gnld ttrn n rl nd prt ardll - ilslila.ilsl.br/pdfs/v54n2a09.pdf · ieaioa oua o eosy ^ l 5 br 2....
TRANSCRIPT

INTERNATIONAL JOURNAL OF LEPROSY^ Volume 54, Number 2
Printed in the U.S.A.
Ganglioside Patterns in Normal andLepromatous Armadillo Tissues'Eugene B. Harris, Yu-Teh Li, and Su-Chen Liz
Gangliosides, also referred to as glyco-sphingolipids, are unique glycolipids pres-ent in the plasma membranes of mamma-lian cells and are particularly abundant inneural tissues. Gangliosides have attractedconsiderable attention in recent years asparticipants in a variety of cellular func-tions. A number of investigations have im-plicated these complex carbohydrates in celldifferentiation and growth, as surface mem-brane receptors, and as glycolipid antigenshaving a role in regulating immune re-sponses ( 4).
Gangliosides, as the most specific lipidsof neurons, very likely have a role in neu-rofunction ( 7). It is well known that Myco-bacterium leprae are associated with pe-ripheral nerves. Electron microscopicstudies, for example, clearly show largenumbers of bacilli within the cytoplasm ofSchwann cells ( 5). Schwann cells are in-volved in the synthesis of the characteristicgangliosides present in peripheral nerves ( 9).
Many nine-banded armadillos developdisseminated (lepromatous) leprosy wheninoculated with M. leprae (6). The ubiquityof gangliosides in animal tissues and theirpossible involvement in neurofunction ledus to investigate ganglioside patterns in M./eprae-infected armadillo tissues.
MATERIALS AND METHODSExperimental animals. Armadillos were
inoculated intravenously with suspensionsof Al. leprae and maintained in the animalcare facility at the National Hansen's Dis-
I Received for publication on 28 February 1985; ac-cepted for publication in revised form on 14 February1986.
2 E. B. Harris, B.S., Research Chemist, BiochemistryResearch Department, Laboratory Research Branch,Gillis W. Long Hansen's Disease Center (NationalHansen's Disease Center), Carville, Louisiana 70721.Y.-T. Li, Ph.D., Professor; S.-C. Li, Ph.D., ResearchProfessor, Department of Biochemistry, Tulane Uni-versity School of Medicine, New Orleans, Louisiana70112, U.S.A.
ease Center, Carville, Louisiana, U.S.A. Af-ter signs of dissemination appeared, the an-imals were sacrificed under anesthesia bycardiac exsanguination. The tissues were re-moved aseptically and stored at —20° or—80°C before processing. The tissues wereexamined histologically by acid-fast stain-ing for the presence of Al. leprae. Tissuefrom normal (non-inoculated) armadillosserved as controls.
Preparation of bacilli. M. leprae wereseparated from the liver tissue of an exper-imentally infected armadillo, as describedpreviously (").
Extraction of tissue ganglioside. Each tis-sue sample and the purified bacilli were ho-mogenized in 10 volumes of acetone, cen-trifuged at 500 x g for 10 min, and theresidue collected. The residue was extractedsuccessively with 10 volumes each of chlo-roform-methanol (2:1), chloroform-meth-anol (1:1), and chloroform-methanol (1:2).The chloroform-methanol extracts werecombined and concentrated in vacuo at 45-50°C, then taken to dryness under a streamof nitrogen. The dried samples (crude lipidfractions) were further extracted accordingto the partitioning procedure of Folch, et al.( 3) unless otherwise noted.
Thin-layer chromatographic (TLC) anal-ysis. Aliquots of the ganglioside extractsalong with the appropriate standards wereapplied to silica gel plates (0.25 mm; E.Merck, Darmstadt, Germany) and devel-oped in chloroform-methanol-water (60:35:8). Gangliosides were located by sprayingthe plates with resorcinol reagent ( 13) andheating them at 110°C for 15 min.
GM3 Ganglioside purification. A crudelipid extract from 100 g Al. /eprae-infectedarmadillo liver was submitted to mild al-kaline hydrolysis by suspending the extractin 10 ml of 0.6 N NaOH in methanol, andincubating at 37°C for 16 hr ('). The samplewas neutralized with 1 N methanolic HC1,then dried, dialyzed thoroughly againstwater, and lyophilized. The Folch partition
289
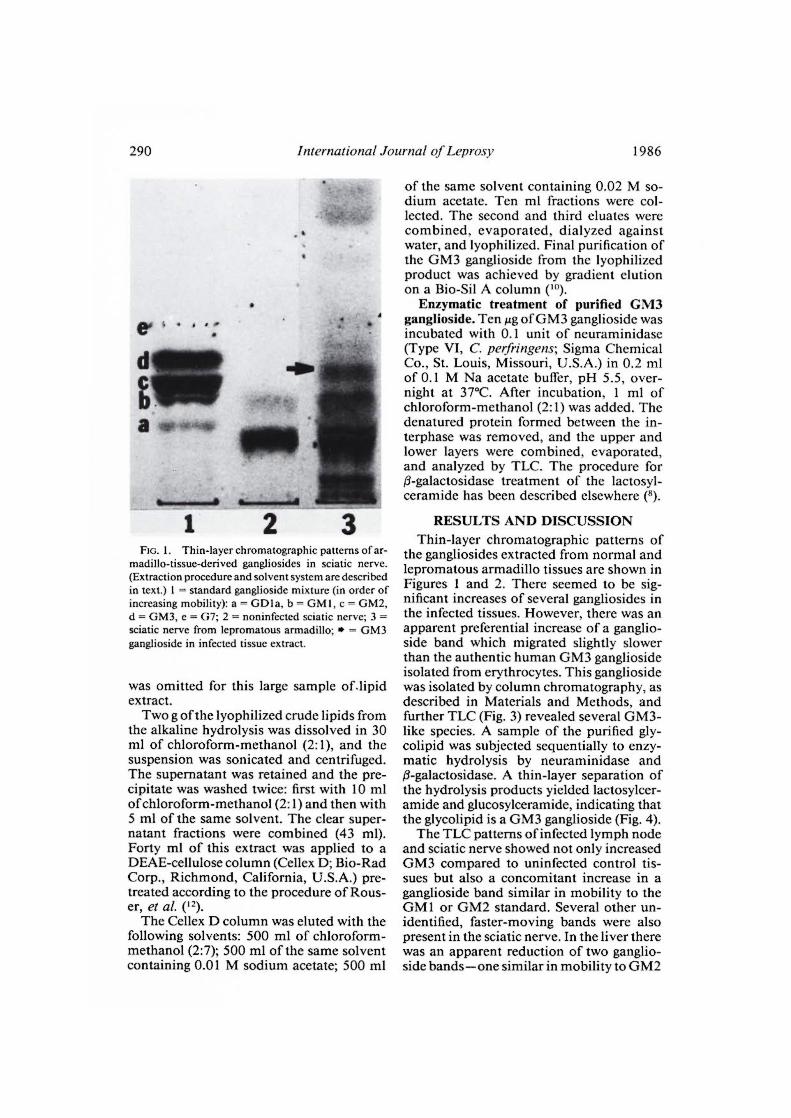
290^ International Journal of Leprosy^ 1986
FIG. 1. Thin-layer chromatographic patterns of ar-madillo-tissue-derived gangliosides in sciatic nerve.(Extraction procedure and solvent system are describedin text.) 1 = standard ganglioside mixture (in order ofincreasing mobility): a = GD1a, b = GM I, c = GM2,d = GM3, e = G7; 2 = noninfected sciatic nerve; 3 =sciatic nerve from lepromatous armadillo; • = GM3ganglioside in infected tissue extract.
was omitted for this large sample of.lipidextract.
Two g of the lyophilized crude lipids fromthe alkaline hydrolysis was dissolved in 30ml of chloroform-methanol (2:1), and thesuspension was sonicated and centrifuged.The supernatant was retained and the pre-cipitate was washed twice: first with 10 mlof chloroform-methanol (2:1) and then with5 ml of the same solvent. The clear super-natant fractions were combined (43 ml).Forty ml of this extract was applied to aDEAE-cellulose column (Cellex D; Bio-RadCorp., Richmond, California, U.S.A.) pre-treated according to the procedure of Rous-er, et al. ( 12).
The Cellex D column was eluted with thefollowing solvents: 500 ml of chloroform-methanol (2:7); 500 ml of the same solventcontaining 0.01 M sodium acetate; 500 ml
of the same solvent containing 0.02 M so-dium acetate. Ten ml fractions were col-lected. The second and third eluates werecombined, evaporated, dialyzed againstwater, and lyophilized. Final purification ofthe GM3 ganglioside from the lyophilizedproduct was achieved by gradient elutionon a Bio-Sil A column ( 1 ").
Enzymatic treatment of purified GM3ganglioside. Ten lig of GM3 ganglioside wasincubated with 0.1 unit of neuraminidase(Type VI, C. perfringens; Sigma ChemicalCo., St. Louis, Missouri, U.S.A.) in 0.2 mlof 0.1 M Na acetate buffer, pH 5.5, over-night at 37°C. After incubation, 1 ml ofchloroform-methanol (2:1) was added. Thedenatured protein formed between the in-terphase was removed, and the upper andlower layers were combined, evaporated,and analyzed by TLC. The procedure for0-galactosidase treatment of the lactosyl-ceramide has been described elsewhere ( 8).
RESULTS AND DISCUSSIONThin-layer chromatographic patterns of
the gangliosides extracted from normal andlepromatous armadillo tissues are shown inFigures 1 and 2. There seemed to be sig-nificant increases of several gangliosides inthe infected tissues. However, there was anapparent preferential increase of a ganglio-side band which migrated slightly slowerthan the authentic human GM3 gangliosideisolated from erythrocytes. This gangliosidewas isolated by column chromatography, asdescribed in Materials and Methods, andfurther TLC (Fig. 3) revealed several GM3-like species. A sample of the purified gly-colipid was subjected sequentially to enzy-matic hydrolysis by neuraminidase and0-galactosidase. A thin-layer separation ofthe hydrolysis products yielded lactosylcer-amide and glucosylceramide, indicating thatthe glycolipid is a GM3 ganglioside (Fig. 4).
The TLC patterns of infected lymph nodeand sciatic nerve showed not only increasedGM3 compared to uninfected control tis-sues but also a concomitant increase in aganglioside band similar in mobility to theGM1 or GM2 standard. Several other un-identified, faster-moving bands were alsopresent in the sciatic nerve. In the liver therewas an apparent reduction of two ganglio-side bands—one similar in mobility to GM2

54, 2^ Harris, et al.: Ganglioside Patterns^ 291
Std 1 2 Std 3FIG. 2. Thin-layer chromatographic patterns in lymph node and liver. Standard: a = GM I, b = GM2, c =
GM3; 1 = lymph node, normal armadillo; 2 = lymph node, lepromatous armadillo; 3 = liver, normal armadillo;4 = liver, lepromatous armadillo; * = GM3 ganglioside in the affected tissues.
and another band somewhat slower thanGM 1. Ganglioside concentrations andspecies distribution normally vary consid-erably among tissues and this variation
might account for a number of the differ-ences observed. The ganglioside patternsmay also be influenced by the degree of M.leprae infection of the tissues. Acid-fast
C 4111ba 41*
Mit
4■•■■•=11•114^■•■■•■■■•10^61■■•■••^
111110■11■14^Lao■11
I II 64 68 74 80 III IVFIG. 3. Thin-layer chromatogram of GM3 ganglioside after column purification (described in text). I =
standard mixture: a = GM I, b = GM2, c = GM3; II = before DEAE and Bio-Sil A chromatography; III = afterDEAE, before Bio-Sil A; IV = standard mixture. Fractions 64, 68, 74, and 80 represent species of GM3 afterBio-Sil A chromatography as differentiated by their mobility on thin-layer plate.

292^ International Journal of Leprosy^ 1986
C
b
411, 411400
at► GNI
1 2 3 4 5 6 7 8 9 10 11 12 13 14FIG. 4. Thin-layer chromatogram showing identification of GM3 ganglioside by enzymatic hydrolysis of
column eluates. a = GM3, b = lactosylceramide, c = glucosylceramide. Columns 1 and 14 are standard gan-glioside mixtures. 2 = F68; 3 = F68 + neu; 4 = F68 + neu + i3-gal; 5 = F74; 6 = F74 + neu; 7 = F74 + neu +(3-gal; 8 = F80; 9 = F80 + neu; 10 = F80 + neu + 13-gal; 11 = standard, GM3; 12 = GM3 + neu; 13 = GM3 +neu + 0-gal (neu = neuramidinase; (3-gal = 0-galactosidase; F = fraction number).
stains revealed numerous bacilli in thelymph node and nerve, while the liver con-tained comparatively fewer organisms.
These findings suggest that host cell me-tabolism was altered by the leprosy bacilliand, consequently, the constituent glyco-sphingolipids of the tissues were changed.(GM3 Gangliosides were not detected inlipids extracted in an identical fashion frompurified M. leprae.) Changes in the ganglio-side composition of cells are known to occuras a result of external stimuli. For example,cultured HeLa cells exposed to the short-chain, fatty acid butyrate show an increasedbiosynthesis of GM3 ganglioside as a resultof induced lactosylceramide sialytransfer-ase activity in the cells ( 2).
Several mechanisms could account for thedifferences in ganglioside patterns betweenlepromatous and normal armadillo tissues.The specific glycosyltransferases responsi-ble for ganglioside synthesis may be inducedby the presence of M. leprae in the tissue,thereby resulting in increased gangliosidebiosynthesis. Cellular infiltrates into the tis-
sue (macrophages, lymphocytes) may ex-press these enzymes as they interact withM. leprae, thereby increasing their ganglio-side synthesis. More simply, the inherentganglioside patterns of the infiltrating cellscould be reflected in altered ganglioside pat-terns of the infected tissues. Finally, thechanges observed may merely be secondaryto tissue degeneration or inflammation as-sociated with infection with M. leprae.
From these limited observations it wouldbe premature to draw any conclusions con-cerning the significance of altered ganglio-side concentrations in lepromatous arma-dillo tissue. However, their association withneurological dysfunction, involvement asspecific, surface membrane receptors, andtheir antigenic properties provide a reason-able basis for continued interest in thesecompounds in leprosy.
SUMMARYGangliosides derived from tissues of nor-
mal and lepromatous armadillos were ex-amined by thin-layer chromatography. The

54, 2^Harris, et al.: Ganglioside Patterns^ 293
ganglioside patterns produced by the My-cobacterium leprae-infected tissues variedfrom that of the normal tissue. Althoughincreased levels of several gangliosides wereobserved in the infected tissues, there wasan apparent preferential increase in GM3gangliosides as determined by column chro-matography and enzymatic hydrolysis.
RESUMENUsando la cromatografia en capa fina, se examinaron
los gangliOsidos derivados de los tejidos de armadillosnormales y los derivados de armadillos lepromatosos.
El patron de los gangliOsidos producidos por los te-jidos infectados con Mycobacterium leprae fue dife-rente del producido por los armadillos normales. Aun-quc varios gangliOsidos derivados de los tejidosinfectados mostraron un incremento en su concentra-ciOn, los gangliOsidos GM3 mostraron un incrementoaparentemente preferencial, segán se determine, porcromatografia en columna c hidrOlisis enzimatica.
RÉSUMÉ
On a examine, par une methode de chromatographieen couches minces, les gangliosides extraits de tissusrecueillis chez des tatous normaux et chez des animauxatteints de lepre. Le profil des gangliosides produitspar des tissues infectes par Mycobacterium leprae etaientdifferents de ceux observes dans les tissus normaux.On a observe une augmentation des taux de plusieursgangliosides dans les tissus infectes. II est toutefois ap-paru que cette augmentation semblait porter davantagesur les gangliosides GM3, tels qu'on peut les mesurerpar la chromatographie sur colonne et par l'hydrolyseenzymatique.
Acknowledgments. The authors wish to express theirappreciation to Miss Lynn Larsen for her expert tech-nical assistance. We are also grateful to Mrs. Mary M.Jackson for secretarial help and the assistance of Mr.Gregory T. McCormick. This study was supported inpart by Grants NS09626 from the National Institutesof Health, PCM 82-19489 from the National SciencesFoundation, and Interagency Agreement no. YO I -AI-50001-00 of the National Institute of Allergy and In-fectious Diseases. Y.-T. Li is a recipient of the JavitsNeuroscience Investigator Award.
REFERENCES1. CHIEN, J.-L., Li, S.-C., LAINE, R. A. and Li, Y.-T.
Characterization of gangliosides from bovine
erythrocyte membranes. J. Biol. Chem. 253 (1978)4031-4035.
2. FISHMAN, P. H., SIMMONS, J. L., BRADY, R. 0. andFREESE, E. Induction of glycolipid biosynthesis bysodium butyrate in HeLa cells. Biochem. Biophys.Res. Commun. 59 (1974) 292-299.
3. FOLCH, J., LEES, M. and SLOANE-STANLEY, G. H.A simple method for the isolation and purificationof total lipids from animal tissues. J. Biol. Chem.226 (1957) 497-509.
4. HAKOMORI, S. Glycosphingolipids in cellular in-teraction, differentiation and oncogenesis. In: An-nual Review of Biochemistry. Snell, E., ed. PaloAlto, California: Annual Reviews, Inc., 1981, pp.733-764.
5. JOB, C. K. Pathology of peripheral nerve lesionsin lepromatous leprosy—a light and electron mi-croscopic study. Int. J. Lepr. 39 (1971) 251-268.
6. KIRCHHEIMER, W. F. and STORKS, E. E. Leprosyin experimentally infected armadillos. Int. J. Lepr.40 (1972) 212-213.
7. LEHNINGER, A. L. The neuronal membrane. Proc.Natl. Acad. Sci. U.S.A. 60 (1968) 1069-1080.
8. Li, Y.-T. and Li, S.-C. Utilization of glycosidasesfor the structural studies of complex carbohydratechains. In: CNRS International Symposium of theStructure and Methodology of Glycoconjugates.Montreil, J., ed. Paris: Editions du Centre Na-tional de la Recherche Scientifique, 1973, vol. 1,pp. 339-350.
9. MANSSON, J.-E., HOLMGREN, J., Li, Y.-T., VANIER,
M.-T. and SVENNERHOLM, L. Chemical and im-munological characterization of the major gluco-samine-containing ganglioside of human tissues.Med. Biol. 52 (1974) 240-243.
10. MoMol, T., ANDO, S. and MAGAI, Y. High reso-lution preparative column chromatographic sys-tem for gangliosides using DEAE-Sephadex and anew porus silica latrobeads. Biochim. Biophys.Acta 441 (1976) 448-497.
11. PRABHAKARAN, K., HARRIS, E. B. and KIRCHHEI-
MER, W. F. Hypopigmentation of skin lesions inleprosy and occurrence of o-diphenoloxidase inMycobacterium leprae. In: Pigment Cell. Riley,V., ed. Basel: S. Karger, 1976, pp. 152-164.
12. ROUSER, G., KRITCHEVSKY, G., HELLER, D. andLIEBER, E. Lipid composition of beef brain, beefliver and the sea anemone: two approaches toquantitative fractionation of complex lipid mix-tures. J. Am. Oil Chem. Soc. 40 (1963) 425-454.
13. SVENNERHOLM, L. Quantitative estimation of sialicacids. II. A colorimetric resorcinol-hydrochloricacid method. Biochim. Biophys. Acta 24 (1957)604-611.