karyotypic evolution in gehyra (gekkonidae: reptilia) iii...
TRANSCRIPT

Aust. J. Zool., 1983, 31, 723-41
Karyotypic Evolution in Gehyra (Gekkonidae: Reptilia) III.* The Gehyra australis Complex
Max King
Department of Population Biology, Research School of Biological Sciences, Australian National University, Canberra, A.C.T. 2601.
Abstract
A karyotypic analysis of the widely distributed Australian gekko Gehyra australis has revealed a complex of chromosome races. Each of these is fixed for a particular karyomorph, and is found over a geographically delimited distribution. The seven chromosome races differ by number, or chromosome morphology, and are defined as: 2n = 44,2n = 42a, 2n = 42b, 2n = 42c, 2n = 40a, 2n = 40b, 2n = 38. In addition, a series of animals from these races possess novel chromosome rearrangements. A model for the evolution of this complex of races is proposed, and the relationship ofthese forms to each other and to members of the Gehyra variegata-punctata complex is discussed.
Introduction
It is now clear that chromosome evolution in terms of gross chromosomal change has been far more extensive in some reptilian lineages than in others. In the Iguanidae, Agamidae and Lacertidae many species share a karyotype format common to that group. Thus, most iguanids have a 12V + 24m karyotype with 2n = 36; most Australian agamids have 2n = 32 made up of 12V + 20m elements, and most Lacertidae have 2n = 38 all acrocentric chromosomes (see Gorman 1973; King 1981a). However, some species within these families do show interspecific variation in their karyotype morphology attributable to fusion, fission or pericentric inversion.
Considerable intraspecific chromosome variation is also present in certain groups of lizards. This variation is expressed in the form of chromosome races, polymorphisms, sex chromosome races and rare mutant animals, and is most commonly found in certain iguanid and gekkonid lizards. Indeed, more chromosome race variation has been encountered in the Australian Gekkonidae than in any other group of reptiles, with species from the genera Diplodactylus [D. tessellatus, 2n = 28, 38 (King 1982a); D. vittatus, 2n = 38A, 38B, 38C, 34, 36 (King 1977a)], Phyllodactylus [P. marmoratus, 2n= 36, 36ZW, 34, 32 (King and Rofe 1976; King and King 1977)] and Hemidactylus [H. frenatus, 2n=46, 40 (King 1978)l providing striking examples of the situation.
Perhaps the most interesting genus in terms of its chromosome race diversity is Gehyra. In an analysis of the G. variegata-punctata complex, King (1979) found that there were five chromosome races (2n = 44, 42, 40a, 40b, 38) each of which was allopatrically distributed. Specimens from these races also showed characteristic differences in their external morphology. In an analysis of specimens of Gehyra australis from a relatively restricted area, King (1982b) was able to show that two chromosomally distinct forms were present (2n = 40 and 2n = 42) and that these were morphologically distinguishable.
* Part 11, Aust. J. Zool., 1982, 30, 93-101. 004-959X/83/050723$02.00

Max King
The present paper presents the results of a detailed karyotypic analysis of Gehyra australis throughout its Australian distribution; the distributions of the karyoptypic variants are described; and the evolutionary, phylogenetic and taxonomic implications are considered.
Materials and Methods In this study a total of 81 specimens collected from 40 localities were karyotyped (see Fig. 1;
Table 1). Chromosomes were obtained from each specimen by means of intestinal epithelial preparations. In
males, testis preparations were also made. The air-drying techniques used are described in detail by King and Rofe (1976). At least 10 cells were examined from each individual to ascertain chromosome number and morphology.
All specimens used in this study have been preserved for morphometric analysis and will be lodged in the relevant State museum collections.
EZ8 G. xenopus Storr
Fig. 1. Map showing the approximate distribution of the chromosome races of G. australis. The closely related G. xenopus is also included. Precise localities of specimens are shown in Table 1.
Results
Gehyra australis Gray shows considerable karyotypic variation throughout its mainland distribution. This occurs in two forms. First, a series of chromosome races are present, each of which is fixed for a particular karyomorph and occurs over a defined geographic distribution. Second, animals possessing novel chromosome rearrangements occur as population isolates, as rare mutants, or as a polymorphism.
Karyotypic Evolution in Gehyra. I11
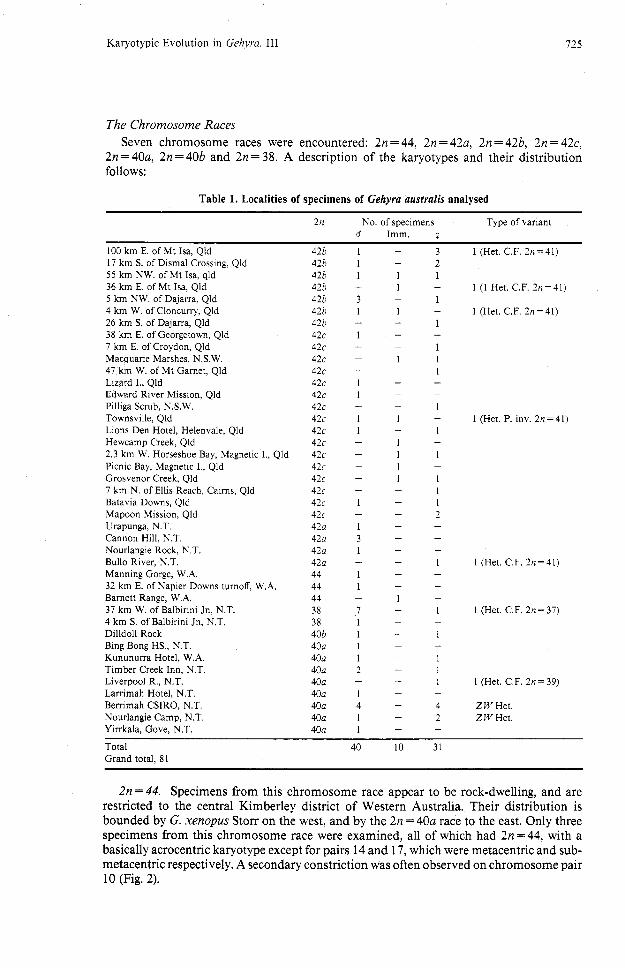
The Chromosome Races Seven chromosome races were encountered: 2n = 44, 2n = 42a, 2n = 42b, 2n = 42c,
2n = 40a, 2n = 40b and 2n = 38. A description of the karyotypes and their distribution follows:
Table 1. Localities of specimens of Gehyra australis analysed
2n No. of specimens Type of variant d Imm. 9
100 km E. of Mt Isa, Qld 42b 1 - 3 1 (Het. C.F. 2n=41) 17 km S. of Dismal Crossing, Qld 426 1 - 2 55 km NW. of Mt Isa, qld 42b 1 1 1 36 km E, of Mt Isa, Qld 42b - 1 - 1 (1 Het. C.F. 2n=41) 5 km NW. of Dajarra, Qld 42b 3 - 1 4 km W, of Cloncurry, Qld 42b 1 1 - 1 (Het. C.F. 2n =41) 26 km S. of Dajarra, Qld 42b - - 1 38 km E. of Georgetown, Qld 42c 1 - - 7 km E. of Croydon, Qld 42c - - 1 Macquarie Marshes, NXW. 42c - 1 I 47 km W. of Mt Garnet, Qld 42c - - 1 Lizard I., Qld 42c 1 - - Edward River Mission, Qld 42c 1 - -
Pilliga Scrub, NXW. 42c - - 1 Townsville, Qld 42c 1 1 - 1 (Het. P. inv. 2n =41) Lions Den Hotel, Helenvale, Qld 42c 1 - 1 Hewcamp Creek, Qld 42c - 1 -
2.3 km W. Horseshoe Bay, Magnetic I., Qld 42c - 1 1 Picnic Bay, Magnetic I., Qld 42c - 1 -
Grosvenor Creek, Qld 42c - 1 1 7 km N. of Ellis Reach, Cairns, Qld 42c - - 1 Batavia Downs, Qld 42c 1 - 1 Mapoon Mission, Qld 42c - - 2 Urapunga, N.T. 42a 1 - -
Cannon Hill, N.T. 42a 3 - - Nourlangie Rock, N.T. 42a 1 - -
Bullo River, N.T. 42a - - 1 1 (Het. C.F. 2n=41) Manning Gorge, W.A. 44 1 - -
32 km E. of Napier Downs turnoff, W.A. 44 1 - - Barnett Range, W.A. 44 - 1 -
37 km W. of Balbirini Jn, N.T. 38 7 - 1 1 (Het. C.F. 2n = 37) 4 km S. of Balbirini Jn, N.T. 38 1 - - Dilldoll Rock 40b 1 - 1 Bing Bong HS., N.T. 40a 1 - - Kununurra Hotel, W.A. 40a 1 - 1 Timber Creek Inn, N.T. 40a 2 - 1 Liverpool R., N.T. 40a - - 1 1 (Het. C.F. 2n= 39) Lammah Hotel, N.T. 40a 1 - -
Berrimah CSIRO, N.T. 40a 4 - 4 Z W Het. Nourlangie Camp, N.T. 40a 1 - 2 Z W Het. Yirrkala, Gove, N.T. 40a 1 - -
Total Grand total, 81
2n = 44. Specimens from this chromosome race appear to be rock-dwelling, and are restricted to the central Kimberley district of Western Australia. Their distribution is bounded by G, xenopus Storr on the west, and by the 2n = 40a race to the east. Only three specimens from this chromosome race were examined, all of which had 2n =44, with a basically acrocentric karyotype except for pairs 14 and 17, which were metacentric and sub- metacentric respectively. A secondary constriction was often observed on chromosome pair 10 (Fig. 2).

Max King
2n=42a. This chromosome race was recently described as Gehyra pamela on morphological grounds (King 1982b). Specimens with this karyomorph were first thought to be restricted to the Arnhem Land escarpment. However, it is now clear that this chromosome race has a wider distribution, occurring in isolated populations as far south as the Roper River and as far west as the Bullo River (Fig. 1 ) . This is also an essentially rock-dwelling form. The karyotype is acrocentric, although pair 17 appears to be submeta- centric in some preparations; however, it is not certain if this is a product of resolution or a polymorphism. Size diminishes gradually from the largest to the smallest elements (King 1982b, fig. 3). The short arms on the larger chromosomes appear to be shorter than those in the 2n = 44 karyotype. Sex chromosomes were not detected.
2n = 42b. A large number of specimens from this rock-dwelling chromosome race have been analysed. Specimens are restricted in their distribution to the mountain ranges in central western Queensland including the towns of Mt Isa, Cloncurry, Dismal Crossing and Dajana (Fig. 1) . These animals are morphologically very distinctive and appear to be allopatrically distributed. Rock outcrops surrounding the above-mentioned sites are inhabited by small specimens of Gehyra variegata (King 1979). The karyotype of this chromosome race is very similar to that of the 2n = 42a type, being basically acrocentric but for pair 17 which is submetacentric. In some cells the centromeric areas on the larger elements appear to be achromatic (Fig. 3). Heteromorphic sex chromosomes were not observed.
2n = 42c. This chromosome race is very widespread and animals can live on either rocks, trees, or human habitations. It is distributed from the tip of Cape York peninsula in Queensland, south to just over the Queensland border with New South Wales. It also reaches the Gulf of Carpentaria in the west (Fig. 1) . The karyotype is acrocentric but for the metacentric pair 14 and submetacentric 17. Pair 3 is acrocentric, but has a pronounced short arm and appears to be the product ofa fusion between pairs 5 and 22 ofthe 2n = 44 karyotype (Fig. 4). A distinct secondary constriction is present on chromosome pair 10. No morphologically recognizable sex chromosomes were encountered.
2n = 40a. Specimens from this widespread chromosome race live on trees or human habitations. They are distributed from Lake Argyle in Western Australia, in a broad band eastward to Bing Bong station, at the base of the Gulf of Carpentaria (Fig. 1). The chromosomes are acrocentric with a gradual diminution in size from the largest to the smallest elements (King 1982b, fig. 2). Sex chromosomes were present in several populations and will be more fully described below.
2n = 40b. Only two specimens from this geographically remote, rock-dwelling Gehyra were collected. The specimens were found on an isolated rock outcrop at the base ofthe Gulf of Carpentaria (Fig. 1). In both cases the karyotype has 2n = 40 chromosomes, the first pair ofwhich is submetacentric and appear to be the product of a fusion between pairs 5 and 19 of a 2n=42 ancestral karyotype. The remainder of the karyotype is acrocentric (Fig. 5). Sex-related differences between chromosomes were not found.
2n = 38. This rock-dwelling chromosome race is also geographically restricted, occurring in the outcrops and mountain ranges associated with the MacArthur River system at the base of the Gulf of Carpentaria (Fig. 1 ) . The karyotype has 2n = 38 elements, all of which are acrocentric but for pairs 5 and 8. Pair 17 is submetacentric (Fig. 6). Chromosome pair 5 appears to be the product of a fusion between pairs 6 and 18 of an ancestral 2n=42 karyotype, whereas pair 8 appear to be the product of a fusion between pairs 16 and 20 in a similar ancestral karyotype. The submetacentric pair 17 is a marker chromosome found in this race and in the 2n = 44, 2n = 42b and 2n = 42c chromosome races. No sex-correlated chromosome heteromorphism was observed.

Karyotypic Evolution in Gehyra. 111
Fig. the Fig. pair
2. The 2n = 44 chromosome race karyotype. This complement is very similar to that of members G. variegata-punctata complex (King 1979). 3. The 2n = 42b chromosome race karyotype. Note the achromatic centromeric areas (arrowed)
.s 1 and 2.

Max IOng
Fig. 4. The 2n = 42c chromosome race karyotype. Note chromosome pair 3 (underlined) which is a presumed fusion product. The pair 10 secondary constriction is arrowed. Fig. 5. The 2n = 40b chromosome race karyotype. Note the submetacentric pair 1 (underlined) which is a presumed fusion product.

Karyotypic Evolution in Gehyra. 111
Fig. 6. The 2n = 38 chromosome race karyotype. Note the metacentric pairs 5 and 8 (underlined) which are presumed fusion products. Fig. 7. A 2n = 42c karyotype ofan animal heteromorphic for a pair 7 pericentric inversion (underlined), The pair 10 secondary constrictions are arrowed.

Max King
Variants Within Chromosome Races This variation takes two forms. First, in the 2n = 40a race certain populations had a
ZZ/Z W sex chromosome heteromorphism. This was more fully described in King (1 977b) and is characterized by a large W-chromosome in female specimens. The Z-chromosome is typically acrocentric, whereas the Wis larger and submetacentric. The short arm ofthe Wis C-band positive and appears to represent additional material. The male karyotype, which is indistinguishable from other members of the 2n=40a race, has normal acrocentric Z-chromosomes (pair 1).
The second form of variation is quite striking and is relatively frequent. A series of seven specimens from different chromosome races were heterozygous for previously undetected chromosome rearrangements. Six of these animals were heterozygous for chromosome fusions and one for a pericentric inversion. In each case a different size group of chromosome was involved in the rearrangement (see Figs 7-9); this was observed in four different chromosome races, namely 2n = 42b, 2n = 42c, 2n = 40a and 2n = 38. Three of these novel rearrangements were seen at disparate localities in the 2n = 42b race, two in the 2n = 40a race, one in the 2n = 42crace and one in the 2n = 38 race. Only heterozygotes for the changes, and standard homozygotes, were observed. Homozygotes of the derived form were not encountered and different chromosomes appeared to be implicated in each fusion. Although samples were small in most instances, a series of eight animals from a population of the 2n=38 race on a small isolated rock outcrop were scored, one of which was heterozygous for a chromosome fusion (2n = 37), all others being normal (Fig. 8).
Novel rearrangements have also been reported in G. pilbara and in the G. variegata- punctata complex (King 1979; and Table 2).
Table 2. Numbers of specimens of Gehyra species karyotyped, and the degree of chromosomal divergence
Taxon Yo. of specimens No. of chromo- No. of novel Reference karyotyped some races rearrangements
G. variegata- punctata 250
G. pilbara 6 G, austrahs 8 1
King 1979: 6 unpublished 1 Unpublished 7 This paper
Discussion
Chromosome Evolution (i) Chromosome race differentiation
The predominant means of gross chromosomal rearrangement observed in most vertebrate species has involved the evolutionary increase or decrease in chromosome number. This pattern has been seen in organisms as diverse as the marsupials (Hayman and Martin 1974), rodents (Matthey 1973; Capanna et al. 1977; Baverstock et al. 1979) and crocodiles (Cohen and Gans 1970). In the majority of these cases, change in number is the result of chromosome fusion or fission.
In the lizard species thus far karyotyped both fusion and fission have also been used to explain many of the chromosome number differences described. Thus, in the North American iguanid genus Sceloporus, Webster et al. (1 972), Hall and Selander (1 973) and Paul et al. (1 976) have all argued for chromosome change based on fission, although presumed instances of chromosome fusion have also been postulated. In contrast, chromosome fusion has undoubtedly been the predominant mode ofkaryotypic change in the chromosome races of gekkonids of the genera Hemidactylus, Phyllodactylus, Diplodactylus and Gehyra analysed by King (summary in King 198 la).
The criteria used for assessing the direction of change in these studies have been based on extensive karyotypic surveys, in which the most common karyotype was regarded as the

Karyotypic Evolution in Gehyra. 111
Fig. 8. A 2n = 37 karyotype of an individual from the 2n = 38 race heteromorphic for a centric fusion (see elements involved underlined). Fig. 9. Cells from animals heteromorphic for separate centric fusions (see fusion products arrowed): (a) 2n = 40a race, het 2n = 39; (b) 2n = 42b race, het 2n = 41; (c) 2n = 38 race, het 2n = 37; (4 2n = 42b race, het 2n = 41.

Tab
le 3
. C
hrom
osom
e m
arke
rs fo
und
in a
ser
ies
of G
ehyr
a sp
ecie
s
Spec
ies
Rac
e M
arke
rs
Fus
ion
prod
ucts
10
sc
Met
a 14
M
eta1
7'
'5 7
3 6
R 3'
2'
5' 8'
\
G'. v
aneg
ata-
punc
tata
2n
= 44
2n
= 40
b 2n
= 40
a 2n
= 38
2n
= 4
2
G. p
ilbar
a 2
n=
44
G. a
ustr
alis

Karyotypic Evolution in Gehyra. 111
ancestral form. The practice of using the dominant karyotype may be open to criticism ifthis is to be the sole criterion. However, additional supporting information such as the phylogenies produced by electrophoretic or morphological analyses can assist in determining the direction of change.
In many of the cases where fusion has been proposed as the means of karyotypic reorganization the fusions in question have occurred between differing, though closely related chromosome races, and in a series of genera. In all these instances a number of different chromosomes have been involved in the rearrangements. If the changes were not fusion but regarded as fission, and if evolution had gone in the direction of metacentricity to acrocentricity, all of the most closely related species complexes would necessarily have had a polyphyletic origin. This is clearly unacceptable when all other criteria are considered (King 1981a, 1982b).
In the genus Gehyra, King (1979) postulated that the 2n = 44 complement was the extant ancestral karyomorph for this group. This karyotype has a series of chromosome markers, viz. metacentric pairs 14 and 17, a secondary constriction on chromosome pair 10, and different short-arm lengths in acrocentric pairs 1-5. Such a karyotype morphology has been observed in Gehyra nana (King 1979), G. pilbara (King, unpublished), a number of forms of the G. variegata-punctata complex (King 1979) and, in the present study, in the 2n=44 chromosome race of Gehyra australis from the Kimberley in Western Australia. In the G. variegata-punctata complex, derived races showed evidence of chromosome fusion within this ancestral format. That is, additional metacentric chromosomes are present (fusion products) accompanying the reduction in chromosome number (see Table 3 for summary).
Ifwe now turn to the chromosome races of G. australis described in this Daver we also see A -
evidence of chromosome fusion being involved in the derivation of particular races. Thus the 2n=42c race has chromosome markers 14, 17 and the secondary constriction on 10 shared with the 2n = 44 karyotype. An increased arm length in pair 3 (a 5/22 fusion product) accompanies the reduction in chromosome number to 2n = 42. Similarly, the 2n = 40b race has an acrocentric karyotype, except for pair 1 which is a presumed 5/19 fusion product evolving from a 2n = 42 karyotype. The chromosome markers 14 and 17 are not visible in this karyotype and may have been inverted to an acrocentric condiiion. The site of the secondary constriction is uncertain. The last example of clear fusion products is seen in the 2n = 38 race. Here, pairs 5 (6/ 18 fusion) and 8 (1 6/20 fusion) have also presumably evolved from an ancestral 2n = 42 karyotype. In this case the pair 17 marker is still present, but the pair 14 metacentrics have been inverted to acrocentrics and the position of the 10 secondary constriction is uncertain.
Although such changes are relatively straightforward, derivation of the 2n=40a karyotype can be most parsimoniously achieved only by either the loss or the incorporation of a pair of microchromosomes (i.e. by mechanisms such as tandem fusion) of the 2n = 42a type from which it probably evolved. Similarly, the 2n = 42b race could have evolved from the 2n = 42c karyotype by a series of pericentric inversions reducing the size of the short arms of pairs 1-5 and making pair 14 acrocentric. The 2n = 42a and 2n = 42b chromosome races are very similar in their chromosome morphology.
The most parsimonious phylogenetic sequence of events, based on the assumption of centric fusion and a series of simple pericentric inversions, involves the following steps (Fig. 10):
(1) Ancestral 2n = 44 race; (2) 2rt = 42c race derived from the 2n = 44 ancestral form by a 5/22 fusion; (3) 2n = 42a and 2n = 42b ancestral races produced by inversions of the 1-5 group and
the inversion of pair 14 of 2n = 42c race; (4) 2n = 42a and 2n = 42b races differentiated morphologically yet remained chrom-
osomally similar;

Max King
(5) 2n = 40a race produced by the loss of a microchromosome pair of the 2n = 42a race;
(6) 2n = 40b race produced by a 5/19 fusion of the 2n = 42b race and an inversion of the metacentric pair 17 to an acrocentric;
(7) 2n = 38 race produced by 6/18 and 16/20 fusions of 2n = 42a race. Clearly, a detailed description of this series of changes cannot be provided from the
present study. It is possible that a G-banding analysis might provide a more definitive inter- pretation of the evolution of this complex, and such an analysis is being currently undertaken.
Fig. 10. A phylogeny showing the relationship of the chromosome races within the G, australis complex, and the relationship to other Gehyra so far examined.
(ii) Novel rearrangements Chromosome polymorphism involving fusion, fission, pericentric inversion and the
addition of supernumerary segments has now been recorded in a number of lizard species. Cole (1970, 1977) has identified locality-specific polymorphisms, involving inversions and addition, in Sceloporus clarkii, S. melanorhinus and S. undulatus. Other iguanid species showing polymorphisms are S. grammicus (Hall and Selander 1973), Tropidurus torquatus Becak et al. (1972) and Anolis monticola (Webster et al. 1972). The varanid Varanus acanthurus also has a geographically delimited pericentric inversion polymorphism (King et al. 1982).
There are a number of situations described in the literature, where there is some uncertainty whether a specimen heterozygous for a chromosomal rearrangement is a rare mutant animal, or forms part of a balanced polymorphism. Olmo and Odierna (1980) encountered a specimen of Cordylidae, Cordylus cataphractus, heterozygous for an apparent fission. Similarly, King (1 98 1 a) described a specimen of Phyllodactylus lanei heteromorphic for a chromosome fusion. In both cases too few specimens were available to determine the nature of the variation. In the G. variegata-punctata complex (King 1979), three specimens heterozygous for presumed pericentric inversions were described. In each ofthese animals a

Karyotypic Evolution in Gehyra. I11
different chromosome was implicated in the change and only heterozygotes were observed. If an inversion polymorphism was involved, its frequency must have been very low.
In the G. australis complex we see a relatively high incidence of animals heterozygous for novel rearrangements (7 in 81), yet different chromosomes appeared to be involved in the rearrangements in each case. Indeed, although it is true that chromosome polymorphisms involving fusion are not a common phenomenon, once again the sample sizes are not large enough to eliminate the possibility that these are locality-specific polymorphisms.
Chromosome Race Distribution and the Mode of Speciation
The most striking feature of the distributions of the chromosome races is their relative integrity. Each of these forms is allopatrically distributed, either geographically or in habitat preference. Additionally, a number of the distributions are restricted to major land forms. Thus, the 2n = 44 race occurs in the mountain ranges of the central Kimberley in Western Australia, the 2n = 38 race is restricted to the ranges surrounding the Macarthur River at the base of the Gulf of Carpentaria, the 2n = 42b race is present in the mountain ranges in and around Mt Isa in central Queensland, and the 2n = 42a race occupies the Arnhem Land escarpment and certain surrounding ranges. All of these forms are rock-dwelling and do not appear to have the capacity to live in an arboreal habitat. Indeed, the extent of the distributions of several of these chromosome races is directly proportional to the area of mountain range or rock outcrop available. The validity of this argument is strengthened by the distributions of the less specialized forms, which can readily live on trees and human habitations (2n = 40a, 2n= 42c) and in some cases also on rocks (2n = 42c). In these instances the distributions are enormous, the 2n = 42c race occupying a wide area from the tip of Cape York peninsula to south of the Queensland border. Similarly, the 2n = 40a race occurs from Western Australia to the middle of the Gulf of Carpentaria. This distribution is of particular interest, because it overlaps the areas occupied by both the 2n=38 and 2n = 42a races. Nevertheless, the different forms remain allopatric due to the arboreal preference of the 2n = 40a race and the rock-dwelling habitat of the others.
Although the presence of chromosome races on major geographic features may reflect a requirement for a certain habitat type, this is not necessarily so. The terrestrial gekko Diplodactylus vittatus has a 2n=34 fusion race restricted to a region including the Hinders-Mt Lofty ranges in South Australia (King 19774. This gekko does not require a montane habitat since it also occurs in adjacent mallee country. It is likely that the mountain ranges provided a radiation pathway during favourable conditions and a refugium during less favourable periods.
The complicated central Australian distribution of the G. vanegata-punctata race complex (King 1979) may be similarly interpreted. Here a series of chromosome races occur on isolated rock outcrops and on ranges surrounded by sandy desert. The distributions are erratic and do not always reflect a proximity to direct ancestors. It was proposed that the present-day distribution is a relic of past radiations, extinctions and recolonization; the outcrops here also being refugia. In other areas of Australia the chromosome races of G. vanegata-punctata are also restricted to major land forms or habitat types. For example, the 2n=40a race is found on the Murray-Darling drainage system in south-eastern Australia, whereas the 2n = 44 form of G. varzegata occurs in the Flinders-Mt Lofty ranges in South Australia, occupying an area very similar to that of the 2n=34 form of D. vittatus.
In the G. australis complex, certain of the derived races are distributed in relatively close proximity to their presumed ancestors. Thus, the 2n = 40b race is adjacent to the 2n = 42b race, and the latter is adjacent to the 2n = 42c race. Similarly, the 2n = 42a race is adjacent to the 2n = 40a race, and the 2n = 38 race is adjacent to the 2n = 42a race. If these distributions are compared to the proposed phylogeny (see Fig. lo), it is clear that the evolution of this complex could be interpreted as a series of colonizing radiations, during which the

Max King
individual chromosome races were established, i.e, primary chromosomal allopatry (sensu King 1981~).
Such an explanation can indeed be applied to the more recent chromosomal changes. However, the distributions of the 2n = 44 and 2n = 42c races, which are chromosomally most similar to each other, presents a major problem. These putatively closest relatives are furthest apart in their distributions; the 2n=44 is in the Kimberley district of Western Australia and the 2n= 42c on the east coast of Australia. For the 2n= 42c race to have evolved from the 2n = 44 ancestral karyotype form, one might have expected a relatively close proximity. The most feasible explanation for the present-day distribution is that the 2n = 44 ancestral form, from which the 2n = 42c evolved, once occurred in eastern Australia but is now extinct, the 2n = 44 race found in the Kimberley being a relic of a form which formerly was widespread across the top of northern Australia. This form may have been eventually restricted to the west, either through being eliminated throughout the eastern and central sections of its range by environmental changes, or through displacement by other chromosome races [i.e. stasipatry, sensu White (1978)l.
The concept ofan east-to-west distribution of a 2n = 44 ancestral G. australis in northern Australia is to some extent supported by the present-day distribution of G. nana. This species is sympatric with the 2n= 44 form of G. australis and also has 2n= 44. G. nana is distributed across most of northern Australia except for an area in the vicinity of the Gulf of Carpentaria. Consequently, its east-to-west distribution must at one time have been continuous, the two isolates being formed by elimination of intervening populations (King 1981b).
The two chromosomally closest 2n = 42 races of G. australis are also isolated on the east and west of the Gulfofcarpentaria. It therefore appears likely that the 2n = 42a and 2n = 42b races may have once been contiguous, but that this distributional continuity has been disrupted by some environmental change which left isolates in central Queensland and the Northern Territory. The 2n = 38,40a and 40b races today occupy the intermediate areas of this once extensive distribution. Moreover, the fact that these chromosomally most highly modified, and hence recently derived forms, occupy the central area at the base of the Gulf of Carpentaria, suggests that the colonization of this area has been a relatively recent event.
Clearly, this distributional sequence, if interpreted as a direct reflection of the speciation process, would support a model of stasipatric speciation. That is, the chromosomally derived forms occupy the central areas, whereas the ancestral forms occupy peripheral areas. The derived forms could therefore be seen as actively displacing the parental forms (see White 1978). However, we know that in a number of periods in northern Australia's geological history the eastern and western sections of the continent were isolated by repeated episodes of environmental devastation. Over the past 250 000 years major fluctuations in sea level are known to have occurred in northern Australia (Chappell and Thom 1977). Over a longer time scale (i.e. the Pliocene to Recent) increased aridity has induced significant vegetational changes in this area (Gill 1975). Even the use of fire by aboriginal man over the past 40 000 years may have been a significant agent in modifying faunal distributions.
It is, therefore, not surprising that the region which appears to have undergone a series of population extinctions is situated at the base of the Gulf of Carpentaria. Today, this is one of the more environmentally fragile areas. A narrow belt of rock outcrops extends from east to west. This is bordered on one side by the flat coastal scrubland extending to the Gulf, and on the other by the treeless Savannah Grasslands of the Barkly Tablelands. Both of these areas are prone to fire and partial inundation. Major environmental fluctuations of the type outlined above could be expected to have readily eliminated populations from this region. Taxa which were previously distributed continuously from west to east may therefore have been eliminated over a large part of their ranges, leaving eastern and western isolates. Subsequent periods may have produced a favourable habitat in which population radiations could once again successfully occur. That is, speciation processes involving the parental

Karyotypic Evolution in Gehyra. 111
species being displaced by chromosomally derived forms (stasipatry) are much less likely than that the chromosomally derived forms evolved during population radiations into essentially uninhabited areas (primary chromosomal allopatry).
That the area at the base of the Gulf of Carpentaria has produced a series of population extinctions is seen in the present-day distribution of certain northern Australian reptiles. For many species a narrow corridor across the base of the Gulf joins eastern and western populations (see Cogger 1975). Most significantly, some 14 species show a complete break in this comdor, leaving eastern and western isolates of each species (see Table 4 for examples).
Serpentes
Table 4. Species of reptiles in northern Australia separated into eastern and western isolates by the Gulf of Carpentaria
Family Species
Sauria Scincidae Carlia fusca (Dumeril & Bibron) Egernia frerei Gunther Sphenomorphus crassicaudus (Dumeril)
Varanidae Varanus indicus ( ~ a u d i n ) ~ Varanus semiremex petersA
Gekkonidae Gehyra nana (Storr) Typhlopidae Typhlina aftinis ( ~ o u l e n ~ e r ) ~
Typhlina wiedii ( ~ e t e r s ) ~ Colubridae Amphiesma mair i~ (Gray)
Dendrelaphis punctulatus (Gray) Enhydris polylepis (Fischer) Stegonotus cucullatus (Dumeril, Bibron & Durneril)
Elapidae Demansia atra (Macleay) Oxyuranus scutellatus (Peters) Simoselaps sem&sciatus (Gunther)
A ~ e c a u s e of specialised habitat or lifestyle the absence of these species from the Gulf region may be due to inadequate collection.
A possible evolutionary sequence of events leading to the diversity and distributions that we see today, and based on the most parsimonious phylogenetic model, follows:
(1) The 2n = 44 ancestral Gehyra australis radiated from west to east across the north of Australia;
(2) In the course of this radiation the 2n=42c race was established and this, in turn, radiated along the east coast of Australia occupying much of its present-day distribution;
(3) Environmental changes in the Gulf area eliminated the 2n = 44 race from the central part of its distribution, leaving an isolate in Western Australia;
(4) A population radiation from east to west originated on the periphery of the 2n = 42c race. The chromosomally modified forms were of the ancestral 2n = 42a and 2n = 42b types and occupied the area between the 2n=42c and 2n=44 races;
(5) Environmental changes affecting the Gulf area eliminated these ancestral 2n=42 forms from the central part of their distribution, leaving an uninhabited void, and producing eastern and western isolates, i.e. the 2n = 42a and 2n = 42b races;
( 6 ) Subsequent, more recent population radiations of chromosomally derived forms (2n =40b and 2n = 38) occurred in the Gulf area, radiating from the 2n = 42b and 2n = 42a races respectively;
(7) The 2n=40a race arose on the periphery of distribution of the 2n=42a race, coinciding with a change in habitat preference. This race then radiated throughout the north of the Northern Temtory and into the Gulf area.
This sequence of events is summarized in Fig. 11.

Max King
Taxonomic Implications (i) Specijc relationships
It is now clear that, in gekkos at least, chromosome analysis at a population level can be of considerable taxonomic value. Taxonomically 'awkward' species can be unambiguously categorized into chromosomal races on the basis of karyotypes. Morphological comparisons within and between these chromosome races may then provide diagnostic characteristics permitting a taxonomic revision.
Fig. 11. A presumed sequential series of chromosome race distributions producing the present-day distribution. Arrows indicate chromosome race radiations. Maps B and D show distributions during populations extinction in the area at the base of the Gulf of Carpentaria.
Such an approach has been successfully used in the Diplodactylus vittatus complex, where five allopatrically distributed chromosome races and their morphological difference were described by King (1 977a). Subsequently, Storr (1 979) was able to describe a series of new species and resurrect a number of older taxa, producing a complex of five species. Similarly, Gehyra pamela was recently described as a new species after a chromosomal and morphometric analysis showed it to be clearly distinguishable from G. australis (King 1982b).

Karyotypic Evolution in Gehyra. I11
One ofthe major problems with Gehyra specimens lodged in museums is that coloration and back pattern fade with the passage of time. In life, the variation in colouring and back pattern is spectacular, and in most cases is directly correlated with chromosome race distribution. In this group it is thus important to observe live specimens. A second difficulty which has hampered traditional taxonomists is that, in groups such as the G. australis complex, where many undescribed species are present, too few specimens from too few localities have been examined. Diagnostic characters have therefore been overlooked, and the 'species' is simply regarded as being highly variable (see Mitchell 1965).
There is good reason to believe that the majority of chromosome races in the G. australis complex are good biological species. Specimens from each race are chromosomally and morphologically consistent throughout their range. There is no evidence of introgression of one form into another when distributions are sympatric or when allopatric races appear to abut. The major problem at the moment is that too few specimens have been analysed in certain of the chromosome races (such as 2n=40b) to make a morphometric analysis. Such an analysis has been performed on the 2n =40a, 2n =42a, 2n= 42b, 2n = 42c and 2n = 38 chromosome races, and all show significant morphological differences. A taxonomic revision of this complex of species has been made and will be published separately (King 1983).
(ii) The relationship between the G. australis and G, variegata-punctata complexes In his osteological and morphological study of Australian Gehyra species, Mitchell (1 965)
suggested that there were two species-groups within Australia. The first of these was an Indo-Australian group containing G. australzs and G. oceanica (the status of the latter being uncertain in Australia). The second contained a series of endemic Australian forms recognized at that time: G. varzegata, G. pzlbara, G. punctata and G. fenestra. The basis of this dichotomy was the sternal rib structure: the Indo-Australian group having two sternal ribs, the endemic group having three. Mitchell (1 965) felt that this provided strong evidence for dividing Gehyra into two genera: an endemic Australian genus Dactyloperus Fitzinger, 1843, and an Indo-Australian genus Gehyra Gray, sensu stricto. A final decision on this matter was deferred.
The chromosomal data, however, suggest that the Australian Gehyra are a continuum of genetically related forms. In both the G. varzegata-punctata complex (King 1979) and the G. australis complex particular chromosome markers and ancestral karyotypes are shared. This indicates that the G. australis complex is part of an endemic Australian group rather than an Indo-Australian group, and that it is intrinsically related to other Australian Gehyra. Indeed, both the chromosome data and the consequent phylogeny suggest that the G. australis complex has evolved from an ancestral 2n=44 state common to both the G. australis and G. variegata-punctata complexes. The relative, evolutionary independence of the complexes since this initial divergence is no more than evidence for a dichotomy into species-groups. These data in no way support a generic subdivision into endemic and non-endemic forms, but suggest that in reality both complexes are endemic and belong to one genus.
Mitchell's proposed generic subdivision of Australian Gehyra was based on only six cleared specimens of G. australis. The distribution data provided by Mitchell (1 965) suggest that these specimens were from just two of the seven known chromosome races. Clearly, a detailed osteological analysis of all the forms of G. australis and of the more recently described Gehyra species is necessary to establish the sternal rib structure, before the universality of this structural dichotomy can be accepted.
Acknowledgments
The author is grateful to the many people who helped in this study collecting specimens, in particular Pamela King, Dennis King, Robert Jenkins, Gregory Mengden, John Wombey,

Max King
Hal Cogger and Craig Moritz. Thanks to Bernard John for his helpful comments on the manuscript.
References Baverstock, P. R., Watts, C. H. S., Hogarth, J. T., Robinson, A. C., and Robinson, J. F. (1979). Chromosome evolution in Australian rodents. 11. The Rattus group. Chromosoma (Berl.) 61, 227-41. Becak, M. L., Becak, W., and Denaro, L. (1972). Chromosome polymorphism, geographic variation and karyotypes in Sauria. Caryologia 25, 3 13-26, Capanna, E., Civitelli, M. V., and Cristaldi, M. (1977). Chromosomal rearrangements, reproductive isolation and speciation in mammals. The case of Mus musculus. Boll. Zool. 44, 213-46. Chappell, J., and Thom, B. G. (1977). Sea levels and coasts. In 'Sunda and Sahul'. (Eds J. Allen, J. Golson and R. Jones.) pp. 275-91. (Academic Press: New York.) Cogger, H. G. (1975). 'Reptiles and Amphibians of Australia.' (A. H. and A. W. Reed: Sydney.) Cohen, M. M., and Gans, C. (1970). The chromosomes of the order Crocodilia. Cytogenetics (Basel) 9, 81-105. Cole, C. J. (1970). Karyotypes and evolution of the Spinosus group of lizards in the genus Sceloporus. Am. Mus. Novit. No. 2431. Cole, C. J. (1977). Chromosomal aberration and chromatid exchange in the North American fence lizard, Sceloporus undulatus (Repti1ia:Iguanidae). Copeia 1977, 53-9. Gill, E. D. (1975). Evolution of Australia's unique flora and fauna in relation to the plate tectonics theory. Proc. R. Soc. Victoria 87, 215-34. Gorman, G. C. (1973). The chromosomes of the Reptilia, a cytotaxonomic interpretation. In 'Cytotaxonomy and Vertebrate Evolution'. (Eds A. B. Chiarelli and E. Capanna.) pp. 349-429. (Academic Press: New York.) Hayman, D. L., and Martin, P. (1974). Mammalia I: Monotremata and Marsupialia. In 'Animal Cytogenetics'. Vol. 4, Chordata 4. (Gebriider Borntraeger: Berlin.) Hall, W. P., and Selander, R. K. (1973). Hybridization of karyotypically differentiated populations in the Sceloporus grammicus complex (Iguanidae). Evolution 27, 226-42. King, M. (1977~). Chromosomal and morphometric variation in the gekko Diplodactylus vittatus (Gray). Aust. J. Zool. 25, 43-57. King, M. (1977b). The evolution of sex chromosomes in lizards. In 'Evolution and Reproduction'. Proc. 4th Int. Conf. Reprod. Evol. pp. 55-60. (Australian Academy of Science.) King, M. (1978). A new chromosome form of Hemidactylus frenatus (Dumeril and Bibron). Herpetologica 34, 2 16-1 8. King, M. (1979). Karyotypic evolution in Gehyra (Gekk0nidae:Reptilia). I. The Gehyra variegata- punctata complex. Aust. J. Zool. 27, 373-93. King, M. (1981~). Chromosome change and speciation in lizards. In 'Evolution and Speciation: essays in Honor of M. J. D. White'. (Eds W. R. Atchley and D. Woodruff.) pp. 262-85. (Cambridge University Press.) King, M. (1981b). Notes on the distribution of Gehyra nana Storr and Gehyra punctata (Fry) in Australia. Aust. J. Herpetol. 2, 1-5. King, M. (1982~) . A case for simultaneous multiple chromosome rearrangements. Genetica (The Hague) 59, 53-60. King, M. (19823). Karyotypic evolution in Gehyra (Gekk0nidae:Reptilia). 11. A new species from the Alligator Rivers Region in Northern Australia. Aust. J. Zool. 30, 93-101. King, M. (1983). The Gehyra australis species complex. Amphibia-Reptilia. (In press.) King, M., and King, D. (1977). An additional chromosome race of Phylodactylus marmoratus (Gray) (Repti1ia:Gekkonidae) and its phytogenetic implications. Aust. J. Zool. 25, 667-72. King, M., Mengden, G. A., and King, D. (1982). A pericentric inversion polymorphism and a ZUZW sex chromosome system in Varanus acanthurus Boulenger analysed by G and C banding and silver staining. Genetica (The Hague) 58, 39-45. King, M., and Rofe, R. (1 976). Karyotypic variation in the Australian gekko Phyllodactylus marmoratus (Gray) (Gekk0nidae:Reptilia). Chromosoma (Berl.) 54, 75-87. Matthey, R. R. (1973). The chromosome formulae of eutherian mammals. In 'Cytotaxonomy and Vertebrate Evolution'. (Eds A. B. Chiarelli and E. Capanna.) pp. 530-616. (Academic Press: New York.)

Karyotypic Evolution in Gehyra. I11
Mitchell, F. J. (1965). Australian geckos assigned to the genus Gehyra Gray (Reptilia, Gekkonidae). Senckenb. Blol. 46, 287-3 19. Olmo, E., and Odierna, G. (1 980). Chromosomal evolution and DNA of cordylid lizards. Herpetologrca 4, 311-15. Paul, D., Williams, E. E., and Hall, W. P. (1976). Lizard karyotypes from the Galapagos Islands: chromosomes in phylogeny and evolution. Brevlora 441, 1-3 1 . Storr, G. M. (1 979). The D~plodactylus vlttatus complex (Lacerti1ia:Gekkonidae) in Western Australia. Rec. West. Aust. Mus. 7, 391-402. Webster, T. P., Hall, W. P., and Williams, E. E. (1972). Fission in the evolution of a lizard karyotype. Sclence (Wash. D.C.) 177, 61 1-1 3. White, M. J. D. (1978). 'Modes of Speciation.' (W. H. Freeman: San Francisco.)
Manuscript received 31 May 1982; accepted 2 1 January 1983