haemoglobin inhibits gtp-hydrolysis and gdp/gtp-exchange activities of a low mr gtp-binding protein,...
TRANSCRIPT

British Journal of Huernutology, 1994, 88, 706-711
Haemoglobin inhibits GTP-hydrolysis and GDP/GTP-exchange activities of a low M, GTP-binding protein, ras p21
KOH-ICHI NAGATA* AND YOSHINORI NOZAWA Department of Biochemistry, Gifu University School of Medicine, Tsukasamachi, Gifu, Japan
Received 21 April 1994; accepted for publication 19 August 2 994
Summary. Haemoglobin was observed to inhibit the GDP/ GTP-exchange activity of rus protein (rus p21) by measure- ment of [3H]GDP-dissociation activity in time- and dose- dependent manners. Haemoglobin also inhibited the [32P]GTP-hydrolysis activity of rus p2 1 time- and dose- dependently. These inhibitory effects of haemoglobin were lost after incubation of haemoglobin at 80°C for 3min. Globin showed limited inhibition on the [ 32P]GTP-hydrolysis activity of rus p21. and haemin had no effect, indicating that the ternary tetrameric structure of haemoglobin is essential
for the inhibitory effects on rus p21 activities. Methaemoglo- bin also inhibited both [3H]GDP-dissociation and [32P]GTP- hydrolysis activities of rus p21 in a very similar manner to that by haemoglobin. The obtained results strongly suggest that haemoglobin suppresses the physiological function(s) of rus p21 in vivo inhibiting both [32P]GTP-hydrolysis and GDP/ GTP-dissociation of rus p21 in erythrocytes.
Keywords: rus p2 1, low M, GTP-binding protein, haemoglo- bin, erythrocyte, GDP dissociation inhibitor.
A family of rus genes consisting of Harvey (H)-, Kirsten (K)- and N-rus was first detected as active transforming genes in a variety of human tumours (Barbacid, 1987: Kaziro et ul, 1991: Satoh et ul, 1992). The cellular rus genes encode low M, GTP-binding proteins of 2 1 000 daltons (rus p2 1) and are localized in the inner surface of plasma membranes. rus p2 1 binds 1 mol of GTP or GDP per mol of protein and hydrolyses the bound GTP to GDP and inorganic phosphate. More than 50 genes of rus-related low M, GTP-binding proteins have been identified and they are believed to be involved in the regulation of a wide variety of cellular processes, including signal transduction across the plasma membrane, cytoskeletal organization, translocation of nascent proteins into the endoplasmic reticulum, and vesicular traffic within the cell (Pfeffer, 1992; Takai et al. 1992).
The rus gene is conserved through evolution and widely distributed among eukaryotes from yeast to mammals (Barbacid, 1987; Kaziro et ul, 1991). Normal rus p21 plays critical roles in the control of normal and transformed cell growth and differentiation. rus p21, as well as other GTP- binding proteins, undergoes two alternative conformations.
* Present address: Department of Molecular Pathobiochemistry. Gifu University School of Medicine, Tsukasamachi-40. Gifu 500, Japan.
Correspondence: Dr Yoshinori Nozawa, Department of Biochemis- try, Gifu University School of Medicine, Tsukasamachi-40, Gifu 500, Japan.
The GTP-bound form is ‘active’ in the sense that it can interact with a target molecule. On the other hand, the GDP- bound form is referred to as ‘inactive’ because it cannot stimulate the downstream target. Therefore the nucteotide exchange and inherent GTP-hydrolysis of rus p21 are essential for determination of their conformation and exertion of their biological functions. Because the GDP- dissociation (GDP/GTP-exchange) and GTP-hydrolysis rates of every low M, GTP-binding protein are very low, it has been suggested that regulatory components that modify guanine nucleotide exchange or GTP-hydrolysis reaction should be present. Indeed, four types of regulatory proteins for rus and rus-related GTP-binding proteins have been reported (Boguski & McCormick. 1993). The first group is the GTPase activating protein (GAP), which increases the intrinsic GTP-hydrolysis activity of some low M, GTP- binding proteins specifically. Among them, the GAP specific for rus p21 has been an object of especially intensive study (McCormick. 1989). Neurofibromin (NF1) has also been clarified to have GAP-activity for mammalian rus p2 1 (Hall, 1992). The second type of regulatory protein is a guanine nucleotide-exchange factor (GEF) which stimulates GDP- dissociation and the subsequent GTP-binding. In mamma- lian cells, Ras-GEF and Sos proteins function as a GEF for rus p21 (Shou et ul. 1992: Botwell et al, 1992). Then, such factors for other low M, GTP-binding proteins were identified or partially purified (Takai et ul. 1992: Boguski &
706

Inhibition of Activities of ras p 2 1 by Haemoglobin 707 McCormick, 1993). The third type of regulatory protein, designated as a GDP dissociation inhibitor (GDI). regulates the GDP/GTP exchange reaction by inhibiting the dissocia- tion of GDP from and subsequent binding of GTP to some low M, GTP-binding proteins (Takai et al, 1092). The fourth type of regulatory protein is a GTPase inhibiting protein (GIP) for ras p21 (Tsai et al. 1990). The notable feature of this protein is that it can be activated by acidic phospholipids and diacylglycerol.
In the present study we have demonstrated that haemoglobin may serve as GDI and (GIP for ras p21 and negatively regulate the protein.
MATERIALS AND METHODS
Materials. Expression plasmid, pJL6cIIras, of human cellular-Ha-ras p21, which is identical to rat Ha-ras p21, was a gift from Dr S. Hattori (National Institute of Neuroscience, Tokyo, Japan) (Hoshino et d, 1987). ras p21 overproduced in Escherichia coli was purified by successive column c:hromatographies of DEAE-Sephacel (Pharmacia Inc., Uppsala, Sweden) and hydroxyapatite HCA-100s (Tosoh Inc., Tokyo, Japan) (data not shown). Rat haemo- globin was purified to homogeneity from rat spleen and identiied ammo acid sequence analysis as briefly described in Discussion. Human haemogbbm ii which was ferrous when reconstituted in 25 m~ Tris/HCI (pH 7.4) containing 150 m NaCl, crystallized and lyophilized human methae- moglobin, and bovine haemin chloride were purchased from Sigma Chemical Co. (St Louis, Mo., U.S.A.). Globin was prepared from human haemoglobin by acid-acetone method as described (Ascoli et al, 1981). GTE', guanosine 5'-(3-0- thio)triphosphate (GTP-y-S), GDP, ArP and adenyl-5'-yl- P,y-imidodiphosphate (AppNHp) were from Boehringer Mannheim (Germany). [y-32P]GTP (specific activity, 222 TBq/mmol), [35S]GTP-y-S (specific activity, 43.4 TBq/mmol) and [8,5'-3H]GDP (340.4 GBq/mmol) were from Amersham Inc. (U.K.). Other reagents were from standard vendors.
[32P]GTP-hydrolysis inhibiting activity assay. To determine the intrinsic GTP-hydrolysis activity, ras p2 1 was preloaded with [T-~,P]GTP and the rate of release of the y-32P was determined by measuring residual bound radioactivity on the nitrocellulase membrane filter (Nagata et al, 1992b). In the first reaction, the purified active ras p21 (2pmol estimated by [35S]GTP-y-S binding) was incubated with 1 p~ [Y-~*P]GTP (4-5 x lo3 cpm/pmol) at 30°C for 30min in a 50 pl of the reaction mixture containing 2 5 m~ Tris/HCl (pH 7 9 , 5 m~ MgCl,, 10 m~ EDTA and 1 m~ dithiothreitol (DTT). The exchanging reaction was stopped by placing the reaction mixture on ice and adding 1 M MgC12 to a final Mg2+ concentration of 10 m ~ . Then the intrinsic GTP-hydrolysis rate was determined by incubating the [y-32P]GTP-loaded protein at 30°C in loop1 of 2 5 m ~ TrislHCl (pH 7.6) containing 1 m~ DTT, 0.5 m ATP, I m~ K2HP04, 1 O m ~ MgCl,, 0 5 m~ GTP and indicated amounts of haemoglobin. The reaclion was terminated at the indicated time points by the addition of 3 ml of ice-cold 20 m~ TrislHCI (pH 7.6) Containing 2 5 m ~ MgC12 and l o o m NaC1, followed by rapid filtrationon on nitrocellulose filter (Advantex Inc.,
Tokyo, Japan). Filters were then washed six times with the same buffer. After the filters were dried, they were dissolved in 8 ml of scintillation mixture for radioactivity counting. In some experiments, indicated amounts of globin, haemin or methaemoglobin were added instead of haemoglobin.
[3H]GDP-dissociation inhibiting activity assay. The rate constant for [3H]GDP-dissociation was measured by dis- placement of r3H]GDP from its complex with ras p21 in the presence of a large excess of GTP (0.5 m ~ ) and m~ order of Mg2+ (Nagata et al, 1992b). The [3H]GDP-bound rus p21 was first prepared by incubating ras p a l (2 pmol of active protein estimated [35S]GTP+-binding activity) with 1 p~ [3H]GDP (0.9-1.1 x lo4 dpm/pmol) for 30min at 30°C in a 25pI mixture containing 2 0 m Tris/HCl (pH 7.5), 5 m MgCl,, 10 m EDTA and 1 mMDTT. After the first incuba- tion, 3.5 pl of 20 m TrislHCl (pH 7.5) containing 106.9 mM MgCI, and 7 . 1 4 m GTP, 2 1 . 5 ~ 1 of solution containing 2 0 m Tris/HCl (pH 7.5) 0 . 5 m EDTA, lm DTT and indicated amounts of haemoglobin were added. Final Mg2+ concentration was adjusted to approximately 5 m. Then a second incubation for various times at 30°C was performed and the dissociation reaction was terminated by the same filtration method as the [32P]GTP-hydrolysis inhibiting activity assay. The radioactivity on the nitrocellulose filter was counted. In some experiments the final concentration of Mg2+ was adjusted to 0.5 p~ in the second incubation, and an indicated amount of methaemoglobin was added instead of haemoglobin.
Other assays. [35S]GTP-y-binding activity was determined as described (Nagata & Nozawa, 1988; Nagata et al, 1989). Briefly, samples were incubated for 20min at 30°C in a 100 pI reaction mixture containing 20 mM Tris/HCI (pH 7.5). 0-0965 m~ MgS04, 0.1 m~ EDTA, 1 mM dithiothreitol and 1 p~ [35S]GTP-y-S (1-2 x lo3 cpm/pmol). The reaction was stopped by the addition of 3 ml of ice-cold 20 m~ Tris( HCl (pH 7.5) containing l 0 0 m ~ NaCl and 2 5 m ~ MgCI,, followed by the rapid filtration on nitrocellulose filter. The radioactivity on the filter was counted. Protein was determined with Bio-Rad protein assay using bovine serum albumin as a standard (Bradford, 1976).
RESULTS
Effect ofhaemoglobin on [32PJGTP-hydrolysis activity of rasp21 The inhibition of [32P]GTP-hydrolysis of ras p21 by haemoglobin was in a time-dependent manner as shown in Fig 1A. The [32P]GTP-hydrolysis rate of ras p21 was decreased from 0-035 to 0.007min-' when 1 . 2 p g of haemoglobin was present in the reaction mixture. The dose-dependency of the inhibiting activity for [32P]GTP- hydrolysis of ras p21 was also demonstrated (Fig 1B). Human haemoglobin also exhibited almost the same time- and dose-dependent effects on [32P]GTP-hydrolysis of ras p21 (data not shown). The inhibiting activity of haemoglo- bin was stable at 4°C for at least 1 week, but diminished completely by heating at 80°C for 3 min.
Efjects of haemoglobin on [3HJGDP-dissociation of ras p21 In the next set of experiments, the effect of rat haemoglobin

708 Koh-khi Nagata and Yoshinori Nozawa
0 5 10 15 20
. ,
- 60
- 40
- 20
0 1 2 3 4 5
h
> c .- .- c Y
Time (min) Haemoglobin (pglassay) Fig 1. Time-course (A) and dose dependency (B) of inhibition of [y-32P]GTP-hydrolysis of rus p21 by haemoglobin. (A) [32P]GTP-bound rm p21 (active 2 pmol) was incubated for various periods of time as described in Materials and Methods without any addition (0) or with 1.2 pg of haemoglobin (0). The results were plotted as a percentage relative to the value for the samples at 0 min. The values at 0 min were 8400 and 8700 cprn for no addition and haemoglobin, respectively. (B) [y-32P]GTP-bound ras p21 (active 2 pmol) was incubated for 20 min with indicated doses of haemoglobin (0) as described in Materials and Methods. The inhibition of [y-32P]GTP-hydrolysis was calculated from the filter binding assay as follows: Z =the binding of t3*PIGTP at O r n i n (9000 and 9200 cpm for control and haemoglobin, respectively), Y = the binding after 20 min at 30°C without any addition, X = the binding after incubation for 20min at 30 "C with indicated amounts of haemoglobin; percentage inhibition = [(X-Y)/(Z-y)] x 100%. Each point is the mean of three independent experiments. Bars indicate SD.
100
- 50 8 W
z m n a
3 0
F Y
I I I I I 10 0 5 10 15 20
Time (min)
100
80
60
40
20
0 1 1 1 1 1 1
0 1 2 3 4 5 Haemoglobin @@assay)
Fig 2. Time-course (A) and dose dependency (B) of the effects of haemoglobin on the [3H]GDP-dissociation of rus p21. (A) Time-course study of the effect of haemoglobin on the [3H]GDP-dissociation of rus p21 (active 2 pmol) was assayed in the absence (0). or presence of 4.0 pg of haemoglobin (0) as described in Materials and Methods. The radioactivity was expressed as a percentage relative to the value at 0 min. The values at 0 min for no addition and haemoglobin were 19 200 and 19 600dpm, respectively. (B) [3H]GDP-bound rus p21 (active 2 pmol) was incubated for 20min with indicated doses of haemoglobin (0) as described in Materials and Methods. The values at Omin for no addition and haemoglobin were 19 600 and 19 200 dpm, respectively. The inhibition of [3H]GDP-dissociation was calculated by the same way as that used in Fig 1B and expressed as a percentage relative to the value at Omin. Each point is the mean of three independent experiments. Bars indicate SD.
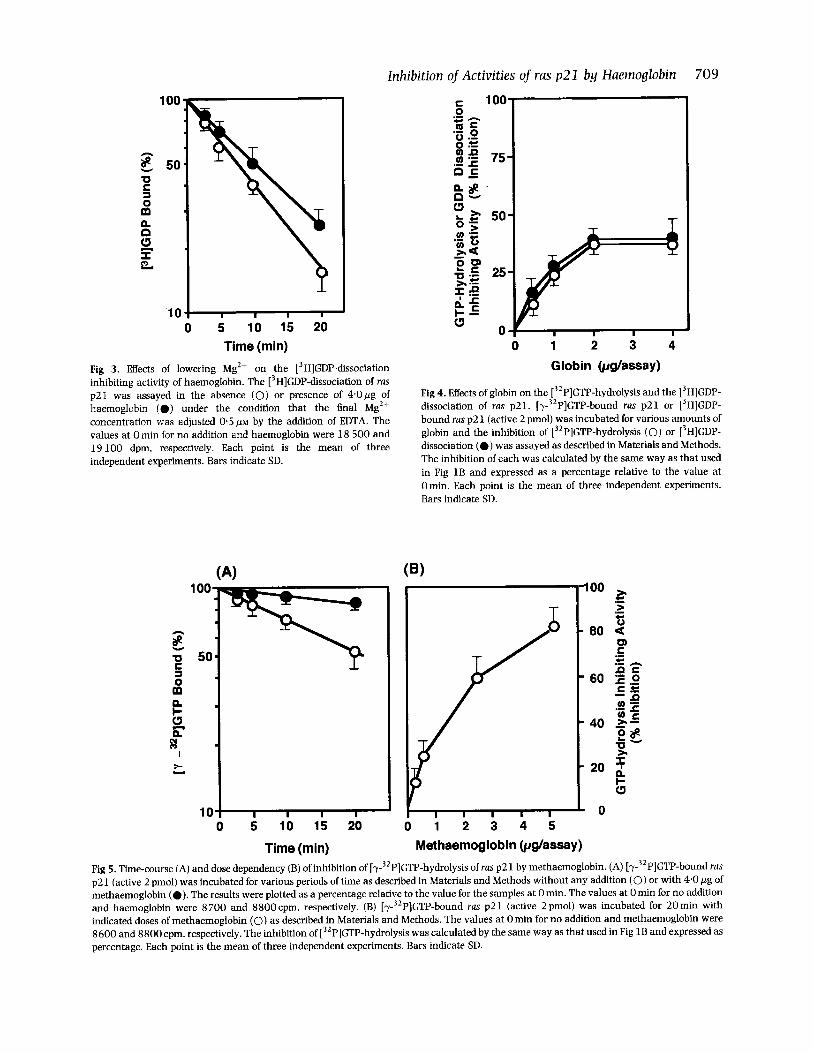
Inhibition of Activities of ras p 2 1 by Haemoglobin 709
100
h
50 U S 3 0 m n n 2 I 0
10 0 5 10 15 20
Time (min) Fig 3. Eflects of lowering Mg2+ on the [3H]GDP-dissociation inhibiting activity of haemoglobin. The [3H]GDP-dissociation of rus p21 was assayed in the absence (0) or presence of 4.0pg of haemoglobin (0) under the condition that the final Mg2+ concentration was adjusted 0.5 ~ L M by the addition of EDTA. The values at 0 min for no addition and haemoglobrn were 18 500 and 19100 dpm, respectively. Each point is the mean of three independent experiments. Bars indicate SD.
(A) 100
E u 50 a m t $z
S
0
P
I 2-
#
I
1c 0 5 110 15 20
Time (min)
0 1 2 3 4
Globin (pglassay)
Fig 4. Effects of globin on the [32P]GTP-hydrolysis and the 13H]GDP- dissociation of rus p21. [y-32P]GTP-bound rus p21 or [3H]GDP- bound rus p21 (active 2 pmol) was incubated for various amounts of globin and the inhibition of [32P]GTP-hydrolysis (0) or ['HIGDP- dissociation (0) was assayed as described in Materials and Methods. The inhibition of each was calculated by the same way as that used in Fig 1 B and expressed as a percentage relative to the value at Omin. Each point is the mean of three independent experiments. Bars indicate SD.
(6)
T
0 1 2 3 4 5 Methaemoglobin (pglassay)
Fig 5. Time-course (A) and dose dependency (B) of inhibition of [y-32P]GTP-hydrolysis of rus p21 by rnethaemoglobin. (A) [y-32P]GTP-bound rus p21 (active 2 pmol) was incubated for vario'us periods of time as described in Materials and Methods without any addition (0) or with 4.0pg of methaemoglobin (0). The results were plotted as a percentage relative to the value for the samples at 0 min. The values at 0 min for no addition and haemoglobrn were 8700 and 8800cpm. respectively. (B) [y-32P]GTP-bound ras p21 (active 2pmol) was incubated for 20min with indicated doses of methaemoglobin (0) as described in Materials and Methods. The values at 0 min for no addition and methaemoglobin were 8600 and 8800 cpm. respectively. The inhibition of [32P]GTP-hydrolysis was calculated by the same way as that used in Fig 1B and expressed as percentage. Each point is the mean of three independent experiments. Bars indicate SD.

710 on [3H]GDP-dissociation of rus p21 was examined. [3H]GDP was slowly dissociated with a rate of 0.035 min-' from ras p21 in a time-dependent manner (Fig 2A). Haemoglobin (4.0 pg of protein per an assay) reduced the dissociation rate to 0.003 min-' as shown in Fig2A. Dose-dependent effect on [3H]GDP-dissociation was also demonstrated in Fig 2B. These results indicate that haemoglobin could function in the presence of physiological concentration of Mg2+. It is of note that the inhibition of haemoglobin on [3H]GDP- dissociation was also observed when the Mg2+ concentra- tion in the reaction mixture was decreased to 0.5 p ~ , under which condition the rate of [3H]GDP-dissociation increased 3-fold (Fig 3). When human haemoglobin was used instead of rat haemoglobin, almost same affects were observed (data not shown).
Koh-Ichi Nagata and Yoshinori Nozawa
Effects of globin or haemin on [32P]Gl'P-hydrolysis and ['HIGDP-dissociation of ras p 2 1 The effects of globin and haemin on [32GTP-hydrolysis or [ 'HIGDP dissociation were examined. Globin prepared from human haemoglobin (Ascoli et al, 1981) was added to the reaction mixture of GTP-hydrolysis or GDP dissociation assay instead of haemoglobin. Weak inhibition (maximal 40%) was observed as shown in Fig 4. Haemin chloride did not inhibit the two activities of rus p21 when its final concentration was adjusted to 1 0 p ~ (data not shown).
Efject of methaemoglobin on [32P]GTP-hydrolysis of rus p 2 1 Methaemoglobin inhibited [32P]GTP-hydrolysis activity of ras p21 time- and dose-dependently in a similar manner to that by haemoglobin (Fig 5). Methaemoglobin also inhibited [3H]GDP dissociation activity of rus p21 time- and dose- dependently like haemoglobin (data not shown).
DISCUSSION
The major function of erythrocytes is well known to be to deliver oxygen to the tissues, and to facilitate the disposal of carbon dioxide and protons formed by tissue metabolism. Therefore, the erythrocyte has a much simpler structure compared to most of mammalian cells. However, the erythrocyte is not metabolically inert, because it synthesizes ATP from glycolysis and contains many other signal transduction components such as G-proteins (Gi and Gs) (Codina et a/. 1984). adenylate cyclase (Kaiser et al, 1974). cyclicAMP-dependent protein kinase (Rubin et ul, 19 72), protein kinase C (Horne et ul, 1985), calmodulin-binding protein (Gardner & Bennett, 1986), unidentified protein tyrosine kinase (Clari & Moret, 1989), and glucose transporter. As for the low M, GTP-binding proteins, rus p21 and unidentified 22kD protein have been reported (Ikeda et ul. 1988). Since haemoglobin forms about 95% of the intracellular protein of the erythrocyte, inhibition of ras p21 activities in the cells may have some importance in physiological condition. It is possible that the function(s) of rus p21, positive control of growth and differentiation, is suppressed in matured erythrocytes, although rus p2 1
should have an important role during the generation of erythrocytes.
In the present study we offered evidence that haemoglobin could serve as a negative regulator of rus p21 in human erythrocytes. One clue in the present study was detection and purification of regulatory protein(s) for ras p21. When various rat organs were screened, spleen cytosol showed the strong inhibiting activity for the intrinsic GTP/GDP- exchange and GTP-hydrolysis activities of rus p21. Accord- ingly, we have purified the inhibitory factor from rat spleen cytosol by measuring the inhibition of ras p2 1-GTP- hydrolysis activity, by using successive column chromato- graphies of anion exchanger DE52 (Whatman Inc., Maidstone, U.K.). hydroxyapatite HCA-100s (Tosoh Inc.)., MonoQ HR5f5 (Pharmacia Inc.) and MonoS HR5f5 (Pharmacia Inc.). The h a 1 preparation obtained from MonoS column yielded a single protein band with M, of 1 4 500 on SDS-polyacrylamide gel electrophoresis (Laemmli, 1970) (data not shown). By its amino acid sequence analysis, it was identified as haemoglobin A (data not shown). We then compared the effects of rat and human haemoglobins on the activities of ras p21.
Haemoglobin does not belong to any category of regulatory proteins of low M, GTP-binding protein, because it exerts inhibitory activity for both GTP-hydrolysis and GDP-dissociation. It is probable that haemoglobin would function in erythrocytes, because it exhibited both GDP- dissociation inhibiting and GTP-hydrolysis inhibiting activ- ities in the presence of physiological concentrations of Mg2+ (Figs I and 2). Moreover, it should be noted that the subunit structure of haemogIobin is crucial for the inhibitory effects, because globin showed maximally 40% inhibition for the [32P]GTP-hydrolysis and [3H]GDP dissociation activities of ras p2 1, but haemin had no effect (Fig 4). Methaemoglobin also inhibited these two activities of rus p21, supporting the hypothesis that the ternary tetrameric structure is important for inhibition. The molecular mechanism of the inhibiting activity of haemoglobin has not been determined, but is possibly similar, if not the same, as that of rubGDI (Takai et al, 1992), because the GDP dissociation inhibiting activity could be observed in the presence of a ~ L M order of MgZf as well as rabGDI (Fig 3).
The inhibitory activities of haemoglobin on GTP-hydro- lysis and GDP-dissociation have been observed in another low M, GTP-binding protein, rum p25 (Nagata et ul, 1990, 1992b) as well as ras p21 (data not shown). These results strongly suggest that some other, if not all, low M, GTP- binding proteins are negatively regulated by haemoglobin in erythrocytes.
Among regulatory proteins for some low M, GTP-binding proteins such as rhoGDI and rhoGEF, the C-terminal domain and/or post-translational prenylation at the C-terminus is essential (Takai et a/, 1992), and they may interact with the terminal domain of the low M, GTP-binding protein. On the other hand, haemoglobin exhibited similar activities for bacterially produced ras p2 1, which is not post-translation- ally modified at the C-terminal region. Therefore the C- terminal region of ras p21 may not be essential for the action of haemoglobin as is rumGDI (Nagata et af, 1992a).

Inhibition of Activities of ras p2 1 by Haemoglobin 71 1 ACKNOWLEDGMENT
This work was supported by the research grant from the Ministry of Education, Science and Culture of Japan.
REFERENCES
Ascoli, F., Rosalia. M., Fanelli, R.R. & Antonini. E. (1981) Preparation and properties of apohemoglobin and reconstituted hemoglobin. Methods in Enzymology, 76, ‘72-87.
Barbacid, M. (1987) ras Genes. Annual Review of Biochemistry. 56,
Boguski, M.S. & McCormick, F. (1993) Proteins regulating Ras and its relatives. Nature, 366, 643-655.
Botwell. D., Fu, P., Simon, M. & Senior, P. “1992) Identification of murine homologues of the Drosophila Son of sevenless gene: potential activators of ras. Proceedings of the National Academy of Sciences o f the United States of America. 89, 6511-6515.
Bradford, M.M. (1976) A rapid and simple method for the quantiation of quantities of protein utilizing the principle of protein-dye binding. Analytical Biochemistry, 72, 248-254.
Clari, G. & Moret, V. (1989) Comparative characterization of membrane-associated and cytosolic Tyr-protein kinases in human erythrocytes. European Journal of Biochemistry, 179,
Codina, J., Hildebrandt, J.D.. Sekura, R.D., Bil-nbaumer, M., Bryan, J., Manclark, C.R.. Iyengar, R. & Brnbaumer, L. (1984) Ns and Ni, the stimulatory and inhibitory regulatory components of adenyl cyclases: purification of the human erythrocyte proteins without the use of activating regulatory ligancls. Journal of Biologicat Chemistry, 259, 5871-5886.
Gardner. K . & Bennett, V. (1986) A new erythrocyte membrane- associated protein with calmodulin binding activity: identification and purification. JournalofBiologicaZChemistry. 261,1339-1348.
Hall, A. (1992) Signal transduction through smalI GTPases: a tale of two CAE’s. Cell. 69, 389-391.
Horne. W.C.. Leto, L.T. & Marchesi, V.T. (1985) Differential phosphorylation of multiple sites in 4.1 and protein 4.9 by phorbol ester-activated and cyclic AMP-dependent protein kinases. Journal of Biological Chemistry, 260, 9073-9066.
Hoshino. M., Clanton. D.J.. Shih, T.Y.. Kawakita, M. & Hattori, S. (1987) Interaction of ras oncogene product p21 with guanine nucleotides. Journal of Biochemistry, 102, 503-511.
Ikeda. K., Kikuchi. A. & Takai, Y. (1988) !$mall molecular weight GTP-binding proteins in human erythryocyte ghosts. Biochemical and Biophysical Research Communications. 156, 889-89 7.
Kaiser. G., Quiring. K., Gauger, D., Palm, D., Recker, H. & Schoeppe,
779-827.
581-588.
W. (1974) Occurrence of adenyl cyclase activity in human erythrocytes Blut. 29, 115-122.
Kaziro. Y., Itoh. H., Kozasa. T., Nakafuku, M. & Satoh, T. (1991) Structure and function of signal-transducing GTP-binding proteins. Annual Review of Biochemistry. 60, 349-400.
Laemmli, U.K.( 1970) Cleavage of structural proteins during the assembly of the head ofbacteriophageT4. Nature, 227,680-685.
McCormick, F. (1989) ras GTPase activating protein: signal transmitter and signal terminator. Cell. 56, 5-8.
Nagata, K.. Itoh, H.. Katada, T.. Takenaka. K.. Ui, M., Kaziro, Y. & Nozawa, Y. (1989) P d c a t i o n , identiJication, andcharacterization of two GTP-binding proteins with molecular weights of 25,000 and 21.000 in human platelet cytosol: one is the rqpl/smg21/Krev-l protein and the other is a novel GTP-binding protein. Journal of Biological Chemistry. 264, 17000-1 7005.
Nagata, K. & Nozawa. Y. (1988) Purification and characterization of two GTP-binding proteins of 22 kDa from human platelet membranes. FEBS Letters. 237, 11 3-1 17.
Nagata, K., Satoh. T.. Itoh, H.. Kozasa. T.. Okano. Y.. Doi. T., Kaziro. Y. & Nozawa, Y. (1990) The ram: a novel low molecular weight GTP-binding protein cDNA from a rat megakaryocyte library. FEBS Letters, 275, 29-32.
Nagata. K.. Suzuki, T., Okano, Y. & Nozawa, Y. (1992a) Presence and some characterization of GDP dissociation inhibitors for a low M, GTP-binding protein, ram p25, in rat spleen cytosol. Life Sciences, 50, 1137-1142.
Nagata, K.. Suzuki. T., Shibagaki, Y.. Mizumoto. K.. Okano. Y.. Kaziro, Y. & Nozawa, Y. (1992b) Characterization and site- directed mutagenesis of a low Mr GTP-binding protein, ram p25. expressed in Escherichia coli. Journal of Biological Chemistry. 267,
Pfeffer, S.R. (1992) GTP-binding proteins in intracellular transport. Trends in Cell Biology, 2.41-46.
Rubin, C.S.. Erlichman, J. & Rosen, O.M. (1972) Cyclic adenosine 3’, 5’-monophosphate-dependent protein kinase of human ery- throcyte membranes. Journal of Biofogical Chemistry, 247, 6 13 5- 6139.
Satoh, T., Nakafuku. M. & Kaziro, Y. (1992) Function of Ras as a molecular switch in signal transduction. Journat of Biofogical Chemistry. 267, 24149-241 52.
Shou. C.. Farnsworth. C.L.. Neel, B.G. &Faig. L.A. (1992) Molecular cloning of cDNAs encoding a guanine nucleotide-releasing factor for Ras p21. Nature, 358, 351-354.
Takai. Y., Kaibuchi, K., Kikuchi. A. & Kawata. M. (1992) Small GTP- binding protein. International Review of Cytology. 133. 187-230.
Tsai, M-H., Yu. C.-L. & Stacey. D.W. (1990) A cytoplasmic protein inhibits the GTPase activity of H-ras in a phospholipid-dependent manner. Science, 16, 982-985.
16000-16006.