forward genetics and reverse genetics forward genetics: from phenotype to gene sequence reverse...
TRANSCRIPT


Forward genetics and reverse genetics
Forward genetics: from phenotype to gene sequence
Reverse genetics : from gene sequence to phenotype
Gene Mutation Phenotype
Mutation Phenotype Gene
There are basically two ways to link the sequence and function of a specific gene: forward and reverse genetics
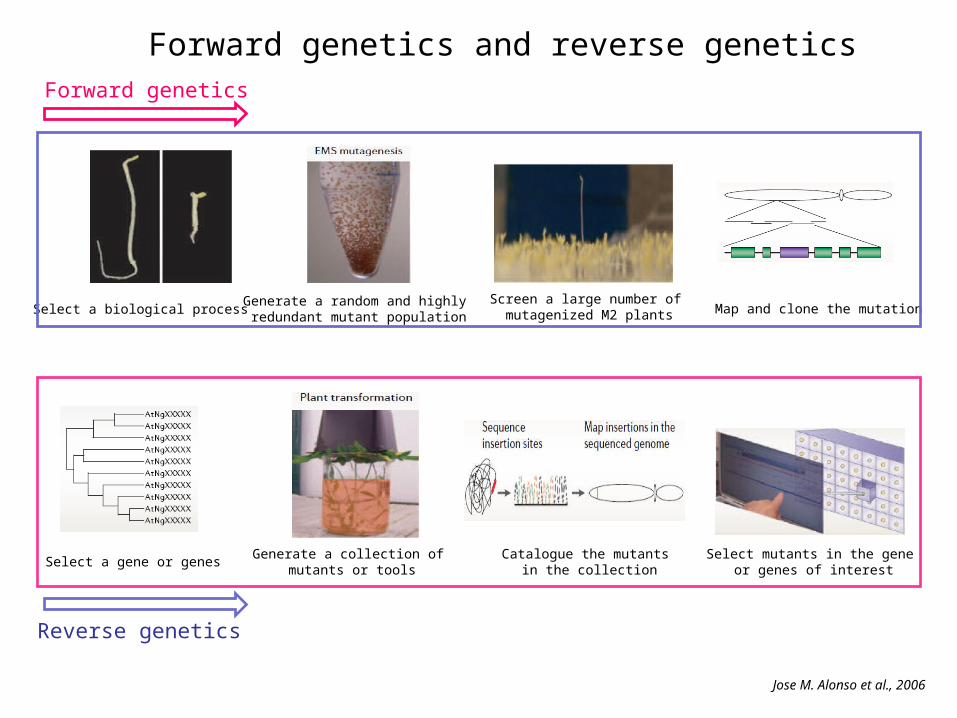
Forward genetics and reverse genetics
Jose M. Alonso et al., 2006
Forward genetics
Reverse genetics
Select a biological processGenerate a random and highly redundant mutant population
Screen a large number of mutagenized M2 plants Map and clone the mutation
Select a gene or genesGenerate a collection of
mutants or toolsCatalogue the mutants
in the collectionSelect mutants in the gene
or genes of interest
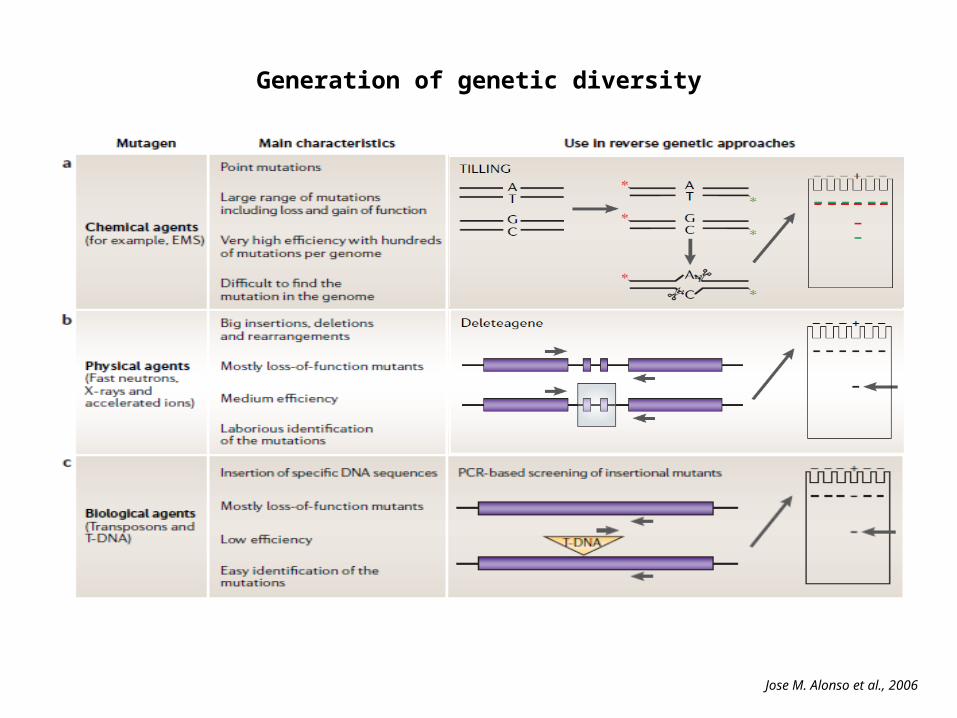
Generation of genetic diversity
Jose M. Alonso et al., 2006

Tandem gene multiplications pose a challenge for functional analysis
In sequenced plant genomes, 15% or more of the identified genes are members of tandem-arrayed gene Families.
Assigning function to unknown plant genes is to study phenotypic changes associated with knockout mutations.
Mutating only one gene in a duplicated pair often produces no measurable phenotype, this poses a particular challenge for functional analysis.
It is often necessary to knock out more than one member of a plant gene family to obtain a mutant phenotype.
Here describe alternate strategies that can be used to identify knockouts of two or more tandem-arrayed plant genes for functional analysis.
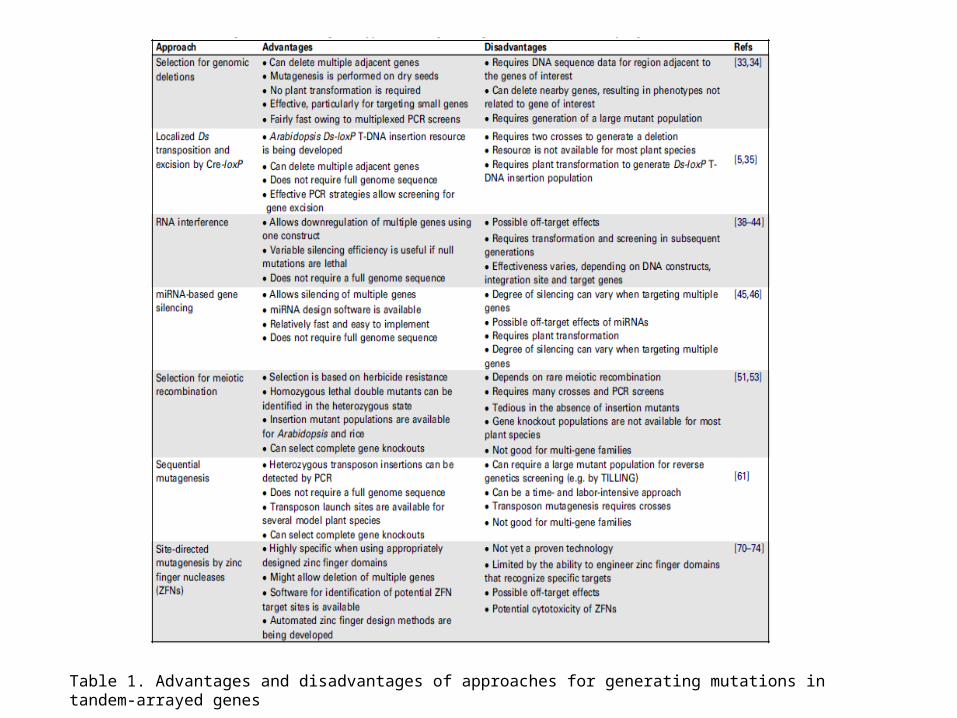
Table 1. Advantages and disadvantages of approaches for generating mutations in tandem-arrayed genes
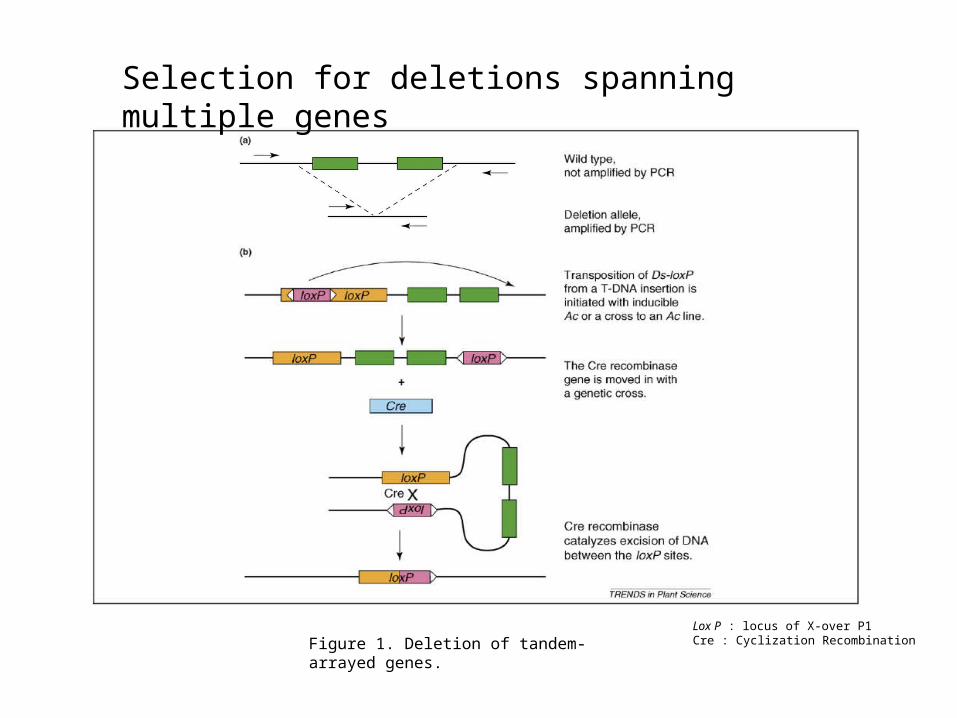
Figure 1. Deletion of tandem-arrayed genes.
Selection for deletions spanning multiple genes
Lox P : locus of X-over P1Cre : Cyclization Recombination

Silencing gene families by RNA interference
RNA interference allows down-regulation of gene expression in transgenic plants, including the genes of interest are located in tandem arrays.
RNA interference was used to silence all members of the gene family.
Silencing efficiency depends on the type of construct that is used, the site of integration, and the particular target genes.
Gene silencing is generally not 100% effective, mutant phenotypes that require the complete knockout of a gene family might not be detected using the described silencing approaches.

Selection for meiotic recombination between adjacent genes
Figure 2. Selection for recombination between T-DNA or transposon insertions

Sequential mutagenesis to create a double mutant
Figure 3. Localized transposon mutagenesis. Ds elements tend to transpose to nearby genetic locations, allowing screens for insertions in adjacent genes. Imperfect DNA repair can leave the gene at the original insertion site inactivated .
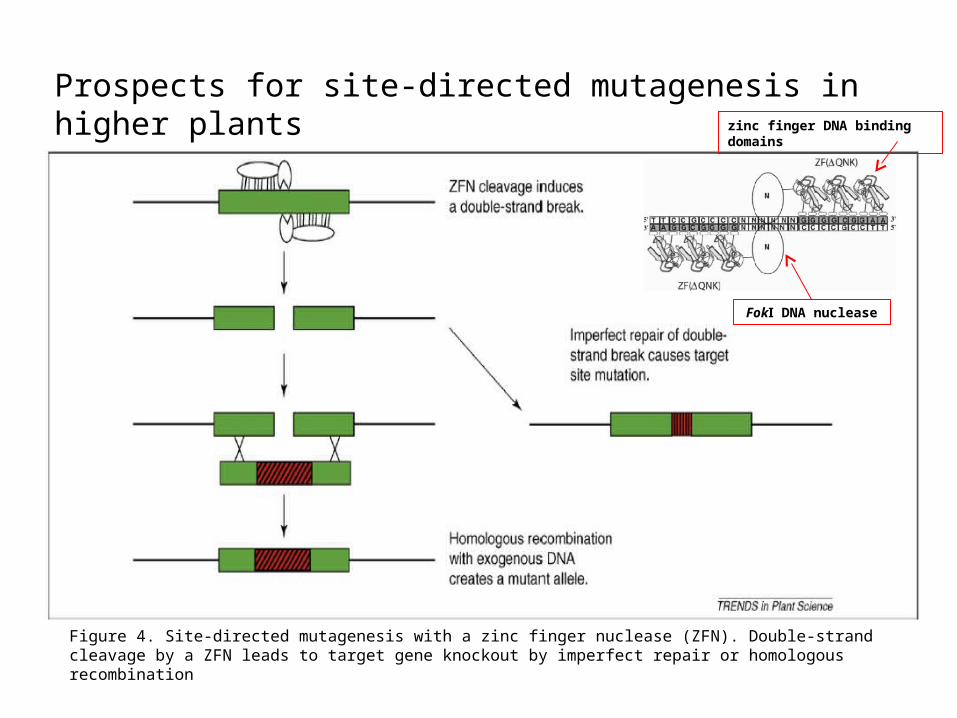
Figure 4. Site-directed mutagenesis with a zinc finger nuclease (ZFN). Double-strand cleavage by a ZFN leads to target gene knockout by imperfect repair or homologous recombination
Prospects for site-directed mutagenesis in higher plants zinc finger DNA binding
domains
FokⅠ DNA nuclease

High-throughput TILLING
Trenton Colbert et al., 2001

Total 50,000 lines
Xin Li et al., 2001