investigation of salmonella enterica in sardinian slaughter pigs: prevalence, serotype and genotype...
TRANSCRIPT

International Journal of Food Microbiology 151 (2011) 201–209
Contents lists available at SciVerse ScienceDirect
International Journal of Food Microbiology
j ourna l homepage: www.e lsev ie r .com/ locate / i j foodmicro
Investigation of Salmonella enterica in Sardinian slaughter pigs: Prevalence, serotypeand genotype characterization
Francesca Piras a,⁎, Derek J. Brown b, Domenico Meloni a, Anna Mureddu a, Rina Mazzette a
a Department of Animal Biology, Faculty of Veterinary Medicine, University of Sassari, Via Vienna 2, 07100, Sassari, Italyb Scottish Salmonella, Shigella & Clostridium difficile Reference Laboratory Stobhill Hospital 133, Balornock Road, Glasgow G21 3UW, Scotland, United Kingdom
⁎ Corresponding author. Tel.: +39 079 229 447; fax:E-mail address: [email protected] (F. Piras).
0168-1605/$ – see front matter © 2011 Elsevier B.V. Alldoi:10.1016/j.ijfoodmicro.2011.08.025
a b s t r a c t
a r t i c l e i n f oArticle history:Received 21 January 2011Received in revised form 19 August 2011Accepted 25 August 2011Available online 31 August 2011
Keywords:SalmonellaSlaughterhousePigPFGE
In order to improve the knowledge about the presence of Salmonella in porkmeat in Sardinia (Italy), the prevalenceand the sources of Salmonella at 5 pig slaughterhouses (slaughtered pigs and environment) were investigated andthe isolates were characterised. A total of 462 samples were collected, 425 from pigs at slaughter and 41 from theslaughterhouse environment. Salmonella was isolated from 26/85 (30.5%) mesenteric lymph nodes, 14/85 (16.4%)colon contents, and from 12/85 (14.1%) carcasses and livers. Salmonella prevalence was 38% (8/21) in samplesfrom surfaces not in contact with meat, and 35% (7/20) in those from surfaces in contact with meat. Thirty-onepigs were identified as carriers of Salmonella in lymph nodes and/or colon content, but of these, only 8 carcasseswere positive. A total of 103 Salmonella isolates were serotyped and genotyped. Eight different serotypes weredetected; themost commonwere S. Derby (44/103, 42.7%) and S. Typhimurium(24/103, 23.3%). Themost prevalentS. Typhimurium phage typewas DT193. Thirty-two isolates were found to be resistant tomore than one antimicro-bial (MDR). Pulse-field gel electrophoresis (PFGE)permitted the resolutionofXbaImacrorestriction fragments of theSalmonella strains into 20 distinct pulsotypes. Combined application of a plasmid profiling assay (PPA) and PFGEgave useful additional information to assist in tracing the routes of Salmonella contamination in abattoirs. To re-duce Salmonella prevalence some preventive measures should be encouraged: the origin of infected slaughteranimals should be identified and direct and cross-contamination of carcasses should be avoided by adhering toHACCP principles in association with good hygiene procedures (GHP).
+39 079 229 458.
rights reserved.
© 2011 Elsevier B.V. All rights reserved.
1. Introduction
Control of Salmonella infection in pigs forms an important part of theRegulation (EC) No. 2160/2003 of the European Parliament. Article 4 ofthis regulation states that Community targets shall be established forthe reduction of the prevalence of Salmonella in herds of slaughterpigs (Anonymous, 2003).
The importance of pigs as a source of salmonellosis has beenshown in a number of investigations (Swanenburg et al., 2001a,b,c;Lo Fo Wong et al., 2002; Botteldoorn et al., 2003; Zheng et al.,2007). Salmonella has been identified in all stages of pork production(Lo Fo Wong et al., 2004), and transmission may occur throughout theproduction chain, directly or indirectly from animal feed, at the farm,within the slaughterhouse or packing plant, inmanufacturing, processingand retailing of food, throughpreparation of food for human consumptionin commercial catering establishments or within the home (Lo Fo Wonget al., 2002). Many opportunities exist within the pig slaughterhouse forthe contamination of pork carcasses with Salmonella and other potential
pathogenic microorganisms, and the process does not contain any pointswhere hazards may be completely eliminated. The sources of contamina-tion during slaughter are environmental or from faecal and/or pharyngealcarriage by infected animals (Bonardi et al., 2003; van der Wolf et al.,2001; Zheng et al., 2007).
Infected pigs generally carry Salmonella asymptomatically in thetonsils, the intestines and the gut-associated lymphoid tissue (GALT)(Boyen et al., 2008). Salmonella is spread to the carcass surface mainlyfrom the carrier animal during the evisceration step and, therefore,good hygienic practises and equipment cleanliness in the abattoircan have a profound influence on the level of carcass contamination(Bonardi et al., 2003).
While numerous investigations have focused on the epidemiologyof salmonellosis in pigs, in-depth studies about the sources of infec-tion or contamination at slaughterhouse are needed (Magistrali etal., 2008).
Sardinia only produces 50% of its pork for consumption whichrepresents a very low self-sufficiency rate. As a consequence there is aneed to import pigmeat and pork products*. Therefore pigs slaughteredin Sardiniamay originate locally ormay have been imported from otherItalian regions and other EC countries. There is a lack of data about Sal-monella prevalence and epidemiology in Sardinia at farm level and inthe finished product.

202 F. Piras et al. / International Journal of Food Microbiology 151 (2011) 201–209
The aim of the present study was the evaluation of Salmonella preva-lence and serotypes in different pig slaughterhouses, by the detection ofSalmonella in animal and environmental samples. Phenotypic andmolec-ular methods were applied in order to characterise the isolates, and gen-otypingwas performed to define clonal relationships between Salmonellastrains, and to investigate their distribution at slaughterhouse level.
2. Materials and methods
2.1. Slaughterhouse description and sample plan
The study was carried out from June 2006 to August 2008 in fiveEC swine abattoirs (S1 to S5) in Sardinia, that slaughtered Sardinianpigs and pigs coming from other parts of Italy and from EU countries.
The slaughterhouses were comparable from the point of view ofthe slaughter techniques and procedures and for the number of pigsslaughtered per hour (approximately 300 carcasses per hour). Thescalding procedure was carried out in vats by the use of recirculatingwater and thewater temperaturewas 62–63 °C. Singeingwas performedat 1000 °C for 10–15 s, so that the carcass surface temperature reachedapproximately 100 °C. Dissection of the rectumwas performedmechan-ically and no plastic bags were used to seal off the rectum duringevisceration.
Experiments took place throughout an entire sampling day (SD)and samples were collected from slaughtered pigs as well as theslaughterhouse environment. The specimens were collected from 85randomly selected pigs (100–120 kg in weight) during 13 samplingvisits: S1, S2 and S5 were visited twice, S4 three times, S3 four times.
2.2. Sample collection
2.2.1. Pig samplesFrom each pig, after evisceration and before chilling, samples were
collected as follows:
• Mesenteric lymphnodes and colon contents: intestineswere collectedin plastic bags in a separate room near the slaughterline and at least5 lymph nodes in the ileocaecal regions were cut out with a sterile,disposable scalpel. The colon was incised and 25 g of its contents wascollected in a sterile plastic stomacher bag;
• Tonsils: Samples of the tonsils were aseptically collected and placedin sterile containers;
• Carcass surface: the carcass surfaces were sampled using sponges(10×10 cm) in the following way:- the upper inner part of both the hind legs including approximately5 cm of the skin and the pelvic entrance (approximately 30 cm×20–25 cm);
- the cut surface area of the abdomen and chest includingapproximately 5 cm of the skin surface; approximately 70–80 cm×8–10 cm was sampled.
A total area of approximately 1400 cm2 of the carcass surface wassampled. Two sterile sponges (10×10 cm) were used, one sponge forthe upper inner part of the hind legs and one for the surface area ofthe abdomen. The two sponges collected from each carcass werepooled prior to testing.
• Liver surface: the liver was sampled with a sterile sponge on bothsides and, with the same sponge, around all lobula. Sampling tookplace in the slaughter line immediately after the removal of thepluck.
2.2.2. Environmental samplesAt the end of the SD, before cleaning, environmental samples were
collected as follows:
• Scalding water: 25 ml was taken with a sterile collection tube andthe temperature was measured.
• Surfaces not in contact with meat:- drains: during pre-chilling, the drain surface was sampled with asterile sponge;
- dirty zone: during stunning-bleeding, a wall surface of 100 cm2
was sampled with a sterile sponge.• Surfaces in contact with meat were sampled with a sterile sponge asfollows:
- dehairing equipment: a surface of 1000 cm2;- knives: from the tip to the base, on both sides;- carcass splitter: on both sides of the blade.
Sponges were immediately put in sterile plastic sponge-bags, andthese were identified with a univocal code for each sample.
All the specimens were transported at +4 °C to the laboratory andanalysed on the same day.
In total 462 samples were collected, 425 from pigs at slaughterand 41 from the slaughterhouse environment.
2.3. Isolation, identification, serotyping, and phage typing of Salmonellaisolates
For the isolation of Salmonella the ISO method 6579/2002 modifiedaccording to EFSA report on “Risk assessment and mitigation opinionof Salmonella in pig production” was used.
Presumptive colonies were submitted to phenotypic identificationwith the API ID 32E system (bioMérieux, Marcy l'Etoile, France).
The isolates were sent to the laboratories of the “Centro Nazionaledi Referenza per le salmonellosi” in Legnaro (Padua, Italy), serotypedby agglutination tests with specific O and H antisera (Staten SerumInstitute, Copenhagen, Denmark) and classified according to theKauffmann–White scheme. Strains of serotypes Typhimurium andEnteritidis were phage typed according to the recommendations ofthe Health Protection Agency (London, UK).
2.4. Characterization of Salmonella isolates
Characterization of Salmonella isolates was carried out at the ScottishSalmonella, Shigella and Clostridium difficile Reference Laboratories(SSSCDRL) in Glasgow (Scotland, UK).
2.4.1. Antimicrobial susceptibility testing (AMR) and detection of class Iintegrons
Antimicrobial susceptibility tests were performed onMueller Hintonagar (Oxoid, Milan, Italy) by agar dilution tests following the NationalCommittee for Clinical Laboratory Standards guidelines (Clinical,Laboratory Standards Institute (CLSI/NCCLS), 2004). Susceptibilities(mg/L) to ampicillin (A, 50), chloramphenicol (C, 20), ciprofloxacin(Cp, 0.5), gentamicin (G, 20), nalidixic acid (Na, 40), streptomycin (S,20), tetracycline (T, 10), trimethoprim (Tm, 10), cefotaxime (Cx, 1), sul-phamethoxazole (Su, 100), cephalothin (Kf, 32), amoxicillin–clavulanicacid (Ax, 16/8), and colistin (Co, 25)were determined in all isolates. Iso-lates demonstrating intermediate resistancewere included in the sensi-tive category. Isolates that were resistant to two or more differentclasses of antimicrobials were defined as multiresistant. Template DNAwas obtained from isolates that showed antibiotic resistance: strainswere inoculated onto cysteine lactose electrolyte deficient (CLED,Oxoid, Milan, Italy) and incubated at 37 °C overnight. Three coloniesfrom each plate were inoculated into a 1.5 ml sterile Eppendorf tubewhich contained 100 μl of sterile water. The tubes were boiled in ablock heater at 105 °C for 12 min and then centrifuged at 13,000 rpmfor 10 min. 80 μl of the supernatant was transferred to a new sterileEppendorf tube and stored at−20 °C indefinitely.
The oligonucleotide primers (VH Bio Ltd, Gateshead, UK) and thecycling parameters, for antibiotic resistance genes and for class 1integrons are shown in Table 1. First, the variable regions of the integronsthat encode specific resistance genes, were amplified by using the 5′-CS

203F. Piras et al. / International Journal of Food Microbiology 151 (2011) 201–209
and the 3′-CS primers. Then, the class 1 integrase-specific Int-1 genewasdetected. Finally the presence of the resistance genes, as indicated inTable 1, was investigated.
All PCR amplifications were carried out by using puReTaq Ready-To-Go PCR beads (GE Healthcare, Little Chalfont, UK) which contained pre-dispensed Taq polymerase, nucleotide and buffer, in accordance to themanufacturer's instruction. PCR amplifications were performed in a25 μl volume, using aGeneAmp9700 Thermocycler (Applied Biosystems,Foster City, CA, USA). Reaction mixtures contained 2.5 μl of each primer(from stock concentration of 10 pmol/μl), 5 μl of template DNA and ster-ile water. PCR products obtained using Int1 primers were analysed on a1% agarose gel. PCR products obtained using CS primers were run on a1.6% agarose gel and the other PCR products in a 1.5% agarose gel.
2.4.2. Plasmid profiling and transfer assayPlasmid DNA was isolated by the alkaline lysis method described
by Kado and Liu (Kado and Liu, 1981). Plasmid containing strainE. coli 39R861 and a supercoiled DNA ladder (Sigma Aldrich, St.Louis, MI, USA) were used to estimate plasmid sizes.
Conjugation experiments were performed in order to determinewhether antimicrobial resistance markers were located on conjugativeplasmids, on n.3 isolates that showed resistance to the followingantibiotics: one strain of S. Derby and one of S. Typhimurium U302(ACSuTTmKf) and one S. Typhimurium unt strain (ACSuTKfAx).
Prior to the analysis both the donor and the recipient were inocu-lated onto CLED and incubated overnight at 37 °C to check purity andobtain a fresh colony.
The selection of transconjugants was carried out by mating thecandidate MDR donor strains with a spontaneous rifampicin resistant E.coli J53-2 strain. The isolates and the donor were grown in 10 ml of BHIbroth (Oxoid, Milan, Italy) in a plastic sterile tube and incubatedovernight at 37 °C. Thereafter, 200 μl of the donor was mixed with800 μl of the recipient, 4 ml of BHIwas addedand the cellswereharvestedby centrifugation at 3000 rpm and themixturewas incubated at 37 °C for4 h.
Transconjugants were selected on CLED agar plates containing onetest antibiotic such as rifampicin (at the same concentrations used for
Table 1Primer sequences and cycling parameters of PCR amplification for antibiotic resistance gen
Gene ofinterest
F or Ra 5′ to 3′DNA sequence of primers Cycling param
Denaturation
Int1 F TCTCGGGTAACATCAAGG 94 °C (45 s)R AGGAGATCCGAAGACCTC
CS F GGCATCCAAGCAGCAAGC 94 °C (45 s)R AAGCAGACTTGACCTGAT
aadA1 F TATCAGAGGTAGTTGGCGTCAT 94 °C (1 min)R GTTCCATAGCGTTAAGGTTTCATT
aadA2 F TGTTGGTTACTGTGGCCGTA 94 °C (1 min)R GATCTCGCCTTTCACAAAGC
bla (Tem) F CATTTCCGTGTCGCCCTTAT 94 °C (1 min)R TCCATAGTTGCCTGACTCCC
bla (Carb2) F GCTTCGCAACTATGACTAC 94 °C (45 s)R GTTCACCATCCAAGACTC
cat1 F CCTATAACCAGACCGTTCAG 94 °C (1 min)R TCACAGACGGCATGATGAAC
cat2 F CCGGATTGACCTGAATACCT 94 °C (1 min)R TCACATACTGCATGATGAAC
dhfr1 F GTGAAACTATCACTAATGGTAGCT 94 °C (1 min)R ACCCTTTTGCCAGATTTGGTAACT
sul1 F TCACCGAGGACTCCTTCTTC 94 °C (1 min)R AATATCGGGATAGAGCGCAG
tetA(A) F GGTTCACTCGAACGACGTCA 94 °C (1 min)R CTGTCCGACAAGTTGCATGA
tetA(B) F CCTCAGCTTCTCAACGCGTG 94 °C (1 min)R GCACCTTGCTCATGACTCTT
tetA(G) F CCGGTCTTATGGGTGCTCTA 94 °C (1 min)R CCAGAAGAACGAAGCCAGTC
a F, forward; R, reverse; 1 Five minutes for the first cycle; 2 Seven minutes for the last cy
the antimicrobial susceptibility tests). These plates were inoculatedwith 100 μl of the mating mixture using a sterile spreader.
Control plates, also containing one of the antibiotics to be tested andrifampicin were inoculated from cultures of the donor, recipient andmating mixture to ensure that the organisms were resistant to theappropriate antibiotic and that both donor and recipient were presentin the mating mixture. All the plates were incubated overnight at 37 °C.The following day potential transconjugant was purified and confirmedto be E. coli by growth onto CLED.
2.4.3. Pulsed field gel electrophoresis (PFGE)Salmonella isolates were inoculated onto CLED and incubated over-
night at 37 °C. Additionally, control strains of S. Braenderup (H9812),which served as a molecular weight standard, and S. Senftenberg(99114159 K), serving as an internal control, were processed with eachbatch of strains.
For chromosomal DNA preparation for subsequent PFGE, theSalmGene protocol by Peters et al. (2003) was used.
Briefly, DNA restrictionwas donewith 50U XbaI enzyme (Invitrogen,Carlsbad, CA, USA) at 37 °C for 4 h. PFGEwas performed with CHEF DRIIsystem (Bio-Rad). Electrophoresis conditions were 6 V/cm for 22 h.Pulse time was ramped from 2 to 64 s. The gel was stained in ethidiumbromide for 10 min (2 μg/ml) (Sigma Aldrich) and band patterns werevisualised under UV light and photographed using a GelDoc XR camerasystem (Bio-Rad, Hercules, CA, USA). Macrorestriction patterns werecompared with the use of Bionumerics software v5.0 (Applied Maths,Sint-Martens-Platen, Belgium). The molecular weights of the restrictionfragments were calculated by comparison with the control strains, andimages were normalised accordingly. Different profiles were assignedto types in accordance with differences in the restriction patterns.
3. Results
3.1. Salmonella prevalence and serotypes
Salmonella spp. was isolated from 64 of 425 samples of slaughteredpigs (15.0%) and from 13 of 41 environmental samples (31.8%).
es and for class 1 integrons.
eters Reference
1 Annealing Extension2
54 °C (30 s) 72 °C (1 min) Leverstein-van Hall et al., 2002
53 °C (30 s) 72 °C (1 min) Lévesque et al., 1995
54 °C (1 min) 72 °C (1 min) Randall et al., 2004
60 °C (1 min) 72 °C (1 min) Randall et al., 2004
54 °C (1 min) 72 °C (1 min) Randall et al., 2004
52 °C (1 min) 72 °C (50 s) Randall et al., 2004
56 °C (1 min) 72 °C (1 min) Randall et al., 2004
56 °C (1 min) 72 °C (1 min) Randall et al., 2004
54 °C (1 min) 72 °C (1 min) Randall et al., 2004
60 °C (1 min) 72 °C (1 min) Randall et al., 2004
56 °C (1 min) 72 °C (1 min) Randall et al., 2004
56 °C (1 min) 72 °C (1 min) Randall et al., 2004
60 °C (1 min) 72 °C (1 min) Randall et al., 2004
cle.

Table 2Prevalence (%) of Salmonella per sample type and slaughterhouse.
S1 S2 S3 S4 S5
Pig samplesLymph nodes 66.6 0 16 60 0Colon content 0 0 8 45 18.8Carcass sponge 0 0 20 35 0Liver sponge 0 0 16 40 0Tonsils 0 0 0 0 0Total 13.3 0 12 36 3.8
Environmental samples 0Contact surfaces with meat 0 0 43 50 25Not contact surfaces with meat 0 0 50 75 0Scalding water 0 0 0 50 0Total 0 0 37.5 60 10
204 F. Piras et al. / International Journal of Food Microbiology 151 (2011) 201–209
Table 2 shows the Salmonella prevalence per slaughterhouse in pigand environmental samples. The prevalence of Salmonella in pig sampleswas 36.0% in slaughterhouse S4, 13.3% in S1, 12.0% in S3 and 3.8%in S5. Further, the prevalence of Salmonella in the samples differedbetween SDwithin slaughterhouses. The overall prevalence of Salmonellain S4 was generally higher than in the other abattoirs in almost all pigsamples. Salmonella was not detected in any of the samples taken at S2.With regards to sampling site, the highest prevalence was observed inmesenteric lymph node samples (30.5%), followed by colon contents(16.4%), carcasses and liver (both 14.1%). Salmonella was not detected inany of the tonsil samples. Thirty one pigs were found to carry Salmonellain lymph nodes and/or colon content and in 8 of these pigs Salmonellacould also be isolated from the carcass.
In environmental samples Salmonella prevalencewas slightly higherin samples from surfaces not in contact with meat (8/21, 38%) than insamples from surfaces in contact withmeat (7/20, 35%). The prevalenceof Salmonella in environmental samples was 37.5% (6/16) in S3, 60.0%(6/10) in S4, and 10.0% (1/10) in S5. Salmonella was not detected inany of the samples taken at S1 and S2. In S3 Salmonella was isolated inall the samples of drain water, in one sample of dehairing equipment(20.0%) and in one sample of carcass splitter equipment (20.0%). In S4Salmonella was detected in all environmental samples, including onesample of scalding water.
A total of 103 Salmonella isolates were isolated and they belonged to8 serotypes. Table 3 shows the detail of the Salmonella serotypes isolatedfrom thepig and environmental samples per slaughterhouse andper SD.The SD is indicated as I, II, III and IV preceded by the slaughterhouse
Table 3Distribution of the Salmonella serotypes isolated in pig and environmental samples per slaucated between brackets.
Samples S1I S1II S3I S3II S3III
Carcass
Colon content Infantis (1)Typhimurium193 (1)
Mesentericlymph nodes
Derby(5)
Livingstone(5)
Typhimurium193 (1)
Derby (2)
Liver Typhimurium104B (1)
Drain water TyphimuriumU302 (1)
Typhimurium59 (1)
Dirty zone wall
Splittingequipment
Typhimurium59 (2)
Dehairingequipment
Typhimurium59 (1)
Scalding water
abbreviation. S. Derby was the most prevalent serotype isolated frompig samples accounting for 47.0% of all isolates. Other Salmonella sero-types that were isolated from the pig samples were Typhimurium(16.1%), Panama (21.9%), Livingstone (5.8%), Infantis (5.8%) and Rissen(4.6%). Serotypes Newport and Bredeney were isolated relatively rarely(1.1%).
Only two serotypes were identified among the 16 strains isolatedfrom environmental samples: S. Typhimurium (71.5%) and S. Derby(28.5%).
Phage typing of S. Typhimurium isolates resulted into 4 differentphage types: DT193 (12 strains), DT59 (4), DT104b (1), and U302(1) (Fig. 2). Six S. Typhimurium strains were untypeable by phagetyping.
3.2. Antimicrobial susceptibility testing and detection of class I integrons
All the 103 isolates were susceptible to ciprofloxacin, gentamicin,nalidixic acid, cefotaxime and colistin. Of all the isolates tested 71(69.0%) were susceptible to all the antimicrobials tested. The remaining32 isolates (31.0%) were found to be resistant to at least one antibiotic.The most common resistance phenotype observed was to cefalothin(100%), followed by ampicillin (90.7%), tetracycline (65.7%), sulpha-methoxazole (62.5%), amoxicillin–clavulanic acid (53.1%), chloram-phenicol (34.4%), trimethoprim (28.2%) and streptomycin (22.0%).
Antimicrobial resistance (AMR) was found among the followingserotypes: S. Typhimurium (95.6%), S. Derby (18.2%) and S. Rissen(25.0%) Among the 32 isolates that showed resistance 30 (93.8%),were found to be resistant to more than one antimicrobial.
Not all the integron-associated resistance genes, investigated inour study, were present in the isolates (Table 4). In fact there was al-ways no correlation between the presence of resistance genes andcorresponding resistance phenotypes. All the strains that werestreptomycin-resistant contained both the genes aadA1 and aadA2;all the strains that were tetracycline-resistant contained the genetetA(G) and either the gene tetA(A) or tetA(B). However, there wereseveral cases in which a strain was resistant to an antibiotic but theidentity of the gene conferring resistance could not be ascribed.None of the strains that were ampicillin-resistant contained thegene bla(carb2) and 8 of 30 strains did not contain the bla(tem)gene. 7 of 11 chloramphenicol-resistant strains contained the cat1gene and none of them contained the cat2 gene. Just 4 of 15 strainsthat were trimethoprim-resistant contained the dhfr1 gene.
ghterhouse (S1→S5) and sampling day (I→ IV); the number of strains isolated is indi-
S3IV S4I S4II S4III S5I
Panama (12)Typhimurium unt (3)
Derby(3)
Derby (4)
Panama (2)Rissen (1)
Derby(5)
Derby (2)Infantis(2)
Typhimurium193 (7)Infantis (2)
Bredeney (1)Newport (1)Typhimurium 193 (1)
Derby(4)
Derby (5)
Panama (5)Rissen (3)
Derby(3)
Derby (5)
Typhimurium unt (3) Derby(1)
Derby (1)
Derby(1)
Derby (1) Typhimurium193 (2)
Derby(1)
91.4
77.2
64.4
PFGE-XbaI100908070
PFGE-XbaI20.0040.00100.00150.00200.00250.00400.00500.00600.00800.0015002000
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
FP3
FP3
FP1
FP2
FP4
FP4
FP5
FP38
FP39
FP19
FP20
FP31
FP32
FP33
FP34
FP35
FP36
FP37
FP44
FP46
FP48
FP49
FP50
FP51
FP52
FP53
FP54
FP55
FP56
FP57
FP58
FP59
FP61
FP62
FP63
FP21
FP22
FP23
FP24
FP25
FP26
FP27
FP28
FP29
FP30
FP62
FP60
Derby
Derby
Derby
Derby
Derby
Derby
Derby
Derby
Derby
Derby
Derby
Derby
Derby
Derby
Derby
Derby
Derby
Derby
Derby
Derby
Derby
Derby
Derby
Derby
Derby
Derby
Derby
Derby
Derby
Derby
Derby
Derby
Derby
Derby
Derby
Derby
Derby
Derby
Derby
Derby
Derby
Derby
Derby
Derby
Derby
Derby
Derby
DerX23
DerX23
DerX21
DerX21
DerX21
DerX21
DerX21
DerX26
DerX26
DerX22
DerX22
DerX22
DerX22
DerX22
DerX22
DerX22
DerX22
DerX22
DerX22
DerX22
DerX22
DerX22
DerX22
DerX22
DerX22
DerX22
DerX22
DerX22
DerX22
DerX22
DerX22
DerX22
DerX22
DerX22
DerX22
DerX22
DerX22
DerX22
DerX22
DerX22
DerX22
DerX22
DerX22
DerX22
DerX22
DerX22
DerX22
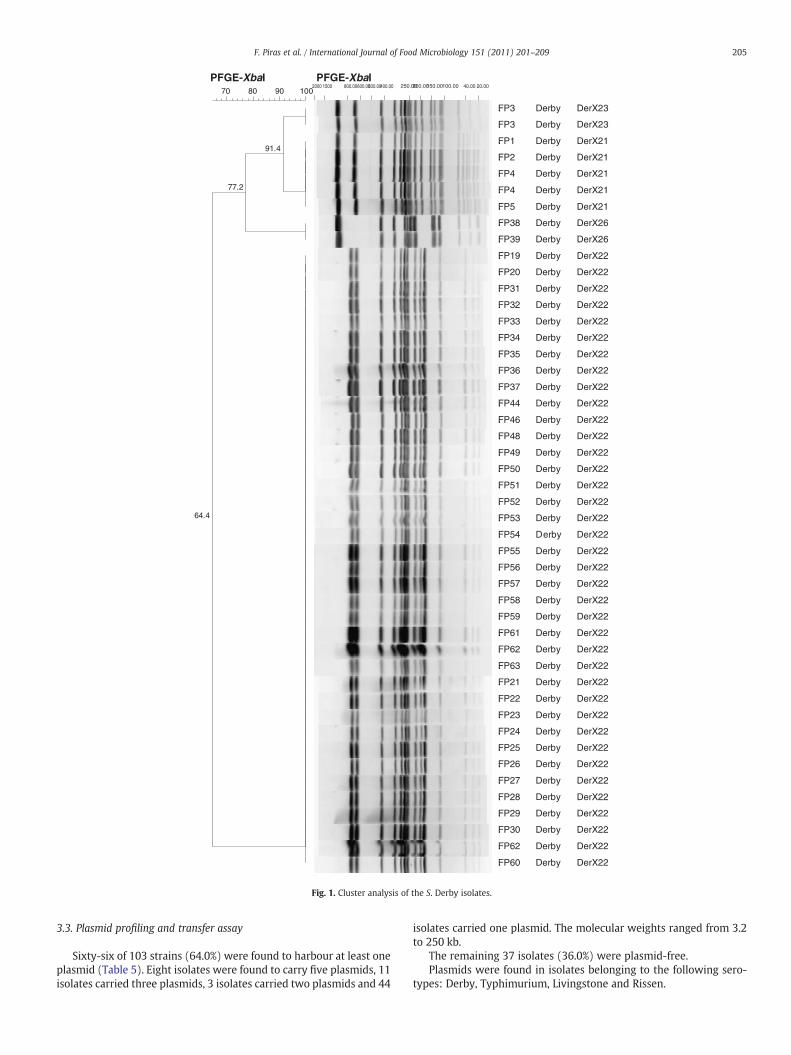
Fig. 1. Cluster analysis of the S. Derby isolates.
205F. Piras et al. / International Journal of Food Microbiology 151 (2011) 201–209
3.3. Plasmid profiling and transfer assay
Sixty-six of 103 strains (64.0%) were found to harbour at least oneplasmid (Table 5). Eight isolates were found to carry five plasmids, 11isolates carried three plasmids, 3 isolates carried two plasmids and 44
isolates carried one plasmid. The molecular weights ranged from 3.2to 250 kb.
The remaining 37 isolates (36.0%) were plasmid-free.Plasmids were found in isolates belonging to the following sero-
types: Derby, Typhimurium, Livingstone and Rissen.

92.0
83.7
93.3
89.7
83.0
67.9
PFGE-XbaI
100908070
PFGE-XbaI20.0040.00100.00150.00200.00250.00400.00500.00600.00800.0015002000
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
FP98
FP99
FP101
FP102
FP104
FP97
FP100
FP103
FP17
FP40
FP41
FP42
FP43
FP76
FP77
FP91
FP92
FP78
FP93
FP18
FP12
FP13
FP14
Typhimurium
Typhimurium
Typhimurium
Typhimurium
Typhimurium
Typhimurium
Typhimurium
Typhimurium
Typhimurium
Typhimurium
Typhimurium
Typhimurium
Typhimurium
Typhimurium
Typhimurium
Typhimurium
Typhimurium
Typhimurium
Typhimurium
Typhimurium
Typhimurium
Typhimurium
Typhimurium
DT193
DT193
DT193
DT193
DT193
DT193
DT193
DT193
DT104b
untyp
untyp
untyp
untyp
untyp
untyp
untyp
untyp
untyp
untyp
U302
DT193
untyp
DT193
STYMXB.0046
STYMXB.0046
STYMXB.0046
STYMXB.0046
STYMXB.0046
STYMXB.0046
STYMXB.0046
STYMXB.0046
Tm120X15
STYMXB.0079
STYMXB.0079
STYMXB.0079
STYMXB.0079
TmuntX16
TmuntX16
TmuntX16
TmuntX16
TmuntX16
TmuntX16
STYMXB.0058
STYMXB.0051
STYMXB.0051
Tm193X13
Fig. 2. Cluster analysis of the S. Typhimurium isolates.
206 F. Piras et al. / International Journal of Food Microbiology 151 (2011) 201–209
Among S. Typhimurium and S. Derby isolates, 7 and 6 plasmid pro-files respectively, were identified. Five S. Derby strains, isolated fromlymph nodes samples of five different animals in slaughterhouse S1during the same sampling day, carried two different plasmids of ca.90 and 3.6 kb, but one was sensitive to all the antimicrobials tested;the other four, which harboured an additional large plasmid of ca.180 kb, showed the MDR profile ACSuTTmKf.
The recipient E. coli J53-2 (mated with the donor S. TyphimuriumU302) exhibited resistance to tetracycline: this could be demonstrat-ed by the fact that on the CLED plate containing tetracycline and thetransconjugant were inoculated, it was possible to see typical E.colicolonies (yellow, opaque colonieswith a slightly deeper coloured centre):this result could indicate that resistance genes tetA (G) and/or tetA (B) arelocated on plasmid DNA.
3.4. PFGE analysis
Genomic DNA was analysed by pulsed field gel electrophoresis,using the XbaI enzyme. Among S. Derby isolates, four PFGE patternswere identified in S1 during the SD I, two pulsetypes (4 designatedas DerX21 and 1 DerX23) were detected in 5 lymph nodes from 5pigs.
Table 4Resistance genetic sequence identified in class 1 integron-carrying multidrug-resistant stra
Serotype AMR pattern n. Int1 CS aadA1 aadA2 bla(t
Derby ACSuTTmKf 4 Pos Pos +Typhimurium DT 193 ASSuTTmKf 2 Neg Neg + + +Typhimurium DT104B ASSuTKf 1 Neg Neg + + +Typhimurium U302 ACSuTTmKf 1 Pos Pos −Derby ASuTTmKfAx 2 Neg Neg +Typhimurium DT59 ASSuTKf 4 Neg Neg + + +Typhimurium unt ACSuTKfAx 6 Pos Pos −Rissen Tkf 1 Neg Neg −Typhimurium DT 193 AKfAx 9 Neg Neg +
The variation of pulsotype DerX23 could be justified by the ab-sence of two bands of the molecular weight of about 180-kb(Fig. 1). This result had been confirmed by the plasmid profilingwhich exhibited, the absence of a large plasmid of 180-kb in pulso-type DerX23. Moreover, this latest pattern showed susceptibility toall the tested antibiotics, suggesting that the AMR markers could belocated on conjugative plasmid.
In S4 the same pulsetype (DerX22) was isolated during two SD,from almost all pig and environmental samples.
Among S. Typhimurium seven PFGE patterns were identified. Itwas not possible to isolate the same pulsotype within the sameslaughterhouse in different sampling days.
The ability to assign a PFGE type to some phage type untypable S.Typhimurium strains (Table 5) was an interesting data, as it can pro-vide evidence of clonal relationships among these isolates. Otherwise,the variation observed in the PFGE patterns for S. Typhimurium iso-lates was in accordance with the results of phage typing.
We identified three different PFGE patterns within isolates ofDT193, from three different slaughterhouses, that provide evidencethat there are different clones of DT193 circulating on the island.This phage type is known to be highly variable (Doran et al., 2005),this variation probably being an indicator that DT193 is polyphyletic,
ins of Salmonella.
em) bla(carb2) cat1 cat2 dhfr1 sul1 tetA(A) tetA(B) tetA(G)
− − − + + − + +− − − + − +− − − + +− + − − + − + +− − − − + +− − − + +− + − − − − + +− − + − +−

Table 5Characteristics of the 103 strains of Salmonella belonging to different serotypes (n. of isolates).
Serotype Slaughterhouse SD Pulsetype PP (kb) Source AMR pattern
Derby (4) S1I DerX21 180;90;3.6 Lymph node ACSuTTmKfDerby (1) DerX23 90;3.6 Lymph node Fully sensitiveDerby (2) S3III DerX24 90;11.0;5.4 Lymph nodes ASuTTmKfAxDerby (2) S4II DerX22 12.0;6.2;5.7 Lymph nodes KfDerby (7) S4II and S4III DerX22 5.7 Lymph nodes Fully sensitiveDerby (7) S4II and S4III DerX22 5.7 Colon content Fully sensitiveDerby (2) S4II DerX22 5.7;5.3;4.2 Liver Fully sensitiveDerby (6) S4II and S4III DerX22 5.7 Liver Fully sensitiveDerby (7) S4II and S4III DerX22 5.7 Carcass Fully sensitiveDerby (1) S4II DerX22 5.7 Scalding water Fully sensitiveDerby (2) S4II and S4III DerX22 5.7 Drain water Fully sensitiveDerby (1) S4II DerX22 5.7 Dirty zone wall Fully sensitiveDerby (1) S4II DerX22 5.7 Dehairing equipment Fully sensitiveDerby (1) S4III DerX22 5.7 Splitting equipment Fully sensitiveTyphimurium 193 (2) S3I STYMXB.0051 110;7.0 Colon content ASSuTTmKfTyphimurium 104B (1) S3II Tm120X15 11.0;6.5;5.8;4.3;3.8 Liver ASSuTkfTyphimurium U302 (1) STYMXB.0058 120;5.2;4.7;4.4;3.7 Drain water ACSuTTmKfTyphimurium 59 (1) S3III STYMXB.0079 like Free Dehairing equipment ASSuTKfTyphimurium 59 (1) STYMXB.0079 like Free Drain water ASSuTKfTyphimurium 59 (1) STYMXB.0079 like Free Splitting equipment ASSuTKfTyphimurium 59 (1) STYMXB.0079 like Free Dirty zone wall ASSuTKfTyphimurium unt (3) S3IV TmuntX16 130;6.7;4.1;3.6;3.3 Carcass ACSuTKfAxTyphimurium unt (3) TmuntX16 130;6.7;4.1;3.6;3.3 Drain water ACSuTKfAxTyphimurium 193 (1) S4I Tm193X13 250;90;3.2 Lymph nodes fully sensitiveTyphimurium 193 (7) S5I STYMXB.0046 55 Colon content AKfAxTyphimurium 193 (2) STYMXB.0046 55 Splitting equipment AKfAxPanama (12) S3IV PanX18 Free Carcass swabs Fully sensitivePanama (2) PanX18 Free Colon content Fully sensitivePanama (5) PanX18 Free Liver Fully sensitiveLivingstone (4) S1II LivX15 Free Lymph nodes Fully sensitiveLivingstone (1) LivX15 85 Lymph nodes Fully sensitiveInfantis (1) S3I SINFXB.0074 Free Colon content Fully sensitiveInfantis (2) S4III SINFXB.0039 Free Colon content Fully sensitiveInfantis (2) S5I SINFXB.0014 Free Colon content Fully sensitiveRissen (1) S3IV RisX10 4,8 Colon content TKfRissen (3) RisX9 Free Liver Fully sensitiveNewport (1) S4I NewX70 Free Lymph nodes Fully sensitiveBredeney (1) BreX16 Free Lymph nodes Fully sensitive
207F. Piras et al. / International Journal of Food Microbiology 151 (2011) 201–209
i.e. the phenotype can be achieved via several different genomic line-ages. The three S. Infantis patterns (SINFXB.0014, SINFXB.0039 andSINFXB.0074), observed in this study, are all relatively close (Fig. 3).This may be indicative of minor genetic variation in the clones ofthis serotype circulating on Sardinia.
4. Discussion
The results of this survey demonstrate that the prevalence ofSalmonella contamination in slaughtered pigs in Sardinia is high.Salmonella was observed at one or more sampling sites in 41.0% of pigssampled. Mesenteric lymph nodes resulted in the highest prevalence(30.5%). The results about Salmonella prevalence are in accordance
81.0
78.7
96.8
58.5
PFGE-XbaI10090807060
PFGE-XbaI2300.00400.00500.00800.001500
Fig. 3. Cluster analysis of the S. In
with other similar studies (Swanenburg et al., 2001c; Botteldoorn et al.,2003).
Differences in Salmonella prevalence were observed in relation toSD and slaughterhouses, that may depend on the unhomogeneousapplication of Slaughtering and Hygienic Practises by the operators,the moment of the sampling and the origin of the animals.
Salmonella was isolated from one water sample of the scaldingtank in slaughterhouse S4. This can indicate that the water tempera-ture was not always above 60 °C or that Salmonellamay have been as-sociated with residual organic material, and consequently protectedagainst high temperature (Swanenburg et al., 2001c). Usually, con-taminated drain water is not considered as a critical point, but carcasscontamination can occur if water from the drains is sprayed during
20.0040.00100.00150.0000.00
.
.
.
.
.
.
.
.
.
FP95
FP96
FP45
FP47
FP11
FP88
FP94
FP90
FP79
Infantis
Infantis
Infantis
Infantis
Infantis
Rissen
Rissen
Rissen
Rissen
SINFXB.0014
SINFXB.0014
SINFXB.0039
SINFXB.003 9
SINFXB.0074
RisX9
RisX9
RisX9
RisX10
fantis and S. Rissen isolates.

208 F. Piras et al. / International Journal of Food Microbiology 151 (2011) 201–209
cleaning procedures, e.g. under high pressure and aerosol formation.The persistence of Salmonella in drain water samples was probablycorrelated with inadequate application of cleaning procedures, allow-ing the persistence of “house strains” in biofilm production (Malcovaet al., 2008). It cannot be excluded that Salmonella was able to multi-ply in the drains (Swanenburg et al., 2001c).
Our study showed that many salmonellae were brought into theslaughterhouses by the delivered pigs. In fact, 31 pigs were carriersof Salmonella in lymph nodes and/or colon content. In 8 of thesepigs, Salmonellawas also isolated from the carcass. For the interpreta-tion of our data, it should be noted that slaughtered carrier pigs (withpositive colon content and/or mesenteric lymph nodes) could havebeen infected on the farm and during transport, or during the waitingperiod in the lairage prior to slaughter. Several authors have reportedthe detection of Salmonella in the faeces and the mesenteric lymphnodes as early as 3 h after infection (Fedorka-Cray et al., 1995;2000). In particular, the lairage and the high contamination levelof the slaughterhouse environment are probably the main sourcefor Salmonella infections prior to slaughter (Hurd et al., 2001a,b;Swanenburg et al., 2001b). However, a direct correlation betweencarrier status and carcass contamination levels is not clear.
Salmonella serotypes, which can survive in certain niches of theslaughterhouse environment, may become part of the resident flora(house strains). This can result in subsequent carcass contaminationof slaughter pigs passing along the slaughter line.
The most prevalent serotypes were S. Derby and S. Typhimurium;these results are in accordance with other similar studies carried outin pig farms (Weigel et al., 2007) and at slaughterhouses (Nollet et al.,2004). It is important to keep in mind that all the serotypes associatedwith swine species are considered potentially dangerous for publichealth, and cases of non typhoidal salmonellosis have been describedin humans.
The prevalence of S. Typhimurium DT 193 (50.0%) is of particularsignificance, as this phage type has been associated with foodborneoutbreaks (in Italy and the United Kingdom) traced to contaminatedpork products (Gebreyes and Altier, 2002; Gebreyes et al., 2004).
Observations in this study on the prevalence of antimicrobialresistance in Salmonella swine isolates are not in accordance withother studies carried out at slaughter (Davies et al., 2004), whereresistance to tetracycline was the most common phenotype observedin 75.6% of 716 isolates of S. enterica. MDR was found in associationwith serotypes Derby and Typhimurium: the increasing multipleantimicrobial resistance, associated with such pork-related serotypes,may become a serious human health hazard in the near future (Boyenet al., 2008).
In our study it was not possible to detect resistance to fluoroqui-nolones and cephalosporins, which represents a favourable situationwith regard to public health.
In general, a high level of resistance was observed to frequentlyused antimicrobials in veterinary and public health practises, includ-ing streptomycin and tetracycline, as was expected (Aragaw et al.,2007).
However, we also observed resistance to antimicrobials that arenot commonly used in animal health and production in Europe,such as ampicillin, ciprofloxacin, nalidixic acid, chloramphenicol,florfenicol, and gentamicin. This suggests the importance of othersources of MDR Salmonella serovars, such as: imported Salmonellacontaminated feed, living animals and foodstuff, use of antimicrobialsin the human health sector and possibly cross-resistance among therelated antimicrobials, or co-selection of resistances carried on thesame DNA element.
We detected differences in the resistance to antibiotics of the ami-noglycoside class: the overall resistance to ampicillin was high(90.7%), while resistance to amoxicillin–clavulanic acid was 53.1%,to streptomycin 22% and no isolate showed resistance to gentamicin.Our results are in accordance with a previous study (Farrington et al.,
2001), which showed that antimicrobials belonging to the same classusually possess slight structural differences to avoid cross-resistance.
The PFGE subtyping provided additional strain information, to en-hance the Salmonella traceability in Sardinia pig slaughterhouses. Alarge variability of Salmonella population at pig abattoir has been con-firmed. Moreover the detection of the same S. Derby pulsotype(DerX22) within the same plant, during two different SD, from bothpig and environmental samples, suggested that the strain had becomepersistent, and therefore a probable source of environment and car-cass cross-contamination. This would also be supported by the factthat the animals slaughtered during the first SD were from Holland,while those slaughtered during the second SD were of local origin,making it highly unlikely that the two pig herds carried the samestrain prior to arrival at the slaughterhouse.
This study provided additional information about the Salmonellaprevalence in Sardinian slaughterhouses, and highlighted the impor-tance of improving the application of some preventive measures inorder to avoid the presence of a residential Salmonella flora in theslaughterhouse. Slaughter should be performed according to HACCPprinciples in association with good hygiene procedures (GHP), inorder to avoid faecal contamination of carcasses by increasing careduring evisceration, and modifying the meat inspection procedures.Regular cleaning and disinfecting of all equipment, also duringslaughter, should be carried out and the effectiveness of cleaningand disinfection should be checked.
References
Anonymous, 2003. Regulation (EC) No 2160/2003 of the European Parliament and ofthe Council of 17 November 2003 on the control of Salmonella and other specifiedfood-borne zoonotic agents. Official Journal of European Union 325, 1–15.
Aragaw, K., Molla, B., Muckle, A., Cole, L., Wilkie, E., Poppe, C., Kleer, J., Hildebrandt, G.,2007. The characterization of Salmonella serovars isolated from apparently healthyslaughtered pigs at Addis Ababa abattoir, Ethiopia. Preventive Veterinary Medicine82, 252–261.
Bonardi, S., Brindani, F., Pizzin, G., Lucidi, L., D'Incau, M., Liebana, E., Morabito, S., 2003.Detection of Salmonella spp., Yersinia enterocolitica and verocytotoxin-producingEscherichia coli O157 in pigs at slaughter in Italy. International Journal of Food Mi-crobiology 85, 101–110.
Botteldoorn, N., Heyndrickx, M., Rijpens, N., Grijspeerdt, K., Herman, L., 2003. Salmonellaon pig carcasses: positive pigs and cross contamination in the slaughterhouse. Journalof Applied Microbiology 95, 891–903.
Boyen, F., Haesebrouck, F.,Maes, D., Van Immerseel, F., Ducatelle, R., Pasmans, F., 2008. Non-typhoidal Salmonella infections in pigs: a closer look at epidemiology, pathogenesis andcontrol. Veterinary Microbiology 130, 1–19.
Clinical, Laboratory Standards Institute (CLSI/NCCLS), 2004. Performance standards forantimicrobial susceptibility testing, Informational Supplement. NCCLS DocumentM100 S14 (ISBN 1 56238 516 X), 14th ed. CLSI/NCCLS, Wayne, PA. USA.
Davies, R.H., Dalziel, R., Gibbens, J.C., Wilesmith, J.W., Ryan, J.M.B., Evans, S.J., Byrne, C.,Paiba, G.A., Pascoe, S.J.S., Teale, C.J., 2004. National survey for Salmonella in pigs,cattle and sheep at slaughter in Great Britain (1999–2000). Journal of Applied Mi-crobiology 96, 750–760.
Doran, G., Morris, D., O'Hare, C., DeLappe, N., Bradshaw, B., Corbett-Feeney, G., Cormican,M., 2005. Cost-effective application of pulsed-field gel electrophoresis to typing ofSalmonella enterica Serovar Typhimurium. Applied and Environmental Microbiology71, 8236–8240.
Farrington, L.A., Harvey, R.B., Buckley, S.A., Droleskey, R.E., Nisbet, D.J., Inskip, P.D.,2001. Prevalence of antimicrobial resistance in salmonellae isolated frommarket-age swine. Journal of Food Protection 64, 1496–1502.
Fedorka-Cray, P.J., Kelley, L.C., Stabel, T.J., Gray, J.T., Laufer, J.A., 1995. Alternate routes ofinvasion may affect pathogenesis of Salmonella Typhimurium in swine. Infectionand Immunity 63, 2658–2664.
Fedorka-Cray, P.J., Gray, J.T., Wray, C., 2000. Salmonella infections in pigs. In: Wray, C.,Wray, A. (Eds.), Salmonella in Domestic Animals. CAB International, Wallingford,pp. 191–207.
Gebreyes,W.A., Altier, C., 2002.Molecular characterization ofmultidrug-resistant Salmonellaenterica subsp. enterica serovar Typhimurium isolates fromswine. Journal of ClinicalMi-crobiology 8, 2813–2822.
Gebreyes, W.A., Thakur, S., Davies, P.R., Funk, J.A., Altier, C., 2004. Trends in antimicrobialresistance, phage types and integrons among Salmonella serotypes from pigs 1997–2000. The Journal of Antimicrobial Chemotherapy 53, 997–1003.
Hurd, H.S., Stabel, T.J., Carlson, S., 2001a. Sensitivity of various fecal sample collectiontechniques for detection of Salmonella Typhimurium in finishing hogs. Proceedingsof the Third International Symposium for Epidemiology and Control of Salmonellain Pork. Washington DC, USA, pp. 63–64.
Hurd, H.S., Gailey, J.K.,McKean, J.D., Rostagno,M.H., 2001b. Rapid infection inmarket-weightswine following exposure to a Salmonella Typhimurium-contaminated environment.American Journal of Veterinary Research 62, 1194–1197.

209F. Piras et al. / International Journal of Food Microbiology 151 (2011) 201–209
Kado, C.I., Liu, S.-T., 1981. Rapid procedure for detection and isolation of large and smallplasmids. Journal of Bacteriology 145, 1365–1373.
Leverstein-van Hall, M.A., Paauw, A., Box, A.T.A., Blok, H.E.M., Verhoef, J., Fluit, A.C., 2002.Presence of integron-associated resistance in the community is widespread andcontributes to multidrug resistance in the hospital. Journal of Clinical Microbiology40, 3038–3040.
Lévesque, C., Piché, L., Larose, C., Roy, P.H., 1995. PCRmapping of integrons reveals severalnovel combinations of resistence genes. Antimicrobial Agents and Chemoterapy 39,185–191.
Lo Fo Wong, D.M.A., Hald, T., van der Wolf, P.J., Swanenburg, M., 2002. Epidemiologyand control measures for Salmonella in pigs and pork. Livestock Production Science76, 215–222.
Lo Fo Wong, D.M.A., Dahl, J., Stege, H., van der Wolf, P.J., Leontides, L., von Altrock, A.,Thorberg, B.M., 2004. Herd-level risk factors for subclinical Salmonella infectionin European finishing-pig herds. Preventive Veterinary Medicine 62, 253–266.
Magistrali, C., Dionisi, A.M., De Curtis, P., Cucco, L., Vischi, O., Scuota, S., Zicavo, A., Pezzotti,G., 2008. Contamination of Salmonella spp. in a pig finishing herd, from the arrival ofthe animals to the slaughterhouse. Research in Veterinary Science 85, 204–207.
Malcova, M., Hradecka, H., Karpiskova, R., Rychlik, I., 2008. Biofilm formation in fieldstrains of Salmonella enterica serovar Typhimurium: identification of a new colonymorphology type and the role of SGI1 in biofilm formation. Veterinary Microbiology129, 360–366.
Nollet, N., Maes, D., De Zutter, L., Duchateau, L., Houf, K., Huysmans, K., Imberechts, H., Geers,R., de Kruif, A., Van Hoof, J., 2004. Risk factors for the herd-level bacteriologic prevalenceof Salmonella in Belgian slaughter pigs. Preventive Veterinary Medicine 65, 63–75.
Peters, T.M., Maguire, C., Threlfall, E.J., Fisher, I.S., Gill, N., Gatto, A.J., Salm-gene project,2003. The Salm-gene project — a European collaboration for DNA fingerprinting.Euro Surveillance 8 (2), 46–50.
Randall, L.P., Cooles, S.W., Osborn, M.K., 2004. Antibiotic resistance genes, integronsand multiple antibiotic resistance in thirty-five serotypes of Salmonella entericaisolated from humans and animals in the UK. The Journal of Antimicrobial Chemo-therapy 23, 208–216.
Swanenburg, M., van der Wolf, P.J., Urlings, H.A.P., Snijders, J.M.A., van Knapen, F.,2001a. Salmonella in slaughter pigs the effect of logistic slaughter procedures ofpigs on the prevalence of Salmonella on pork. International Journal of Food Micro-biology 70, 231–242.
Swanenburg, M., Urlings, H.A.P., Keuzenkamp, D.A., Snijders, J.M.A., 2001b. Salmonellain the lairage of pig slaughterhouses. Journal of Food Protection 64, 12–16.
Swanenburg, M., Urlings, H.A.P., Snijders, J.M.A., Keuzenkamp, D.A., van Knapen, F.,2001c. Salmonella in slaughter pigs: prevalence, serotypes and critical controlpoints during slaughter in two slaughterhouses. International Journal of Food Mi-crobiology 70, 243–254.
van der Wolf, P.J., Elbers, A.R.W., van der Heijden, H.M.J.F., van Schie, F.W., Hunneman,W.A., Tielen, M.J.M., 2001. Salmonella seroprevalence at the population and herdlevel in pigs in The Netherlands. Veterinary Microbiology 80, 171–184.
Weigel, R.M., Nucera, D., Qiao, B., Teferedegne, B., Kyun Suh, D., Barber, D.A., Bahnson,P.B., Isaacson, R.E., White, B.A., 2007. Testing an ecological model for transmissionof Salmonella enterica in swine production ecosystems using genotyping data. Pre-ventive Veterinary Medicine 81, 274–289.
Zheng, D.M., Bonde, M., Sørensen, J.T., 2007. Associations between the proportion ofSalmonella seropositive slaughter pigs and the presence of herd level risk factorsfor introduction and transmission of Salmonella in 34 Danish organic, outdoor(non-organic) and indoor finishing-pig farms. Livestock Science 106, 189–199.