haploid
TRANSCRIPT

PRODUCTION AND UTILIZATION PRODUCTION AND UTILIZATION OF DOUBLE HAPLOIDS IN OF DOUBLE HAPLOIDS IN
FLOWER CROPSFLOWER CROPS
K.Ravindra KumarRoll No. 10461
Division of Floriculture & Landscaping

Introduction Haploids - defined as the sporophyte plants that contain a gametic chromosome number (n). Monohaploid: e.g. Rose n = x = 7 Polyhaploid: e.g. French Marigold n = 2x = 24

The history of DHs began with the observation of natural sporophytic haploid in Datura stramonium L. reported by Bergner in 1921.
This was followed by similar discoveries in other plant species e.g. Nicotiana tabacum and Triticum compactum
First report on haploid production was published by Blakeslee et al. (1922) in Datura stramonium.
Guha and Maheshwari (1964, 1966) developed an in vitro anther culture technique for the production of haploid Datura innoxia plants.
Haploids were reported in many other species, i.e. barley (Kasha and Kao, 1970), tobacco (Burk et al., 1979), rice, maize, brassica.
History

At present DHs production via anther culture has been reported in more than 250 plant species belonging to 100 genera and 40 families. (V.K.Misra et al., 2014)
The first in vitro induced haploid plants of female tissues origin was achieved by San Noeum (1976) in barley.
Commercial varieties developed through DH protocols - reported for many crops and more than 290 varieties have already been released (Ferrie and Caswell, 2011).
Compared to agronomic species, there has been very little work on DH in ornamental species despite the tremendous potential benefits.

Methods to Induce DHs In vivo occurrence of haploid plants
Spontaneous haploidsHybridization
a) Intraspecific hybridization b) Wide hybridization
ParthenogenesisIn vitro Methods
Haploids from male gametes a) Anther culture b) Pollen/Microspore culture
Haploids from female gametes a) Ovary slice culture b) Ovule culture

Spontaneous HaploidsNaturally occurring haploids in 71 species, representing 39 genera in 16 families of angiosperms. e.g. agropyron, alfalfa, citrus, peach and trillium
Gymnosperms (Pohlheim, F. 1968) e.g. Unique haploid from of Thuja plicata called Thuja gigantea
‘gracilis’ Beissn
But the exact source of haploids in these species, in terms of their embryological origin, is often unknown (Dunwell 2010)
Cultivar development has utilized these naturally occurring haploids.
e.g. ‘Kleine Liebling’ is a haploid cultivar of Pelargonium (Darker
1966), ‘Marglobe’ is a DH cultivar of tomato (Morrison 1932)
The frequency of naturally occurring haploids is generally low, consequently, this is not an efficient method for breeding.

Haploids through hybridization Intraspecific hybridization
Observed generation of haploid from diploid material (Haplopappus), tetraploid material (Pathenium, Sorghum, Sisymbrium and alfalfa) allopolyploid (Aegilotriticum)
In citrus haploid progeny observed in the interploidy cross between diploid and triploids (Oiyama et.al. 1993)
Haploids of sugar beet were identified by crossing diploid male sterile plants with green hypocotyls and tetraploid fodder beets homozygous for red hypocotyls (Bosemark 1971)
There is a new method to produce haploid from the seed of plants by manipulating a centromere-specific histone CENH3. When the cenh3 mutant expressing abnormal CENH3 is crossed to the wild type, chromosomes from the mutant are eliminated, and then eventually the haploids are produced. This method could be applied to any plant because the CENH3 is universal in eukaryotes (Ravi and Chan, 2010)

In vivo DH induction in Maize
CIMMYT
Step 1: Induction Cross Step 2: Haploid kernel identification with the embryo & endosperm marker (R1-nj)
Step 3: Artificial genome doubling 0.06% Colchicine, 0.5% DMSO solution for 8 h
Step 4: Recovery of seeds from D0 plants
Pollination with pollen of the same species (e.g. maize) No in vitro culture is needed, since kernels containing haploid embryos display a normal
germination rate and lead to viable haploid seedlings.Haploid embryos can be selected early in the breeding process, based on morphological
and physiological markers (Dominantly inherited purple coloration from inducer line).

Wide HybridizationInterspecific or intergeneric crosses in which the pollinator chromosomes are eliminated have been used successfully to produce maternal haploids.
1: Barley DHsHordeum vulgare (cultivated) x H. bulbosum (wild )
(2n=2x=14) (female) (2n=2x=14) (male)Kasha, K.J. and Kao, K.N., 1970
2 : Wheat , Triticale, Rye, Oat Wheat (Triticum aestivum) x Maize (Zea mays L.)Triticale (Triticosecale) x Maize (Z. mays L.)Rye (Secale cereale) x Maize (Z. mays L.)Oat (Avena sativa) x Maize (Z. may s L.)
Palmer, et. al., 2005
Embryo rescue is a necessary part of this technique as the endosperm does not develop and therefore cannot provide nutrients for continued development of the embryo

ParthenogenesisIn this method the egg cell developes into an embryo without fertilization by the sperm nucleus.This can be achieved by pollination with irradiated pollen or with the addition of chemicals (Khush et. al., 1996)There is generally a low frequency of haploid recovery and therefore this method is not widely used for breeding purpose.
Ferrie Alison et.al. 2011

ANDROGENESIS
Androgenesis is defined as the process of embryo development from the male gametophytes (i.e. microspores or anthers), with the subsequent regeneration of haploid and doubled haploid plants from these cells.

Half anther culture in Anthurium
PCTO, Budi Winarto et.al., 2012

Microspore culture – Zantedeschia aethiopica
80 pollen grains/dropMedium
MicrosporesFilter paper
Anthers
Pollen in hanging dropsIsolated microspore culture
Afri. J. Bio., Shimin Wang et.al., 2011

Gynogenesis is the culture of unfertilized female gametophytes (i.e. ovules, ovaries).This method is used when plants do not respond to androgenic methods, there is a problem with regenerating albino plants from anther culture, or the donor plants are male sterileGynogenesis is usually less efficient than androgenesis.Used in plant families that do not respond to androgenesis
- Liliaceae - Compositae
GYNOGENESIS

Gynogenesis – Spathiphyllum wallisii
PCTO, Tom Eeckhaut et.al., 2001

Factors affecting haploid inductionDonor plant genotypePhysiological condition of donor plants & Pre-treatmentCulture medium compositionPhysical factors during tissue culture (light, temperature)Organic addenda/sugar/sugar alcoholDevelopmental stage of gametes, microspores and ovules.Other miscellaneous factors.

Stage and alternatives of microspores
J.Exp.Bot. Jose et.al., 2007

In vitro generated haploids in ornamental crops
POP, Ferrie et.al.,2011

Identification of HaploidsMorphological observation
One year old seedlings of Dendrocalamus latiflorus a, C & d Anther regenerated plants of 3x, 6x and 12x b) Seedling sample 6x In vitro cell Dev. Biol. Guirong Qiao et al.,2013
Differences of leaf morphology a) Haploid, B) Diploid and C) Triploid in Anthurium andreanum cv. ‘Tropical ‘ PCTO, Winarto et.al., 2011
Acta.Hort. Gao et.al., 2011

Identification of Haploids cont.Chloroplast number in stomatal guard cells
Anthurium
Chrysanthemum
Haploid, Tetraploid (4x) and Pentaploid (5x)
D) Haploid cell with 19, E) Diploid cell with 30, F) Triploid cell with 52 chloroplasts

Identification of Haploids cont.Chromosomal count
& Flow cytometry
Determination of ploidy level of regenerated plants of Popular by flow cytometry (a-c) and chromosome counting (d-f).
a and d plant displaying a haploid histogram (2n=x=19)
b and e plant displaying a diploid histogram (2n=2x=38)
c and f plant displaying a triploid histogram (2n=3x=57)
PCTO, Ying Li., et. al., 2013

Identification of Haploids cont.DNA finger printing
Anemone coronariaZantedeschia aethiopica
A section of AFLP banding patterns:Lanes 1-4 are from 4 diploid plants from anther culture, and 5-8 from 4 individual donor plants. Arrows indicate polymorphic bandsScien. Hort. Xiyan Zhang et.al.,2011
RAPD profiles generated by primers OPA18, OPA4, OPA17 and OPA18 from DNA of the donor plant (DP) and three regenerants (AP) of the family A,B and C, respectively, M: ᶲx174 DNA (Hae III fragment)Plant Breed. Laura et.al., 2006

Double haploids in
Crop Improvemen
t

Rapid technique homozygous plants can be achieved in one generation
(inbred lines)Production of homozygous lines of the cross pollinating species and hybrids are highly desirable
(to avoid inbreeding depression)The conventional method of inbreeding is impractical for self-incompatible, male sterile and tree species.Protocols already developed for Albizza lebbek, Azadirachta indica, Cassia siamea, Ceratonia siliqua, Hevea brasiliensis, Peltophorum pterocarpum, Populus sp.
Biotech. Advances. Priyanka et.al., 2008

Shortens the breeding cycle
Normally, in a hybridization programme evaluation of lines is possible only after 4-5 years of pedigree breeding and it takes another 4-5 years to release a new variety.
By anther culture of F1 hybrids the various genotypes of gametes can be fixed and evaluated in the first generation
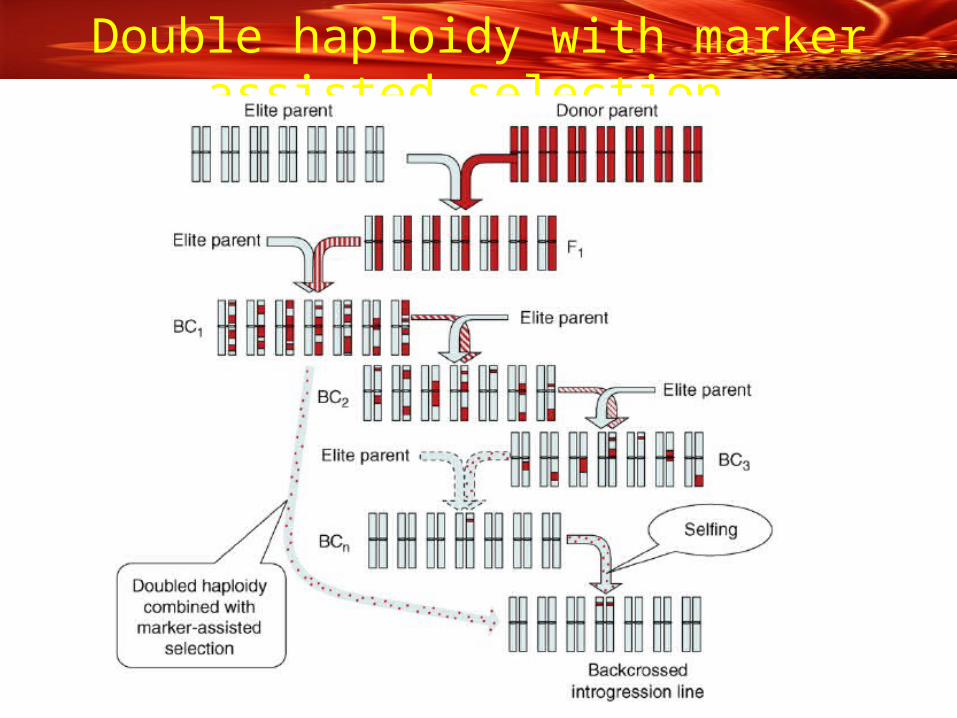
Double haploidy with marker assisted selection

New plant breeding techniques (NPBT) Homozygous parental lines of a selected heterozygous plant are reproduced.The genes involved in the meiotic recombination process are silenced through transgenesis.Consequently, nonrecombined haploid lines are obtained from the heterozygous plant and their chromosomes are doubled through the double-haploid technique.The doubled haploids obtained are screened to find a pair that, would reconstitute the original heterozygous plants.Only nontransgenic plants are selected, thus the offspring of the selected parental lines would not carry any additional genomic charge.

Recessive mutants Haploids are extremely useful for detecting recessive mutants which may not express themselves in the heterozygous diploid background and therefore can be easily lost.
Different spate colors produced from flowered plants derived from anther culture of Anthurium. a) spate and spadix color of A.andreanum cv. Tropical as the donor plant. B-h Spate and spadix color variation of regenerants

Other advantages of Double Haploids (DH)
DHs can be represent as new variety (self-pollinated crops)
Induction of mutations
DH lines are also valuable tools in marker-trait association studies,
molecular marker-assisted or genomic selection – based breeding and
functional genomics
Useful in cytogenetic research
Genetic engineering
Useful in development of chromosome substitution lines.
QTL mapping, genetic and physical mapping

Objective:
To develop comprehensive anther culture method for double haploid production in Primula.
Case study - 1

IntroductionPrimula is an ornamental plant popular for its early spring blooms and diversity of colors.
The species exhibits desirable horticultural traits such as long flowering season, plentiful flowers and a pleasant fragrance.
Producing homozygous lines by conventional methods in this species is time consuming and difficult, because the species is self-incompatible.
Anther culture has been successfully applied in many field crops, but its use is still limited in ornamentals and not yet reported in Primula.
In this study special attention was given to several factors influencing androgenesis, such as the correlation of bud morphology and size, microspore developmental stages, plant growth regulator concentrations.

Materials and MethodsPlant material : 200 glass house raised Baby prime rose plants were used as anther donor plants Determination of microspore development stageThe relationship between the flower bud morphology, bud size, microspore developmental stage and viability was determined firstly.Preparation of induction media and induction of callusPreparation of differentiation media and plant RegenerationPlantlet rooting and transplantation
Abbreviated
number
PGR (mg/l) No. of anthers cultured
BAP 2,4-D
I1 1.0 0.5 603
I2 1.0 1.0 600
I3 1.0 1.5 596
I4 1.0 2.0 598
I5 2.0 0.5 600
I6 2.0 1.0 595
I7 2.0 1.5 605
I8 2.0 2.0 602
Abbreviated
number
PGR (mg/l) No. of calli cultured
BAP NAA
D1 0.1 0.01 60
D2 0.2 0.01 58
D3 0.5 0.01 56
D4 1.0 0.1 57
D5 1.0 0.2 60
D6 1.0 0.5 56
D7 2.0 0.1 58
D8 2.0 0.2 60
D9 2.0 0.5 59
Abbreviated number
PGR (mg/l)
IBA
R1 0.0
R2 0.05
R3 0.1
R4 0.2

i) Flow cytometry : Leaf samples of in vitro-grown plants derived from anther culture
were collected And added to Lysis buffer to release the nuclei. The extract was stained with 2 µl DAPI. Analysis of nuclei was conducted using a Partec CA II flow
cytometer. Tissue from a diploid plant of P.forbesii was used as an internal
standard. The ploidy level of each plant was determined based on a
comparison between the fluorescence peak value of the tested plants with that of the diploid control.
ii) Cytological analysis: Actively growing root tips (5-8) approximately 2 mm in length
were used for cytological analysis Samples were stained with Carbol fuchsin solution and observed
under microscope (100 x)
Determination of ploidy level

Results:

Callus induction of P.forbesii via anther culture

Shoot regeneration of P.forbesii via anther cultureShoot regeneration of P.forbesii via anther culture

Comparison of regenerated plants with different ploidy levels in cultureComparison of regenerated plants with different ploidy levels in cultureMorphological Flow cytometry histograms Chromosomal count


Inference
Identified buds of 4.0 – 5.0 mm long for in vitro culturing.The callus induction rate was significantly high in the media containing 1.0 mg/l BAP and 0.5 mg/l 2,4 – D.Adventitious buds were observed only on shoot induction medium with 0.2 mg/l BAP and 0.01 mg/l NAA.MS without PGRs was suitable for rooting of baby primrose plantlets.
Regenerated plants in this study showed multiple ploidy levels viz. haploid, diploid, triploid, tetraploid, hexaploid and mixoploid. Only 2% of plants observed as haploids.
Reasons might be spontaneous, repeated chromosome doubling to generate polyploid plants include nuclear fusion during early microspore divisions, endomitosis, endoreduplication or multipolar mitosis during the callus phase.

Case study - 2
Objective:1.To study the effect of cold pre-treatment on unfertilized ovule culture.2.To investigate the ability of embryo like structure (ELS) production through gynogenesis and reproducibility of the haploids and DHs through this technique.

Two Japanese Gentian species, G.triflora and G.scabra are one of the most important plants for cut-flower and pot plant use in Japan.Production of homozygous lines are indispensable for F1 hybrid breeding, but it is difficult In this species owing to their intense inbreeding depression.Efficiency of embryogenesis in anther culture was quite low and large genotypic variation was observed (Mostly triploids). More over this method is not effective for G.scabra.Unfertilized ovule culture has more advantageous than anther culture from the point of view of embryogenic efficiency, limited influence of donor plant genotype and a high frequency of haploids production.Cold pretreatment of flower buds or inflorescences before culture has been reported to enhance the frequency of embryogenesis in several species.
Introduction

Plant materials: 43 genotypes of gentians consisted of 12 cultivars and 31 lines of G.triflora (2n = 26), G.scabra (2n = 26), G.triflora var. japonica f. montana (2n = 26) and their hybrids.Cold pretreatment: Four genotypes (G.triflora cv. Ashiro-no-Aki, G.scabra line 17-260, G.triflora x G.scabra lines 17-386 and 17-260) were tested.Anthers removed from each flower, inflorescences were stored at low temperature (40C) for 3,7 and 14 days in dark.Unfertilized ovule culture and plant regeneration: Ovules excised from a pistil were cultured in the medium of ½ NLN containing 0.8 % agar-solidified and 10 % sucrose.The ELSs developed from ovules were transferred to modified agar (1.0%) – solidified MS medium with concentration of major salts reduced by 50% (1/2 MS) and supplemented with 3% sucrose and 1.0 mg/l GA3.
Materials and Methods

Flow cytometry:Young leaf samples were analyzed by using Partec CyFlow PA and Cell Lab Quanta SC.Chromosome doubling treatment:Chromosome doubling of haploids was performed as described by Morgan et al. (2003). Shoots containing axillary buds were cut and subcultured to propagation medium (Pr) containing 50 µM oryzalin for 1,2,3 and 4 weeks and elongated axillary shoots were transferred to Propagation medium without BA after 6 weeks.
a) Unfertilized ovules in an ovary. b) Ovules cultured on ½ NLN soild medium c) ELSs emerged from ovules after 50 days of culture d) A magnified ELS in c plate
Determination of ploidy level

Cold pretreatment produced more ELSs than non-treatment in 3 genotypes except for line 17-386. In particular 7 and 14 days tended to exhibit a higher response.
e, f : A plantlet regenerated from ELS
Results



W14 and W15 genes are allelic and encode proteins related to the α/β hydrolase fold superfamily.Donor plants of 17-386 and 17-488 carried both alleles W14/W15 (heterozygous), whereas regenerated diploid plants carried one of them (homozygous)On the other hand, strain 17-260 carried only W14 allele (homozygous). Therefore, for strain 17-260, the FLS, which is flavonoid biosynthetic genes, was used.FLS amplified two bands (heterozygous), in the 17-260 strain. When 43 diploid plants were examined, 41 plants showed a single band: however, 2 plants of Go963 and Go990 had the same two bands as the donor plant.
W14 and W15 genes are allelic and encode proteins related to the α/β hydrolase fold superfamily.Donor plants of 17-386 and 17-488 carried both alleles W14/W15 (heterozygous), whereas regenerated diploid plants carried one of them (homozygous)On the other hand, strain 17-260 carried only W14 allele (homozygous). Therefore, for strain 17-260, the FLS, which is flavonoid biosynthetic genes, was used.FLS amplified two bands (heterozygous), in the 17-260 strain. When 43 diploid plants were examined, 41 plants showed a single band: however, 2 plants of Go963 and Go990 had the same two bands as the donor plant.

Cold pretreatment for 1-2 weeks at 40C found to be beneficial
In the present study using 43 genotypes 40 genotypes produced ELSs
Though variations are present, unfertilized ovule culture was affected less by genotypes compared with of anther culture.
A large number of regenerated plantlets, which were obtained through gynogenesis, consisted of haploids (57.8%) and diploids (34.6%).
Oryzalin is effective chemical for chromosomal doubling.
W14/W15 and FLS genes were used as genetic markers in order to confirm whether the diploid plant obtained from unfertilized ovule culture is a DH or not.
Inference

Conclusion
Doubled haploids (DH) technology is highly desirable can be implemented in all environments.Rapid production of homozygous lines.Speed up the breeding by fixing of the elite inbreds.Shortened breeding cycle and cost saving.Elimination of unfavorable genes and enrichment of favorable genes.Improvement of germplasm lines.Early release of commercial hybrids and resource saving.Combination with other tools including precision phenotyping, bioinformatics, genomic selection and modeling can improve the efficiency of the breeding process.

Thank you!