genetic polymorphism of the ipah multicopy antigen gene in
TRANSCRIPT

University of Nebraska - Lincoln University of Nebraska - Lincoln
DigitalCommons@University of Nebraska - Lincoln DigitalCommons@University of Nebraska - Lincoln
Public Health Resources Public Health Resources
1995
Genetic polymorphism of the Genetic polymorphism of the ipaHipaH multicopy antigen gene in multicopy antigen gene in
ShigellaShigella spps. and enteroinvasive spps. and enteroinvasive Escherichia coli Escherichia coli
Jerry M. Buysse Walter Reed Army Institute of Research
Antoinette B. Hartman Walter Reed Army Institute of Research
Nancy Strockbine Center for Disease Control
Malabi Venkatesan Walter Reed Army Institute of Research
Follow this and additional works at: https://digitalcommons.unl.edu/publichealthresources
Part of the Public Health Commons
Buysse, Jerry M.; Hartman, Antoinette B.; Strockbine, Nancy; and Venkatesan, Malabi, "Genetic polymorphism of the ipaH multicopy antigen gene in Shigella spps. and enteroinvasive Escherichia coli" (1995). Public Health Resources. 232. https://digitalcommons.unl.edu/publichealthresources/232
This Article is brought to you for free and open access by the Public Health Resources at DigitalCommons@University of Nebraska - Lincoln. It has been accepted for inclusion in Public Health Resources by an authorized administrator of DigitalCommons@University of Nebraska - Lincoln.

Microbial Pathogenesis 1995; 19:335-349
Genetic polymorphism of the ipaH multicopy antigen gene in Shigella spps. and enteroinvasive Escherichia coil
Jerry M. Buysse, TM Antoinette B. Hartman, z Nancy Strockbine 3 and Malabi Venkatesan 1'2
1 Department of Bacterial Immunology and 2 Department of Enteric Infections, Division of Communicable Diseases and Immunology, Walter Reed Army Institute of Research, Walter Reed Army Medical Center, Washington, D.C., 20307-5100 and z Enteric Diseases Laboratory Section, Center for Disease Control, Atlanta, GA 30333, U.S.A.
(Received June 27, 1995; accepted in revised form September 1, 1995)
Buysse, J. H. (Department of Bacterial Immunology, Division of Communicable Diseases and Immunology, Walter Reed Army Institute of Research, Walter Reed Army Medical Center, Washington D.C. 20307-5100, U.S.A.), A. B. Hartman, N. Strockbine and M. Vankatesan. Genetic polymorphism of the ipaHmulticopy antigen gene in Shigella spps. and enteroinvasive Escher- ichia coil Microbial Pathogenesis 1995; 19: 335-349.
The ipaH loci comprise a multicopy antigen gene family unique to Shigella species and enteroinvasive Escherichia coli (EIEC). DNA probes derived from the Shigella flexneriserotype 5 ipaH~.8 gene were used to compare the molecular arrangement of ipaH alleles in a variety of Shigella and EIEC strains. Multiple copies of ipaH-homologous sequences were detected in all invasion plasmids examined. Oligonucleotide probes covering discrete 24 bp segments of the ipaH~.8 gene and sequences flanking the ipaH,.s (probe H25) and ipaH2.5 (probe H24) loci were used to define the extent of homology among invasion plasmid copies of ipaH in S. flexneri serotypes 1, 2 and 5 and in S. sonnei. IpaHalleles carried by these invasion plasmids were not structurally equivalent and showed sequence divergence at their amino- and carboxy-terminal ends. The H25 probe was shown to correspond to an IS629 sequence genetically linked to the ipaH alleles, while the H24 probe defined a DNA sequence found only in Shigella invasion plasmids. Chromosomal DNA from invasion plasmid-cured S. flexneri and S. sonnei strains hybridized a core ipaH?.8 gene segment, indicating that portions of the ipaH?.8 structural gene were reiterated and contained within the shigellae chromosomes. Based on the specificity of the ipaH~.8 core probe and the detection of ipaH sequences on the invasion plasmids and chromosomes of Shigella strains, three polymorphic groups within a collection of forty S. dysenteriae 1 isolates received by the United States Centers for Disease Control in 1988 were identified using this probe. These results suggest that ipaH restriction fragment length polymorphisms may be useful in genetic lineage and epidemiologic studies of virulent shigellae. © 1995 Academic Press Limited
Key words: Shigella; enteroinvasive Escherichia co~i; ipaH; invasion plasmid.
I n t r o d u c t i o n
Infection wi th Shigella spps. or entero invas ive Escherichia coil (EIEC), the et io logic agents of baci l lary dysentery, results in the product ion of immune serum conta in ing
* Current address and correspondence to: Jerry M. Buysse, Molecular Biology Research, Upjohn Lab- oratories, 301 Henrietta Street, Kalamazoo, MI 49007, U.S.A.
0882-4010/95/110335+15 $12.00/0 © 1995 Academic Press Limited

336 J.M. Buysse et aL
antibodies that recognize surface-expressed and secreted proteins of the bacteria. TM
Several immunogenic proteins are encoded by genes on the 220 kb invasion plasmid found in all virulent strains of these organisms. 5"6 Within this group of proteins, invasion plasmid antigens (Ipas) have been identified that are critical determinants of the invasive phenotype (IpaB, IpaC, and IpaD), the lysis of phagocytic vacuoles (IpaB), the induction of apoptosis in macrophages (IpaB) and the inter- and intra- cellular spreading phenotype (IcsA or VirG). 7-1s In addition, immunogens whose function in the pathogenesis of the disease is unknown have been identified, includ- ing IpaA and products of the multicopy ipaH gene family. 1'~s-~8
The ipaH gene was originally isolated from a ;~gt11 expression library of the S. flexneri serotype 5 (strain M90T) invasion plasmid, pWR100. 8 Hybridization studies of pWR100 using ,~gtllipaH gene probes revealed multiple copies of ipaH on this invasion plasmid, in contrast to the unit copy representation of ipaBCDA and icsA antigen genes. DNA sequence and Southern blot analysis of a 2.9 EcoRI insert (~gtllSfl-S39) encoding the 60 kDa IpaH antigen showed that the fragment over- lapped two Hindlll segments (7.8 and 4.5 kb in size), each of which carried a copy of the ipaH gene. ~7 The five copies of ipaH subsequently detected on pWR100 were designated ipaHg.8, ipaHT.8, ipaH4.s, ipaH2.5, and ipaHi.4, corresponding to the size of the Hindlll fragment harboring each allele. TM
The molecular genetic structure of these genes in S. flexneri serotype 5 has been examined in detail. TM Oligonucleotide hybridization data and DNA sequence analysis demonstrated that a conserved region of ipaH~.8 (bp 720-1557) was carried by each of the five pWR100 ipaH copies, however the DNA sequence for each gene diverged outside of this common core region. Comparison of the ipaHT.8 and ipaH4.s open reading frames (ORFs) revealed that amino acid residues in the amino-end of the proteins conformed to a consensus leucine-rich repeat motif found in a number of polypeptides, most notably the YopM polypeptide of Yersinia pestis and the leucine- rich glycoproteins GPIBc~, GPIb/~, and GPIX of human platelets. ~9 Interestingly, the antigenic domain of the IpaH proteins encoded by ipaHT.8 and ipaH4.s was found to reside in the conserved core region of the polypeptides and not in the sequence variable amino-end of the molecules. Although the five pWR100 ipaH alleles have not been completely mapped, the location for two alleles has been established; ipaH~.8 and ipaH4.s are contiguous and arranged in a head-to-tail fashion roughly 10 kb from the 3' end of the virB gene (Fig. 1). It should be noted that at this point no virulence function has been assigned to the ipaH alleles.
Nucleic acid sequences derived from the S. flexneri ipaHT.8 and ipaH4.s alleles have been shown to be effective probes for the specific detection of virulent and avirulent shigellae in clinical and environmental samples. 2°-24 Consistent ipaH colony hybrid- ization of avirulent shigellae isolates, cured of the invasion plasmid or containing substantial deletions of invasion plasmid DNA, suggests that multiple copies of the ipaHgene are present in the chromosome and invasion plasmids of various Shigella and EIEC strains. Preliminary analysis of S. flexneri serotype 5 strain M90T demon- strated copies of the ipaH gene on the invasion plasmid (five copies) and the chro- mosome (six copies), supporting a model of ipaH dispersion to both replicons. ~8
The aim of this study was to extend our molecular analysis of ipaHgenetic structure in a variety of Shigella and EIEC strains, in order to have a better understanding of ipaH heterogeneity in these organisms. These studies provide a starting point for efforts to specifically inactivate ipaH alleles within a given Shigella species. Addition- ally, results from this analysis suggest that ipaH polymorphism, defined with an ipaHT.8 core sequence probe, may provide a useful tool in epidemiological inves- tigations of shigellosis outbreaks.

Genetic polymorphism of ipaH 337
E S E I I I
icsA
I I 1 0 k b
H SS S S
II JL' T S S B E H E B B I EE H H E E H E I I I I I I I I I I I I I t t , ~ i = , ,
I I m l m
IpaH4.5 I vireipaADCB mxlHIJMEDA spa47.spa40
I ~H/Sp "~ P . . S " I [ P v l A s A E . P I I I I III I I I I l l I I
. . . . . . . . . . . . S39 probe m a
< - ' - ~ " ' - l k b
1599 bp Sp 0 bD
H4 H10 H15 H8 H7 H2 = / 1 / 1 1 1 1 l l ,
ipaH core probe
Fig. 1. Genetic and restriction map of the S. flexneri pWR100 invasion plasmid. The top line depicts a partial restriction map of a 70 kb section of the plasmid, encoding genes essential for invasion (virB, ipaBCDA, rnxiHIJMEDA, spa47-spa40) and intercellular spreading (icsA), in relation to two copies of ipaH designated ipaH4.s and ipaHT.8. The 4.5 kb BamHI/EcoRI fragment encoding these alleles is presented in an expanded view in the second line, with the location of the head-to-tail ipaHgenes indicated below the line along with the extent of the $39 probe; the arrows under the ipaH genes indicate the direction of tran- scription. The third line encompasses an expanded view of the molecular structure of the ipaH~.8 and ipaH4.5 genes; a Sail restriction site marks the separation of the genes' LPX-repeat regions from the conserved sections of these related loci. The location of the H25 oligonucleotide probe is indicated with a triangle below the line and 3' to the end of the ipaH4.s gene. The bottom line depicts a restriction map of the ipaHT.8 gene and the locations of the H2, H4, H7, H8, H10 and H15 oligonucleotide probes along the DNA sequence of ipaH~.a; also shown is the extent of the ipaH core sequence probe derived from ipaH~.e. Abbreviations for restriction enzyme sites are: Aval (A), BamHI (B), EcoRI (E), Hindlll (/-/), Psi1 (P), Pvull (Pv), Sail (S), Sphl (Sp).
R e s u l t s
IpaH distribution in shigellae invasion plasmid and chromosomal DIVAs Invasion plasmid DNAs from several Shigella and EIEC strains were digested with Hindlll and screened by Southern blot analysis, using the ipaH core sequence probe (Fig. 1); plasmids tested were obtained from the WRAIR strain collection and from a variety of other sources worldwide (Table 1). The ipaH core probe encompassed a 627 bp region of the ipaHT.8 gene that is conserved in all of the ipaHalleles maintained by S. flexneri serotype 5, strain M90T. This experiment revealed that multiple copies of ipaH were present in all of the plasmids tested (Fig. 2). Since the ipaH core sequence did not contain a Hindlll restriction site, the number of fragments hybridized
338
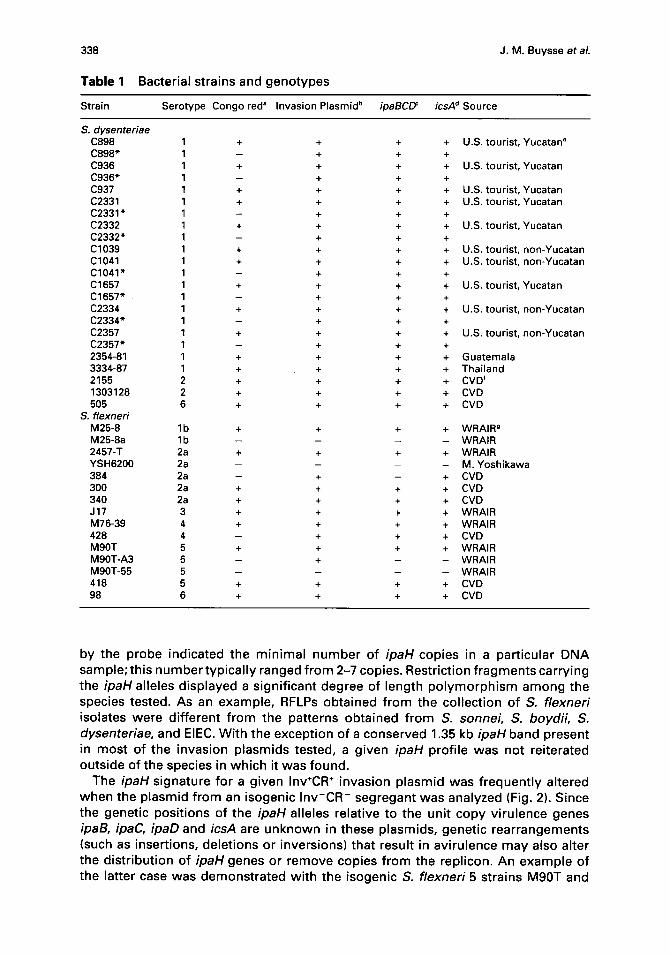
Table 1 Bacterial strains and genotypes
J. M . B u y s s e et al.
St ra in S e r o t y p e C o n g o red" I n v a s i o n P l a s m i d b ipaBCD ~ icsA d S o u r c e
S. dysenteriae C898 1 + + + +
C 8 9 8 " 1 - + + +
C936 1 + + + +
C 9 3 6 " 1 - + + +
C937 1 + + + +
C2331 1 + + + +
C 2 3 3 1 " 1 - + + +
C2332 1 + + + +
C 2 3 3 2 " 1 - + + +
C1039 1 + + + +
C1041 1 + + + +
C 1 0 4 1 " 1 - + + +
C1657 1 + + + +
C 1 6 5 7 " 1 - + + +
C2334 1 + + + +
C 2 3 3 4 " 1 - + + +
C2357 1 + + + +
C 2 3 5 7 " 1 - + + +
2354-81 1 + + + +
3334-87 1 + + + +
2155 2 + + + +
1303128 2 + + + +
505 6 + + + +
S. f lexned M 2 5 - 8 l b + + + +
M 2 5 - 8 a l b . . . .
2 4 5 7 - T 2a + + + +
Y S H 6 2 0 0 2a . . . .
384 2a - + - +
300 2a + + + +
340 2a + + + +
J17 3 + + + +
M 7 6 - 3 9 4 + + + +
428 4 - + + +
M 9 0 T 5 + + + +
M 9 0 T - A 3 5 - + - -
M 9 0 T - 5 5 5 . . . .
418 5 + + + +
98 6 + + + +
U.S. t o u r i s t , Y u c a t a n e
U.S. t ou r i s t , Y u c a t a n
U.S. t ou r i s t , Y u c a t a n
U.S. t o u r i s t , Y u c a t a n
U.S. t o u r i s t , Y u c a t a n
U.S. t o u r i s t , n o n - Y u c a t a n
U.S. t ou r i s t , n o n - Y u c a t a n
U.S. t o u r i s t , Y u c a t a n
U.S. t ou r i s t , n o n - Y u c a t a n
U.S. t ou r i s t , n o n - Y u c a t a n
G u a t e m a l a
T h a i l a n d CVD f
CVD
CVD
W R A I R 0
W R A I R
W R A I R
M. Y o s h i k a w a
CVD
CVD
CVD
W R A I R
W R A I R
CVD
W R A I R
W R A I R
W R A I R
CVD
CVD
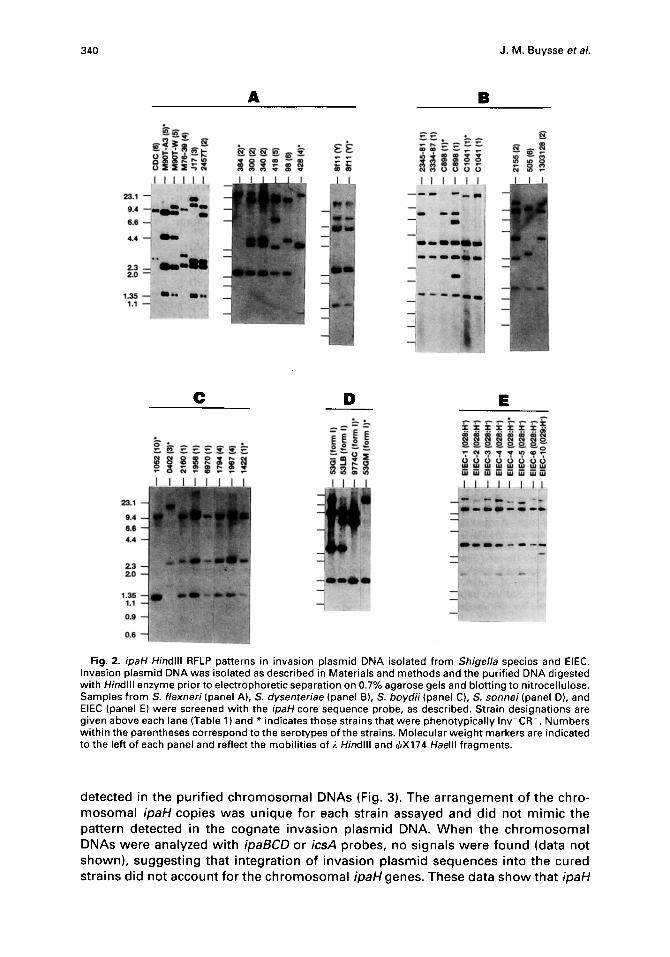
by the probe indicated the minimal number of ipaH copies in a particular DNA sample; this number typically ranged from 2-7 copies. Restriction fragments carrying the ipaH alleles displayed a significant degree of length polymorphism among the species tested. As an example, RFLPs obtained from the collection of S. flexneri isolates were different from the patterns obtained from S. sonnei, S. boydii, S. dysenteriae, and EIEC. With the exception of a conserved 1.35 kb ipaH band present in most of the invasion plasmids tested, a given ipaH profile was not reiterated outside of the species in which it was found.
The ipaH signature for a given Inv*CR * invasion plasmid was frequently altered when the plasmid from an isogenic Inv-CR- segregant was analyzed (Fig. 2). Since the genetic positions of the ipaH alleles relative to the unit copy virulence genes ipaB, ipaC, ipaD and icsA are unknown in these plasmids, genetic rearrangements (such as insertions, deletions or inversions) that result in avirulence may also alter the distribution of ipaH genes or remove copies from the replicon. An example of the latter case was demonstrated with the isogenic S. flexneri 5 strains M90T and

Genetic po l ymorph ism of ipaH
Table 1 continued
339
Strain Serotype Congo red ° Invasion Plasmid b ipaBCET icsA ~ Source
CDC 6 + + + + CDC h Sfl l Y + + + + A. Lindberg Sfl l Y - + - - A. Lindberg
S. sonnei 53GI Form I + + + + WRAIR 53GII Form II . . . . WRAIR 53LB Form I + + + + WRAIR 9774C Form I - + - + WRAIR 53GM Form I - + - + WRAIR
S. boydii 2160 1 + + + + CVD 1956 " 1 + + + + CVD 6970 1 + + + + CVD 1422 1 - + - - CVD 0402 3 - + - - CVD 1794 4 + + + + CVD 1967 4 + + + + CVD 1052 10 - + - - CVD
EIEC 1 028:H- + + + + CVD 2 028:H- + + + + CVD 3 028:H- + + - - CVD 4 028:H + + + + CVD 4 028:H- - + + + CVD 5 028:H + + + + CVD 6 028:H- + + + + CVD 10 029:H- + + + +i CVD
a Congo red posit ive cells were Inv" and Congo red negative cells were Inv- (HeLa cell invasion assay). b Invasion plasmid DNA content determined by the procedure of Cassie. 44 c Hybridization with 4.7 kb Hindlll f ragment probe of pHC17. 2s'48
Hybridization with 1.6 kb EcoRV-Hindlll f ragment probe of pHS3192. TM
eS. dysenteriae 1 strains recovered f rom U.S. shigellosis patients report ing travel to Mexico (CDC collection, 1988). A total of twenty-e ight Yucatan and eleven non-Yucatan isolates were screened.
fCenter for Vaccine Development, Universi ty of Maryland School of Medicine. ~Walter Reed Army Institute of Research. h Centers for Disease Control. Enteroinvasive E. coil These strains were provided by the CVD and were collected from various locations
in the U.S. (EIEC-3), Japan (EIEC-2,6), and South America (EIEC-1,4,5,10). i lncrease in Sail virG band size. * Indicates Inv-CR- isogenic segregant.
M 9 0 T - A 3 (F ig . 2A ) . M 9 0 T - A 3 c o n t a i n s a n e x t e n s i v e d i s c o n t i n u o u s d e l e t i o n t h a t
r e m o v e s t h e icsA, ipaBCDA, a n d mxi/spa l oc i . 2s'26 S o u t h e r n a n a l y s i s o f Hindlll- d i g e s t e d i n v a s i o n p l a s m i d f r o m t h e s e s t r a i n s i n d i c a t e d t h a t t h e ipaHg.8 a l l e l e w a s
d e l e t e d f r o m M 9 0 T - A 3 . A l t e r a t i o n s in t h e ipaH RFLP p a t t e r n s o f i s o g e n i c S. dysen- teriae 1 ( C 8 9 8 v s C 8 9 8 " ; C 1 0 4 1 vs . C 1 0 4 1 " ; F ig . 2B) , S. sonnei ( 53GI v s 5 3 G M ; F ig .
2D) a n d EIEC ( 0 2 8 : H - #4 ) (F ig . 2E) s t r a i n s w e r e f o u n d c o i n c i d e n t w i t h t h e l oss o f t h e
a b i l i t y t o i n v a d e c u l t u r e d e p i t h e l i a l ce l l s . It s h o u l d b e n o t e d , h o w e v e r , t h a t a l t e r a t i o n s
in p l a s m i d ipaH RFLPs w e r e n o t a b s o l u t e m a r k e r s o f t h e a v i r u l e n c e p h e n o t y p e s i n c e
a v i r u l e n t s e g r e g a n t s , s o m e o f w h i c h h a d l o s t t h e ipaBCD o r icsA g e n e s , d i d n o t
a l w a y s c o n t a i n a n a l t e r e d ipaH p a t t e r n (eg . S. flexneriY, s t r a i n S f l l ; F ig . 2A ) .
In a d d i t i o n t o t h e p l a s m i d - b o r n e c o p i e s o f ipaH, w e w a n t e d t o d e t e r m i n e i f ipaH s e q u e n c e s w e r e p r e s e n t in t h e c h r o m o s o m a l D N A o f Shigella s t r a i n s o t h e r t h a n S.
flexneri M 9 0 T . G e n o m i c D N A f r o m i n v a s i o n p l a s m i d - c u r e d s t r a i n s o f S. flexneri 5, S.
flexneri 2, S. flexneri 1, a n d S. sonnei w a s e x a m i n e d w i t h t h e ipaH c o r e s e q u e n c e
p r o b e d e s c r i b e d a b o v e . In a l l o f t h e s t r a i n s t e s t e d , f o u r t o s i x c o p i e s o f ipaH w e r e
340 J.M. Buysse et al.
i i i i
p l l l l l l :~..::.. ;.~ .,~,~
9,4 + . . . . :~;~.
A B
I I I I I I . . . . " ~ i -
ll~-,.~,
C D E
" - ~Eoo
I _ J _ _ L _ I _ J ~ I I
Fig. 2. ipaH Hindlll RFLP patterns in invasion plasmid DNA isolated from Shigella species and EIEC. Invasion plasmid DNA was isolated as described in Materials and methods and the purified DNA digested with Hindlll enzyme prior to electrophoretic separation on 0.7% agarose gels and blotting to nitrocellulose. Samples from S. flexneri (panel A), S. dysenteriae (panel B), S. boydii (panel C), S. sonnei (panel D), and EIEC (panel E) were screened with the ipaH core sequence probe, as described. Strain designations are given above each lane (Table 1) and * indicates those strains that were phenotypically Inv CR-. Numbers within the parentheses correspond to the serotypes of the strains. Molecular weight markers are indicated to the left of each panel and reflect the mobilities of ~. Hindlll and ~bX174 Haelll fragments.
de tec ted in the pu r i f i ed c h r o m o s o m a l DNAs (Fig. 3). The a r r a n g e m e n t o f t he chro- m o s o m a l ipaH cop ies w a s u n i q u e fo r each s t ra in assayed and d id no t m i m i c the pa t te rn de tec ted in the c o g n a t e i n v a s i o n p l a s m i d DNA. W h e n the c h r o m o s o m a l DNAs w e r e ana l yzed w i t h ipaBCD or icsA probes , no s igna l s w e r e f o u n d (data no t shown) , s u g g e s t i n g tha t i n t e g r a t i o n o f i n v a s i o n p l a s m i d s e q u e n c e s in to t he cured s t ra ins d id no t a c c o u n t fo r the c h r o m o s o m a l ipaHgenes. These da ta s h o w tha t ipaH

Genetic polymorphism of ipaH 341
kb f
P
1 2 3 !1 I I
C P C P
4 I I
C C
23.1 9.4
6.6
' m '1
W
4.4 i m '
2.3 2.0
m , l I
. . . i m
1,0
lira
Fig. 3. ipaH Hindlll RFLP patterns in chromosomal DNA prepared from invasion plasmid-cured Shigella species. Chromosomal DNA (C) from plasmid-cured species was electrophoresed next to the cognate plasmid DNA (P) and represented S. flexneri 5 (M90T/M90T-55) lane 1, S. flexneri 2a (2457T/YSH6200) lane 2, S. sonnei (53GI/53GII) lane 3, and S. flexneri lb (M25-8a) lane 4. The 627 bp ipaH core sequence probe was used.
genes were represented in more than one copy on both major replicons of the Shigella cell.
Analysis of ipaH gene structure in S. flexneri and S. sonnei invasion plasmids In a previous study it was shown that the five pWR100 ipaH alleles contained a conserved DNA sequence extending 627 bp between oligonucleotides H8 and H15 on the ipaHT~ gene (Fig. 1). 18 Outside of this conserved region, the nucleotide sequence diverged in the 5' and 3' ends of the ipaH4.5, ipaH25 and ipaH~.4 genes. To investigate whether ipaHsequence diversity observed in S. flexneri5 was also found in other sero- types of Shigella, oligonucleotides corresponding to various sections of the ipaHT.8 gene, including the core sequence, were end-labelled and hybridized to Hindlll- digested plasmid DNA from S. flexneri 1, S. flexneri 2, and S. sonnei, along with a S. flexneri 5 control (Fig. 4). A pattern of ipaH sequence variation, similar to that observed with S. flexneri 5, was seen with these strains. Oligonucleotide H2, cor- responding to the 5' end of pWR100 ipaH78, hybridized single ipaH copies in S. flexneri serotypes 1 and 2 and in S. sonnei. The H7 probe, overlapping a conserved sequence linked to the amino-terminal Sail site of ipaH78 and ipaH45 (Fig. 1), detected two copies of ipaH in S. flexneri 1, S. flexneri2, and S. sonnei. The H4 sequence was incorporated in ipaHg., and ipaHT.8 DNA sequences and partially overlapped the 3' end of ipaH1.4 in pWR100; used against these invasion plasmids, H4 detected two

342 J.M. Buysse et al.
A . So
_L_CLLL.L 23.1 23.1 - ' L ~ ~ - ~ " ~ l
9.4 9.4
6.6 6.6
4.4 4.4
2.3 2.3 2.0 2.0
1.0 1.0
C =
23 .1 - ~ ~ 1 9 . 4 - - II,
6 6 - '
. . , . ~!~
2 . 3 - 2 . 0 -
1.0-- ': i~.~
I
2.3-- ]" 1.0 2
i
Om ff Fig. 4. An analysis of ipaH gene structure in the invasion plasmids of S. flexneri serotype 1, strain M25-
8 (panel A), S. flexneri, serotype 2, strain 2457-T (panel B), S. flexneri serotype 5, strain M90T (panel C), and S. sonnei Form I, strain 53GI (panel D). The core probe used in each case was a 32P-labelled PCR amplification product obtained with primers H8 and H15, as in Fig. 3; 24-mer oligonucleotide probes H2, H7, H10, and H4, along with probes H24, and H25 (Fig. 1 and see Materials and methods) were also used to screen the plasmids.
large ipaH Hindlll f ragments in S. flexneri 1, S. flexneri 2 and S. sonnei, and a small 1.35 kb band in S. flexneri 2. As expected, the H10 sequence def ined the 3' l imit of the conserved core region and hybr id ized the major i ty of ipaH copies in the four p lasmids tested. These results indicate that ipaH genes carr ied by o ther invasion p lasmids are genet ica l ly d ivergent and are not s imple re i terat ions of a s ingle gene locus. Sequence d ivergence outs ide of the conserved core region may there fore be a c o m m o n feature of ipaH alleles in Shigella species.
DNA sequences immed ia te ly f lanking the stop codons of ipaH4s and ipaH25 were

Genetic polymorphism of ipaH 343
identical for at least 250 bp downstream of their respective 3' ends. TM Sequence identity was also noted for the regions upstream of methionine start codons in ipaH2.5 and ipaHi.4. To determine if this molecular configuration was found in other Shigella invasion plasmids, oligonucleotide probes H24 and H25 (Fig. 1) were generated for Southern blot analysis of Hindlll-digested invasion plasmid DNAs (Fig. 4). H24, derived from the shared sequence upstream of the ipaH2.s and ipaH~.4 start codons, hybridized a limited number of ipaH bands identified by the core sequence probe and, in addition, at least one other band not recognized by the core probe. The H24 oligonucleotide probe was found to be Shigella-specific, in that colony blots using this probe to detect homologous sequences in ETEC, EPEC, and RDEC E. coli strains were negative, as were attempts to find the element in a panel of 200 enteric strains that contained representative isolates of Salmonella, Aeromonas, Proteus, Cam- pylobacter, Klebsiella, Citrobacter, Yersinia, and Vibrio (18; see Materials and methods). A 33 bp sequence contiguous with the 3' end of ipaH4.5 and ipaH2.s (Fig. 1) comprised the H25 probe. Hybridization with H25 mimicked in part the core sequence hybridization patterns, with the addition of other unique bands in each case exam- ined. Thus, for S. flexneri 5, the 4.5 and 2.5 kb ipaH bands were detected with the H25 probe along with four new bands not detected with the core sequence probe. Examination of the GENBANK database using the 250 bp segment that incorporates H25 and flanks the 3' end of ipaH4.s demonstrated that this sequence was identical with the 5' end of insertion sequence IS629, a 1.2 kb iso-lS3 homologue originally described in S. sonneL 27'28 Further, colony blot hybridization experiments indicated that H25 sequences were present on ETEC, EPEC and RDEC E. coli strains (data not shown), but were not found in the enteric strain panel described above. These results suggest that certain ipaH alleles are genetically linked with distinct repetitive elements in S. flexneri serotypes and S. sonneL
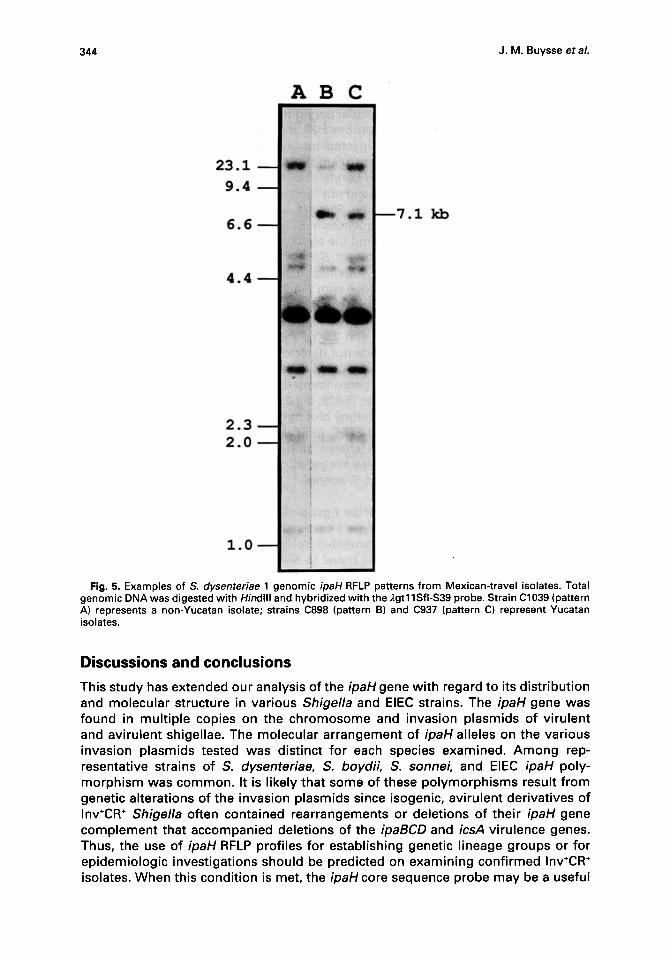
Analysis of ipaH RFLPs in S. dysenteriae 1 genomic DNA During 1988, 51 S. dysenteriae serotype 1 cases were identified in the U.S. through an enhanced surveillance effort implemented by the Centers for Disease Control. 29'3° Forty of the 51 patients had reported travel to Mexico and 29 of these had visited Cancun or other Yucatan Peninsula resort areas. We wanted to determine if genomic ipaH RFLP profiles might be useful in assessing clonal and geographic relationships among the isolates. Total genomic DNA was prepared from 39 of the Mexican-travel isolates, digested with Hindlll and probed with S39 ipaH DNA. Three patterns of ipaH hybridization were detected among these samples (Fig. 5), with ipaH Hindlll fragments of 1.4 kb, 2.1 kb, 3.0 kb, 3.8 kb, 4.6 kb and >23 kb common in each pattern type. S. dysenteriae 1 cultures recovered from travellers to the Yucatan peninsula (n=28) and from those .that had visited other areas in Mexico (n=11) were then screened in the HeLa cell invasion assay to obtain isogenic Inv+CR ÷ and Inv-CR- segregants. Genomic DNA from these strains was examined with the ipaH core sequence probe and the 4.7 kb Hindlll ipaBCD probe; an example of the hybridization results is presented in Fig. 6. Yucatan isolates C2331 and C2332 contained a 7.1 kb ipaH Hindlll fragment that was found in each of the Yucatan isolates analyzed. The ipaH RFLP patterns of the 28 Yucatan strains examined corresponded either to pattern B (19/28) or pattern C (9/28), whereas 9/11 non-Yucatan strains, represented by isolates such as C2334 and C2357, did not harbor the 7.1 kb fragment and matched pattern A (Fig. 5; Fig. 6). With the exceptions of C898 and C1041 (Fig. 2), no changes in ipaH RFLP profiles were detected between isogenic Inv÷CR ÷ and Inv-CR- S. dysen- teriae 1 clones, the latter retaining the ipaBCD and icsA sequences in all cases (Fig. 6B; Table 1).
344 J.M. Buysse et al.
AB C
2 3 . 1
9 .4
6 . 6 k b
4 . 4
2 . 3 2 . 0
I-1 Fig. 5. Examples of S. dysenteriae 1 genomic ipaH RFLP patterns from Mexican-travel isolates. Total
genomic DNA was digested with Hindlll and hybridized with the ),gtl 1Sfi-$39 probe. Strain C1039 (pattern A) represents a non-Yucatan isolate; strains C898 (pattern B) and C937 (pattern C) represent Yucatan isolates.
Discussions and conclusions
This study has extended our analysis of the ipaHgene with regard to its distribution and molecular structure in various Shigella and EIEC strains. The ipaH gene was found in multiple copies on the chromosome and invasion plasmids of virulent and avirulent shigellae. The molecular arrangement of ipaH alleles on the various invasion plasmids tested was distinct for each species examined. Among rep- resentative strains of S. dysenteriae, S. boydii, S. sonnei, and EIEC ipaH poly- morphism was common. It is likely that some of these polymorphisms result from genetic alterations of the invasion plasmids since isogenic, avirulent derivatives of Inv÷CR ÷ Shigella often contained rearrangements or deletions of their ipaH gene complement that accompanied deletions of the ipaBCD and icsA virulence genes. Thus, the use of ipaH RFLP profiles for establishing genetic lineage groups or for epidemiologic investigations should be predicted on examining confirmed Inv+CR ÷ isolates. When this condition is met, the ipaH core sequence probe may be a useful

Genetic polymorphism of ipaH 345
A. B.
Yucatan n~-Yucatan I . . I I , ~1
23.1 ~
Y u c a t a n non-Yucatan I II [ I
I I I I I I I . I I
-~,, i ~
Fig. 6. Analysis of ipaH RFLP profiles in Inv*CR* and Inv CR S. dysenteriae 1 isolates. Genomic DNA from Yucatan and non-Yucatan S. dysenteriae 1 cultures was digested with Hindlll and probed with ipaH core sequence DNA (panel A) and pHC17 ipaBCDinsert DNA (panel B). Strain designations and genotypes are reported in Table 1; * indicates Inv CR segregants. Genomic DNA from M90T was included as a positive control in each panel.
tool to complement various molecular markers used in monitoring the origin and spread of particular shigellosis outbreaks. As a demonstration of this potential, ipaH hybridization profiles obtained by Southern blot analysis of a collection of S. dysenteriae 1 genomic DNAs revealed reproducible and distinct RFLP patterns that correlated with the geographic origin of the strains and the travel histories of the patients from which the strains were collected.
IpaHalleles display significant gene polymorphism and are located on both major replicons of the Shigella cell. In addition, the multiple copies of ipaH detected in a given Shigella species' invasion plasmid were not structurally identical and contained significant sequence divergence from the prototypical ipaH78 gene of S. flexneri serotype 5. These observations suggest a recombination mechanism for the dis- semination of ipaH genes within and between the invasion plasmid and chro- mosome. Recombination processes are responsible for the remarkable diversity of pilin and Opa sequences documented in Neisseria gonnorheae 33"34 and probably generate antigenic variation in Vmp proteins of Borrelia species. 35 Additional mech- anisms that account for gene polymorphism include SOS repair fol lowing DNA damage and insertion/excision of mobile genetic elements, especially those closely associated with the target gene. The latter mechanism is interesting in view of the direct linkage of IS629 with ipaH45 and ipaH2.5 in S. flexnerL It is possible that IS629 serves as a recombination vehicle, shuffling the ipaH array as a result of IS629 transposition. The products of such recombination events might also determine which ipaH gene copies are expressed within the cell, much as IS I transposition can influence virF gene expression and the virulence phenotype in S. flexnerL 36
The high level of ipaH reiteration within shigellae genomes broadens the list of repeated elements found in Shigella species. Numerous copies of IS I, IS2, IS3, IS629, IS600, IS630, and IS911 have been documented in S. flexneri, S. sonnei and S. dysenteriae. 27'~7 In some instances, 30 or more copies of a particular insertion

346 J.M. Buysse et al.
sequence have been detected, in comparison to E. coli K12 which contains less than 10 of each element. Oligonucleotides H24 and H25 revealed additional sets of repeated elements on the invasion plasmids tested. The H24 probe hybridized a low copy number repeat (2-3) unique to Shigella species that was not found in the enteric bacteria panel examined in this study. The H25 probe, corresponding to IS629, defined six or more copies of this insertion sequence in the Shigella plasmids tested. IS629 linkage with the ipaH4.5 and ipaH2.s genes is reminiscent of the genetic linkage between repetitive elements and virulence genes that has been noted before, with the best examples found for the Vibrio chlorae 01 CT× genetic element and the vlp gene clusters in Mycoplasma hyorhinis. 31'32 It should be noted however, that a virulence-related function for the ipaH genes has not been established to date. Shigella species are also distinguished by their collection of iso-insertion sequences, that is, elements that have sequence similarity to a progenitor insertion element; S. dysenteriae alone carries over 200 copies of iso-lS 1 elementsY The heterogeneity and sequence divergence of ipaH copies within a given strain follows the pattern established for IS and iso-lS elements in Shigella.
Genetic polymorphism of the ipaH locus can be exploited to develop a Shigella- specific DNA probe useful in epidemiologic studies of Shigella transmission. The most commonly used epidemiolog!cal techniques rely on biochemical tests for Shi- gella identification coupled with serotyping and determination of antimicrobial sus- ceptibility patterns in isolates obtained from an infected population, z8 These techniques have been complemented by the use of various molecular markers to monitor the origin and spread of particular shigellosis outbreaks, including plasmid fingerprinting and plasmid restriction fragment analysis, zs'4° Geotypic methods that increase the level of specificity in Shigella epidemiologic investigations include the examination of RFLPs with defined gene probes (e.g. rRNA) and the use of species- specific gene probes to rapidly and efficiently detect shigellae in clinical samples. 21'3°'41'42 By using an ipaH core sequence probe, the combination of RFLP analysis with a species-specific gene probe is feasible. IpaH RFLP analysis, in con- junction with similar analyses using Shiga toxin gene (stx/slt) or rRNA gene probes, might be expected to provide good discrimination among Shigella isolates and perhaps point out clonal differences between strains isolated from distinct geo- graphic locations. As an example, the collection of S. dysenteriae 1 isolates examined in this study had been previously shown to have uniform RFLP patterns with either Shiga toxin or rRNA probes. 29'3° By contrast, analysis of the same S. dysenteriae 1 samples using the ipaHT.s core probe revealed three distinct RFLP patterns. The ipaHT.8 core probe thus provides a reliable molecular tool for the specific detectibn of shigellae (virulent and avirulent) and may be useful in studying the molecular epidemiology of Shigella infection and spread.
Materials and methods
Bacteria and microbiological procedures. Representative S. flexneri, S. sonnei, S. boydii, S. dysenteriae and EIEC strains were obtained from a variety of sources, summarized in Table 1. After recovery from lyophile or agar slant by overnight growth in L-broth (37°C), each culture was individually plated onto tryptic soy agar plates supplemented with 0.05% Congo red dye and 0.2% galactose and incubated overnight at 39°C. Congo red positive (CR ÷) and Congo red negative (CR-) isolates from each strain were then tested in the HeLa cell invasion assay, as previously described, 43 to obtain invasion-positive (Inv ÷) and invasion-negative (Inv-) clones. Serotyping of the bacteria was done with typing antisera obtained from Difco Laboratories (Detroit, MI, U.S.A.) specific for serotypes of S. flexneri, S. sonnnei, S. boydii, S. dysenteriae, and EIEC U-serogroup antigens 028 and 029. A panel of enteric bacteria, described in previous

Genetic polymorphism of ipaH 347
reports, 24."2 containing 200 isolates representing several gram-negative pathogens was used in DNA colony blot hybridizations to test the various probes described below.
DNA isolation. Invasion plasmid DNA was prepared from Inv*CR ÷ and Inv-CR- cells by the procedure of Cassie et al. 44 and was purified by two cycles of CsCI-ethidium bromide density gradient ultracentrifugation. Total genomic DNA and chromosomal DNA from plasmid-cured cells was prepared using an extraction technique that incorporates a hexadecyltrimethyl ammonium bromide precipitation of polysaccharides and proteins fol lowed by extraction with phenol/chloroform/isoamyl alcohol and isopropanol precipitation of the DNAo 4s.
DNA probes and hybridization conditions. Purified DNA (total genomic, chromosomal or invasion plasmid) was digested with Hindlll (Boehringer-Mannheim), according to manu- facturer's recommendations, and the fragments separated on 0.7% agarose gels using Tris- borate/l~DTA (pH 8.3) buffer. The DNA fragments were transferred to nitrocellulose filters and hybridized with selected DNA probes (Fig. 1). S. flexneri 5 pWR100 gene sequences were used to probe the various DNA samples and included: (i) a 2.9 kb EcoRI insert from recombinant phage ~gtl 1Sfi-$39 that contains the entire ipaH?.8 sequence and 600 bp of the amino terminal end of the contiguous ipaH~.s gene (probe $39)8.'.18; (ii) a 627 bp PCR-amplification product from an interior, conserved region of the ipaHT.8 gene (the ipaH core probe), extending from the amino-end primer H8 (5'-GCGI-rCCI-rGACCGCCTTTCCGATACCG-3') to the carboxy-end primer H15 (5'C'I-I-I'CAGCCGGTCAGCCACCCCTCTGAGAG-3'); (iii) oligonucleotide probes H2, H7, H10, and H4, corresponding to 24 bp oligomers derived from the ipaHT.8 DNA sequence; 18 (iv) H24 (5'-TGI-ICTGGAGCGGAAATTGTGGCAGAAI-I-FCCC-3') and H25 (5'-TGAACCGCC CCGGGAATCCTGGAGACTAAAC-3') probes; H24 overlaps the first ten codons of ipaHz6, which is not depicted in Fig. 1; '(v) a 4.7 kb Hindlll fragment of pHC1725.28 carrying the ' ipgB-ipgC-ipaB- ipaC-ipaD' genes; and (vi) a 1.6 kb EcoRV-Hindlll segment of the icsA gene excised from the icsA-recombinant plasmid pHS3192. TM Nick translation was used to radiolabel double-stranded DNA probes and Southern hybridization was performed using standard conditions described earlier. ~? Oligonucleotide probes were end-labelled at 37°C for 30 min with 66 pmoles of ~- [32P]-ATP in a l X forward reaction buffer using 10 units of T4 polynucleotide kinase (5' DNA terminus labelling kit, BRL). Hybridizations with the labelled oligonucleotide probes were carried out using previously standardized conditions. TM The ipaH core sequence probe was generated using Taq polymerase, 10 ng of ~gt l lSf l-S39 template DNA, 50 pmoles each of primers H8 and H15 and reaction components provided in the Perkin-Elmer PCR Amplif ication Kit. All probes derived from plasmid or phage recombinants or from PCR reactions were gel- purified over a Gene Clean matrix (BIO101).
The authors wish to acknowledge Ben Tall for strains received from the Center for Vaccine Department, The University of Maryland School of Medicine. We also thank AIf Lindberg for the gift of strain Sf l l , Masanosuke Yoshikawa for strain YSH6200 and Phillipe Sansonettii for the donation of plasmid pHS3192. We thank Kathy Kronke for expert preparation of this manuscript.
References
1. Hale TL. Genetic basis of virulence in Shigella species. Microbiol Rev 1991; 55: 206-24. 2. Oaks EV, Hale TL Formal SB. Serum immune response to Shigella protein antigens in rhesus monkeys
and humans infected with Shigella spp. Infect Immun 1986; 53: 57-63. 3. Oaks EV, Turbyfill KR. Myosin-cross-reactive epitope of Shigella flexneriinvasion plasmid antigen B.
Infect Immun 1992; 60: 557-64. 4. Barzu S, Nato F, Rouyre S, Mazie J-C, Sansonetti P J, Phalipon A. Characterization of B-cell epitopes
on IpaB, an invasion-associated antigen of Shigella flexneri: identification of an immunodominant domain recognized during natural infection. Infect Immun 1993; 61: 3825-31.
5. Sansonetti P J, Kopecko D J, Formal SB. Shigella sonnei plasmids: evidence that a large plasmid is necessary for virulence. Infect Immun 1981; 34: 75-83.
6. Sansonetti P J, Kopecko D J, Formal SB. Involvement of a plasmid in the invasive ability of Shigella flexneri. Infect Immun 1982; 35: 852-60.
7. Baudry B, Maurelli AT, Clerc P, Sadoff JC, Sansonetti PJ. Localization of plasmid loci necessary for the entry of Shigella flexneri into HeLa cells, and characterization of one locus encoding four immunogenic polypeptides. J Gen Microbio11987; 133: 3403-13.
8. Buysse JM, Stover CK, Oaks EV, Venkatesan MM, Kopecko DJ. Molecular cloning of invasion plasmid

348 J.M. Buysse et al.
antigen (ipa) genes from Shigella flexneri: analysis of ipa gene products and genetic mapping. J Bacteriol 1987; 169: 2561-9.
9. Sasakawa C, Kamata K, Sakai T et aL Virulence-associated genetic regions comprising 31 kilobases of the 230-kilobase plasmid in Shigella flexneri 2a. J Bacteriol 1988; 170: 2480-4.
10. Mdnard R, Sansonetti P J, Parsot C. Nonpolar mutagenesis of the ipa genes defines IpaB, IpaC and IpaD as effectors of Shigella flexneri entry into epithelial cells. J Bacteriol 1992; 175: 5899-906.
11. High N, Mounier J, Pr6vost MC, Sansonetti PJ. IpaB of Shigella flexneri causes entry into epithelial cells and escape from the phagocytic vacuole. EMBO J 1992; 11: 1991-9.
12. Zychlinsky A, Kenny B, M~nard R, Prdvost MC, Holland IB, Sansonetti PJ. IpaB mediates macrophage apoptosis induced by Shigella flexneri. Molec Microbiol 1994; 11: 619-27.
13. Bernardini ML, Mounier J, D'Hauteville H, Coquis-Rondon M, Sansonetti PJ. Identification of icsA, a plasmid locus of Shigella flexneri that governs bacterial intra- and intercellular spread through interaction with F-actin. PNAS 1989; 86: 3867-71.
14. Goldberg MB, Barzu O, Parsot C, Sansonetti PJ. Unipolar localization and ATPase activity of IcsA, a Shigella flexneri protein involved in intracellular movement. J Bacteriol 1993; 175: 2189-96.
15. Lett MC, Sasakawa C, Okada N e t aL virG, a plasmid-coded virulence gene of Shigella flexneri: Identification of the virG protein and determination of the complete coding sequence. J Bacteriol 1989; 171: 353-9.
16. Venkatesan MM, Buysse JM. Nucleotide sequence of invasion plasmid antigen gene ipaA from Shigefla flexneri 5. NAR 1990; 18: 1648.
17. Hartman AB, Venkatesan M, Oaks EV, Buysse JM. Sequence and molecular characterization of a multicopy invasion plasmid antigen gene, ipaH, of Shigella flexneri. J Bacteriol 1990; 172: 1905-15.
18. Venkatesan MM, Buysse JM, Hartman AB. Sequence variation in two ipaHgenes of Shigella flexneri 5 and homology to the LRG-like family of proteins. Molec Microbiol 1991; 5: 2435-45.
19. Kobe B, Deisenhofer J. The leucine-rich repeat: a versatile binding motif. Trends Biochem Sci 1994; 19: 415-21.
20. Islam MS, Hasan MK, Miah MA et aL Use of the polymerase chain reaction and fluorescent-antibody methods for detecting viable but nonculturable Shigella dysenteriae type 1 in laboratory microcosms. Applied Environ Microbiol 1993; 59: 536-40.
21. Oberhelman RA, Kopecko D J, Venkatesan MM et aL Evaluation of alkaline phosphatase-labelled ipaH probe for diagnosis of Shigella infections. J Clin Microbiol 1993; 31: 2101-4.
22. Sethabutr O, Venkatesan M, Murphy GS, Eampokalap B, Hoge CW, Echeverria P. Detection of shigellae and enteroinvasive Escherichia coli by amplification of the invasion plasmid antigen H DNA sequence in patients with dysentery. J Infect Dis 1993; 167: 458-61.
23. Venkatesan MM, Buysse JM, Kopecko DJ. DNA sequence homology among ipa genes of Shigella spp. and enteroinvasive Escherichia coil In: Talwar GP, ed. Progress in Vaccinology. New York: Springer-Verlag, 1989; 406-422.
24. Venkatesan MM, Buysse JM, Kopecko DJ. Use of Shigella flexneri ipaC and ipaH gene sequences for the general identification of Shigella spp. and enteroinvasive Escherichia coil J Clin Microbiol 1989; 27: 2687-91.
25. Buysse JM, Venkatesan M, Mills JA, Oaks EV. Molecular characterization of a trans-acting, positive effector (ipaR) of invasion plasmid antigen synthesis in Shigella flexneri serotype 5. Microb Pathog 1990; 8:197-211.
26. Venkatesan MM, Fernandez-Prada C, Buysse JM, Formal SB, Hale TL. Virulence phenotype and genetic characteristics of the T32-1STRATI Shigella flexneri2a vaccine strain. Vaccine 1991; 9: 358-63.
27. Matsutani S, Ohtsubo H, Maeda Y, Ohtsubo E. Isolation and characterization of IS elements repeated in the bacterial chromosome. J Mol Biol 1987; 196: 445-55.
28. Ishiguro N, Sato G. Nucleotide sequence of insertion sequence IS3411, which flanks the citr#te utilization determinant of transposon Tn341 I. J Bacteriol 1988; 170: 1902-6.
29. Parsonnet J, Greene KD, Gerber AR, Tauxe RV, Vallejo-Aguilar O J, Blake PA. Shigella dysenteriae type 1 infections in US travellers to Mexico. Lancet 1989; 2: 543-5.
30. Stockbine NA, Parsonnet J, Greene K, Kiehlbauch JA, Wachsmuth IK. Molecular epidemiologic tech- niques in analysis of epidemic and endemic Shigella dysenteriaetype 1 strains. J Infect Dis 1991; 163: 406-9.
31. Waldor MK, Mekalanos I-F. Emergence of a new cholera pandemic: molecular analysis of virulence determinants in Vibrio cholerae O139 and development of a live vaccine prototype. J Infect Dis 1994; 170: 278-83.
32. Wise K. Adaptive surface variation in mycoplasmas. Trends Microbiol 1993; 1: 59-63. 33. Wainwright LA, Hoikka-Pritchard K, Seifert HS. A conserved DNA sequence is required for efficient
gonococcal pilin antigenic variation. Molec Microbiol 1994; 13: 75-87. 34. Meyer T. In: Iglewski BH, Clark VL, eds. Molecular Basis of Bacterial pathogenesis. San Diego:
A,'ademic Press, 1990; 137-53. 35. Rosa PA, Schwan T, Hogan D. Recombination between genes encoding major outer surface proteins
A and B of Borrelia burgdorferi. Molec Microbiol 1992; 6: 3031-40. 36. Mills JA, Venkatesan MM, Baron LS, Buysse JM. Spontaneous insertion of an IS/-like element into
the virF gene is responsible for avirulence in opaque colonial variants of Shigella flexneri 2a. Infect Immun 1992; 60: 175-82.

Genetic polymorphism of ipaH 349
37. Ohtsubo E, Ohtsubo H, Doroszkiewicz W, Nyman K, Allen D, Davison D. An evolutionary analysis of ISO-IS 1 elements from Escherichia coli and Shigella strains. J Gen Appl Microbiol 1984; 30: 359-76.
38. Wharton M, Spiegel RA, Horan JM et al. A large outbreak of antibiotic-resistant Shigellosis at a mass gathering. J Infect Dis 1990; 162: 1324-8.
39. Haider K, Kay BA, Talukder KA, Huq MI. Plasmid analysis of Shigella dysenteriae Type 1 isolates obtained from widely scattered geographical locations. J Clin Microbiol 1988; 26: 2083-6.
40. Litwin CM, Storm AL, Chipowsky S, Ryan KJ. Molecular epidemiology of Shigella infections: Plasmid profiles, serotype correlation, and restriction endonuclease analysis. J Clin Microbiol 1991, 29: 104- 8.
41. Frankel G, Riley L, Giron JA et al. Detection of Shigella in feces using DNA amplification. J Infect Dis 1990; 161: 1252-6.
42. Venkatesan MM, Buysse JM, Vandendries E, Kopecko DJ. Development of testing of invasion-associ- ated DNA probes for detection of Shigella spp. and enteroinvasive Escherichia coli. J Clin Microbiol 1988; 26: 261-6.
43. Elsinghorst EA. Measurement of invasion by gentamicin resistance. Methods Enzymol 1994; 236: 405-2.0.
44t Cassie F, Boucher C, Julliot JS, Michael M, Denaire J. Identification and characterization of large plasmids in Rhizobium meliloti using agarose gel electrophoresis. J Gan Microbiol 1979; 113: 229- 42.
45. Wilson K. Preparation of genomic DNA from bacteria. In: Ausubel FM, Brant R, Kingston RE, Moore DD, Seidman JG, Smith JA, Struhl K, eds. Current protocols in Molecular Biology. New York: John Wiley and Sons, 1987; 2.4.1.-2.4.5.
46. Venkatesan MM, Buysse JM, Kopecko DJ. Characterization of invasion plasmid antigen genes (ipaBCD) from Shigella flexneri. Proc Natl Acad Sci 1988; 85: 9317-21.