inhibitors of crayfish glutamic acid decarboxylase - north carolina
TRANSCRIPT

Neurochemical Research, Vol. 9, No. 7, 1984
INHIBITORS OF CRAYFISH GLUTAMIC ACID DECARBOXYLASE
R O B E R T M . G R O S S F E L D 1, STEVEN W . Y A N C E Y 1, AND
C L A U D E F . BAXTER 2 1 Department of Zoology
North Carolina State University Raleigh, North Carolina 27650
and
2 V.A. Medical Center 151B2, 16111 Plummer St.
Sepulveda, California 91343
Accepted March 20, 1984
Crayfish glutamic acid decarboxylase (GAD), like the homologous enzymes from other species, is inhibited by carbonyl-trapping agents (e.g. aminooxyacetic acid; AOAA) and sulfhydryl reagents (e.g. 5,5'-dithiobis-(2-nitrobenzoic acid); DTNB). It also is inhibited by the product GABA, many anions (e.g. SCN- and C1- ), and some cations (e.g. Zn+2). The inhibition by AOAA, but not that by DTNB, was prevented by increasing the concentration of the pyridoxal phosphate (PLP) coen- zyme. GABA blocked the effects of PLP on enzyme activity. The inhibition by AOAA, DTNB, GABA, and chloride all were competitive with substrate. The effect of GABA occurs at physiological concentrations and may contribute to the regulation of GAD activity in vivo. The quantitative effect of anions is dependent on the cation with which they are administered. ATP stimulated GAD activity in homogenates prepared with potassium phosphate or Tris-acetate buffer, even when no exogenous PLP was provided.
INTRODUCTION
The glutamic acid decarboxylase enzymes (GAD; L-glutamic acid l-car- boxy-lyase, EC 4.1.1.15) of crayfish (10) and lobster (21) CNS tissues are
Address reprint request to: Robert M. Grossfeld, Department of Zoology, North Carolina State University, Campus Box 7617, Raleigh, North Carolina 27695-7617.
947 0364-3190/84/0700-0947503.50/0 �9 1984 Plenum Publishing Corporation

948 GROSSFELD ET AL.
stimulated in vitro by pyridoxal phosphate (PLP), 2-mercaptoethanol, po- tassium phosphate, and alkaline pH. The biochemical similarity of these enzymes suggests that the compounds which inhibit lobster GAD also might be inhibitors of crayfish GAD. This study was performed to de- termine whether the activity of crayfish GAD is altered by the same classes of compounds that affect the activity of GAD from other species and to determine the probable modes of action of some of the most ef- fective inhibitory substances. We expect that some of the compounds identified as inhibitors of crayfish GAD, or analogs of them, will prove to be useful biochemical or pharmacological probes of the enzyme's mech- anism of action or its function in vivo.
EXPERIMENTAL PROCEDURE
Homogenates were prepared from the abdominal segment of the ventral nerve cord (VNC) of the crayfish, Procambarus clarkii (10). The 20 ~xl of the standard reaction mixture con- tained 0.2 ram pyridoxal phosphate (PLP), 25 mM 2-mercaptoethanol, 0.13% (v/v) Triton X-100, 18 mM L-glutamic acid, about 0.5 IxCi of purified H3-glutamate, 100 mM potassium phosphate buffer, pH 7.2, and 5 txl tissue homogenate containing about 30 txg of protein (tissue samples). For each variation in the composition of the reaction mixture, correspond- ing blanks also were prepared without homogenate to permit correction of the tissue sample data for radioactivity not produced by GAD. The reaction tubes were incubated in the clark at 32~ for 8-10 hr to give the small tissue samples adequate time to generate sufficient [3H]GABA to be measured accurately. Under these conditions, [14C]GABA and C~402 are formed in equimolar amounts by lobster and crayfish GAD from a [U-J4C]glutamic acid substrate; moreover, GABA is not metabolized and GAD activity is linear with time (10, 21). The activity of the crayfish enzyme remains linear for 9.5 hr and the product ratio is unchanged when the reaction mixture contains sufficient carbonyl trapping reagent, sulfhy- dryl reagent, non-radioactive GABA, or anion to reduce the enzyme activity by 50%. After the enzyme was inactivated by addition of 0.5 ml 5% (w/v) trichloroacetic acid, the samples were applied to 0.6 • 4 cm beds of AG 50 W-X4 (100• mesh), Na form resin. Radioactive GABA was recovered from the columns with 3 ml 0.2 M sodium acetate, pH 5.5 after the radioactive glutamate had been removed with 7 ml 0.2 M sodium citrate, pH 3.5 (10, 17).
The mode of inhibition by a compound and the apparent Ki value, under variable substrate conditions, were determined by analysis of Lineweaver-Burk (16) graphs of the data. The 0.1 mM glutamate contributed to the reaction mixture by the tissue homogenate was neg- ligible in comparison with the exogenous glutamate added. The mode of inhibition of a compound under variable cofactor conditions was not assessed in the same way because of the complex effect of endogenous and exogenous PLP on enzyme activity (10); rather, the probable effect of the compound was deduced from the change in inhibition of enzyme activity as the PLP concentration was varied.
In each experiment, the enzyme activity is expressed as the total cpm of [H3]GABA formed by samples incubated for the same time with a constant quantity of tissue protein. Each figure illustrates the data from a single representative experiment. Duplicate analyses of a tissue sample usually differed by no more than 5% and the quantitative effect of an inhibitor typically varied by no more than 10-15% between experiments.
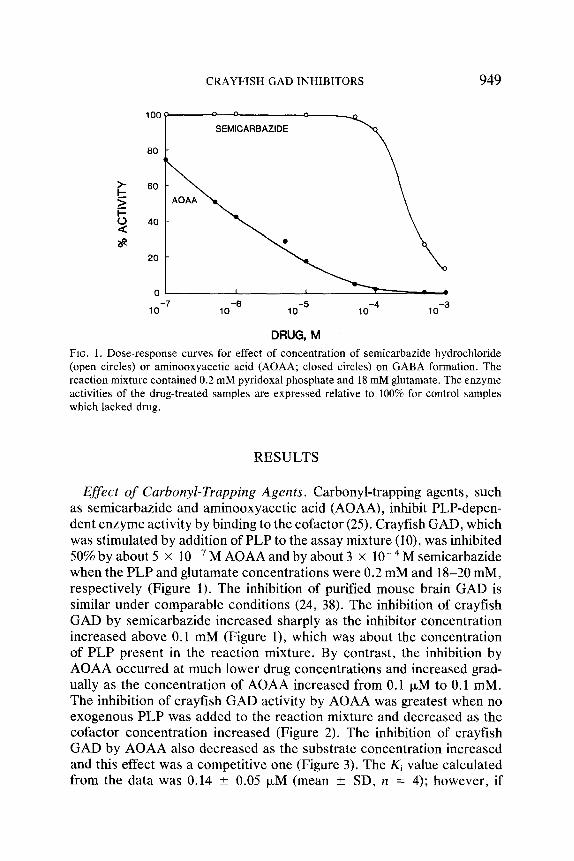
CRAYFISH GAD INHIBITORS 949
>- I--
I-- 0 .<
z~
100
80
60
40
20
0
10-
E ARBAZIDE
l I ~ - �9
- 6 - 5 10 10 10 -4 10 -3
DRUG, M FIG. 1. Dose-response curves for effect of concentration of semicarbazide hydrochloride (open circles) or aminooxyacetic acid (AOAA; closed circles) on GABA formation. The reaction mixture contained 0.2 mM pyridoxal phosphate and 18 mM glutamate. The enzyme activities of the drug-treated samples are expressed relative to 100% for control samples which lacked drug.
RESULTS
Effect of Carbonyl-Trapping Agents. Carbonyl-trapping agents, such as semicarbazide and aminooxyacetic acid (AOAA), inhibit PLP-depen- dent enzyme activity by binding to the cofactor (25). Crayfish GAD, which was stimulated by addition of PLP to the assay mixture (10), was inhibited 50% by about 5 • 10 - 7 M AOAA andby about 3 x 10 - 4 M semicarbazide when the PLP and glutamate concentrations were 0.2 mM and 18-20 mM, respectively (Figure 1). The inhibition of purified mouse brain GAD is similar under comparable conditions (24, 38). The inhibition of crayfish GAD by semicarbazide increased sharply as the inhibitor concentration increased above 0.1 mM (Figure 1), which was about the concentration of PLP present in the reaction mixture. By contrast, the inhibition by AOAA occurred at much lower drug concentrations and increased grad- ually as the concentration of AOAA increased from 0.1 IxM to 0.1 mM. The inhibition of crayfish GAD activity by AOAA was greatest when no exogenous PLP was added to the reaction mixture and decreased as the cofactor concentration increased (Figure 2). The inhibition of crayfish GAD by AOAA also decreased as the substrate concentration increased and this effect was a competitive one (Figure 3). The Ki value calculated from the data was 0.14 _+ 0.05 ~xM (mean _+ SD, n = 4); however, if

950 GROSSFELD ET AL.
10
8
o o o 6
X
:g 4 Q. 0
O 1X10-TM A O A A o ~ + ~ ~ ____-A ~
�9 I 1 ~ = L A
0 10 -7 10 -6 10 -5 10 -4 10 -3
PYRIDOXAL PHOSPHATE, M
FIG. 2. The effect of concentration of exogenous pyridoxal phosphate (PLP) on the inhibition of GABA formation by aminooxyacetic acid (AOAA). The concentration of exogenous PLP was varied from 0 to 1 mM whereas the concentration of glutamate was constant at 18 raM. The samples contained 1 x 10- 7 M AOAA (open circles) or no added AOAA (closed circles).
some AOAA was rendered inactive by binding to excess free PLP in the reaction mixture, then this value would be an over-estimate of the ap- parent Ki.
Effect of Sulfhydryl Reagents. Sulfhydryl reagents, such as DTNB (5,5'- dithiobis-(2-nitrobenzoic acid)) and PCMS (p-chloromercuriphenyl sul- fonate), alter the activity of some proteins by binding to one or more free sulfhydryl groups essential for the protein's binding or biological functions (5, 36). Crayfish GAD, which is stimulated by addition of 0.5-50 mM 2- mercaptoethanol to the reaction mixture (10), was inhibited 50% by about 0.6 mM DTNB or PCMS when the added glutamate and 2-mercaptoeth- anol concentrations both were 10 mM (Figure 4). The inhibition by 0.25- 5 mM DTNB decreased as the substrate concentration increased and this effect was a competitive one (the data at 0.5 and I mM DTNB are illus- trated in Figure 5). By contrast, the inhibition by DTNB was not signif- icantly dependent on the concentration of cofactor for cofactor concen- trations up to 10 mM (Figure 6). Both in the presence and absence of DTNB, the addition of low levels (0.2 mM) of PLP to the reaction mixture stimulated GAD activity whereas higher levels (10 mM) inhibited enzyme activity. This dual response to PLP probably results from the preferential binding of PLP at low concentrations to a lysine residue at the enzyme's active site and the additional binding of PLP at high concentrations to a lysine residue at a secondary site in the enzyme (10).

CRAYFISH GAD INHIBITORS 951
10
O X 8
L~
AOAA
pM AOAA
AOAA
0 1 2 3 4
l l G L U T A M A T E , mM X 0.1
FIG. 3. The effect of glutamate concentration on the inhibition of GABA formation by ami- nooxyacetic acid (AOAA). The samples contained 3 x 10 -6 M AOAA (open squares), 0.4 • 10 -6 M AOAA (closed squares), 0.1 x 10 -6 M AOAA (open circles) or no added AOAA (closed circles). The glutamate concentration was varied from 2.5 to 20 mM at constant specific radioactivity. The PLP concentration was constant at 0.2 mM.
Effect of GABA. Many enzymes are inhibited by the product of their catalytic function, a response which may serve to regulate the physio- logical levels of the product. Crayfish GAD was inhibited by addition of non-radioactive GABA to the reaction mixture (Figure 7). Activity was reduced 50% by 10-15 mM GABA when the glutamate concentration was 25 mM and by 1-5 mM GABA when the glutamate concentration was 2.5-10 mM. Only 0.05 mM GABA was contributed to the reaction mixture by the tissue homogenate. The inhibition by 1-50 mM GABA was com- petitive with the substrate glutamate (Figure 8 illustrates the data obtained with 5 mM GABA). The apparent Ki value was estimated as 1.3 + 0.2 mM (mean _+ SD, n = 4) .
The addition of 10 mM PLP to the reaction mixture did not relieve the product inhibition. However, in the presence of GABA, the effects of PLP on enzyme activity were altered (Figure 9). When GABA was present
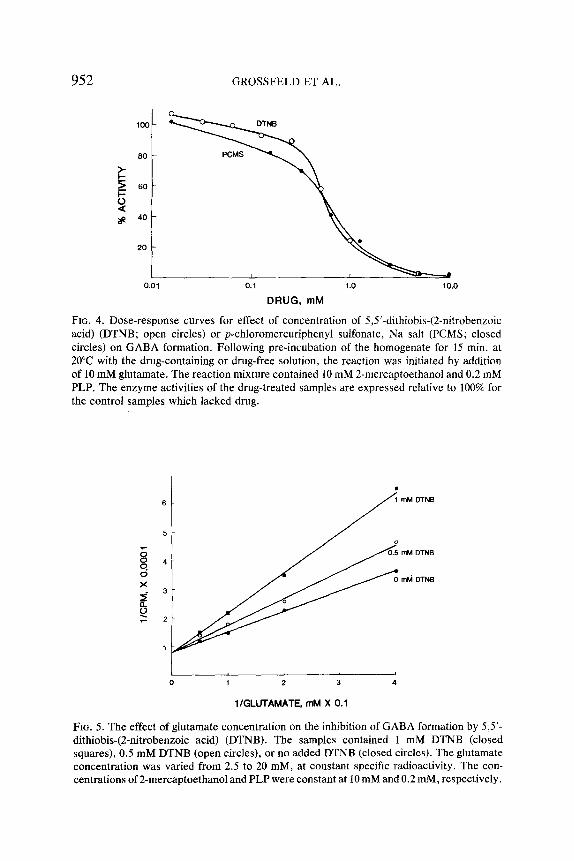
952 GROSSFELD ET AL.
80 >-
> 60 I'- o ,<
40
2o
0.01 0.1 1.0 10.0
DRUG, mM
FIG. 4. Dose-response curves for effect of concentration of 5,5'-dithiobis-(2-nitrobenzoic acid) (DTNB; open circles) or p-chloromercuriphenyl sulfonate, Na salt (PCMS; closed circles) on GABA formation. Following pre-incubation of the homogenate for 15 min. at 20~ with the drug-containing or drug-free solution, the reaction was initiated by addition of 10 mM glutamate. The reaction mixture contained 10 mM 2-mercaptoethanol and 0.2 mM PLP. The enzyme activities of the drug-treated samples are expressed relative to 100% for the control samples which lacked drug.
0 o. 0 X
I1.
d 6 J l mM DTNB
J o 5
0.5 rnM DTNB
mM OTNB 3
2
1
i i , i
1 2 3 4
1/GLUTAMATE, mM X 0.1
FIG. 5. The effect of glutamate concentration on the inhibition of GABA formation by 5,5'- dithiobis-(2-nitrobenzoic acid) (DTNB). The samples contained 1 mM DTNB (closed squares), 0.5 mM DTNB (open circles), or no added DTNB (closed circles). The glutamate concentration was varied from 2.5 to 20 mM, at constant specific radioactivity. The con- centrations of 2-mercaptoethanol and PLP were constant at 10 mM and 0.2 raM, respectively.

CRAYFISH GAD INHIBITORS 953
0 0 0
X
0- 0
i• 0.775 mM DTNB
0 2 4 6 8
PYRIDOXAL PHOSPHATE, mM
i
1 0
FIG. 6. The effect of concentration of exogenous pyridoxal phosphate (PLP) on the inhibition of GABA formation by 5,5'-dithiobis-(2-nitrobenzoic acid) (DTNB). The concentration of exogenous PLP in the reaction mixture was 0, 0.2, 2, or 10 mM whereas the glutamate and mercaptoethanol concentrations were constant at 10 mM. The samples contained 0.775 mM DTNB (open circles) or no added DTNB (closed circles).
at 2.5, 10, 15 or 25 mM, there was little or no stimulatory effect of low concentrations (<0.2 mM) of PLP and little or no inhibitory effect of high concentrations (>1 mM) of PLP.
Effect of Ions. The activity of GAD from many species is affected by the cation and anion content of the assay medium. Crayfish GAD was particularly sensitive to inhibition by Zn +2 and also was inhibited by 10 mM Mn +2 (Table I). At 10 mM, the chloride salts of Li +1, Na +1, K +l Mg +z, Ca +2, and B a +2 w e r e ineffective inhibitors. The chloride salts of Cu +2 and Co +2 were not tested because they formed a precipitate with a component, probably the thiol compound, of the reaction mixture. Cray- fish GAD also was inhibited by the potassium salts of all monovalent anions tested at 200 mM (Table II). The order of inhibition was SCN- > I - > NO3- > Br- > CI- > acetate- > F - , the same order of potency as has been reported for mouse brain GAD (31, 38). By contrast, when the crayfish nerve homogenates were incubated with the sodium salts of the same anions at 200 raM, NaF and Na acetate stimulated enzyme activity while NaC1 had no effect (Table II). Thus, the cation affected the overall response to an anion. Note that the relative order of enzyme ac- tivity was the same with 200 mM K and Na salts of the anions but that the samples containing 200 mM Na salts had about twice as much enzyme activity as the samples containing 200 mM of the corresponding K salts.

954 GROSSFELD ET AL.
100q
80
40
20
M GLU
mM GLU
I I I | I
10 20 30 40 50
GABA, rnM FIG. 7. Dose-response curves for effect of concentration of exogenous GABA on GABA formation. The reaction mixture contained 0, 1, 10, or 50 mM non-radioactive GABA and 2.5 mM glutamate (open circles) or 25 mM glutamate (closed circles). The enzyme activities (total [H3]GABA formed) of the GABA-containing samples are expressed relative to 100% for the control samples which lacked non-radioactive GABA.
6
5 GABA
S 4 O d X 3
2 0 4 0 6 0 8 0 1 0 0
1/GLUTAMATE, mM X 0.01
FIG. 8. The effect of glutamate concentration on the inhibition of GABA formation by ex- ogenous, non-radioactive GABA. T he samples contained 5 mM non-radioactive GABA (open circles) or no added GABA (closed circles). The glutamate concentration was varied from 1 to 25 raM, at constant specific radioactivity.

CRAYFISH GAD INHIBITORS 955
0 0 0
X
Q. O
NO GABA
15 mM GABA
~j . . . . . . .
0 2 4 6 8 10
PYRIDOXAL PHOSPHATE, mM
Fio. 9. The effect of concentration of exogenous pyridoxal phosphate (PLP) on the inhibition of GABA formation by exogenous, non-radioactive GABA. The concentration of exogenous PLP in the reaction mixture was 0, 0,2, 2, or 10 mM, whereas the glutamate concentration was constant at 25 raM, The samples contained 15 mM non-radioactive GABA (open circles) or no added GABA (closed circles).
This suggests that the primary effect of a K ~alt added to the reaction mixture at 200 mM was an anion-induced inhibition of enzyme activity whereas there was a dual response to a Na salt added at 200 raM, i.e. an anion-induced inhibition and a cation-induced stimulation of enzyme ac- tivity. This interpretation is consistent with the stimulation of crayfish
T A B L E I EFFECT OF CATiOnS On G A B A FORMATION
% of Control Activity
Salt Added 5 mM 10 rnM
None I00 100 LiC1 98 98 NaC1 105 105 KC1 101 99 MgCI2 104 96 CaC12 111 106 BaC12 114 111 ZnCI2 5 1 MnCI2 95 77
Effect of cations on GABA formation. The solution used for homogenizing the tissue had 5raM Tris-acetate buffer, pH 7.2 substituted for 100 mM potassium phosphate buffer, pH 7.2.

956 GROSSFELD ET AL.
TABLE II EFFECT OF ANIONS ON GABA FORMATION
% of Control Activity
K Salt Na Salt
Anion Added 50 mM 200 mM 50 mM 200 mM
None 100 100 100 100 $O4- 2 104 102 - - - - F - 1 98 68 127 130 CH3COO- 1 90 61 129 122 CI- 1 74 41 118 99 B r - 1 73 36 109 87 NO3 i 70 33 - - - - I - 1 66 29 103 69 SCN - l 43 18 79 37
Effect of anions on GABA formation. The solution used for homogenizing the tissue con- tained 100 mM potassium phosphate buffer, pH 7.2. All samples contained about 18 mM additional K added with the substrate. The sulfate salt was added at a maximum concen- tration of I00 mM to yield the same maximum cation concentration as the other samples.
GAD activity by addition of Na phosphate, but not K phosphate, to a reaction mixture containing 100 mM K phosphate buffer (10).
We examined the mode of action of Cl- on crayfish GAD since this ion competes with substrate in inhibiting mouse brain GAD (Susz et al., 1966) whereas its stimulatory effect on E. coli GAD results from an in- creased interaction between the coenzyme and apoenzyme and is non- competitive with substrate (8, 9, 22, 23, 27). The activity of the crayfish enzyme was reduced 50% by about 100 mM KCI when the glutamate concentration was 20 mM (Figure I0). The inhibition was competitive with the substrate (Figure 11). The apparent K~ value was estimated as 63 _+ 18 mM (mean _+ SD, n = 3) with KCI and 311 _+ 120 mM (mean + SD n = 3) with NaC1.
Effect of ATP. Mammalian brain GAD is inhibited by ATP and this effect is dependent upon the concentrations of PLP and Pi in the reaction mixture. By contrast, crayfish GAD was not inhibited by ATP. When the assay was performed with 100 mM potassium phosphate buffer containing either 0 or 0.2 mM exogenous PLP, GAD activity was increased slightly by 1-10 mM ATP (Table III,A,C). Part of this effect may result from the Na added with ATP (10). When the assay was performed with 5 mM Tris- acetate buffer, ATP had an even greater effect in increasing the basal enzyme activity (Table IIi,B,D) but note that the enzyme activity meas-
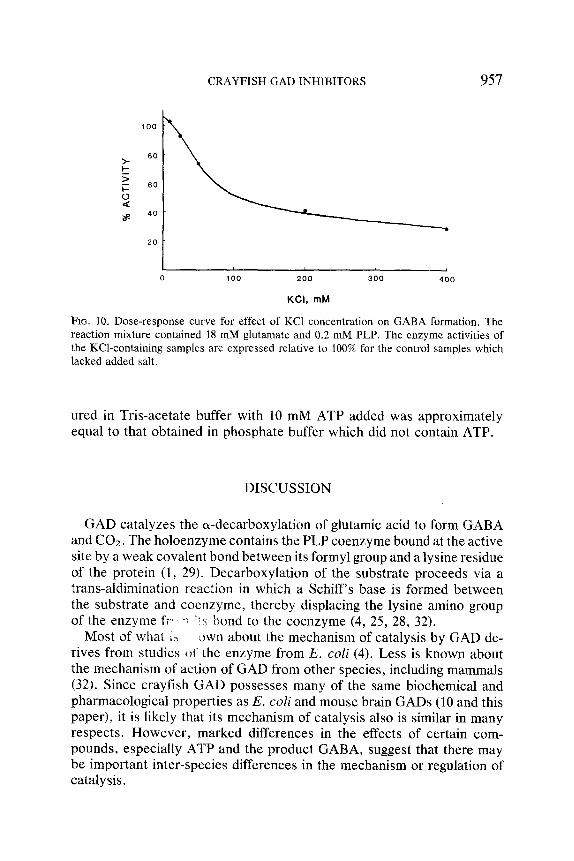
CRAYFISH GAD INHIBITORS 957
>.- I.-
C- o
100
80
60
40
2O
i i t i
0 100 2 0 0 3 0 0 4 0 0
KCI, mM
Fie. 10. Dose-response curve for effect of KC1 concentration on GABA formation. The reaction mixture contained 18 mM glutamate and 0.2 mM PLP. The enzyme activities of the KCl-containing samples are expressed relative to 100% for the control samples which lacked added salt.
ured in Tris-acetate buffer with 10 mM ATP added was approximately equal to that obtained in phosphate buffer which did not contain ATP.
DISCUSSION
GAD catalyzes the a-decarboxylation of glutamic acid to form GABA and COz. The holoenzyme contains the PLP coenzyme bound at the active site by a weak covalent bond between its formyl group and a lysine residue of the protein (1, 29). Decarboxylation of the substrate proceeds via a trans-aldimination reaction in which a Schiff's base is formed between the substrate and coenzyme, thereby displacing the lysine amino group of the enzyme fF ~-~ !~s bond to the coenzyme (4, 25, 28, 32).
Most of what ~ own about the mechanism of catalysis by GAD de- rives from studies of the enzyme from E. coli (4). Less is known about the mechanism of action of GAD from other species, including mammals (32). Since crayfish GAD possesses many of the same biochemical and pharmacological properties as E. coli and mouse brain GADs (10 and this paper), it is likely that its mechanism of catalysis also is similar in many respects. However, marked differences in the effects of certain com- pounds, especially ATP and the product GABA, suggest that there may be important inter-species differences in the mechanism or regulation of catalysis.

958 GROSSFELD ET AL.
400 mM KCI 32
28
24
w 2o
o d
200 mM KCI
O
12
8
~ mM KCI
"i NO KCI
1 2 3 4
1 / G L U T A M A T E , mM X 0 .1
FIc. 11. The effect of glutamate concentration on the inhibition of GABA formation by KCI. The samples contained 400 mM KC1 (open squares), 200 mM KC1 (closed squares), 50 mM KCI (open circles), or no added KC1 (closed circles). The glutamate concentration was varied from 2.5 to 20 raM, at constant specific radioactivity whereas the PLP concentration was constant at 0.2 raM.
Effect of Carbonyl-Trapping Agents. Carbonyl trapping agents such as AOAA can form a covalent derivative with the aldehyde carbon of PLP thereby blocking the binding of the substrate to the coenzyme in GAD. Carbonyl trapping agents which possess an acidic group, as does AOAA, are particularly effective inhibitors of mouse brain GAD, probably be- cause they also interact with an essential, positively charged residue of the enzyme protein (24, 25).
Crayfish GAD also was inhibited by AOAA and semicarbazide and the inhibition was prevented by increasing the concentration of coenzyme or substrate. The reduction of inhibition by PLP could mean that the pres- ence of excess coenzyme causes AOAA or the AOAA-PLP complex to dissociate from the holoenzyme, with the restitution of active enzyme, or that excess free PLP binds free AOAA to reduce the amount of drug bound to the enzyme (24, 25, 33), The difference in the shapes of the dose-response curves for the inhibition of crayfish GAD by semicarbazide

CRAYFISH GAD INHIBITORS 959
TABLE III EFFECT OF DI-SOD~UM ATP ON GABA FORMATION
Assay Conditions
GAD Activity, cpm (%)
lmM 5mM 10mM No ATP ATP ATP ATP
CA) Phosphate, 0.2mM PLP 4042 (100) 4059 (101) 4318 (108) 4549 (114) (B) Tris, 0.2raM PLP 1173 (100) 1676 (143) 2970 (253) 4288 (366) (C) Phosphate, No PLP 3120 (100) 3771 (121) 3628 (116) 3616 (116) (D) Tris, No PLP 962 (100) 1224 (127) 1782 (185) 2389 (248)
Effect of di-sodium ATP on GABA formation. A homogenate of CNS tissue was prepared, without exogenous PLP, either with 100 mM potassium phosphate buffer, pH 7.2 (A, C) or with 5 mM Tris-acetate buffer, pH 7.2 (B, D). Aliquots of each homogenate were assayed with no PLP added (C, D) or with 0.2 mM PLP added (A, B).
and A O A A suggests that, in the assay, AOAA interacts more effectively with enzyme-bound PLP than does semicarbazide at drug concentrat ions below 0.1 mM.
Effect of Sulfhydryl Reagents. A free sulfhydryl residue of bacterial and mammal ian brain GADs may part icipate in the binding of coenzyme or in its interact ion with substrate (6, 22, 25, 28, 38). The binding of DTNB to such an essential free sulfhydryl group located at or near the active site may physical ly obst ruct the access of the substrate to the catalytic site of mouse brain GAD (38). Alternatively, the binding of DTNB near or far f rom the active site could produce a change in the enzyme which prevents the interaction between the substrate and the coenzyme or a p o e n z y m e at the act ive site (12, 22, 25, 28, 36). Either situation could explain the compet i t ive kinetics obtained.
The inhibition of crayfish GAD by D T N B also was compet i t ive with the substrate , but it was virtually unaffected by changes in the coenzyme concentrat ion. An alternate interpretat ion of the latter data that we cannot rule out, however , is that PLP does counteract D T N B ' s pr imary inhibition of catalysis but that this effect of PLP is overshadowed by the secondary inhibition of enzyme activity produced when the PLP concentrat ion is increased f rom 0.2 to 10 mM. The inhibition of crayfish GAD by sulfhydryl reagents is not likely to be the result of chelation of an essential metal ion in the ho loenzyme since E D T A did not inhibit the enzyme at similar concentra t ions (10).
Effect of GABA. The GADs from lobster (21) and cockroach (2) are inhibited compet i t ive ly by physiological concentrat ions of GABA. This product inhibition of G A B A synthesis may regulate the intracellular

960 GROSSFELD ET AL.
GABA concentration (15). By contrast, the GADs from guinea pig brain (7) and bacteria (30) are not inhibited by even 200-1000 mM GABA.
The inhibition of crayfish GAD by GABA is competitive with glutamate. The failure of 10 mM PLP to reverse the inhibition by GABA suggests that GABA and the coenzyme form a relatively stable complex that is not readily displaced by excess coenzyme. Moreover, the lack of effect of exogenous PLP on enzyme activity in the presence of GABA suggests that the complex formed between free or bound PLP and GABA is more stable than that formed between PLP and lysine residues of the enzyme. We believe that the inhibition by GABA probably is of physiological rel- evance because the concentration at which it occurs in vitro probably is similar to the concentration to which the enzyme is exposed in situ.
Effect of Ions. The divalent cations Zn, Cu, Cd, and Hg are particularly potent inhibitors of mammalian brain GAD, while the monovalent cations K, Na, and Li are comparatively ineffective at equivalent concentrations (7, 38). The monovalent anion SCN- also is a potent inhibitor of mouse brain GAD, followed in order by I - , NO3-, SO4-2, Br - , CI- , acetate- , and F - (31, 38). By contrast, bovine heart GAD is stimulated by C1- (37) and E. coli GAD is stimulated by I - , S C N - , B r - , C1 , and SO4 2 (9, 22, 27). SO4 -2 has no effect on cockroach nerve GAD (2) but is a potent inhibitor of bovine brain GAD (11).
The inhibitory effects of simple ions on crayfish GAD were similar to their effects on mammalian brain GAD and were distinct from their effects on bacterial GAD. The inhibition of crayfish GAD by anions and cations, with the possible exception of zinc, probably occurs at too high an ion concentration to be physiologically important. The competitive inhibition by chloride may result from an interaction of this anion with a substrate- binding site on the holoenzyme or may result from a modification of the enzyme which makes it incapable of binding substrate. It.is possible that anions other than chloride (e.g. SCN- and I - ) and cations (e.g. Zn +2) also may affect the activity of crayfish GAD by interfering with the chem- ical interaction between substrate, coenzyme, and apoenzyme but their modes of action were not examined. For example, heavy metal ions may inhibit the decarboxylation of amino acids by chelation of the carboxyl group of the substrate (28). Also, heavy metal ions can form stable com- plexes with free sulfhydryl groups, thereby preventing participation by the sulfhydryl groups in substrate binding or catalysis (36).
Effect of ATP. If mammalian brain GAD is not completely saturated with PLP, then the binding of substrate may initiate a side reaction whose consequence is the production of inactive apoenzyme upon dissociation of the altered coenzyme (18, 19, 26). ATP and ADP can bind to a regu- latory site on the apoenzyme and thereby block its reactivation by PLP

CRAYFISH GAD INHIBITORS 961
(18-20, 26, 34, 35). This inhibition is time-dependent and is not compet- itive with substrate or coenzyme. By contrast, carrot GAD is not inhibited by ATP and the inhibition by ATP of E. coli GAD is less pronounced than for rat brain GAD and is independent of PLP concentration (35). Crayfish GAD also was not inhibited by ATP under similar conditions. In fact, ATP increased the activity of the crayfish enzyme. We surmise that the apparently greater stimulation of crayfish GAD activity by ATP in Tris-acetate buffer compared to that in phosphate buffer resulted from a restoration by the Na ATP of the enzyme activity lost reversibly when the tissue was homogenized in Tris-acetate buffer without PLP present.
CONCLUSION
Crayfish GAD is similiar to E. coli and mouse brain GADs in its re- sponse to several classes of potential inhibitory compounds but differs significantly from each in its response to others. Further studies will be necessary with a purified crayfish GAD to confirm the results obtained with the crude homogenate and to reveal the details of the chemical prop- erties of the enzyme.
ACKNOWLEDGMENT
We are grateful to Diana Gibson, Karen Lee, and Yvette Gray for their technical assist- ance, to Evan Jones for HPLC analysis of tissue glutamate and GABA contents, to Nancy Cochran for help in preparing the manuscript, and to Betty Black and Michael Kazarinoff for helpful comments. This work has been supported by grants to R.M.G. from the N.C. Board of Science and Technology (Project 2036) and the N.C.S.U. Agricultural Research Servide (Project NCO 5490) and to C.F.B. from the V.A. Medical Research Service. This is paper number 5490 of the Journal Series of the N.C. Agricultural Research Service, Raleigh, N.C. 27650.
REFERENCES
1. ANDERSON, J. A., and CHANG, H.-F. W. 1965. Borohydride reduction of L-glutamate decarboxylase. Arch. Biochem. Biophys. 110:346-349.
2. BAXTER, C. F., and TORRALBA, G. F. 1975. 3,-aminobutyric acid and glutamate decar- boxylase (L-glutamate 1-carboxy-lyase E.C. 4.1.1.15) in the nervous system of the cock- roach, Periplaneta americana. I. Regional distribution and properties of the enzyme. Brain Res. 84:383-397.
3. BLINDERMANN, J.-M., MAITRE, M., OSSOLA, L., and MANDEL, P. 1978. Purification and some properties of L-glutamate decarboxylase from human brain. Eur. J. Biochem. 86:143-152.

962 GROSSFELD ET AL.
4. BOEKER, E. A. and SNELL, E. E. 1972. Amino acid decarboxylases, Pages 217-253, in BOYER, P. D. (ed.), The Enzymes, Vol. VI, Academic Press, New York.
5. BOYER, P. D. 1959. Sulfhydryl and disulfide groups of enzymes, Pages 511-588, in BOYER, P. D., LARDY, H., and MYRBACK, K. (eds.), The Enzymes, Vol. 1, Academic Press, New York.
6. COZZANI, I., SANTONI, C., JORI, G., GENNARI, G., and MAMBURRO, A. M. 1974. Pyridoxal phosphate-sensitized photoinactivation of glutamate decarboxylase from Clostridium perfringens. Biochem. J. 141:463-468.
7. FISHER, S. K., and DAVIES, W. E. 1974. Some properties of guinea pig brain glutamate decarboxylase and its inhibition by the convulsant allylglycine (2-amino-4-pentenoic acid). J. Neurochem. 23:427-433.
8. FONDA, M. L. 1971. Interaction ofpyridoxal analogues with glutamate apodecarboxylase and aspartate apoaminotransferase. J. Biol. Chem. 246:2230-2240.
9. GERIC, J. T., and KwocK, L. 1973. Bromide ion probe N.M.R. studies of glutamate decarboxylase (E. coli). Biochem. Biophys. Res. Commnn. 54:937-941.
10. GROSSFELD, R. M., YANCEY, S. W., and BAXTER, C. F. 1984. Assay and properties of glutamic acid decarboxylase in homogenates of crayfish nervous tissue. Comp. Biochem. Physiol. In press.
l l . HADJIAN, R. A., and STEWART, J. A. 1977. Immunological quantitation of glutamic acid decarboxylase in developing mouse brain. J. Neurochem. 28:1249-1257.
12. JACOBSON, G. R., and STARK, G. R. 1973. Aspartate transcarbamylases, Pages 225-308, in BOYER, P. D. (ed.), The Enzymes, Vol. 9, Group Transfer Part B, Academic Press, New York.
13. KRAVITZ, E. A., KUFFLER, S. W., POTTER, D. D., and VAN GELDER, N. M. 1963. Gamma- aminobutyric acid and other blocking compounds in crustacea. II. Peripheral nervous system. J. Neurophysiol. 26:729-738.
14, KRAVITZ, E. A., and POTTER, D. D. 1965. A further study of the distribution of ~/- aminobutyric acid between excitatory and inhibitory axons of the lobster. J. Neurochem. 12:323-328.
15. KRAVITZ, E. A., MOLINOEF, P. B., and HALL, Z. W. 1965. A comparison of the enzymes and substrates of gamma-aminobutyric acid metabolism in lobster excitatory and in- hibitory axons. Proc. Nat. Acad. Sci. U.S.A. 54:778-782.
16. LINEWEAVER, H., and BURK, D. 1934. The determination of enzyme dissociation con- stants. J. Amer. Chem. Soc. 56:658-666.
17. MADERDRUT, J. L. 1979. A radiometric microassay for glutamic acid decarboxylase. Neurosci. 4:995-1005.
18. MARTIN, D. L., and MARTIN, S. B. 1982. Effect of nucleotides and other inhibitors on the inactivation of glutamate decarboxylase. J. Neurochem. 39:1001-1008.
19. MILLER, L. P., MARTIN, D. L., MAZUMDER, A., and WALTERS, J. R. 1978. Studies on the regulation of GABA synthesis: Substrate-promoted dissociation of pyridoxal-5'- phosphate from GAD. J. Neurochem. 30:361-369.
20. MILLER, L. P., and WALTERS, J. R. 1979. Effects of depolarization on cofactor regulation of glutamic acid decarboxylase in substantia nigra synaptosomes. J. Neurochem. 33:533-539.
21. MOLINOFF, P. B., and KRAVITZ, E. A. 1968. The metabolism of ~-aminobutyric acid (GABA) in the lobster nervous system-glutamic decarboxylase. J. Neurochem. 15:391- 409.
22". O'LEARY, M. H., and BRVMMOND, W. B., JR. 1974. pH jump studies of glutamate de- carboxylase. J. Biol. Chem. 249:3737-3745.
23. O'LEARY, M. H., and MALIK, J. M. 1972. Kinetics and mechanism of the binding of pyridoxal 5'-phosphate to apoglutamate decarboxylase. J. Biol. Chem. 247:7097-7105.

CRAYFISH GAD INHIBITORS 963
24. ROBERTS, E., and SIMONSEN, D. G. 1963. Some properties of L-glutamic decarboxylase in mouse brain. Biochem. Pharmacol. 12:113-134.
25. ROBERTS, E,, WEIN, J., and SIMONSEN, D. G. 1964. 7-aminobutyric acid (7-ABA), vi- tamin B6 and neuronal function-A speculative synthesis. Vit. and Horm. 22:505-559.
26. SELIGMANN, B., MILLER, L. P., BROCKMAN, D. E., and MARTIN, D. L. 1978. Studies on the regulation of GABA synthesis: The interaction of adenine nucleotides and glu- tamate with brain glutamate decarboxylase. J. Neurochem. 30:371-376.
27. SHUKUYA, R., and SCHWERT, G. W. 1960. Glutamic acid decarboxylase--I. Isolation procedures and properties of the enzyme. J. Biol. Chem. 235:1649-1652.
28. SNELL, E. E. 1958. Chemical structure in relation to biological activities of vitamin B6, Vit. and Horm. 16:77-125.
29. STRAUSBAUCH, P. H., and FISCHER, E. H. 1970. Structure of the binding site of pyridoxal 5'-phosphate to Escherichia coli glutamate decarboxylase. Biochem. (USA) 9:233-238.
30. SUKHAREVA, B. S., and MALIKOVA, L. G. 1977. Substrate specificity ofE. coli glutamate decarboxylase. Molec. Biol. (Russ.) 11:302-307.
31. Susz, J. P., HABER, B., and ROBERTS, E. 1966. Purification and some properties of mouse brain L-glutamic decarboxylase. Biochem. (USA) 5:2870-2876.
32. SZE, P. Y. 1979. L-glutamate decarboxylase, in Adv. Exp. Med. and Biol., Vol. 123, GABA-Biochemistry and CNS Functions, pp. 59-78. Plenum Press, New York.
33. TAPIA, R., and SANDOVAL, M. E. 1971. Study on the inhibition of brain glutamate de- carboxylase by pyridoxal phosphate oxime-O-acetic acid. J. Neurochem. 18:2051-2059.
34. TURSKY, T. 1970. Inhibition of brain glutamate decarboxylase by adenosine triphos- pbate. Eur. J. Biochem. 12:544-549.
35. TURSKY, T., and LASSANOVA, M. 1978. Inhibition ef different molecular forms of brain glutamic acid decarboxylase (GAD) with ATP. J. Neurochem. 30:903-905,
36. WEBS, J. L. 1966. Enzyme and Metabolic Inhibitors, Vol. 2, pp. 635-653. Academic Press, New York.
37. Wu, J.-Y. 1977. A comparative study of L-glutamate decarboxylase from mouse brain and bovine heart with purified preparations. J. Neurochem. 28:1359-1367.
38. Wu, J.-Y., and ROBERTS, E. 1974. Properties of brain L-glutamate decarboxylase: In- hibition studies. J. Neurochem. 23:759-767.