identification of phosphorylation sites within the sh3 domains of tec family tyrosine kinases
TRANSCRIPT

Identification of phosphorylation sites within the SH3 domains
of Tec family tyrosine kinases
Beston F. Norea,*, Pekka T. Mattssona,b, Per Antonssona, Carl-Magnus Backesjoa,Anna Westlundc, Johan Lennartssond, Henrik Hanssone, Peter Lowf,
Lars Ronnstrandd, C.I. Edvard Smitha
aKarolinska Institutet, Clinical Research Center (CRC) at Novum, Huddinge University Hospital, SE-141 86 Huddinge, SwedenbDepartment of Biochemistry and Food Chemistry, University of Turku, FIN-20014 Turku, Finland
cKaroBio AB, SE-141 57 Huddinge, SwedendLudwig Institute for Cancer Research, SE-751 24 Uppsala, Sweden
eDepartment of Biotechnology, Royal Institute of Technology (KTH), SCFAB, SE-106 91 Stockholm, SwedenfDepartment of Neuroscience, Karolinska Institutet, SE-171 77 Stockholm, Sweden
Received 19 July 2002; received in revised form 2 October 2002; accepted 6 November 2002
Abstract
Tec family protein tyrosine kinases (TFKs) play a central role in hematopoietic cellular signaling. Initial activation takes place through
specific tyrosine phosphorylation situated in the activation loop. Further activation occurs within the SH3 domain via a transphosphorylation
mechanism, which for Bruton’s tyrosine kinase (Btk) affects tyrosine 223. We found that TFKs phosphorylate preferentially their own SH3
domains, but differentially phosphorylate other member family SH3 domains, whereas non-related SH3 domains are not phosphorylated. We
demonstrate that SH3 domains are good and reliable substrates. We observe that transphosphorylation is selective not only for SH3 domains,
but also for dual SH3SH2 domains. However, the dual domain is phosphorylated more effectively. The major phosphorylation sites were
identified as conserved tyrosines, for Itk Y180 and for Bmx Y215, both sites being homologous to the Y223 site in Btk. There is, however,
one exception because the Tec-SH3 domain is phosphorylated at a non-homologous site, nevertheless a conserved tyrosine, Y206. Consistent
with these findings, the 3D structures for SH3 domains point out that these phosphorylated tyrosines are located on the ligand-binding
surface. Because a number of Tec family kinases are coexpressed in cells, it is possible that they could regulate the activity of each other
through transphosphorylation.
D 2002 Elsevier Science B.V. All rights reserved.
Keywords: Immunodeficiency; Tyrosine kinase; Btk; Itk; Tec; Bmx; Signal transduction; X-linked agammaglobulinemia; XLA
1. Introduction
Tec family protein tyrosine kinases (TFKs) together with
Src family kinases (SFKs) are multifunctional non-receptor
protein tyrosine kinases (PTKs), making up about 40–45%
of all cytoplasmic PTKs identified in the human genome
[1,2]. There are five related members of Tec PTKs in
humans, Bruton’s tyrosine kinase (Btk), Itk, Tec, Bmx and
Txk/Rlk. The domain architecture of these kinases consists
of five domains, three of which, SH3, SH2 and SH1 (kinase
domain), are found in the SFKs [2–5]. Apart from Txk,
which lacks the pleckstrin homology (PH) domain [6], the
presence of a PH domain and a Tec homology domain (TH)
at the N-terminal end distinguishes this family from SFKs
[2–5]. TH domains are comprised of a Btk motif and a
poly-proline (PP) region, which are thought to regulate and/
or stabilize kinase activity [7].
The central role of TFKs in cellular signaling was
directly recognized after identification of the BTK gene,
mutations in which cause the human immunodeficiency
1570-9639/02/$ - see front matter D 2002 Elsevier Science B.V. All rights reserved.
doi:10.1016/S1570-9639(02)00524-1
Abbreviations: XLA, X-linked agammaglobulinemia; Xid, X-linked
immunodeficiency; Btk, Bruton’s tyrosine kinase; PTK(s), protein tyrosine
kinase(s); PPSH3, extended Btk-SH3 domain with N-terminal poly-proline
regions; PP, poly-proline; Amp, Amphiphysin; IVK, in vitro kinase assay;
IP, immunoprecipitation; pY, phosphotyrosine; rhBtk, full-length recombi-
nant human Btk; 2D-mapping, two-dimensional tryptic phosphopeptide
mapping
* Corresponding author. Tel.: +46-8-5858-3657;
fax: +46-8-5858-3650.
E-mail address: [email protected] (B.F. Nore).
www.bba-direct.com
Biochimica et Biophysica Acta 1645 (2003) 123–132

disease, X-linked agammaglobulinemia (XLA) [8,9] and a
milder form, X-linked immunodeficiency (Xid), in mouse
[10,11]. XLA exhibits an intrinsic defect in B-cell develop-
ment and maturation at level pre-B transition, leading to a
dramatic reduction of peripheral B-cells [12]. The role of
Tec kinases in signal transduction is best illustrated for Btk,
particularly in the context of B-cell antigen receptor (BCR)
engagement. Itk plays an analogous role in T-cell signaling,
as shown for Itk knockout mice where T-cell development
and T-cell receptor signaling responses are impaired [13,14].
SFKs are ubiquitous PTKs regulating multiple cellular
signaling pathways [15,16]. It has been shown that SFKs
directly phosphorylate TFKs in the activation loop [17–19].
This phosphorylation site was mapped for human Btk on
Y551 [17] and for mouse Itk on Y511 [19]. Data from
multiple sequence alignments of TFKs deduced putative
homologous sites for other family members in human,
mouse and other species [20].
For Btk activation, membrane recruitment is a crucial
step associated with rapid, but transient, phosphorylation of
two major regulatory tyrosine residues, Y551 in the activa-
tion loop and subsequent autophosphorylation at Y223 in
the SH3 domain [17,21]. SH3 domain phosphorylation
provides a negative charge (as well as steric exclusion) to
this region, which can be severely unfavorable for the
interaction with ligand(s) [22]. The role of Btk-SH3 phos-
phorylation on Y223 in regulation of protein–protein inter-
actions and signal transduction has not yet been resolved,
despite the fact that numerous interacting proteins have been
identified [2,5,23]. It has been postulated that the two poly-
proline motifs in the TH domain stabilize and could interact
with the adjacent SH3 domain [24,25]. Recently, further
evidence provided an explanation to why the TH region of
Btk makes intermolecular (trans) interactions, whereas the
corresponding interaction in the related Itk kinase with only
one poly-proline motif is intramolecular (cis) [26–28].
However, it remains to be shown whether such an inter-
action takes place and/or influences signaling in vivo. In
Btk, the SH3 domain mutation Y223F results in enhanced
fibroblast transformation, implying that the SH3 domain
may play a negative regulatory role [21].
To determine a reliable substrate and to examine the
specificity for TFKs activity, we utilized a transphosphor-
ylation assay with recombinant SH3 proteins of Btk, Itk, Tec
and Bmx. For this purpose, we applied a steady-state in vitro
kinase assay (IVK) to follow the extent of g32P-ATP label-
ing of different SH3 domains at identical conditions. Our
initial analysis showed that transphosphorylation activity of
Btk is not restricted to its own SH3 domain, but also found
toward other family member SH3 domains. Like Btk, we
found that both Itk and Tec phosphorylate their own SH3
domains, but differentially transphosphorylate other TFK
members. The IVK activity of Btk, Itk and Tec is restricted
to SH3 domains of family members, whereas non-related
SH3 domains, such as Grb2 [29–31] and Amphiphysin
(Amp) [32], are not phosphorylated at any other tyrosine.
Our results show that the specificity in tyrosine phosphor-
ylation of TFK-SH3 domains in vitro is high. Therefore, we
believe that SH3 domain phosphorylation is required for a
distinct function in signaling.
2. Materials and methods
2.1. Cell culture and antibodies
The human B-cell line Ramos and T-cell Line Molt4
were cultured in RPMI 1640 supplemented with 10% FBS,
50 Ag/ml streptomycin and 50 units/ml penicillin (Gibco-
Invitrogen). Anti-Btk and anti-Itk were polyclonal rabbit
antibodies raised against their C- and N-terminal regions,
respectively. Anti-Tec was a polyclonal antibody kindly
provided by Dr. H. Mano (Jichi Medical School, Japan).
2.2. Cloning
The human Btk, Tec, Itk and Bmx SH3 domain con-
structs were cloned in pGEX-4T-3 vector as described
earlier [22]. A DNA fragment encoding residues 212–275
of hBtk containing tyrosine 223 mutated to phenylalanine
was amplified by PCR from pBluescript-hBtk Y223A. The
mutation was created by the overlap extension PCR techni-
que [33]. The fragment was cloned into pGEX-4T-3 vector
(Pharmacia). The sequence and presence of the mutation
was verified by sequencing. The human Btk gene was
cloned in baculovirus expression system pFASTBAC HT
with N-terminal 6xHis tag (Invitrogen).
2.3. Expression and purification of SH3 domains
Expression of protein was induced by the addition of
isopropyl D-thiogalactopyranoside (IPTG) to the XL1-Blue
strain of Escherichia coli [34]. After induction, the cells were
harvested and lysed by sonication at 4 jC in PBS buffer
containing 1% Triton X-100, 0.1 mM aprotinin, 1 mM
leupeptin and 1 mM EDTA. The GST-SH3 fusion products
were bound to glutathione-Sepharose, and digested with 100
units thrombin (Sigma-Aldrich). The GST-SH3 fusion pro-
teins were directly concentrated for gel filtration on a Super-
dex-200 column (Pharmacia). The purified Tec family SH3
proteins were finally concentrated and the protein solution
was stored at � 70 jC. The Btk (Y223A) SH3, Grb2-SH3
(AA159–217) and Amp-SH3 (AA545–694) proteins were
purified using the same procedure.
2.4. Expression and purification of human Btk (hBtk)
Spodoptera frugiperda (Sf9) cells (Invitrogen) were
maintained as suspension cultures in siliconized (Sig-
macotek, Sigma-Aldrich) 5 L shake flasks in 1 L of Sf900II
serum-free medium (Gibco-Invitrogen). When the culture
reached a cell density of 2� 106 cells ml� 1, the cells were
B.F. Nore et al. / Biochimica et Biophysica Acta 1645 (2003) 123–132124

infected at a multiplicity of infection of 5 with a recombinant
Autographa californica nuclear polyhedrosis baculovirus
containing the Btk gene. After 48 h, the cells were harvested
and then immediately frozen in liquid nitrogen and stored at
� 70 jC until processed. The production of full-length
recombinant human Btk (rhBtk) was performed by melting
the insect cell pellet lysate buffer containing 50 mM HEPES,
pH 8.0, 10 mM h-mercaptoethanol, 1 mM EDTA, 1 mM
PMSF, 1% NP-40, 0.5% Na-deoxycholate, 0.1 mM aproti-
nin, 1 mM leupeptin, 300 mM NaCl and two EDTA-free
Complete inhibitor cocktail tablets (Roche). The cells were
disrupted by mild sonic oscillation (Raytheon, USA) and the
cell debris was pelleted by centrifugation. The 6x polyhis-
tidine stretch in the N-terminal rhBtk facilitated the Btk
purification using Ni-affinity column (Qiagen, USA) (J.A.
Marquez, manuscript in preparation). The polyhistidine
stretch was cleaved by rTEV protease and Btk was further
purified on a Superdex-200 column (Pharmacia).
2.5. Immunoprecipitations (IP) and in vitro kinase assay
Molt4 were used for immunoprecipitation of Itk, and
Ramos cells for Btk and Tec. One to 5� 107 cells were
lysed in modified RIPA buffer (50 mM HEPES-buffer pH
7.5, 1% digitonine, 1 mM Na3VO4, 1 mM Na2MoO4, 1 mM
PMSF, 1 mM DTT) and one tablet of EDTA-free Complete
inhibitor cocktail (Roche) per 5 ml solution. Btk, Itk or Tec
was immunoprecipitated by incubation with 5 Ag of poly-
clonal antibodies for 1.5 h with rotation, at 4 jC. Followingaddition of 40 Al Protein A SepharoseR, the suspensions
were rocked for 2 h at 4 jC. Beads were washed five times
for 5 min each with modified RIPA buffer without digito-
nine. Subsequent to the IP procedure, immunoprecipitated
beads with Btk, Itk and Tec were washed twice with kinase
buffer (50 mM HEPES-buffer, pH 7.5; 10 mM MnCl2, 4
mM MgCl2, 0.05 mM Na3VO4, 0.1 mM Na2MoO4, 0.1 mM
PMSF, 1 mM DTT). The beads were resuspended in 20 Alkinase buffer containing 50 AM cold ATP and 1–10 Agrecombinant proteins. In a similar manner, in vitro kinase
with purified hBtk, 200 ng of purified Btk combined with
20 Al kinase buffer containing 50 AM cold ATP and 5–20
Ag recombinant proteins. Finally, 1–20 ACi g32P-ATP was
added to the reaction mixtures with gentle vortex for 20
min. Reactions were terminated by the addition of SDS-
polyacrylamide gel loading buffer. Proteins were resolved
on 16% SDS-PAGE and transferred to either HQ-PVDF or
nitrocellulose membranes (Millipore).
2.6. Analysis of tryptic phosphopeptide and radio-Edman
sequencing
Tryptic phosphopeptides were digested on nitrocellulose
membranes according to methods previously described by
Hansen et al. [35] and they were separated on a Hunter thin
layer electrophoresis system (HTLE-700) at 2000 V for 40
min under water cooling. The second dimension chromatog-
raphy was performed in isobutyric acid buffer for 16 h
[36,37]. Briefly, nitrocellulose membrane slices of 32P-
labeled SH3 protein were excised, washed extensively with
double distilled H2O, and then soaked in 0.5% polyvinylpyr-
olidone (Sigma) for 30 min at 37 jC. The membranes were
then washed four times with 50mMNH4HCO3. Immobilized
proteins were digested twice with 1 Ag trypsin (Promega) in
200 Al 50 mM NH4HCO3. Supernatants were separated and
the membranes were washed twice with NH4HCO3 and then
vacuum dried. Dried pellets were dissolved in 7 Al, pH 1.9
buffer (4.6 formic acid:15.6 acetic acid:18 H2O). The phos-
phopeptides were detected with a Fuji phosphoimager. Eluted
phosphopeptides [35] were coupled to Sequelonk-AAmem-
brane (Millipore) by use of carbodiimide coupling, according
to standard procedures as described by the manufacturer.
Radio-Edman degradation was performed using an Applied
Biosystems gas-phase sequencer (Model 477A) as described
by Blume-Jensen et al. [38].
3. Results
3.1. Purification of SH3 domains
Introducing purified SH3 domains of Btk, Itk, Tec and
Bmx enable us to study transphosphorylation and to identify
a reliable substrate in vitro for quantifying kinase activity.
The SH3 domains were expressed and purified to homoge-
neity on glutathione-Sepharose columns followed by gel
filtration (Fig. 1A). Three Btk-SH3 domain variants were
Fig. 1. Purification of recombinant proteins. (A) Proteins were fractionated
on the Phastgels (8–20% gradient gels) system and stained with Coomassie
blue G-250. SH3 domains of GST-Grb2 (C-terminal) and GST-Amphiphy-
sin (Amp-SH3) domains were thrombin cleaved without further purifica-
tion. The quantity of protein loaded was between 9 and 12 Ag per lane. (B)
Autophosphorylation of rhBtk in vitro. The g32P-ATP-labeled rhBtk was
separated on 7.5% SDS-PAGE and blotted on PVDF membrane. The
amount of Btk loaded was 100 ng per lane. Lanes 1–3 represent three
batches of purified human Btk. Radiolabeled rhBtK were detected and
visualized by autoradiography.
B.F. Nore et al. / Biochimica et Biophysica Acta 1645 (2003) 123–132 125
produced, a wild-type SH3 domain, a Y223A mutation form
lacking the transphosphorylation site [21] and an extended
version of Btk-SH3 domain (PPSH3), containing both PP
region (GSSHRKTKKPLPPTPEEDQILKKPLPPEPA-
AAPV-) at the N-terminal. For recombinant Btk production,
human full-length Btk protein (rhBtk) was expressed in Sf9
insect cells using a baculovirus vector. The rhBtk was
purified with a Ni-affinity resin and was found to have
intact kinase activity (Fig. 1B).
3.2. Transphosphorylation specificity of Btk
Initial examination of SH3 domain phosphorylation was
done by IVK assay using immunoprecipitation of endoge-
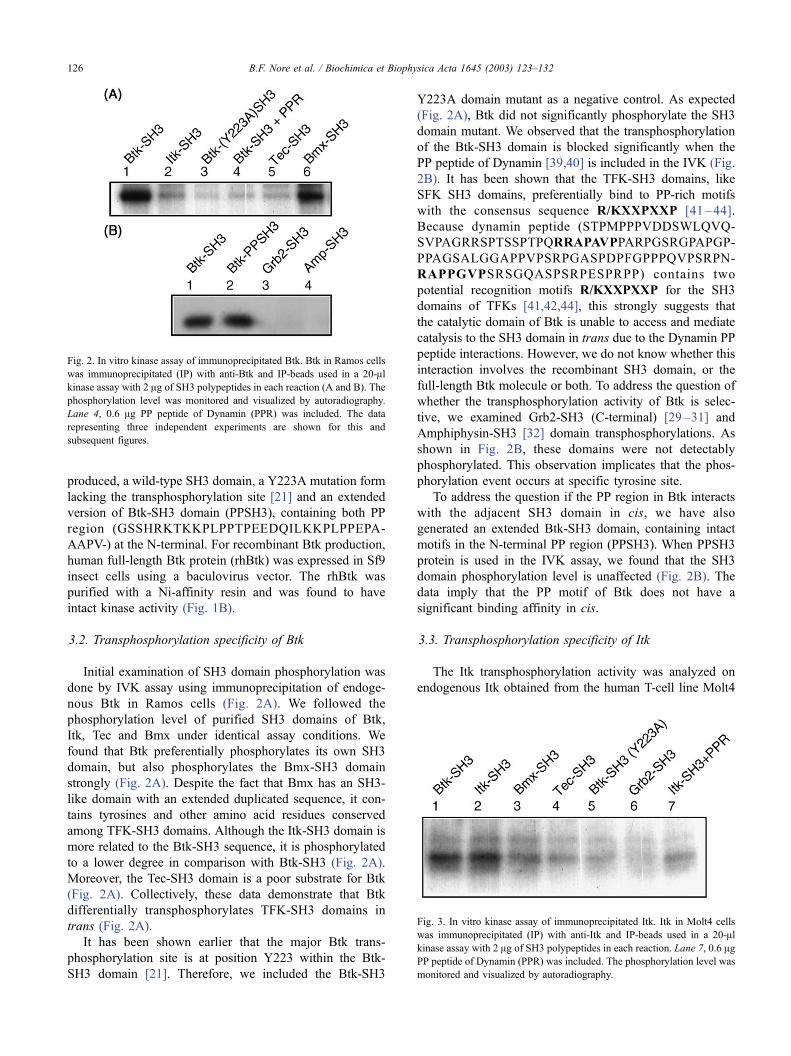
nous Btk in Ramos cells (Fig. 2A). We followed the
phosphorylation level of purified SH3 domains of Btk,
Itk, Tec and Bmx under identical assay conditions. We
found that Btk preferentially phosphorylates its own SH3
domain, but also phosphorylates the Bmx-SH3 domain
strongly (Fig. 2A). Despite the fact that Bmx has an SH3-
like domain with an extended duplicated sequence, it con-
tains tyrosines and other amino acid residues conserved
among TFK-SH3 domains. Although the Itk-SH3 domain is
more related to the Btk-SH3 sequence, it is phosphorylated
to a lower degree in comparison with Btk-SH3 (Fig. 2A).
Moreover, the Tec-SH3 domain is a poor substrate for Btk
(Fig. 2A). Collectively, these data demonstrate that Btk
differentially transphosphorylates TFK-SH3 domains in
trans (Fig. 2A).
It has been shown earlier that the major Btk trans-
phosphorylation site is at position Y223 within the Btk-
SH3 domain [21]. Therefore, we included the Btk-SH3
Y223A domain mutant as a negative control. As expected
(Fig. 2A), Btk did not significantly phosphorylate the SH3
domain mutant. We observed that the transphosphorylation
of the Btk-SH3 domain is blocked significantly when the
PP peptide of Dynamin [39,40] is included in the IVK (Fig.
2B). It has been shown that the TFK-SH3 domains, like
SFK SH3 domains, preferentially bind to PP-rich motifs
with the consensus sequence R/KXXPXXP [41–44].
Because dynamin peptide (STPMPPPVDDSWLQVQ-
SVPAGRRSPTSSPTPQRRAPAVPPARPGSRGPAPGP-
PPAGSALGGAPPVPSRPGASPDPFGPPPQVPSRPN-
RAPPGVPSRSGQASPSRPESPRPP) contains two
potential recognition motifs R/KXXPXXP for the SH3
domains of TFKs [41,42,44], this strongly suggests that
the catalytic domain of Btk is unable to access and mediate
catalysis to the SH3 domain in trans due to the Dynamin PP
peptide interactions. However, we do not know whether this
interaction involves the recombinant SH3 domain, or the
full-length Btk molecule or both. To address the question of
whether the transphosphorylation activity of Btk is selec-
tive, we examined Grb2-SH3 (C-terminal) [29–31] and
Amphiphysin-SH3 [32] domain transphosphorylations. As
shown in Fig. 2B, these domains were not detectably
phosphorylated. This observation implicates that the phos-
phorylation event occurs at specific tyrosine site.
To address the question if the PP region in Btk interacts
with the adjacent SH3 domain in cis, we have also
generated an extended Btk-SH3 domain, containing intact
motifs in the N-terminal PP region (PPSH3). When PPSH3
protein is used in the IVK assay, we found that the SH3
domain phosphorylation level is unaffected (Fig. 2B). The
data imply that the PP motif of Btk does not have a
significant binding affinity in cis.
3.3. Transphosphorylation specificity of Itk
The Itk transphosphorylation activity was analyzed on
endogenous Itk obtained from the human T-cell line Molt4
Fig. 2. In vitro kinase assay of immunoprecipitated Btk. Btk in Ramos cells
was immunoprecipitated (IP) with anti-Btk and IP-beads used in a 20-Alkinase assay with 2 Ag of SH3 polypeptides in each reaction (A and B). The
phosphorylation level was monitored and visualized by autoradiography.
Lane 4, 0.6 Ag PP peptide of Dynamin (PPR) was included. The data
representing three independent experiments are shown for this and
subsequent figures.
Fig. 3. In vitro kinase assay of immunoprecipitated Itk. Itk in Molt4 cells
was immunoprecipitated (IP) with anti-Itk and IP-beads used in a 20-Alkinase assay with 2 Ag of SH3 polypeptides in each reaction. Lane 7, 0.6 AgPP peptide of Dynamin (PPR) was included. The phosphorylation level was
monitored and visualized by autoradiography.
B.F. Nore et al. / Biochimica et Biophysica Acta 1645 (2003) 123–132126

(Fig. 3). We found that Itk efficiently phosphorylates both
Btk and Itk SH3 domains (Fig. 3). In other words, both Itk
and Btk SH3 domains are good substrates for Itk in vitro.
This observation raises the possibility that Itk and Btk might
regulate each other in cells coexpressing these kinases, such
as mast cells. Itk phosphorylated Bmx-SH3 to a low extent
(Fig. 3), in contrast to Btk, which efficiently phosphorylates
Bmx-SH3 (Fig. 2A). The Tec-SH3 domain is the least
suitable Itk substrate of the four TFK-SH3 domains used
(Fig. 3, lane 4). Furthermore, we observe that the Btk-SH3
mutant Y223A was not phosphorylated by Itk (Fig. 3, lane
5). Itk was unable to phosphorylate two non-conserved
tyrosines at the C-terminal Grb2-SH3 domain (Fig. 3, lane
6). This is in accordance with the idea that TFK-SH3
domains are the preferred substrates. The phosphorylation
level is reduced considerably by addition of Dynamin PP
peptide (Fig. 3, lane 7).
3.4. Transphosphorylation specificity of Tec
The endogenous kinase activity of Tec was investigated
from immunoprecipitation of Tec in Ramos cell extracts. In
a similar manner, we performed the kinase assay using
purified TFK-SH3 domains to determine a potential sub-
strate. Here, we noticed that Btk-SH3 is phosphorylated to a
similar extent as the Tec-SH3 domain (Fig. 4), but Btk does
not phosphorylate the Tec-SH3 domain appreciably (Fig.
2A). This observation may suggest that Tec has a broader
substrate specificity reflecting the broader tissue expression
of Tec. In addition, we found that Itk-SH3 and Bmx-SH3
domains are also phosphorylated by Tec (Fig. 4), but to a
smaller extent. Moreover, we could show that Tec does not
phosphorylate the Amphiphysin-SH3 domain, implying that
the location of tyrosine phosphorylation is conserved (Fig.
4, lane 3).
3.5. Transphosphorylation activity of rhBtk
For the subsequent experiments, we performed the IVK
assay using purified full-length rhBtk (Fig. 1B) to deter-
mine the transphosphorylation specificity and the consis-
tency with IP experiments of endogenous Btk. When we
performed the IVK assay for rhBtk with SH3 domain
substrates at identical concentrations as in the IP experi-
ments (Fig. 2A), namely 1–2 Ag, we found that the
transphosphorylation pattern is equivalent (not shown).
Moreover, the transphosphorylation pattern was found to
be consistent, even if the substrates are scaled-up 20 times
(Fig. 5). These observations demonstrate that the catalytic
activity of rhBtk has an identical substrate preference and
specificity as endogenous protein obtained from Ramos
cells (Fig. 2A).
3.6. Identification of transphosphorylation site(s)
To further characterize the recognition motif of phos-
phorylation, we decided to identify the phosphotyrosine
(pY) residue(s) in the SH3 domains. For this purpose, we
applied two-dimensional tryptic phosphopeptide mapping
(2D-mapping) using Hunter thin layer electrophoresis to
separate and resolve short phosphopeptides on TLC cellu-
lose plates.
In Fig. 6, we have summarized the results from 2D-
mapping experiments for Btk-SH3, Itk-SH3, Bmx-SH3 and
Tec-SH3 domains. The Btk-SH3 tryptic phosphopeptides
resolved on the 2D-TLC plate to three peptides labeled with32P (Fig. 6A). Among these phosphopeptides, only two show
a clear peptide sequence of VVALYDYMPMNANDLQL
and the pY was only identified in cycle 5, corresponding to
Y223 (Fig. 6A, inset) and this is in agreement with earlier
findings of the major transphosphorylation site within the
Btk-SH3 domain [21]. Here, we could confirm that Y223 is
the only site in the Btk-SH3 domain being detectably phos-
phorylated. In particular, we could exclude the possibility that
Y225 becomes phosphorylated, because the corresponding
tyrosine in Itk is conserved [20].
In a similar manner, we have analyzed Itk-SH3 tryptic
phosphopeptides (Fig. 6B) and the 2D-mapping was
resolved to one strong spot and a diffuse smeared band.
Fig. 4. In vitro kinase assay of immunoprecipitated Tec. Tec in Ramos cell
was immunoprecipitated (IP) with anti-Tec and IP-beads used in a 20-Alkinase assay with 2 Ag of SH3 polypeptides in each reaction. The
phosphorylation level was monitored and visualized by autoradiography.
Fig. 5. In vitro kinase assay of rhBtk. Identical quantities of full-length
rhBtk protein (200 ng) were applied in a 20-Al kinase assay together with
20 Ag of SH3 polypeptides as substrates in each reaction. The
phosphorylation level was monitored and visualized by autoradiography.
Phosphopeptide bands were digested with trypsin and subjected to 2D-
mapping analysis (described in Materials and methods).
B.F. Nore et al. / Biochimica et Biophysica Acta 1645 (2003) 123–132 127
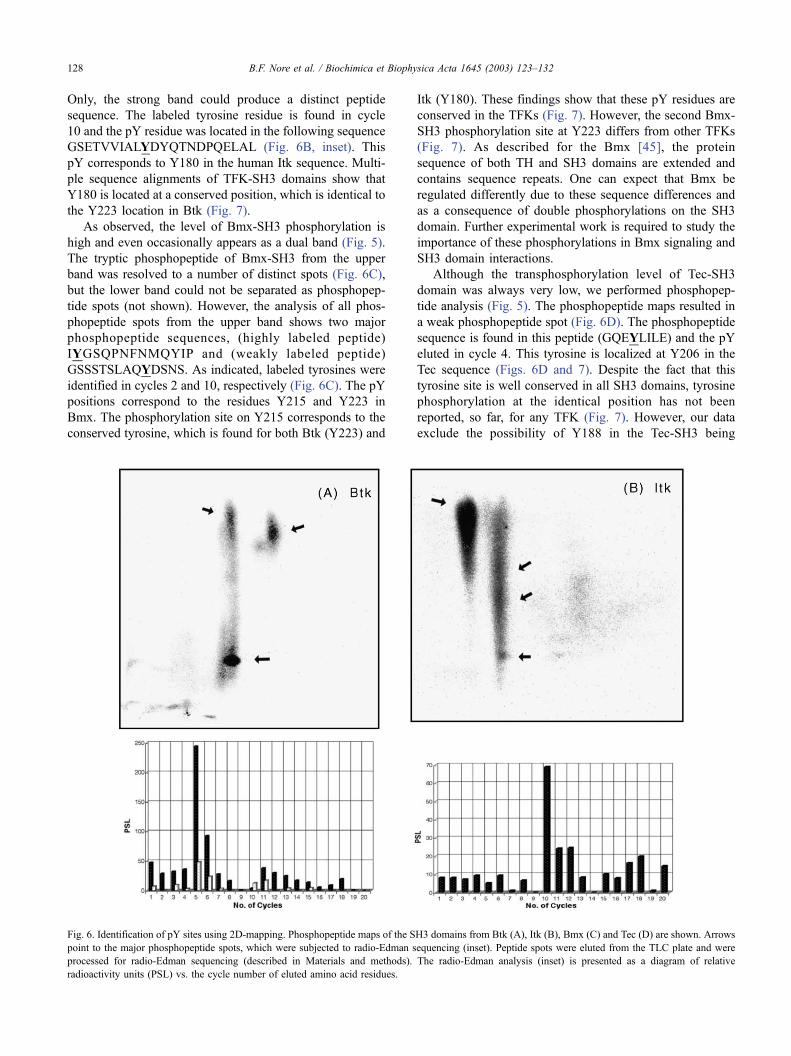
Only, the strong band could produce a distinct peptide
sequence. The labeled tyrosine residue is found in cycle
10 and the pY residue was located in the following sequence
GSETVVIALYDYQTNDPQELAL (Fig. 6B, inset). This
pY corresponds to Y180 in the human Itk sequence. Multi-
ple sequence alignments of TFK-SH3 domains show that
Y180 is located at a conserved position, which is identical to
the Y223 location in Btk (Fig. 7).
As observed, the level of Bmx-SH3 phosphorylation is
high and even occasionally appears as a dual band (Fig. 5).
The tryptic phosphopeptide of Bmx-SH3 from the upper
band was resolved to a number of distinct spots (Fig. 6C),
but the lower band could not be separated as phosphopep-
tide spots (not shown). However, the analysis of all phos-
phopeptide spots from the upper band shows two major
phosphopeptide sequences, (highly labeled peptide)
IYGSQPNFNMQYIP and (weakly labeled peptide)
GSSSTSLAQYDSNS. As indicated, labeled tyrosines were
identified in cycles 2 and 10, respectively (Fig. 6C). The pY
positions correspond to the residues Y215 and Y223 in
Bmx. The phosphorylation site on Y215 corresponds to the
conserved tyrosine, which is found for both Btk (Y223) and
Itk (Y180). These findings show that these pY residues are
conserved in the TFKs (Fig. 7). However, the second Bmx-
SH3 phosphorylation site at Y223 differs from other TFKs
(Fig. 7). As described for the Bmx [45], the protein
sequence of both TH and SH3 domains are extended and
contains sequence repeats. One can expect that Bmx be
regulated differently due to these sequence differences and
as a consequence of double phosphorylations on the SH3
domain. Further experimental work is required to study the
importance of these phosphorylations in Bmx signaling and
SH3 domain interactions.
Although the transphosphorylation level of Tec-SH3
domain was always very low, we performed phosphopep-
tide analysis (Fig. 5). The phosphopeptide maps resulted in
a weak phosphopeptide spot (Fig. 6D). The phosphopeptide
sequence is found in this peptide (GQEYLILE) and the pY
eluted in cycle 4. This tyrosine is localized at Y206 in the
Tec sequence (Figs. 6D and 7). Despite the fact that this
tyrosine site is well conserved in all SH3 domains, tyrosine
phosphorylation at the identical position has not been
reported, so far, for any TFK (Fig. 7). However, our data
exclude the possibility of Y188 in the Tec-SH3 being
Fig. 6. Identification of pY sites using 2D-mapping. Phosphopeptide maps of the SH3 domains from Btk (A), Itk (B), Bmx (C) and Tec (D) are shown. Arrows
point to the major phosphopeptide spots, which were subjected to radio-Edman sequencing (inset). Peptide spots were eluted from the TLC plate and were
processed for radio-Edman sequencing (described in Materials and methods). The radio-Edman analysis (inset) is presented as a diagram of relative
radioactivity units (PSL) vs. the cycle number of eluted amino acid residues.
B.F. Nore et al. / Biochimica et Biophysica Acta 1645 (2003) 123–132128

phosphorylated as residue, which is equivalent to the
conserved Btk-Y223, Itk-Y180 and Bmx-Y215 (Fig. 7).
Taken together, these data show that Tec-SH3 is not
phosphorylated at the common site found in the other
family members.
In addition to previously identified structures of Btk-SH3
[22] and Itk-SH3 domain [25], the Tec-SH3 domain struc-
ture has recently been published [46]. These structural
coordinates were used to visualize the conserved tyrosines
from these SH3 domains and the phosphorylated tyrosine
sites found in this work are decorated (Fig. 8). From the
structural model (Fig. 8), we found the Y206 on Tec-SH3
domain is positioned at the same side as Y223 in Btk and
Y180 in Itk. Our SH3 domain models show that the pY
identified in this work is located on the same surface. This
implies that these conserved tyrosines define a common
binding surface required for a specific interaction, which is
conserved between the TFK members.
3.7. Transphosphorylation of the SH3SH2 tandem domain
compared to the single SH3 domain
As described in earlier sections, the TFK-SH3 domains
are specifically phosphorylated, although the degree of
Fig. 7. Multiple sequence alignments of pY site in the Tec family SH3 domains. Multiple alignments of SH3 domains indicating phosphorylation sites (black
box) identified in this study. Sequence alignment was performed as described in Nore et al. [20].
Fig. 6 (continued ).
B.F. Nore et al. / Biochimica et Biophysica Acta 1645 (2003) 123–132 129
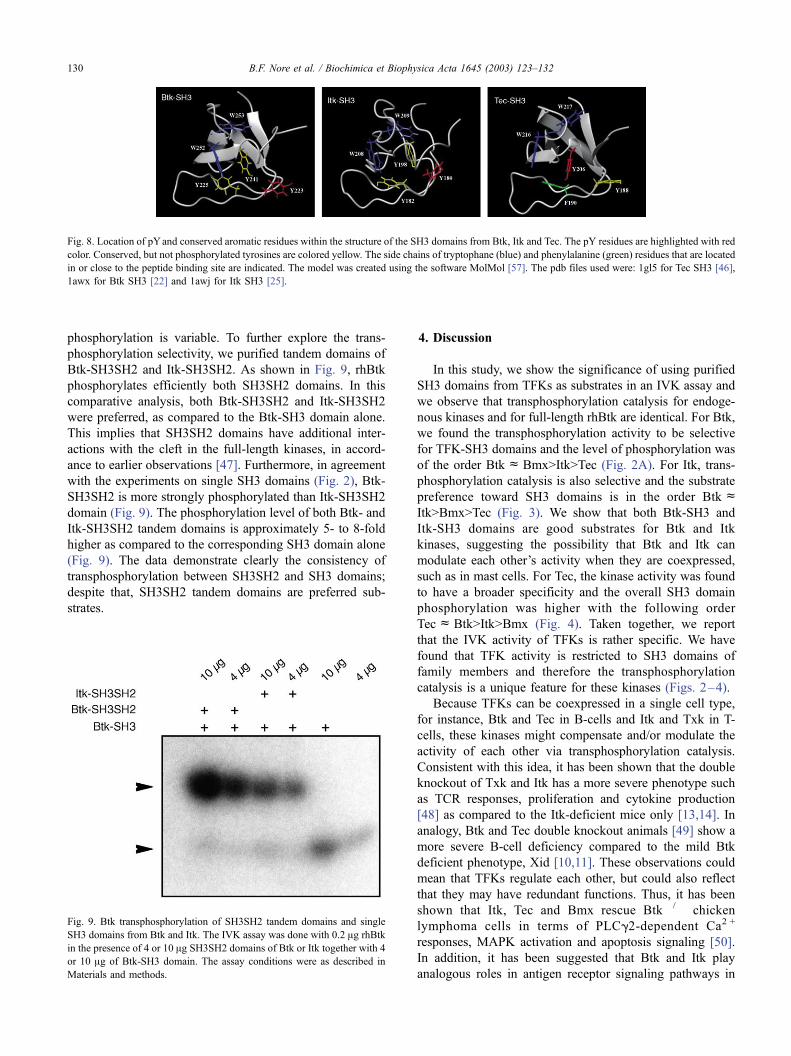
phosphorylation is variable. To further explore the trans-
phosphorylation selectivity, we purified tandem domains of
Btk-SH3SH2 and Itk-SH3SH2. As shown in Fig. 9, rhBtk
phosphorylates efficiently both SH3SH2 domains. In this
comparative analysis, both Btk-SH3SH2 and Itk-SH3SH2
were preferred, as compared to the Btk-SH3 domain alone.
This implies that SH3SH2 domains have additional inter-
actions with the cleft in the full-length kinases, in accord-
ance to earlier observations [47]. Furthermore, in agreement
with the experiments on single SH3 domains (Fig. 2), Btk-
SH3SH2 is more strongly phosphorylated than Itk-SH3SH2
domain (Fig. 9). The phosphorylation level of both Btk- and
Itk-SH3SH2 tandem domains is approximately 5- to 8-fold
higher as compared to the corresponding SH3 domain alone
(Fig. 9). The data demonstrate clearly the consistency of
transphosphorylation between SH3SH2 and SH3 domains;
despite that, SH3SH2 tandem domains are preferred sub-
strates.
4. Discussion
In this study, we show the significance of using purified
SH3 domains from TFKs as substrates in an IVK assay and
we observe that transphosphorylation catalysis for endoge-
nous kinases and for full-length rhBtk are identical. For Btk,
we found the transphosphorylation activity to be selective
for TFK-SH3 domains and the level of phosphorylation was
of the order BtkcBmx>Itk>Tec (Fig. 2A). For Itk, trans-
phosphorylation catalysis is also selective and the substrate
preference toward SH3 domains is in the order BtkcItk>Bmx>Tec (Fig. 3). We show that both Btk-SH3 and
Itk-SH3 domains are good substrates for Btk and Itk
kinases, suggesting the possibility that Btk and Itk can
modulate each other’s activity when they are coexpressed,
such as in mast cells. For Tec, the kinase activity was found
to have a broader specificity and the overall SH3 domain
phosphorylation was higher with the following order
TeccBtk>Itk>Bmx (Fig. 4). Taken together, we report
that the IVK activity of TFKs is rather specific. We have
found that TFK activity is restricted to SH3 domains of
family members and therefore the transphosphorylation
catalysis is a unique feature for these kinases (Figs. 2–4).
Because TFKs can be coexpressed in a single cell type,
for instance, Btk and Tec in B-cells and Itk and Txk in T-
cells, these kinases might compensate and/or modulate the
activity of each other via transphosphorylation catalysis.
Consistent with this idea, it has been shown that the double
knockout of Txk and Itk has a more severe phenotype such
as TCR responses, proliferation and cytokine production
[48] as compared to the Itk-deficient mice only [13,14]. In
analogy, Btk and Tec double knockout animals [49] show a
more severe B-cell deficiency compared to the mild Btk
deficient phenotype, Xid [10,11]. These observations could
mean that TFKs regulate each other, but could also reflect
that they may have redundant functions. Thus, it has been
shown that Itk, Tec and Bmx rescue Btk� / � chicken
lymphoma cells in terms of PLCg2-dependent Ca2 +
responses, MAPK activation and apoptosis signaling [50].
In addition, it has been suggested that Btk and Itk play
analogous roles in antigen receptor signaling pathways in
Fig. 9. Btk transphosphorylation of SH3SH2 tandem domains and single
SH3 domains from Btk and Itk. The IVK assay was done with 0.2 Ag rhBtk
in the presence of 4 or 10 Ag SH3SH2 domains of Btk or Itk together with 4
or 10 Ag of Btk-SH3 domain. The assay conditions were as described in
Materials and methods.
Fig. 8. Location of pYand conserved aromatic residues within the structure of the SH3 domains from Btk, Itk and Tec. The pY residues are highlighted with red
color. Conserved, but not phosphorylated tyrosines are colored yellow. The side chains of tryptophane (blue) and phenylalanine (green) residues that are located
in or close to the peptide binding site are indicated. The model was created using the software MolMol [57]. The pdb files used were: 1gl5 for Tec SH3 [46],
1awx for Btk SH3 [22] and 1awj for Itk SH3 [25].
B.F. Nore et al. / Biochimica et Biophysica Acta 1645 (2003) 123–132130

accordance with a reciprocal pattern of expression in B-cells
and T-cells, respectively [2,51]. However, human B cells,
such as the Ramos cell line used in this study, coexpress Btk
and Tec, and yet the phenotype in XLA is very severe. This
argues against redundant functions although the expression
level may be a significant factor.
To determine the location of SH3 phosphorylation sites,
we carried out phosphopeptide mapping of Btk, Itk, and
Bmx and Tec SH3 domains subsequent to transphosphor-
ylation. From tryptic peptide mapping analysis of Btk, we
confirmed that the Btk-SH3 phosphorylation site is located at
Y223 (Fig. 6A), a result identical to that achieved by the
previous phospho-Btk mapping [21]. We demonstrate here
that the transphosphorylation site of Itk-SH3 is located at
Y180 (Fig. 6B). In addition, sequence homology alignments
show that Itk-Y180 corresponds to Btk-Y223 [20]. For Bmx,
we obtained two phosphorylated sites, Y215 and Y223 (Fig.
6C). The Bmx-Y215 is a conserved tyrosine, which is
homologous to Btk-Y223 and Itk-Y180. In contrast, Bmx-
Y223 site is a unique site with no homology to SH3 domains
of other TFKs (Fig. 7). The Bmx-SH3 domain, unlike other
family members, has an extended repeat at the N-terminal
region [2,45] and the overall phosphorylation level is differ-
ent. On the other hand, Tec is widely expressed in different
tissues and was found to have a broad preference for the
TFK-SH3 domains, whereas Tec-SH3 protein was a poor
substrate for Btk and Itk kinases (Fig. 5). The weak trans-
phosphorylation site of Tec-SH3 domain was found at Y206
(Fig. 6D). The Y206 site is still a conserved tyrosine, which
has not been shown phosphorylated in any family member
SH3 domain (Fig. 7). On other hand, the common phos-
phorylated tyrosine found in Btk-Y223, Itk-Y180 and Bmx-
Y215 is not phosphorylated in the Tec-SH3 domain. The
majority of phosphorylation sites are conserved tyrosines
between all TFK members. They are located at a common
binding surface site, as illustrated from the three-dimensional
model for Btk, Itk, and Tec SH3 domains (Fig. 8). When the
Btk-SH3 domain is autophosphorylated, its ability to take
part in protein interactions, such as with WASP, is altered as
compared to the non-phosphorylated fusion protein [52]. In
addition, the BCR-induced calcium mobilization and PLCg2
phosphorylation are abrogated partially by Btk mutation on
tyrosine 223 to phenylalanine (Y223F) [53]. Therefore, we
believe that SH3 domain phosphorylations regulate the
protein–ligand interactions for all Tec family members. In
accordance to the single SH3 domain phosphorylations, Btk-
SH3SH2 tandem domains are more strongly phosphorylated
than Itk-SH3SH2. This could mean that SH3SH2 tandem
domains have an additional interaction(s) with the substrate-
binding cleft in the kinase domain, or other parts of the
kinase, or, alternatively, that the SH3SH2 doublet alters
conformation so that the target tyrosine residue is more
exposed. Furthermore, we have not ruled out the possibility
of new phosphorylation sites in the doublet. However, the
isolated SH2 domain of Btk is not a substrate (unpublished
observations).
Taken together, our data show that transphosphorylation
of SH3 domains of the TFKs is a conserved mechanism,
which is not established in other SH3 domain-containing
proteins [54]. However, it has been demonstrated that two c-
Src SH3 domain mutants, Y133 and Y138, inhibit mito-
genic signaling, phosphorylation presumably being a pre-
requisite for the function [55,56]. On the other hand, these
two tyrosines in the Src SH3 domain are located at the SH3
domain C terminus, which is not homologous to the
phosphotyrosines reported in this work. Here, we show
the TFK transphosphorylation site to be specific and con-
served. Despite the fact that the role of SH3 domain
phosphorylations is still unclear, our data imply that TFK-
SH3 domains serve a conserved function. It has been shown
previously that the PXXP motif in the TH region of Itk may
associate in cis with its SH3 domain [25]. In the case of Btk,
such interactions can be observed in trans [26]. Further
examination of PP peptide interactions in two independent
studies [27,28] showed that Btk, in contrast to the Itk, forms
a dimer. In fact, this reflects the existence of double PP
stretches in Btk favoring asymmetric homodimer forma-
tions, whereas Itk containing a single PP motif allows
interactions in cis [27,28]. The identification of the pY
targets in the SH3 domain of the TFKs demonstrates another
level of regulation with both conserved and unique features.
Acknowledgements
We are indebted to Christer Wernstedt for radio-Edman
sequencing. The Swedish Cancer Foundation, Swedish
Medical Research Council, the European Union Grant
QLRT-2000-01395, the Paivikki and Sakari Sohlberg
Foundation, and Sigurd and Elsa Goljes Stiftelse supported
this work. Lars Ronnstrand holds a position as Senior
Researcher funded by the Swedish Research Council.
References
[1] D.R. Robinson, Y.M. Wu, S.F. Lin, Oncogene 19 (2000) 5548–5557.
[2] C.I.E. Smith, T.C. Islam, P.T. Mattsson, A.J. Mohamed, B.F. Nore, M.
Vihinen, BioEssays 23 (2001) 436–446.
[3] W.C. Yang, Y. Collette, J.A. Nunes, D. Olive, Immunity 12 (2000)
373–382.
[4] H. Mano, Cytokine Growth Factor Rev. 10 (1999) 267–280.
[5] C.M. Lewis, C. Broussard, M.J. Czar, P.L. Schwartzberg, Curr. Opin.
Immunol. 13 (2001) 317–325.
[6] J. Debnath, M. Chamorro, M.J. Czar, E.M. Schaeffer, M.J. Lenar-
do, H.E. Varmus, P.L. Schwartzberg, Mol. Cell. Biol. 19 (1999)
1498–1507.
[7] M. Vihinen, B.F. Nore, P.T. Mattsson, C.M. Backesjo, M. Nars, S.
Koutaniemi, C. Watanabe, T. Lester, A. Jones, H.D. Ochs, C.I.E.
Smith, FEBS Lett. 413 (1997) 205–210.
[8] D. Vetrie, I. Vorechovsky, P. Sideras, J. Holland, A. Davies, F. Flinter,
L. Hammarstrom, C. Kinnon, R. Levinsky, M. Bobrow, C.I.E. Smith,
D.R. Bentley, Nature 361 (1993) 226–233.
[9] S. Tsukada, D.C. Saffran, D.J. Rawlings, O. Parolini, R.C. Allen, I.
Klisak, R.S. Sparkes, H. Kubagawa, T. Mohandas, S. Quan, J.W.
B.F. Nore et al. / Biochimica et Biophysica Acta 1645 (2003) 123–132 131

Belmont, M.D. Cooper, M.E. Conley, O.N. Witte, Cell 72 (1993)
279–290.
[10] J.D. Thomas, P. Sideras, C.I.E. Smith, I. Vorechovsky, V. Chapman,
W.E. Paul, Science 261 (1993) 355–358.
[11] D.J. Rawlings, D.C. Saffran, S. Tsukada, D.A. Largaespada, J.C.
Grimaldi, L. Cohen, R.N. Mohr, J.F. Bazan, M. Howard, N.G. Cope-
land, N.A. Jenkins, O.N. Witte, Science 261 (1993) 358–361.
[12] P. Sideras, C.I.E. Smith, Adv. Immunol. 59 (1995) 135–223.
[13] K.Q. Liu, S.C. Bunnell, C.B. Gurniak, L.J. Berg, J. Exp. Med. 187
(1998) 1721–1727.
[14] X.C. Liao, D.R. Littman, Immunity 3 (1995) 757–769.
[15] C.L. Abram, S.A. Courtneidge, Exp. Cell Res. 254 (2000) 1–13.
[16] G. Superti-Furga, S.A. Courtneidge, BioEssays 17 (1995) 321–330.
[17] D.J. Rawlings, A.M. Scharenberg, H. Park, M.I. Wahl, S. Lin, R.M.
Kato, A.C. Fluckiger, O.N. Witte, J.P. Kinet, Science 271 (1996)
822–825.
[18] H. Mano, Y. Yamashita, A. Miyazato, Y. Miura, K. Ozawa, FASEB J.
10 (1996) 637–642.
[19] S.D. Heyeck, H.M. Wilcox, S.C. Bunnell, L.J. Berg, J. Biol. Chem.
272 (1997) 25401–25408.
[20] B.F. Nore, A.J. Mohamed, L. Vargas, L.J. Branden, C.M. Backesjo,
M. Vihinen, B. Christensson, C.I.E. Smith, Allergy Clin. Immunol.
Int. 12 (2000) 126–133.
[21] H. Park, M.I. Wahl, D.E. Afar, C.W. Turck, D.J. Rawlings, C.
Tam, A.M. Scharenberg, J.P. Kinet, O.N. Witte, Immunity 4
(1996) 515–525.
[22] H. Hansson, P.T. Mattsson, P. Allard, P. Haapaniemi, M. Vihinen,
C.I.E. Smith, T. Hard, Biochemistry 37 (1998) 2912–2924.
[23] A.T. Miller, L.J. Berg, Curr. Opin. Immunol. 14 (2002) 331–340.
[24] C.I.E. Smith, K.B. Islam, I. Vorechovsky, O. Olerup, E. Wallin, H.
Rabbani, B. Baskin, L. Hammarstrom, Immunol. Rev. 138 (1994)
159–183.
[25] A.H. Andreotti, S.C. Bunnell, S. Feng, L.J. Berg, S.L. Schreiber,
Nature 385 (1997) 93–97.
[26] H. Hansson, M.P. Okoh, C.I.E. Smith, M. Vihinen, T. Hard, FEBS
Lett. 489 (2001) 67–70.
[27] H. Hansson, C.I.E. Smith, T. Hard, FEBS Lett. 508 (2001) 11–15.
[28] A. Laederach, K.W. Cradic, K.N. Brazin, J. Zamoon, D.B. Fulton,
X.Y. Huang, A.H. Andreotti, Protein Sci. 11 (2002) 36–57.
[29] J. den Hertog, S. Tracy, T. Hunter, EMBO J. 13 (1994) 3020–3032.
[30] R.B. Birge, B.S. Knudsen, D. Besser, H. Hanafusa, Genes Cells 1
(1996) 595–613.
[31] J. Schlessinger, Curr. Opin. Genet. Dev. 4 (1994) 25–30.
[32] P. Wigge, H.T. McMahon, Trends Neurosci. 21 (1998) 339–344.
[33] S.N. Ho, H.D. Hunt, R.M. Horton, J.K. Pullen, L.R. Pease, Gene 77
(1989) 51–59.
[34] W.O. Bullock, J.M. Fernandez, J.M. Short, BioTechniques 5 (1987)
376–379.
[35] K. Hansen, M. Johnell, A. Siegbahn, C. Rorsman, U. Engstrom, C.
Wernstedt, C.H. Heldin, L. Ronnstrand, EMBO J. 15 (1996)
5299–5313.
[36] P. van der Geer, T. Hunter, Electrophoresis 15 (1994) 544–554.
[37] K.X. Luo, T.R. Hurley, B.M. Sefton, Methods Enzymol. 201 (1991)
149–152.
[38] P. Blume-Jensen, C. Wernstedt, C.H. Heldin, L. Ronnstrand, J. Biol.
Chem. 270 (1995) 14192–14200.
[39] J.P. Liu, A.T. Sim, P.J. Robinson, Science 265 (1994) 970–973.
[40] J.M. Sontag, E.M. Fykse, Y. Ushkaryov, J.P. Liu, P.J. Robinson, T.C.
Sudhof, J. Biol. Chem. 269 (1994) 4547–4554.
[41] H. Yu, J.K. Chen, S. Feng, D.C. Dalgarno, A.W. Brauer, S.L. Schreib-
er, Cell 76 (1994) 933–945.
[42] K. Alexandropoulos, G. Cheng, D. Baltimore, Proc. Natl. Acad. Sci.
U. S. A. 92 (1995) 3110–3114.
[43] C.H. Lee, K. Saksela, U.A. Mirza, B.T. Chait, J. Kuriyan, Cell 85
(1996) 931–942.
[44] H.V. Patel, S.R. Tzeng, C.Y. Liao, S.H. Chen, J.W. Cheng, Proteins
29 (1997) 545–552.
[45] L. Tamagnone, I. Lahtinen, T. Mustonen, K. Virtaneva, F. Francis, F.
Muscatelli, R. Alitalo, C.I.E. Smith, C. Larsson, K. Alitalo, Oncogene
9 (1994) 3683–3688.
[46] S.E. Pursglove, T.D. Mulhern, J.P. Mackay, M.G. Hinds, G.W. Book-
er, J. Biol. Chem. 277 (2002) 755–762.
[47] K.N. Brazin, D.B. Fulton, A.H. Andreotti, J. Mol. Biol. 302 (2000)
607–623.
[48] E.M. Schaeffer, J. Debnath, G. Yap, D. McVicar, X.C. Liao, D.R.
Littman, A. Sher, H.E. Varmus, M.J. Lenardo, P.L. Schwartzberg,
Science 284 (1999) 638–641.
[49] W. Ellmeier, S. Jung, M.J. Sunshine, F. Hatam, Y. Xu, D. Baltimore,
H. Mano, D.R. Littman, J. Exp. Med. 192 (2000) 1611–1624.
[50] M.G. Tomlinson, T. Kurosaki, A.E. Berson, G.H. Fujii, J.A. Johnston,
J.B. Bolen, J. Biol. Chem. 274 (1999) 13577–13585.
[51] C.D. Tsoukas, J.A. Grasis, K.A. Ching, Y. Kawakami, T. Kawakami,
Trends Immunol. 22 (2001) 17–20.
[52] L.M. Morrogh, S. Hinshelwood, P. Costello, G.O. Cory, C. Kinnon,
Eur. J. Immunol. 29 (1999) 2269–2279.
[53] T. Kurosaki, M. Kurosaki, J. Biol. Chem. 272 (1997) 15595–15598.
[54] M. Nishida, K. Nagata, Y. Hachimori, M. Horiuchi, K. Ogura, V. Man-
diyan, J. Schlessinger, F. Inagaki, EMBO J. 20 (2001) 2995–3007.
[55] M.A. Broome, T. Hunter, J. Biol. Chem. 271 (1996) 16798–16806.
[56] M.A. Broome, T. Hunter, Oncogene 14 (1997) 17–34.
[57] R. Koradi, M. Billeter, K. Wuthrich, J. Mol. Graph. 14 (1996) 51–55,
29–32.
B.F. Nore et al. / Biochimica et Biophysica Acta 1645 (2003) 123–132132