, Ogunshola O, Gassmann M. Hypoxia and high altitude: The molecular response. Adv Exp Med Biol 2003; 543: 89 – 115.
10. Do.e Z, Fukumoto Y, Takaki A, Tawara S, Ohashi J, Nakano M, et al. Evidence for Rho-kinase activation in patients with pulmonary arterial hypertension. Circ J 2009; 73: 1731 – 1739.
11. Koyama T, Ono K, Watanabe H, Ohba T, Murakami M, Iino K, et al. Molecular and electrical remodeling of L- and T-type Ca2+ channels in rat right atrium with monocrotaline-induced pulmo-nary hypertension. Circ J 2009; 73: 256 – 263.
12. Zhang E, Jiang B, Yokochi A, Marytama J, Mitani Y, Ma N, et al. Effect of all-trans-retinoic acid on the development of chronic hypoxia-induced pulmonary hypertension. Circ J 2010; 74: 1696 – 1703.
13. Abbracchio MP, Burnstock G, Boeynaems JM, Barnard EA, Boyer JL, Kennedy C, et al. Update of the P2Y G protein-coupled nucleo-tide receptors: From molecular mechanisms and pathophysiology to therapy. Pharmacol Rev 2006; 59: 281 – 341.
14. North RA. Molecular physiology of P2X receptors. Physiol Rev 2002; 82: 1013 – 1067.
15. Yamamoto K, Korenaga R, Kamiya A, Ando J. Fluid shear stress activates Ca2+ influx into human endothelial cells via P2X4 purino-ceptors. Circ Res 2000; 87: 385 – 391.
16. Yamamoto K, Korenaga R, Kamiya A, Qi Z, Sokabe M, Ando J. P2X4 receptors mediate ATP-induced calcium influx in human vascular endothelial cells. Am J Physiol Heart Circ Physiol 2000; 279: 285 – 292.
17. Yamamoto K, Sokabe T, Ohura N, Nakatsuka H, Kamiya A, Ando J. Endogenously released ATP mediates shear stress-induced Ca2+ influx into pulmonary artery endothelial cells. Am J Physiol Heart Circ Physiol 2003; 285: H793 – H803.
18. Yamamoto K, Sokabe T, Matsumoto T, Yoshimura K, Shibata M, Ohura N, et al. Impaired flow-dependent control of vascular tone and remodeling in P2X4-deficient mice. Nat Med 2006; 12: 133 – 137.
19. Shen JB, Pappano A, Liang BT. Extracellular ATP-stimulated cur-rent in wild type and P2X4 receptor transgenic mouse ventricular myocytes: Implication for a cardiac physiologic role of P2X4 recep-tors. FASEB J 2006; 20: 277 – 284.
20. Hu B, Mei Q, Smith E, Barry WH, Liang BT. A novel cardiac ino-tropic phenotype with cardiac transgenic expression of human P2X4 receptor transgenic mouse. FASEB J 2001; 15: 2739 – 2741.
21. Sonin D, Zhou SY, Cronin C, Sonina T, Wu J, Jacobson KA, et al. Role of P2X purinergic receptors in the rescue of ischemic heart failure. Am J Physiol Heart Circ Physiol 2008; 295: 1191 – 1197.
22. Schlüter H, Offers E, Brüggemann G, Giet M, Tepel M, Nordhoff E, et al. Diadenosine phosphates and the physiological control of blood pressure. Nature 1994; 367: 186 – 188.
23. Hou M, Malmsjö M, Möller S, Pantev E, Bergdahl A, Zhao XH, et al. Increase in cardiac P2X1- and P2Y2-receptor mRNA levels in congestive heart failure. Life Sci 1999; 65: 1195 – 1206.
24. Bo X, Kim M, Nori SL, Schoepfer R, Burnstock G, North RA. Tis-sue distribution of P2X4 receptors studied with an ectodomain anti-body. Cell Tissue Res 2003; 313: 159 – 165.
25. Rice WR, Burton FM, Fiedeldey DT. Cloning and expression of the alveolar type II cell P2u-purinergic receptor. Am J Respir Cell Mol Biol 1995; 12: 27 – 32.
26. Shen JB, Cronin C, Sonin D, Joshi BV, Gongora Nieto M, Harrison D, et al. P2X purinergic receptor-mediated ionic current in cardiac myocytes of calsequestrin model of cardiomyopathy: Implications for the treatment of heart failure. Am J Physiol Heart Circ Physiol 2007; 292: 1077 – 1084.
27. Wang L, Andersson M, Karlsson L, Watson MA, Cousens DJ, Jern S, et al. Increased mitogenic and decreased contractile P2 receptors in smooth muscle cells by shear stress in human vessels with intact endothelium. Arterioscler Thromb Vasc Biol 2003; 23: 1370 – 1376.
28. Erlinge D, Burnstock G. P2 receptors in cardiovascular regulation and disease. Purinergic Signal 2008; 4: 1 – 20.