human midbrain dopamine neurons express serotonin 2a receptor: an immunohistochemical demonstration
TRANSCRIPT

Ž .Brain Research 853 2000 377–380www.elsevier.comrlocaterbres
Short communication
Human midbrain dopamine neurons express serotonin 2A receptor: animmunohistochemical demonstration
Keiko Ikemoto a,), Akiyoshi Nishimura b, Nobuo Okado c, Masahiko Mikuni d, Katsuji Nishi b,Ikuko Nagatsu a
a Department of Anatomy, School of Medicine, Fujita Heath UniÕersity, Toyoake, Aichi 470-1192, Japanb Department of Legal Medicine, Shiga UniÕersity of Medical Science, Otsu 520-2192, Japan
c Department of Anatomy, Institute of Basic Medical Science, UniÕersity of Tsukuba, Tsukuba, Ibaraki 305-0018, Japand Department of Neuropsychiatry, Gumma UniÕersity School of Medicine, Maebashi 371-8511, Japan
Accepted 12 October 1999
Abstract
Ž . Ž .We demonstrated intense serotonin 5-HT 2A receptor immunoreactivity in the human ventral tegmental area VTA using by aŽ .recently raised antibody against 5-HT2A receptor. The substantia nigra SN neurons also showed 5-HT2A receptor immunoreactivity.
Ž .Double immunohistochemistry of 5-HT2A receptor and tyrosine hydroxylase TH revealed many neurons doubly labeled by 5-HT2Areceptor and TH in the VTA and SN. It is suggested that activity of human midbrain dopaminergic neurons might be strongly regulatedvia 5-HT2A receptors at the level of their originating nuclei. q 2000 Elsevier Science B.V. All rights reserved.
Keywords: Human; Substantia nigra; Ventral tegmental area; Oculomotor nucleus; Serotonin 2A receptor; Serotonin; Tyrosine hydroxylase; Dopamine;Immunohistochemistry
Ž . Ž .Serotonin 5-HT –dopamine DA interaction via 5-HT2A receptors has been implicated in etiology of psy-choses and therapeutic actions of antipsychoticsw x2,9,15,16,18 . Although there are numerous animal studiesthat show inhibitory actions of 5-HT on DA neuronsw x9,10,16,18 , little has been known in human materialsw x17 . Recently, 5-HT2A receptor immunoreactivity hasbeen reported in a small number of neurons of the rat
Ž . Ž .ventral tegmental area VTA and substantia nigra SNw x1,3 , the originating nuclei of mesolimbic or mesostriataldopaminergic neurons. These 5-HT2A receptor immunore-
Ž .active -ir neurons might be DA neurons. However, con-sidering some species differences in mammalian
w xmonoaminergic neuronal systems 4,5,7,11,13 , expression
) Corresponding author. Fax: q81-562-93-2649; e-mail:[email protected]
of 5-HT2A receptors in the human homologous areasmight be different from that in the rat.
The present study was conducted to elucidate morpho-logical interaction between DA neurons and 5-HT2A re-ceptors in the VTA and SN in the human, with immunohis-tochemical method by using antibodies against 5-HT2A
w x Ž . w xreceptor 3 and tyrosine hydroxylase TH 12 .Human brains were obtained from four autopsied cases
Ž24–47 years old, post-mortem interval to fixation: 4–16.h in Department of Legal Medicine, Shiga University of
Medical Science, Japan. These cases died from naturalcauses and had no known clinically and pathologicallydetectable neurological and psychiatric diseases. Brainswere immediately sliced into 1 cm slabs and immersed in
Ž .the fresh fixative pH 7.4 containing 5% glutaraldehyde orŽ .4% paraformaldehyde in 0.1 M phosphate buffer PB at
48C for 5–72 h. The slices were then transferred to PBcontaining 15% sucrose and 0.1% sodium azide for storageat 48C. The brain sections were made using a cryostat in50 mm thick in coronal planes. The sections were treated
0006-8993r00r$ - see front matter q 2000 Elsevier Science B.V. All rights reserved.Ž .PII: S0006-8993 99 02237-4

( )K. Ikemoto et al.rBrain Research 853 2000 377–380378
with 40% methanol and 1% H O for 20 min to inhibit2 2w xendogenous peroxidase 4 .
The sections of the human brains were incubated inŽantibodies against N-terminal of 5-HT2A receptor pro-
. w xvided by Dr. N. Okado, Japan 3 diluted 1: 200–400 orw xTH 12 diluted 1:10,000–70,000 in PBS containing 0.3%
Triton X-100 at 48C for 1 week. Then they were incubatedin secondary antibodies for 12 h at room temperature;
Žbiotinylated anti-chicken IgG Vector, Burlingame, CA,.USA; 1:1000 was used for the 5-HT2A receptor antibody,
Ž .biotinylated anti-rabbit IgG Vector; 1:1000 for the THantibody. Finally, they were incubated in avidin–biotin–
Ž .peroxidase complex Vector; 1:1000 for 1 h at roomtemperature. Peroxidase activity was then revealed with 50
Ž .mM Tris–HCl buffer pH 7.6 containing 0.0003% H O ,2 2Ž .0.01% 3,3-diaminobenzidine DAB –4 HCl and 1% nickel
Ž .ammonium sulfate DAB–nickel method . Details of theproduction, characterization, and specificity of the 5-HT2Areceptor and TH antisera have been described elsewherew x3,12 . To confirm the specificity of 5-HT2A receptorantiserum, an absorption test was performed. A synthe-sized peptide, corresponding to amino acids 14–33 in the
w xN-terminal extracellular region of rat 5-HT2A receptor 3was used as an antigen of the absorption test. An atlas of
w x w xNieuwenhuys et al. 14 and our previous study 6 wereused to determine the anatomical territories.
For dual labeling of 5-HT2A receptor and TH, thesections were incubated in a mixture of 5-HT2A receptor
w x Ž . w x Žantibody 3 diluted 1:200 and TH antibody 12 diluted.1:10,000 for 1 week. A half of the sections were incu-
bated in biotinylated anti-chicken IgG, followed by incuba-Ž .tion in avidin–biotin–peroxidase complex Vector; 1:1000
for 1 h at room temperature, and peroxidase activity wasŽ .revealed with 50 mM Tris–HCl buffer pH 7.6 containing
0.0015% H O and 0.05% DAB. Then, the sections were2 2
incubated in biotinylated anti-rabbit IgG, followed by incu-Žbation in avidin–biotin–peroxidase complex Vector;
.1:1000 , and peroxidase activity was revealed by DAB–nickel method. The other half of the sections were incu-bated in ultrasmall gold conjugate goat anti-rabbit IgGŽAurion, GAR GP-US, Wageningen, Netherlands; diluted
.1: 100 at 48C overnight, and TH immunoreactivity wasŽvisualized by a silver intensification kit IntenSEe Silver
Enhancement Kit, Amersham, RPN 491, Bucking-.hamshire, England . The sections were incubated in bio-
tinylated anti-chicken IgG, then in avidin–biotin–per-Ž .oxidase complex Vector; 1:1000 , and DAB–nickel
method was applied.
The absorption test resulted in no immunoreactivity instandard immunohistochemical procedures, which con-firmed the specificity of the 5-HT2A receptor antiserum.
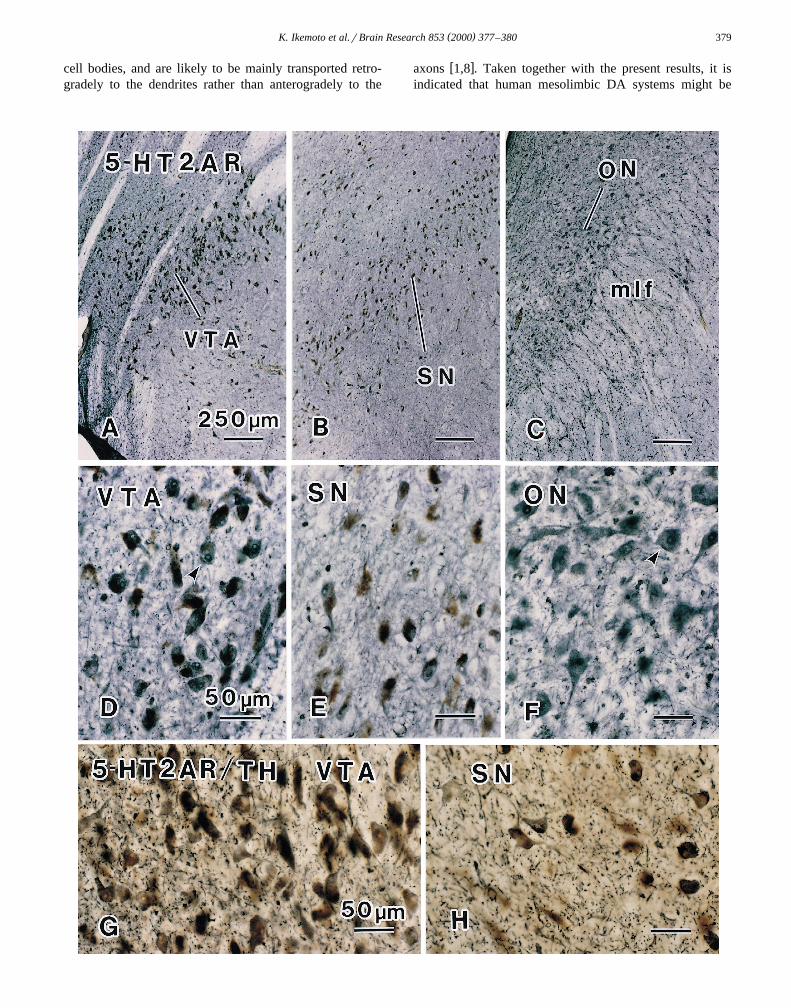
In the VTA, intense 5-HT2A receptor immunoreactivityŽwas observed in neuromelanin-pigmented neurons Fig.
.1A,D . In the SN, neuromelanin-pigmented neurons alsoŽ .showed 5-HT2A receptor immunoreactivity Fig. 1B,E .
The 5-HT2A-ir neurons in the VTA and SN were round,ovoid, bipolar or multipolar in shape and 20–35 mm indiameter. These 5-HT2A receptor-ir neurons possessed one
Ž .to two processes and well-stained nuclei Fig. 1D,E . Thenucleoli of these neurons showed weak immunoreactivityŽ .Fig. 1D,E . In the marginal area of the SN, 5-HT2Areceptor-ir neurons with no neuromelanin pigment wereoccasionally seen. In the neurons of the oculomotor nu-
Ž .cleus ON , intense 5-HT2A receptor immunoreactivityŽ .was observed Fig. 1C . In these 5-HT2A receptor-ir neu-
rons possessed some processes and well-stained nucleiŽ .Fig. 1F . 5-HT2A receptor immunoreactivity was espe-cially intense in the marginal regions of some cell bodies
Ž .of 5-HT2A receptor-ir neurons Fig. 1D,F arrow heads .In double immunohistochemistry for 5-HT2A receptor
and TH, many neurons were doubly stained for 5-HT2AŽ .receptor and TH in the VTA and SN Fig. 1G,H . In the
marginal area of the SN, some neurons were singly stainedŽ .for 5-HT2A receptor data not shown . Such 5-HT2A
receptor-only-ir neurons lacked neuromelanin pigment.The present study showed 5-HT2A receptor-ir neurons
in neuromelanin-pigmented neurons, putative DA neurons,in the human VTA and SN. The fact that 5-HT2A receptorand TH were doubly immunostained in the human VTAand SN is another evidence that human midbrain dopamin-ergic neurons express 5-HT2A receptors. In the rat VTAand SN, 5-HT2A receptor-ir neurons have been described,
w xhowever, the quantity is small 1,3 . In the human homolo-gous area, the density of 5-HT2A receptor-ir neuronsseems to be far higher than that in the rat. The speciesdifference is evident.
On the other hand, the existence of a number of 5-HT2Areceptor-ir neurons is likely to be common in the ON both
w xin the rat 1,3,19 and human.In the present results, 5-HT2A receptor immunoreactiv-
ity was more intense in the neurons in the VTA than thoseŽ .in the SN compare Fig. 1D with E . The VTA is an
originating nucleus of the mesolimbic DA system, whichrelates to motivation, attention and reward. The rat studieshave been shown that 5-HT2A receptors in the midbrainand dorsal raphe nucleus are synthesized in the neuronal
Fig. 1. Photomicrographs of immunostained brain sections obtained by an autopsy of a 35-year-old female. In this case, post-mortem period to fixation wasŽ .16 h, and fixation was performed with 5% glutaraldehyde in 0.1 M PB for 5 h. A–F Single-staining of 5-HT2A receptor in a section through the
Ž . Ž . Ž . Ž .midbrain. A,D The VTA. B,E The SN. C,F The ON. In the VTA and SN, many neuromelanin-pigmented neurons brown show 5-HT2A receptorŽ . Ž . Ž .immunoreactivity purple . The marginal regions of some 5-HT2A receptor-ir neurons are well-stained D,F arrow heads . G,H . Double-staining ofŽ . Ž . Ž . Ž .5-HT2A receptor brown and TH purple . TH-staining was intensified by nickel. G The VTA. H The SN. Many neurons are doubly stained for
5-HT2A receptor and TH. Abbreviations: VTA, ventral tegmental area; SN, substantia nigra; ON, oculomotor nucleus, mlf; medial longitudinal fasciculus.
( )K. Ikemoto et al.rBrain Research 853 2000 377–380 379
cell bodies, and are likely to be mainly transported retro-gradely to the dendrites rather than anterogradely to the
w xaxons 1,8 . Taken together with the present results, it isindicated that human mesolimbic DA systems might be

( )K. Ikemoto et al.rBrain Research 853 2000 377–380380
powerfully regulated by 5-HT via 5-HT2A receptors at thelevel of the midbrain originating nuclei.
Acknowledgements
This study was supported by Grant-in-Aid for ScientificResearch on Priority Areas, Ministry of Education, Sci-
Ž .ence, Sports and Culture of Japan to K.I. C-1 10680713 ,Ž . Ž .A.N., M.M. B 11470200 , K.N. C-2 11670413 and I.N.,
and Fujita Health University in Japan.
References
w x1 V. Cornea-Habert, M. Riad, C. Wu, S.K. Singh, L. Descarries,Cellular and subcellular distribution of the serotonin 5-HT2A recep-tor in the central nervous system of adult rat, J. Comp. Neurol. 409Ž .1999 187–209.
w x2 B. Dean, T. Hussain, W. Hayes, E. Scarr, S. Kitsoulis, C. Hill, K.Opeskin, D.L. Copolov, Changes in serotonin and GABA recep-2A A
tors in schizophrenia: studies on the human dorsolateral prefrontalŽ .cortex, J. Neurochem. 72 1999 1593–1599.
w x3 S. Hamada, K. Senzaki, K. Hamaguchi-Hamada, K. Tabuchi, H.Yamamoto, T. Yamamoto, S. Yoshikawa, H. Okano, N. Okado,Localization of 5-HT2A receptor in rat cerebral cortex and olfactorysystem revealed by immunohistochemistry using two antibodies
Ž .raised in rabbit and chicken, Mol. Brain Res. 54 1998 199–211.w x4 K. Ikemoto, K. Kitahama, A. Jouvet, R. Arai, A. Nishimura, K.
Nishi, I. Nagatsu, Demonstration of L-dopa decarboxylating neuronsŽ .specific to human striatum, Neurosci. Lett. 232 1997 111–114.
w x5 K. Ikemoto, I. Nagatsu, K. Kitahama, A. Jouvet, A. Nishimura, K.Nishi, T. Maeda, R. Arai, A dopamine-synthesizing cell groupdemonstrated in the human basal forebrain by dual labeling immuno-histochemical technique of tyrosine hydroxylase and aromatic L-
Ž .amino acid decarboxylase, Neurosci. Lett. 243 1998 129–132.w x6 K. Ikemoto, I. Nagatsu, A. Nishimura, K. Nishi, R. Arai, Do all of
human midbrain tyrosine hydroxylase neurons synthesize dopamine?,Ž .Brain Res. 805 1998 255–258.
w x7 K. Ikemoto, K. Kitahama, T. Maeda, M. Jouvet, I. Nagatsu, Cholin-
Ž .ergic neurons with monoamine oxidase type B MAOB -activity inthe laterodorsal tegmental nucleus of the mouse, Neurosci. Lett. 271Ž .1999 53–56.
w x8 A. Jansson, B. Tinner, H.W.M. Steinbusch, L.F. Agnati, K. Fuxe,On the relationship of 5-hydroxytryptamine neurons to 5-hydroxy-tryptamine 2A receptor-immunoreactive neuronal processes in thebrain stem of rats. A double immunolabelling analysis, NeuroReportŽ .9 1998 2505–2511.
w x9 S. Kapur, G. Remington, Serotonin–dopamine interaction and itsŽ .relevance to schizophrenia, Am. J. Psychiatry 153 1996 466–476.
w x10 M.D. Kelland, A.S. Freeman, L.A. Chiodo, Serotonergic afferentregulation of the basic physiology and pharmacological responsive-ness of nigrostriatal dopamine neurons, J. Pharmacol. Exp. Ther.
Ž .253 1990 803–811.w x11 L. Kung, M. Force, D.J. Chute, R.C. Roberts, Immunocytochemical
localization of tyrosine hydroxylase in the human striatum: a post-Ž .mortem ultrastructural study, J. Comp. Neurol. 390 1998 52–62.
w x12 I. Nagatsu, Y. Kondo, S. Inagaki, N. Karasawa, T. Kato, T. Nagatsu,Immunofluorescent studies on tyrosine hydroxylase: application for
Ž .its axoplasmatic transport, Acta Histochem. Cytochem. 10 1977494–499.
w x13 S. Nakamura, I. Akiguchi, J. Kimura, A subpopulation of mousestriatal cholinergic neurons show monoamine oxidase activity, Neu-
Ž .rosci. Lett. 161 1993 141–144.w x14 R. Nieuwenhuys, J. Voogd, Chr. van Huijzen, The Human Central
Nervous System, A Synopsis and Atlas, Springer-Verlag, Berlin,1979.
w x15 S. Nyberg, B. Eriksson, G. Oxenstierna, C. Halldin, L. Farde,Suggested minimal effective dose of risperidone based on PET-mea-sured D2 and 5-HT2A receptor occupancy in schizophrenic patients,
Ž .Am. J. Psychiatry 156 1999 869–875.w x16 M.F. O’Neill, C.L. Heron-Maxwell, G. Shaw, 5-HT2 receptor antag-
onism reduces hyperactivity induced by amphetamine, cocaine, andMK-801 but not D1 agonist C-APB, Pharmacol. Biochem. Behav.
Ž .63 1999 237–243.w x17 A. Pazos, A. Probst, J.M. Palacios, Serotonin receptors in the human
brain: IV. Autoradiographic mapping of serotonin-2 receptors, Neu-Ž .roscience 21 1987 123–139.
w x18 L. Ugedo, J. Grenhoff, T.H. Svensson, Ritanserin, a 5-HT receptor2
antagonist, activates midbrain dopamine neurons by blocking sero-Ž .tonergic inhibition, Psychopharmacology 98 1989 45–50.
w x19 C. Wu, E.J. Yoder, J. Shih, K. Chen, P. Dias, L. Shi, X.-D. Ji, J.Wei, J.M. Conner, S. Kumar, M.H. Ellisman, S.K. Singh, Develop-ment and characterization of monoclonal antibodies specific to the
Ž .serotonin 5-HT receptor, J. Histochem. Cytochem. 46 19982A
811–824.