iai.asm.org · 16 extended n-terminal a -helix and a globular domain. the pilins have a ladle shape...
TRANSCRIPT

1
Expression of a Clostridium perfringens type IV pilin by Neisseria gonorrhoeae mediates
adherence to muscle cells
Katherine Rodgers1, Cindy Grove Arvidson
2, Stephen Melville
1*
Running title: C. perfringens pilins in N. gonorrhoeae
Keywords: Clostridium perfringens, type IV pili, pathogenesis
1. Department of Biological Sciences, Virginia Tech, Blacksburg, VA
2. Department of Microbiology and Molecular Genetics, Michigan State University,
East Lansing, MI
*Corresponding author: Mailing address: Life Sciences I, Virginia Tech, Blacksburg,
VA 24061; Phone: 540-231-1441; email: [email protected]
Copyright © 2011, American Society for Microbiology and/or the Listed Authors/Institutions. All Rights Reserved.Infect. Immun. doi:10.1128/IAI.00909-10 IAI Accepts, published online ahead of print on 6 June 2011
on February 18, 2020 by guest
http://iai.asm.org/
Dow
nloaded from

2
Abstract
Clostridium perfringens is an anaerobic, Gram-positive bacterium that causes a range of
diseases in humans, including lethal gas gangrene. We have recently shown that strains
of C. perfringens move across the surface of agar plates using a unique type IV pili
(TFP)-mediated social motility that had not been previously described. Based on
sequence homology to pilins in Gram-negative bacteria, C. perfringens appears to have
two pilin subunits, PilA1 and PilA2. Structural prediction analysis indicated PilA1 is
similar to the pseudopilin found in Klebsiella oxytoca, while PilA2 is more similar to true
pilins found in the Gram-negative pathogens Pseudomonas aeruginosa and Neisseria
gonorrhoeae. Strains of N. gonorrhoeae that are genetically deficient in the native pilin,
PilE, but supplemented with inducible expression of PilA1 and PilA2 of C. perfringens,
were constructed. Genetic competence, wild-type twitching motility, and attachment to
human urogenital epithelial cells were not restored by expression of either pilin.
However, attachment was observed with the N. gonorrhoeae strain expressing PilA2 to
mouse and rat myoblast (muscle) cell lines. Significantly, wild-type C. perfringens cells
adhered to mouse myoblasts under anaerobic conditions and adherence was 10-fold lower
in a pilT mutant which lacks functional TFP. These findings implicate C. perfringens
TFP in the ability of C. perfringens to adhere to and move along muscle fibers in vivo,
which may provide a therapeutic approach to limiting this rapidly-spreading and highly
lethal infection.
on February 18, 2020 by guest
http://iai.asm.org/
Dow
nloaded from

3
Introduction 1
Clostridium perfringens is a Gram-positive anaerobic pathogen that causes gas 2
gangrene and food poisoning (32). Recently, we have shown that C. perfringens 3
possesses a unique gliding motility that is mediated by type IV pili (TFP) (39). TFP in 4
other bacteria are implicated in attachment to and invasion of host cells, microcolony and 5
biofilm formation, attachment to abiotic surfaces, twitching motility, bacteriophage 6
susceptibility, and genetic competence (1, 13, 17, 23, 28, 39, 40). TFP are long, flexible 7
filaments that extend from the bacterial cell surface and are homopolymers composed of 8
thousands of copies of a single protein, pilin (4, 15). Pilins are synthesized as precursor 9
proteins with an N-terminal leader sequence that is cleaved by a bifunctional 10
endopeptidase (PilD), usually at a phenylalanine residue immediately preceded by a 11
glycine residue; the phenylalanine residue is also methylated by the same endopeptidase 12
(30, 38). 13
The crystal structures of several type IV pilins from Gram-negative bacteria have 14
been solved (3, 7, 8, 12, 29). Pilin structures reveal a conserved core comprised of an 15
extended N-terminal α-helix and a globular domain. The pilins have a ladle shape due to 16
the protrusion of the N-terminal helix from the globular domain (8). Two regions flank 17
the conserved structural core and are exposed on the surfaces of the pilus filaments: the 18
αβ-loop and the D-region, delineated by two cysteines linking the C-terminal segment to 19
the β-sheet via a disulfide bond (6). These regions are believed to perform many of the 20
pilus-associated functions (10). 21
Two gene products, PilA1 and PilA2, were identified as putative pilins that serve 22
as structural subunits of TFP in C. perfringens (36, 39). In a previous report, we 23
on February 18, 2020 by guest
http://iai.asm.org/
Dow
nloaded from

4
described results obtained from the FUGUE protein structure prediction program (39) 24
comparing the C. perfringens PilA proteins to those in the database (available on the 25
FUGUE server at http://www-cryst.bioc.cam.ac.uk/~fugue/). PilA1 proteins had only a 26
single match to the pseudopilin PulG from Klebsiella oxytoca. However, the PilA2 27
proteins had matches to both PulG and pilins from P. aeruginosa and N. gonorrhoeae 28
(39). Since pseudopilins have similar folds to pilins, but lack the D-loop domain, the 29
similarity of the PilA2 protein to both the pseudopilins and the pilins is logical, since 30
pseudopilins comprise a subset of the larger pilin structures. The conserved cysteine pair 31
seen in the Gram-negative type IV pilins is not present in PilA2, but the C-terminal 32
segment is still present and is predicted to form a loop similar to the D-region of PilE 33
(39). 34
The N-terminal α-helices comprise the pilin polymerization domain, based on 35
cryo-electron microscopy (cryoEM) reconstruction of the N. gonorrhoeae type IV pilus 36
(8). The N-terminal α-helices are arranged in a helical array inside the filament core, 37
making extensive contacts with one another to provide mechanical strength. The 38
globular domains line the outer core of the pilus, exposing the αβ-loop and D-regions (6). 39
The C. perfringens pilus PilA2, along with all type IV pili, likely shares the same helical 40
architecture as other pilins, based on conserved sequence, structural and biological 41
functions associated with all TFP systems analyzed thus far. 42
The structure of the N-terminal truncated PulG pseudopilin from Klebsiella oxytoca 43
(18), which is similar to the predicted PilA1 structure, shows a similar fold in the N-terminal 44
domain and globular head region as that seen in true pilins but lacks the D-loop containing 45
the hypervariable region between the disulfide bond. The 3-dimensional structure allowed a 46
on February 18, 2020 by guest
http://iai.asm.org/
Dow
nloaded from

5
model for pseudopilus stacking in a filament to be constructed, where 17 monomers 47
comprised four helical turns in a left-handed filament (18). The pseudopilin filaments are 48
predicted to be much shorter than those in TFP, which corresponds to the observation that 49
pseudopilins do not extend past the outer membrane in Gram-negative bacteria. 50
Additional evidence supporting the assignment of PilA1 as a pseudopilin and 51
PilA2 as a true pilin comes from a phylogenetic analysis of the conserved region (i.e., 52
first 65-69 amino acids) of pilin and pseudopilin proteins (Fig. 1A and Fig. S1). The 53
analysis was carried out using previously described methods (31). The bootstrap analysis 54
shows pilins and pseudopilins fall into two distinct families of proteins and that C. 55
perfringens PilA1 is in the pseudopilin family and PilA2 in the pilin family (Fig. 1A). In 56
addition, the bootstrap values for PilA1 and PilA2 have a top score, suggesting they are 57
both derived from an independent line of descent within the pilin or pseudopilin group 58
(Fig. 1A). This supports our previous hypothesis that Clostridia TFP are ancient and 59
likely the ancestral forms from which all Eubacteria TFP systems have evolved (39). 60
With this structural information in hand, we wanted to assign a specific function 61
to each pilin. Because C. perfringens pili are short (200-300 nm) and difficult to 62
distinguish from one another on intact cells (39), we decided to determine if PilA1 and 63
PilA2 can complement a deficiency in PilE, the native pilin, in N. gonorrhoeae strain 64
MS11. N. gonorrhoeae was chosen because (i) the pili assembly apparatus has been 65
well-characterized, (ii) heterologous expression of Pseudomonas aeruginosa PilA (43) 66
and Francisella tularensis PilA (33) in N. gonorrhoeae resulted in formation of intact 67
pili, (iii) N. gonorrhoeae exists as single cells or diplococci, which makes visual 68
observations significantly easier in comparison to C. perfringens, in which motile 69
bacteria are found in long chains of cells with an end-to-end conformation (39). Our 70
on February 18, 2020 by guest
http://iai.asm.org/
Dow
nloaded from

6
results show that PilA2 from C. perfringens was assembled into pili but PilA1 was not. 71
While neither pilin from C. perfringens complemented the absence of native pilin for 72
genetic competence or adherence to human urogenital cells, PilA2 pili enabled the N. 73
gonorrhoeae strain expressing this protein to adhere to myoblasts (muscle cells) derived 74
from rodent cell lines. C. perfringens, under anaerobic conditions, was also able to 75
adhere to mouse myoblasts in a TFP-dependent manner, suggesting the results seen with 76
N. gonorrhoeae are relevant to the host species itself. 77
on February 18, 2020 by guest
http://iai.asm.org/
Dow
nloaded from

7
Materials and Methods 78
Bacterial strains and growth conditions. The bacterial strains used in this study are 79
listed in Table 1. E. coli DH10B and DH5α were used for cloning steps and were grown 80
in Luria-Bertani (LB) broth, supplemented with 15 g/l agar (Difco) for solid media. 81
Twenty µg/ml chloramphenicol and 100 µg/ml kanamycin were added when necessary 82
for selection. C. perfringens strains 13, SM125 (pilT mutant), and SM125(pSM271) 83
(complemented pilT mutant) were grown anaerobically as previously described (39). 84
N. gonorrhoeae strains were derivatives of MS11A (34). N. gonorrhoeae strain 85
MS11-307 (∆pilE1::erm; ∆pilE2) served as the non-revertible non-piliated control (2, 86
26). In liquid culture, all N. gonorrhoeae strains were grown at 37oC and 5% CO2 in GC 87
broth supplemented with 0.042% sodium bicarbonate and Kellogg’s supplements I and II 88
(16). Solid media was prepared by supplementing GC medium base (Difco) with 1.25 g/l 89
agar and Kellogg’s supplements. Three µg/ml erythromycin and 10 µg/ml 90
chloramphenicol were added as needed. Piliation and Opa- phenotypes were assessed 91
microscopically. 92
Plasmid construction. Plasmids and oligonucleotides used in this study are summarized 93
in Table 1. C. perfringens pilA1 and pilA2 genes were amplified by polymerase chain 94
reaction (PCR) using primer pairs OKR011/OKR019 and OKR013/OKR014, 95
respectively. Amplification yielded a 484 base pair pilA1 gene and a 609 base pair pilA2 96
gene. PCR products were digested with HindIII and XbaI and ligated into similarly 97
digested pKH35 (11). This placed the pilA1 and pilA2 genes immediately downstream of 98
the lac promoter, yielding plasmids pKR107 and pKR106. Clones were verified by DNA 99
sequencing. 100
on February 18, 2020 by guest
http://iai.asm.org/
Dow
nloaded from

8
Generation of N. gonorrhoeae strains expressing C. perfringens PilA1 and PilA2. A 101
derivative of the N. gonorrhoeae strain MS11 (MS11C3, ∆pilE2) was transformed with 102
pKR106 and pKR107. Resulting transformants were then transformed with genomic 103
DNA from MS11-307 (∆pilE1::Erm) (26) to yield KR106 and KR107, non-piliated 104
mutants that harbor pilA2 and pilA1, respectively. 105
Induction and detection of pilA1 and pilA2 expression. N. gonorrhoeae strains were 106
grown on GC agar (GCA) plates as described above with IPTG added to a concentration 107
of 0.5 mM Colonies grown overnight were swabbed from plates, resuspended in SDS-108
PAGE loading buffer (100 mM DTT, 50 mM Tris-HCl, 2% SDS, 0.1% bromophenol 109
blue, and 10% glycerol) and boiled for 20 minutes. Samples were centrifuged for 8 110
seconds at 12,500 rpm in a microcentrifuge, and 15 µl of the supernatant subjected to 111
SDS-PAGE. Western blots using antibodies to PilA1 and PilA2 were carried out as 112
previously described (39). 113
Growth curves. One colony each from strains KR106 and KR107 was inoculated into 20 114
ml GCB supplemented with 0.042% sodium bicarbonate and incubated at 37oC with 115
shaking at 300 rpm. Overnight cultures were diluted 1/20 with fresh medium and 116
incubated at 37oC with shaking. IPTG (0.5 mM) was added to one 20 ml culture of 117
KR106 and one KR107 culture and 1.0 mM IPTG was added to a separate culture of each 118
strain. One ml of each culture was removed every hour, serially diluted with fresh 119
medium, and plated on GCA. Plates were incubated at 37oC overnight, and colonies 120
were counted to calculate colony forming units (CFU)/ml. 121
Attempted pilT mutagenesis. The pilT gene was amplified from MS11 chromosomal 122
DNA with oligonucleotides OKR063 and OKR064, using Phusion High-Fidelity DNA 123
on February 18, 2020 by guest
http://iai.asm.org/
Dow
nloaded from

9
polymerase (New England Biolabs). The PCR product was cloned into pGEM-T easy 124
(Promega) resulting in plasmid pKR112. The kanamycin resistance gene was amplified 125
by PCR from chromosomal DNA of an N. gonorrhoeae strain carrying the 126
EZ::Tn5<KAN-2> transposon using primers OKR065 and OKR066 and was then cloned 127
into the pilT gene to produce plasmid pKR113. The linear fragment consisting of the pilT 128
gene interrupted by the kanamycin resistance cassette was used for electroporation with 129
recipient N. gonorrhoeae strains MS11 and KR106, as previously described (9). 130
Immunofluorescence. N. gonorrhoeae strains KR106 and KR107, grown for 24 hours 131
with and without IPTG (1 mM), were swabbed and resuspended in GC broth to an OD600 132
of 0.1. Ten µl of each were placed on glass coverslips and incubated at 37oC for 1.5 133
hours and then fixed by adding 700 µl of 2.5% paraformaldehyde in Dulbecco’s 134
phosphate buffered saline (DPBS) for 10 minutes. Coverslips were then washed in 2% 135
bovine serum albumin (BSA) in DPBS (BSA/DPBS) and blocked in 10% goat serum in 136
DPBS at 37oC for 30 minutes. The coverslips were washed twice in BSA/DPBS and then 137
incubated with affinity-purified antibodies (39) against PilA1 or PilA2 in BSA/DPBS at 138
37oC for 30 minutes. Coverslips were then washed twice in 2% BSA in DPBS and 139
incubated with 1:100 goat anti-rabbit serum conjugated to Alexafluor 594 (Molecular 140
Probes). Coverslips were subjected to a final wash with 2% BSA in DPBS and images 141
captured using an Olympus IX81 upright microscope linked to a Hamamatsu Model 142
C4742 CCD camera. Slidebook 4.1 imaging software (Intelligent Imaging Innovations) 143
was used to collect images. 144
Transformation assays. N. gonorrhoeae strains were swabbed from plates grown for 18 145
hours into 1 ml GC broth supplemented with 5 mM MgCl2 and Kellogg’s supplements I 146
on February 18, 2020 by guest
http://iai.asm.org/
Dow
nloaded from

10
and II (GCB+Mg+supps). Concentration of bacteria was determined 147
spectrophotometrically and adjusted to 108 CFU/ml. Twenty µl of this culture and 200 148
ng of genomic DNA from N. gonorrhoeae strain MS11-310 (∆pilE1::Kan) were added to 149
180 µl GCB+Mg+supps and incubated at 37oC and 5% CO2 for one hour to allow DNA 150
uptake. The samples were then diluted 10-fold with GCB+Mg+supps and incubated for 151
four hours at 37oC with 5% CO2. Serial dilutions of the transformation mixes were plated 152
on nonselective and selective (100 µg/ml kanamycin) GCA and incubated 48 hours at 153
37oC and 5% CO2. Transformation efficiency was calculated by dividing the number of 154
KanR transformants by the total number of bacteria present at the end of the experiment. 155
N. gonorrhoeae adherence assays. For A431 (human epidermal carcinoma cell line) 156
and MDCK (canine kidney cell line), 104 cells per well were seeded into a 24-well plate 157
in DMEM for A431 cells or Eagle’s Minimum Essential Medium (EMEM) for MDCK 158
cells and incubated at 37oC and 5% CO2 until approximately 90% confluent. GC strains 159
were swabbed from plates grown for 24 hours and then used to inoculate 1 ml prewarmed 160
supplemented GCB. KR106 and KR107 were swabbed from plates without IPTG as well 161
as plates containing 0.5 mM IPTG and diluted to a concentration of 4 X 106 CFU/ml
in 162
DMEM for adherence to A431 cells or EMEM for adherence to MDCK cells. Cell 163
culture media were supplemented with 10% heat treated FBS and Kellogg’s supplement 164
II. Spent cell culture medium was removed from the seeded eukaryotic cells, and each 165
well inoculated with 250 µl of the appropriate bacterial strain. Six wells were inoculated 166
per N. gonorrhoeae strain and incubated for three hours at 37oC and 5% CO2. Three 167
wells per strain were designated “total bacteria” wells, and three were designated 168
“adherent bacteria” wells. Total bacteria were calculated as follows: 250 µl of 169
on February 18, 2020 by guest
http://iai.asm.org/
Dow
nloaded from

11
supernatant (total liquid volume) was removed from the well and set aside. Then, 750 µl 170
PBS with 0.25% EDTA was added to each well and incubated at room temperature for 10 171
minutes, after which cells that remained attached were removed by vigorous pipetting. 172
This 750 µl of bacteria and detached cells were added to the original 250 µl removed 173
from the well and dilutions made in PBS followed by plating on GCA. Colonies were 174
counted after 48 hours of incubation at 37oC and 5% CO2. For adherent bacteria, the 175
supernatant was aspirated and each well was washed five times with PBS. Then 1 ml of 176
PBS containing 0.25% EDTA was added to each well and allowed to sit for 10 minutes. 177
Cells that remained attached were lifted by vigorous pipetting. Following mixing with a 178
vortexer for 20 seconds, dilutions were prepared in PBS and plated on GCA 179
supplemented with Kellogg’s supplements I and II. Colonies were counted after 48 hours 180
of incubation at 37oC and 5% CO2. The percent adherence of each strain was determined 181
by dividing the number of adherent bacteria by the number of total bacteria per well and 182
then multiplying by 100. Induced expression of PilA1 and PilA2 was confirmed by 183
Western blot, as described above. 184
For C2C12 myoblasts (46) and L6 myoblasts (22), cells were seeded into 48-well 185
plates containing Dulbecco's modification of Eagle's Medium with 10% fetal bovine 186
serum (DMEM/FBS) and incubated at 37oC and 5% CO2 until approximately 90% 187
confluence was observed. N. gonorrhoeae strains were swabbed from plates incubated 188
for 24 hours and used to inoculate 1 ml of prewarmed supplemented GCB. KR106 and 189
KR107 were swabbed from plates without IPTG as well as plates containing 0.5 mM 190
IPTG. Each strain was diluted in DMEM supplemented with 10% heat-treated FBS and 191
Kellogg’s supplement II to a concentration of 4 X 108 CFU/ml. The spent cell culture 192
on February 18, 2020 by guest
http://iai.asm.org/
Dow
nloaded from

12
medium was removed from the seeded myoblasts, and each well inoculated with 250 µl 193
of the appropriate bacterial strain. A total of 11 wells were inoculated per GC strain. 194
These were incubated at 37oC and 5% CO2 for three hours. Total bacteria and adherent 195
bacteria were calculated as described above, with the exception that myoblasts were lifted 196
from wells by scraping in three directions with a pipette tip, and then pipetting vigorously 197
up and down. Expression of PilA1 and PilA2 from strains KR107 and KR106, 198
respectively, was confirmed by western blot on induced cultures used in these assays. 199
C. perfringens adherence assays. Adherence of C. perfringens strains to C2C12 cells 200
took place in a Coy anaerobic chamber with an atmosphere of 85% N2, 10% CO2, and 5% 201
H2. C2C12 cells were grown to confluency (~2 days) in 0.5 ml DMEM/FBS in 48-well 202
tissue culture plates and then placed in a 37 ºC incubator inside the anaerobic chamber. 203
C. perfringens strains, grown overnight on PGY medium with agar (24) under anaerobic 204
conditions, were removed from the anaerobic chamber, scraped off the plates and 205
suspended in 1 ml DPBS. Bacteria were pelleted in a centrifuge, suspended in DPBS and 206
the suspensions diluted in DPBS to give ~2 x 107 CFU/ml. After the C2C12 cells were in 207
anaerobic conditions for 2 h, 5 µl of the bacterial suspension (~1 x 105 CFU) was added 208
to each well and incubated anaerobically at 37 ºC for 75 min. The plates were then 209
removed from the anaerobic chamber and each well was washed 3 times with 0.5 ml 210
aerobic DPBS to remove unattached bacteria. After the final wash, one half ml of 211
distilled water was added to the wells to lyse the myoblasts. The cells and bacteria were 212
scraped off the bottom of the well, placed in a microcentrifuge tube and subjected to 213
vortex mixing for 20 sec. The bacteria in the sample were then quantified by serial 214
dilution and plating on PGY medium. Attachment assays for each strain were performed 215
on February 18, 2020 by guest
http://iai.asm.org/
Dow
nloaded from

13
on quintuplicate samples from two separate experiments (10 samples total). C2C12 cell 216
viability before and after incubation under anaerobic conditions was tested using the 217
trypan blue dye exclusion assay, as previously described (27). 218
Video microscopy. All video was obtained using an Olympus IX81 upright microscope 219
linked to a Hamamatsu Model C4742 CCD camera. Slidebook 4.1 (Intelligent Imaging 220
Innovations) imaging software was used compile motility videos. To determine if strains 221
KR106 and KR107 exhibited twitching motility, colonies of each strain were removed 222
from plates incubated overnight and resuspended in GCB that was prewarmed to 37oC 223
and supplemented with Kellogg’s supplements and 0.042% sodium bicarbonate. Ten µl 224
of the bacterial suspensions were added to microscope slides, covered with coverslips, 225
and immediately inverted. Slides were maintained at 37oC during microscopy. 226
To examine the interactions of C. perfringens with C2C12 cells under anaerobic 227
conditions, C2C12 cells were first grown aerobically in DMEM/FBS in 50 ml tissue 228
culture flasks until reaching ~50% confluency. They were then transferred to a 37 ºC 229
incubator inside a Coy anaerobic chamber which contained an atmosphere of 85% N2, 230
10% CO2 and 5% H2. After two hours under anaerobic conditions, a 1:1 multiplicity of 231
infection (MOI) of C. perfringens cells was added to the flask. The cap on the flask was 232
tightened to prevent oxygen from getting inside and placed on a heated stage set at 37 ºC 233
for video microscopy. 234
Electron microscopy. For field emission scanning electron microscopy (FE-SEM) to 235
visualize pili, N. gonorrhoeae strains were grown overnight on GC agar and Kellogg’s 236
supplements, then spread on 0.2 µm GTTP filters (Millipore) on the surface of fresh GC 237
agar plates with 1 mM IPTG and incubated at 37 ºC in a CO2 incubator for 2 h. After 2 h 238
on February 18, 2020 by guest
http://iai.asm.org/
Dow
nloaded from

14
of growth, the filters were lifted from the plates and mounted on top of aluminum sample 239
holders embedded in dry ice, which quickly froze the samples. The filters were then 240
sputter-coated with gold to a thickness of 1 nm and pili were imaged using a Helios 600 241
NanoLab SEM (FEI). For SEM of N. gonorrhoeae strains attached to C2C12 myoblasts, 242
attachment was obtained by seeding C2C12 myoblasts on 13 mm round glass coverslips 243
layered on glass beads in a 12-well tissue culture plate and allowed to grow to 244
approximately 90% confluency. N. gonorrhoeae strains were added and incubated as 245
described above for attachment assays, and samples on glass coverslips were transferred 246
to 6-well plates containing 2 ml of 2.5% glutaraldehyde in 0.1 M sodium cacodylate (pH 247
7.2) and fixed overnight at 4oC. Samples were post-fixed for 30 min in 1% osmium 248
tetroxide in cacodylate buffer. The coverslips were then washed in deionized water, 249
dehydrated in ethanol and critical point dried using a Ladd critical point dryer. Dried 250
samples were mounted onto sample holders, lightly sputter-coated with gold and 251
examined with a Phillips 505 scanning electron microscope. 252
on February 18, 2020 by guest
http://iai.asm.org/
Dow
nloaded from

15
Results 253
Pilin alignments. Alignment of PilA1 and PilA2 from C. perfringens strain 13 254
with PilE from N. gonorrhoeae strain MS11 are shown in Fig. 1B. Overall sequence 255
homology between PilA1/PilA2 and PilE is low, except in the N-terminal region (Fig. 256
1B). Even less sequence similarity was seen when PilA1 and PilA2 were compared to 257
the minor pilins PilX and PilV of N. gonorrhoeae (data not shown). Both PilA1 and 258
PilA2 from C. perfringens possess conserved N-terminal leader sequences at the site of 259
pre-pilin peptidase (PilD) cleavage (Fig. 1B), which suggests they are likely to be 260
processed into mature pilins. 261
Expression of PilA1 and PilA2 in N. gonorrhoeae. N. gonorrhoeae strains 262
KR107 and KR106 were constructed to express C. perfringens PilA1 and PilA2, 263
respectively, under an IPTG-inducible promoter in a pilE mutant background using 264
previously described methods (11). Addition of 0.5 mM or 1 mM IPTG had no effect on 265
the growth rate of either strain (Fig. S2). In the presence of IPTG, strain KR107 (PilA1 266
expression) had a colony pilin phenotype identical to the PilE mutant strain, MS11-307, 267
while KR106 (PilA2 expression) had a colony pilin phenotype somewhere between 268
MS11-307 and the MS11 (PilE+). Pilin proteins were detected by western blot using 269
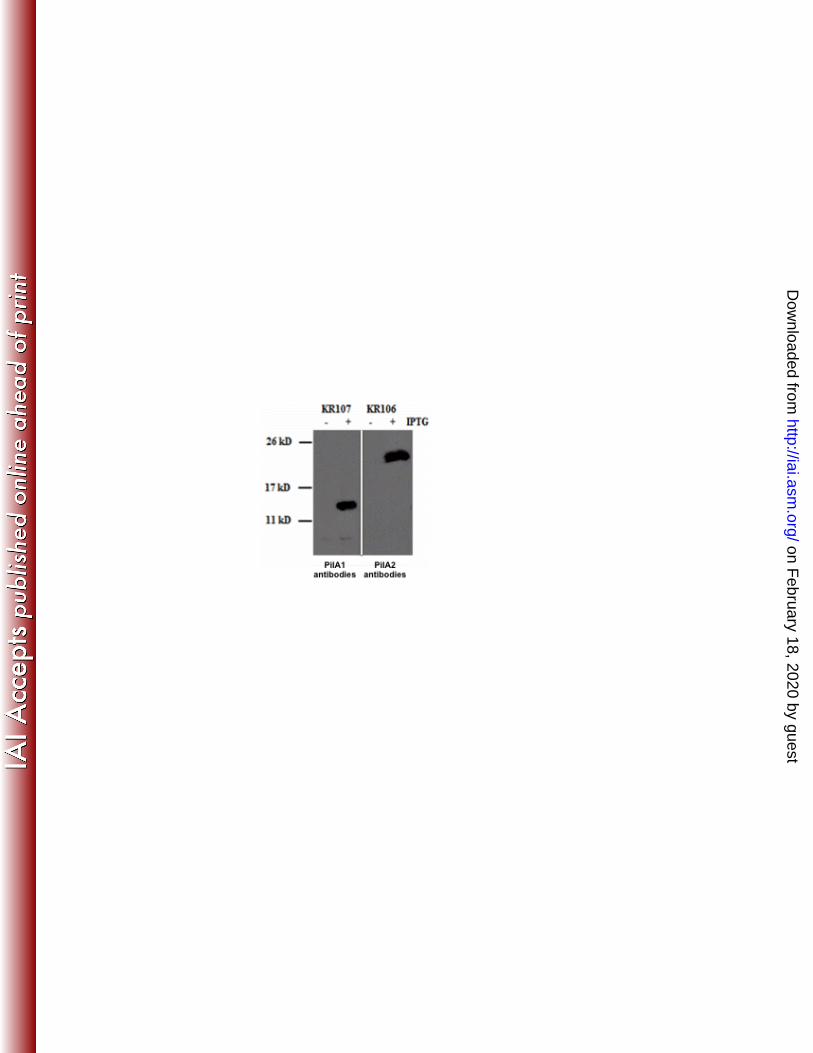
affinity purified anti-PilA1 and anti-PilA2 antibodies on whole cell lysates prepared from 270
GC strains KR107 and KR106, respectively, after induction with IPTG. Neither protein 271
was detected in the absence of IPTG (Fig. 2). PilA1 expressed in N. gonorrhoeae 272
appeared to be 14 kD, which runs as slightly larger than the predicted size of the 273
processed protein, at 12.6 kD. However, PilA2 expressed in N. gonorrhoeae is 23 kD, 274
larger than the predicted size of 19.5 kD. To determine if PilA2 was post-translationally 275
on February 18, 2020 by guest
http://iai.asm.org/
Dow
nloaded from

16
modified in N. gonorrhoeae, PilA2-containing cell free extracts were treated with O-276
glycosidase, N-glycosidase, and sialidase, in series and individually and with both 277
Antarctic phosphatase and alkaline phosphatase. There was no change in the apparent 278
mass of PilA2 after any of these treatments (data not shown). These results suggest 279
PilA2 is either covalently modified with an unknown moiety or migrates somewhat 280
slower than predicted in SDS-PAGE gels. 281
Immunofluorescence using affinity purified antibodies against PilA1 and PilA2 282
showed increased expression of each of these proteins on the surface of N. gonorrhoeae 283
cells after induction by IPTG (Fig. 3). The fluorescence appeared in specific regions on 284
the surface of the cells (Fig. 3), which may correspond to the location of pili on the 285
surface of the bacteria, but pili themselves, if present would not be resolved using light 286
microscopy due to their small size. 287
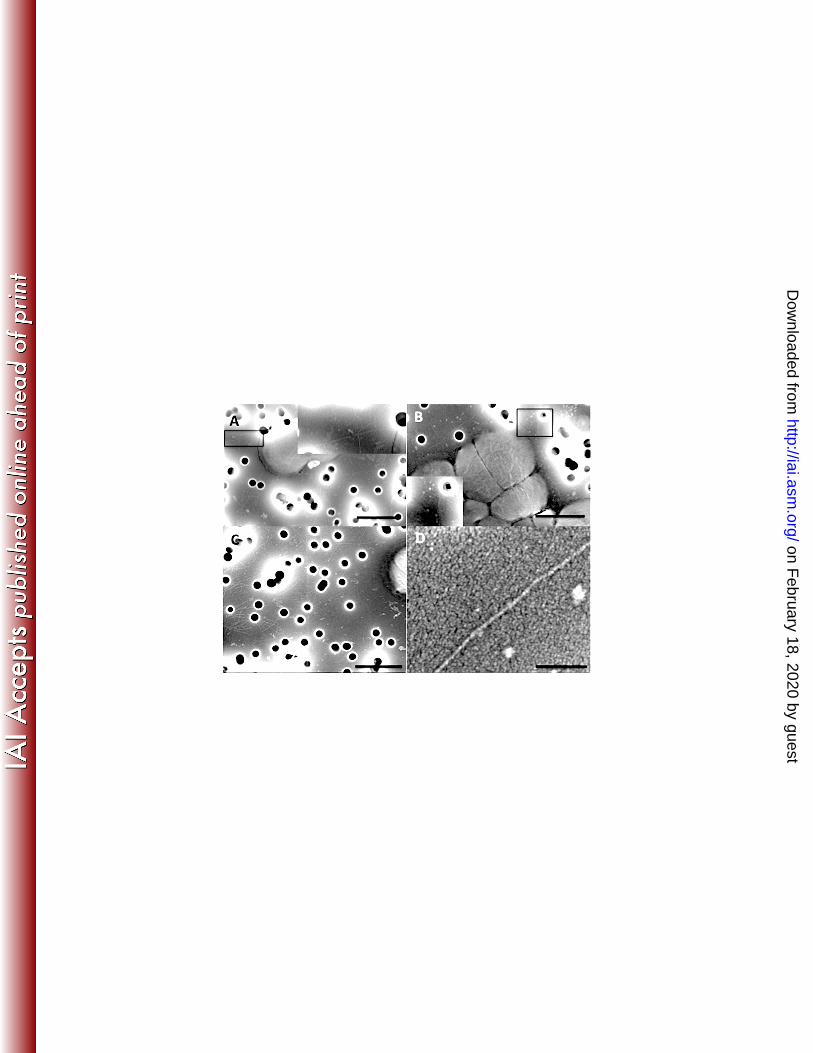
Pili were visible by FE-SEM on strain MS11, the N. gonorrhoeae wild-type 288
strain, grown on GTTP filters (Fig. 4A). Despite examining thousands of cells of each 289
type, no pili could be seen with the pilE mutant strain, MS11-307, or the strain expressing 290
C. perfringens pilA1, KR107 (data not shown). Strain KR106, expressing the pilA2 gene 291
in the presence of IPTG, produced numerous pili associated with ~10-15% of the cells 292
(data not shown and Fig. 4B). Measurements of the length of pili that could be clearly 293
defined as being attached to bacteria showed the PilA2 fibers were similar in length to 294
PilE fibers, 1.06 +/- 0.35 µm and 1.08 +/- 0.29 µm, respectively. Many PilA2 fibers were 295
visible on the surface of filters in regions lacking bacteria (Fig. 4C), suggesting they had 296
detached during growth or while processing the filters for electron microscopy. A high 297
on February 18, 2020 by guest
http://iai.asm.org/
Dow
nloaded from

17
magnification image of an isolated PilA2 fiber (Fig. 4D) shows the fibers are ~7 nm in 298
diameter, similar to that seen with fibers on the surface of C. perfringens (39). 299
Complementation of pilin functions. Because TFP are necessary for genetic 300
transformation in N. gonorrhoeae, strains KR106 and KR107 were assayed for their 301
ability to restore transformation in the absence of pili. Wild-type strain MS11 302
transformed at a frequency of 2 x 10-3
transformants per CFU using genomic DNA from a 303
kanamycin-resistant N. gonorrhoeae strain. However, no transformants were detected in 304
the same experiment with the pilE mutant (MS11-307), or N. gonorrhoeae expressing 305
PilA1 or PilA2 from C. perfringens (data not shown). 306
TFP are also needed for twitching motility in N. gonorrhoeae. Video microscopy 307
was used to determine if PilA1 and PilA2-expressing strains exhibited twitching motility. 308
The cells in Video 1 clearly demonstrate the characteristic twitching motility of wild-type 309
N. gonorrhoeae on a glass coverslip. N. gonorrhoeae strain MS11-307, a pilE mutant, 310
did not show twitching motility and the cells of this strain are motionless (Video 2). 311
Similarly, strain KR107 bacteria can be seen binding to both the coverslip (in-focus cells) 312
and the glass slide (out of focus cells) but exhibited very little motion in the presence of 313
IPTG (Video 3) , indicating that expression of PilA1 from C. perfringens does not restore 314
twitching motility in the absence of native PilE. N. gonorrhoeae strain KR106 possesses 315
a twitching motility phenotype that resembles neither the twitching motility of wild-type 316
N. gonorrhoeae nor the motionless phenotype of the pilE mutant. In the presence of 317
IPTG, some KR106 cells appear to adhere to the glass coverslip and spin in place (Video 318
4). We hypothesize that the diplococci can extend a pilus composed of C. perfringens 319
PilA2, but that the pilus cannot easily be retracted, so the cell remains tethered to the 320
on February 18, 2020 by guest
http://iai.asm.org/
Dow
nloaded from

18
glass coverslip and rotates due to Brownian motion. While only a fraction of the cells in 321
any given video frame exhibited this phenotype (Video 4), this fraction roughly 322
corresponds to the fraction of cells possessing pili, as seen by FE-SEM (Fig. 4B). 323
Adherence of N. gonorrhoeae expressing PilA1 and PilA2 to mammalian 324
cells. N. gonorrhoeae strains expressing PilA1 and PilA2 were tested for their ability to 325
mediate adherence to human epithelial cells, which in N. gonorrhoeae is partially 326
dependent on TFP (21). Wild-type N. gonorrhoeae adhered to A431 cells (a vulval 327
carcinoma cell line) at an average frequency of 33% (Figure 5A). The pilE mutant strain 328
and strain KR106 in the absence or presence of IPTG did not adhere appreciably to this 329
cell line (Figure 5A). We observed nonspecific binding of strain KR107 bacteria, but 330
none of the other strains tested, to the plastic tissue culture plates in the absence of 331
mammalian cells (data not shown), and therefore, adherence data for this strain is not 332
included. 333
Since C. perfringens strain 13 is an etiologic agent of gas gangrene, which is an 334
infection of muscle tissue, we rationalized that C. perfringens was likely to encounter 335
muscle cells and connective tissue cells during a gangrene infection. Therefore, a N. 336
gonorrhoeae strain expressing PilA2 was tested for its ability to adhere to two different 337
myoblast cell lines, C2C12 and L6, derived from mice and rats, respectively. Wild-type 338
N. gonorrhoeae adhered to C2C12 cells at a frequency of 0.0005%, which was not 339
statistically different from the adherence of the non-piliated mutant at 0.02% (Fig. 5B). 340
KR106, adhered to C2C12 cells at a rate of 0.15% in the presence of IPTG but adherence 341
dropped to 0.02%, in the absence of IPTG (Fig. 5B). This was significantly higher than 342
on February 18, 2020 by guest
http://iai.asm.org/
Dow
nloaded from

19
that of wild-type N. gonorrhoeae, the non-piliated mutant, and uninduced KR106 343
(P<0.001). 344
Surprisingly, wild-type N. gonorrhoeae adhered to L6 myoblasts (rat) at a rate of 345
2.13% (Figure 5C). The pilE mutant adhered at a rate of 0.02%, which was 100-fold less 346
and statistically significant (P<0.0066). PilA2 expression in strain KR106 resulted in 347
adherence to L6 cells at a rate of 0.17%, which is significantly higher adherence than 348
observed strain KR106 without IPTG (P<0.0256), which adhered at a rate of 0.012%, a 349
14-fold difference. We also tested cells derived from a type of connective tissue, primary 350
mouse fibroblasts, and found that both strains MS11 and KR106 with IPTG adhered 351
significantly better than the corresponding strains, MS11-307 and KR106 without IPTG 352
(Fig. 5D), suggesting either pilus could mediate adherence. 353
In order to assess the adherence of each strain to a cell type in which it was 354
expected that neither N. gonorrhoeae native pili nor C. perfringens pili would bind, 355
assays were performed with MDCK (canine kidney) cells (Figure 5E). As projected, 356
adherence to this cell line was less than 0.003% for all strains tested, and no strain 357
adhered at a significantly higher frequency than any other. 358
Visual confirmation of adherence to mouse muscle cells (C2C12) was obtained by 359
SEM of C2C12 cells cultured on glass cover slips. Cells of strain KR106 appeared as 360
either microcolonies (Fig. 6A) or isolated diplococci (Fig. 6B). Strain KR107 can be 361
seen binding to both the smooth surface of the cover slip and C2C12 cells in SEM images 362
(Fig. 6C), confirming our observation that PilA1 expression was causing N. gonorrhoeae 363
to adhere to inert surfaces such as plastic plates and glass coverslips in the absence of 364
mammalian cells. 365
on February 18, 2020 by guest
http://iai.asm.org/
Dow
nloaded from

20
Adherence of C. perfringens to C2C12 cells. Since PilA2 expression allowed N. 366
gonorrhoeae to adhere to rodent myoblasts, we performed adherence assays with live C. 367
perfringens strain 13 bacteria, the source of the pilA1 and pilA2 genes used in this study. 368
Under anaerobic conditions in liquid medium (DMEM/FBS) >95% of the C2C12 369
myoblasts were viable for 12 hours (data not shown), as measured using trypan blue 370
exclusion. Initial attachment assays using cells with fresh DMEM/FBS showed 371
undetectable adherence of C. perfringens bacteria to C2C12 cells under anaerobic 372
conditions (data not shown). Video microscopy of the C. perfringens-C2C12 interactions 373
under anaerobic conditions showed the bacteria were not lined up in the characteristic 374
end-to-end fashion seen in motile C. perfringens cells (39) but rather floated in the tissue 375
culture medium mostly as individual cells (Video 5). We theorized that glucose, present 376
in fresh DMEM at 4.5 g/l, might be inhibiting the expression of motility genes, as 377
previously reported (25, 39). Therefore, we carried out the same assay using C2C12 cells 378
that had been grown for 2 days in DMEM to deplete the glucose before placement in an 379
anaerobic chamber. Under these conditions, C. perfringens strain 13 adhered at a 380
relatively high level of 3.8% (Fig. 7). A strain 13 mutant with an inactivated pilT gene, 381
SM125, adhered ~10-fold less efficiently (Fig. 7). When strain SM125 was 382
complemented with a multicopy plasmid carrying an intact pilT gene, adherence 383
increased ~2-fold, but not to the same level as the wild type strain (Fig. 7). Video 384
microscopy of the C2C12 cells and strain 13 bacteria under low glucose conditions 385
showed the bacteria now formed chains and aggregates that appear to be connected in a 386
head-to-tail manner (Video 6), similar to what was seen on agar surfaces (39). In 387
on February 18, 2020 by guest
http://iai.asm.org/
Dow
nloaded from

21
contrast, the pilT mutant strain, while composed of somewhat longer cells than those seen 388
with strain 13, did not form long chains or aggregates (Video 7). 389
on February 18, 2020 by guest
http://iai.asm.org/
Dow
nloaded from

22
Discussion 390
Structural prediction programs provided evidence that C. perfringens PilA1 is a 391
pseudopilus while PilA2 is a true pilin (39). In an effort to obtain information about the 392
actual functions of these pilins in C. perfringens, we constructed strains of N. 393
gonorrhoeae that express these proteins. The pilA1 gene was expressed and the protein 394
made (Fig. 2), but no pili could be seen on the surface of the cells using FE-SEM (data 395
not shown). In contrast, the PilA1 protein could be found on the surface of intact N. 396
gonorrhoeae using immunofluorescence (Fig. 3). We interpret these results as indicating 397
PilA1 protein is somehow exposed on the surface to allow antibodies to bind but is not 398
present as intact pili. This may be due to its predicted pseudopilin-like structure, which 399
might not allow the N. gonorrhoeae pilus assembly apparatus to polymerize it into pili. 400
The observation that PilA1 expression mediated attachment to artificial surfaces such as 401
plastic and glass (Fig. 6) may correlate with the presence of this protein on the surface of 402
the bacteria. 403
Pseudopili are often associated with type II secretion systems (14). High level 404
heterologous expression of the Klebsiella pneumoniae pseudopilus PulG in E. coli 405
resulted in the formation of bundled extracellular pili (41). However, unlike E. coli, N. 406
gonorrhoeae does not possess a type II secretion system of its own, which may explain 407
why the results from our experiments differed from those obtained by Vignon et al (41). 408
Altogether, these results indicate the function of PilA1 in C. perfringens remains to be 409
determined. 410
The N. gonorrhoeae strain expressing pilA2, KR106, formed pili that resemble 411
those visible by FE-SEM on the surface of C. perfringens in width (5-7 nm (39)). 412
on February 18, 2020 by guest
http://iai.asm.org/
Dow
nloaded from

23
However, the pili visible on the surface of C. perfringens are 0.2-0.3 µm long (39) while 413
PilA2 made by N. gonorrhoeae are ~1 µm long (Fig. 4) . Because we were unable to 414
determine what protein comprised the surface pili on C. perfringens using immunogold 415
bead labeling due to their short length and high density (39), it is possible that these pili 416
fibers were not comprised of PilA2 but some other protein. Alternatively, it may be that 417
pilus polymerization terminates more quickly in C. perfringens than N. gonorrhoeae, 418
which would account for the difference in lengths seen with these species. 419
While able to form pili, N. gonorrhoeae strain KR106 could not complement the 420
natural transformation and twitching motility phenotypes seen with wild-type N. 421
gonorrhoeae. However, strain KR106 did provide a unique motility phenotype. In 422
contrast to wild-type N. gonorrhoeae cells, which appear to crawl across the glass 423
coverslip (Video 1), a fraction of the KR106 cells examined appear to adhere to the 424
coverslip and rotate in place (Video 4), possibly due to Brownian motion of bacteria 425
suspended at the end of a tether. Because competence and normal twitching motility are 426
not restored in KR106, and both of these functions require the action of the retraction 427
ATPase PilT (44, 45), we hypothesize that PilA2 is sufficiently compatible with the TFP 428
apparatus in N. gonorrhoeae to be assembled into pili, but that PilA2 is unable to interact 429
with PilT, so pili cannot be retracted. This hypothesis was tested by attempting to create 430
a pilT mutation in strain KR106 by introducing recombinant DNA by electroporation (see 431
Materials and Methods), since strain KR106 is not competent. Despite numerous 432
attempts, no mutants were obtained using this method, perhaps due to the sensitivity of 433
strain KR106 to the high voltages required for electroporation (data not shown). 434
on February 18, 2020 by guest
http://iai.asm.org/
Dow
nloaded from

24
Expression of PilA2 mediated attachment of N. gonorrhoeae to mouse and rat 435
myoblasts as well as mouse fibroblasts but not to human vulval carcinoma cells or canine 436
kidney cells (Fig. 5). Interestingly, this pattern of attachment corresponds to cell types 437
that C. perfringens would likely encounter in a gangrene infection (i.e., muscles and 438
connective tissue), but not to other cell types. To our knowledge, this is the first specific 439
adherence factor that has been identified in C. perfringens. 440
We have also demonstrated that TFP mediate adherence of C. perfringens to 441
C2C12 cells if the conditions are optimal (Fig. 7). These conditions include anaerobiosis 442
and low glucose levels, both of which are likely to be found in the environment favorable 443
for gas gangrene to develop and spread. The pilT mutant derivative of strain 13, SM125, 444
showed a 10-fold decrease in adherence, suggesting 90% of the adherence is dependent 445
on PilT-related functions. The levels of PilA1 and PilA2 on the surface of strain SM125 446
was lower than those seen with strain 13, which differs from what has been observed in 447
most other TFP systems in Gram-negative bacteria, where pilT mutants are hyperpiliated 448
(39). Recently, a pilT mutant of the Gram-negative pathogen Francisella tularensis was 449
shown to lack surface pili (5), indicating this phenotype exists in Gram-negative bacteria 450
as well. 451
Lack of PilA1 and/or PilA2 on the surface may account for the decreased 452
adherence we observed with strain SM125. We do not know why the plasmid carrying a 453
wild-type copy of the pilT gene failed to fully complement the adherence phenotype but 454
did allow for motility on agar plates (39). One possibility is that multiple copies of the 455
pilT gene produced abnormally high levels of the PilT protein, a retraction ATPase, 456
on February 18, 2020 by guest
http://iai.asm.org/
Dow
nloaded from

25
thereby shifting the equilibrium of pilin polymerization more to the retracted state with 457
fewer pili on the surface of the cells. 458
A distinct characteristic of gas gangrene is the remarkable extent to which muscle 459
tissue is degraded (37). Adherence to muscle fibers may allow C. perfringens to move 460
along a muscle bundle and not be detached by the physical action of muscle contraction 461
and relaxation. Also, by mediating both adherence and motility, TFP may permit the 462
bacteria to surround muscle fibers in three dimensions. This would allow them to break 463
down the fibers more effectively with degradative enzymes than if they were confined to 464
a single side of a muscle bundle. 465
N. gonorrhoeae adhered to rat muscle cells and mouse fibrobalsts in a PilE-466
dependent manner (Fig. 5B), although adherence to the rat cells was much more efficient 467
(Fig. 5C-D). N. gonorrhoeae is considered to be a human-specific pathogen and does not 468
naturally infect rodents so, although of little pathogenic significance, we postulate that 469
there is a gratuitous receptor present on L6 cells but not on C2C12 cells with which 470
native N. gonorrhoeae pili can interact. This unknown receptor is likely not the same one 471
that PilA2 binds, since PilA2 mediated attachment to both L6 and C2C12 cells (Fig. 5B-472
C). 473
Our finding that N. gonorrhoeae can polymerize PilA2 from C. perfringens into 474
an intact pilus is a testament to the extreme level of conservation TFP systems have 475
retained throughout evolutionary history. Given the apparent independent line of descent 476
of PilA2 from Gram-negative pilins (Fig. 1A) and the proposal that the Clostridia are 477
likely the ancestral form of Eubacteria TFP (39), polymerization of PilA2 into intact pili 478
on February 18, 2020 by guest
http://iai.asm.org/
Dow
nloaded from

26
by N. gonorrhoeae suggests this TFP assembly system has maintained the same basic 479
structural properties since the earliest stages of bacterial evolution. 480
on February 18, 2020 by guest
http://iai.asm.org/
Dow
nloaded from

27
Acknowledgements 481
We thank Gemma Reguera for constructing the pilin phylogenetic tree, Kathy Lowe for 482
assistance with scanning electron microscopy and Steve McCartney and John McIntosh 483
for help with the FE-SEM. We also thank William Huckle for providing MDCK cells, 484
Matthew Hulver for providing L6 myoblasts and Liwu Li for mouse primary fibroblasts. 485
on February 18, 2020 by guest
http://iai.asm.org/
Dow
nloaded from

28
Literature Cited
1. Aas, F. E., M. Wolfgang, S. Frye, S. Dunham, C. Lovold, and M. Koomey. 2002.
Competence for natural transformation in Neisseria gonorrhoeae: components of DNA
binding and uptake linked to type IV pilus expression. Mol Microbiol 46:749-60.
2. Arvidson, C. G., R. Kirkpatrick, M. T. Witkamp, J. A. Larson, C. A. Schipper, L. S.
Waldbeser, P. O'Gaora, M. Cooper, and M. So. 1999. Neisseria gonorrhoeae mutants
altered in toxicity to human fallopian tubes and molecular characterization of the genetic
locus involved. Infect Immun 67:643-52.
3. Audette, G. F., R. T. Irvin, and B. Hazes. 2004. Crystallographic analysis of the
Pseudomonas aeruginosa strain K122-4 monomeric pilin reveals a conserved receptor-
binding architecture. Biochemistry 43:11427-35.
4. Bullitt, E., and L. Makowski. 1998. Bacterial adhesion pili are heterologous assemblies
of similar subunits. Biophys J 74:623-32.
5. Chakraborty, S., M. Monfett, T. M. Maier, J. L. Benach, D. W. Frank, and D. G.
Thanassi. 2008. Type IV pili in Francisella tularensis: roles of pilF and pilT in fiber
assembly, host cell adherence, and virulence. Infect Immun 76:2852-61.
6. Craig, L., and J. Li. 2008. Type IV pili: paradoxes in form and function. Curr Opin
Struct Biol.
7. Craig, L., R. K. Taylor, M. E. Pique, B. D. Adair, A. S. Arvai, M. Singh, S. J. Lloyd,
D. S. Shin, E. D. Getzoff, M. Yeager, K. T. Forest, and J. A. Tainer. 2003. Type IV
pilin structure and assembly: X-ray and EM analyses of Vibrio cholerae toxin-
coregulated pilus and Pseudomonas aeruginosa PAK pilin. Mol Cell 11:1139-50.
on February 18, 2020 by guest
http://iai.asm.org/
Dow
nloaded from

29
8. Craig, L., N. Volkmann, A. S. Arvai, M. E. Pique, M. Yeager, E. H. Egelman, and J.
A. Tainer. 2006. Type IV pilus structure by cryo-electron microscopy and
crystallography: implications for pilus assembly and functions. Mol Cell 23:651-62.
9. Dillard, J. P. 2006. Genetic manipulation of Neisseria gonorrhoeae. In R. Coico, T.
Kowalik, J. M. Quarles, B. Stevenson, R. K. Taylor, and A. E. Simons (ed.), Current
Protocols in Microbiology. John Wiley and Sons, Hoboken, New Jersey.
10. Forest, K. T. 2005. Structure and assembly of type IV pilins. In G. Waksman, M.
Caparon, and S. Hultgren (ed.), Structural Biology of Bacterial Pathogenesis. ASM Press,
Washington, D.C.
11. Hamilton, H. L., N. M. Dominguez, K. J. Schwartz, K. T. Hackett, and J. P. Dillard.
2005. Neisseria gonorrhoeae secretes chromosomal DNA via a novel type IV secretion
system. Mol Microbiol 55:1704-21.
12. Hazes, B., P. A. Sastry, K. Hayakawa, R. J. Read, and R. T. Irvin. 2000. Crystal
structure of Pseudomonas aeruginosa PAK pilin suggests a main-chain-dominated mode
of receptor binding. J Mol Biol 299:1005-17.
13. Herdendorf, T. J., D. R. McCaslin, and K. T. Forest. 2002. Aquifex aeolicus PilT,
homologue of a surface motility protein, is a thermostable oligomeric NTPase. J Bacteriol
184:6465-71.
14. Johnson, T. L., J. Abendroth, W. G. Hol, and M. Sandkvist. 2006. Type II secretion:
from structure to function. FEMS Microbiol Lett 255:175-86.
15. Keizer, D. W., C. M. Slupsky, M. Kalisiak, A. P. Campbell, M. P. Crump, P. A.
Sastry, B. Hazes, R. T. Irvin, and B. D. Sykes. 2001. Structure of a pilin monomer from
on February 18, 2020 by guest
http://iai.asm.org/
Dow
nloaded from

30
Pseudomonas aeruginosa: implications for the assembly of pili. J Biol Chem 276:24186-
93.
16. Kellogg, D. S., Jr., W. L. Peacock, Jr., W. E. Deacon, L. Brown, and D. I. Pirkle.
1963. Neisseria gonorrhoeae. I. Virulence Genetically Linked to Clonal Variation. J
Bacteriol 85:1274-9.
17. Kirn, T. J., M. J. Lafferty, C. M. Sandoe, and R. K. Taylor. 2000. Delineation of pilin
domains required for bacterial association into microcolonies and intestinal colonization
by Vibrio cholerae. Mol Microbiol 35:896-910.
18. Kohler, R., K. Schafer, S. Muller, G. Vignon, K. Diederichs, A. Philippsen, P.
Ringler, A. P. Pugsley, A. Engel, and W. Welte. 2004. Structure and assembly of the
pseudopilin PulG. Mol Microbiol 54:647-64.
19. Larkin, M. A., G. Blackshields, N. P. Brown, R. Chenna, P. A. McGettigan, H.
McWilliam, F. Valentin, I. M. Wallace, A. Wilm, R. Lopez, J. D. Thompson, T. J.
Gibson, and D. G. Higgins. 2007. Clustal W and Clustal X version 2.0. Bioinformatics
23:2947-8.
20. Mahony, D. E., and G. G. Kalz. 1968. A temperate bacteriophage of Clostridium
perfringens. Can J Microbiol 14:1085-93.
21. Maier, B. 2005. Using laser tweezers to measure twitching motility in Neisseria. Curr
Opin Microbiol 8:344-9.
22. Mandel, J. L., and M. L. Pearson. 1974. Insulin stimulates myogenesis in a rat
myoblast line. Nature 251:618-20.
23. Mattick, J. S. 2002. Type IV pili and twitching motility. Annu Rev Microbiol 56:289-
314.
on February 18, 2020 by guest
http://iai.asm.org/
Dow
nloaded from

31
24. Melville, S. B., R. Labbe, and A. L. Sonenshein. 1994. Expression from the
Clostridium perfringens cpe promoter in C. perfringens and Bacillus subtilis. Infect
Immun 62:5550-8.
25. Mendez, M., I. H. Huang, K. Ohtani, R. Grau, T. Shimizu, and M. R. Sarker. 2008.
Carbon catabolite repression of type IV pilus-dependent gliding motility in the anaerobic
pathogen Clostridium perfringens. J Bacteriol 190:48-60.
26. Merz, A. J., D. B. Rifenbery, C. G. Arvidson, and M. So. 1996. Traversal of a
polarized epithelium by pathogenic Neisseriae: facilitation by type IV pili and
maintenance of epithelial barrier function. Mol Med 2:745-54.
27. O'Brien, D. K., and S. B. Melville. 2000. The anaerobic pathogen Clostridium
perfringens can escape the phagosome of macrophages under aerobic conditions. Cellular
Microbiol 2:505-519.
28. O'Toole, G. A., and R. Kolter. 1998. Flagellar and twitching motility are necessary for
Pseudomonas aeruginosa biofilm development. Mol Microbiol 30:295-304.
29. Parge, H. E., K. T. Forest, M. J. Hickey, D. A. Christensen, E. D. Getzoff, and J. A.
Tainer. 1995. Structure of the fibre-forming protein pilin at 2.6 A resolution. Nature
378:32-8.
30. Pepe, J. C., and S. Lory. 1998. Amino acid substitutions in PilD, a bifunctional enzyme
of Pseudomonas aeruginosa. Effect on leader peptidase and N-methyltransferase
activities in vitro and in vivo. J Biol Chem 273:19120-9.
31. Reguera, G., K. D. McCarthy, T. Mehta, J. S. Nicoll, M. T. Tuominen, and D. R.
Lovley. 2005. Extracellular electron transfer via microbial nanowires. Nature 435:1098-
101.
on February 18, 2020 by guest
http://iai.asm.org/
Dow
nloaded from

32
32. Rood, J. I. 1998. Virulence genes of Clostridium perfringens. Annu Rev Microbiol
52:333-60.
33. Salomonsson, E., A. Forsberg, N. Roos, C. Holz, B. Maier, M. Koomey, and H. C.
Winther-Larsen. 2009. Functional analyses of pilin-like proteins from Francisella
tularensis: complementation of type IV pilus phenotypes in Neisseria gonorrhoeae.
Microbiology 155:2546-59.
34. Segal, E., E. Billyard, M. So, S. Storzbach, and T. F. Meyer. 1985. Role of
chromosomal rearrangement in N. gonorrhoeae pilus phase variation. Cell 40:293-300.
35. Shi, J., T. L. Blundell, and K. Mizuguchi. 2001. FUGUE: sequence-structure homology
recognition using environment-specific substitution tables and structure-dependent gap
penalties. J Mol Biol 310:243-57.
36. Shimizu, T., K. Ohtani, H. Hirakawa, K. Ohshima, A. Yamashita, T. Shiba, N.
Ogasawara, M. Hattori, S. Kuhara, and H. Hayashi. 2002. Complete genome
sequence of Clostridium perfringens, an anaerobic flesh-eater. Proc. Natl. Acad. Sci. U S
A 99:996-1001.
37. Stevens, D. L. 1997. Necrotizing Clostridial soft tissue infections, p. 141-152. In J. I.
Rood, B. A. McClane, J. G. Songer, and R. W. Titball (ed.), The Clostridia: Molecular
biology and pathognesis. Academic Press, San Diego, CA.
38. Strom, M. S., and S. Lory. 1992. Kinetics and sequence specificity of processing of
prepilin by PilD, the type IV leader peptidase of Pseudomonas aeruginosa. J Bacteriol
174:7345-51.
on February 18, 2020 by guest
http://iai.asm.org/
Dow
nloaded from

33
39. Varga, J. J., V. Nguyen, D. K. O'Brien, K. Rodgers, R. A. Walker, and S. B.
Melville. 2006. Type IV pili-dependent gliding motility in the Gram-positive pathogen
Clostridium perfringens and other Clostridia. Mol Microbiol 62:680-94.
40. Varga, J. J., B. Therit, and S. B. Melville. 2008. Type IV pili and the CcpA protein are
needed for maximal biofilm formation by the gram-positive anaerobic pathogen
Clostridium perfringens. Infect Immun 76:4944-51.
41. Vignon, G., R. Kohler, E. Larquet, S. Giroux, M. C. Prevost, P. Roux, and A. P.
Pugsley. 2003. Type IV-like pili formed by the type II secreton: specificity, composition,
bundling, polar localization, and surface presentation of peptides. J Bacteriol 185:3416-
28.
42. Wilgenbusch, J. C., and D. Swofford. 2003. Inferring evolutionary trees with PAUP*.
Curr Protoc Bioinformatics Chapter 6:Unit 6 4.
43. Winther-Larsen, H. C., M. C. Wolfgang, J. P. van Putten, N. Roos, F. E. Aas, W. M.
Egge-Jacobsen, B. Maier, and M. Koomey. 2007. Pseudomonas aeruginosa Type IV
pilus expression in Neisseria gonorrhoeae: effects of pilin subunit composition on
function and organelle dynamics. J Bacteriol 189:6676-85.
44. Wolfgang, M., P. Lauer, H. S. Park, L. Brossay, J. Hebert, and M. Koomey. 1998.
PilT mutations lead to simultaneous defects in competence for natural transformation and
twitching motility in piliated Neisseria gonorrhoeae. Mol Microbiol 29:321-30.
45. Wolfgang, M., J. P. van Putten, S. F. Hayes, D. Dorward, and M. Koomey. 2000.
Components and dynamics of fiber formation define a ubiquitous biogenesis pathway for
bacterial pili. Embo J 19:6408-18.
on February 18, 2020 by guest
http://iai.asm.org/
Dow
nloaded from

34
46. Yaffe, D., and O. Saxel. 1977. Serial passaging and differentiation of myogenic cells
isolated from dystrophic mouse muscle. Nature 270:725-7.
on February 18, 2020 by guest
http://iai.asm.org/
Dow
nloaded from
35
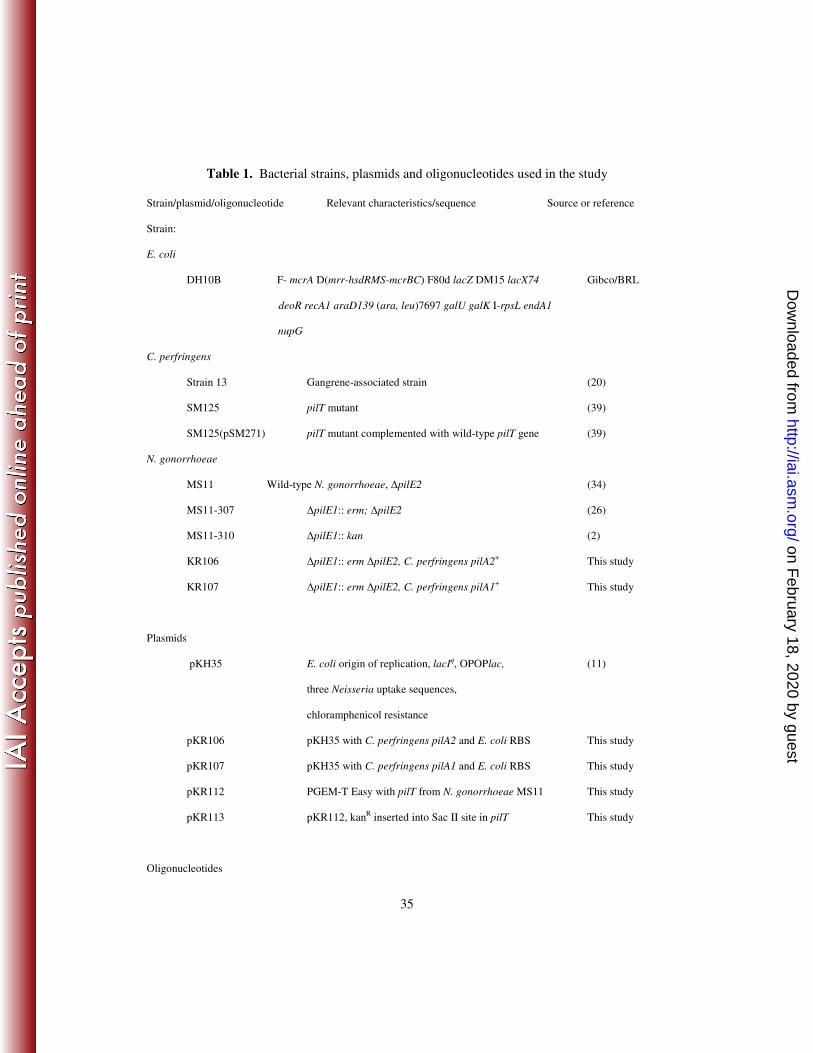
Table 1. Bacterial strains, plasmids and oligonucleotides used in the study
Strain/plasmid/oligonucleotide Relevant characteristics/sequence Source or reference
Strain:
E. coli
DH10B F- mcrA D(mrr-hsdRMS-mcrBC) F80d lacZ DM15 lacX74 Gibco/BRL
deoR recA1 araD139 (ara, leu)7697 galU galK I-rpsL endA1
nupG
C. perfringens
Strain 13 Gangrene-associated strain (20)
SM125 pilT mutant (39)
SM125(pSM271) pilT mutant complemented with wild-type pilT gene (39)
N. gonorrhoeae
MS11 Wild-type N. gonorrhoeae, ∆pilE2 (34)
MS11-307 ∆pilE1:: erm; ∆pilE2
(26)
MS11-310 ∆pilE1:: kan
(2)
KR106 ∆pilE1:: erm ∆pilE2, C. perfringens pilA2+ This study
KR107 ∆pilE1:: erm ∆pilE2, C. perfringens pilA1+
This study
Plasmids
pKH35 E. coli origin of replication, lacIq, OPOPlac, (11)
three Neisseria uptake sequences,
chloramphenicol resistance
pKR106 pKH35 with C. perfringens pilA2 and E. coli RBS This study
pKR107 pKH35 with C. perfringens pilA1 and E. coli RBS This study
pKR112 PGEM-T Easy with pilT from N. gonorrhoeae MS11 This study
pKR113 pKR112, kanR inserted into Sac II site in pilT This study
Oligonucleotides
on February 18, 2020 by guest
http://iai.asm.org/
Dow
nloaded from

36
OKR011 5′-CTTTAAAAAATAAAGCTTAGGAGGCATAAATGTTATTACTGAAAGC-3′
OKR013 5’-CATATTAAGCTTAGGAGGAAAACCAATGAATACAAAAAAAC-3’
OKR014 5′-CACCTTATTATTTTACTAATTCTAGAACTTTTAATACTATTGATTATTTC-3’
OKR019 5′-GCCCCTATGAAATCTAGATAATTAAAGTATATTTATACTATAGACACC-3′
OKR063 5’-CGAAACCATATGCTAAACAGAGCCGCA-3’
OKR064 5’-GCCTTGCCATGGCGCC-3’
OKR065 5’-CCGCGGCCGTCTGAACTCAAAATC-3’
OKR066 5’-CCGCGGTTGATGAGAGCTTTGTTGTAG-3’
on February 18, 2020 by guest
http://iai.asm.org/
Dow
nloaded from

37
Figure Legends
Figure 1. A. Phylogenetic tree showing relatedness of the conserved N-terminal domains of
pilin and pseudopilin proteins. Common colors denote evolutionarily related pilins. Bootstrap
values are shown on individual branches. They were calculated from 100 replicates using the
neighbor method in the PAUP 4.0 4a software (42). Note that PilA1 and PilA2 sequences
branched with a top score, suggesting they are an independent line of descent within the
pseudopilin or pilin group. The alignments used to generate this tree are shown in Fig. S1. B.
Alignment of pilin proteins PilA1 and PilA2 from C. perfringens strain 13 with PilE from N.
gonorrhoeae strain MS11 using the Clustal W 2.0 program (19). Residues with an asterisk are
identical in all three sequences, a colon are similar in all three sequences, a period are similar or
identical in 2 of 3 sequences. The arrow indicates the site of pre-pilin peptidase cleavage
(known in PilE; proposed in PilA1 and PilA2).
Figure 2. Western blot analysis showing heterologous expression of C. perfringens pilins in N.
gonorrhoeae. Whole cell lysates of colonies swabbed from plates were subjected to SDS-PAGE
followed by immunoblot with anti-PilA1 and anti-PilA2 antibodies. The location of molecular
weight standards are indicated on the left. (+) indicates the presence of IPTG; (-) the absence of
IPTG.
Figure 3. Immunofluorescence (red color) shows increased expression of PilA1 and PilA2 on
the surface of N. gonorrhoeae strains KR106 and KR107 in the presence of IPTG. All scale bars
= 2 µm. DIC, differential interference contrast microscopy.
on February 18, 2020 by guest
http://iai.asm.org/
Dow
nloaded from

38
Figure 4. Field emission-scanning electron microscopy of N. gonorrhoeae strains cultured on
0.2 µm membrane filters. A. Wild-type N. gonorrhoeae strain MS11. B. N. gonorrhoeae strain
KR106, a pilE- mutant expressing C. perfringens PilA2. C. Surface of a filter near strain KR106
cells; unattached pili can be seen on the surface of the filter. D. High magnification image of an
isolated PilA2 pilus from strain KR106. Region inside boxes are shown in higher magnification
in the insets in panels A and B. Bar = 1 µm for panels A-C; 100 nm for panel D.
Figure 5. Adherence of N. gonorrhoeae strains expressing C. perfringens pili. Adherence of N.
gonorrhoeae strains to (A) A431 cells, a human vulval carcinoma cell line, (B) C2C12 cells, a
mouse myoblast cell line, (C) L6 cells, a rat myoblast cell line (D) Mouse primary fibroblasts,
(E) MDCK cells, a canine kidney cell line. Values shown represent the mean and SD of at least
3 independent samples. Numerical values above each bar represent the P values, calculated
using a two tailed unpaired t test with Welch correction. Note differences in scale used in each
panel.
Figure 6. SEM of N. gonorrhoeae strains KR106 and KR107 adhering to mouse muscle cells.
C2C12 cells were seeded on glass coverslips and incubated with (A-B) N. gonorrhoeae strain
KR106 or (C) N. gonorrhoeae strain KR107, all with IPTG added. Bar = 1 µm.
Figure 7. Adherence of C. perfringens strains to C2C12 cells under anaerobic conditions.
Values shown represent the mean and SD of 10 independent samples. Numerical values above
on February 18, 2020 by guest
http://iai.asm.org/
Dow
nloaded from

39
each bar represent the P values, calculated using a two tailed unpaired t test with Welch
correction.
on February 18, 2020 by guest
http://iai.asm.org/
Dow
nloaded from