fundamental characteristics of the immunoglobulin vh ... · fundamental characteristics of the...
TRANSCRIPT

of July 11, 2018.This information is current as
and Camelidsin Comparison with Those of Humans, Mice,
Repertoire of ChickensHImmunoglobulin VFundamental Characteristics of the
Orla Cunningham and William J. J. FinlayTchistiakova, Laird Bloom, Janet Paulsen, Davinder Gill, Shúilleabháin, Bénédicte Autin, Emma Cummins, LioudmilaJ. Fennell, Alfredo Darmanin-Sheehan, Deirdre Ní Leeying Wu, Katarzyna Oficjalska, Matthew Lambert, Brian
http://www.jimmunol.org/content/188/1/322doi: 10.4049/jimmunol.1102466November 2011;
2012; 188:322-333; Prepublished online 30J Immunol
MaterialSupplementary
6.DC1http://www.jimmunol.org/content/suppl/2011/11/30/jimmunol.110246
Referenceshttp://www.jimmunol.org/content/188/1/322.full#ref-list-1
, 15 of which you can access for free at: cites 70 articlesThis article
average*
4 weeks from acceptance to publicationFast Publication! •
Every submission reviewed by practicing scientistsNo Triage! •
from submission to initial decisionRapid Reviews! 30 days* •
Submit online. ?The JIWhy
Subscriptionhttp://jimmunol.org/subscription
is online at: The Journal of ImmunologyInformation about subscribing to
Permissionshttp://www.aai.org/About/Publications/JI/copyright.htmlSubmit copyright permission requests at:
Email Alertshttp://jimmunol.org/alertsReceive free email-alerts when new articles cite this article. Sign up at:
Print ISSN: 0022-1767 Online ISSN: 1550-6606. Immunologists, Inc. All rights reserved.Copyright © 2011 by The American Association of1451 Rockville Pike, Suite 650, Rockville, MD 20852The American Association of Immunologists, Inc.,
is published twice each month byThe Journal of Immunology
by guest on July 11, 2018http://w
ww
.jimm
unol.org/D
ownloaded from
by guest on July 11, 2018
http://ww
w.jim
munol.org/
Dow
nloaded from

The Journal of Immunology
Fundamental Characteristics of the Immunoglobulin VH
Repertoire of Chickens in Comparison with Those ofHumans, Mice, and Camelids
Leeying Wu,*,1 Katarzyna Oficjalska,†,1 Matthew Lambert,† Brian J. Fennell,†
Alfredo Darmanin-Sheehan,† Deirdre Nı Shuilleabhain,† Benedicte Autin,† Emma Cummins,†
Lioudmila Tchistiakova,* Laird Bloom,* Janet Paulsen,* Davinder Gill,* Orla Cunningham,†
and William J. J. Finlay†
Examination of 1269 unique naive chicken VH sequences showed that the majority of positions in the framework (FW) regions
were maintained as germline, with high mutation rates observed in the CDRs. Many FW mutations could be clearly related to the
modulation of CDR structure or the VH–VL interface. CDRs 1 and 2 of the VH exhibited frequent mutation in solvent-exposed
positions, but conservation of common structural residues also found in human CDRs at the same positions. In comparison with
humans and mice, the chicken CDR3 repertoire was skewed toward longer sequences, was dominated by small amino acids (G/S/
A/C/T), and had higher cysteine (chicken, 9.4%; human, 1.6%; and mouse, 0.25%) but lower tyrosine content (chicken, 9.2%;
human, 16.8%; and mouse 26.4%). A strong correlation (R2 = 0.97) was observed between increasing CDR3 length and higher
cysteine content. This suggests that noncanonical disulfides are strongly favored in chickens, potentially increasing CDR stability
and complexity in the topology of the combining site. The probable formation of disulfide bonds between CDR3 and CDR1, FW2,
or CDR2 was also observed, as described in camelids. All features of the naive repertoire were fully replicated in the target-
selected, phage-displayed repertoire. The isolation of a chicken Fab with four noncanonical cysteines in the VH that exhibits 64 nM
(KD) binding affinity for its target proved these constituents to be part of the humoral response, not artifacts. This study supports
the hypothesis that disulfide bond-constrained CDR3s are a structural diversification strategy in the restricted germline v-gene
repertoire of chickens. The Journal of Immunology, 2012, 188: 322–333.
Monoclonal Abs have become the most rapidly expandingarea in biotherapeutic drug discovery (1, 2). In Ab drugdiscovery programs, panels of lead Abs are often re-
quired that exhibit high affinity, react with multiple orthologs ofthe target, have broad epitope coverage, and most importantly,potently modulate the biological function of their target. It is alsobeneficial for downstream development into viable therapeuticagents that the resulting Abs should have uniform stability andexpression characteristics. A relatively untapped source of suchAbs is the immune repertoire of animals whose v-gene germlinediversity is restricted, but still highly homologous to humansequences. A number of biotherapeutic research groups have,therefore, developed an interest in an underused option in bio-therapeutic drug discovery: high-affinity, potently neutralizingrAbs from chickens (3–5). This approach exploits the fact that
avians are phylogenetically distant from mammals but have the-oretically uniform, humanizable v-gene sequences (3, 4).The generation of Ab diversity in chickens is well characterized
at the genomic level (6–8), but despite many years of study of thismodel species, the true amino acid content of the final functionalrepertoire has not been extensively examined in the way that it hasfor primates and rodents (9–14). The VH domain of Abs is typi-cally critical to Ab specificity, and the humanization of Ab ther-apeutics is greatly aided by understanding the structural charac-teristics of the VH repertoire, from which the lead clone is derived.We felt, therefore, that a detailed understanding of the funda-mental amino acid content of the adult chicken repertoire wouldshed light on the similarities and differences between the v-genediversification mechanisms of humans, rodents, and avians. Itwould also greatly inform our ability to harness some of theunique properties of chicken Abs. In addition, a clear view of thediversification mechanisms and final amino acid content used inthe VH CDRs is critical to the development of novel design fea-tures in synthetic Ab libraries (14–16).The Ig system of chickens is distinct from that of humans and
mice. Although chickens have structural and functional equivalentsof mammalian IgM, IgA, and IgG, they have not been found toproduce direct homologs of IgE or IgD, with IgM being the majorisotype expressed on the surface of their B cells (6). Indeed, it hasbeen proposed that chicken IgG, with its four Cg domains, may bea structural relative of both mammalian IgG and IgE subclasses(17). Studies on the underlying molecular biology of chickenv-gene diversification have led to some fascinating insights. Thesimple avian v-gene germline repertoire is in marked contrast to
*Global Biotherapeutics Technologies, Pfizer, Cambridge, MA 02140; and †Global Bio-therapeutics Technologies, Pfizer, Dublin 22, Ireland
1L.W. and K.O. contributed equally to this study.
Received for publication August 26, 2011. Accepted for publication November 1,2011.
Address correspondence and reprint requests to Dr. William J. J. Finlay, GlobalBiotherapeutics Technologies, Development Laboratory, Pfizer, Grange Castle Busi-ness Park, Clondalkin, Dublin 22, Ireland. E-mail address: [email protected]
The online version of this article contains supplemental material.
Abbreviations used in this article: FW, framework; IMGT, International Immunoge-netics Information System; scFv, single-chain Ab.
Copyright� 2011 by TheAmericanAssociation of Immunologists, Inc. 0022-1767/11/$16.00
www.jimmunol.org/cgi/doi/10.4049/jimmunol.1102466
by guest on July 11, 2018http://w
ww
.jimm
unol.org/D
ownloaded from

the system used by humans, mice, and primates, which involvea large set of sequences that are highly diverse in both sequence andstructure, and are predominantly capable of functional VDJ recom-bination (18, 19). In chickens, single functional v-genes exist forboth the L and H chains, containing unique VL–JL, VH–JH, andD segments (8, 20, 21). To use a restricted v-gene germline rep-ertoire and yet achieve the same level of Ig protection as in miceand primates, the chicken has evolved a distinctly different set ofdiversification mechanisms, including the gene conversion process(8, 22). The gene conversion process in chickens is a system similarto that observed in rabbits (23) in which a template v-gene is di-versified by the incorporation of segments from numerous upstreampseudogenes that lack recombination signal sequences.Despite the use of this very simple (essentially single framework
[FW]) germline v-gene system (6, 17, 24–26), chickens havea very adaptable Ab repertoire that is capable of generating high-affinity interactions with a broad range of Ags including proteins,peptides, and haptens (3, 27–29). This broad Ag recognition ca-pability, combined with simple isolation of multiortholog cross-reactive mAbs via technologies such as phage display (3, 30),have made chicken Abs a potentially valuable source of Ab ther-apeutics. In this study, we have therefore undertaken bioinfor-matics analyses to define the fundamental amino acid diversity ofVH sequences from the adult chicken spleen and bone marrowB cell repertoire of both naive and immunized animals.
Materials and MethodscDNA library generation and sequencing
Spleen and femur bone marrow were harvested from each of three adultWhite Leghorn chickens (Lampire). Total RNA was then prepared fromeach tissue sample using TRIzol reagent (Invitrogen), and cDNA wasgenerated from each RNA sample using oligo(dT) priming and the Su-perscript III First Strand Synthesis kit, according to the manufacturer’sinstructions (Invitrogen). Each tissue cDNAwas then used to create clonedlibraries of the VH gene repertoire, essentially as described previously (31,32). DNA sequencing was subsequently performed on randomly chosenclones from each tissue sample library (GATC).
Generation of VH repertoire amino acid sequence databases
To identify H chain v-gene sequences, Hidden Markov Model profiles wereconstructed using the HMMERBUILD program. The GENEWISE programwas then used to compare the sequenced cDNAs against the H chain HiddenMarkov Models and to produce multiple translated predictions. The besttranslated VH sequences were manually evaluated to remove partial, du-plicated, frameshifted, and stop codon-containing sequences to derive a setof full-length unique sequences. To compare the chicken CDR-H3 reper-toire with those of humans and mice, we also downloaded all availablesequences from the International Immunogenetics Information System(IMGT) database and ran the same predictions as above. Once the chicken,human, and mouse v-gene databases had been completed, amino acidusage at each position of the sequences were determined and visualizedusing an in-house developed Excel macro.
Selection of target-specific Abs from immune phage displaylibraries
Single-chain Ab (ScFv) libraries were generated from the spleen and bonemarrow cDNA samples of chickens immunized with 12 mammalian andnonmammalian protein Ags including cytokines, chemokines, extracellu-lar receptors, and purified Igs. Library generation, rescue, and selectionmethods used were essentially as described previously (33). All librarieswere rescued and selected separately on each target protein. Target-bindingspecificity in resulting phage clones was determined via scFv ELISAanalyses as described previously (34).
Conversion of cysteine variant scFv clone to human chimericFab format
ScFv clones of interest were synthesized as bicistronic Fab cassettes in theoptimal L chain–H chain orientation, essentially as previously described(31, 35), for cloning into the pWRIL-1 vector (32). The final Fab cas-
settes were synthesized using human Cl and CH1 sequences in the L andH chains, respectively. On both chains, the sequences encoding for C-terminal cysteine residues were omitted to maximize expression rate inEscherichia coli, because the natural affinity between the L and H chains issufficient to create functional Fabs (36).
Expression and purification of Fab proteins
Fab expression and purification was carried out as follows: E. coli clones ofinterest were inoculated into 5 ml 2YT broth containing 0.1% (w/v) glu-cose and 100 mg/ml carbenicillin. Cultures were allowed to grow shakingat 220 rpm at 37˚C for 16 h. These cultures were used to inoculate 500 ml2YT broth containing 100 mg/ml carbenicillin until OD600 values reached0.8–1.2. Protein expression was induced with 0.25 mM isopropyl b-D-thiogalactoside, and cultures were incubated at 28˚C shaking at 220 rpmfor 16 h. Cells were pelleted by centrifugation at 4000 3 g and resus-pended in 10 ml ice-cold periplasmic buffer (50 mM HEPES, 0.5 mMEDTA, and 20% [w/v] sucrose [pH 7.5]). A total of 10 ml of a 1/5 dilutionof periplasmic buffer was then added, and the samples were incubated onice for 30 min. Samples were centrifuged at 15,000 3 g, 4˚C for 15 min,and the supernatant was recovered. Fab proteins were purified in a two-stepprocedure via nickel affinity chromatography and size-exclusion chroma-tography on an Akta Explorer System as described previously (34).
Binding kinetics analysis
Surface plasmon resonance-based kinetic analysis was carried out at am-bient temperature on a Biacore T100 instrument using a CM5 chip coatedwith 2000 resonance units of IgM. Kinetics analyses were performed ata flow rate of 50 ml/min with a contact phase of 180 s and a dissociationphase of 180 s. Purified H2 Fab was used in the concentration range 125–7.81 nM. Affinity constants were determined by 1:1 global–fit analysis ofthe binding curves using Biacore evaluation software. All sensorgram datafitted well to the Langmuir 1:1 interaction models, and x2 values for allanalyses were ,0.3.
ResultsAnalysis of naive diversity in the FW regions of the chicken VH
repertoire
Spleen and bone marrow cDNA from three healthy adult chickenswere used to generate a naive repertoire rAb sequence library inE. coli. From each of these libraries, single clones were randomlypicked and subjected to DNA sequencing. A total of 1269 unique,in-frame, nonredundant clones were identified and used to performVH sequence alignments, identification of the FW and CDR regionsvia a hidden Markov model, and numbering according to the IMGTsystem for Ig structural definition (37).To evaluate the extent of diversification in the VH repertoire, the
percent deviation from the germline VH1 gene (8) was calculatedfor each amino acid position. This analysis indicated that thehighest mutation rates in the repertoire were observed within theCDRs (Fig. 1). More notable, however, was the marked conser-vation in the FW regions in which substitutions were generallyrare and predominantly conservative. In FW 1, only two fre-quently substituted positions were observed, namely 18A (33% G,9% T, and 1% V) and 25A (16% G). Interestingly, a third position,17R, was found to be predominantly glycine and not the germline-encoded arginine (95% G and 5% R).In FW 2, position 39M was found to be very highly conserved.
This residue is known to be buried in human VH structures (38, 39),so it is unsurprising that on the rare occasions where it was mu-tated in chickens, the substituting residues were usually also hy-drophobic (2% V, 1% I, and 0.5% L). In FW 2, only three po-sitions are substituted frequently: 40G, 52F, and 55G. Position 52Fis substituted at 19% Y and 38% W. Each of these three residueshave also been commonly observed at position 52 in human andmurine VH sequences, and 52F is known to be a key residue af-fecting the VH–VL interface (40). Position 52 is also a “Vernier”residue, which affects the structure of the CDR2 (41). Residues40G and 55G are both positions that are included in CDR1 and
The Journal of Immunology 323
by guest on July 11, 2018http://w
ww
.jimm
unol.org/D
ownloaded from
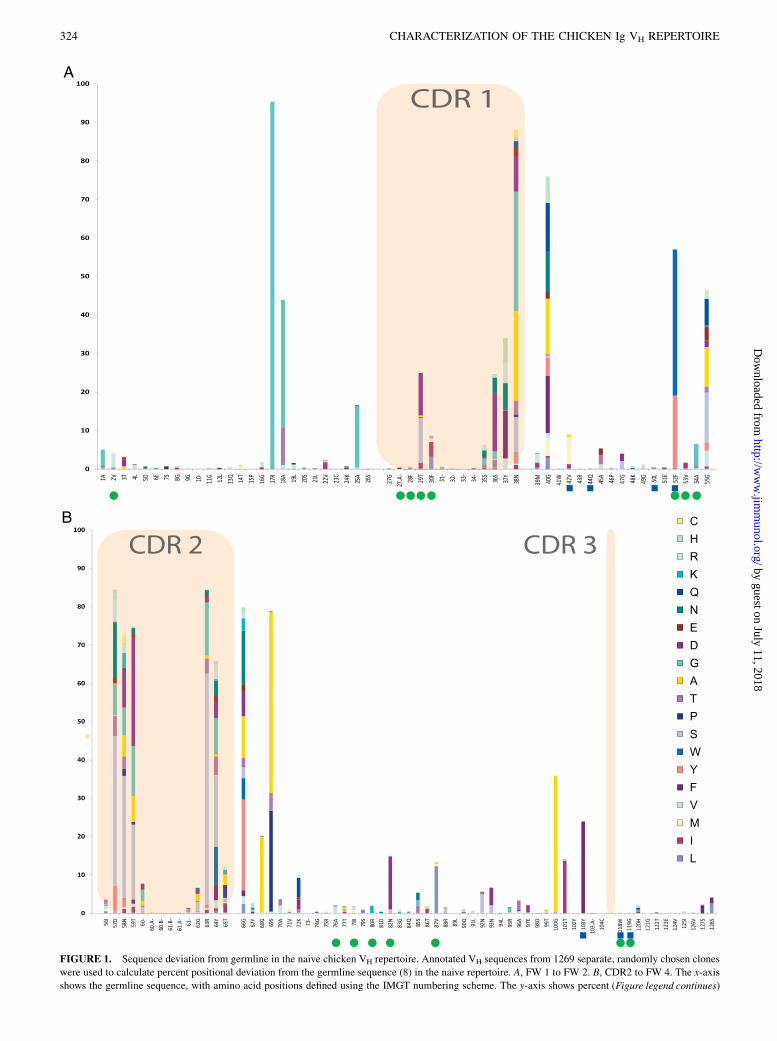
FIGURE 1. Sequence deviation from germline in the naive chicken VH repertoire. Annotated VH sequences from 1269 separate, randomly chosen clones
were used to calculate percent positional deviation from the germline sequence (8) in the naive repertoire. A, FW 1 to FW 2. B, CDR2 to FW 4. The x-axis
shows the germline sequence, with amino acid positions defined using the IMGT numbering scheme. The y-axis shows percent (Figure legend continues)
324 CHARACTERIZATION OF THE CHICKEN Ig VH REPERTOIRE
by guest on July 11, 2018http://w
ww
.jimm
unol.org/D
ownloaded from

CDR2 (respectively) under the Kabat numbering scheme (42) andwere both found to be substituted in the majority of clones (.75and .45% in the naive population, respectively). Although thesepositions are both designated as part of FW 2 under the IMGTnumbering scheme, their hypervariability is not surprising becausethey are known to be solvent exposed and frequently mediatecontacts with Ag in cocrystal structures for Abs from mammals(38, 39).FW 2 residue 42V is also associated with VH–VL interface
interactions and was mutated, but at a lower rate (7% M, 1% A,and 1% I). The other two FW 2 residues that are known to beimportant VH–VL interface positions (44Q and 50L) were highlyconserved. Vernier residues 53V and 54A are known to affectCDR2 conformation and were rarely substituted to 1% I and 6%G, respectively.In the FW 3 region, position 66G was found to be hypervariable.
This residue is designated as a constituent of CDR2 in the Kabatnumbering scheme (42) and is also known to be solvent exposedand to frequently mediate contact with Ag in mammals (39).Several other CDR2-proximal positions were also variable: 68G(20% A), 69S (47% A, 26% P, and 5% T), and 72K (5% Q, 3% D,and 1% T). CDR2 supporting Vernier residues 76A, 78I, and 80Rwere all highly conserved, whereas positions 82N and 87V weresubstituted more frequently to D (14%) and L (12%), respectively.In the case of 87V, homologous substitution to L may still be ofconsiderable significance because this residue has been found tomake CDR contacts and to be a critical position that had to bemaintained to retain target-binding affinity during the humaniza-tion of two previously described chicken Abs (3, 4). In addition,FW 3 VH–VL interface residue 103Y was frequently substitutedfor F (24%).The FW 4 region appeared to be the most highly conserved, with
the VH–VL interface residues 118W and 119G mutating veryrarely, if ever. Position 120H, which is variant in other species,was rarely mutated (,5%) in chickens. This generally low inci-dence of mutations in the FW 4 region may be attributed to thepresence of only a single functional J-region in the chicken VH
locus, which is not thought to be targeted during the gene con-version process (8).To allow comparison of the naive and immunized repertoires,
phage-display libraries of scFv were generated from chickensimmunized with a variety of Ags. After phage-display selectionsand screening on 12 separate protein targets, 318 total uniquesequences were successfully identified that exhibited specific bind-ing to their cognate Ags in direct ELISA screening of scFvs.On analysis of a plot of the positional diversity in these 318 VH
sequences, generated as above, it was found that the profileof amino acid substitutions across the V-gene sequence was highlysimilar to that of the naive repertoire (Supplemental Fig. 1).
Analysis of naive diversity in CDRs 1 and 2
In the CDR1 region, considerable diversity was observed at allpositions other than the highly conserved 27G and 28F (Fig. 1).CDR1 positions that were found to be highly mutated (29T-38N)were generally those known to be solvent exposed and whichcommonly interact with Ag (43). 37Y occupies a position that canbe buried (39) but in chickens was diversified ∼35% of the time,either to hydrophobic (V and F) or charged residues (R, H).
The CDR2was found to be a region of high diversity, particularlyin the solvent-exposed residues between positions 57 and 65 thatare known to frequently contact Ag (Fig. 1) (43). Position 56I washighly conserved, as expected, because it is an important, buried,structural residue that is highly conserved in multiple species (16).On the few occasions that 56I was mutated (,5%), it was sub-stituted for either homologous hydrophobic (V, L, and M) or morerarely, small residues (S and T).Interestingly, at a number of solvent-exposed positions in the
CDR1 (position 38), FW 2 (position 55) and the CDR2 (position58), noncanonical cysteine insertions were observed at low fre-quency (2.1, 0.8, and 2.4%, respectively). Importantly, only a singlenoncanonical cysteine was ever found at any of these three posi-tions in any individual sequence, and no clones were identified thatcontained a cysteine in both CDR1 and CDR2 simultaneously.Interestingly, in Ab structural models, each of these residues havecommonly been mapped as packing closely to the HCDR3, whereunpaired cysteines were also observed (see below) (16, 38).
Length distribution in the CDR3 repertoire
To compare the length distribution of the CDR3 regions, 11,423human, 5,054 mouse, and 1,269 naive chicken sequences wereexamined (Fig. 2A). As observed previously, the range of humanCDR3 lengths was approximately normally distributed and ex-tremely broad (5–37 aa; mean, 16.16 4.1) (14). The mouse CDR3repertoire was much shorter (5–26 aa; mean, 11.8 6 2.4). Thechicken naive repertoire was not normally distributed and favoredmuch longer sequences, despite a similar mean length to humans(8–32 aa; mean, 16.2 6 3.2). The majority (89%) of chickenCDR3 lengths were between 15 and 23 aa. Shorter CDR3 lengths(,15 aa, 8 aa being the shortest observed) were found to be rel-atively rare (each ,2% of the repertoire), with a major increase infrequency beginning at lengths 15 (6.5%) and 16 (16.7%), withlength 16 being the highest frequency length overall.In the selected chicken repertoire, the length distribution profile
closely mimicked that of the naive repertoire (Fig. 2B). One no-table difference was the identification of a sequence with a CDR3length of 6 aa, two residues shorter than the shortest sequence inthe naive repertoire. In addition, no binding sequences with CDR3longer than 29 aa were observed, whereas lengths up to 32 aacould be found (albeit at low frequency) in the naive repertoire.Abs with such exceptionally long CDR3s are rarely selected fromnaive human Ab libraries. To estimate whether clones containingsuch VH CDR3s could be functionally expressed in E. coli froma phagemid system, a chimeric chicken–human Fab construct wasgenerated containing the longest CDR3 found in the naive data-base. This 32-aa CDR3 contained two noncanonical cysteine resi-dues (“Type 1”; Fig. 3). Under standard expression conditions, thisFab was found, by Western blot analysis, to be expressed into theperiplasm (data not shown).
Amino acid usage in the CDR3 loop repertoire
Analyses of amino acid distribution and Shannon entropy (44)(amino acid variability in a given position in aligned sequences)were performed for sequences at each CDR length, includingresidues 105–117 of both the naive and selected repertoires (Fig.4A, 4B, Supplemental Fig. 2). Examination of the Shannon en-tropy plots indicated that positions 105–106 and 115–117 are
deviation. CDRs 1 and 2 are highlighted in shaded boxes. The CDR3 is excluded for clarity (and covered separately in Fig. 4). Amino acid substitutions at
each position are color coded. Also illustrated are the locations of Vernier positions (green circle) and known L chain interface residues (blue column) as
defined previously (40, 41).
The Journal of Immunology 325
by guest on July 11, 2018http://w
ww
.jimm
unol.org/D
ownloaded from

extremely conserved. Particularly highly conserved are positions106 (K/R) and 116 (D). The maintenance of these residues isstrongly indicative that the vast majority of chicken CDR3 struc-tures involve a stabilizing salt bridge at the bottom of the loop,theoretically creating the “Kinked Base” motif (45), although arecent study suggests that this structural motif is not guaranteed(46). The amino acid distribution plots in Fig. 4 also suggest thefrequent use of the motifs CAK and CAR at positions 104–106(CAK being predominant), which are common sequences alsofound at these positions in primates and rodents. The IDA motif atpositions 115–117 was found to be extremely highly conserved,which is due to these residues being donated by the J region.Interestingly, although there was a high level of entropy at allpositions between 107 and 114 in all lengths examined, position114 was repeatedly a position of high entropy values (∼4.0) andwas often preceded by reduced entropy (,3.0) at positions 112and 113 (Fig. 4A, 4B, Supplemental Fig. 2). When examined
across all CDR3 lengths, cysteine use was observed at all positionsbetween 107 and 114, with a general trend toward the highestfrequencies in the middle of the loops and progressively loweringtoward the base of the loop (Supplemental Fig. 2).To increase our understanding of CDR3 repertoire content,
analysis of the total amino acid usage in chicken, mouse, andhuman CDR3s was subsequently performed for all residues be-tween positions 107 and 114 (Fig. 5A). Positions 105–106 and115–117, which are also part of the CDR under the IMGT defi-nition, were excluded from the analysis to avoid germline FW andJ region-related biases, respectively. Several major differenceswere observed between the CDR3 aa content of chickens andhumans/mice. First, the prevalence of cysteine in the chickenrepertoire was much higher than for either mammal (chicken,9.4%; human, 1.6%; and mouse, 0.25%). That chickens should beincorporating ∼38 times more cysteine than mice and ∼6 timesas much as human is clearly reflective of the universal content of
FIGURE 2. CDR3 length distribution
in chickens. A, Comparison of CDR3
length distribution for the naive chicken
VH repertoire (red column) to that found
in public database sequences of mouse
(gray column) and human (black column).
B, Comparison of CDR3 length distribu-
tion in the naive (black column) and se-
lected (white column) chicken VH reper-
toires.
326 CHARACTERIZATION OF THE CHICKEN Ig VH REPERTOIRE
by guest on July 11, 2018http://w
ww
.jimm
unol.org/D
ownloaded from

that amino acid in the genomic D regions in reading frame 1 (21)and its regular use in D-like pseudogene sequences (8). Second, itwas observed that tyrosine content in the chicken CDR3 region
was markedly lower (9.2%) than that of humans (16.8%) or mice(26.4%).Further differences in repertoire content were observed when
examining specific groupings of amino acid chemistries. Smallamino acids such as G, S, A, and Twere all overrepresented in thechicken repertoire (54% total) relative to human (36% total) andmouse (35% total). For G, S, and A, this is not surprising given theirhigh representation in chicken D regions, but T is not found in anygenomic D region and may be introduced either through somaticmutation or gene conversion (21). Proline is underrepresented,however, which is surprising because the chicken repertoire hasa large percentage of long CDR3s, and this residue has been ob-served to increase in frequency as human CDRs get longer (14).Proline is also predicted to add structure to longer human CDR3loops (14) but may not be required for the structural stabilizationof chicken CDR3 loops, because of the high incidence of intra-and inter-CDR disulfide formation. Furthermore, the content ofpositively charged residues (R, K, and H) was distinctly lower inthe chicken repertoire (2.9% total) than in either mouse (8.7%total) or human (8.4% total). Negatively charged residue D wasequally represented in chicken as it is in mouse and human (6.8–7%), whereas E was lower, with N approximately equal. Hydro-phobic residues other than tryptophan (I, F, L, M, P, and V) werealso significantly lower in the chicken repertoire (8.5% total)versus mouse (13.1% total) and human (19.8% total). When com-pared with the naive repertoire, the target-specific binding se-quences again showed an extremely similar profile (Fig. 5B). Al-though some residues were found at moderately increased fre-quency (e.g., L, I, Y, T, and H), others were also slightly reducedin their representation (e.g., G, S, and C).
Cysteine usage in chicken VH CDRs
To further examine the influence of high cysteine content in thenaive and selected CDR repertoires of chickens, we calculatedthe average number of clones having noncanonical cysteines ineach VH CDR. In the CDR3 of the naive repertoire, it was foundthat 33.5% of all clones contained no cysteines, 53% containedtwo, and 0.7% contained four (Fig. 6A). That ∼87% of all clonesshould have an even number of cysteine residues would appearlogical, given the potential negative influence of unpaired thiolgroups on protein folding and secretion (47). Of greater interest,however, may be the observation that 11.4 and 1.4% (12.8% total)of all clones have one or three cysteines, respectively, in theCDR3. This odd number of thiol groups may be partially com-pensated, however, by the presence of extra noncanonical cysteinegroups in the CDR-H1 (2.2%) or CDR-H2 (3.4%), which may pairwith and stabilize the CDR3 via a disulfide bond. Importantly, the
FIGURE 3. Representative CDR sequences
for chicken VH structural types 1–6. Potential
disulfide linkages between cysteine residues are
denoted by black bars. In this figure, the CDR
definition according to Kabat (42) is used, solely
to ensure the inclusion of all noncanonical cys-
teines, including that frequently found at IMGT-
defined position 55. No noncanonical cysteines
were observed outside these regions.
FIGURE 4. Plots of Shannon entropy (upper panel) and amino acid
frequency (lower panel) for VH CDR3 positions 105–117, at the average
length of 16 aa. A, A total of 211 naive sequences. B, A total of 47 selected
target-binding sequences. Percent amino acid content at each position is
color-coded. Shannon entropy is plotted on the scale 0–4.
The Journal of Immunology 327
by guest on July 11, 2018http://w
ww
.jimm
unol.org/D
ownloaded from

cysteine residues observed in the CDRs H1 and H2 were found atpositions 38, 55, and 58, which are typically solvent exposed andare also structurally proximal to the CDR3, making them theo-retically available to form inter-CDR disulfide bonds (38, 39).When the average number of cysteines in the CDR3 was plottedfor each CDR3 length, a strong correlation (R2 = 0.97) is observedbetween increasing length and higher cysteine content (Fig. 6B).This indicates that disulfide linkage is strongly favored in longerclones, which may increase the stability of the CDR structure.When these analyses were extended to the selected repertoire,
some distinct differences were observed (Fig. 6C, 6D). The fre-quencies of cysteines in different CDRs changed: CDR3s withzero cysteines became dominant at 50.1%, with two cysteineslowered to 44.2%, one cysteine at 3.3%, three cysteines at 1.8%,and four cysteines at 0.7%. In CDRs 1 and 2, the frequency of cyste-ines dropped to 1.8 and 1.3%, respectively. Importantly, CDRswith unpaired cysteines (i.e., lacking a potential partner for intra-or inter-CDR disulfide bonding) were reduced in the selectedrepertoire when compared with the naive population. Thesechanges in cysteine pattern were underlined by a reduction in thecorrelation between CDR3 length and cysteine frequency (R2 =0.78), when compared with the naive repertoire.
Proposal of structural subtypes in chicken VH domains
For both camelid VHH (48) and shark IgNAR molecules (49),sequence attributes including the pattern of noncanonical cysteineusage in the V domain have allowed the description of structural“types,” which may be found in the repertoire. Our analyses of thechicken VH sequence database, therefore, led us to describe thefollowing types 1–6, based on the presence or absence of non-canonical cysteines and listed in approximate frequency by whichthey are found in the naive repertoire (Fig. 3): type 1, containingtwo cysteines in the CDR3 (55% of the naive repertoire); type 2,containing no noncanonical cysteines (33%); type 3, containingone cysteine in the CDR1 at position 38 plus one cysteine in theCDR3; type 4, containing one cysteine in the CDR2 at positions55 (type 4a) or 58 (type 4b) plus one cysteine in the CDR3; type 5,containing one cysteine in the CDR1 at position 38 plus threecysteines in the CDR3; and type 6, containing four cysteines in theCDR3. Each of types 3–6 represent ,5% of the total naive rep-ertoire.
Expression and analysis of an IgM-specific type 5 chicken Fab
Selection and screening of a scFv library generated from chickensimmunized with dogfish IgM identified a clone (H2), which is anexample of one of the atypical structural types (type 5; Fig. 3)outlined above. H2 exhibits strong, specific IgM binding, despitehaving four noncanonical disulfides in the VH (none in the VL).Intrigued by this structure, we analyzed it further.To examine the expression and affinity properties of H2, we first
converted the V genes from scFv format into a bicistronic, chimericFab expression construct (31). The E. coli expression, nickel pu-rification, and analytical size exclusion chromatography analysisof this construct proved that the Ab is produced as a stable, mo-nomeric Fab. No evidence was observed for either covalent ornoncovalent aggregate formation (Fig. 7A). Because the H2 Fabwas soluble and monomeric in solution, this facilitated the de-termination of its affinity for IgM. Purified IgM protein was co-valently coupled to a Biacore CM5 chip, and purified H2 Fab wasflowed in the mobile phase, generating binding curves that fit wellto a 1:1 Langmuir kinetics model, with a globally fit KD of 64 nM(Fig. 7B).
DiscussionAs in all higher vertebrates, primary diversity in chicken VH
regions is created by V-D-J recombination, combined with so-matic hypermutation (8, 20, 50). It has become clear, however,that there are other mechanisms of VH diversification in the chickenthat are distinctly different from those observed in mice andhumans (8, 51). The greatest influence on the diversification ofthe repertoire appears to be mediated by “gene conversion,”a process in which multiple upstream CVH pseudogenes mayundergo recombination into the VH gene sequence, after a func-tional V-D-J rearrangement event (8, 50). This process occursduring diversification of the H chain, with an analogous processoccurring in the L chain, leading to mutation of the CDRs and alsothe modulation of FW sequence. For this process to be efficient, itrelies on high DNA homology between the pseudogene and thegermline gene, which acts as the acceptor in the recombinationevent (6).The intrinsic need for high overall sequence homology between
germline and pseudogene may explain our observation that the FWregions of the chicken VH repertoire are highly conserved incomparison with germline. We observed that: hypervariability waspredominantly contained within the CDRs; the majority of di-versity in CDRs 1 and 2 was found in the solvent-exposed resi-dues, and this was coupled with strong maintenance of common
FIGURE 5. Comparison of total amino acid usage bias in VH CDR3
positions 107–114 of chickens, mice, and humans. A, The percentage
content of each of the natural amino acids was calculated for the CDR3s of
the chicken naive (white column), human (black column), and mouse (gray
column) repertoires. For ease of comparison, the amino acids are grouped
according to gross chemical character (e.g., hydrophobic [LIMV], aro-
matic [FYW], small [GSACTP] and charged [DEKRH]). B, The percent-
age content of amino acids found in the CDR3s of the naive (black
column) and selected (white column) chicken VH repertoires.
328 CHARACTERIZATION OF THE CHICKEN Ig VH REPERTOIRE
by guest on July 11, 2018http://w
ww
.jimm
unol.org/D
ownloaded from

structural residues that have also been observed in mice, humans,and other mammals (14, 38, 52). It is logical that the solvent-exposed residues in the CDRs should be the primary focus fordiversification, because they are most likely to make Ag contacts(38), but the observation that several important structural residuesare highly conserved might suggest limited conformational di-versity. Clear indications could be seen, however, that regions ofthe FW regions are modulated, which are of specific importance tothe generation of a functional repertoire. These regions includedresidues associated with affecting VH–VL interaction (40) andVernier regions that directly affect CDR structure (41). Variabilityin the Vernier regions was observed proximally to all three CDRs.The VH repertoire therefore remains essentially “single frame-work” but adds significant variability at appropriate positions toincrease the diversity of CDR presentation modes. It will be ofconsiderable interest to see, in future studies, whether these ob-servations also hold for other species that use a gene conversionmethod of v-gene diversification, such as rabbits.Although the FW modulations described above aid the gener-
ation of a complex repertoire, the majority of all diversity in the VH
was still observed to be in the CDR3. As the most hypervariableregion in any Ab, the VH CDR3 is also likely to be a center uponwhich novel complexity might be built. This is clearly of criticalimportance to a repertoire that is primarily reliant upon a sin-
gle germline gene. In chickens, V-D-J recombination leads to amuch more limited primary CDR3 diversity than in humans andmice, because chickens have only 15 functional D segments, all ofwhich are highly homologous. Indeed, some chicken D segments(e.g., D9/12/13 plus D4/8/11) are identical in amino acid sequence(21). In addition, the choice of D segment reading frame isstrongly enforced by molecular control mechanisms (53). Readingframe 1 predominates in chickens, because reading frame 2 cre-ates sequences containing stretches of hydrophobic residues suchas V and L. Because reading frame 2 lacks common structural andcontact residues such as glycine, tyrosine, and tryptophan, it is lesslikely to provide a functional Ab combining site than readingframe 1 (18). Reading frame 3 leads to the incorporation of stopcodons and cannot create a functional encoding sequence (21, 23).This general style of control mechanism appears to be universaland has been observed (albeit in different reading frames) for rab-bits, sharks, mice, primates, and humans (13, 18, 53).The chicken D segment repertoire is heavily biased toward the use
of reverse turn/hydrophilic residues such as G, S, and Y, just as it isin all other vertebrate species studied to date (14, 18). In contrastto humans and mice, however, chicken D segments also obligatelyuse C. This is partly because, in reading frame 1, the 15 genomicD segments lead to a consensus sequence 9–10 aa in length: G-S-(A/G)-Y-C-(G/C)-(S/W)-X-A-(Y/E) (X, nonconserved) (21). The
FIGURE 6. Comparative analysis of noncanonical cysteine usage patterns in chicken VH domains. A, Percentage of naive clones containing cysteines in
CDR1, 2, or 3. B, Cysteine usage in the naive CDR3 repertoire as loop length increases (R2 = 0.97). C, Percentage of selected clones containing cysteines in
CDR1, 2, or 3. In both plots, the observation of 0 (red column), 1 (black column), 2 (gray column), 3 (white column), or 4 (green column) cysteine residues
in individual sequences is represented. No sequences were observed that had cysteines in CDR1 and 2 simultaneously. In addition, the CDR3 was the only
region ever observed to contain more than one noncanonical cysteine residue at any one time. In the plots for CDR1 and 2, therefore, only values of “1” are
represented for the sake of clarity. D, Cysteine usage in the selected CDR3 repertoire (R2 = 0.78). Analyses in C and D performed for CDR3 lengths
representing .1% of the total population.
The Journal of Immunology 329
by guest on July 11, 2018http://w
ww
.jimm
unol.org/D
ownloaded from

final repertoire is more complex than this, however, because the Dsegment repertoire is subsequently further diversified not just bysomatic mutation but also by insertion of new sequences via geneconversion. These D-like sequences may replace the entire D seg-ment or only a small section (8, 21).Our analyses of CDR3 content show very clearly that the bias
toward small amino acids (G/S/A/C) suggested by the genomicD segments is indeed recapitulated in the adult repertoire. As aclear indication of a preference for small amino acids, T (not ob-served in D segments) is also overrepresented in comparison withmice and humans. Large aromatic and hydrophobic residues arestrongly disfavored in the chicken repertoire, including an un-usually low representation of Y, which is dominant in the rep-ertoires of mice and humans (14). This finding may have sig-nificant biological relevance as several synthetic Ab studies havesuggested that tyrosine is a critical amino acid for forming afunctional CDR3 repertoire (54–57). Indeed, structural analysisof both natural and synthetic Abs has suggested that tyrosine andtryptophan are capable of playing a dominant role in specificinteraction with Ag (58). In our analysis of the CDR3 repertoire,however, tryptophan was used at 3–4% by all three species (Fig.5). No increase in tryptophan use in the selected chicken reper-toire was observed that might compensate for the low tyrosinelevel. This observation would also suggest that the use of Dsegments that encode directly for tryptophan, such as D4, D7,D8, D11, and D14 (21), is either low in the chicken repertoire, orthe tryptophan residue is commonly removed via hypermutation.In addition, the chicken CDR3 repertoire has a reduced content ofpositively charged residues in comparison with human and mu-rine sequences. This may be of considerable practical importancefor the biotechnological use of chicken Abs, because excesspositive charge in the Ab combining site has been associated with
polyreactivity (59) and poor pharmacokinetic profile in in vivoanimal models (60).Taken together, these findings suggest that the molecular basis
for target interaction may differ between humans, mice, andchickens. Tyrosine, tryptophan, and arginine are all residues thathave been found to make critical contacts with Ag in mammalianAbs (58). The relatively low representation of Y/W/R and biastoward G/A/S in the chicken CDR3 repertoire may indicate thatAg specificity is mediated to a greater extent by hydrogen bondingvia the smaller residues than by the hydrophobic/charge inter-actions commonly observed for Y/W/R (61). Alternatively, it isalso possible that the higher incorporation of small residues in thechicken repertoire is a necessity, because these residues are re-quired to generate increased complexity in the secondary structureof CDR3 loops (58). This increased CDR3 complexity may helpto compensate for the limited FW diversity observed in thechicken repertoire. In future studies, the derivation of cocrystalstructures for chicken Abs and their cognate Ags may help tofurther dissect this issue.Our analyses of the CDR3 led to the important observation that
chickens use much more C in their repertoire than do mice orhumans and that C usage increases in direct correlation with CDRlength. The obligate use of C in the majority of unmutated chickenD segments and in D-like sequences (21) appears to lead to .50%of all B cell clones in the naive repertoire containing cysteine intheir VH CDR3. Both humans and rhesus have been shown to usethe gene segment D2, which can lead to the generation of func-tional CDR3 sequences containing a pair of cysteines, similar tothose found in our proposed chicken type 1 VH sequences (14, 18).Despite these observations, however, CDR3 sequences containingcysteine are observed at relatively low frequency in the matureB cells of humans and primates, and they are very rare in mice
FIGURE 7. Size exclusion chromatography and Bia-
core characterization of the IgM-binding clone H2. A,
Size-exclusion chromatograph of 100 mg/ml of the
clone H2 shown in red and molecular mass markers
shown in blue (kDa for each marker). Chromatograph
inset represents an Instant Blue-stained reducing SDS-
PAGE gel of 8 mg purified H2. Both analyses show
a highly purified protein of the expected size. B, 1:1
Langmuir model fitted sensorgrams of the binding ki-
netics analysis for clone H2.
330 CHARACTERIZATION OF THE CHICKEN Ig VH REPERTOIRE
by guest on July 11, 2018http://w
ww
.jimm
unol.org/D
ownloaded from

(14, 53). The high incorporation rate of cysteine in the chickenCDR3 may be balanced, however, by the frequent use of D-Djunctions (i.e., incorporation of two D segments in one CDR3)(21). Our analyses would suggest that this D-D junction forma-tion, along with high C content and the intrinsic flexibility ofsequences biased toward G and S, can create long CDR3s that cancontain intra or even inter-CDR disulfide bridges. These covalentbonds between CDRs are analogous to those observed at highfrequency in the Igs of other animals such as camels (48), sharkIgNARs (49, 62), cows (63–65), pigs (66), and even the duckbilledplatypus (67).Although VH CDR3s from chickens having two C residues and
probable intra-CDR disulfide bonding have been previously de-scribed (28, 31), it is fascinating to note the presence of clonesthat appear to contain disulfide linkage between the CDR3 andthe CDR1, the CDR2, or FW 2. Indeed, the positioning of non-canonical C residues observed in the CDRs or FW 2 of chickens,combined with an uneven number of C residues in the CDR3,imply structural convergence with similar CDR features describedfor both camels and llamas (48). The observed placement of thesenoncanonical C residues is maintained across multiple species,irrespective of whether the VH domain is expected to pair with anL chain, in the case of cow, pig, and platypus IgG (63, 66, 67), oris independent in the case of camelid VHH (48). A noticeableexception to this observation is the rabbit repertoire. While alsolimited predominantly to a single highly expressed VH germline(68), rabbits exhibit a VH CDR3 that has a shorter length–distri-bution profile (mean, 13.6 aa) than chickens and humans andtypically lacks cysteine incorporation (69). It is likely, however,that the rabbit has developed an alternative diversification mech-anism that focuses on the Vk, rather than the VH, because cysteineresidues are encoded within the CDR3 of some rabbit Vk germ-lines and cDNA sequences containing probable intraloop disulfidebonds have been described previously (70). Furthermore, func-tional target-specific Abs have been isolated from immune rabbitsthat exhibit considerably longer Vk CDR3 usage than typicallyfound in humans, as well as noncanonical cysteines in both theCDR1 and 2 of the VH (69, 70). Through mutagenesis studies, ithas been suggested that inter- or intra-CDR disulfides may be es-sential for both V-domain stability and to maintain binding func-tion in IgG and single-domain Abs (47, 71).In our analysis of CDR3 length distribution, it was found that the
vast majority of chicken B cells are producing loops $ 15 aa inlength. It is intriguing to note that a CDR of 15 aa is approxi-mately equivalent to a single V-D-J recombination event, if theconsensus D segment is combined with the heavily conservedpositions donated by FW 3 and the J region (105–106 and 115–117, respectively). Longer CDR3 lengths must therefore be cre-ated by a combination of n-nucleotide addition, D-D joiningevents (21), or the formation of “mosaic CDR3 sequences” byrecombination events with pseudogene-derived “D-like” sequen-ces that are introduced during gene conversion (8). When com-bined with the prevalence of multicysteine containing CDR3s,these observations are strongly suggestive that B cells containingeither V-D-D-J or (CVD)-D-J arrangements represent $60% ofthe total adult repertoire. This is surprising as V-D-D-J rear-rangements have been described as being frequent in the embry-onic bursa of fabricius but strongly selected against prior to theonset of gene conversion (6).The isolation of 318 unique chicken scFv Abs from our im-
munized phage-display libraries, recognizing 12 separate proteinAgs, allowed us to gain some clear insights into the Ag-specificrepertoire. The selected repertoire appears to closely mimic thatof the naive repertoire in CDR and FW mutation rates, CDR3
length distribution, and amino acid content. These are importantobservations in a practical sense, because they show that the se-lected repertoire may be a relatively accurate representation of theAb repertoire in the immunized animal. One deviation from thisoverall picture is the observation that the selected repertoire ex-hibits a lower correlation between CDR3 length and cysteinecontent than is found in the naive repertoire. It is possible, however,that this phenomenon is driven by the influence of the E. coli expres-sion machinery used in the phage–display process, which may se-lect against clones with complex disulfide bonding profiles or free,unpaired cysteine residues.The observations outlined above have allowed us the opportu-
nity to define six specific structural types of chicken VH domains,based on their noncanonical cysteine content. These type defi-nitions may aid further characterization of the chicken reper-toire in future studies. Although types 1 and 2 dominate the reper-toire in both the naive and selected sequence populations, theless prevalent members may still be of considerable importance.To illustrate that the less common types 3–6 may be a fullyfunctional part of the Ag-specific response, we examined our se-lected repertoire for functional target-binding clones with raremotifs. Fab H2 was generated, which exhibited efficient, mono-meric expression in E. coli and 64 nM affinity for IgM, despitecontaining four noncanonical cysteine residues in the CDRs. Be-cause the H2 VH domain is a member of the rare type 5 family(,2% of the total naive repertoire), these data clearly illustratedthat such sequences are produced during the response to foreignAgs and are not merely nonfunctional sequence artifacts. In addi-tion, the successful expression of a chimeric Fab containing a CDR3of 32 aa in E. coli suggests that the full extent of the chickenrepertoire may be sampled using phage display.All current evidence suggests that chickens use different amino
acid architecture than humans and mice to generate novel Abs intheir VH CDRs, commonly using long CDR3 loops that are sta-bilized by disulfide bonding. We propose that this process createsCDR3s of great stability and structural complexity and isa mechanism for creating maximal diversity in a single-FW rep-ertoire. The observed predictability of VH subtypes that can bederived from immune chickens, coupled with clearly limitedVernier residue and VL interface residue complexity, will aid therapid and reproducible humanization of chicken Abs for thera-peutic indications.
AcknowledgmentsWe thank Caroline Barrelle and John Steven for the provision of purified
dogfish IgM protein, Thad Welch for assistance with data analyses, and
members of Global Biotherapeutics Technologies, past and present, for
the provision of target-specific chicken Ab sequences.
DisclosuresThe authors have no financial conflicts of interest.
References1. Fuh, G. 2007. Synthetic antibodies as therapeutics. Expert Opin. Biol. Ther. 7:
73–87.2. Hoogenboom, H. R. 2005. Selecting and screening recombinant antibody li-
braries. Nat. Biotechnol. 23: 1105–1116.3. Nishibori, N., H. Horiuchi, S. Furusawa, and H. Matsuda. 2006. Humanization of
chicken monoclonal antibody using phage-display system. Mol. Immunol. 43:
634–642.4. Tsurushita, N., M. Park, K. Pakabunto, K. Ong, A. Avdalovic, H. Fu, A. Jia,
M. Vasquez, and S. Kumar. 2004. Humanization of a chicken anti–IL-12 mono-
clonal antibody. J. Immunol. Methods 295: 9–19.5. Akamatsu, Y., K. Pakabunto, Z. Xu, Y. Zhang, and N. Tsurushita. 2007. Whole
IgG surface display on mammalian cells: application to isolation of neutralizing
chicken monoclonal anti–IL-12 antibodies. J. Immunol. Methods 327: 40–52.
The Journal of Immunology 331
by guest on July 11, 2018http://w
ww
.jimm
unol.org/D
ownloaded from

6. Ratcliffe, M. J. 2006. Antibodies, immunoglobulin genes and the bursa ofFabricius in chicken B cell development. Dev. Comp. Immunol. 30: 101–118.
7. Reynaud, C. A., V. Anquez, A. Dahan, and J. C. Weill. 1985. A single rear-rangement event generates most of the chicken immunoglobulin light chaindiversity. Cell 40: 283–291.
8. Reynaud, C. A., A. Dahan, V. Anquez, and J. C. Weill. 1989. Somatic hyper-conversion diversifies the single Vh gene of the chicken with a high incidence inthe D region. Cell 59: 171–183.
9. Ivanov, I. I., R. L. Schelonka, Y. Zhuang, G. L. Gartland, M. Zemlin, andH. W. Schroeder, Jr. 2005. Development of the expressed Ig CDR-H3 repertoireis marked by focusing of constraints in length, amino acid use, and charge thatare first established in early B cell progenitors. J. Immunol. 174: 7773–7780.
10. Link, J. M., M. A. Hellinger, and H. W. Schroeder, Jr. 2002. The Rhesus monkeyimmunoglobulin IGHD and IGHJ germline repertoire. Immunogenetics 54: 240–250.
11. Link, J. M., J. E. Larson, and H. W. Schroeder. 2005. Despite extensive similarityin germline DH and JH sequence, the adult Rhesus macaque CDR-H3 repertoirediffers from human. Mol. Immunol. 42: 943–955.
12. Schroeder, H. W., Jr., J. L. Hillson, and R. M. Perlmutter. 1987. Early restrictionof the human antibody repertoire. Science 238: 791–793.
13. Schroeder, H. W., Jr., G. C. Ippolito, and S. Shiokawa. 1998. Regulation of theantibody repertoire through control of HCDR3 diversity. Vaccine 16: 1383–1390.
14. Zemlin, M., M. Klinger, J. Link, C. Zemlin, K. Bauer, J. A. Engler, H. W. Schroeder,Jr., and P. M. Kirkham. 2003. Expressed murine and human CDR-H3 intervals ofequal length exhibit distinct repertoires that differ in their amino acid compositionand predicted range of structures. J. Mol. Biol. 334: 733–749.
15. Sidhu, S. S., and F. A. Fellouse. 2006. Synthetic therapeutic antibodies. Nat.Chem. Biol. 2: 682–688.
16. Sidhu, S. S., B. Li, Y. Chen, F. A. Fellouse, C. Eigenbrot, and G. Fuh. 2004.Phage-displayed antibody libraries of synthetic heavy chain complementaritydetermining regions. J. Mol. Biol. 338: 299–310.
17. Parvari, R., A. Avivi, F. Lentner, E. Ziv, S. Tel-Or, Y. Burstein, and I. Schechter.1988. Chicken immunoglobulin gamma-heavy chains: limited VH gene reper-toire, combinatorial diversification by D gene segments and evolution of theheavy chain locus. EMBO J. 7: 739–744.
18. Schroeder, H. W., Jr. 2006. Similarity and divergence in the development andexpression of the mouse and human antibody repertoires. Dev. Comp. Immunol.30: 119–135.
19. Schroeder, H. W., Jr., J. L. Hillson, and R. M. Perlmutter. 1990. Structure andevolution of mammalian VH families. Int. Immunol. 2: 41–50.
20. Parvari, R., E. Ziv, F. Lantner, D. Heller, and I. Schechter. 1990. Somatic di-versification of chicken immunoglobulin light chains by point mutations. Proc.Natl. Acad. Sci. USA 87: 3072–3076.
21. Reynaud, C. A., V. Anquez, and J. C. Weill. 1991. The chicken D locus and itscontribution to the immunoglobulin heavy chain repertoire. Eur. J. Immunol. 21:2661–2670.
22. Reynaud, C. A., V. Anquez, H. Grimal, and J. C. Weill. 1987. A hyperconversionmechanism generates the chicken light chain preimmune repertoire. Cell 48:379–388.
23. Weill, J. C., and C. A. Reynaud. 1992. Early B-cell development in chickens,sheep and rabbits. Curr. Opin. Immunol. 4: 177–180.
24. Parvari, R., E. Ziv, F. Lantner, S. Tel-Or, Y. Burstein, and I. Schechter. 1987. Afew germline genes encode the variable regions of chicken immunoglobulin lightand g-heavy chains. Prog. Clin. Biol. Res. 238: 15–26.
25. Parvari, R., E. Ziv, F. Lentner, S. Tel-Or, Y. Burstein, and I. Schechter. 1987.Analyses of chicken immunoglobulin light chain cDNA clones indicate a fewgermline V lambda genes and allotypes of the C lambda locus. EMBO J. 6: 97–102.
26. Reynaud, C. A., A. Dahan, and J. C. Weill. 1983. Complete sequence ofa chicken lambda light chain immunoglobulin derived from the nucleotide se-quence of its mRNA. Proc. Natl. Acad. Sci. USA 80: 4099–4103.
27. Finlay, W. J., N. C. deVore, E. N. Dobrovolskaia, A. Gam, C. S. Goodyear, andJ. E. Slater. 2005. Exploiting the avian immunoglobulin system to simplify thegeneration of recombinant antibodies to allergenic proteins. Clin. Exp. Allergy35: 1040–1048.
28. Finlay, W. J., I. Shaw, J. P. Reilly, and M. Kane. 2006. Generation of high-affinitychicken single-chain Fv antibody fragments for measurement of the Pseudo-nitzschia pungens toxin domoic acid. Appl. Environ. Microbiol. 72: 3343–3349.
29. Yamanaka, H. I., T. Inoue, and O. Ikeda-Tanaka. 1996. Chicken monoclonalantibody isolated by a phage display system. J. Immunol. 157: 1156–1162.
30. Finlay, W. J., L. Bloom, and O. Cunningham. 2011. Phage display: a powerfultechnology for the generation of high specificity affinity reagents from alterna-tive immune sources. Methods Mol. Biol. 681: 87–101.
31. Andris-Widhopf, J., C. Rader, P. Steinberger, R. Fuller, and C. F. Barbas, III.2000. Methods for the generation of chicken monoclonal antibody fragments byphage display. J. Immunol. Methods 242: 159–181.
32. Finlay, W. J., O. Cunningham, M. A. Lambert, A. Darmanin-Sheehan, X. Liu,B. J. Fennell, C. M. Mahon, E. Cummins, J. M. Wade, C. M. O’Sullivan, et al.2009. Affinity maturation of a humanized rat antibody for anti-RAGE therapy:comprehensive mutagenesis reveals a high level of mutational plasticity bothinside and outside the complementarity-determining regions. J. Mol. Biol. 388:541–558.
33. Finlay, W. J., L. Bloom, and O. Cunningham. 2011. Optimized generation ofhigh-affinity, high-specificity single-chain Fv antibodies from multiantigen im-munized chickens. Methods Mol. Biol. 681: 383–401.
34. Cummins, E., D. P. Luxenberg, F. McAleese, A. Widom, B. J. Fennell,A. Darmanin-Sheehan, M. J. Whitters, L. Bloom, D. Gill, and O. Cunningham.
2008. A simple high-throughput purification method for hit identification inprotein screening. J. Immunol. Methods 339: 38–46.
35. Kirsch, M., M. Zaman, D. Meier, S. Dubel, and M. Hust. 2005. Parameters af-fecting the display of antibodies on phage. J. Immunol. Methods 301: 173–185.
36. Rothlisberger, D., A. Honegger, and A. Pluckthun. 2005. Domain interactions inthe Fab fragment: a comparative evaluation of the single-chain Fv and Fabformat engineered with variable domains of different stability. J. Mol. Biol. 347:773–789.
37. Pommie, C., S. Levadoux, R. Sabatier, G. Lefranc, and M. P. Lefranc. 2004.IMGT standardized criteria for statistical analysis of immunoglobulin V-REGIONamino acid properties. J. Mol. Recognit. 17: 17–32.
38. Lee, C. V., W. C. Liang, M. S. Dennis, C. Eigenbrot, S. S. Sidhu, and G. Fuh.2004. High-affinity human antibodies from phage-displayed synthetic Fab li-braries with a single framework scaffold. J. Mol. Biol. 340: 1073–1093.
39. Hoet, R. M., E. H. Cohen, R. B. Kent, K. Rookey, S. Schoonbroodt, S. Hogan,L. Rem, N. Frans, M. Daukandt, H. Pieters, et al. 2005. Generation of high-affinity human antibodies by combining donor-derived and synthetic comple-mentarity-determining-region diversity. Nat. Biotechnol. 23: 344–348.
40. Padlan, E. A. 1994. Anatomy of the antibody molecule. Mol. Immunol. 31: 169–217.
41. Foote, J., and G. Winter. 1992. Antibody framework residues affecting theconformation of the hypervariable loops. J. Mol. Biol. 224: 487–499.
42. Kabat, E. A., T. Wu, H. M. Perry, K. S. Gottesman, and C. Foeller. 1991.Sequences of Proteins of Immunological Interest, 5th Ed. National Institutes ofHealth, Bethesda, MD.
43. MacCallum, R. M., A. C. Martin, and J. M. Thornton. 1996. Antibody-antigeninteractions: contact analysis and binding site topography. J. Mol. Biol. 262:732–745.
44. Shannon, C. E. 1948. A mathmatical theory of communication. Bell Syst. Tech. J.27: 623–656.
45. Shirai, H., A. Kidera, and H. Nakamura. 1996. Structural classification of CDR-H3 in antibodies. FEBS Lett. 399: 1–8.
46. North, B., A. Lehmann, and R. L. Dunbrack, Jr. 2011. A new clustering of an-tibody CDR loop conformations. J. Mol. Biol. 406: 228–256.
47. Fennell, B. J., A. Darmanin-Sheehan, S. E. Hufton, V. Calabro, L. Wu,M. R. Muller, W. Cao, D. Gill, O. Cunningham, and W. J. Finlay. 2010. Dis-section of the IgNAR V domain: molecular scanning and orthologue databasemining define novel IgNAR hallmarks and affinity maturation mechanisms. J.Mol. Biol. 400: 155–170.
48. Harmsen, M. M., R. C. Ruuls, I. J. Nijman, T. A. Niewold, L. G. Frenken, andB. de Geus. 2000. Llama heavy-chain V regions consist of at least four distinctsubfamilies revealing novel sequence features. Mol. Immunol. 37: 579–590.
49. Stanfield, R. L., H. Dooley, M. F. Flajnik, and I. A. Wilson. 2004. Crystalstructure of a shark single-domain antibody V region in complex with lysozyme.Science 305: 1770–1773.
50. Reynaud, C. A., B. Bertocci, A. Dahan, and J. C. Weill. 1994. Formation of thechicken B-cell repertoire: ontogenesis, regulation of Ig gene rearrangement, anddiversification by gene conversion. Adv. Immunol. 57: 353–378.
51. McCormack, W. T., L. W. Tjoelker, and C. B. Thompson. 1991. Avian B-celldevelopment: generation of an immunoglobulin repertoire by gene conversion.Annu. Rev. Immunol. 9: 219–241.
52. Rader, C., G. Ritter, S. Nathan, M. Elia, I. Gout, A. A. Jungbluth, L. S. Cohen,S. Welt, L. J. Old, and C. F. Barbas, III. 2000. The rabbit antibody repertoire asa novel source for the generation of therapeutic human antibodies. J. Biol. Chem.275: 13668–13676.
53. Raaphorst, F. M., C. S. Raman, B. T. Nall, and J. M. Teale. 1997. Molecularmechanisms governing reading frame choice of immunoglobulin diversity genes.Immunol. Today 18: 37–43.
54. Fellouse, F. A., P. A. Barthelemy, R. F. Kelley, and S. S. Sidhu. 2006. Tyrosineplays a dominant functional role in the paratope of a synthetic antibody derivedfrom a four amino acid code. J. Mol. Biol. 357: 100–114.
55. Fellouse, F. A., K. Esaki, S. Birtalan, D. Raptis, V. J. Cancasci, A. Koide,P. Jhurani, M. Vasser, C. Wiesmann, A. A. Kossiakoff, et al. 2007. High-throughput generation of synthetic antibodies from highly functional minimal-ist phage-displayed libraries. J. Mol. Biol. 373: 924–940.
56. Fellouse, F. A., B. Li, D. M. Compaan, A. A. Peden, S. G. Hymowitz, andS. S. Sidhu. 2005. Molecular recognition by a binary code. J. Mol. Biol. 348:1153–1162.
57. Fellouse, F. A., C. Wiesmann, and S. S. Sidhu. 2004. Synthetic antibodies froma four-amino-acid code: a dominant role for tyrosine in antigen recognition.Proc. Natl. Acad. Sci. USA 101: 12467–12472.
58. Birtalan, S., R. D. Fisher, and S. S. Sidhu. 2010. The functional capacity of thenatural amino acids for molecular recognition. Mol. Biosyst. 6: 1186–1194.
59. Li, H., Y. Jiang, E. L. Prak, M. Radic, and M. Weigert. 2001. Editors and editingof anti-DNA receptors. Immunity 15: 947–957.
60. Boswell, C. A., D. B. Tesar, K. Mukhyala, F. P. Theil, P. J. Fielder, andL. A. Khawli. 2010. Effects of charge on antibody tissue distribution and phar-macokinetics. Bioconjug. Chem. 21: 2153–2163.
61. Birtalan, S., Y. Zhang, F. A. Fellouse, L. Shao, G. Schaefer, and S. S. Sidhu.2008. The intrinsic contributions of tyrosine, serine, glycine and arginine to theaffinity and specificity of antibodies. J. Mol. Biol. 377: 1518–1528.
62. Dooley, H., M. F. Flajnik, and A. J. Porter. 2003. Selection and characterizationof naturally occurring single-domain (IgNAR) antibody fragments from immu-nized sharks by phage display. Mol. Immunol. 40: 25–33.
63. Aitken, R., J. Gilchrist, and M. C. Sinclair. 1997. A single diversified VH genefamily dominates the bovine immunoglobulin repertoire. Biochem. Soc. Trans.25: 326S.
332 CHARACTERIZATION OF THE CHICKEN Ig VH REPERTOIRE
by guest on July 11, 2018http://w
ww
.jimm
unol.org/D
ownloaded from

64. O’Brien, P. M., R. Aitken, B. W. O’Neil, and M. S. Campo. 1999. Generation ofnative bovine mAbs by phage display. Proc. Natl. Acad. Sci. USA 96: 640–645.
65. Sinclair, M. C., J. Gilchrist, and R. Aitken. 1997. Bovine IgG repertoire isdominated by a single diversified VH gene family. J. Immunol. 159: 3883–3889.
66. Li, F., and R. Aitken. 2004. Cloning of porcine scFv antibodies by phage displayand expression in Escherichia coli. Vet. Immunol. Immunopathol. 97: 39–51.
67. Johansson, J., M. Aveskogh, B. Munday, and L. Hellman. 2002. Heavy chain Vregion diversity in the duck-billed platypus (Ornithorhynchus anatinus): longand highly variable complementarity-determining region 3 compensates forlimited germline diversity. J. Immunol. 168: 5155–5162.
68. Friedman, M. L., C. Tunyaplin, S. K. Zhai, and K. L. Knight. 1994. NeonatalVH, D, and JH gene usage in rabbit B lineage cells. J. Immunol. 152: 632–641.
69. Popkov, M., R. G. Mage, C. B. Alexander, S. Thundivalappil, C. F. Barbas, III, andC. Rader. 2003. Rabbit immune repertoires as sources for therapeutic monoclonalantibodies: the impact of k allotype-correlated variation in cysteine content onantibody libraries selected by phage display. J. Mol. Biol. 325: 325–335.
70. Sehgal, D., G. Johnson, T. T. Wu, and R. G. Mage. 1999. Generation of theprimary antibody repertoire in rabbits: expression of a diverse set of Igk-V genesmay compensate for limited combinatorial diversity at the heavy chain locus.Immunogenetics 50: 31–42.
71. Lee, C. V., S. G. Hymowitz, H. J. Wallweber, N. C. Gordon, K. L. Billeci,S. P. Tsai, D. M. Compaan, J. Yin, Q. Gong, R. F. Kelley, et al. 2006. Syntheticanti-BR3 antibodies that mimic BAFF binding and target both human and mu-rine B cells. Blood 108: 3103–3111.
The Journal of Immunology 333
by guest on July 11, 2018http://w
ww
.jimm
unol.org/D
ownloaded from