influence of osmotic stress on heat shock proteins 25 and 72 in mouse mesangial cells
TRANSCRIPT

Influence of osmotic stress on heat shock proteins 25 and 72 inmouse mesangial cells
EVA MULLER, ANKE BURGER-KENTISCHER, WOLFGANG NEUHOFER, ANDREAS SCHOBER, and FRANZ-X. BECK
Department of Physiology, University of Munich, Munich, Germany
Influence of osmotic stress on heat shock proteins 25 and 72 inmouse mesangial cells. Previous studies have shown intensestaining for heat shock protein 25 (HSP25) in the extraglomerularmesangium (EGM). Because relationships are believed to existbetween osmotic stress, expression of HSP25, and protectionagainst stress and because the EGM may be exposed to high localtonicity, we examined the expression of HSP25 and the majorstress-inducible and cytoprotective HSP72 in mouse mesangialcells and embryonic lung fibroblasts (3T3) after exposure tohypertonic stress (addition of 150 mM NaCl to the medium for twoto seven days). Mesangial, but not 3T3, cells expressed high levelsof HSP25 already under control conditions, whereas neither cellline contained HSP72. Hypertonic treatment neither enhanced(mesangial cells) or induced (3T3 cells) HSP25 expression.HSP72, however, was induced strongly in 3T3 cells, but onlyminimally in mesangial cells. The high level of HSP25 in mesan-gial cells thus seems not to be a consequence of high tonicity in theEGM because cultured mesangial cells express HSP25 alreadyunder control conditions, and osmotic stress did not induceHSP25 in either cell line. Furthermore, high amounts of HSP25seem to reduce the requirement for HSP72 after stress exposure,suggesting that, in mesangial cells, HSP25 might assume somefunctions of HSP72.
High levels of heat shock protein 25 (HSP25) are detect-able in the extraglomerular mesangial (EGM) region of thejuxtaglomerular apparatus [1]. HSP25 expression is regu-lated physiologically by hormones, cytokines, state of dif-ferentiation, etc., but is also highly inducible by stressors(heat, arsenite, oxidants, etc.) [2] including osmotic stress[3]. Because the EGM region may be exposed to high localtonicity [4], this osmotic stress might be responsible for thehigh levels of HSP25 in this region. To test this hypothesis,we exposed a mesangial cell line [5] for varying periods tohypertonic stress and analyzed HSP25 expression. To de-fine mesangial cell-specific effects, cells of an embryoniclung fibroblast line (3T3) were subjected to the sameexperimental protocols.
Increased HSP25 contents protect cells against environ-mental stressors [6]. Another HSP, HSP72, is also cytopro-tective, but is detectable only after stress exposure [7]. Toexamine whether the requirement for HSP72 after stressexposure is influenced by high levels of HSP25, HSP72induction after stress exposure was also determined.
METHODS
Mouse glomerular mesangial cells (SV40 MES13 [5];ATCC CRL 1927) and embryonic lung fibroblasts (BALB/3T3: ATCC CCL-163) were grown in Dulbecco’s modifiedEagle’s medium supplemented with 2.5% and 10% fetalcalf serum, respectively, plus antibiotics (all Gibco, Paisly,UK). In hyperosmotic stress experiments, the medium wassupplemented with 150 mM NaCl; for heat stress experi-ments, cells were exposed to 41.5°C for five hours. Forprotein isolation, cells were lysed in a solution describedpreviously [1] by three cycles of thawing and freezing andfive minutes sonication. After centrifugation (10,000 g, 5min, 4°C), the supernatant was kept as the “cytoplasmicfraction.” Protein in the pellet was extracted by shaking (in6 M urea containing 0.1% Triton X-100, 30 min, 4°C),sonication (5 min), and centrifugation (16,000 g, 20 min,4°C). This supernatant represents the “pellet fraction.”Western blotting and immunodetection were performed asdescribed elsewhere [1] using the previously describedantibodies and antisera and an antiserum specific for bactin (A-2066; Sigma, Deisenhofen, Germany). b-Actinvalues were used to correct for variations in protein load-ing.
A synthetic oligonucleotide (30-mer) complementary tothe sequence 59-CC ATG GTG CTG ACC AAG ATGAAG GAG ATC G-39 was used as a selective probe forHSP72 mRNA [8] and was end labeled with digoxigeninusing a DIG Oligonucleotide Tailing Kit (BoehringerMannheim, Mannheim, Germany). Specific probes forHSP25 and b-actin mRNAs were synthesized by reversetranscriptase-polymerase chain reaction (RT-PCR). TotalRNA from heat-shocked mesangial cells was reverse tran-scribed with “Ready To Go T-Primed First Strand” Kit(Pharmacia, Freiburg, Germany), and PCR was carried out
Key words: extraglomerular mesangium, embryonic lung fibroblasts, jux-taglomerular apparatus, b-actin, HSP25 and HSP72, cytoprotection, cellstressors.
© 1998 by the International Society of Nephrology
Kidney International, Vol. 54, Suppl. 67 (1998), pp. S-162–S-164
S-162
on 2 ml of the RT reaction using a nucleotide solution with0.35 mM digoxigenin-dUTP (Boehringer Mannheim). A385-bp HSP25 fragment (nucleotides 8 to 392) was ampli-fied by PCR (94°C denaturation, 61°C annealing, 72°Cextension, 25 cycles, MWG Taq polymerase; MWG, Eber-sberg, Germany) with specific primers (sense, 59-AGCGCC GCG TGC CCT TCT C-39; antisense, 59-TGC CTTTCT TCG TGC TTG CCA GTG-39) and a 350-bp b-actinfragment (nucleotides 384 to 734; PCR, 94°C denaturation,56°C annealing, 72°C extension, 35 cycles) with specificprimers (sense, 59-AAC CGC GAG AAG ATG ACC CAGATC ATG TTT-39; antisense, 59-AGC AGC CGT GGCCAT CTC TTG CTC GAA GTC-39). RNA was isolatedusing TriReagent (Biozol, Eching, Germany). Northernanalysis, hybridization (56°C), and detection were carriedout using standard methods [9]. b-Actin values were usedto correct for variations in RNA loading.
RESULTS AND DISCUSSION
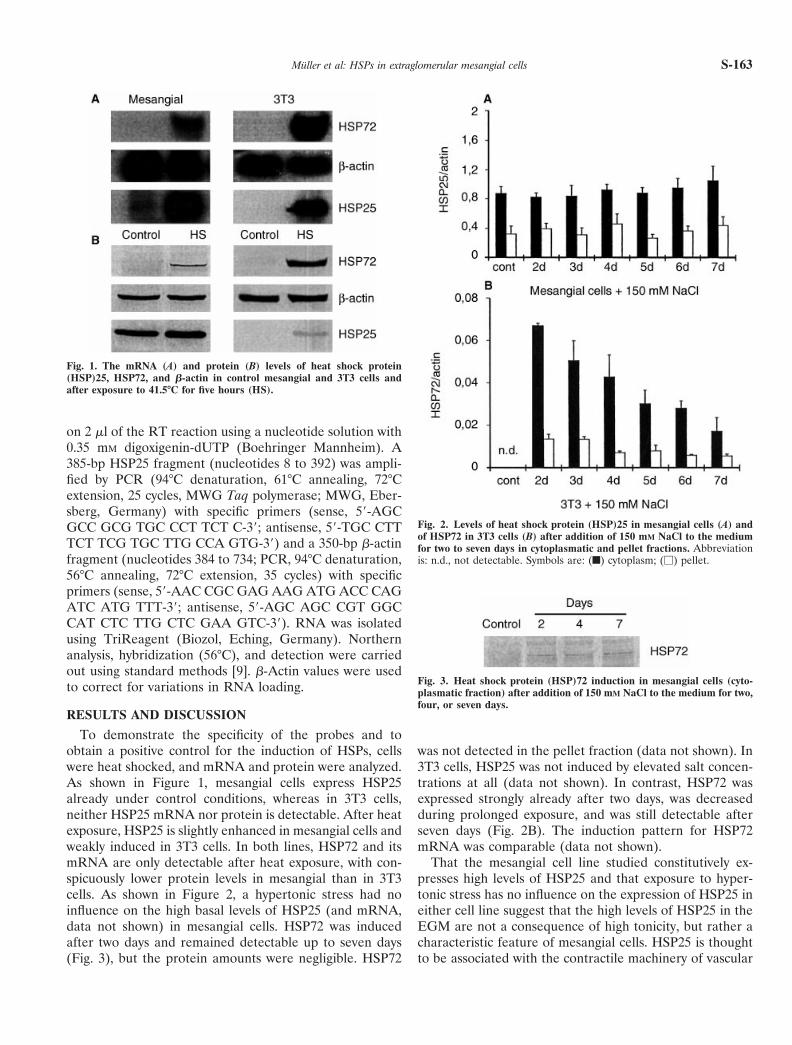
To demonstrate the specificity of the probes and toobtain a positive control for the induction of HSPs, cellswere heat shocked, and mRNA and protein were analyzed.As shown in Figure 1, mesangial cells express HSP25already under control conditions, whereas in 3T3 cells,neither HSP25 mRNA nor protein is detectable. After heatexposure, HSP25 is slightly enhanced in mesangial cells andweakly induced in 3T3 cells. In both lines, HSP72 and itsmRNA are only detectable after heat exposure, with con-spicuously lower protein levels in mesangial than in 3T3cells. As shown in Figure 2, a hypertonic stress had noinfluence on the high basal levels of HSP25 (and mRNA,data not shown) in mesangial cells. HSP72 was inducedafter two days and remained detectable up to seven days(Fig. 3), but the protein amounts were negligible. HSP72
was not detected in the pellet fraction (data not shown). In3T3 cells, HSP25 was not induced by elevated salt concen-trations at all (data not shown). In contrast, HSP72 wasexpressed strongly already after two days, was decreasedduring prolonged exposure, and was still detectable afterseven days (Fig. 2B). The induction pattern for HSP72mRNA was comparable (data not shown).
That the mesangial cell line studied constitutively ex-presses high levels of HSP25 and that exposure to hyper-tonic stress has no influence on the expression of HSP25 ineither cell line suggest that the high levels of HSP25 in theEGM are not a consequence of high tonicity, but rather acharacteristic feature of mesangial cells. HSP25 is thoughtto be associated with the contractile machinery of vascular
Fig. 1. The mRNA (A) and protein (B) levels of heat shock protein(HSP)25, HSP72, and b-actin in control mesangial and 3T3 cells andafter exposure to 41.5°C for five hours (HS).
Fig. 2. Levels of heat shock protein (HSP)25 in mesangial cells (A) andof HSP72 in 3T3 cells (B) after addition of 150 mM NaCl to the mediumfor two to seven days in cytoplasmatic and pellet fractions. Abbreviationis: n.d., not detectable. Symbols are: (f) cytoplasm; (M) pellet.
Fig. 3. Heat shock protein (HSP)72 induction in mesangial cells (cyto-plasmatic fraction) after addition of 150 mM NaCl to the medium for two,four, or seven days.
Muller et al: HSPs in extraglomerular mesangial cells S-163

smooth muscle cells [10]. EGM cells contain also highamounts of microfilaments, exhibit a structure similar toarteriolar smooth muscle cells [11], and are also contractilein vitro [5]. Thus, high HSP25 contents may result fromassociation of this protein with the contractile apparatus ofEGM cells.
HSP25 is known to be cytoprotective when present inhigh amounts. Should HSP25 exert this function also inmesangial cells, one might expect that the stress-inducedexpression of the major protective HSP, HSP72, would bereduced. Although the induction of HSP72 in mesangialcells was very weak after both heat and hypertonic stress, itwas enhanced greatly in 3T3 cells. Thus, in mesangial cells,the high basal levels of HSP25 seem to reduce the require-ment for HSP72 after stress exposure, suggesting thatHSP25 may assume protective functions in mesangial cells,which are usually exerted by HSP72.
ACKNOWLEDGMENTS
This study was supported by the Deutsche Forschungsgemeinschaft (Be963/4-4).
Reprint requests to Eva Muller, Ph.D., Physiologisches Institut der Univer-sitat Munchen, Pettenkoferstrabe 12, D-80336 Munchen, Germany.E-mail: [email protected]
REFERENCES1. MULLER E, NEUHOFER W, OHNO A, RUCKER S, THURAU K, BECK FX:
Heat shock proteins HSP25, HSP60, HSP72, HSP73 in isoosmotic
cortex and hyperosmotic medulla of rat kidney. Pflugers Arch 431:608–617, 1996
2. CIOCCA DR, OESTERREICH S, CHAMNESS GC, MCGUIRE WL, FUQUA
SA: Biological and clinical implications of heat shock protein 27,000(Hsp27): A review. J Natl Cancer Inst 85:1558–1570, 1993
3. KATO K, BRIJLALL D, ADLER SA, KATO S, HERZ F: Effect ofhyperosmolality on alkaline phosphatase and stress-response protein27 of MCF-7 breast cancer cells. Breast Cancer Res Treat 23:241–249,1992
4. PERSSON B, SAKAI T, MARSH DJ: Juxtaglomerular interstitial hyper-tonicity in Amphiuma: Tubular origin-TGF signal. Am J Physiol254:F445–F449, 1988
5. MACKAY K, STRIKER LJ, ELLIOT S, PINKERT CA, BRINSTER RL,STRIKER GE: Glomerular epithelial, mesangial, and endothelial celllines from transgenic mice. Kidney Int 33:677–684, 1988
6. LAVOIE JN, GINGRAS-BRENTON G, TANGUAY RM, LANDRY J: Induc-tion of Chinese hamster HSP27 gene expression in mouse cells confersresistance to heat shock. J Biol Chem 268:3420–3429, 1993
7. BUREL C, MEZGER V, PINTO M, RALLU M, TRIGON S, MORANGE M:Mammalian heat shock protein families. Expression and functions.Experientia 48:629–634, 1992
8. NOWAK TSJ, BOND U, SCHLESINGER MJ: Heat shock RNA levels inbrain and other tissues after hyperthermia and transient ischemia.J Neurochem 54:451–458, 1990
9. ENGLER-BLUM G, MEIER M, FRANK J, MULLER GA: Reduction ofbackground problems in nonradioaktive northern and southern blotanalyses enables higher sensitivity than 32P-based hybridizations. AnalBiochem 210:235–244, 1993
10. BITAR KN, KAMINSKI MS, HAILAT N, CEASE KB, STRAHLER JR:HSP27 is a mediator of sustained smooth muscle contraction inresponse to bombesin. Biochem Biophys Res Commun 181:1192–1200,1991
11. KRIZ W, ELGER M, LEMLEY K, SAKAI T: Structure of the glomerularmesangium: A biomechanical interpretation. Kidney Int 38:S2–S9,1990
Muller et al: HSPs in extraglomerular mesangial cellsS-164