la adhesión es un fenómeno esencial durante la fertilización · 2018-09-07 · la adhesión es...
TRANSCRIPT

Wassarman et al, Nature Cell Biol 2001; Prymakoff & Myles, Science 2002; Vjujina & Evans Front Biosci 2008; Satouh & Ikawa TIBS 2018
La adhesión es un fenómenoesencial durante la fertilización
células
del
cúmulo
zona pelúcida
adhesión
fusión

La adhesión es esencial para mantener la cohesión del embrión en desarrollo
BMoC 2008; Damsky et al Cell 1983
mórula
mórula
+ anti-E-caderinaEl bloqueo de E-caderinas (uvomorulina) con un anticuerpo
monoclonal desesambla las adhesiones intercelulares,
inhibe la compactación y desorganiza la mórula.
compactación

CR1
CR1
Trp
Trp
El triptofano (Trp) conservado del CR1 de un
monómero interacciona con un bolsillo
hidrofóbico del CR1 de otro monómero en trans.
Bases estructurales de la adhesión mediada por caderinas
Boggon et al., Scien ce 2002;
Brasch et al TICB 2012;
adaptado en Alberts et al MBC 2008
Trp

La adhesiones proporcionan cohesión a los tejidos y son fundamentalespara la organización y arquitectura multicelular
Alberts et al BMoC 2008
filamentos
intermedios

Las adhesión es mediada por proteínas de transmembrana, que acoplan físicamente ligandos extracelulares con el citoesqueleto
Alberts et al MBC 2002
citoplasma
ligando
extracelular
citoesqueleto
Los receptores de adhesión median la transmisión de fuerzas entre la célula y su entorno.

El reconocimiento de ligandos extracelulares involucra diversos mecanismos moleculares
uniones homofílicas
Ej. caderina-caderina,
NCAM-NCAM
uniones a través de una molécula
extracelular puente. Ej.
integrinas-proteoglicanos-integrinasuniones heterofílicas
Ej. integrinas-ICAMs,
selectinas-glicanos
Alberts et al MBC 2002
Los dominios extracelulares de las moléculas de adhesión han evolucionado para reconocer
moléculas del mismo tipo (unión homofílica) o moléculas diferentes (unión heterofílica).

Lodish et al MBC 2004
Los procesos de reconocimiento y adhesión están mediados por cuatro familias de proteínas
(~115) (~700) (~26)

KNOCK OUT INTEGRINS & CADHERINS
La depleción de moléculas de adhesión provoca letalidad o severos defectos en el desarrollo, estructura y funcionamiento de los tejidos
N-cadE10, neurulation/somito
genesis defects, heart
does not form
E-cad
die at time of
implantation
N-CAM
Nervous system
defectsL1
claudin
die at birth

Lodish et al MBC 2004; Alberts et al MBC 2015
Los receptores de adhesión cumplen funciones mecánicas
Los receptores de adhesión se anclan al citoesqueleto mediante diversas proteínas
adaptadoras y de “scaffold”. Las interacciones son de naturaleza no covalentes y dinámicas.

Fuerzas transmitidas a través de las adhesiones inducen cambios conformacionales en proteínas del citoplasma
El proceso de conversión de fuerzas en señales bioquímicas y
respuestas biológicas se denomina mecanotransducción.
stretching!
Las fuerzas intracelulares
dependen de actina y miosina
La elongación de
-catenin
promueve su
interacción con
vinculina.
Lodish et al MBC 2004; Alberts et al MBC 2015

Roca-Cusachs et al JCS 2012; Wang et al Cell Molec Bioing 2016
Fuerzas generadas por las células alteran la conformación de proteínas de la matriz extracelular y del citoplasma
extended
closed
conformation
open
conformation
compact
FN modules
La elongación de talina
promueve su interacción
con vinculina.
stretching!
sitios de oligomerización
tensionado
módulo
relajado
módulo de FN III
Alberts et al MBC 2015

Chiquet et al, BBA 2009
Métodos para estudiar las respuestas mecánicas de las células
Empleo de substratos deformables (ej. silicona, poliacrilamida, etc)
célula en reposo célula ejerciendo tracción
substrato substrato
substrato substrato
Empleo de trampas ópticas
rayo lásermicroesfera
cubierta con
proteína adhesiva
filamentos
de actina
fuerza
desplazamiento
video disponible

Provenzano et al CPCB 2010; Schwarz & Gardel, JCS 2012; Paluch et al BMC Biol 2015; Alberts et al BMC2008
Métodos para estudiar las respuestas mecánicas de las células
Contracción de geles de colágeno
1 mm
Alineamiento de fibras de colágeno
Respuesta a la rigidez del substrato
mapa de fuerzas

Hynes Cell 2002
Los receptores de adhesión también activan vías de señalización intracelulares
Diversos procesos celulares fundamentales (ej. proliferación, apoptosis, etc) son regulados por la adhesión
a la matriz extracelular. La adhesión activa diversas enzimas y proteínas asociadas a los complejos de
adhesión. Esta función se realiza de manera sinérgica con otros tipos de receptores.

Diferentes estructuras y funciones de los complejos de adhesión
dendritic cell
T-cell
sinapsis inmunológica
Comunicación y señalización
neuronasepitelialesepiteliales
Anclaje
epiteliales
Barrera
Barrera (hemostasis)
malla de fibrina
colágeno
plaquetas activadas
Señalización

Diferentes estructuras y funciones de los complejos de adhesión
matriz
extracelular
(fibroblasto)
Anclaje
Migración
fotos tomadas cada 1 hora
Doyle et al, Nature Commun 2015

Los epitelios exhiben mútiples tipos de uniones intercelulares
Uniones estrechas: Constituyen una
barrera al pasaje de moléculas entre la
cara apical y basolateral.
Uniones adherentes: Anclan los
citoesqueletos contráctiles y son
esenciales durante la morfogénesis.
Desmosomas: Anclan los citoesqueletos
no contráctiles y son esenciales para
mantener la estructura del epitelio.
Uniones en hendidura: Permiten el pasaje
de iones, segundos mensajeros, y otras
moléculas pequeñas. Contribuyen a la
coordinación funcional del epitelio.
Hemidesmosomas: Anclan las células,
mediante un citoesqueleto no contráctil
(fil. intermedios), a la membrana basal
que limita con el tejido subyacente.
Adhesiones focales: Anclan las células,
mediante un citoesqueleto contráctil
(actin), a la membrana basal.
Las células epiteliales maduras se polarizan formando un dominio apical (= luminal) y uno basolateral.
(estrechas)
(adherentes y
desmosomas)
(hendidura)
(focales)

Las uniones estrechas impiden la libre difusión de moléculas a través del epitelio (transporte paracelular) y
actúan como barrera a la difusión de proteínas de membrana entre los dominios apical y basolateral.
Microscopía electrónica que ilustra la barrera formada
por las uniones estrechas. En este experimento se
incubó un epitelio con hidróxido de lantanio (opaco a los
electrones). El lantanio no pasa las uniones estrechas.
lantanio
lantanio
Uniones estrechas (Tight/Occluding junctions)
transporte
vectorial
(transcelular)
de glucosa
En el intestino la polarización de
las células permite un flujo
vectorial de nutrientes.

Visualización de uniones estrechas por microscopía
electrónica. Técnica de congelación y fractura
Al microscopio electrónico las uniones estrechas se visualizancomo una costura en la porción apical de las células epiteliales
Alberts et al MBC 2002
cara apical
cara basolateral

célula 1 célula 2
Las principales proteínas de transmembrana de las uniones estrechas son las Claudinas y Ocludinas
Los dominios extracelulares de claudinas y
ocludinas median interacciones homofílicas
independientes de calcio. Los dominios
intracelulares se anclan al citoesqueleto de
actina por medio de proteínas adaptadoras
denominadas ZO ("Zonula Occludens").
Claudinas, ocludinas, JAMs y tricelulinas son componentes de transmembrana de las uniones estrechas.
Las proteínas de “scaffold” ZO poseen varios dominios
que les permiten interaccionar con proteínas de
transmembrana, actina y otros adaptadores.
JAMs

Uniones en hendidura (Gap Junctions)
La mayoría de las células poseen uniones en hendidura. En estos complejos las membranas se
separan unos 2-4 nm (panel de la izquierda). En una vista frontal las uniones en hendidura forman
agregados de estructuras cilíndricas (panel de la derecha).
Lodish et al MBC 2004
conexones
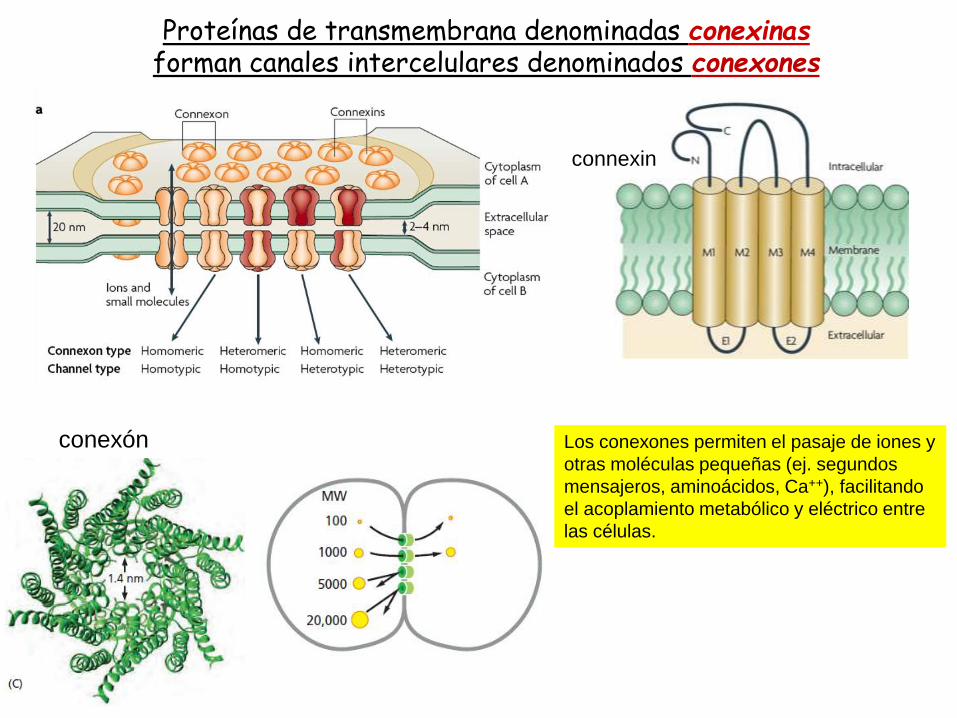
Proteínas de transmembrana denominadas conexinasforman canales intercelulares denominados conexones
Los conexones permiten el pasaje de iones y
otras moléculas pequeñas (ej. segundos
mensajeros, aminoácidos, Ca++), facilitando
el acoplamiento metabólico y eléctrico entre
las células.
connexin
conexón

Conectividad funcional
entre neuronas de la retina.
Liberación del neuro-
transmisor dopamina.
Las uniones en hendidura permiten el acople metabólico y eléctrico entre células
MBC, 2008
Inhibición de la
permeabilidad de
las uniones en
hendidura.
Inyección de un trazador
fluorescente que pasa por
las uniones en hendidura
folículo
ovárico
La permeabilidad de los conexones es regulada por varios factores como calcio, voltaje, pH.
Moléculas de cGMP sintetizado en las células foliculares
pasan a través de uniones en hendidura al ovocito e
inhiben su maduración (arresto en profase meiótica).
células foliculares

Uniones de anclaje
Las uniones de anclaje incluyen a las
uniones adherentes, desmosomas ,
hemidesmosomas y complejos focales.
Se componen de proteínas de
transmembrana que anclan los
ligandos extracelulares con el
citoesqueleto.
Alberts et al MBC 2002

Uniones adherentes (Adherens Junctions)
En epitelios, las uniones adherentes forman bandas
adhesivas (“adhesion belt”) cerca del dominio apical.
Alberts et al MBC 2002
~ 20 nm
Las proteínas de transmembrana de las uniones
adherentes son las caderinas. Los dominios
extracelulares de las caderinas de células
adyacentes interaccionan entre sí en presencia
de calcio. Los dominios intracelulares de las
caderinas se anclan a los filamentos de actina
contráctiles.
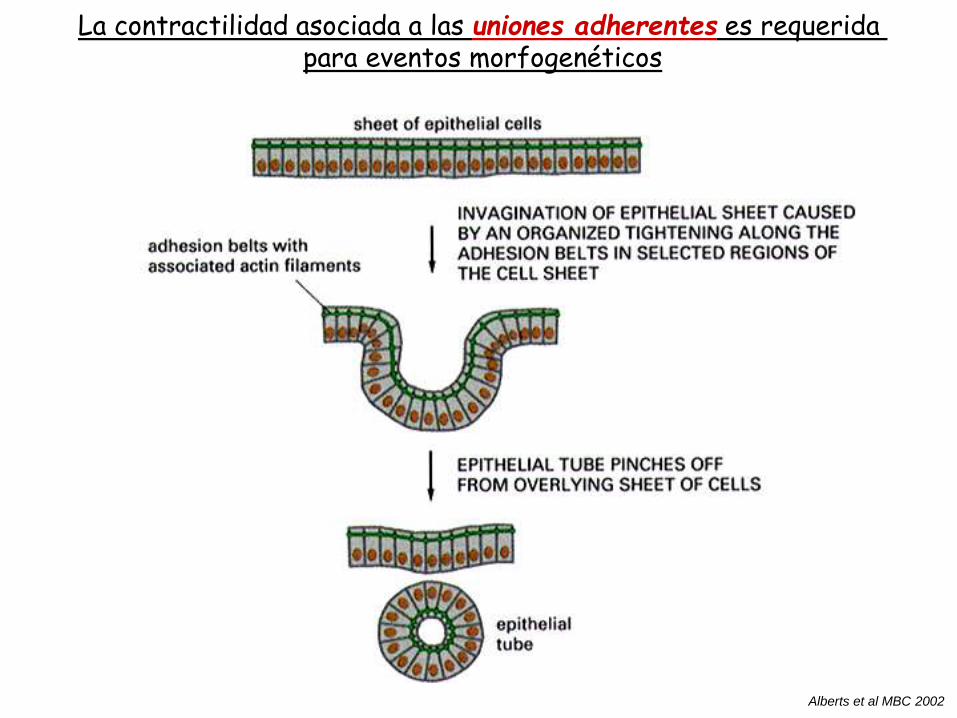
La contractilidad asociada a las uniones adherentes es requerida para eventos morfogenéticos
Alberts et al MBC 2002

Microscopía electrónica de barrido. Formación del tubo neuraly de la vesícula óptica
Alberts et al MBC 2002
Gilbert Developmental Biology 2000
invaginación de ectodermo

DESMOSOMAS Y HEMIDESMOSOMAS
Los desmosomas y hemidesmosomas son uniones de
anclaje que contribuyen a la estructura de los epitelios.
Intracelularmente se acoplan a los filamentos
intermedios los cuales no son contráctiles. Los
desmosomas anclan células epiteliales adyacentes,
mientras que los hemidesmosomas anclan la cara
basal de las células epiteliales a la lámina basal.
Alberts et al MBC 2002
integrin
receptors

Organización y componentes moleculares de los desmosomas
Las proteínas de transmembrana de los desmosomas son las desmogleínas y desmocolinas, integrantes de
la superfamilia de las caderinas. Los dominios extracelulares de desmogleínas y desmocolinas de células
adyacentes interaccionan entre sí en una manera dependiente de calcio. Sus dominios intracelulares se unen
a placoglobinas y placofilinas que mediante la unión a desmoplaquinas, anclan el complejo a los
filamentos intermedios.
Alberts et al, MBC 2015

Organización y componentes moleculares de los hemidesmosomas
Las principales proteínas de transmembrana de los hemidesmosomas son las integrinas 6β4 y el
colágeno de transmembrana tipo XVII. Estas proteínas interaccionan entre sí y se unen a la laminina de la
membrana basal. Sus dominios intracelulares interaccionan con las proteínas adaptadoras plectina y
BP230 que a su vez se unen a los filamentos intermedios.
Alberts et al, MBC 2015

Lodish et al MBC 2004
Cohen et al., JCB 2004
La formación de las uniones de anclaje y estrechas son esenciales para lapolarización de las células epiteliales
Las células epiteliales MDCK se polarizan cuando se cultivan confluentes sobre soportes porosos cubiertos
con una lámina basal sintética. Microscopía de fluorescencia y electrónica revelan que las células adquieren
una forma columnar, y diferencian un dominio apical con microvellosidades, y un dominio basolateral.
MDCK: Madin-Darby Canine Kidney cells

Las Caderinas tienen un rol preponderante en las uniones intercelulares
Las caderinas son proteínas de transmembrana cuyo
dominio extracelular contiene repeticiones de ~110
aminoácidos, denominadas “cadherin repeats” (CR). Las
caderinas clásicas son una familia de proteínas de
transmembrana de tipo I a la cual pertenecen las
caderinas tipo E, P y N. Se caracterizan por tener cinco
CR en su dominio extracelular y un dominio citoplasmático
conservado que interacciona con proteínas citosólicas
denominadas cateninas. La unión de Ca2+ entre los CR
favorece la forma rígida y extendida del ectodominio.
Molecule Predominant Cellular Distribution
E-cadherin Preimplantation embryos, non-neural epithelial tissue
P-cadherin Trophoblast
N-cadherin Nervous system, lens, cardiac and skeletal muscle
SOURCE: M. Takeichi, 1988, Development 102:639; M. Takeichi, 1991, Science 251:1451; H. Inuzuka et al., 1991, Neuron 7:69; and M. Donalies et al., 1991, Proc. Nat'l. Acad. Sci. USA 88:8024.
caderinas clásicas
Molecule Predominant Cellular Distribution
E-cadherin Preimplantation embryos, non-neural epithelial tissue
P-cadherin Trophoblast
N-cadherin Nervous system, lens, cardiac and skeletal muscle
SOURCE: M. Takeichi, 1988, Development 102:639; M. Takeichi, 1991, Science 251:1451; H. Inuzuka et al., 1991, Neuron 7:69; and M. Donalies et al., 1991, Proc. Nat'l. Acad. Sci. USA 88:8024.
Alberts et al, MBC2008; Pokutta & Weiss, ARCDB 2007
Estructura
tridimensional del
dominio extracelular
(ectodominio).
CR1
CR2
CR3
CR4
CR5
Ca2+
carbohydratos
catenins

Los dímeros en trans intercambian hojas beta
entre los CR1. La unión es de baja afinidad (~
0,7mM) y la fuerza para romper la interacción de
un dímero es de ~60pN. Sin embargo las
uniones adherentes involucran la interacción
entre varios miles de moléculas de caderinas.
Interacción de los ectodominios de las caderinas y agregación en la membrana
MP
MP
interacción de
múltiples
ectodominios
en trans
CR1
CR1
Trp
Trp
El triptofano (Trp) conservado
de un CR1 interacciona con un
bolsillo hidrofóbico del CR1 de
la molécula en trans.

El reconocimiento homofílico de las caderinas es un mecanismoque facilita la segregación de grupos o poblaciones celulares
células sin caderinas
incubadas con y sin Ca2+
células transfectadas con
caderinas e incubadas en
presencia y en ausencia de Ca2+
conclusión: la adhesión
requiere de caderinas
y calcio.
resultado: las células
que expresan las mismas
caderinas se segregan.
conclusión: la adhesión
es homotípica.
resultado: células que
expresan diferentes
niveles del mismo tipo
de caderina se segregan.
conclusión: diferencias
cuantitativas en la
expresión de caderinas
juegan un rol en la
organización de los
tejidos
estado
inicial
estado
final
incubación
con agitación +Ca2+
células transfectadas
con E-caderinas (azul)
y N-caderinas (naranja)
resultado:
adhesión
resultado:
no adhesión
mezcla de células que
expresan diferentes niveles
del mismo tipo de caderina
resultado:
no adhesión
+Ca2+ +Ca2+

Hibridización in situ revela la expresión de diferentes
caderinas en áreas específicas del cerebro de un
embrión de ratón.
El neuroectodermo expresa E-caderinas. Durante
la formación del tubo neural el neuroepitelio cambia
la expresión de E-caderinas por N-caderinas.
E-cad
N-cad
tubo
neural
ectodermo
La interacción homofílica mediada por caderinas facilita la segregación celular durante el desarrollo
Hirano & Takeichi, Physiol Rev 2012

Las cateninas anclan las caderinas al citoesqueleto y la estabilizan en la superficie
La b-catenina interacciona directamente con
las caderinas y con -catenina. Alfa catenina
interacciona con otras proteínas (vinculina, -
actinina) que unen actina. Las interacciones
son dinámicas y reguladas por fosforilación.
Alberts et al, MBC2008; Hirano & Takeichi, Physiol Rev 2012
sin anclaje al citoesqueleto
adhesión débil
con anclaje al citoesqueleto
adhesión fuerte
(-catenin,
vinculin,
-actinin)
~~extracelular
domain
En desmosomas la proteína adaptadora equivalente
a la β-catenina es placoglobina.
anclaje

La unión de la catenina p120 bloquea las señales de endocitosis (en amarillo) presentes en el dominio
citoplasmático de las caderinas e impide su internalización mediada por clatrina. La fosforilación de la
caderina inhibe la unión de p120 y facilita la endocitosis de las caderinas.
La catenina p120 es requerida para estabilizar la caderina en la superficie celular
Reynolds, Cell 2010
endocytic
signal
DEE

La β-catenina no unida a las adhesiones puede actuar como una señalque promueve la actividad transcripcional
Las caderinas reclutan β-catenina a las adhesiones intercelulares. La β-catenina disociada de las
adhesiones puede seguir dos destinos: a) degradación en proteasomas; b) estabilización y translocación al
núcleo donde forma complejos con los factores de transcripción TCF y LEF y promueve la transcripción de
genes pro-invasivos y mitogénicos (ciclina D, metaloproteasas, fibronectina, myc, etc).

La regulación transcripcional de las caderinas también modifica la adhesión
Kang & Masagué, Cell 2004
transición epitelio-mesénquima o TEM
receptores
TGFβFGF
Wnt
extracelular
intracelular
TEM
La estimulación de receptores de crecimiento y
citoquinas activan y promueven la expresión de
factores de transcripción, como Twist, Snail,
SIP1 y Slug, que reprimen la expresión de E-
caderina y cateninas y estimulan la expresión
de proteínas involucradas en motilidad.
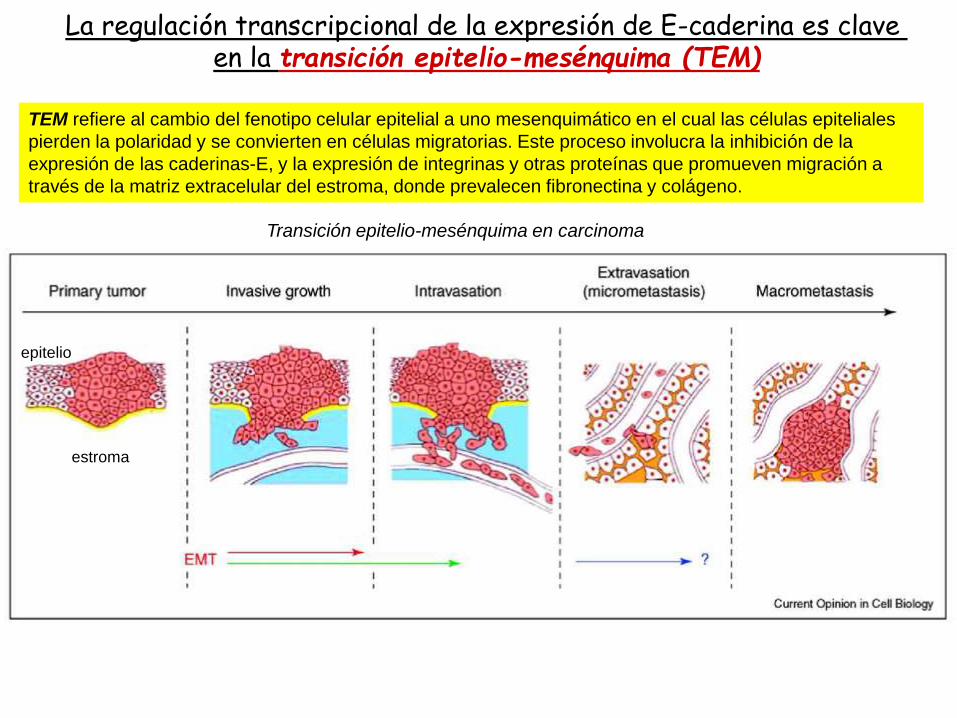
La regulación transcripcional de la expresión de E-caderina es clave en la transición epitelio-mesénquima (TEM)
TEM refiere al cambio del fenotipo celular epitelial a uno mesenquimático en el cual las células epiteliales
pierden la polaridad y se convierten en células migratorias. Este proceso involucra la inhibición de la
expresión de las caderinas-E, y la expresión de integrinas y otras proteínas que promueven migración a
través de la matriz extracelular del estroma, donde prevalecen fibronectina y colágeno.
Transición epitelio-mesénquima en carcinoma
estroma
epitelio

epitelio
tejido
conectivo
(estroma)
membrana
basal
normal carcinoma
La flecha en (F) indica células invasivas que han atravesado la membrana basal y se diseminan por el estroma.
Durante el proceso de TEM las células tumorales epiteliales degradan la membrana basal e invaden el estroma

TEM es regulado durante el desarrollo embrionario y permite la formación de estructuras nuevas
↓ N-caderina
↑ integrinas
↑ N-caderina
↑ N-caderina
Las células de la cresta neural se originan
por TEM. Después de migrar a lo largo del
embrión se agregan y diferencian para
formar distinas estructuras corporales.

La adhesión mediada por caderinas puede ser alterada rápidamente por mecanismos post-traducción
Balzac et al., JCS 2005
A B
Las uniones entre caderinas requiere de calcio. La
eliminación de Ca2+ con un quelante (EGTA) inhibe
reversiblemente la adhesión.
células unidas separadas unidas unidas separadas
El anclaje de las caderinas al citoesqueleto es inhibido
por tirosina quinasas como Src y VEGFR. Inhibidores de
fosfatasas (PAO) provocan la pérdida de la adhesión.
La endocitosis de las caderinas ocurre de
manera constitutiva pero es estimulada por
diferentes factores extracelulares. La
endocitosis ocurre por mecanismos
dependientes e independientes de clatrina.
La unión de la catenina p120 al
tallo citoplasmático de la
caderina inhibe su endocitosis.
EGF, HGF, VEGF

La diapédesis requiere de una inactivación temporaria y rápida de la función adhesiva de las caderinas vasculares
Shaw et al, J. Immunol. 2001
Visualización de la migración trans-epitelial de leucocitos. Células endoteliales fueron transfectadas con VE
caderina-GFP y activadas con TNF. Leucocitos polimorfonucleares marcados con Cell Tracker orange (rojo) se
sembraron sobre la monocapa y se registró el pasaje entre dos células endoteliales adyacentes (flecha en 3:00).
0 0:45 3:00 3:15 3:30
3:45 4:00 4:15 4:30 9:15
video disponible
El factor de crecimiento vascular, VEGF, estimula receptores (VEGFR) con actividad quinasa de la
membrana endotelial. El VEGFR fosforila las caderinas vasculares provocando la pérdida de la adhesión.

Las caderinas y otras moléculas de adhesión funcionan de manera coordinadadurante el establecimiento de las sinapsis neuronales
sinapsis
madura
sinaptogénesiscadherin

Hynes. Cell 2002,
Anthis & Campbell, TIBS 2011
Las integrinas son receptores de moléculas de la matriz extracelular
12
10
11
collagen receptors
Fibronectin,
Vitronectin,
Fibrinogen
El receptor es un heterodímero formado por la unión no covalente entre una subunidad y una b
de una familia de subunidades y b. Ambas subunidades son proteínas de transmembrana tipo I.
El sitio de unión al ligando está compuesto por el extremo N-terminal de las dos subunidades.
β-propeller
ligando
Mg2+, Mn2+

La actividad de las integrinas depende de cambios conformacionalesmodulados por ligandos extra e intracelulares
Las integrinas existen en dos conformaciones alostéricas que determinan la afinidad por el ligando: inactiva
(= baja afinidad por el ligando) y activa (= alta afinidad por el ligando). La conformación inactiva es
estabilizada por interacciones entre las cadenas y b. Interacciones específicas con ligandos extracelulares
o intracelulares inducen cambios conformacionales que activan el receptor y su señalización.
Shattill et al NRMBC2010
microscopía electrónica
de los ectodominios en
la conformación inactiva
(A) y activa (B).
El tripéptido RGD está presente en
proteínas de la matriz extracelular,
como fibronectina, y es reconocido
por ciertas integrinas celulares
interacciones
entre α y β
inactiva
activa activa
regulación del
citoesqueleto,
supervivencia,
transcripción
ensamble y
organización
de la ECMligando extracelular
ligando
intracelular

La unión de las integrinas al ligando extracelular induce su agregacióny la formación de complejos de adhesión
Las agregación de integrinas incrementa la avidez (múltiples pares de interacción receptor/ligando). La
agregación es facilitada por la naturaleza polimérica de las proteínas de la matriz extracelular y por el
efecto de fuerzas ejercidas por el citoesqueleto de actina y miosina. Varias proteínas “scaffold” tales
como talina, vinculina y -actinina participan en el anclaje de las integrinas a los filamentos de actina.
Marcación fluorescente de b1-integrina
y actina: complejos de adhesiónLa microscopía de super-resolución revela diferentes
capas de proteínas en los complejos de adhesión
Kanchanawong et al, Nature 2010

A través de los complejos de adhesión las células ejercen fuerzas de tracción sobre el substrato
tracción mediada por miosina II inhibición de miosina y la tracción
(A y C) muestran la deformación de un substrato de silicona cuando se estimula la GTPasa
RhoA. La inhibición de la actividad de miosina elimina las fuerzas de tracción (B).
A BC
La estimulación de integrinas
activa RhoA, que a su vez
activa a miosina y promueve la
contractilidad actina-miosina.
Ballestrem et al 2001;
Morimatsu et al 2013
Determinación de fuerzas
mediadas por las integrinas.
Se emplea un sensor de
fuerzas compuesto por pares
de FRET en los extremos de
una secuencia peptídica
elástica, y de un motivo RGD
para la unión de integrinas. El
rango de fuerzas de una
integrina es de 2-40 pN.
high force

Las fuerzas generadas por el citoesqueleto y los complejos de adhesión promueven el ensamble y organización de la matriz extracelular
Las fuerzas ejercidas por fibroblastos
sobre moléculas de fibronectina
extracelular inducen la exposición de
sitios de oligomerización ocultos que
promueven el ensamble de las fibras.
Alineamiento de las fibras de
colágeno entre dos colonias de
fibroblastos inmersos en un gel
de colágeno.
1 mm
coloniacolonia
gel de colágeno
fibras alineadas
secuencias de oligomerización
tensionado
módulo
relajado
módulo de FN III

Modelo de ensamble de las fibras de fibronectina
Singh et al Annu Rev Cell Dev Biol 2010
Dímero de FN unido
a la integrina.
Fuerzas celulares alteran la
conformación de moléculas de FN.
La agregación de integrinas expone
nuevos sitios de unión entre
moléculas de FN y cambios
adicionales en la conformación.
polimerización y formación de
una matriz de FN insoluble.

La agregación de las integrinas en los complejos de adhesiónactiva tirosina quinasas y promueve fosforilación
Fibroblastos adheridos sobre fibronectina activan la fosforilación de numerosas proteínas en los
complejos de adhesión. Esto puede apreciarse por inmunofluorescencia con un anticuerpo
anti-fosfotirosina. Las flechas señalan los focos de adhesión marcados con el anticuerpo.

FAK y Src son las principales quinasas activadas en los complejos de integrinas
integrinas FAK Cas Crk JNK
integrinas FAK Grb2 Sos Ras Erk
(MAP kinasa)
(MAP kinasa)
expresión de ciclinas D;
proliferación
PTK adaptor adaptor MAP
PTK adaptor GEF GTPase MAP

Integrinas localizadas en rafts lipídicos activan una vía de señalización dependiente de caveolina y Fyn
integrina caveolina Fyn Shc Grb2 Sos Ras ErkPTK adaptor adaptor GEF GTPase MAP

Las quinasas Src y FAK activadas en las adhesiones cumplen diferentes roles reguladores del citoesqueleto y motilidad
Las células normales producen
focos de adhesión alargados
(flechas en (A). Las células
deficientes en FAK (B) producen
múltiples focos pequeños.
Src-FAK
E-cadherin
Rac
polimerización de
actina y extensión
de lamelas y
filopodios
crecimiento y
recambio de
los contactos
adhesivos
+
+
-
+
A
B
(Schlaepfer, PBMB 1999)
Rho
+
++
generación
de fuerza,
contractilidad
miosinaArp2/3, forminas

Las células normales se polarizan formando una lamela de avance en la dirección de la migración. Las células deficientes
en FAK no se polarizan eficientemente, cambian de dirección frecuentemente y exhiben alteraciones quimiotácticas.
Tilghman et al JCS2005; Owen et al JCB 2007
Tavora et al EMBO Mol Med 2010
FAK es requerido para la polarización y migración direccional
células que expresan FAK
células que no expresan FAK
cambios de forma de macrófagostiempo
FAK wt
FAK KO
Inhibición de la respuesta
quimiotáctica de células
endoteliales al VEGF

Las adhesiones regulan el dinamismo del citoesqueleto y la migración celular
Durante la migración, nuevas adhesiones se ensamblan en el margen de avance (a) y se desensamblan
en el polo posterior (p).
0 min 5 min 10 min 15 min
Burdiso et al JCS 2013; Pankov et al JCB 2005
p
a a aa
p
Mutaciones en integrinas que afectan la activación de GTPasas también provocan alteraciones en los patrones de migración
WT mut
trayectorias azarosas direccionadas

Un nivel de adhesión intermedio es óptimo para la migración
Holly et al., Exp. Cell Res. 2000
La fuerza de la adhesión de las células al substrato depende del tipo celular y del tipo
de substrato. Un nivel intermedio de adhesión es compatible con mayores velocidades
de migración. Niveles de adhesión bajos o muy altos inhiben la migración.
Nivel de adhesión

La interacción de las integrinas epiteliales con la membrana basal es requerida para la polarización y diferenciación de los epitelios
Estructura de un acino de glándula mamaria
polarización normal
O’Brien et al, NCB 2001; Bissell et al, COCB 2003
La inhibición del ensamble de
la lamina basal produce una
polarización invertida (rojo)
En rojo se muestra un marcador del dominio apical
lámina
basal
lamina
basal
lumen
actina

La estimulación de las integrinas promueve la supervivencia celular mediante la inhibición de la apoptosis
APAF 1: Apoptotic Protease Activating Factor 1
integrina
ECM
FAK
Ausencia de estimulación apoptosis estimulación apoptosis
integrina activa

La mayoría de las células dependen del anclaje a la matriz extracelular para su crecimiento, proliferación y supervivencia
El incremento del área celular generado
por contactos entre la célula y el substrato
es un factor crítico que promueve el
crecimiento, supervivencia y la proliferación.
= área total
de fibronectina
Growth
fibronectina
Chen et al., Science 1997
integrinas
caderinas
sobrevivencia,
crecimiento y
proliferación
diferenciación apoptosis
. ..... . ..... . ..... . .....
La estimulación de integrinas activan vías de
señalización que promueven el crecimiento,
la proliferación y la supervivencia de las células.
extendidaextendida
sobrevivencia
y crecimiento
Las células epiteliales requieren del contacto con la
matriz para su diferenciación. Durante la involución
de la glándula mamaria y de la próstata, la lámina
basal se degrada y las células mueren por apoptosis.

matriz extracelular
Erk
Ras
caveolina
ciclo celular
Rho
cki
Raf/MEK
ciclina D
PI-3k
Akt
caspasa 9
Bad
apoptosis
Rac
JNK
p53
Red de vías de señalización activadas por adhesión
CDK
motility
p130Cas/
paxilina
ShcGrb2
FAK/Src
integrinasmembrana

CAMs (Cell Adhesion Molecules)
Alberts et al MBC 2002
Se caracterizan por poseer repeticiones de motivos homólogos a los encontrados en inmunoglobulinas
(Ig-like) en la porción extracelular. Otros módulos frecuentes en los dominios extracelulares son los FN-
III, homólogos a los encontrados en fibronectina. Algunos miembros como las ICAM y VCAM median
interacciones heterofílicas, otras como las NCAM median interacciones homofílicas.
Ig-like domain
N-terminus
C-terminus
N-terminus
C-terminus
FN-III domain
A diferencia de las caderinas
la mayoría de las CAMs no
median anclaje entre células,
y no requieren calcio para
su función adhesiva.

La N-CAM y otras CAMs relacionadas cumplen funciones importantes en reconocimiento intercelular durante el desarrollo
Durante la formación del sistema nervioso los axones neuronales se fasciculan y recorren
distancias considerables juntos. NCAM y L1 contribuyen en este proceso. El agregado de
ácido polisiálico (PSA) en NCAM inhibe su función adhesiva y produce defasciculación.
Las N-CAMs fueron las
primeras CAMs caracterizadas.
Existen ~ 20 isoformas de N-
CAM generadas por corte y
empalme de un mRNA comun.
Fascículos de axones de neuronas
ganglionares de la retina. Los fascículos
axonales convergen en el sitio de salida
del nervio óptico (asterisco rojo).
*
Villegas-Perez et al., J. Neurosci 1988
N-CAM con
repeticiones de
ácido siálico (PSA)
PSA: Poly Sialic Acid

(A) Las ICAMs y VCAMs se expresan en el endotelio e interaccionan con integrinas (Lβ2) de linfocitos T activados durante
procesos inflamatorios. La interacción entre integrinas/CAMs acercan las membranas adheridas favoreciendo la transmisión
de señales intercelulares (ej. quimioquinas) y eventualmente la diapédesis. (B) Las ICAMs expresadas en células dendríticas
se unen a integrinas en las membranas de linfocitos T y promueven la sinapsis inmunológica.
A
ICAM y VCAM cumplen funciones importantes en la interacción de células del sistema inmune y el endotelio
B
LFA-1: Leukocyte Function Antigen = integrin L/β2
Sinapsis inmunológica
Mecanismos dependientes e independientes de citoquinas secretadas
por las células endoteliales, activan integrinas de los linfocitos y
establecen una unión fuerte entre las células y el endotelio.
Constanin et al Nature Immunol 2012
signaling &
activation
of talin
signaling &
activation
of talin
Lβ2

Las selectinas poseen un dominio de lectina en el extremo distal
de la región extracelular, que reconoce de manera específica y
dependiente de calcio a oligo/polisacáridos en glicolípidos y
glicoproteínas (ej. PSGL-1). El dominio intracelular se asocia al
citoesqueleto de actina. Las selectinas median adhesiones de
baja afinidad entre leucocitos y las células endoteliales.
Las Selectinas interaccionan con carbohidratos en proteínas y lípidos
PSGL-1 (P-Selectin Glycoprotein Ligand-1)
Marth & Grewal, Nature Rev Immunol 2008; MBC Alberts et al 2015; Tinoco et al Trends Immunol 2017
Alineamiento de
ectodominios de
selectinas E y P
Ca2+
extracelular
intracelular
PSGL-1
sLex: tetrasacárido
Las selectinas intervienen en las primeras
etapas de la extravasación de leucocitos.

Función coordinada de selectinas, integrinas y caderinas durante la extravasación de leucocitos
Estímulos inflamatorios (ej. TNFα) inducen la expresión de selectinas en la superficie endotelial que se unen a glicanos en la
membrana de leucocitos (“rolling”). La unión leucocito-endotelio permite la señalización mediada por quimioquinas liberadas
por las células endoteliales y que estimulan receptores en los leucocitos (“activation”). La activación de integrinas en los
leucocitos permite su interacción con ICAM, VCAM, y en forma coordinada con la inactivación de VE-caderinas y PECAM
(Platelet Endothelial Cell Adhesion Molecule) facilitan el pasaje trans-endotelial de los leucocitos.
Ley et al Nature Rev Immunol2007
VE-cadherin, JAMs
ICAM, PECAMselectins
integrins
“inside-out”
integrins
“outside-in”

La señalización de diferentes moléculas de adhesión es integrada en el citoplasma
integrinasRTKs
factores de
crecimientomatriz
extracelular
E-caderinas
b-catenina
Frizzled
Wnt
Erk
ciclo celular
cki
ciclina D
núcleo
uniones estrechas
Zona B
ZO-1
ZO-1 en uniones estrechas
secuestra los factores de
transcripción Zona B y la
kinasa Cdk4, inhibiendo la
proliferación.
b-catenina es reclutada
a los complejos de
caderinas inhibiendo la
proliferación.
Cdk4estabilización
en citoplasma
retención
en
adhesiones
retención en adhesiones
activación
síntesis
síntesis
fosforilación,
inactivación
La estimulación de integrinas activa
quinasas que promueven la síntesis
de ciclina D e inhiben la función y
expresión de inhibidores de Cdks.
FAK
CDKs