indicadores da magnitude da diversidade e … · em áreas inundáveis há plantas aquáticas e...
TRANSCRIPT

1
INDICADORES DA MAGNITUDE DA DIVERSIDADE E ABUNDÂNCIA DE
VERTEBRADOS SILVESTRES DO PANTANAL NUM MOSAICO DE
HÁBITATS SAZONAIS
CLEBER J. R. ALHO1, CHRISTINE STRÜSSMANN2 e LUÍS A. S.
VASCONCELLOS3
RESUMO: O Pantanal é uma planície de inundação (138.000 km2) alimentada pelos
tributários da margem esquerda do rio Paraguai e por ele mesmo, em sua grande maioria
no Brasil. Essa inundação varia de 11.000 a 110.000 km2 com média de 53.000 km2 .
Tal grau de inundação cria anualmente um gradiente de hábitats complexos em
mosaico. Os rios e outros cursos d'água são seguidos por matas ciliares ou de galeria e
há outros hábitats arbóreos nas áreas mais elevadas. O restante é formado por campos
sazonalmente inundáveis intercalados por capões de cerrado em diferentes fisionomias.
Em áreas inundáveis há plantas aquáticas e emergentes. A diversidade de vertebrados
ocorrendo no Pantanal é comparável à dos Cerrados, mas com pouco endemismo. A
produtividade anual da planície propicia a abundância sazonal de vertebrados. O ciclo
anual de seca e enchente favorece o aparecimento de nichos alimentares, reprodutivos e
quarteirão de passagem para vertebrados terrestres, aquáticos e semi-aquáticos. Os
processos ecológicos anuais, como a cheia, são vitais em diferenciar a fauna da planície
daquela dos planaltos vizinhos. O pulso de enchente dos rios, em conseqüência das
estações de chuva e seca, resulta em sazonalidade hidrológica, com produtividade de
ambientes reprodutivos e alimentares, produzindo heterogeneidade espacial pelos
diferentes graus de inundação da paisagem. Os Cerrados e o Pantanal, em geral,
compartilham da mesma fauna, sem haver diferenças interespecíficas relevantes quanto
ao grau de especiação ecológica, mas os recursos usados por espécies indicadoras do
Pantanal são local e sazonalmente mais abundantes, posto que os processos ecológicos
anuais ocasionam maior produtividade. A competição por alimento e espaço para
1 Departamento de Ecologia, Universidade de Brasília. Correio eletrônico: [email protected] Curso de Pós-Graduação em Biociências, Doutoranda em Zoologia, Pontificia Universidade Católica do
Rio Grande do Sul. Correio eletrônico [email protected] Correio eletrônico: [email protected]

2
reprodução proporciona a formação de distintas guildas de espécies, nas quais há fortes
interações interespecíficas, enquanto que as interações entre guildas diferentes são
fracas. Jacarés têm ampla distribuição local, capivaras e cervos são vistos forrageando
nos campos inundáveis na estação seca. Aves paludícolas são abundantes na seca,
formando colônias. Durante a estação de chuva populações espetaculares de rãs e sapos
são vistas e ouvidas. Essas espécies servem como indicadoras da riqueza e vitalidade do
Pantanal.

3
INDICATORS FOR THE MAGNITUDE OF DIVERSITY AND ABUNDANCE
OF THE VERTEBRATE WILDLIFE WITHIN A MOSAIC OF HABITATS IN
THE PANTANAL
ABSTRACT: The Pantanal is an enormous wetland (138,000 km2) that is fed by
tributaries of the upper Paraguay river in the center of South America, mainly in Brazil.
Inundation in the region varies from 11,000 to 110,000 km2 averaging 53,000 km2. The
degree of inundation creates a range of major habitats in a complex mosaic with annual
seasonallity. The rivers and streams are lined with gallery forests, and other arboreal
habitats appear on the more elevated areas. The remainder is either grasslands or
seasonally flooded grasslands interspersed with patches of different kinds of savanna
(cerrado). Floating, emergent and aquatic plants are abundant in inundable areas. The
vertebrate species diversity within the Pantanal is comparable to the diversity found in
the Cerrado biome, with low degree of endemism. The wetland's seasonal productivity
allows local abundance of diferent species of vertebrates. The annual cycle of dry and
wet seasons provides feeding and reproductive niches or stopping grounds for
terrestrial, aquatic and semi-aquatic species. Ecological processes, such as annual
inundation, are vital to differenciate the Pantanal's wildlife from the surrounding
savanna of the plateaus. The distinct annual pulsing of the rivers, as a result of the wet
and dry seasons, results in hydrological seasonality with productivity of feeding and
reproductive grounds, causing a pattern of spatial heterogeneity by different degrees of
inundation in the landscape. Both Cerrado and Pantanal share the same fauna, with no
relevant interspecific differences in the degree of ecological speciation, but the
resources used by some indicators species in the Pantanal are locally and seasonally
more abundant since the ecological annual process provides more productivity.
Competion for food and reproductive grounds suggests the formation of guilds of
different species, within which interspecific interactions are strong and interactions of
species belonging to different guilds are weak. Caymans are widespread and capybaras
and marsh deers can easily be seen foraging on grasslands during the dry season.
Waterfowl are abundant in the dry season, forming colonies. During the wet season

4
spectacular populations of frogs and toads are seen and heard. These species serve as
indicators of the richness and health of the Pantanal.

5
INTRODUÇÃO
Com o advento da Biologia da Conservação os pesquisadores em Ecologia
passaram a enfatizar o papel da diversidade e abundância de espécies da fauna silvestre
em seus hábitats naturais como a expressão de interações ecológicas, particularmente
competição por recursos alimentares, reprodutivos, de uso de espaço e outros. Os
processos ecológicos que anualmente - ou em períodos esporádicos maiores –
influenciam nos ciclos de vida da fauna silvestre, como os períodos sazonais de seca e
enchente do Pantanal, atuam nas escalas temporal e espacial, estabelecendo diferenças
locais ou regionais, quando comparadas, em larga escala biogeográfica, com os biomas
vizinhos, particularmente os Cerrados.
Há consenso em Ecologia de que a diversidade de espécies é fruto de uma
variedade de processos ecológicos e evolutivos, causas históricas e geográficas.
Contudo, o número de espécies coexistindo numa escala espacial ampla, como nos
Cerrados e Pantanal, varia significantemente em abundância local em função de
recursos sazonais regionais.
A oferta sazonal desses recursos leva a dois fenômenos: (a) associações e
agregações de espécies diferentes que interagem entre si, resultando numa
compartimentação em unidades ecológicas chamadas guildas, com a característica de
que há forte interação entre essas espécies diferentes dentro de cada guilda, mas com
interações fracas entre guildas diferentes; e (b) sincronização na escala de tempo de
agregação de espécies (abundância) com a oferta de recursos sazonais, notadamente
alimento e nichos reprodutivos.
Há indícios de que a cadeia trófica, nesse caso, faculta maior abundância de
espécies da fauna, e no caso de vertebrados, principalmente aves, anfíbios e alguns
répteis e mamíferos, onde os elos alimentares sazonais atuam mais no nível da
abundância local, sendo mais ou menos independentes da diversidade.
Desse modo, a distribuição de espécies, numa escala ampla, varia desde os
limites máximos por onde a espécie ocorre (distribuição geográfica) até uma área de
maior ocorrência da espécie (distribuição regional). Já o termo abundância é usado para

6
designar o tamanho (ou densidade) da população local. Na prática, o termo “local”
refere-se sempre à área amostral de um determinado estudo.
É bem conhecido que as espécies que são capazes de explorar um leque amplo
de recursos tornam-se, no decorrer de sua evolução, de distribuição geográfica ampla e
localmente abundantes. Algumas espécies de vertebrados do Pantanal parecem seguir
essa tendência: têm distribuição geográfica ampla, ocorrendo em toda a região de
cerrado e em outros biomas brasileiros, mas ocorrem em altas densidades em certos
períodos sazonais da região. Em outras palavras, parece não haver diferenças
interespecíficas de vertebrados entre o Pantanal e outros biomas brasileiros,
principalmente os Cerrados (não parece haver diferença no grau de especiação
ecológica), mas algumas populações que ocorrem no Pantanal usam recursos que são
mais abundantes ou mais produtivos.
O Pantanal como um sistema produtivo
Uma planície inundável, como o Pantanal, é universalmente definida como um
sistema ecologicamente produtivo. São hábitats em zonas de transição entre terras altas
não inundáveis (planaltos) e planícies inundáveis. A produtividade ecológica é
determinada pela: (a) dinâmica ou pulso hidrológico dos rios situados à margem
esquerda do rio Paraguai e por ele mesmo; (b) propriedades físicas, químicas,
microbiológicas do substrato; e (c) comunidade de plantas e animais especializados a
condições de solos saturados ou inundados. É um sistema que depende de inundação
mais ou menos rasa, recorrente, com saturação perto da superfície do substrato, na
época de cheia. Há, portanto, inundação recorrente ou saturação do substrato, com a
presença de características físicas, químicas e biológicas que refletem as condições de
inundação periódica, em solos hídricos e vegetação hidromórfica. Desse modo, três
elementos caracterizam o Pantanal como planície de inundação: água, substrato
(características fisico-químicas) e biota.
São três os critérios para definir a planície inundável: (a) o critério hidrológico
do pulso do rio Paraguai e seus afluentes da margem esquerda; (b) o critério do
substrato baseado nas condições física e química que refletem a natureza de inundação

7
periódica e (c) a presença de organismos, incluindo espécies de vertebrados, como aves
paludícolas, anfíbios, répteis e mamíferos, adaptados ao regime recorrente de
inundação. Assim, água e a inundação recorrente são elementos importantes. O regime
hidrológico é fundamental para o funcionamento do sistema, mesmo que os elementos
físico-químicos do substrato ou os biológicos persistam por algum tempo, em caso de
perturbação.
Baixo nível de oxigênio na água e nos solos saturados é uma das características
das planícies inundáveis. No solo saturado isto se deve à presença de microorganismos,
raízes e outros organismos. No Pantanal , a água do rio Paraguai que drena do planalto
e encontra na planície um impedimento para seu fluxo apresenta redução de oxigênio
dissolvido (02), sobressaturação de dióxido de carbono (C02) e metano (CH4), além da
perda de sedimentos em suspensão e redução de nitrogênio e fósforo (Hamilton et al.,
1995; Hamilton et al., 1997). Essas águas que inundam a planície, quando encontram
ambientes com plantas vasculares emergentes aquáticas, tornam-se pobres em oxigênio
dissolvido com excesso de produção de CO2.
Enchente e vazante: ritmos sazonais
A planície é formada pelo rio Paraguai e seus tributários da margem esquerda,
grandemente no Brasil (Bento Gomes, Cuiabá, São Lourenço, Itiquira, Taquari, Negro,
Aquidauana-Miranda, Nabileque e Apa). As margens do rio Paraguai constituem o
limite oeste do Pantanal, tocando a Bolívia ao norte e, o Paraguai, ao sul. O rio Paraguai
e seus tributários têm drenagem lenta quando descem do planalto e encontram a
planície. Em anos chuvosos, como em 1984 e 1995, o rio Paraguai transborda,
expandindo sua extensão na planície em até 20 km.
A altitude da planície varia de 60 a 150 m e o gradiente topográfico é fraco, com
desnível variando entre 30 a 50 cm por quilômetro no sentido leste-oeste, e 3 a 15 cm
por quilômetro no sentido norte-sul (Franco e Pinheiro, 1982). Esse pequeno desnível na
planície constitui impedimento para o escoamento da água que chega no leito dos rios.
As terras acima de 200 m são consideradas planalto, podendo atingir 800 e 1.200 m de
altura, como a Chapada dos Guimarães.

8
O Pantanal é formado por uma enorme planície aluvial, drenada por uma
complexa cadeia de rios e córregos entremeados por cerrado, matas e campos
inundáveis. O período de chuva (com variações entre a região norte e sul da região)
ocorre entre outubro-dezembro e março-maio, variando entre 1.200 a 1.300 mm por
ano. Inundação máxima foi detectada o mais cedo em fevereiro, no norte, e a mais tardia
em junho, no sul (Hamilton et al., 1996). As chuvas que ocorrem no planalto ocasionam
rápida resposta na planície, inundando a região. O Pantanal está situado a 1.500 km da
costa atlântica e localiza-se em área de clima seco, com classificação Aw (clima de
savana), conforme Köppen (Brasil, 1997). Segundo ainda Brasil, enquanto a
precipitação na planície é de 800 a 1.200 mm, o potencial de evaporação é de 1.300 a
1.600 mm, com balanço hídrico negativo, com retenção de 30 a 60% de água na planície
inundável.
Há períodos longos de seca e inundação registrados na literatura (Cadavid
Garcia e Castro, 1986). Dados históricos do nível do rio Paraguai em Ladário, perto de
Corumbá, MS, mostram que os níveis máximos de enchente alcançaram 4 metros de
1900 a 1960, mas apenas 2 m de 1960 a 1972. Portanto, de 1960 a 1972, na época de
seca, muitas terras ficaram sem inundação, voltando a serem inundadas outra vez a
partir de 1973, quando o nível do rio atingiu 5 m na enchente (Sá et al., 1998). Outros
dados diários indicaram variações anuais na enchente e variações em intervalos de dois
a cinco anos. Na seca a água que não escoa para os leitos dos rios pelos canais de
drenagens (vazantes e corixos) fica retida nas baías e campos de inundação, evaporando
e infiltrando-se no solo.
Produtividade de peixes e outros recursos
Há 263 espécies de peixes catalogados para o Pantanal (Britski et al., 1999).
Peixes são recursos ecológicos importantes para essa região, como compartimento
biótico do sistema. Constituem alimento sazonal em torno do qual há agregação de
espécies de vertebrados: aves paludícolas, que se agregam em vazantes ou baías, para se
alimentarem, ou em ninhais, colônias de reprodução em comportamento cooperativo
para onde os pais trazem o alimento, geralmente peixes, e onde se congregam grandes

9
quantidades de predadores que se aproveitam da oferta de alimento (filhotes de aves que
caem dos ninhos), como sucuris, jacarés, lobinhos, mão-pelada, coatis e outros
oportunistas.
A produtividade anual de alimentos dos vertebrados está ligada ao tamanho do
substrato, ao regime de descarga dos rios, ao hábito de predação, a diferenças anuais de
temperatura, ao fluxo intermitente de enchente e ao tipo de bioma, processos que
influenciam a produtividade local (Vinson e Hawkins, 1998; Wantzen e Junk, 2000). O
ambiente muda diante de cada descarga hídrica com sedimentos e nutrientes
dissolvidos, criando novos microhábitats, com depósitos de substâncias inorgânicas e
orgânicas que o enriquecem e favorecem a proliferação de invertebrados e a migração
de peixes. Ao mesmo tempo, plantas bem adaptadas a ocuparem seus espaços na época
da seca ou as plantas aquáticas rapidamente se proliferam. As árvores interagem com o
regime hídrico por meio de suas raízes. A matéria orgânica morta ou viva vai-se
alternando e provendo microhábitats para os invertebrados. Por exemplo, as áreas com
mais sombra no interior das matas ciliares produzem menos algas e biomassa de
macrófitas que áreas mais ensolaradas, mas recebem muito mais detritos de folhas,
galhos e troncos mortos, os quais desempenham papel crucial no balanço energético do
ecossistema (Wantzen e Junk, 2000). Citando outros autores, Wantzen e Junk (2000)
afirmam que substratos originados de árvores são fontes de diversidade e abundância
para invertebrados.
O ciclo hidrológico do Pantanal está intimamente relacionado com o ciclo de
vida dos peixes e à conseqüente oferta sazonal de alimento. Três fases distintas são
identificadas com respeito ao ciclo de vida dos peixes em relação ao regime de águas do
Pantanal:
a) Estação da cheia, período de chuva contínua com inundação da planície,
geralmente de outubro a abril. O movimento dos peixes começa com a piracema,
quando os cardumes sobem os rios no início da estação da enchente. Mais tarde, os
peixes migram dos leitos dos rios e se deslocam para as áreas inundadas adjacentes,
em busca de alimento nos campos inundados e baías. A reprodução ocorre no início
da estação de cheia, com a rodada, movimentação de peixes desde o leito dos rios
para as áreas inundadas. Desse modo, a rodada ocorre logo após a piracema.

10
b) Vazante, período coincidindo com o fim da estação cheia, geralmente de abril a
maio. Os peixes dispersam-se das áreas alagadas e voltam aos leitos dos rios por
meio de canais de drenagens (vazantes e corixos). Essa migração é conhecida como
lufada. Nesse período, os peixes são capturados por predadores naturais e aqueles
que ficam aprisionados em baías e campos de inundação, que estão em processo de
seca, servem de alimento para predadores que aí se agregam.
c) Estação de seca, geralmente de junho a outubro. As espécies de peixes sedentárias,
isto é, aquelas que não migram, enfrentam baixos níveis de oxigênio nas águas rasas
e mornas, e algumas espécies permanecem dormentes na lama durante toda a
estação seca.
O ciclo de vida dos peixes é determinante na oferta de alimento para muitas
espécies de vertebrados do Pantanal. Sincronicamente, o ciclo hidrológico faculta picos
de abundância de invertebrados, como caramujos e moluscos, que também têm
relevância na cadeia trófica.
Mosaico de hábitats
O regime de seca-cheia faculta eventos de grande produção primária quando, na
seca, os campos inundáveis são rapidamente tomados por plantas oportunísticas de
estratégia “r”, cuja biomassa serve de alimento para herbívoros. Igualmente, o ritmo
fenológico faculta a oferta de brotos novos, sementes, flores e frutos, alimento para os
vertebrados arborícolas.
Segundo Hamilton et al. (1996), o ciclo sazonal de inundação não é uniforme em
todo o Pantanal, sendo mais regular no norte do que no sul da região. Estes autores
detectaram, ainda, diferenças em grau de inundação nas diferentes sub-regiões do
Pantanal. Em ano muito seco analisado, como em 1986, 75% da área normalmente
inundável permaneceu seca.
O Pantanal é subdivido em sub-regiões, que recebem nomes locais com base no
regime de inundação e tipo de cobertura vegetal (Silva e Abdon, 1998). As diversas
unidades de paisagens presentes na região e adotadas neste trabalho estão descritas na
literatura, em Hernandez Filho, Ponzoni e Pereira, 1998, em Brasil (1997) e em Silva et

11
al. (1998), para fitofisionomias arbustivas, arbóreas e florestadas, e em Abdon et al.
(1998) para as plantas aquáticas.
A planície inundável do Pantanal é formada por depósitos aluviais acumulados
desde o Pleistoceno (Brasil, 1997). Nos dias de hoje esses depósitos estão cobertos por
florestas (matas de galeria e outras formas florestadas), cerrado e cerradão e várias
formas de campos, incluindo campos inundáveis. Fatores antropogênicos, como a
conversão de cerrado do planalto onde nasce o rio Taquari, em campos de agricultura
(principalmente cultivo de soja), nos últimos 20 anos, têm ocasionado assoreamento do
rio, com descargas excessivas de sedimentos trazidos pelo rio do planalto para a planície
(Alho et al., 1988b; Silva et al., 1998).
O Pantanal é caracterizado por ter limite não distinguível entre área inundável e
terra livre de inundação. Todo ano, contudo, grande parte de ambientes terrestres
transformam-se em ambientes aquáticos. Estando inserido no bioma Cerrados, o
Pantanal localiza-se num ambiente seco, distanciando-se 1.500 km da costa atlântica.
A paisagem natural de hoje é resultado de: (a) processos evolutivos ocorridos
desde o Pleistoceno; (b) sazonalidade bem pronunciada, com ciclos anuais de cheia e
seca e períodos excepcionais de cheias ou expansão do Pantanal (com evidentes reflexos
sobre a produtividade primária e os processos de sucessão ecológica), e (c) áreas
relacionadas com a intervenção humana, principalmente pela expansão de áreas de
pastagem, desmatamentos, implantação de estradas e outras atividades.
Indicadores. Cada um dos três grandes compartimentos que compõem o
Pantanal – hidrologia, substrato e biota – pode ser interpretado e desdobrado em
indicadores dos processos ecológicos e ciclos do ecossistema. Para efeito deste trabalho,
o enfoque será dado no terceiro elemento citado – biota – particularmente anfíbios,
répteis, aves e mamíferos e seus hábitats associados. Os peixes são vertebrados que têm
distribuição muito particular e não são aqui abordados.
O monitoramento de fauna por meio de indicadores designados é um processo
de avaliação contínua, de observação analítica e documentação de ações ou atividades,
produtos, resultados, impactos, tendências para investigar as razões de expectativas que
podem ser presumidas ou de hipóteses que podem ser formuladas. Os indicadores são

12
sinais ou evidências que tornam possível para o pesquisador verificar qual a extensão do
fenômeno observado e se o mesmo está sofrendo variações por causa dos processos ou
intervenções específicas. Por exemplo, embora a capivara tenha uma ampla distribuição
geográfica, quais os indicadores do hábitat que podem ser tomados para analisar a
abundância dessa espécie no Pantanal? Desse modo, vários tipos de indicadores podem
ser estabelecidos: indicadores de contexto – quando se deseja monitorar fatos ou
situações relacionados com uma região ampla; indicadores de processos – quando o
que se torna importante é a observação da seqüência de ações, ou interações de
comportamento numa escala de tempo; indicadores de impacto – quando se quer
verificar os efeitos relativos a objetivos gerais, como o número de espécies numa área
de pastagem implantada.
O uso de espécies indicadoras para avaliar e monitorar processos biológicos nos
ecossistemas data do início do século. Esse conceito, posteriormente, teve amplo
desenvolvimento, para uso no controle da poluição em rios e lagos. O conceito de
espécies indicadoras baseia-se no uso daquelas que estão presentes na comunidade e
sujeitas a algum tipo de impacto evolutivo no passado, ou antropogênico, no presente. O
valor de indicação da espécie pode ser pelo fato de serem intolerantes às condições
modificadas ou degradadas (sendo, portanto, as primeiras a desaparecerem após o
impacto antropogênico), ou porque são tolerantes às condições de impacto.
Conhecendo a natureza da interação entre a presença e/ou abundância da espécie
em relação ao tipo de impacto, sua resposta ao mesmo pode ser usada como indicadora.
Os impactos causados induzem a alterações na natureza das relações entre as espécies
na comunidade. Pode-se esperar, em função das deduções citadas, que grupos ou
conjunto de espécies indicadoras podem definir o “status” do sistema em termos do
impacto corrente. Nesse contexto, é extremamente importante ter conhecimento prévio
dos efeitos dos fatores impactantes, da espécie, população e comunidade.
Nesse sentido, quais os critérios que são usados para escolher objetivamente as
espécies indicadoras?
(a) o primeiro critério é que um conjunto de espécies relacionadas seja
preferível;

13
(b) as espécies devem ser estenotópicas ou sensíveis, com pequena variabilidade
na resposta ao fator ou fatores impactantes, de modo que uma mudança na
abundância relativa da espécie indica as condições do hábitat;
(c) sejam residentes ao longo ou em parte do ano;
(d) sejam fáceis de monitorar;
(e) que apresentem curto tempo de geração de modo que a população mude
rapidamente quando exposta aos fatores ambientais;
(f) sejam abundantes para que variações no tamanho populacional sejam de
magnitude suficiente para responder aos impactos ambientais.
O primeiro critério nos remete aos conceitos de espécie-chave e guilda. No caso
de espécie-chave, está se referindo à espécie de cuja presença uma série de outras delas
dependem, direta ou indiretamente. A espécie-chave pode não ser a mais abundante,
mas seu efeito é maior do que se espera. É o caso de espécies-topo da cadeia trófica,
como predadores importantes.
O conceito de guilda está ligado à idéia de espécies indicadoras, exceto pelo fato
de que as indicadoras foram transformadas em guildas indicadoras. Estas respondem
similarmente aos fatores impactantes.
A ferramenta de bioindicadores ou de indicadores ambientais tem sido
amplamente utilizada, inclusive pelo Banco Mundial para avaliar seus projetos, através
dos indicadores de performance (Segnestam, 1999).
Objetivos deste trabalho. Este trabalho tem por objetivo discutir a distribuição
e abundância de anfíbios, répteis, aves e mamíferos no Pantanal comparando-as com
aquelas em biomas vizinhos, particularmente os Cerrados. As seguintes questões são
postuladas:
As comunidades de vertebrados vivendo nos diferentes hábitats do Pantanal são
similares em riqueza e diversidade aos hábitats de outros biomas brasileiros,
particularmente dos Cerrados da região do planalto da Chapada dos Guimarães?
- Se são diferentes, é possível analisar essas diferenças em termos de
processos ecológicos e ciclos anuais ?

14
- Essas diferenças podem ainda ser interpretadas em termos de nichos
alimentares, reprodutivos, espaciais e outros que favoreçam abundâncias regionais?
- Espécies generalistas, com distribuição ampla, tendem a ser localmente
mais abundantes que as especialistas?
- A abundância regional pode ser resumida em simples agregação em torno
de recursos sazonais ?
- Que medidas devem ser tomadas para incentivar formas de
desenvolvimento sustentável, como o ecoturismo, e como os processos ecológicos,
as paisagens naturais e as espécies devem ser protegidas?
MATERIAL E MÉTODOS
Este trabalho é baseado na experiência pessoal de cada um dos autores no
Pantanal e em áreas de Cerrados no Brasil Central (não estando limitado,
necessariamente, a um dado período de tempo ou local) e em informações disponíveis
na literatura científica.
ANFÍBIOS E RÉPTEIS
Dentre os vertebrados ocorrentes na Bacia do Alto Paraguai (BAP), anfíbios e
répteis estão entre os grupos menos conhecidos, tanto do ponto de vista da composição
de distintas faunas locais como da história natural das espécies e de seu grau de
associação às unidades de paisagem. Uma listagem compreensiva dos taxa da
herpetofauna presentes na bacia foi disponibilizada, pela primeira vez na literatura, no
documento final referente ao componente biótico do Plano de Conservação da Bacia do
Alto Paraguai – PCBAP (Brasil, 1997). Um total de 22 espécies de anfíbios anuros e 83
de répteis foram listadas para a planície de inundação, em documento com reduzida
circulação e de difícil acesso para a comunidade científica em geral.
No que se refere aos anuros, a abrangência da lista apresentada em Brasil (1997)
está limitada a áreas da porção sul da planície, em particular os arredores da área urbana

15
de Corumbá e a região central do Pantanal da Nhecolândia (Brasil, 1997). A lista inclui,
também, dezoito espécies de anuros de áreas elevadas das bordas leste e oeste do
Pantanal de Mato Grosso do Sul, incluindo as serras do Urucum, Bodoquena e
Maracaju. Já a lista de répteis da BAP inclui, além das espécies presentes na planície,
outras 84 formas com ocorrência limitada a porções mais elevadas, no entorno de
praticamente toda a Bacia (Brasil, 1997).
Peixes, aves e mamíferos tiveram as listagens de espécies do Pantanal revistas
e/ou ampliadas a partir daquelas apresentadas por Brasil com a conseqüente
disponibilização de listagens mais recentes (por exemplo, peixes – Britsky et al., 1999;
aves – Tubelis e Tomás, no prelo; mamíferos – Mauro, apud Adámoli e Pott, 1999).
Para anfíbios e répteis, não houve atualização das listagens constantes em Brasil (1997).
O exame de material testemunho depositado em grandes coleções científicas,
como as do Museu de Zoologia da Universidade de São Paulo (MZUSP) e do Museu
Nacional do Rio de Janeiro (MNRJ), poderá revelar importantes registros adicionais
para a área da BAP. Dada a escassez de inventários em localidades na Bacia,
amostragens pontuais, sejam elas avaliações ecológicas rápidas (por exemplo,
Strussmann et al., 2000), sejam diagnósticos faunísticos mais completos (como aqueles
realizados na área de influência do Aproveitamento Múltiplo de Manso, no município
de Chapada dos Guimarães, Mato Grosso; Strussmann, 2000), podem também aumentar
significativamente o conhecimento sobre a herpetofauna da região. Em conjunto, por
exemplo, os estudos de diagnóstico e de resgate faunístico em Manso evidenciaram a
presença de doze espécies de répteis anteriormente não apontadas para a BAP.
Neste trabalho, é feita uma análise comparativa de alguns indicadores da
magnitude da diversidade e da abundância de anfíbios e répteis em áreas da planície
inundável e de seu entorno, particularmente os cerrados da região de Chapada dos
Guimarães. Tendo por base as informações apresentadas em Brasil (1997) e
observações de campo adicionais, são brevemente discutidos aspectos relativos à
riqueza de espécies, padrões de distribuição e de associação a distintas unidades de
paisagem, abundância relativa, vigor e sazonalidade das populações desses organismos.

16
Riqueza de espécies
Exames adicionais da literatura e de espécimes depositados em coleções e,
principalmente, inventários faunísticos recentes, como aqueles realizados na região do
Aproveitamento Múltiplo de Manso (APM Manso), na borda nordeste da BAP,
revelaram como presentes em áreas de planalto da Bacia, além das formas enumeradas
em Brasil (1997), o quelônio Geochelone denticulata, os lagartos Anolis cf.
fuscoauratus, Micrablepharus atticolus, Tupinambis quadrilineatus e uma nova espécie
de Cnemidophorus, as cobras-cegas Amphisbaena fuliginosa e Cercolophia roberti, as
serpentes Dipsas indica, Drymoluber brazili, Liophis longiventris, L. paucidens e uma
subespécie adicional de Bothrops neuwiedi.
Na planície inundável, além das formas listadas por Brasil (1997), há registro
positivo apenas para uma espécie adicional de lagarto, do gênero Anolis (próxima a e
em sintopia com A. meridionalis), no Pantanal de Barão de Melgaço, e de uma serpente
(cuja identidade taxonômica necessita ser melhor estudada, podendo constituir espécie
ainda não descrita de Hydrops4), na região de Corumbá, MS.
Há, assim, 85 espécies de répteis ocorrentes na planície inundável (sem serem
exclusivas da região) e outras 94 somente registradas, até o momento, em áreas do
entorno, num total de 179 espécies de répteis para a BAP. Quanto aos anfíbios, além das
formas registradas em Brasil (1997), são conhecidas pelo menos treze espécies
adicionais na planície e outras 27 aparentemente exclusivas dos planaltos de entorno. A
anurofauna da BAP estaria constituída, por pelo menos 80 espécies, das quais mais de
metade (45 espécies) parece ocorrer exclusivamente em áreas altas no entorno (FIG. 1).
As listas de herpetofauna para a Bacia, no entanto, são ainda incompletas. Quelônios,
por exemplo, constituem um grupo muito mal amostrado na área, havendo,
seguramente, espécies adicionais àquelas citadas em Brasil (1997).
4 V.L. Ferreira, comunicação pessoal.
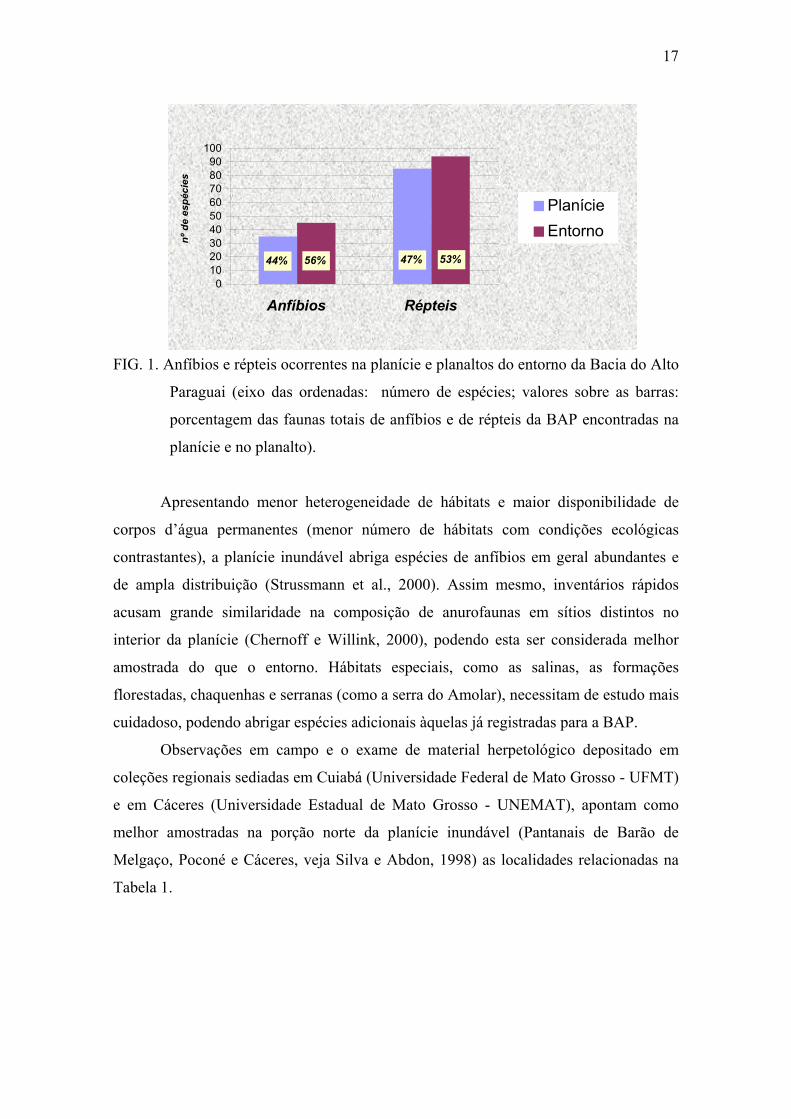
17
FIG. 1. Anfíbios e répteis ocorrentes na planície e planaltos do entorno da Bacia do Alto
Paraguai (eixo das ordenadas: número de espécies; valores sobre as barras:
porcentagem das faunas totais de anfíbios e de répteis da BAP encontradas na
planície e no planalto).
Apresentando menor heterogeneidade de hábitats e maior disponibilidade de
corpos d’água permanentes (menor número de hábitats com condições ecológicas
contrastantes), a planície inundável abriga espécies de anfíbios em geral abundantes e
de ampla distribuição (Strussmann et al., 2000). Assim mesmo, inventários rápidos
acusam grande similaridade na composição de anurofaunas em sítios distintos no
interior da planície (Chernoff e Willink, 2000), podendo esta ser considerada melhor
amostrada do que o entorno. Hábitats especiais, como as salinas, as formações
florestadas, chaquenhas e serranas (como a serra do Amolar), necessitam de estudo mais
cuidadoso, podendo abrigar espécies adicionais àquelas já registradas para a BAP.
Observações em campo e o exame de material herpetológico depositado em
coleções regionais sediadas em Cuiabá (Universidade Federal de Mato Grosso - UFMT)
e em Cáceres (Universidade Estadual de Mato Grosso - UNEMAT), apontam como
melhor amostradas na porção norte da planície inundável (Pantanais de Barão de
Melgaço, Poconé e Cáceres, veja Silva e Abdon, 1998) as localidades relacionadas na
Tabela 1.
47%44% 53%56%
0102030405060708090
100
Anfíbios Répteis
nº d
e es
péci
es
PlanícieEntorno

18
TABELA 1. Localidades na porção norte do Pantanal (Mato Grosso) melhor amostradas
quanto à composição da herpetofauna, com base em observações de campo
e em material depositado até 1995 em duas coleções regionais (v. texto).Coordenadas
Localidade Latitude(S)
Longitude(O)
Répteis
(nº spp.)
Anfíbios
(nº spp.)
1. Fazenda. Santa Inês (Pantanal de Poconé) 16º 30’ 56º 45’ 37 262. Reserva Particular do Patrimônio Natural (RPPN)SESC Pantanal (Pantanal de Barão de Melgaço)
16º 46’ 56º 10’ 26 23
3. Parque Nacional do Pantanal 17º 52’ 57º 27’ 20 234. Base de Pesquisas de Fauna do IBAMA, RodoviaTranspantaneira
17º 10’ 57º 00’ 17 23
O número de anuros presentes nas localidades consideradas como relativamente
bem amostradas, entre 23 e 26 espécies, aproxima-se do valor estimado para o número
total de espécies potencialmente ocorrentes num mesmo sítio, algo em torno de 30
espécies. Já as áreas de entorno da BAP, especialmente as altas cabeceiras, são
virtualmente desconhecidas do ponto de vista de suas anurofaunas, os poucos
inventários existentes indicando haver, aí, maior número de espécies num dado sítio e
maior “turnover” de espécies entre sítios.
Possivelmente, muitas das formas de regiões de cabeceiras (assim como plantas
e peixes desses sítios) apresentam distribuição restrita, com diferentes sítios
configurando-se como ilhas com microbiotas distintas (Chernoff e Willink, 2000). Em
relatório não publicado, A.F.B. de Araújo e G. Colli (Universidade de Brasília) estimam
a ocorrência de 45 espécies de anuros em uma localidade qualquer em cerrado, em um
raio de 20 km. Em Manso, cuja fauna de anuros é, atualmente, a mais bem conhecida da
Bacia do Alto Paraguai, foram registradas 47 espécies, em área de tamanho equivalente.
Répteis são mais difíceis de amostrar em curto prazo, especialmente serpentes, e
ainda não há informações suficientes sobre distintas faunas locais para estimar o número
de espécies co-ocorrendo em uma determinada região na planície. Na Tabela 1, por
exemplo, o número de espécies conhecidas para a região da Fazenda Santa Inês, no
Pantanal de Poconé (30 % mais espécies de répteis do que na segunda localidade melhor
amostrada), incorpora dados sobre a ofiofauna local obtidos durante dois anos de estudo
de campo intensivo. Em conseqüência, o número de espécies de serpentes registradas
para a região da fazenda Santa Inês é superior (60%) ao número de espécies conhecidas

19
na segunda localidade melhor amostrada. Nesta, apenas um inventário rápido foi
realizado, no começo do período chuvoso. Para anfíbios do Pantanal, este é o período
em que a maior parte das espécies iniciam suas atividades reprodutivas, o que facilita o
reconhecimento das espécies pelo registro de suas vocalizações e pela formação de
agregações reprodutivas.
Padrões de distribuição
Formado durante o Pleistoceno por reativação tectônica, que rebaixou a porção
central de uma planície de erosão pré-existente, o Pantanal desenvolveu-se, nos últimos
30 mil anos, com a contribuição de sedimentos das chapadas circundantes (Ab’Sáber,
1988). Intensos e generalizados processos erosivos em porções superficiais dos
planaltos circundantes e um progressivo e continuado acúmulo de detritos no interior da
planície oportunizaram “poucas possibilidades para o desenvolvimento de ecossistemas
e homogeneização de revestimentos florísticos” (Ab’Sáber, 1988, p.41).
A ocupação biótica do Pantanal é, assim, relativamente recente, tendo ocorrido
após uma fase seca, ao final do Pleistoceno, em que houve grande expansão das
caatingas em território brasileiro, com “importante penetração de climas e floras semi-
áridas, no interior e bordos da depressão pantaneira” (Ab’Sáber, 1988, p. 41, 44). A
recomposição da tropicalidade na depressão, com umidificação do clima e invasão por
floras e faunas oriundas de refúgios situados em diferentes sítios das terras
circunvizinhas, permitiu então a expansão dos cerrados, predominantemente pelo
grande leque do Taquari, a penetração de componentes florísticos periamazônicos, pelo
norte, e do Chaco Oriental, pelo extremo sudoeste e sul (Ab’Sáber, 1988). Adámoli
(1982) e Adámoli e Pott (1999) reconhecem a presença adicional de influências da
floresta atlântica, na borda sudeste da planície.
Ao constituir uma mescla relativamente recente de elementos oriundos de
províncias fitogeográficas vizinhas, os quais foram capazes de se adaptar a um novo
cenário ambiental regido por inundações periódicas, a herpetofauna da planície
pantaneira apresenta baixo nível de endemismos locais, em contraste com os planaltos
adjacentes. Mesmo espécies com hábitos aquáticos e imagem indiscutivelmente
associada à região pantaneira, como a víbora-do-pantanal (ou jacuruxi, Dracaena

20
paraguayensis) e a sucuri-amarela (ou sucuri-do-Pantanal, Eunectes notaeus), atingem
outras áreas de inundação, contíguas mas definitivamente situadas fora dos limites da
BAP, ou mais para o norte, na região do vale do Guaporé, ou ao sul, em região do
território argentino sob a influência do baixo curso do rio Paraguai.
O número de províncias fitogeográficas incluídas na área de distribuição de uma
espécie qualquer tem sido empregada, algumas vezes, como medida indireta de sua
versatilidade ecológica (veja Henderson et al., 1995). Uma análise neste sentido revela
que a maioria das espécies de répteis registradas para a porção norte do Pantanal não é
exclusiva da região (FIG. 2a).
FIG. 2a. Ocorrência, em outros biomas sul-americanos, de espécies da fauna de répteis
do Pantanal.
a
19 %(12 spp.)
11 % (7 spp.)
9 % (6 spp.)
8 % (5 spp.)
24 % (15 spp.)
29 % (18 spp.) Distribuição continental
Diagonal de formações abertas
Pantanais e Chaco inundável
Cerrado
Áreas abertas e Floresta Atlântica
Áreas abertas e Floresta Amazônica

21
FIG. 2b. Ocorrência, em outros biomas sul-americanos, de espécies da fauna de répteis
de uma localidade de planalto, no entorno da Bacia do Alto Paraguai
(Aproveitamento Múltiplo de Manso, Chapada dos Guimarães, MT).
Em conjunto, espécies com ocorrência somente ao longo da “diagonal de
formações abertas sul-americana” (caatinga, cerrado, pantanal, chaco) perfazem mais de
50 % da fauna de répteis da porção norte do Pantanal. Aí se incluem tanto espécies
amplamente distribuídas ao longo de toda a diagonal como formas com ocorrência ora
apenas em sua porção sul, do cerrado ao chaco (como as serpentes Hydrodynastes gigas
e Phalotris tricolor), ora restritas à porção norte, da caatinga ao pantanal (o lagarto
Micrablepharus maximiliani, o anfisbênio Amphisbaena vermicularis e a serpente
Philodryas nattereri, por exemplo). Há, também, formas restritas ao cerrado (por
exemplo, o lagarto Coleodactylus brachystoma e a serpente Xenopholis undulatus) e às
áreas inundáveis sob influência do sistema Paraguai-Paraná (chegando, em alguns
casos, a áreas de cerrado periféricas ao Pantanal) e/ou pantanais do alto Guaporé (como,
o quelônio Acanthochelys macrocephala, o jacaré Caiman yacare, as serpentes
Eunectes notaeus, Micrurus tricolor, Clelia bicolor, os lagartos Dracaena
paraguayensis e Mabuya guaporicola).
Pelo menos quinze espécies registradas para a porção norte do Pantanal (c. 25 %
do total) têm distribuição praticamente continental, ocorrendo em cinco ou mais
províncias distintas, que incluem tanto outras formações abertas como áreas dos
b27 %
(27 spp.)
16 % (16 spp.)
17 % (17 spp.)
14 % (14 spp.)
26 %(26 spp.)
Distribuição continental
Diagonal de formações abertas
Endêmicas do Cerrado
Áreas abertas e Floresta Atlântica
Áreas abertas e Floresta Amazônica

22
domínios florestais amazônico e/ou atlântico (como o quelônio Geochelone carbonaria,
os lagartos Iguana iguana, Hemidactylus mabouia, o anfisbênio A. alba e as serpentes
Boa constrictor, Liophis almadensis, Mastigodryas bifossatus, Spilotes pullatus).
Das espécies presentes em ambientes abertos e em um dos dois grandes biomas
florestados, cinco podem ser consideradas como “amazônicas” (o quelônio G.
denticulata, os lagartos Tupinambis teguixin, Mabuya nigropunctata e as serpentes
Oxyrhopus aff. petola e Corallus hortulanus); outras oito espécies aparecem em biomas
abertos e em áreas da floresta atlântica (como os lagartos Tropidurus torquatus, Mabuya
frenata, ou a serpente Philodryas patagoniensis) (FIG. 2a).
Uma análise comparativa da distribuição, nos grandes biomas, das espécies de
répteis presentes na área da usina hidrelétrica de Manso, no entorno da porção nordeste
da planície pantaneira, revela padrões de ocorrência distintos daqueles observados na
planície (FIG. 2b). Embora espécies com ampla distribuição representem proporções
semelhantes em faunas de répteis do Pantanal e de seu entorno, a contribuição de
formas exclusivas de biomas abertos e daquelas que podem ser encontradas tanto nestes
como em biomas florestados é distinta nas duas porções da Bacia. Na planície, formas
exclusivas de biomas abertos representam c. 54 % da fauna total de répteis, enquanto no
planalto esta contribuição equivale a apenas 42 %. Já o número de formas
“amazônicas”, por exemplo, é duas vezes maior na fauna de répteis de localidade no
planalto do que entre os répteis registrados para o Pantanal.
O caráter “amazônico” de parte da herpetofauna de Manso é mais evidente entre
espécies e subespécies de hábitos aquáticos ou semi-aquáticos, entre as quais se registra
a presença de diversas formas vicariantes em relação àquelas presentes no Pantanal
(Tabela 2). O papel dos sistemas hidrográficos parece ter sido fundamental na
distribuição dessas espécies, tanto no passado como nos tempos atuais.

23
TABELA 2. Exemplos de espécies e subespécies vicariantes nas herpetofaunas do
Pantanal e dos cerrados no entorno, na porção norte da planície. Espécies
de hábitos aquáticos ou semi-aquáticos estão assinaladas com (A).Pantanal Manso
Hydrodynastes gigas (A) Hydrodynastes bicinctus (A)Eunectes notaeus (A) Eunectes murinus (A)
Helicops angulatus (A)Helicops leopardinus (A)H. polylepis (A)
Caiman yacare (A) Caiman crocodilus (A)Bufo granulosus ssp. 1 Bufo granulosus ssp. 2Bufo aff. margaritifer sp. 1 Bufo aff. margaritifer sp. 2Leptodactylus podicipinus Leptodactylus petersiiElachistocleis ovalis Elachistocleis sp.
Em conjunto, as espécies de répteis presentes na porção norte do Pantanal
parecem apresentar distribuições mais amplas do que aquelas de localidades no planalto
(FIG. 3). A predominância, no Pantanal, é de répteis com ocorrência anotada para três
biomas, enquanto em Manso, no planalto, predominam formas com ocorrência em
apenas dois biomas.
FIG. 3. Distribuição das espécies de répteis da porção norte do Pantanal e de uma
localidade do entorno (Manso, Chapada dos Guimarães, MT) segundo sua
ocorrência em um, dois, ou mais biomas distintos.
No planalto, a multiplicidade de hábitats, incluindo fisionomias florestadas
estruturalmente heterogêneas, e de ecótones parece favorecer um maior número de
1 2 3 4 5 6
0
5
10
15
20
25
nº e
spéc
ies
nº biomas distintos
Planície (Pantanalnorte)Planalto (Manso)

24
especializações e, consequentemente, maior número de espécies que na planície
(Strussmann, 2000). A herpetofauna registrada até o momento para a região sob
influência do reservatório de Manso é uma das mais ricas já estudadas na região
neotropical, superando, em alguns casos, o número de espécies registrado em
localidades amazônicas (veja Duellman, 1990; O’Shea, 1998). A região abrange não só
fisionomias características de cerrados abertos como formações florestadas peculiares
(Conceição, 2000), que abrigam fauna especializada e servem como corredores de
dispersão para formas amazônicas.
Embora as espécies de répteis da planície sejam, em geral, mais amplamente
distribuídas do que aquelas ocorrentes nos planaltos do entorno, ainda assim podem ser
reconhecidas faunas locais diferenciadas, em distintos “pantanais”. Entre as proposições
de subdivisão da BAP em “zonas ecológicas”, “sub-regiões fisiográficas” ou
“Pantanais” diferenciados destacam-se aquelas de Adámoli (1982) e, mais
recentemente, Silva e Abdon (1998), que reconheceram e delimitaram onze sub-regiões
distintas no Pantanal brasileiro.
Nas sub-regiões pantaneiras, as diferenças em composição faunística parecem
estar associadas, primariamente, à maior ou menor influência de elementos chaquenhos,
pré-amazônicos, da floresta atlântica ou do cerrado sobre a biota. Entre as serpentes do
Pantanal de Poconé, por exemplo, na porção norte do Pantanal (aproximadamente entre
16° - 17°30’S e 56° – 57°30’W), Strussmann e Sazima (1993) apontaram como mais
abundantes as espécies Eunectes notaeus, Helicops leopardinus, Hydrodynastes gigas,
Liophis poecilogyrus, Thamnodynastes cf. strigilis. Já na região da Nhecolândia (entre
18° – 20° S e 55° – 57°W), município de Corumbá, porção sul do Pantanal, Leptodeira
annulata, Liophis typhlus e Lystrophis mattogrossensis seguramente estão entre as
formas mais comuns (Brasil, 1997). Tais espécies, relativamente bem representadas na
coleção regional da EMBRAPA-Pantanal, em Corumbá, não foram sequer registradas
para o Pantanal de Poconé, estando até o momento restritas aos cerrados do entorno
deste.
O Pantanal de Poconé foi incluído por Adámoli (1982) no grupo de pantanais
onde “os Cerrados ocupam só 10% da superfície, [estando] situados na periferia”
(Adámoli, 1982, p. 116). Já o da Nhecolândia, segundo o mesmo autor, apresenta entre

25
40 e 50% de sua superfície ocupada por cerrados (sendo evidentes, também, as
influências chaquenhas e amazônicas), o que pode explicar as diferenças em
composição ou, pelo menos, em abundância relativa das espécies de répteis ali
detectadas.
Também entre os anfíbios deve haver diferenças nos conjuntos faunísticos dos
diferentes pantanais. Em área inundável do Pantanal de Barão de Melgaço
(aproximadamente entre 16° - 17°30’N e 55° - 56°30’W), incluído por Adámoli (1982)
no grupo de pantanais “nitidamente pertencentes aos Cerrados, nos quais estes ocupam
aproximadamente 70% da superfície” (Adámoli, 1982, p. 115), um levantamento rápido
acusou espécies frequentemente encontradas nos cerrados periféricos ao Pantanal norte
(como Physalaemus nattereri, Leptodactylus mystacinus, Elachistocleis sp.). Ao longo
de dez anos de observações de campo, nenhuma dessas espécies foi ainda registrada
para regiões nucleares do Pantanal de Poconé5. Por outro lado, pela acentuada influência
chaquenha, é nos pantanais do Nabileque e de Porto Murtinho que, entre os répteis e
anfíbios, deve ocorre a maior proporção de formas caracteristicamente associadas ao
chaco oriental.
A subdivisão de faunas de anuros e de répteis em comunidades ou grupos de
espécies caracteristicamente associadas a unidades de paisagem ou a comunidades
vegetais peculiares consiste em exercício de grande importância e aplicabilidade em
conservação. Entre outros aspectos, pode contribuir para a caracterização e seleção de
sítios prioritários para conservação. Na FIG. 4 são mostrados perfis esquemáticos das
unidades de paisagem mais conspícuas na porção norte do Pantanal (FIG. 4a) e em
Manso (FIG. 4b), com as respectivas taxocenoses de anfíbios mais frequentemente
associadas a cada unidade.
5 C. Strussmann, dados não publicados.

26
FIG. 4a. Perfil esquemático das principais unidades de paisagem e das espécies deanuros mais freqüentes em cada unidade, na porção norte (Poconé, MT) daplanície inundável do Pantanal. Espécies arborícolas e subarborícolasrepresentadas acima da linha da superfície; espécies terrícolas e criptozóicasrepresentadas abaixo da linha da superfície.
FIG. 4b. Perfil esquemático das principais unidades de paisagem e das espécies deanuros mais freqüentes em cada unidade, em uma região de planalto, na bordanordeste do Pantanal, sob a influência do reservatório do AproveitamentoMúltiplo de Manso (Chapada dos Guimarães, MT). Espécies arborícolas esubarborícolas representadas acima da linha da superfície; espécies terrícolas ecriptozóicas representadas abaixo da linha da superfície.
Espécies ocorrentes em fisionomias abertas representam, aproximadamente,
50% da herpetofauna, tanto no Pantanal norte como em Manso. Entretanto, parece haver
menor compartimentação entre os répteis e anuros da planície do que entre aqueles de

27
regiões do planalto. Na porção norte do Pantanal, por exemplo, distintas fisionomias
abertas (campos inundáveis, bordas de baías, corixos, vazantes e outros) abrigam
anurofaunas bastante semelhantes. Estas incluem, além de formas terrícolas e
subarborícolas, duas espécies de hábitos semi-aquáticos, com grande habilidade para a
natação e o mergulho (Lysapsus limellus e Pseudis paradoxa).
A grande maioria das espécies de anuros do Pantanal suporta o período de seca
abrigada em locais úmidos (sob bancos de macrófitas em decomposição, troncos caídos,
em meio a emaranhados de raízes e outros) e rapidamente dispersa-se e coloniza os
campos, uma vez inundados (por exemplo, Hyla nana, H. raniceps, Scinax acuminatus,
S. fuscomarginatus). Espécies como Pseudopaludicola sp., Leptodactylus fuscus, L.
podicipinus, Bufo granulosus e Elachistocleis cf. ovalis permanecem nas proximidades
da linha de contato entre os biótopos aquáticos e terrestres (o chamado "moving
littoral", sensu Junk et al., 1989). Os pequenos Physalaemus vocalizam em águas rasas,
próximos à borda e há, também, espécies habitualmente associadas à vegetação
flutuante em corpos d’água permanentes (rios, corixos, baías), como Hyla punctata,
Scinax nebulosus.
Apenas três espécies de anuros da porção norte do Pantanal parecem estar
restritas a ambientes florestais (especialmente matas ciliares e as formações localmente
denominadas capões e cordilheiras): Bufo aff. margaritifer, Leptodactylus elenae e
Chiasmocleis mehelyi. Outras espécies, como Bufo paracnemis e Physalaemus cuvieri,
são euritópicas, podendo ser encontradas tanto no interior de capões e de cordilheiras
como em fisionomias abertas.
Em Manso, diferentemente do que ocorre no Pantanal, fisionomias abertas
distintas abrigam anurofaunas também diferenciadas. Há, assim, espécies
caracteristicamente associadas aos cerrados estabelecidos sobre areia (cerrado sentido
restrito), aos campos sujos de cerrado sobre pedra, outras exclusivas de veredas e
campos úmidos. Também entre as formas ocorrentes em ambientes florestados há clara
distinção entre os conjuntos de espécies de “matas de brejo”, de cerradões e de matas de
vales, por exemplo, com poucas espécies em comum entre esses ambientes.
A FIG. 5 apresenta a proporção de espécies utilizando diferentes substratos em
herpetofaunas da porção norte do Pantanal e da região de Manso. Em ambas as regiões,

28
tanto entre anfíbios como entre os répteis, formas terrícolas constituem maioria (entre
55 -63% do número total de espécies).
FIG. 5. Proporção de utilização de distintos substratos em herpetofaunas da planície
(porção norte do Pantanal) e do planalto adjacente (região da Usina
Hidrelétrica de Manso, Chapada dos Guimarães, Mato Grosso).
O segundo conjunto melhor representado, independente de grupo taxonômico ou
região, é o das espécies com hábitos arborícolas ou semi-arborícolas. Já a importância
relativa de formas com hábitos aquáticos ou semi-aquáticos e de formas com hábitos
fossoriais distingue as herpetofaunas da porção norte do Pantanal e da localidade no
planalto. Enquanto, na primeira, a representatividade de anfíbios e de répteis com
hábitos aquáticos ou semi-aquáticos é superior à de formas fossórias, no planalto ocorre
o inverso, sendo as formas de hábitos fossoriais relativamente mais numerosas do que
na planície.
Para serpentes da porção norte do Pantanal, o favorecimento da presença de
espécies aquáticas/semi-aquáticas foi tentativamente atribuído por Strussmann e Sazima
(1993) às condições ecológicas imperantes na planície, notadamente a grande
disponibilidade de corpos d’água e recursos associados (Strussmann e Sazima, 1993).
Para os cerrados, Vanzolini (1971) menciona como principais condicionantes do
favorecimento de formas fossórias a penetrabilidade do solo até níveis profundos (“o
60 %25 %6 %
9 %
55 %
30 %10 %
5 %
0 10 20 30 40 50 60
% do nº total de espécies
PLANALTO (Manso)
PLANÍCIE (Pantanalnorte)
Terrícolas/criptozóicas Arborícolas/subarborícolas Aquáticas/semiaquáticas Fossórias

29
volume e profundidade dos sistemas radiculares e dos troncos subterrâneos facultam
densa penetração da fauna até níveis de boa estabilidade térmica e favoráveis teores de
umidade”; Vanzolini, 1971, p.270-271), e a intensidade das trocas de radiação ao nível
do solo, com as temperaturas, durante o dia, podendo atingir valores extremamente
altos. Em Manso foram registradas, até agora, oito espécies simpátricas de
anfisbenídeos, tanto em fisionomias florestadas como abertas, havendo também
serpentes e lagartos de hábitos semifossórios. No Pantanal, a estrutura arenosa dos
solos, seu encharcamento periódico, a cobertura graminosa rala e uniforme, a reduzida
disponibilidade e o isolamento de áreas permanentemente secas possivelmente
constituem limitações para a vida subterrânea (Strussmann, 1992). As únicas espécies
de répteis fossórios ou semifossórios até agora registrados para o Pantanal de Poconé,
Amphisbaena vermicularis, Typhlops brongersmianus, Micrurus tricolor e Phalotris
tricolor, foram sempre encontrados em capões ou cordilheiras.
Padrões de abundância
Com diversidade de espécies e grau de endemismo inferiores aos dos planaltos
do entorno, é na presença sazonal de grandes contingentes populacionais, com elevada
biomassa, que a herpetofauna da planície apresenta seu maior diferencial. Para anfíbios,
que apresentam estreita dependência de água livre no ambiente, a maior disponibilidade
de corpos d’água permanentes e a periódica transformação de enormes extensões de
biótopos terrestres em biótopos aquáticos possibilitam o estabelecimento de populações
vigorosas na planície pantaneira, especialmente durante a estação chuvosa.
Favorecimento semelhante pode ser visto entre os répteis de hábitos aquáticos ou
semi-aquáticos, sendo particularmente abundantes na planície o jacaré-do-pantanal
(Caiman yacare), a jaracussu-do-brejo (Hydrodynastes gigas), a sucuri-amarela
(Eunectes notaeus) e a cobra-d’água (Helicops leopardinus). Brown Junior (1986)
sugere uma tendência ao favorecimento, na planície, de predadores generalistas e de
organismos “móveis, grandes, resistentes a longas estiagens e friagens e ligados a
ambientes aquáticos”. Caiman yacare, E. notaeus e H. gigas satisfazem tais condições.
A maior abundância de presas é, reconhecidamente, uma das principais causas
de aumento na atividade de serpentes após o início do período chuvoso. A sucuri-

30
amarela alimenta-se, basicamente, de aves paludícolas, recurso sazonalmente abundante
e previsível no Pantanal. Na América do Sul, diversas outras áreas sazonalmente
inundáveis apresentam, ocasionalmente, uma avifauna paludícola rica e diversificada
(Scott e Carbonell, 1986). Na costa leste do Amapá e da Ilha de Marajó, por exemplo,
assim como nos llanos da Venezuela e no Departamento de Beni, na Bolívia, grande
número de aves aquáticas residentes e migratórias reunem-se anualmente para
reprodução, alimentação ou “desasagem” (fide Sick, s.d.). Em todas essas áreas,
conforme anteriormente observado por Strussmann (1992), foi assinalada a presença de
serpentes do gênero Eunectes (Scott e Carbonell, 1986; Nascimento et al., 1991), por
vezes em elevadas densidades, como nos llanos (Rivas, 1999) e na ilha de Marajó6.
A serpente mais abundante na parte norte da planície, o colubrídeo H. gigas (v.
Strussmann e Sazima, 1993), parece ser favorecida por sua grande plasticidade
adaptativa: utiliza tanto hábitats abertos como florestados, terrenos secos como
inundados, sendo capaz de explorar, alternativamente, os recursos mais abundantes em
cada estação. Na seca, procura por roedores, sapos (Bufo paracnemis) e outros pequenos
vertebrados sob coivaras ou em meio a emaranhados de gramíneas e vegetação rasteira;
durante as cheias, explora o “moving littoral” à procura de rãs (Leptodactylus spp.); no
período de vazante, foi observada caçando pequenos peixes aprisionados em poças em
vias de secar.
Também muito comum no Pantanal, a cobra d´água não satisfaz a premissa de
Brown Junior (1986) por seu pequeno tamanho (em geral, < 50 cm de comprimento
total). Entretanto, H. leopardinus alimenta-se basicamente de peixes, recurso abundante
tanto na planície como na calha dos rios e outros corpos d´água permanentes, onde estas
serpentes permanecem durante a estação seca.
Anfíbios anuros, cuja abundância é drasticamente aumentada no Pantanal,
durante o período chuvoso (em resposta a aumento na produção primária e nas
populações de consumidores de primeira ordem), também constituem presas
importantes para serpentes da planície. Cerca de 42% das espécies do Pantanal de
Poconé utilizam anfíbios como alimento principal. Algumas, como Liophis
6 L. Dirksen, comunicação pessoal

31
poecilogyrus e Thamnodynastes sp., figuram também entre as mais abundantes da
ofiofauna local (Strussmann e Sazima, 1993).
Dentre as cerca de 30 espécies de anfíbios conhecidas da porção norte da
planície, Physalaemus albonotatus é uma das mais constantes e conspícuas durante o
período chuvoso, quando forma grandes coros, mesmo durante o dia. Outra espécie
diurna localmente abundante é Lysapsus limellus, diminuta rã verde de hábitos semi-
aquáticos. Comumente encontrada em meio a aguapés (Eichhornia spp., Salvinia
auriculata), L. limellus possivelmente é favorecida pela grande disponibilidade de
invertebrados fitófagos que participam do processo de decomposição de grandes
aglomerados dessas macrófitas, em ambientes aquáticos em vias de secar. Igualmente
diurna e abundante no Pantanal, mas com hábitos e desovas terrestres, é Adenomera cf.
hylaedactyla, encontrada em capões, cordilheiras e outras formações florestadas.
A grande diversidade e a intensidade das vocalizações de anuros em agregações
reprodutivas, após as primeiras chuvaradas no Pantanal, foram registradas por Lutz
(1946) para uma localidade no Pantanal de Miranda, Mato Grosso do Sul. A região
apresenta influência de cerrado de magnitude semelhante à observada no Pantanal da
Nhecolândia (entre 40 e 50% da superfície coberta por cerrados, segundo Adámoli,
1982).
Apesar da crescente descaracterização por que vêm passando enormes extensões
de cerrado, especialmente nos arredores de grandes cidades, agregações reprodutivas
comparáveis às descritas por Lutz (tanto em composição como na abundância relativa
das espécies participantes) têm sido repetidamente observadas em áreas parcialmente
urbanizadas (originalmente cobertas por cerrados) na cidade de Cuiabá, borda noroeste
do Pantanal norte.
Assim, enquanto chegam notícias, de diversas partes do mundo, sobre extinções
locais e declínios populacionais de anfíbios (GAD – “Global Amphibian Decline”, v.
Beebee, 1996), parece haver indicações consistentes de que, tanto na porção norte do
Pantanal como em seu entorno imediato, as comunidades herpetofaunísticas ainda se
apresentam razoavelmente bem conservadas. Inventários rápidos na região das altas
cabeceiras do rio Taquari, em Mato Grosso do Sul, evidenciaram alto grau de
empobrecimento das comunidades locais, especialmente de anuros, em decorrência de

32
profundas modificações ambientais resultantes de práticas inadequadas de agricultura e
pecuária extensivas (Strussmann et al., 2000). Assim, embora não possam ser
generalizadas para toda a BAP, as evidências sobre a situação das populações de
anfíbios e de répteis em áreas do Pantanal norte e seu entorno revelam a necessidade e o
potencial de estudos de monitoramento de populações de répteis e de anfíbios, como
eficiente medida para a conservação de áreas no Pantanal e no entorno.
AVES
Estão registradas 444 espécies de aves para a planície pantaneira, entendida aqui
como somente a parte inundável do Pantanal (v. Tubelis e Tomás, neste volume)
enquanto que para os Cerrados há 837 espécies das quais 759 (90,7%) se reproduzem no
bioma (Silva, 1995). Contudo, para o Pantanal, como a região é definida neste trabalho
(veja Silva e Abdon, 1998), há 661 espécies. Em dois levantamentos realizados, um no
Pantanal, na região de Barão de Melgaço, outro na região do planalto, área do rio Manso
na Chapada dos Guimarães, foram registradas, respectivamente, 264 espécies (Alho et
al., 1998) e 259 espécies de aves (Vasconcellos, 2000). Essas observações de campo,
bem como a consistente literatura publicada para ambas as regiões, mostram que, no
Pantanal, as aves paludícolas concentram-se ou agregam-se, no período de seca, em
torno de alimento (guildas alimentares) e em locais de reprodução, os ninhais (guildas
reprodutivas). Para muitas espécies paludícolas, o Pantanal é considerado como o
principal ambiente aquático do interior do continente sul-americano, abrigando
populações de tuiuiús (Jabiru mycteria) em toda sua extensão (Antas e Nascimento,
1996). Esta espécie, como diversas outras que se alimentam e se reproduzem no
Pantanal, aproveitando-se do regime de enchente e vazante, deixa anualmente a região,
em movimentos migratórios entre a planície e o planalto ou em direção ao sul como é o
caso, também, do cabeça-seca Mycteria americana. O tuiuiú, por exemplo, mostra
grande capacidade de deslocamento quando os animais são marcados por
rádiotransmissores (Antas e Nascimento, 1996). Mesmo dentro da planície as
populações deslocam-se em busca de pontos onde haja maior disponibilidade de
alimento, onde se concentram. Outro aspecto relevante é o sincronismo existente entre o

33
regime de enchente e vazante e o sucesso reprodutivo desta e de outras espécies, como o
cabeça-seca. Em casos de enchentes atípicas tem sido observado abandono de ninhos
(Antas e Nascimento, 1996). Os resultados das investigações no campo desses autores
indicam que essa correspondência pode ser evidenciada pela existência de período
reprodutivo sazonal e pelas flutuações no sucesso reprodutivo entre anos. Os ninhos são
territórios reprodutivos, localizados em árvores mortas ou vivas de piúvas e manduvis,
defendidas ativamente pelos casais que nelas nidificam, ano após ano. Os hábitats
preferidos, segundo os resultados desses estudos, são áreas inundáveis próximas a
capões de mata. Das 661 espécies que ocorrem no Pantanal, 77% (508 espécies)
ocorrem em hábitats florestados ou arborizados, como os diversos tipos de matas,
cerradão, cerrado e áreas alteradas; 13% (89 espécies) dependem de hábitats com a
presença de rios, baías, campos inundáveis e de outros abertos e, finalmente, 10% (64
espécies) distribuem-se em hábitats também abertos de rios, baías e campos inundáveis
(FIG. 6). De fato, o Pantanal concentra uma das maiores abundâncias de aves
paludícolas da região neotropical (Stotz et. al., 1996). São treze espécies de garças,
pertencentes aos gêneros Casmerodius, Egretta e Pilherodius, além de cabeças-secas e
jaburus (gêneros Mycteria, Euxenura e Jabiru), acrescidos de colhereiros, íbis, martins-
pescadores e outros de hábitos aquáticos.

34
FIG. 6. Hábitats utilizados por espécies de aves do Pantanal. Em azul, espécies de aves
(89) com ocorrência exclusiva no ambiente aquático; em laranja, espécies (64)
com ocorrência predominante em ambiente aquático; em amarelo, espécies
(508) com ocorrência predominante em ambientes florestados.
A adaptação aos diversos tipos de hábitats leva a diferentes níveis de
distribuição de dietas (FIG. 7), onde somente um percentual baixo de espécies de aves
(5%) dependem de alimentação piscívora.
FIG. 7. Distribuição das dietas da avifauna do Pantanal. O maior valor percentual
corresponde à guilda dos insetívoros. Entretanto, na guilda dos piscívoros
encontra-se a grande abundância da avifauna do Pantanal.
Insetívora39%
Nectarívora4%
Necrófaga1%
Omnívora18%
Piscívora5%
Granívora8%
Carnívora7%
Frugívora18%
77%
13%10%

35
Igualmente, a análise da estratificação das espécies que ocorrem no Pantanal
quanto às suas adaptações a hábitats evidencia que apenas 13% das espécies são
estritamente terrestres, enquanto outros 15% ocupam ocasionalmente esse estrato (FIG.
8).
FIG. 8. Utilização dos estratos (hábitats) pela avifauna do Pantanal. A – água; T –
terrestre; T/A – terrestre + água; T/S – terrestre + sub-bosque; T/Em – terrestre
+ estrato médio; T/D – terrestre + dossel; T/Ar – terrestre + aéreo; S – sub-
bosque; S/Em – sub-bosque + estrato médio; S/D – sub-bosque + dossel; Em –
estrato médio; Em/D – estrato médio + dossel; D – dossel; D/Ar – dossel +
aéreo; Ar – aéreo.
Em algumas regiões, como na sub-região de Poconé, durante o período de
garimpo intenso de ouro, algumas espécies de aves que se alimentam de peixes,
moluscos e crustáceos foram encontradas contaminadas por mercúrio, por causa do
processo de biomagnificação desse metal na cadeia trófica (Alho e Vieira, 1997). Entre
essas espécies estão o biguá (Phalacrocorax olivaceus), o carão (Aramus guarauna), a
aninga (Anhinga anhinga), o gavião-caramujeiro (Rosthramus sociabilis) e a garça-
branca (Casmerodius alba).
T13%
S/D4%
Em3%
Em/D9%
D28%
D/Ar1%
Ar3%
S/Em9%
T/A5%
A3%
T/S3%
T/Em1%
T/D5%
T/Ar1%
S12%

36
MAMÍFEROS
Para procurar entender o padrão presente da diversidade e abundância da fauna
de mamíferos do Pantanal é necessário considerar os fatores gerais que controlam esse
padrão nas escalas temporal e espacial, isto é, evidências históricas ou evolutivas e
processos ecológicos atuais que governam esses padrões, principalmente os ciclos
anuais de enchente-vazante.
Sob a ótica evolutiva, o panorama presente de diversidade é relativamente
recente. A radiação ou dispersão de mamíferos deu-se no Pleistoceno posterior e início
do Holoceno, obedecendo o eixo de altas latitudes, como da região austral da América
do Sul, para baixas latitudes, após a chegada dos diferentes estratos faunísticos desde o
Cenozóico, acompanhando a deriva continental (Webb, 1992). O "corredor" de
dispersão das savanas ocorreu no Plioceno posterior e início do Pleistoceno, com
intercâmbio de mamíferos nas Américas. Ocorreu, então, uma extinção em massa dos
mamíferos de grande porte, começando a dispersão e ocupação de nichos pelos
presentes mamíferos de médio e pequeno portes. No Pleistoceno estima-se que a fauna
dos Cerrados/Pantanal pudesse ter cerca de 35% das espécies conhecidas hoje. Isso
significa dizer que o panorama de diversidade conhecido hoje é relativamente recente,
com ocupação de nichos e especiação recentes.
Os Cerrados contam com 195 espécies de mamíferos enquanto que o Pantanal
tem 132 espécies (FIG. 9). A maioria das espécies de ambos os biomas depende de
hábitats florestados.

37
FIG. 9. Número de espécies de mamíferos no Cerrado (195) e no Pantanal (132),
destacando as preferências por hábitats florestados e abertos.
A maior parte da heterogeneidade observada em uma comunidade ecológica é
determinada pela vegetação, principalmente a arbórea ou florestal, que forma uma
matriz de grande complexidade subdividida em diferentes estratos, ocupados pelos
mamíferos. A fauna do Pantanal e dos Cerrados apresenta uma forte dependência da
complexidade dos hábitats arbóreos e florestados. Essa dependência pode ser avaliada
em função das características do macronicho de cada uma das espécies de mamíferos.
Para cada uma das espécies analisadas atribui-se um valor em função de sua
dependência de hábitat arbóreo ou florestado em termos do tamanho do mamífero, do
requisito de hábitat, da dieta e do tipo de locomoção. Para essa análise, foram utilizadas
as quinze espécies de mamíferos mais abundantes (isto é, mais encontradas em censos
por avistamentos, transectos, captura-recaptura ou por sinais de presença como pegadas,
vocalização e outros indícios). Para cada uma das características do macronicho é
atribuído um valor numérico ordinal que aumenta na medida em que sua dependência
também aumenta. O tamanho da espécie de mamífero é considerado um elemento de
dependência visto que a vagilidade, isto é, a capacidade do animal em deslocar-se,
aumenta com o seu tamanho. A capacidade de deslocamento de um mamífero tende a
aumentar inversamente com o seu tamanho. O tamanho do animal, para efeito dessa
análise, foi considerado em termos de classe de peso. Para o hábitat os mamíferos foram
classificados como: (a) dependentes de mata; (b) medianamente dependentes e (c) não
dependentes. No valor para as categorias de dieta considera-se que a dependência da
13262
5515
19595
8614
0 20 40 60 80 100 120 140 160 180 200
PANTANAL
CERRADO
Total de espécies no bioma VoantesDependentes de hábitats florestados Dependentes de hábitats abertos e aquáticos

38
espécie em termos de dieta aumenta na medida em que o mamífero usa recursos mais
abundantes na floresta, como brotos novos, frutas e outros. Assim, as seguintes
categorias são consideradas, com seus valores, para a análise:
carnívoro/folívoro/frugívoro (6); frugívoro/omnívoro (5); frugívoro/herbívoro (4);
herbívoro/pastador (3); insetívoro/omnívoro (2) e mirmecófago (1). As categorias de
locomoção analisadas são: arborícola (5); escansorial (4); semifossorial ou fossorial (3);
terrestre (2) e semi-aquático (1). A medida da dependência da floresta (ou intolerância
ao desmatamento) é calculada em duas etapas. Na primeira, é feita a soma total dos
valores do macronicho para cada uma das quinze espécies de mamíferos (Vi). Depois é
aferida a diferença do valor de cada espécie do valor médio de todas as espécies
utilizadas na análise (X), se Vi-X é maior que 0 a espécie é considerada dependente.
Quando Vi-X é menor que 0, é não dependente. Com essa análise são identificadas as
espécies do Pantanal e do planalto (cerrado da região de Manso) que apresentam alto
grau de dependência de hábitats arbóreos e florestados (FIG. 10).
FIG. 10. Dependência de hábitats florestados (mata de galeria, mata ciliar,
cambarazal, paratudal, mata semi-decídua, cerradão, cerrado denso, e
outras formas florestadas), ou intolerância ao desmatamento, das 15
espécies de mamíferos mais abundantes no Pantanal.

39
Quando quinze espécies mais comuns (mais encontradas por diversos métodos
em censos realizados) de mamíferos são examinadas quanto à abundância em ambientes
do Pantanal e dos Cerrados de Manso, há diferenças marcantes em número de animais,
expressos em percentagens (FIG. 11). A FIG. 11 mostra de maneira marcante a
diferença em abundância de espécies comuns ocorrendo na planície e no planalto. O
coati Nasua nasua foi o animal mais observado no pantanal da Nhecolândia (Alho et al.,
1988a). É animal diurno e escansorial, forrageando no chão ou áreas abertas na estação
seca e também em hábitat arbóreo. Alimenta-se oportunisticamente de caranguejos e
outros invertebrados na época seca e de frutas e outros recursos da floresta na época
cheia. O lobinho Cerdocyon thous foi a segunda espécie mais observada em censo
conduzido na região da Nhecolândia, ocorrendo também com freqüência na região de
Manso. Essas diferenças em número relativos de animais de ampla distribuição
geográfica são favorecidas por ofertas de requisitos no hábitat, principalmente de
alimento e espaço, expressos em termos de abundância (FIG. 12). O bugio Alouatta
caraya é visto com freqüência no dossel das matas, principalmente ao longo dos rios do
Pantanal. São animais herbívoros, que dependem dos ritmos fenológicos da floresta, isto
é, da produção de brotos novos, flores, frutos e semente.
FIG. 11. Abundância relativa de espécies de mamíferos do Pantanal (mais
avistadas/anotadas em diferentes tipos de censos), expressa em termos de
hábitats disponíveis.

40
FIG. 12. Abundância relativa de espécies de mamíferos de Manso, Chapada dos
Guimarães, MT (mais avistadas/anotadas em diferentes tipos de censos),
expressa em termos de hábitats disponíveis.
Essa diferença em abundância pode ser ainda visualizada em termos de
adaptação ao tipo de substrato ou hábitat (FIG. 13) e adaptação ao hábito ou nicho
alimentar (FIG. 14). O cervo-do-pantanal Blastocerus dichotomus é susceptível às
mudanças sazonais visto que a maior oferta de alimento está disponível na estação seca
(Tomás, 1993). Todos os ambientes onde os cervos são avistados em censos são hábitats
sazonalmente inundáveis. Essas mudanças causam diferenças significativas na
densidade e distribuição dos animais. Do mesmo modo, o regime de enchente e vazante
influencia significantemente as populações de capivaras no Pantanal (Alho et al., 1989).
O uso de hábitat varia sazonalmente. Na seca, as capivaras passam grande parte da noite
nas matas (capões de cerrado) e forrageiam nos campos durante o dia. Na cheia, passam
mais tempo nas matas, inclusive mudando seu regime alimentar, com maior consumo de
espécies encontradas nas matas e nas áreas inundadas. O comportamento social e a
estrutura social do grupo também varia sazonalmente. Os grupos sociais são maiores e
de estrutura diferente na estação da seca. Igualmente, o espaço domiciliar de cada grupo
social é maior na época seca.

41
FIG. 13. Estratificação das quinze espécies de mamíferos de médio e grande portes mais
abundantes no Pantanal e no Cerrado de acordo com a adaptação ao hábitat
(uso do substrato).
FIG. 14. Estratificação das quinze espécies de mamíferos de médio e grande portes mais
abundantes no Pantanal e no Cerrado de acordo com o hábito alimentar
(adaptação ao nicho alimentar).
0
50
100
150
200
250
300
350
aquático e semi-aquático
terrestre arborícola
substrato utilizado
abun
dânc
ia re
lativ
a
Pantanal
Cerrado
0
100
200
300
400
500
600
herbívoro/folívoro/frugívoro frugívoro/granívoro/omnívoro
nicho alimentar
abun
dânc
ia re
lativ
a Pantanal
Cerrado

42
Os pequenos mamíferos (roedores silvestres e marsupiais) dos Cerrados
mostram maior diversidade em matas de galerias, embora alguns cricetídeos como
Bolomys lasiurus sejam abundantes em áreas abertas com campos (FIG. 15).
FIG. 15. Distribuição de pequenos mamíferos em hábitats de cerrado (região de
Brasília) e do Pantanal (região da Nhecolândia).
Os pequenos mamíferos do Pantanal também são encontrados em áreas arbóreas,
como os capões de cerrado, sendo que algumas espécies se especializam em explorar os
ecótones desses ambientes com os campos, como é o caso de Clyomys laticeps (FIG.
15). A inundação periódica dos campos é um impedimento para os habitantes dessas
áreas essencialmente abertas. Somente 1,6% dos animais são capturados em armadilhas
de censos em áreas inundáveis. Enquanto nos ambientes abertos de Cerrado Bolomys
lasiurus é o animal mais capturado, no Pantanal da Nhecolândia, Oryzomys fornesi é o
roedor mais generalista, encontrado em três tipos diferentes de hábitats (Lacher e Alho,
1989). Enquanto os pequenos mamíferos do Pantanal concentram-se nos capões de
cerrado ou nas matas de galeria, nos Cerrados de Manso, dez de dezenove espécies
estudadas são restritas a um tipo de hábitat e seis são restritas a dois tipos. Há espécies

43
identificadas como generalistas, como Bolomys lasiurus, ocorrendo em quatro dos sete
tipos de hábitats que ocorrem em Manso, Essa espécie é praticamente ausente dos
campos inundávies do Pantanal. Os marsupiais de Manso (seis espécies estudadas)
mostram grande especialização por hábitats de florestas, como os do Pantanal. Em
Manso, só Monodelphis domestica mostra ser mais generalista, habitando também
hábitats abertos (Schneider, 2000).
CONSERVAÇÃO E OPÇÕES SOCIOECONÔMICAS
A fauna do Pantanal tradicionalmente representa motivo para atrair o turista. O
ecoturismo, se bem orientado, é no momento a melhor opção socioeconômica para a
região e pode trazer benefícios para a conservação.
O programa conhecido como Projeto Pantanal, liderado pelo Ministério do Meio
Ambiente, em cooperação com os governos de Mato Grosso e Mato Grosso do Sul, e
com financiamento de 400 milhões de dólares pelo Banco Interamericano de
Desenvolvimento, considera o componente ecoturismo. Nesse componente há a
previsão de implementação de estradas-parque, nos dois Estados.
O projeto de construção da hidrovia Paraná-Paraguai para o trajeto de grandes
comboios de carga, particularmente no trecho do Pantanal entre Corumbá e Cáceres,
tem sido ultimamente rediscutido para implantação, apesar das conseqüências
ambientais apontadas como prejudiciais (Hamilton, 1999; Huszar et al., 1999). Conta-se
com a oportunidade desse grande projeto para destacar seguintes ameaças correntes ao
Pantanal:
a) fogo - Os incêndios sem controle têm sido um problema, trazendo
alterações na vegetação e ameaçando diretamente a fauna local;
b) desmatamento - A substituição de ambientes arbóreos, principamente
capões de cerrado e cerradão e outros ambientes florestados, em pastagens para o
gado, tem sido documentado na literatura (Mourão, 1996; Silva et al., 1998). Como
indicado neste trabalho, a diversidade da fauna do Pantanal depende grandemente de
hábitats florestados;

44
c) gado - A pecuária pantaneira atravessa nos últimos anos crise pela
dificuldade de competir com a pecuária mais produtiva de outras regiões fora do
Pantanal. Um elemento importante, para o turismo organizado, é o aspecto cultural e
tradicional das fazendas pantaneiras, como atrativo socioeconômico e cultural;
d) contaminantes ambientais - O Pantanal tem sido foco de vários tipos de
contaminação que afetam diretamente a fauna ou indiretamente pela modificação de
seus hábitats. Um problema é biomagnificação de mercúrio na cadeia trófica,
oriundo dos garimpos de ouro e, também, o uso sem controle de agrotóxicos (Alho
et al., 1988b; Alho e Vieira, 1997). Outro problema sério é o despejo de esgoto das
grandes cidades nos rios que inundam o Pantanal, como é o caso da cidade de
Cuiabá, no rio Cuiabá;
e) projetos de infra-estruturas altamente impactantes como a hidrovia
Paraná-Paraguai e o centro petroquímico de Corumbá voltam, com força política, a
serem discutidos para implantação;
f) turismo desorganizado - O potencial turístico do Pantanal continua a ser
explorado pelo turismo anárquico das grandes barcas-hotel transportando
pescadores, os acampamentos de pescadores em matas ciliares e outros. Espera-se
que o componente ecoturismo do Projeto Pantanal do MMA-BID possa trazer
disciplina para este setor, essencial para a região;
g) atropelamento de fauna silvestre - As recém-abertas estradas
pavimentadas no Pantanal têm trazido atropelamentos de animais (Fisher, 1997); a
implantação das estradas-parque necessariamente deve abordar a questão, nos
planos de manejo;
h) áreas protegidas e espécies ameaçadas - Esse também é um componente
importante do Projeto Pantanal, que se espera traga implementação consistente das
as unidades de conservação e corredores ecológicos identificados.

45
OBSERVAÇÕES CONCLUSIVAS
1. Há 85 espécies de répteis ocorrentes na planície inundável (sem serem
exclusivas da região) e outras 94 somente registradas, até o momento, em áreas do
entorno, num total de 179 espécies de répteis para a BAP. A anurofauna da BAP,
por sua vez, está constituída por pelo menos 80 espécies, das quais mais da metade
(45 espécies) parece ocorrer exclusivamente em áreas altas no entorno;
2. Apresentando menor heterogeneidade de hábitats e maior disponibilidade
de corpos d’água permanentes (menor número de hábitats com condições
ecológicas contrastantes), a planície inundável abriga espécies de anfíbios em geral
abundantes e de ampla distribuição. Já as áreas de entorno da BAP, especialmente
as altas cabeceiras, são virtualmente desconhecidas do ponto de vista de suas
anurofaunas, os poucos inventários existentes indicando haver, aí, maior número de
espécies num dado sítio e maior “turnover” de espécies entre sítios;
3. Na planície, formas exclusivas de biomas abertos representam c. 54 % da
fauna total de répteis, enquanto no planalto essa contribuição eqüivale a apenas 42
%. O número de formas “amazônicas”, por sua vez, é duas vezes maior na fauna de
répteis de localidade no planalto do que entre os répteis registrados para o Pantanal;
4. Distintas sub-regiões do Pantanal abrigam herpetofaunas também
diferenciadas;
5. A importância relativa de formas com hábitos aquáticos ou semi-
aquáticos e de formas com hábitos fossoriais distingue as herpetofaunas da porção
norte do Pantanal e da localidade no planalto. Enquanto, na primeira, a
representatividade de anfíbios e de répteis com hábitos aquáticos ou semi-aquáticos
é superior àquela de formas fossórias, no planalto ocorre o inverso, sendo as formas
de hábitos fossoriais relativamente mais numerosas do que na planície;
6. A grande abundância documentada da avifauna do Pantanal corresponde a
aves paludícolas que se agregam em torno de recursos sazonais alimentares ou
reprodutivos, formando, em muitos casos, guildas de espécies;
7. O comportamento reprodutivo de algumas espécies mostra sincronismo
entre o regime de enchente e vazante, com reflexos no sucesso reprodutivo;

46
8. Há deslocamento sazonal das aves paludícolas entre a planície e o
planalto, ou mesmo entre diferentes sub-regiões da planície, embora esses dados
ainda sejam insipientes para muitas espécies e consubstanciadas para outras, como
é o caso do tuiuiú;
9. Tanto no Pantanal quanto nos Cerrados a grande diversidade de espécies
resulta, em grande parte, da presença de ambientes florestados nesses dois biomas,
tais como matas ciliares, matas de galeria, cerradão e demais formações
arborizadas;
10. Desmatamento ou alteração de hábitats do Pantanal podem interferir na
diversidade de avifauna da região, já que a maior diversidade concentra-se em
hábitats florestados ou arborizados;
11. A concentração espetacular de aves no Pantanal constitui elemento
fundamental para o ecoturismo da região, com fortíssimo apelo ao uso sustentável
deste recurso para a região e como incentivo econômico à conservação dos hábitats
e suas espécies associadas;
12. Mesmo mamíferos com ampla distribuição encontram na planície
pantaneira ambientes mais propícios de oferta de nichos alimentares e reprodutivos,
dentre outros, que favorecem grande abundância, principalmente em áreas abertas
próximas a áreas arbóreas, casos da capivara e do cervo-do-Pantanal;
13. As espécies de mamíferos menos conspícuas, isto é, aquelas menos
avistadas, estão associadas a hábitats florestados, dos quais dependem para sua
sobrevivência. O atual nível de desmatamento de ambientes arbóreos para
transformação em pastos pode trazer ameaças à diversidade de mamíferos do
Pantanal, beneficiando somente umas poucas espécies, abundantes em áreas
abertas;
14. As análises que mostram as espécies de mamíferos mais dependentes de
ambientes florestados podem identificar espécies indicadoras ou espécies-chave
para esse tipo de ambiente, como é o caso do bugio Alouatta caraya para as matas
ciliares ao longo dos rios do Pantanal. Por outro lado, as espécies indicadoras de
ambientes abertos, como a capivara e o cervo, têm grande apelo para o ecoturismo
da região;

47
15. O regime de enchente e vazante tem uma influência direta no uso do
espaço, na densidade das populações, na estratégia alimentar, na estrutura social de
mamíferos abundantes no Pantanal, como está bem documentado para capivaras e
cervos;
16. Embora, no Pantanal, ambientes arbóreos sejam frequentemente
transformados em pastagens para o gado, a fauna silvestre de mamíferos ainda está
bem conservada, com espécies oficialmente declaradas ameaçadas sendo
observadas com relativa facilidade, visto que os animais silvestres transitam
livremente pelos interstícios de pastagens implantadas e hábitats arbóreos e
florestados naturais.

48
REFERÊNCIAS BIBLIOGRÁFICAS
AB’SABER, A.N. O Pantanal Mato-grossense e a teoria dos refúgios. Revista
Brasileira de Geografia, v.50, n.especial, p.9-57, 1988.
ABDON, M. de M.; SILVA, J.S.V.; POTT, V.J.; SILVA, M.P. Utilização de dados
analógicos do Landsat-TM na discriminação da vegetação de parte da sub-região da
Nhecolândia no Pantanal. Pesquisa Agropecuária Brasileira, v.33, n.especial,
p.1799-1813, out., 1998.
ADÁMOLI, J.; POTT, A. Las fuentes de diversidad en el Pantanal. In: MATTEUCCI,
S.D.; SOLBRIG, O.T.; MORELLO, J.; HALFTER, G. (Ed.) Biodiversidad y uso
de la tierra: conceptos y ejemplos de Latinoamérica. Buenos Aires: Eudeba, 1999.
p.317-360.
ADÁMOLI, J.. O Pantanal e suas relações fitogeográficas com os cerrados: discussão
sobre o conceito “Complexo do Pantanal”. In: CONGRESSO NACIONAL DE
BOTÂNICA,32., 1982, Teresina. Anais... Teresina:SBB, 1982. p. 109-119.
ALHO, C.J.R.; CAMPOS, Z.M.S.; GONÇALVES, H.C. Ecology, social behavior, and
management of the Capybara (Hydrochaeris hydrochaeris) in the Pantanal of
Brazil. In: REDFORD, K.F.; EISENBERG, J.F. (Ed.) Advances in neotropical
mammalogy. Gainesville: University of Florida/Sandhill Crane, 1989. p.163-194.
ALHO, C.J.R.; CONCEIÇÃO, P.N.; STRUSSMANN, C.; VASCONCELLOS, L.A.S.;
SCHNEIDER, M. Plano de manejo da RPPN SESC-Pantanal. Brasília:
FUNATURA, 1998. 121p.
ALHO, C.J.R.; LACHER JR, T.E.; CAMPOS, Z.M.S.; GONÇALVES, H.C.
Mamíferos da Fazenda Nhumirim, sub-região de Nhecolândia, Pantanal do Mato
Grosso do Sul: levantamento preliminar de espécies. Revista Brasileira de
Biologia, v.48, n.2, p.213-225, 1988a.

49
ALHO, C.J.R.; LACHER JR, T.E.; GONÇALVES H.C. Environmental degradation in
the Pantanal ecosystem of Brazil.. BioScience, v.38, n.3, p.164-171, 1988b.
ALHO, C.J.R.; VIEIRA, L.M. Fish and wildlife resources in the Pantanal wetlands of
Brazil and potential disturbances from the release of environmental contaminants.
Environmental Toxicology and Chemistry, v.16, n.1, p.71-74, 1997.
ANTAS, P.T.Z.; NASCIMENTO, I.L.S. Tuiuiú: sob os céus do Pantanal. Brasília:
SEMAVE e Monsanto, 1996. 169p.
BEEBEE, T.J.C. Ecology and conservation of amphibians. London: Chapman e
Hall, 1996. 214p.
BRASIL. Ministério do Meio Ambiente dos Recursos Hídricos e da Amazônia Legal.
Plano de Conservação da Bacia do Alto Paraguai-PCBAP, v.II,t.III. Diagnóstico
dos meios fisicos e biótico, meio biótico. Brasília: MMA/PNMA, 1997. 433p.
BRITSKI, H.A.; SILIMON, K.Z. de S.; LOPES, B.L. Peixes do Pantanal: manual de
identificação. Corumbá: Embrapa Pantanal, 1999. 184p.
BROWN JUNIOR, K.S. Zoogeografia da região do Pantanal Mato-grossense. In:
SIMPÓSIO SOBRE RECURSOS NATURAIS E SÓCIO-ECONÔMICOS DO
PANTANAL, 1., 1984, Corumbá. Anais... Brasilia: EMBRAPA-DDT, 1986.
p.137-178. (EMBRAPA-CPAP. Documentos, 5).
CADAVID GARCIA, E.A.; CASTRO, L.H.R. Análise da frequencia de chuvas no
Pantanal Mato-Grossense. Pesquisa Agropecuária Brasileira, v.21, n.9, p.909-
925, 1986.
CHERNOFF, B.; WILLINK, P.W. Biodiversity patterns within the Pantanal, Mato
Grosso do Sul, Brazil. In: WILLINK, P.W.; CHERNOFF, B.; ALONSO, L.E.;
MONTAMBAULT, J.R. (Ed.) A biological assessment of the aquatic ecosystems
of the Pantanal, Mato Grosso do Sul, Brasil. Washington: Conservation
International, 2000. p.103-106. (Bulletin of Biological Assessment, 17).

50
MOURÃO, G. Uso de levantamento aéreo para estudo de distribuição e
abundância de grandes vertebrados no Pantanal Mato-grossense.
Manaus:UFA/INPA, 1996. Tese Doutorado.
CONCEIÇÃO, P.N. da Os habitats da área de influência do APM Manso. In: ALHO,
C.J.R. et al. (Ed) Fauna silvestre da região do Rio Manso, MT. Brasília:
IBAMA/Centrais Elétricas do Norte do Brasil, 2000. p.31-127.
DUELLMAN, W.E. Herpetofaunas in neotropical rainforests: comparative
composition, history, and resource use. In: GENTRY, A. (Ed.) Four neotropical
rainforests. New Haven: Yale University Press, 1990. p.455-505.
FISHER, W.A. Efeitos da BR-262 na mortalidade de vertebrados silvestres:
síntese naturalística para a conservação da região do Pantanal-MS. Campo
Grande,MS; UFMS, 1997. 44p. Dissertação Mestrado.
FRANCO, M. do S.M.; PINHEIRO, R. Geomorfologia. In: BRASIL. Ministério das
Minas e Energias. Projeto RADAMBRASIL: folha SE. 21 Corumbá e SE 20. Rio
de Janeiro: Departamento Nacional de Produção Mineral, 1982. p.161-224.
(Levantamento de Recursos Naturais, 27).
HAMILTON, S.K. Potential effects of a major navigation project (Paraguay-Parana
Hidrovia) on inundation in the Pantanal floodplains. Regulated rivers: research
and management, n.15, p.289-299, 1999.
HAMILTON, S.K.; SIPPEL, S.J.; CALHEIROS, D.F.; MELACK, J.M. An anoxic
event and other biogeochemical effects of the Pantanal wetland on the Paraguay
river. Limnology Oceanography, v.42, n.2, p.257-272, 1997.
HAMILTON, S.K.; SIPPEL, S.J.; MELACK, J.M. Oxygen depletion and carbon
dioxide and methane production in waters of the Pantanal wetland of Brasil.
Biogeochemistry, n.30, p.115-141, 1995.

51
HAMILTON, S.K.; SIPPEL, S.J.; MELACK, J.M. Inundation patterns in the Pantanal
wetland of South America determined from passive-microwave remote sensing.
Archiv Hydrobiology, v.137, n.1, p.1-23, 1996.
HENDERSON, R.W.; MICUCCI, T.W.P.; PUORTO, G.; BOURGEOIS, R.W.
Ecological correlates and patterns in the distribution of neotropical boines
(Serpentes: Boidae) a preliminary assessment. Herpetological Natural History,
v.3, n.1, p.15-27, 1995.
HERNANDES FILHO, P.; PONZONI, F.J.; PEREIRA, M.N. Mapeamento da
fitofisionomia e do uso da terra de parte da bacia do Alto Taquari mediante o uso de
imagens TM/Landsat e HRV/Spot. Pesquisa Agropecuária Brasileira, v.33,
n.especial, p.1755-1762, 1998.
HUSZAR, P.; PETERMANN, P.; LEITE, A.; RESENDE, E.; SCHNACK, E.;
SCHNEIDER, E.; FRANCESCO, F.; RAST, G.; SCHNACK, J.; WASSON, J.;
GARCIA LOZANO, L.; DANTAS, M.; OBRDLIK, P.; PEDRONI, R. Fact or
fiction: a review of the hydrivua Paraguay-Parana official studies. Toronto: World
Wildlife Fund/World Wide Fund for Nature (WWF), 1999. 217p.
JUNK, W.J.; BAYLEY, P.B.; SPARKS, R.E. The flood pulse concept in river-
floodplain systems. Canadian Special Publication in Fisheries and Aquatic
Sciences, n.106, p.110-127, 1989.
LACHER JR, T.E.; ALHO, C.J.R. Microhabitat use among small mammals in the
Brazilian Pantanal. Journal of Mammalogy, v.70, n.2, p.396-401, 1989.
LUTZ, B.A. Notable frog chorus in Brazil. Copeia, v.1946, n.3, p.153-155, 1946.
NASCIMENTO, F.P.; ÁVILA-PIRES, T.C.S.; SANTOS, I.N.F.F.; LIMA, A.C.M.
Répteis da Marajó e Mexiana, Pará, Brasil. I. Revisão bibliografica e novos
registros. Boletim do Museu Paraense Emílio Goeldi, Ser. Zoologia, n.7, p.25-
41, 1991.

52
O’SHEA, M.T. The reptilian herpetofauna of the ilha de Maracá. In: MILLIKEN, W.;
RATTER, J.A. (Ed.) Maracá: the biodiversity and environment of an Amazonian
rainforest. Edinburgh: John Wiley e Sons, Royal Botanic Garden, 1998. p.231-
262.
RIVAS, J.A. The life history of the green anaconda(Eunectes murinus), with
emphasis on its reproductive biology. Knoxville: The University of Tennessee,
1999. 154p. Tese Doutorado.
SÁ, L.D.A.; SAMBATTI, S.B.M.; GALVÃO, G.P. Ondeleta de Morlet aplicada ao
estudo da variabilidade do nível do rio Paraguay em Ladário, MS. Pesquisa
Agropecuária Brasileira, v.33, n.especial, p.1775-1785, 1998.
SCHNEIDER, M. Mastofauna. In: ALHO, C.J.R. et al. (Eds) Fauna silvestre da
região do Rio Manso, MT. Brasília: IBAMA/Centrais Eletricas do Norte do
Brasil, 2000. p.217-238.
SCOTT, D.A.; CARBONELL, M. A directory of neotropical wetlands. Cambridge:
IUCN, 1986. 684p.
SEGNESTAM, L. Environmental performance indicators. Washington: The World
Bank, 1999. 38p. (Environment Department Papers, 71).
SICK, H. Migrações de aves na América do Sul Continental. Brasília:
IBDF/CEMAVE. (Publicação Tecnica, 2).
SILVA, J. dos S.V. da ; ABDON, M. de M. Delimitação do Pantanal brasileiro e suas
sub-regiões. Pesquisa Agropecária Brasileira, v.33, n.especial, p.1703-1711,
1998.
SILVA, J. dos S.V. da ; ABDON, M. de M.; SILVA, M.P.; ROMERO, H.R.
Levantamento do desmatamento no Pantanal brasileiro até 1990/91. Pesquisa
Agropecuária Brasileira, v.33, n.especial, p.1739-1745, 1998.

53
SILVA, J.M.C. Birds of the cerrado region, South America. Steenstrupia, n.21, p.69-
92, 1995.
STOTZ, D.F.; FITZPATRICK, J.W.; PARKER, T.A.; MOSKOVITS, D. Neotropical
birds: ecological and conservation. Chicago: University of Chicago Press, 1996.
478p.
STRUSSMANN, C. Herpetofauna. In: ALHO, C.J.R. et al. (Eds) Fauna silvestre da
região do Rio Manso, MT. Brasília: IBAMA/Centrais Eletricas do Norte do
Brasil, 2000. p.153-189.
STRUSSMANN, C. Serpentes do Pantanal de Poconé, Mato Grosso: composição
faunística, história natural e ecologia comparada. Campinas: INICAMP, 1992.
125p.
STRUSSMANN, C.; ALMEIDA PRADO, C.P. de; UETANABARO, M.; FERREIRA,
V.L. Amphibians and reptiles of selected localities in the southern Pantanal
floodplains and neighboring cerrado areas, Mato Grosso do Sul, Brasil. In:
WILLINK, P.W.; CHERNOFF, B.; ALONSO, L.E.; MONTAMBAULT, J.R. (Eds)
A biological assessment of the aquatic ecosystems of the Pantanal, Mato Grosso
do Sul, Brasil. Washington: Conservation International, 2000. p.98-102. (Bulletin
of Biological Assessment, 17).
STRUSSMANN, C.; SAZIMA, I. The snake assemblage of the Pantanal at Pocone,
Western Brazil: faunal composition and ecological summary. Studies on
Neotropical Fauna and Environment, v.28, n.3, p.157-168, 1993.
TOMÁS, W.M. Status and ecology of a Marsh Deer (Blastocerus dichotomus)
population in southern Pantanal, Brazil. Brasília: WWF, 1993.
TUBELIS, D.P.; TOMÁS, W.M. Caracterização da avifauna da planicie do
Pantanal. (no prelo).

54
VANZOLINI, P.E. Problemas faunísticos do cerrado. In: SIMPÓSIO SOBRE O
CERRADO, 1., 1971. São Paulo: Edgard Blucher/EDUSP, 1971. p.267-281.
VASCONCELLOS, L.A.S. Avifauna. In: ALHO, C.J.R. et al. (Eds.) Fauna silvestre
da região do Rio Manso,MT. Brasília: IBAMA/Centrais Eletricas do Norte do
Brasil, 2000. p.191-216.
VINSON, M.R.; HAWKINS, C.P. Biodiversity of stream insects: variation at local,
basin and regional scales. Annual Review of Entomology, n.43, p.271-293, 1998.
WANTZEN, K.M.; JUNK, W.J. The importance of stream-wetland-systems for
biodiversity: a tropical perspective. In: GOPAL, B.; JUNK, W.J.;DAVIS, J.A.
(Eds.) Biodiversity in wetlands: assessment, function and conservation.
Leiden: Backhuys Bublishers, 2000. v.1, p.11-34.
WEEB, S.D. Isolation and interchange: a deep history of South American Mammals.
In: EISENBERG, J.F.; REDFORD, K.H. (Eds.) Mammals of the neotropics: the
central neotropics. Chicago: University of Chicago Press, 1992. p.13-19.