grehan, 2012
TRANSCRIPT

Bulletin of the Buffalo Society of Natural Sciences 42, 2012 33
MORPHOLOGICAL EVIDENCE FOR PHYLOGENETIC RELATIONSHI PS WITHIN THE HEPIALIDAE (LEPIDOPTERA: EXOPORIA)
John R. Grehan
Research Associate, Section of Invertebrate Zoology, Carnegie Museum of Natural History Email [email protected]
ABSTRACT - Of the 70 currently recognized genera of Hepialidae, 58 were examined to test earlier evidence (Grehan, 2010) for a Latin American (tergal lobe) clade comprising Druceiella and three other genera, and to identify character states that may be informative of broader generic relationships within the Hepialidae. Thirty six characters were considered in this study and the relationships were analyzed between 45 genera where informative character states were shared between two or more genera. Derived states were defined with respect to an outgroup comprising other Hepialoidea and the Mnesarchoidea. Character state analysis (PAUP) showed significant bootstrap (78%) support for the genera Afrotheora, Antihepialus, Fraus, Gazoryctra (‘primitive Hepialidae’ of authors) as the sistergroup for the remaining six clades (74%) of which the tergal lobe clade was corroborated (75%). The ‘oxycanine’ genera of Australasia were grouped together, but with low bootstrap support (52%) and several characters were involved in clustering four mostly European genera together, but without significant bootstrap support. The lack of resolution for these and other genera is attributed to the lack of character states identified in the current survey. These results are therefore considered provisional and contingent upon future exhaustive survey of morphological features in the Hepialidae and especially for the outgroup whose members are poorly documented at present for many features. The morphological evidence presented here is considered to represent independent genetic (heritable) evidence to that of DNA sequence similarity and therefore not necessarily falsifiable by any future incongruent patterns of molecular sequence similarity.
INTRODUCTION The biology of ghost moths (Hepialidae) is poorly known for most species and their phylogenetic relationships are even less understood despite having a nearly global distribution range and including some major economic pests in forestry and agriculture (Grehan, 1989; Tobi & Grehan, 1993). The 70 genera and over 500 species (Nielsen et al., 2000; Mielke & Grehan, 2012) make the Hepialidae the largest family in the Hepialoidea whose remaining members are represented by only four families with seven genera for a total of 21 species. These families, together with the Mnesarchaeoidea with a single genus (Mnesarchaea, Meyrick, 1886) and eight species comprise the suborder Exoporia. The Exoporia is the sistergroup to the suborder Ditrysia with about 157,500 species (Van Nieukerken et al., 2011) and most of the generally better known moth and butterfly groups.
Earlier comprehensive analysis of phylogenetic relationships between hepialid genera is limited to faunal studies for New Zealand (Dugdale, 2004) and southern South
America (Nielsen and Kristensen, 1984), and the generic revisions of Fraus (Nielsen & Kristensen, 1989), Bipectilis (Nielsen, 1988), Afrotheora (Nielsen & Scoble, 1988) and Phymatopus (Wagner, 1985). A revision of Australian genera is currently underway (Thomas Simonsen, personal communication) and a revised taxonomic checklist of the Latin American ghost moth fauna was recently completed (Mielke & Grehan (2012) as a prelude to future phylogenetic evaluation of ghost moths in this region.
The first principal effort to define a major
phylogenetic division within the Hepialidae was made by Tindale (1941: IV) who proposed the subfamily Zenophassinae to accommodate the monotypic genus Zenophassus Tindale, 1941. This taxonomic separation was based on a distinctive combination of primitive features in Zenophassus schamyl (Christof, 1888). The subfamily did not gain general acceptance and was later considered by Nielsen & Robinson (1983) to be invalid because the characters did not support such a primary division in the Hepialidae. A subfamily division proposed by

Bulletin of the Buffalo Society of Natural Sciences 42, 2012 34
Dumbleton (1966) distinguished between ‘Oxycaninae’ of Australasia where the wing vein R4 branches directly from the stem of R2 & R3, and the remaining ‘Hepialinae’ where R4 has a common stem with R5. Nielsen & Robinson (1983) regarded this distinction to be problematic because the oxycanine venation was also present in two South American genera that showed no other evidence of a close relationship with the Australasian genera (see also Dugdale, 1994).
The first explicitly phylogenetic approaches
for the Hepialidae (Nielsen, 1988; Nielsen& Kristensen, 1989; Nielsen & Scoble 1988) pointed out that the monophyly of the Hepialidae could not be upheld unless four ‘primitive’ genera (Afrotheora, Bipectilis, Fraus, Gazoryctra) were removed. The monophyly of the remaining Hepialidae sensu stricto appeared to be supported by at least three features (Appendix 1). Supporting evidence for the monophyly of the Hepialidae sensu lato remains unresolved.
Two subsequent studies of generic
relationships only considered limited groups of genera. Wagner & Rosovsky (1991) considered the relationships of ten genera with respect to16 characters, but the character states were not substantiated with respect to their presence or absence within the Hepialidae or any outgroup (Appendix 2). A cladistic study of the inter- and intra-generic relationships of oxycanine genera in New Zealand by Brown et al. (2000) limited their outgroup to a single local species of Aenetus and three species of Aoraia, but without substantiating the character state distribution for other genera and a hepialid outgroup which would be necessary for identifying derived character states for relationships among hepialid genera (Appendix 3).
The phylogenetic review presented here is
part of an ongoing taxonomic study of the Central-South American genus Druceiella. In order to identify characters supporting systematic relationships within the genus and corroborate its monophyly, it was desirable to compare this genus with every other genus of the Hepialidae (possible at least for genitalic features). To establish the phylogenetic relationships between Druceiella and other
genera it was necessary to explore the generic relationships of all genera within the Hepialidae and that in turn required, at the very least, comparisons with immediate outgroup taxa (other Exoporia).
This goal of this morphological study is to
begin to address the two major limitations of previous studies: (1) explicit designation of an outgroup, (2) limit informative character states to those that are only present within the Hepialidae sensu lato (i.e. the Hepialidae as currently recognized), and (3) document the presence or absence of informative character states for almost all hepialid genera (not examined at this time were Hepialiscus, Parahepialiscus, Neohepialiscus, and Palpifer). Morphological and molecular evidence A purely morphological analysis may seem rather quaint and pointless against the many, and technologically sophisticated, methods now available for analyzing thousands of DNA sequence similarities. According to most evolutionary biologists, the sheer number of similarities in DNA sequence comparisons is sufficient cause to characterize molecular results as both necessary and sufficient for phylogenetic reconstruction. Under this viewpoint morphology is relegated to a mere shadow that is only phylogenetically meaningful when it corresponds to molecular results. Other approaches regard morphological and molecular characters as being cladistically equivalent and that combining the data sets into a ‘total evidence’ result is a valid approach.
The molecular assumption has its most prominent application to the proposition that humans and chimpanzees are most closely related to each other because they share more DNA sequence similarities in common that with any other primate. This relationship has been elevated to the statues of a scientific fact even though there is almost nothing in morphology that is uniquely shared in common between humans and chimpanzees whereas at least 28 unique or specialized features are shared between humans and orangutans. This genetic similarity between humans and orangutans extends to the fossil record where early hominids look more like orangutans than

Bulletin of the Buffalo Society of Natural Sciences 42, 2012 35
chimpanzees (Schwartz, 1987, 2004, Grehan & Schwartz, 2009).
Even though arguments for the superiority of
molecular evidence have appealed to the large number of DNA base pair comparisons involved (the law of large numbers), molecular sequence similarities, by their very nature, cannot provide cladistic character states as in morphology where it is possible to restrict the data set to be analyzed to only those characters that demonstrate a derived condition within the ingroup. Molecular sequence data represents a combination of primitive and derived states that cannot be individually recognized as such. Molecular approaches rely on the resulting tree to generate those states rather than analysis being used to sort out the best set of corroborating derived character states in morphology. Essentially, molecular approaches are phenetic.
Combining morphological and molecular patterns of similarity (see Simonsen et al., 2012 for Lepidoptera) as ‘total evidence’ does not necessarily render a more accurate result than morphology alone since it represents a combination of cladistic and phenetic characters. The ‘total evidence’ approach assumes that morphological and molecular sequence characters are cladistically equivalent even though it is only in morphology that the derived state of each individual character can be independently proposed and the data set therefore be restricted to hypothesized apomorphies.
There are many such problems in molecular
methods where the use of complex statistical modeling serves to emphasize that molecular approaches are more about overall similarity that may obscure actual patterns of relationship (Mooi & Gill, 2010; Ebach, 2011; Livezey, 2011; Schwartz, 2005, 2012; Schwartz & Maresca, 2007). Supporters of molecular hegemony will quite happily contradict their position when using fossils where morphology becomes transformed into reliable evidence (Schwartz, 2004).
Molecular results have generated many phylogenetic results that are biogeographically cohesive and informative (Heads, 2012) and
more often than not molecular results may be reliable - but this is only with the caveat that this is no less true of morphology and that molecular results do not automatically falsify contrasting morphological results (or vice versa).
The current analysis is based on the
assumption that comparative morphology represents an independent and valid science and that there is no empirical evidence to substantiate the claims that molecular evidence is necessarily more reliable or cladistically equivalent to morphological evidence. The relationships proposed here are subject to future morphological corroboration or falsification, but not to whether or not they will correspond to some future molecular sequence pattern. Morphological characters Finding uniquely shared features between hepialid genera and having sufficient sampling of both outgroup and ingroup taxa represent two of the major changes for this study. At the very least, all species of Hepialidae and other Exoporia should be examined and documented for every character under consideration. At this time this goal is not possible and the characters proposed here may be problematic depending on the scope of comparison.
For some genera, such as in New Zealand and southern South America, there is extensive information for all species of each endemic genus, particularly for the genitalia. Genitalia and wing venation are among the most widely documented and illustrated (and therefore potentially relatable) features in both the Hepialidae and the outgroup taxa. Taxonomic groups The Hepialidae is the ingroup, but in the absence of one or more defining features being generally recognized for this family, the Hepialidae is treated here as an unconfirmed, but de facto monophyletic entity (but see note below on the second abdominal sternite). Most taxonomic treatments of the Hepialidae comprise species and generic descriptions of varying quality and detail.
Most of the 70 recognized genera (Nielsen et al., 2000; Mielke & Grehan, 2012) are poorly defined and lack one or more defining features

Bulletin of the Buffalo Society of Natural Sciences 42, 2012 36
to support their monophyly. This is particularly problematic for broadly distributed and species-rich genera such as Endoclita and Thitarodes. The current analysis is, therefore, applicable only to the extent that the member species of each genus are sampled or documented. Phassus guienensis is referred to by its current name, but is incertae sedis (Mielke & Grehan, 2012) because it lacks features to support it as a member of the genus Phassus. Korscheltellus is defined here to include three species as characterized by Grehan (2012).
The immediate reference outgroup comprises the four other families of Hepialoidea (Anomosetidae, Neotheoridae, Prototheoridae, Palaeosetidae) and the Mnesarchaeidae (Mnesarchaeoidea).
With only eight genera and 29 species this outgroup provides a relatively narrow range of comparison to identify shared derived features within the Hepialidae. Inclusion of the exoporian sistergroup Ditrysia would be desirable, but its shear size makes detailed comparison impractical for this study. Many ditrysian features such as genitalia are sufficiently well documented to be able to recognize hepialid character states as distinct. The Hepialidae is treated here as a monophyletic group even though there is at this time no unequivocal character or set of characters to support that assumption.
. METHODS
Sixty two genera were available for comparison, but phylogenetic analysis was applied to only 47 genera where two or more genera shared one or
more shared derived character states (Appendix 4). Characters with missing values were not included. Relationships were analyzed with a heuristic search of 36 characters using PAUP® 4.0b10 (Swofford, 2005) for 200 trees and assuming both the ingroup and outgroup to be monophyletic. Character states were coded as absent (0) or present (1), with outgroup character states coded (0) for all characters. Only unequivocal derived character states were used for the analysis by excluding characters that were variable for the outgroup. Dissections by the author are referred to by the initials ‘JRG’ followed by the dissection number.
The following character list illustrates all character states, but space limitations preclude illustrations for all species. The primary emphasis is to illustrate the derived state for some or all of the represented taxa. Specimens imaged are from the following collections: AMNH – American Museum of Natural History; BF – Bernard Francois collection; CMNH – Carnegie Museum of Natural History; CNC – Canadian National Collection; CUIC – Cornell University Insect Collection; RMBH – R.M. Bohart Museum, University of Los Angeles, Davis; JMM – John Michael Maes collection; JRG – John R. Grehan collection; LACM – Los Angeles County Museum; NZAC - New Zealand Arthropod Collection; PMNH – Peabody Museum of Natural History. Character descriptions Examples of derived character state are illustrated except for tergal lobe, wing vein and 8th abdominal sternite variation characters that are reviewed for the family as a whole.
Character 1. Pseudotegumenal arms and valves with transverse ridges All other genera and outgroup taxa have smooth surfaces in the same position on the valves.
Cibyra dorita Schaus, 1901 [JRG: M89]
Aepytus guarani (Pfitzner, 1914) [JRG: M180]
Cibyra ferruginosa Walker, 1856 [M154: JRG]
Cibyra stigmatica (Pfitzner, 1937) [JRG: M141]
Bulletin of the Buffalo Society of Natural Sciences 42, 2012 37
Character 2. Elongate intermediate plate extends to anterior edge of pseudotegumen This feature needs to be better defined in future treatments. IP: intermediate plate.
Hepialyxodes rileyi Viette, 1951 [NMNH: M184]
Cibyra stigmatica Pfitzner, 1937) [JRG: M141]
Schaefferiana epigramma (Herrich-Schäffer, [1854]) [JRG: M183]
Aepytus guarani (Pfitzner, 1914) [JRG: M180]
Character 3. Tergosternal bar triangular . Refer to Grehan (2010). Only Mnsearchaea and Prototheora examined for other Exoporia. Character 4. Tergal lobe with posterior-lateral knob. As for character 3 (Grehan, 2010). Character 5. Anterio-dorsal margin of tergal lobe with broken inlet . As for character 3 (Grehan, 2010). Character 6. Anteriorly extended anterio-lateral peudotegumen and intermediate plate. The anterior margin (blue arrow) of the lateral pseudotegumen and intermediate plate is anterior to the medial fusion of the pseudotegumen.
Druceiella sp. [RMBM: JRG M96]
Trichophassus giganteus (Herrich-Schäffer, [1853]) [NZAC: JSD593]
Phassus guianensis Schaus, 1940 [AMNH: M84]
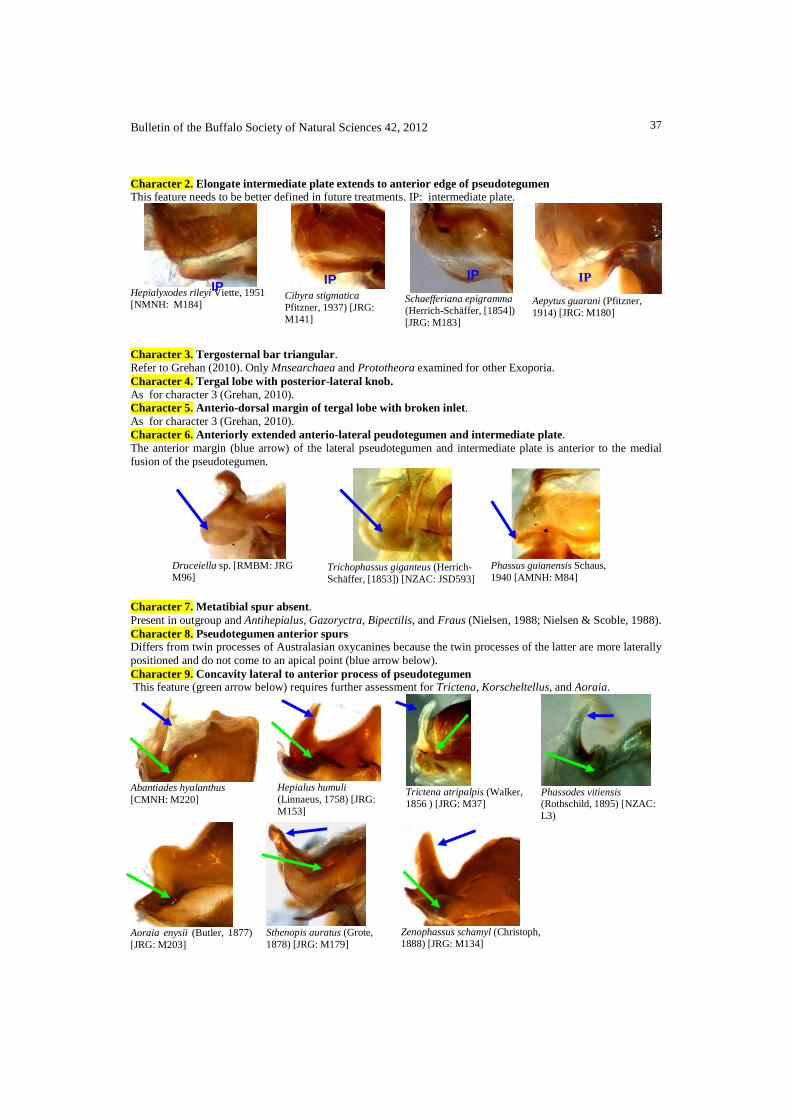
Character 7. Metatibial spur absent. Present in outgroup and Antihepialus, Gazoryctra, Bipectilis, and Fraus (Nielsen, 1988; Nielsen & Scoble, 1988). Character 8. Pseudotegumen anterior spurs Differs from twin processes of Australasian oxycanines because the twin processes of the latter are more laterally positioned and do not come to an apical point (blue arrow below). Character 9. Concavity lateral to anterior process of pseudotegumen This feature (green arrow below) requires further assessment for Trictena, Korscheltellus, and Aoraia.
Abantiades hyalanthus [CMNH: M220]
Hepialus humuli (Linnaeus, 1758) [JRG: M153]
Trictena atripalpis (Walker, 1856 ) [JRG: M37]
Phassodes vitiensis (Rothschild, 1895) [NZAC: L3)
Aoraia enysii (Butler, 1877) [JRG: M203]
Sthenopis auratus (Grote, 1878) [JRG: M179]
Zenophassus schamyl (Christoph, 1888) [JRG: M134]
IP IP IP IP

Bulletin of the Buffalo Society of Natural Sciences 42, 2012 38
Character 10. Trulleum apex a triangular projection The apex of the trulleum has also been characterized as free of the pseudotegumen (Nielsen, 1988; Nielsen & Kristensen, 1988), but what constitutes a ‘free’ condition has not been specified. In all these cases the apex of the trulleum projects away from the pseudotegumen with a triangular apex.
Gazoryctra sciophanes (Ferguson, 1979) [JRG: M313]
Antihepialus antarcticus (Wallengren, 1860) [CNC: M132 ]
Afrotheora jordani (Viette, 1956), (BMNH). Trulleum as a double bar
Character 11. Lateral pseudoteguminal lobes sclerotized from origin with intermediate plate Variable in morphology. Requires further analysis.
Hampsoniella sp., [LACM: M221]
Gymelloxes terea (Schaus, 1892), [JMM: M155]
Pseudodalaca serta (Schaus, 1894), M199 (PMNH)
Pseudophassus mahagoniatus Pfitzner, 1914 [JRG: M157]
Trichophassus giganteus (Herrich-Schäffer, [1853]), JSD593 (NZAC)
Character 12. Tegumenal apex articulates with scleritozied posterior vincular margin This feature is highly variable in detail. In Druceiella and Gymelloxes there are two lateral sclerotized knobs along the vincular margin where as Pfitzneriana has a single knob and Phassus guianensis has an extended sclerotized margin whereas sclerotization is minimal in the remaining genera.
Alloaepytus tesselloides (Schaus, 1901) [CMNH: M182
Gymelloxes terea (Schaus, 1892) [JMM: M155]
Pfitzneriana sp. [AMNH: M167]
Phassus guianensis Schaus, 1940 [AMNH: M84]
Pseudophassus mahagoniatus Pfitzner, 1914 [JRG: M157]
Roseala tessellatus (Herrich-Schäffer, [1854]) [JRG: M187]
Druceiella n. sp. [JRG: M145]
Bulletin of the Buffalo Society of Natural Sciences 42, 2012 39
Character 13. Apex of phallus sclerotized. Below for Vietteogorgopis and Tricladia (blue arrow) Character 14. Valve with spine shaped apex and setose basal lobe Basal lobe not very clear in these images, but indicated by red arrow.
Vietteogorgopis sp. [JRG: M164]
Tricladia prytanes (Schaus, 1892),[AMNH: M197]
Trichophassus giganteus Le Cerf, 1919 [NZAC: JSD593]
Pseudophassus mahagoniatus Pfitzner, 1914 [JRG: M157]
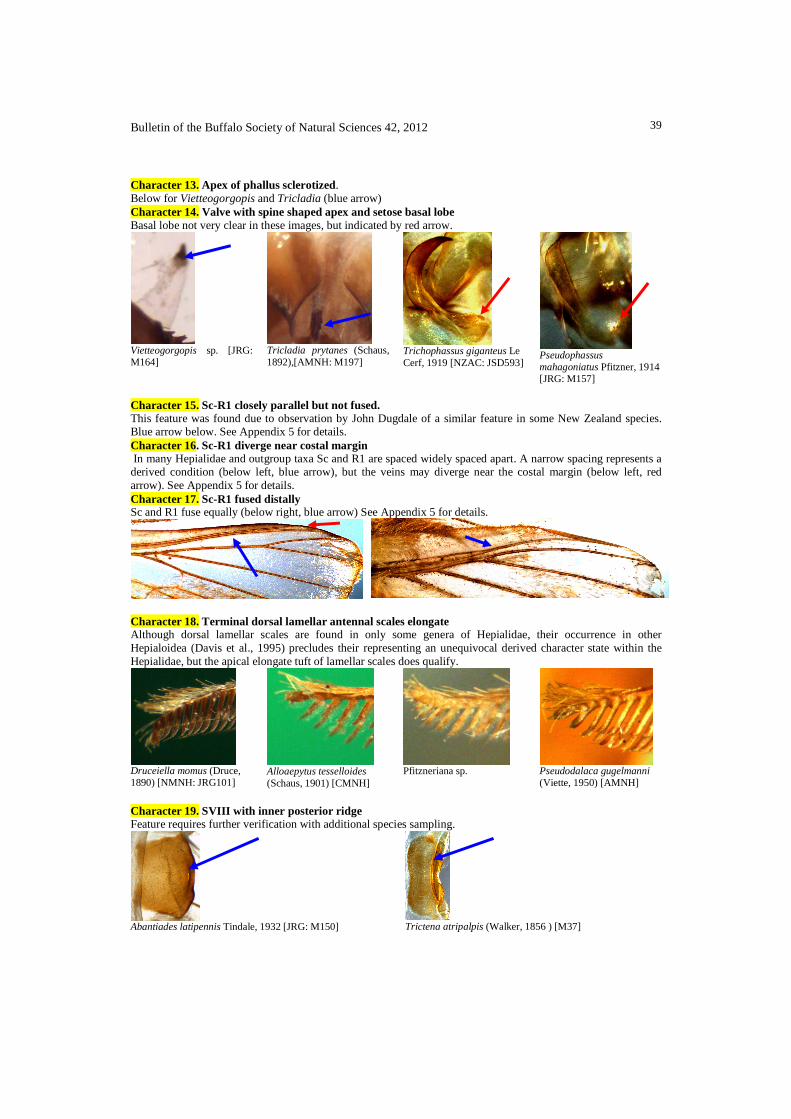
Character 15. Sc-R1 closely parallel but not fused. This feature was found due to observation by John Dugdale of a similar feature in some New Zealand species. Blue arrow below. See Appendix 5 for details. Character 16. Sc-R1 diverge near costal margin In many Hepialidae and outgroup taxa Sc and R1 are spaced widely spaced apart. A narrow spacing represents a derived condition (below left, blue arrow), but the veins may diverge near the costal margin (below left, red arrow). See Appendix 5 for details. Character 17. Sc-R1 fused distally Sc and R1 fuse equally (below right, blue arrow) See Appendix 5 for details.
Character 18. Terminal dorsal lamellar antennal scales elongate Although dorsal lamellar scales are found in only some genera of Hepialidae, their occurrence in other Hepialoidea (Davis et al., 1995) precludes their representing an unequivocal derived character state within the Hepialidae, but the apical elongate tuft of lamellar scales does qualify.
Druceiella momus (Druce, 1890) [NMNH: JRG101]
Alloaepytus tesselloides (Schaus, 1901) [CMNH]
Pfitzneriana sp.
Pseudodalaca gugelmanni (Viette, 1950) [AMNH]
Character 19. SVIII with inner posterior ridge Feature requires further verification with additional species sampling.
Abantiades latipennis Tindale, 1932 [JRG: M150]
Trictena atripalpis (Walker, 1856 ) [M37]

Bulletin of the Buffalo Society of Natural Sciences 42, 2012 40
Character 20. SVIII reduced to lateral posterior margin Posterior margin of SVIII entire for outgroup and other Hepialidae.
Pfitzneriana sp. [AMNH: M167]
Pseudophassus mahagoniatus Pfitzner, 1914 [JRG: M157]
Tricladia prytanes (Schaus, 1892) [AMNH: M197]
Vietteogorgopis sp. [JRG M164]
Character 21. Hindleg tarsi missing) See Wagner (1985) Character 22 Hindleg tibia swollen See Wagner (1985) Character 23. Lightly sclerotized region (ls) between tergosternal bar (tb) and posterior tergal lobe (ptl) See Grehan, 2010 for detailed comparisons for Hepialidae
Dumbletonius characterifera (Walker, 1865) [JRG: M173]
Oxycanus dirempta (Walker, 1865) [CMNH: M219]
Wiseana sp. [JRG: M175]
Character 24. Twin process (tp) of the anterior pseudogegumen See also figures in Dugdale (1994).
Oxycanus sp. [CNC: M224]
Dioxycanus fusca (Philpott, 1914) [JRG: M172]
Character 25. TIII with lateral bar (blue arrow) A lateral bar is present in TII and TII in Antihepialus and Afrotheora (arrows). In all other Hepialidae and outgroups a lateral bar is present only on TII as illustrated here for Zenophassus schamyl. Character proposed by Nielsen & Scoble (1986) and corroborated in this study.
Afrotheora jordani (BMNH)
Antihepialus antarcticus M132
Zenophassus schamyl (Christoph, 1888) (SII bar missing) [JRG: M134]
ptl
tb
ptl
tb
ls ls ls
tp tp

Bulletin of the Buffalo Society of Natural Sciences 42, 2012 41
Character 26. Valve bilobate Valves with a laterally projecting basal lobe. Size and position in Gazoryctra appear to be unique. Bilobate condition in some Fraus (cf. Nielsen & Kristensen, 1989) has a different configuration.
Gazoryctra rosicaput (Ferguson, 1979) [nzac: JSD603]
Gazoryctra sciophanes (Ferguson, 1979) [JRG: M313]
Antihepialus antarcticus (Wallengren, 1860) [CNC: M132]
Character 27. Broad, shallow ‘V’ shaped anterior SII margin With the exception of ‘primitive’ Hepialidae – Afrotheora, Antihepialus, Fraus, Gazoryctra – see character 28) the anterior margin of SII forms a shallow V as the lateral bar (LB) is oriented, to a greater or lesser degree, laterally. In some genera such as Korscheltellus and Triodia (see character 30 below) this character state may not be applicable, and further assessment and additional taxa need to be sampled - especially for the non-hepialid Exoporia. SII in the outgroup genus Prototheora (Hepialoidea) appears to lack a distinctly sclerotized anterior margin and the lateral bar is narrow and forwardly oriented. See Appendix 6 for generic comparisons.
Prototheora sp. [PMNH: M199] Outgroup
Druceiella songoensis [NMNH: M198]
Andeabatis chilensis (Ureta, 1951) [M211, NMNH]
Character 28. Broad, anteriorly oriented SII lateral bar The lateral SII bar extends directly forward rather than also laterally as in other Hepialidae. There was insufficient detail for Bipectilis by Nielsen (1988: Fig. 38) for characterizing this feature.
Afrotheora jordani (BMNH) Narrow, U-shaped anterior margin
Antihepialus antarcticus M132 (CNC) Narrow, U-shaped anterior margin
Fraus simulans Walker, 1856, F31 (JRG) Narrow, U-shaped anterior margin
Gazoryctra sciophanes (Ferguson,1979) [JRG: M131]
LB LB

Bulletin of the Buffalo Society of Natural Sciences 42, 2012 42
Character 29 Tegumen apex spout-like This projection may involve the trulleum in some genera (Grehan, 2012).
Hepialus humuli (Linnaeus, 1758 ) [JRG: M153]
Pharmacis aemilianus (Constantini, 1911) [JRG: M45]
Korscheltellus gracilis (Grote, [1865]) [JRG: M147]
Triodia sylvina (Linnaeus, 1761) [JRG: M130]
Zenophassus schamyl (Christoph, 1888 ) [JRG: M134]
Character 30 SII lateral ridge angled posterioro-medially Most Hepialidae lack a lateral ridge on SII. A ridge is found in Prototheora and other Lepidoptera and may represent the primitive condition. The medially oriented posterior ridge in Korscheltellus and Triodia appears distinctive in its concave shape (cf some Aenetus species, Appendix 6).
Korscheltellus gracilis (Grote, [1865 ]) [JRG: M196]
Triodia sylvina (Linnaeus, 1761) [JRG: M130]
Character 31. Anterior pseudotegumen with medial sclerotized ridge
Korscheltellus gracilis (Grote, [1865]) [JRG: M196]
Korscheltellus lupulina (Linnaeus, 1758) [JRG: M230]
Triodia sylvina (Linnaeus, 1761) [JRG: M130]
Character 32 Anterior vinculum with U shaped anterior margin
Hepialus humuli (Linnaeus, 1758) [JRG: M153]
Pharmacis aemilianus (Constantini, 1911) [JRG: M45]
Pharmacis bertrandi (Le Cerf, 1936) [JRG: M156]
Triodia sylvina (Linnaeus, 1761) [JRG: M158]
Zenophassus schamyl (Christoph, 1888) [JRG: M134]

Bulletin of the Buffalo Society of Natural Sciences 42, 2012 43
Character 33. Anterior vincular margin forms lateral pocket
Triodia sylvina (Linnaeus, 1761 ) M158 (JRG)
Zenophassus schamyl (Christoph, 1888) M134 (JRG)
Character 34. Wing vein R4 branches directly from stem of R2 & R3 ‘Oxycanine” pattern. Included in the present study are Aepytus, Cladoxycanus, Dioxycanus, Dumbletonius, Heloxycanus, Jeana, Elhamma, Tricladia, and Vietteogorgopis. Also applicable to Napialus, Hepialiscus, and Parahepialiscus (Ueda, 1988; Dugdale, 1994; Grehan, 2011). Character 35. Pseudotegumen apex fused and sclerotized
Druceiella amazonensis, Viette, 1950 [BF: M192]
Phassus guienensis Schaus, 1940 [AMNH: M84]
Andeabatis chilensis (Ureta, 1951) [NMNH: M212]
Parapielus luteicornis (Berg, 1882) (JRG: M142]
Character 36. Pseudotegumen with projecting valvaller (v) and pseudoteguminal apex (pa) In these lateral views the pseudoteguminal apex is sometimes partially obscured by the valve.
Dioxycanus fusca (Philpott, 1914), M227 (JRG)
Hepialus humuli (Linnaeus, 1758 ), M153 (JRG)
Korscheltellus gracilis (Grote, [1865 ]), [JRG: M196]
v v
pa pa pa
v

Bulletin of the Buffalo Society of Natural Sciences 42, 2012 44
Phassodes vitiensis (Rothschild, 1895) [NZAC: L35]
Endoclita excresens (Butler, 1877), (NZAC: JSD606)
Zenophassus schamyl (Christoph, 1888), [JRG: M134]
Triodia sylvina (Linnaeus, 1761) [JRG: M130]
Wiseana sp. [ JRG: M175]
RESULTS
Parsimony analysis resulted in 69 trees and a strict consensus tree comprising four principal clades that are named here with respect to a characteristic morphological or geographic feature (Fig. 1). These names are intended only for descriptive reference and do not imply any necessary taxonomic rank or certainty. Bootstrap values over 50% are included in the consensus tree. The consensus tree separates the ‘primitive Hepialidae of Neilsen et al (2000) comprising Afrotheora, Antihepialus, Fraus, and Gazoryctra. While this group has significant bootstrap support, the number of characters involve are few and the arrangement must still be regarded as tentative. 1. Primitive Hepialidae: Supported by a bootstrap value of 78.The genera Gazoryctra, Fraus, Antihepialus, and Afrotheora were identified as ‘primitive’ or basal Hepialidae (Nielsen, 1988; Nielsen & Scoble, 1988; Nielsen & Kristensen, 1989) but their precise relationship with other Hepialidae was unresolved. The current analysis supports their monophyly although there is only one character state shared by all genera (the broad anteriorly directed sternal bar). The genus Bipectilis may also be a member of this clade, or represent another basal lineage because it retains the tibal spur (as in Gazoryctra, Fraus, and Antihepialus, but not Afrotheora), and the pseudotegumen of
Fig. 1. Consensus tree and bootstrap values for seven generic groups of Hepialidae. See text for discussion of numbered groups. Bipectilis has several elongate projections that may be homologous to those found in the other hepialoid families (Nielsen, 1988). There was no resolution of relationships between these genera in this analysis.
(2) Western Pacific clade. No bootstrap support. The Australian root feeding genera Abantiades and Trictena are supported as sister taxa (Fig. 2). The Fijian Phassodes may also be a root feeder since there are no reports of stem boring by this moth (although feeding on live or dead foliage on the ground cannot be ruled out).
2. W. pacific 1. primitive
5. W. Eurasia
4. Leto 3. tergal lobe
Outgroup
7. oxycanine
6. Andean
78
74
72 52
75
pa
v v
pa
v
v
pa
pa
v
pa

Bulletin of the Buffalo Society of Natural Sciences 42, 2012 45
Genera in this clade share the presence of a laterally projecting first abdominal tergal lobe and a closely parallel Sc-R1 venation similar to that of the cibyrine genera. They also share the presence of an anterio-dorsal process of the pseudotegumen that is also present in some members of the hepialinine clade (see clade 4). In the absence of a single character state to define this group its relationship with the cibyrines and hepialines will require further corroboration.
Fig. 2. Western Pacific clade (Abantiades and Trictena in Australia, and Phassodes in Fiji). No significant bootstrap support. (3) Tergal lobe clade Supported with a bootstrap value of 75 and including 13 of the 17 genera (as recognized by Mielke & Grehan, 2012) previously subsumed by Nielsen et al. (2000) under Cibyra (Fig. 3). The clade also includes Phassus guianensis for which the holotype has been examined and corresponds to the cibyrine taxa rather than other Phassus (incertae sedis by Mielke & Grehan, 2012).
The tergal lobe group appears to have robust
support with all members exhibiting a triangular tergosternal bar, a tergal lobe with lateral knob, and the anterior tergal lobe margin interrupted by an invagination extending laterally as a ‘T’ shaped inlet (Grehan, 2011). In addition, the second abdominal sternite is similar in all genera and may constitute an additional apomorphy. The tergal lobe clade represents an unresolved polytomy with respect to the Western Pacific clade and Leto, but without significant bootsrap support. It was previously noted (Grehan, 2010) that Callipielus also shares the presence of a lateral knob, but in the absence of other unique similarities this genus was not grouped with the tergal lobe clade (or any other non-primitive Hepialidae) in this analysis. (4) Leto clade. The monotypic Leto is currently anomalous with respect to the remaining African hepialid fauna with which it shows no obvious
close relationships. The larvae of Leto are stem borers whereas the other African hepialids are subterranean and feed on roots or surface foliage (Grehan, 1989). The larva of Leto is also the only hepialid known so far that lacks prothoracic sensory pits, a condition that is probably a unique or rare derivation (Grehan & Rawlins, 2003). The relationship of Leto with the tergal lobe clade is based on the share presence of the proximate Sc-R1 veins and their divergence towards the costal margin.
Fig. 3. Tergal lobe clade. See text for discussion. (5) West Eurasian clade. This group lacks significant bootstrap support, but includes several genera that are principally or exclusively western Eurasian in distribution with in an unresolved relationship with the North American Sthenopis and New Zealand Aoraia (Fig. 4).
The inclusion of Aoraia in this clade appears to be considered an artifact of the clustering procedure as there are no features shared
Trichophassus
Roseala
Hepialyxodes
Druceiella
Tricladia 54
Pseudodalaca
Pseudophilaenia
Pseudophassus
Pfitzneriana P. guianensiss
Hampsoniella
Cibyra
Aepytus
Alloaepytus
Gymelloxes
Schaefferiana
Vietteogorgopis
Abantiades
Trictena
Phassodes

Bulletin of the Buffalo Society of Natural Sciences 42, 2012 46
between Aoraia and the hepialine genera that are not also shared with other non-hepialine genera. The inclusion of Sthenopis is less problematic, although in the current analysis there are no uniquely shared features with the remaining hepialines which together share in common a unique spoutlike pseudteguminal apex, and there are also several other characters shared in common between different genera.
Fig. 4. West Eurasian group (Zenophassus, Triodia, Pharmacis, Hepialus) with an unresolved relationship with the North American Sthenopis and New Zealand Aoraia.
The inclusion of Zenophassus schamyl with several principally western Eurasian genera in this group was not anticipated as Z. schamyl is similar in biology to the North American Sthenopis where larvae first tunnel and feeding within host-plant roots followed by tunneling into the lower stem as the larva matures and it is from the stem that the adult emerges (Grehan, 1989). This shared pattern of development, along with a superficial similarity of the adults led to the suggestion that these genera were closely related (Grehan & Rawlins, 2003) but this now seems unlikely. The adult of Z. schamyl is noticeably larger than the other western Eurasian species and has a forewing pattern superficially similar to Sthenopis purpurescens and S. argenteomaculatus, but not S. thule or S. auratus. Although not strongly supported in this study, the several uniquely shared features between various member genera may predict future corroboration from other characters. (6) Andean clade. Supported with a bootstrap value of 72, but two characters states (absence of tibial spur, broad anterior SII margin) are shared with many other genera. The other character state, a melanized pseudotegumen is also shared with some other genera. Nielsen & Robinson (1983) also note this similarity along with specialization of SVIII in males (not
considered in the current study), but the female genitalia were also noted to differ in several characters. (6) Oxycanine clade. This group has low bootstrap support but conforms to the classic Old World oxycanine ghost moths of New Zealand and Australia (sensu Dumbleton, 1966; see also Brown et al., 2000). Not included in this analysis are the Asian and North African genera Neohepialiscus, Parahepialiscus, Hepialiscus that also share an oxycanine wing venation pattern. Oxycanus may be polyphyletic as some species share the presence of unique paranal sclerites with at least some Hepialiscus species (Grehan, 2011). This group was found to be monophyletic by Brown et al (2000), but those results are problematic because the outgroup was limited to two other hepialid genera rather than outside the Hepialidae. The character states used in their study may be valid, but require corroboration through comparison with all other hepialid genera and outgroup comparison with other Exoporia (Appendix 3). DISCUSSION The close relationship of the tergal lobe and Western Pacific clades conforms to an earlier inference by Grehan (2010) for a trans-Pacific relationship that also included the genus Callipielus, but no additional supporting evidence was found for the latter genus in this study. The possible addition of Leto will require future corroboration to asses the biogeographic implications. The tergal lobe clade represents a major evolutionary radiation from Mexico to southern Patagonia that is largely or entirely east of the Andes south of Ecuador. This distribution overlaps Schausiana in Mexico and Phassus in southern Mexico and Central America north of Colombia, and in southeastern Brazil. Phassus and Schausiana may be closely related to the tergal lobe clade, or allied with the east Asian Endoclita and Australasian Aenetus as suggested by a shared similarity in the larval prothoracic sensory pit and reddish-brown metatibial scent scales on the adult hind leg (Grehan and Rawlins, 2003). Other hepialid genera south of the Mexico-US border are in southern South America entirely or mostly to the west of the Andes (Nielsen & Robinson, 1983) or within the Andean system of Peru-Ecuador (at least two
Triodia
Sthenopis
Pharmacis
Zenophassus
Aoraia
Hepialus

Bulletin of the Buffalo Society of Natural Sciences 42, 2012 47
genera) where the tergal lobe clade appears to be absent.
Other clades found in this study must
be regarded as tentative given the low number of character states involved. A key requirement for further study will be to document and illustrate character states in the hepialid outgroups to substantiate or refute derived character states presented in this study, and to identify additional character states that will contribute to the resolution of all generic relationships within the Hepialidae. ACKNOWLEDGEMENTS I am extremely grateful to colleagues in various collections (listed in Methods) that have allowed long-term and renewed loans of material for over a decade, and for continuing this level of support following loss of former institutional resources. Without this sustained support this study would have been impossible. I am also most appreciative for specimens contributed by friends and colleagues over the many years I have been interested in the Hepialidae. These specimens have proved to be extremely helpful to my efforts. For continued insights and discussion on the systematics of Hepialidae. I am grateful to Don Davis, John Dugdale, and Carlos Mielke, Thomas Simonsen for comments on the manuscript. I am also greatly appreciate discussions on hepialid and Lepidoptera systematics with these individuals and John Rawlins.
REFERENCES Brown, B., Dugdale, J.S., Emberson, R.M. &
Paterson, A.M. 2000. Phylogeny of New Zealand hepialid moths (Lepidoptera: Hepialidae) inferred from a cladistic analysis of morphological data. Systematic Entomology 25, 1-14.
Common, I.1990. Moths of Australia. E.J. Brill, New York and Melbourne. University Press, Melbourne, Australia.
Davis, D. R. 1996. A revision of the southern African family Prototheoridae (Lepidoptera: Hepialoidea). Entomologica Scandanavica 27, 393-439.
Davis, D. R., Karsholt, O., Kristensen, N. P. and Nielsen, E. S., 1995, Revision of the genus Ogygioses (Palaeosetidae), Invertebrate Taxonomy 9, 1231-1263.
Dugdale, J.S. 1994. Hepialidae. Fauna of New Zealand 30, 1-163.
Dumbleton, L.J. 1966. Genitalia, classification and zoogeography of the New Zealand Hepialidae (Lepidoptera). New Zealand Journal of Science 9: 920-981.
Ebach, M.C., De Carvalho, M.R. & Williams, D.M. 2011. Opening pandora’s molecular box. Zootaxa 2946, 60–64.
Grehan, J. R. 1989. Larval feeding habits of the Hepialidae. Journal of Natural History (British Museum) 33, 469-485.
Grehan, J.R. 2010. Structural variants in the morphology of the first abdominal tergite supporting the monophyly of the Latin American genera Cibyra, Druceiella, Pfitzneriella, and Trichophassus (Lepidoptera: Hepialidae). Bulletin of the Buffalo Society of Natural Sciences 39, 43-63.
Grehan, J.R. 2011. Notes on the biogeography of eastern Asian ghost moths (Lepidoptera: Hepialidae). Bulletin of the Buffalo Society of Natural Sciences 40, 59-74.
Grehan, J.R. 2012. Evidence for the taxonomic validity of the ghost moth Korscheltellus fusconebulosa (De Geer, 1778) (Lepidoptera: Hepialidae) and the monophyly of Korscheltellus Börner, 1920. Bulletin of the Buffalo Society of Natural Sciences 41 (in press).
Grehan, J.R. & Rawlins, J.E. 2003. Larval description of a New World ghost moth, Phassus sp., and the evolutionary biogeography of wood-boring Hepialidae (Lepidoptera: Exoporia: Hepialoidea). Proceedings of the Entomological Society of Washington 105, 733-755.
Grehan, J.R., & Schwartz, J.H. 2009. Evolution of the second orangutan: phylogeny and biogeography of hominid origins. Journal of Biogeography 36, 1823–1844.
Heads, M.J. 2012. Molecular Panbiogeography of the Tropics. University of California Press, Berkeley.
Janse, A.J.T. 1942. The moths of South Africa. Volume IV, part 1. Jugatae. Published by the Author and University of Pretoria. Pretoria.
Kristensen, N. P. 1998. The homoneurous glossata. Handbook of Zoology 4 (35), 51–63
Livezey, B. 2011. Progress and obstacles in the phylogenetics of modern birds (Neornithes). In: Dyke, G. and Chiappe, L. (eds),

Bulletin of the Buffalo Society of Natural Sciences 42, 2012 48
Evolution of modern birds. Univ. California Press, Berkeley, pp. 117-145.
Mielke, C.G.C. & Grehan, J.R. 2012. Catalogue of the Latin American Hepialidae with taxonomic remarks (Lepidoptera). Nachrichten des Entomologische Vereins Apollo, N. F. 32, 131–158.
Mooi, R.D. & Gill, A.C. 2010. Phylogenies without Synapomorphies—A Crisis in Fish Systematics: Time to show some character. Zootaxa 2450, 26–40.
Nielsen, E.S. 1988. The peculiar Asian ghost moth genus Bipectalis Chu & Wang: taxonomy and systematic position (Lepidoptera: Hepialidae s.str.). Systematic Entomology 13, 171-195.
Nielsen, E.S & Kristensen, N.P. 1989. Primitive ghost moths. Morphology and taxonomy of the Australian genus Fraus Walker (Lepidoptera: Hepialidae s.lat.). Monographs of the Australian Lepidoptera 1, 1-206.
Nielsen, E.S, & Robinson, G.S. 1983. Ghost moths of southern South America. Entomonograph 4, 1-192.
Nielsen, E.S, & Robinson, G.S. & Wagner, D.L. 2000. Ghost moths of the world: a global inventory and bibliography of the Exoporia (Mnesarchaeoidea and Hepialoidea) (Lepidoptera). Journal of Natural History 34, 823-878.
Nielsen, E. S. & Scoble, M.J.1986. Afrotheora, a new genus of primitive Hepialidae from Africa (Lepidoptera: Hepialoidea). Entomologica Scandanvica 17, 29-54.
Schwartz, J.H. 1987. The Red Ape: Orang-utans and Human Origins. Houghton Mifflin Company, Boston.
Schwartz, J.H. 2004. Barking up the wrong ape – australopiths and the quest for chimpanzee characters in hominid fossils. Collegium Antropologicum 28, 87–101.
Schwartz, J.H. 2005. The Red ape: Orang-utans and Human Origins. Second revised edition. Westview Press, Boulder, CO.
Schwartz, J.H. In press. Organismal biology, molecular systematics, and phylogenetic reconstruction. In Leaping ahead: Advances in Prosimian Biology (ed. by J. Masters, M. Gamba and F. Génin). Springer Science, New York.
Schwartz, J.H. & Maresca, B. 2007. Do molecular clocks run at all? A critique of molecular systematics. Biological Theory 1, 357–371.
Thomas J. Simonsen, T.J., de Jong, R., Heikkilä, M. & Kaila, K. 2012. Butterfly morphology in a molecular age: Does it still matter in butterfly systematics? Arthropod Structure & Development 41, 307-322.
Swofford, D.L. 2005. Phylogenetic analysis using parsimony (*and other methods), Version 4. Sinauer Associates, Sunderland, MA.
Tindale, N.B.1932. Revision of the ghost moths (Lepidoptera Homoneura, family Hepialidae) Part I. Records of the South Australian Museum 4, 497-536.
Tindale, N.B.1935. Revision of the ghost moths (Lepidoptera Homoneura, family Hepialidae) Part II. Records of the South Australian Museum 5, 13-43.
Tindale, N.B.1941. Revision of the ghost moths (Lepidoptera Homoneura, family Hepialidae) Part IV. Records of the South Australian Museum 7, 15-46.
Tobi, D. & Grehan, J.R. 1993. Review of the ecological and economic significance of forest Hepialidae (Lepidoptera). Forest Ecology and Management 56, 1-12.
Ueda, K. 1988. New species of the genus Hepialiscus, Hampson (Lepidoptera, Hepialidae) from Taiwan. Bulletin of the Kitakyushu Museum of Natural History 15, 35-41.
Van Nieukerken, E.J. 2011. Order Lepidoptera Linnaeus, 1758. Zootaxa 3148, 212-221.
Wagner, D. L., 1985. The biosystematics of the Holarctic Hepialidae, with special emphasis on the Hepialus californicus species group. PhD Dissertation. University of California, Berkeley.
Wagner, D.L. & Rosovsky, J. 1991. Mating systems in primitive Lepidoptera, with emphasis on the reproductive behavior of Korscheltellus gracilis (Hepialidae). The Zoological Journal of the Linnean Society 102, 277-303.
Zhu, H., Wang, L. & Han, H. 2003. Lepidoptera Hepialidae Epiplemidae. Fauna Sinica 38, 1-167.

Bulletin of the Buffalo Society of Natural Sciences 42, 2012 49
Appendix 1. Comments on previously published character states separating Fraus, Gazoryctra, Antihepialus, and Afrotheora from the Hepialidae sensu stricto (Nielsen, 1988)
1. Male genitalia with trulleum fused or hinged to ventral part of pseudotegumen rather than being ‘free’ in the ‘primitive Hepialidae’. The ‘free’ state of the trulleum has never been precisely described and because it is problematic to recognize, it is not included in the current study.
2. Pretarsus lacks or has only a ‘small’ pseudoempodial seta (Nielsen& Kristensen 1988; Nielsen & Scoble 1988). This feature may be correct, but was not examined in this study due to its small size exceeding available magnification.
3. Pupa with an unbroken ventral transverse row of spines on segment VII (Nielsen & Kristensen, 1989). Because the pupa is not documented for many genera this feature was not included in this study as it would introduce numerous missing values that could affect the analysis.
4. Short abdominal tergum I where the length to width ratio is less than 0.3. No information was provided on the species or measurements. Ratios observed in this study for Bipectilis (0.24) Gazoryctra (0.27) and Dalaca (0.26) and Eudalaca (0.23) suggest that the ratio proposed for separating Hepialidae sensu stricto is not valid. Further measurements for the Hepialidae were not possible in this study due to an institutional restructuring.
Appendix 2. Comments on character states identified for relationships of 10 genera in the
Hepialidae by Wagner & Rosovsky (1991). Absence of documentation for all or most genera render the following features subject to future corroboration. Characters 8-12 not considered here as they apply to the relationship of Phymatopus of North America and Europe which was accepted for the present study. Separation of Fraus from remaining genera:
1. Retention of proboscis in Fraus (as the basal hepialid genus). Probably correct, but not included in this study due to lack of time to examine specimens confirm the presence or absence of this feature in all Hepialidae.
2. Less than 39 antennal segments except for Fraus. Considered here to be invalid. Examination of Druceiella showed an average of 34 and a range of 28-43.
Separation of Fraus and Gazoryctra from remaining genera: 3. Absence of tibial spurs. Valid and included in the current study. 4. Complete caudal band of teeth on A7 of the pupa. Possibly correct but not included
because the pupa is not described for most genera. 5. Fused trulleum in the male genitalia (after Nielsen & Scoble, 1986). Possibly
correct, but not included in this study because of difficulty of characterizing presence or absence of fusion.
Separation of Fraus, Gazoryctra, Triodia, Pharmacis, and Korscheltellus from remaining genera:
6. Metatibiae not swollen. Metatibiae are noticeably swollen in Phymatopus of North America and Europe, but whether this distinction does not seem to be more applicable to Hepialus, Zenophassus, and Sthenopis compared with other Hepialidae in general. Further evaluation is required.
7. Rosette-like metatibial hair pencils. In addition to Phymatopus, Hepialus, Zenophassus, and Sthenopis, metatibial hair pencils are present in Endoclita, Phassus, Schausiana, Puermytrans, some Thitarodes, and a yet to be described new genus from Peru. In the absence of a precise definition of ‘rosette-like’, this feature was not included in the current study. The presence of metatibial hair pencils is a prime face apomorphy although it includes genera that show no other apparent close relationship (see also, Grehan & Rawlins, 2003).
8. Loss of protibial epiphysis. Loss of this feature occurs in various other genera. Hepialidae, including within individual genera

Bulletin of the Buffalo Society of Natural Sciences 42, 2012 50
These three features do characterize the remaining genera (Phymatopus, Hepialus, Zenophassus, and Sthenopis).
13. Broad forewing scales with rounded apices. This feature may define a monophyletic group within the Hepialidae and needs to be examined for all genera as well as other Exoporia.
14. At least some forewing scales with reduced secondary ridging . This feature may define a monophyletic group within the Hepialidae and needs to be examined for all genera and other Exoporia.
15. Subtriangular forewings with falcate apices. This feature appears to also apply to other Hepialidae. It is variable within Endoclita and it does not apply to all Sthenopis (e.g. S. thule). This feature requires further investigation.
16. Horn-like twin processes in the genitalia. This appears to be a valid apomorphy that also applies to some other genera and is included in the current study.
Appendix 3. Comments on character states defining the monophyly of New Zealand oxycanines
by Brown et al. (2000) who examined 63 characters to assess the relationships of New Zealand Cladoxycanus, Dioxycanus, Dumbletonius, Heloxycanus, and Wiseana. The outgroup comprised three species of Aoraia and Aenetus virescens. With respect to this outgroup there were nine unambiguous apomorphic states supporting the monophyly of all the oxycanine genera. Lack of time and resources in the current study did not provide the opportunity to consider all of these features, but they are all contingent upon future corroboration of their absence outside of the Hepialidae.
9. Prelabial palp insertion is not raised. In the absence of comparison with the outgroup and other Hepialidae this feature was not included in the current study.
13. Antennal microsensilla present. May be valid. Resources for examining this feature not available for the current study.
15. Forewing veins R4 and R5 arise separately from a combined R2+3 stem. This is valid and was included in the current study.
36. Valvae without acuminate processes. This feature is problematic as many hepialids lack acuminate processes as do other hepialoids and Mnesarchaea.
45. Ovipore on weakly to strongly bilobed and erect papilla. Ovipore not known for most genera and therefore excluded from the current study.
47. Antrum floor of the copulatory pore ventral to the intergenital lobes is membranous in Aenetus and Aoraia, but sclerotized within the oxycanine genera. This condition is not documented for most genera and therefore not included in the current study.
49. On the caudal margin of tergum 8 there is a broad tuft of long hair-like scales in the oxycanine genera but not in Aenetus or Aoraia. This feature was not assessed in the present study, but may prove to be valid upon future examination.
50. Stemmata on head capsule arranged in two parallel rows in Aenetus and Aoraia, and in one straight and one curved row or in two parallel arcs in oxycanine genera. Stemmata arrangement not documented for many genera and requires further assessment.
55. Position of metathoracic seta L3 in relation to SD1 and SD2. Not documented for larvae of most genera and not included in the present study.
Appendix 4. Phylogenetic character matrix. Characters as in text. Taxa: 1 – Outgroup, represented where possible, by species of Mnesarchaeidae, Anomoses, Neotheora, Ogygioses, Palaeoses, Genustes and Prototheora; 2 – Gazoryctra; 3 – Antihepialus; 4 – Fraus; 5 – Afrotheora; 6 – Abantiades; 7 – Andeabatis; 8 – Aoraia; 9 – Callipiellus; 10 – Cladoxycanus; 11 – Dioxycanus; 12 – Druceiella; 13 – Aepytus; 14 – Alloaepytus; 15 – Cibyra; 16 – Gymelloxes; 17 – Hampsoniella; 18 – Hepialyxodes; 19 – Pfitzneriana; 20 – Phassus guianensis; 21 – Pseudodalaca; 22 – Pseudophassus; 23 – Pseudophilaenia; 24 – Roseala; 25 – Schaefferiana; 26 – Trichophassus; 27 – Tricladia; 28 – Vietteogorgopis; 29 – Dumbletonius; 30 – Elhamma; 31 – Heloxycanus; 32 – Hepialus; 33 – Korscheltellus; 34 – Leto; 35 – Oxycanus; 36 – Parapielus; 37 – Pharmacis; 38 – Phassodes; 39

Bulletin of the Buffalo Society of Natural Sciences 42, 2012 51
– Phassus; 40 – Phymatopus Europe; 40 - Phymatopus USA; 41 – Sthenopis; 42 – Trictena; 43 – Triodia; 44 – Wiseana; 45 – Zenophassus.
1 2 3 4 5 6 7 8 9 10
11
12
13
14
15
16
17
18
19
20
21
22
23
24
25
26
27
28
29
30
31
32
33
34
35
36
1 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0
2 0 0 0 0 0 0 0 0 0 1 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 1 0 1 0 0 0 0 0 0 0 0
3 0 0 0 0 0 0 0 0 0 1 0 0 0 0 0 0 0 0 0 0 0 0 0 0 1 1 0 1 0 0 0 0 0 0 0 0
4 0 0 0 0 0 0 0 0 0 1 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 1 0 0 0 0 0 0 0 0
5 0 0 0 0 0 0 1 0 0 1 0 0 0 0 0 0 0 0 0 0 0 0 0 0 1 0 0 1 0 0 0 0 0 0 0 0
6 0 0 0 1 0 0 1 1 1 0 0 0 0 0 1 1 0 0 1 0 0 0 0 0 0 0 1 0 0 0 0 0 0 0 0 1
7 0 0 0 0 0 0 1 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 1 0 0 0 0 0 0 0 1 0
8 0 0 0 0 0 0 1 0 1 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 1 0 0 0 0 0 0 0 0 1
9 0 0 0 1 0 0 1 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 1 0 0 0 0 0 0 0 0 0 10 0 0 0 0 0 0 1 0 0 0 0 0 0 0 0 0 1 0 0 0 0 0 0 0 0 0 1 0 0 0 0 0 0 1 0 0 11 0 0 0 0 0 0 1 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 1 0 0 1 0 0 0 0 0 0 1 0 1 12 0 0 1 1 1 1 1 0 0 0 0 1 0 0 1 1 0 1 0 0 0 0 0 0 0 0 1 0 0 0 0 0 0 0 0 0 13 1 1 1 1 1 0 1 0 0 0 0 0 0 0 1 1 0 0 0 0 0 0 0 0 0 0 1 0 0 0 0 0 0 1 0 0 14 0 0 1 1 1 1 1 0 0 0 0 1 0 0 1 0 0 1 0 0 0 0 0 0 0 0 1 0 0 0 0 0 0 0 0 0 15 1 1 1 1 1 0 1 0 0 0 0 0 0 0 1 1 0 0 0 0 0 0 0 0 0 0 1 0 0 0 0 0 0 0 0 0 16 0 0 1 1 1 1 1 0 0 0 1 1 0 0 1 1 0 0 0 0 0 0 0 0 0 0 1 0 0 0 0 0 0 0 0 0 17 0 0 1 1 1 1 1 0 0 0 1 0 0 0 1 1 0 0 0 0 0 0 0 0 0 0 1 0 0 0 0 0 0 0 0 0 18 0 1 1 1 1 0 1 0 0 0 0 0 0 0 1 1 0 1 0 0 0 0 0 0 0 0 1 0 0 0 0 0 0 0 0 0 19 0 0 1 1 1 1 1 0 0 0 0 1 0 0 1 1 0 1 0 1 0 0 0 0 0 0 1 0 0 0 0 0 0 0 0 0 20 0 0 1 1 1 1 1 0 0 0 0 1 0 0 1 0 0 0 0 0 0 0 0 0 0 0 1 0 0 0 0 0 0 0 0 0 21 0 0 1 1 1 1 1 0 0 0 1 0 0 0 1 1 0 1 0 0 0 0 0 0 0 0 1 0 0 0 0 0 0 0 0 0 22 0 0 1 1 1 0 1 0 0 0 1 1 1 0 1 1 0 0 0 1 0 0 0 0 0 0 1 0 0 0 0 0 0 0 0 0 23 0 0 1 1 1 0 1 0 0 0 0 0 0 0 1 1 0 0 0 0 0 0 0 0 0 0 1 0 0 0 0 0 0 0 0 0 24 0 0 1 1 1 1 1 0 0 0 0 1 0 0 1 1 0 0 0 0 0 0 0 0 0 0 1 0 0 0 0 0 0 0 0 0 25 1 1 1 1 1 0 1 0 0 0 0 0 0 0 1 0 0 0 0 0 0 0 0 0 0 0 1 0 0 0 0 0 0 0 0 0 26 0 0 1 1 1 1 1 0 0 0 1 0 0 0 1 0 0 0 0 0 0 0 0 0 0 0 1 0 0 0 0 0 0 0 0 0 27 0 0 1 1 1 1 1 0 0 0 0 0 1 0 1 1 0 0 0 1 0 0 0 0 0 0 1 0 0 0 0 0 0 1 0 0 28 0 1 1 1 1 1 0 0 0 0 0 0 0 1 1 0 0 0 1 0 0 0 0 0 0 1 0 0 0 0 0 0 1 0 0 29 0 0 0 0 0 0 1 0 0 0 0 0 0 0 0 0 1 0 0 0 0 0 1 1 0 0 1 0 0 0 0 0 0 1 0 1 30 0 0 0 0 0 0 1 0 0 0 0 0 0 0 1 1 0 0 0 0 0 0 0 0 0 0 1 0 0 0 0 0 0 1 0 0 31 0 0 0 0 0 0 1 0 0 0 0 0 0 0 0 0 1 0 0 0 0 0 0 1 0 0 1 0 0 0 0 0 0 1 0 0 32 0 0 0 0 0 0 1 1 1 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 1 0 1 0 0 1 0 0 0 1 33 0 0 0 0 0 0 1 0 1 0 0 0 0 0 0 0 0 0 0 0 0 0 0 1 0 0 1 0 1 1 1 0 0 0 0 1 34 0 0 0 0 0 0 1 0 0 0 0 0 0 0 1 1 0 0 0 0 0 0 0 1 0 0 1 0 0 0 0 0 0 0 0 0 35 0 0 0 0 0 0 1 0 0 0 0 0 0 0 0 0 1 0 0 0 0 0 1 1 0 0 1 0 0 0 0 0 0 1 0 0 36 0 0 0 0 0 0 1 0 0 0 0 0 0 0 1 0 0 0 0 0 0 0 0 0 0 0 1 0 0 0 0 0 0 0 0 0 37 0 0 0 0 0 0 1 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 1 0 1 0 0 1 0 0 0 1

Bulletin of the Buffalo Society of Natural Sciences 42, 2012 52
38 0 0 0 1 0 0 1 1 0 0 0 0 0 0 1 1 0 0 0 0 0 0 0 0 0 0 1 0 0 0 0 0 0 0 0 1 39 0 0 0 0 0 0 1 0 0 0 0 0 0 0 0 0 0 0 0 0 1 1 0 0 0 0 1 0 0 0 0 0 0 0 0 0 40 0 0 0 0 0 0 1 0 0 0 0 0 0 0 0 0 0 0 0 0 1 1 0 0 0 0 1 0 0 0 0 0 0 0 0 0 41 0 0 0 0 0 0 1 1 1 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 1 0 0 0 0 0 0 0 0 0 42 0 0 0 1 0 0 1 1 0 0 0 0 0 0 1 0 0 0 1 0 0 0 0 0 0 0 1 0 0 0 0 0 0 0 0 1 43 0 0 0 0 0 0 1 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 1 0 1 1 1 1 1 0 0 1 44 0 0 0 0 0 0 1 0 0 0 0 0 0 0 0 0 1 0 0 0 0 0 1 1 0 0 1 0 0 0 0 0 0 1 0 1 45 0 0 0 0 0 0 1 1 1 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 1 0 1 0 0 1 1 0 0 1
Appendix 5. Anterior hindwing position of Sc and R. Specimens were prepared by brushing scales from the relevant area of the hindwing. This approach was considered preferable to slide mounts that may not always accurately represent the original condition. In some cases it was necessary to flatten the wing under considerable pressure to show the position of veins that may otherwise be obscured by the pleated topography of the wing veins. Outcond. = outgroup condition for Sc and R being relatively far apart. Derived conditions highlighted in yellow as ‘narrow’ where Sc and R are adjacent or within about the width of a vein apart, ‘basally narrow’ where Sc and R are adjacent or within about the width of a vein apart towards the base of the wing, but further apart distally, and ‘div.’ refers to a wider separation of Sc & R towards the costa. (A) Outgroup Examples
Mnesarchaeidae Mnesarchaea hamadelpha (Hudson, 1928 Plate A)
Paleosetidae Palaeoses scholastica (Common, 1990)
Outgroup taxon: Neotheoridae Neotheora chiloides (Kristensen, 1978)
Outgroup taxon: Prototheoridae Prototheora petrosema (Janse, 1940; Davis, 1996)
Outgroup taxon: Prototheoridae Prototheora corvifera (Philpott, 1928 Fig. 2)
Outgroup taxon: Anomosetidae Anomoses hylecoetes (Common, 1990)
(B) Hepialidae (in group)
Bipectilus unimaculata (Daniel, 1940) (Nielsen, 1988) Outcond.
Fraus griseomaculata Nielsen and Kristensen, 1989 [NZAC] Outcond.
Gazoryctra sciophanes (Ferguson, 1979) [JRG] Outcond.
Antihepialus antarcticus (Wallengren, 1860) [M132: CNC] Outcond.

Bulletin of the Buffalo Society of Natural Sciences 42, 2012 53
Dalaca pallens (Blanchard, 1852) [NZAC: JSD595] Outcond.
Andeabatis chilensis (Ureta, 1951) [NMNH: M211] Outcond.
Korscheltellus gracilis (Grote, [1865]) [JRG] Outcond.
Korsheltellus lupulina (Linnaeus, 1758) [JRG] Outcond.
Oncopera intricata Walker, 1856 [PMNH] Outcond.
Eudalaca infumata (Janse, 1942) [MCZ] Outcond
Eudalaca orthocosma (Janse, 1942) [PMNH] Outcond.
Thitarodes eberti Viette, 1968 [CNC] Outcond.
Hepialus humuli (Linnaeus, 1758) [JRG: M20] Outcond.
Pharmacis bertrandi (Le Cerf, 1936) [JRG] Outcond.
Triodia sylvania (Linnaeus, 1761) [JRG: M130] Outcond.
Phymatopus californicus (Boisduval, 1868) [JRG: M33] Outcond.
Phymatopus hectoides (Boisduval, 1868) [JRG] Outcond.
Phymatopus hecta (Linnaeus, 1758) [CMNH] Outcond.
Gorgopis libania (Stoll, 1781) [PMNH] Outcond.
Pfitzneriella monticola (Maassen, 1890) [CMNH: M136] Outcond.

Bulletin of the Buffalo Society of Natural Sciences 42, 2012 54
Pfitzneriella remota (Pfitzner, 1906) (Pfitzner, 1906) [JRG: M200] Outcond.
Puermytrans chilensis Viette, 1951 [JRG: F32] Outcond.
Blanchardinella venosus (Blanchard, 1852) [CMNH] Outcond.
Zelotypia stacyii Scott, 1859 [NZAC] Outcond.
Endoclita sp. [CMNH] Outcond.
Sthenopis argenteomaculatus(Harris, 1841) [JRG] Outcond.
Sthenopis auratus (Grote, 1878) [JRG] Outcond.
Sthenopis purpurascens (Packard, 1863) [CNC] Outcond.
Zenophassus schamyl (Christoph, 1888) [JRG] Outcond.
Dioxycanus fusca (Philpott, 1914) [JRG: M225] Outcond.
Phassus chrysodidyma Dyar, 1915 [AMNH: M113] Outcond.
Phassus huebneri (Geyer, [1838]) [M228] Outcond.
Phassus pharus (Druce, 1887) [JRG] Outcond.
Pseudophassus mahagoniatus Pfitzner, 1914 [RMBH] Narrow, dist. divergent
Hampsoniella sp. [FSM] Narrow, dist. divergent
Pseudodalaca gugelmani (Viette, 1950) [SAMA: M189] Narrow, dist. divergent

Bulletin of the Buffalo Society of Natural Sciences 42, 2012 55
Pseudophilaenia nr omagua (Pfitzner, 1937) [JRG] Narrow, dist. divergent
Gymelloxes terea (Schaus, 1892) [CMNH: M19] Narrowly parallel
Alloaepytus tesselloides (Schaus, 1901) [AMNH] Narrow.
Schaefferiana epigramma(Herrich-Schäffer, [1854]) [JRG: M183] Narrow.
Pfitzneriana sp. [AMNH] Narrow, dist. divergent Cibyra dorita Schaus, 1901 [JRG: M226] Narrow, dist. divergent
Cibyra ferruginosa Walker, 1856 [JRG:M140] Narrow, dist. divergent
Cibyra n.sp. [JRG] Narrow, dist. divergent
Cibyra stigmatica (Pfitzner, 1937) [JRG: M141] Narrow, div.
Hepialyxoides rileyi Viette, 1951 [NMNH] Narrow, div.
Druceiella n.sp. [JRG: s106] Narrow, dist. div.
Druceiella metellus (Druce, 1890) [JRG: s050] Narrow, div.
Druceiella momus (Druce, 1890) [AMNH: M101] Narrow, div.
Druceiella n. sp. [JRG: M94] Narrow, div.
Callipielus arenosus (JRG: M125) Narrow, div.
Abantiades hyalanthus (CMNH) Narrow, div.

Bulletin of the Buffalo Society of Natural Sciences 42, 2012 56
Abantiades labyrinthicus (Donovan, 1805) [CMNH] Narrow, div.
Trictena atripalpis (Walker, 1856 ) [CMNH] Narrow
Aoraia dinodes (Meyrick, 1890) [NZAC] Outcond.
Aoraia oreobolae Dugdale, 1994 [JRG: M223] Outcond.
Aoraia enysii (Butler, 1877) [JRG] base narrow
Aoraia rufivena Dugdale, 1994 [JRG: M135] base narrow, div.
Phassodes vitiensis (Rothschild, 1895) [NZAC: L34) Narrow, div.
Leto venus (Cramer, 1780) [PMNH] Narrow, div.
Aepytus guarani (Pfitzner, 1914) [JRG: M180] Narrow, div.
Tricladia prytanes (Schaus, 1892) [JRG: M197] Narrow, div.
Vietteogorgopis jordani (Viette, 1956) [JRG: M88] Narrow, div.
Elhamma australasiae (Walker, 1856) [CMNH] Narrow, div.
Parapielus leuticornis (Berg, 1882) [CNC] Narrow
Aenetus cohici Viette, 1961 [JRG: M205] Outgroup condition
Aenetus marginatus Rothschild, 1896 [AMNH: M205] outgroup condition
Aenetus nr toxopeusi Viette, 1956 outgroup condition

Bulletin of the Buffalo Society of Natural Sciences 42, 2012 57
Aenetus ombraloma (Lower, 1902) [NZAC] Distally narrow
Aenetus astathes (Turner, 1915) [NZAC] Distally narrow
Aenetus ligniveren (Lewin, 1805) [NZAC: JSD578] Distally narrow
Aenetus eximia (Scott, 1869) [JRG: M207] Distally narrow
Aenetus virescens (Doubleday, 1843) [JRG] Distally narrow
Aenetus virescens (JRG: M146) Fusion Sc-R1 near R2-R3
Cladoxycanus minos (Hudson, 1905) [JRG] Fusion Sc-R1
Dumbletonius characterifera (Walker, 1865) [JRG: M31] Fusion Sc-R1
Dumbletonius unimaculata (Salmon, 1948) [JRG] Fusion Sc-R1
Heloxycanus patricki Dugdale, 1994 [JRG] Fusion Sc-R1
Oxycanus sphragidius (Meyrick, 1890) [JRG] Fusion Sc-R1
Oxycanus sp. [CNC] Fusion Sc-R1
Wiseana signata (Walker, 1856) [JRG] Fusion Sc-R1
Wiseana jocosa (Meyrick, 1912) [JRG] Fusion Sc-R1

Bulletin of the Buffalo Society of Natural Sciences 42, 2012 58
Wiseana umbraculatus (Guenée, 1868) [JRG] Fusion Sc-R1
Appendix 6. Comparative morphology of second abdominal ventral sternite (SII) for Prototheora (hepialid outgroup genus in the Prototheoridae) and Hepialidae. Note variation in the shape of the anterior margin and the lateral bar (LB ) with respect to its anterior and lateral orientation. If the narrow shape of the lateral bar in Prototheora is characteristic of non-hepialid Exoporia, the broad dimensions of the lateral bar in Hepialidae may constitute an apomorphy for the family.
Prototheora sp. (outgroup) [PMNH: M199]
Afrotheora jordani [BMNH]
Antihepialus antarcticus (Wallengren, 1860) [CNC: M132]
Fraus simulans Walker, 1856 [JRG: F31]
Gazoryctra sciophanes Ferguson, 1979 [JRG: M131]
Druceiella basirubra (of authors) [NMNH: M198]
Druceiella amazonensis Viette, 1950 [BF: M192]
Druceiella metellus (Druce, 1890) [CUIC: M160]
Cibyra feruginosa Walker, 1856 [JRG: M154 ]
Pfitzneriana sp. [AMNH: M167]
Trichophassus giganteus (Herrich-Schäffer, [1853]) [NZAC: JSD593]
Aepytus gaurani (Pfitzner, 1914) [JRG: M180]
Alloaepytus tesselloides (Schaus, 1901) [CMNH: M182]
LB LB

Bulletin of the Buffalo Society of Natural Sciences 42, 2012 59
Gymelloxes) terea (Schaus, 1892) [JMM: M155]
Vietteogorgopsis sp. [JRG: M164]
Pseudophilaenia omagua (Pfitner, 1937) [LACM: M186]
(Pseudodalaca) gugelmanni (Viette, 1950) [UNAM: M87]
Schaefferiana epigramma (Herrich-Schäffer, [1854]) [JRG: M183]
Roseala tessellatus (Herrich-Schäffer, [1854]) [JRG: M187]
Pseudophassus mahagoniatus Pfitzner, 1914 [JRG: M157]
Cibyra monoargenteus (Viette, 1951) [JRG: M165]
Pfitzneriella monticola (Maassen, 1890) [CMNH: M193]
Phitzneriella remota (Pfitzner, 1906) [JRG: M200]
Andeabatis chilensis (Ureta, 1951) [NMNH: M211]
Puermytrans chilensis Viette, 1951 [JRG: F32]
Cladoxycanus minos (Hudson, 1905) [JRG: M137]
Dioxycanus fusca (Philpott, 1914) [JRG: M172]
Heloxycanus patricki Dugdale, 1994 [JRG M151]
Wiseana sp. [JRG: M174]

Bulletin of the Buffalo Society of Natural Sciences 42, 2012 60
Wiseana copularis (Meyrick, 1912) [JRG: M216]
Callipielus arenosus Butler, 1882 [JRG: M148]
Pfitzneriella monticola (Maassen, 1890) [CNMH M193]
Aoraia enysii (Butler, 1877) [JRG: M230]
Aoraia rufivena Dugdale, 1994 [JRG; M175]
Korscheltellus gracilis (Grote, [1865]) [JRG: M147]
Triodia sylvania (Linnaeus, 1761) [JRG: M158]
Pharmacis aemilianus (Constantini, 1911) [JRG: M45]
Pharmacis bertrandi (Le Cerf, 1936) [JRG: M156]
Hepialus humuli (Linnaeus, 1758) (JRG: M153]
Zenophassus schamyl (Christoph, 1888) [JRG: M134]
Sthenopis auratus (Grote, 1878) [JRG: M179]
Sthenopis purpurascens (Packard, 1863) [CNC: M119]
Sthenopis thule (Strecker, 1875) [ CMNH: M214]
Parapielus leuteicornis (Berg, 1882) [JRG: M152]
Blanchardinella venosus (Blanchard, 1852) [CNC: M106]

Bulletin of the Buffalo Society of Natural Sciences 42, 2012 61
Dalaca sp. [JRG:F30]
Abantiades latipennis Tindale, 1932 [JRG:M150]
Trictena atripalpis (Walker, 1856) [M37]]
Leto venus (Cramer, 1780) [PMNH:M127]
Zelotypia stacyii Scott, 1869 [NZAC:766]
Oxycanus dirempta (Walker, 1865) [JRG:M177]
Oxycanus rufescens Walker, 1856 [JRG:M177]
Oxycanus sp. [CMNH: M218] Merged image of SII from cut specimen
Jeana sp.[JRG: M194]
Oncopera rufobrunnea Tindale, 1933 [JRG: M185]
Thiterodes sp. [JRG: M201]
Phymatodes hecta (Linnaeus, 1758) [CMNH: M217]
Phymatopus californicus (Boisduval, 1868) [JRG: M33]
Phymatodes hectoides (Boisduval, 1868) [JRG: M210]
Phassodes vitiensis (Rothschild, 1895) [NZAC: L34]
Schausiana trojesa Schaus, 1901 [CMNH: M75]
Phassus heubneri (Geyer, [1838]) [M228]
Phassus pharus (Druce, 1887) [JRG: M213]
Phassus triangularis Edwards, 1885 (JRG: M159]
Endoclita signifier (Walker, 1856) [CNMH: M149]

Bulletin of the Buffalo Society of Natural Sciences 42, 2012 62
Aenetus cohici Viette, 1961 [JRG: M206]
Aenetus dulcis (Swinhoe, 1892) [M104, JRG]
Aenetus scotti (Scott, 1869) [JRG: F33]
Aenetus sp. (New Guinea) [JRG: M208]
Aenetus virescens (Doubleday, 1843)[JRG: M195]