lactose and melibiose metabolism in erwinia chrysanthemijb.asm.org/content/168/2/886.full.pdf ·...
TRANSCRIPT

JOURNAL OF BACTERIOLOGY, Nov. 1986, P. 886-8910021-9193/86/110886-06$02.00/0Copyright © 1986, American Society for Microbiology
Vol. 168, No. 2
Lactose and Melibiose Metabolism in Erwinia chrysanthemiJAMES S. S. GRAY,'* WILLIAM A. LINDNER,' JOHN M. BRAND,1 AND JOHN P. MILDENHALL2
Department ofBiochemistryl and Department of Genetics and Plant Protection,2 University ofFort Hare, Alice 5700,Republic of Ciskei, South Africa
Received 12 August 1985/Accepted 5 August 1986
A Lac' mutant of Erwinia chrysanthemi was isolated from the Lac- wild type on lactose agar. (3-Galactosidase was expressed independently of lactose transport in both the mutant and the wild type, andneither strain expressed thiogalactoside transacetylase. Lactose transport and a-galactosidase, constitutive inthe Lac' strain, were coordinately induced in the Lac- strain by melibiose and raffinose but not byisopropyl-,B-D-thiogalactopyranoside or thiomethyl-3-D-galactopyranoside. Melibiose was a strong inhibitor ofboth the melibiose- and the raffinose-induced lactose permeases, whereas raffinose was a strong inhibitor ofonly the raffinose-induced lactose permease.
Many members of the family Enterobacteriaceae possessoperons that code for enzymes involved in lactose andmelibiose metabolism. In Escherichia coli, the lacZ genecodes for ,B-galactosidase, the lacY gene codes for lactosepermease, and the lacA gene codes for thiogalactosidetransacetylase (36). The mel operon of E. coli contains the Aand B genes which code for a-galactosidase and melibiosepermease, respectively (30). Klebsiella aerogenes possessesboth a plasmidborne and a chromosomal lac operon (1, 21,22, 24). Both K. aerogenes and Salmonella typhimuriumpossess mel operons similar to the E. coli operon (12, 23).The substrate specificities of the lac and mel permeases
differ among the various members of the Enterobacteria-ceae. The lac permease of E. coli has wide specificity andtransports a variety of a- and P-galactosides, includinglactose, isopropyl-p-D-thiogalactopyranoside (IPTG),thiomethyl-,3-D-galactopyranoside (TMG), melibiose, andgalactose (26). The mel permeases of E. coli and S.typhimurium are similar in specificity and can transportTMG, melibiose, and galactose, but not lactose or IPTG (12,19, 26, 30). The mel permease of K. aerogenes differs fromthose of E. coli and S. typhimurium in being able to transportlactose in addition to TMG, melibiose, and galactose (23,35).The rafoperon is plasmidborne in E. coli (18). It codes for
the three genes involved in raffinose metabolism: a-galactosidase, raffinose permease, and invertase (29, 32). Allthree activities are coordinately induced by melibiose andraffinose (29, 32). Lactose transport by the raffinosepermease is competitively inhibited by melibiose and raf-finose (32).Erwinia chrysanthemi pv. zeae, the causal agent of soft rot
in corn, is a member of the Enterobacteriaceae (34). Onlytwo reports dealing with lactose metabolism in E.chrysanthemi have appeared (6, 8) despite the use of lactosefermentation as a taxonomic marker in the classification ofthe Enterobacteriaceae. The 322 strains of E. chrysanthemistudied by Dickey (3) were all able to hydrolyze o-nitrophenyl-p-D-galactopyranoside (,B-ONPG), althoughmost of them were classified as Lac-. Other workers haveclassified lactose fermentation by E. chrysanthemi as beingeither variable or delayed (5, 11).
* Corresponding author.
Our studies on lactose metabolism by a locally isolatedstrain of E. chrysanthemi showed that it was normallylactose nonutilizing (Lac-). However, lactose-utilizing(Lac') mutants are readily isolated from the parent strain byextended incubation on lactose agar (6). The Lac- and Lac'strains were found to be physiologically and pathogenicallysimilar except for their growth on lactose (6). Since theactivity of the [-galactosidase from the Lac- and Lac'strains was similar on both lactose and P-ONPG, we sug-gested that the Lac- strain differed from the Lac' strain bythe absence of a lactose permease (6). The present studyconfirms that the Lac- strain does differ from the Lac'strain by the absence of a lactose permease coordinatelyinduced with ,3-galactosidase. Evidence is presented that E.chrysanthemi possesses two permeases involved in lactosetransport. One permease appears to be associated with themel operon and the other with the rafoperon. It is concludedthat the Lac' phenotype is due to a mutation allowingconstitutive expression of the mel operon.
MATERIALS AND METHODS
Chemicals. D-[glucose-1-14C]lactose (specific activity, 57.7mCi mmol-1) was purchased from the Radiochemical Centre(Amersham, England). [methyl-14C]TMG ([14C]TMG) (spe-cific activity, 35 mCi mmol-1) was obtained fromSchwarz/Mann (Orangeburg, N.Y.). [isopropyl-14C]IPTG([14C]IPTG) (specific activity, 28 mCi mmol-1) was pur-chased from Service des Molecules Marqudes, Gif-Sur-Yvette, France. P-ONPG and a-ONPG, nonradioactiveIPTG and TMG, and all carbohydrates were purchased fromthe Sigma Chemical Co. (St. Louis, Mo.). Dithiothreitol wasobtained from Boehringer GmbH (Mannheim, Federal Re-public of Germany). Sodium dodecyl sulfate was purchasedfrom the Pierce Chemical Co. (Rockford, Ill.).
Media. The yeast-salts medium described by Mildenhall etal. (14) was adjusted to pH 7.3 before autoclaving (15 lb/in2,121°C, 20 min). The cooled medium was supplemented with5 g of filter-sterilized (0.22-,um-pore-size Millex filter; Mili-pore Corp., Bedford, Mass.) carbon source liter-'. Thismedium was solidified by the addition of Difco agar (15 gliter-'). Cells for transport studies were grown in M63medium (15) supplemented with 50 mM NaCl as describedby Robbie and Wilson (25) and adjusted to pH 7.0 beforeautoclaving. Sterile medium received the following sterilesupplements: 1 mM MgSO4, 0.9 g of yeast extract liter-',
886
on June 19, 2018 by guesthttp://jb.asm
.org/D
ownloaded from

E. CHRYSANTHEMI LACTOSE AND MELIBIOSE METABOLISM 887
and 4 g of glycerol liter-1. When the effects of differentsugars on enzyme production were measured, they wereadded to the medium to a final concentration of 4 g liter-'.Filter-sterilized IPTG and TMG (0.22-,um-pore-size Millexfilter) were used at a final concentration of 0.5 mM.
Organisms. A strain of E. chrysanthemi (FH1) isolatedlocally from corn (13) was used in these studies and has beendeposited, as strain SR260, in the collection of ArthurKelman (Department of Plant Pathology, University of Wis-consin, Madison). This strain, which is Lac-, has beendesignated EC-C. A Lac' mutant, designated EC-S, wasisolated from the wild type by growth on yeast-salts-lactoseagar (6). Both strains were maintained on Difco nutrient agarat 20 to 22°C and subcultured monthly.Enzyme assays. P-Galactosidase was assayed by the
method of Miller (15). The cells were lysed by a modificationof the lysis method of Putnam and Koch (20) involvingreplacement of the sodium dodecyl sulfate with sodiumdeoxycholate.
ot-Galactosidase was assayed in toluene-permeabilizedcells, since the lysis mixture completely inhibited enzymeactivity. The details of the assay were similar to thosedescribed by Schmitt and Rotman (31). The effects of 5 mMdithiothreitot and 5 mM Mn2+, reported by Schmitt andRotman (31) and Burstein and Kepes (2) to stabilize a-
galactosidase, were tested. In the presence of these com-pounds, the reaction was stopped by the addition of 1 ml of1 M Na2CO3-10 mM EDTA, since the colored complexformed between dithiothreitol and Mn2+ in alkaline solutioninterferes with the assay (31).A molar extinction coefficient of 21,300 M-1 cm-' at 420
nm and pH 10.3 for o-nitrophenol (9) was used to calculateenzyme activity, 1 unit of which is defined as the hydrolysisof 1 nmol of a- or 1-ONPG min-'.
Thiogalactoside transacetylase was assayed as describedby Miller (15). One unit of thiogalactoside transacetylaseactivty isdedil as an increase in A412 of 0.01 unit h-'.Preparati of cells for transport studies. Overnight cul-
tures were grown either in nutrient broth (Difco Laborato-ries, Detroit, Mich.) or in M63 medium supplemented withthe.a o carbon source on an orbital shaker (30°C,200 rp.} an then diluted (1:50) into M63 medium supple-mented with 4 g of glycerol per liter-' or various sugars orboth. as ibed in the text. After growth on the orbitalsIhV C fdal- (late log or early stationary phase), thecells were sedimented by centrifugation (10,000 x g, 40C, 10min), suspended to the original culture volume in M63medium without glycerol or yeast extract but containingchloramphenicol (0.15 mg ml-'), and recentrifuged. Thewash was repeated, and the cells were suspended to an A600of about 4.0 and stored on ice.
Transport of 14C-labeled IPTG, TMG, and lactose. Allassays were performed in M63 medium without glycerol or
yeast extract in a final volume of 1 ml in tubes (16 by 100mm). After equilibration at the experimental temperature for5 mn,, uptake was initiated by the addition of 100 ,uM[14ClWEG6f(.25 mCi mmoP1), [14C]TMG (0.5 mCi mmol),or [''CIctcu(2_5 or 0.125 mCi mmol-1). Samples (0.5 ml)werezinred after 10 min, filtered through a 0.45-,um-pore-size HAW? filter (Millipore Corp.) on a Millipore filtrationmanifold, and washed with five 0.5-ml aliquots of suspensionmediumatr-rn temperature (20 to 22°C). The filtration andwash. pocess wwas completed within 15 s. The filters werecounted -i either 10 ml of Beckman EP scintillation fluid(Beckn= mInuruments International S.A., Geneva, Switzer-land) or 10 mlof Packard Filtercount (Packard Instruments
TABLE 1. P-Galactosidase activity of strains EC-C and EC-Sgrown on different carbon sources
P-Galactosidase activityb (U/109 cells)Carbon sourcea
EC-C (Lac-) EC-S (Lac')
Glycerol 6.0 6.1Glycerol + raffinose 5.8 3.7Glycerol + melibiose 9.4 7.3Glycerol + galactose 11.9 8.7Glycerol + lactose 13.9 66.0Glycerol + IPTG 66.5 72.3Glycerol + TMG 64.8 73.8
a Strains EC-C and EC-S were grown to the late exponential phase inyeast-salts-glycerol medium. Galactose, lactose, melibiose, and raffinose wereadded at 4 g liter-', whereas IPITG and TMG were present at 0.5 mM.
b The assay mixture contained 0.1 ml of cells, 0.9 ml of Z buffer, and 25 RIof lysis mixture. After vortexing for 10 s, the assay mixture was equilibratedat 30°C for 5 min, and the reaction was started by the addition of 0.2 ml of1-ONPG (13.2 mM in H20). The reaction was stopped 10 to 15 min later by theaddition of 1.0 ml of 1 M Na2CO3, and after dilution of the mixture with 2 mlH20, the A420 and A550 were determined. A correction for turbidity was madeby subtracting 1.711 x A550 from the A420. Each value represents the mean ofthree determinations. Variation in all instances was less than 5%.
International S.A., Zurich, Switzerland) in a Beckmanmodel 8100 liquid scintillation counter.The effect of different sugars on substrate uptake was
determined by adding them, together with the substrate, tothe assay mixture. The effects of 4 mM 2,4-dinitrophenol and20 mM sodium azide on substrate uptake were determinedafter preincubation of the cells with these compounds for 30min. The effect of 70 mM formaldehyde on substrate uptakewas determined by adding it, together with the substrate, tothe assay mixture (10).
RESULTS
Induction of I8-galactosidase in EC-C (Lac-) and EC-S(Lac') by different carbon sources. ,-Galactosidase wasproduced by both EC-C and EC-S strains when they weregrown on glycerol as a carbon source (Table 1). The levelswere increased 10-fold by growth in the presence of 0.5 mMIPTG or TMG, but lactose was effective as an inducer onlywith strain EC-S. Melibiose, raffinose, and galactose in-duced P-galactosidase activity weakly or not at all in eitherstrain.
Thiogalactoside transacetylase activity in EC-C and EC-S.No thiogalactoside transacetylase activity was detected ineither strain EC-C or EC-S after induction by 1 mM IPTGeven though high levels of P-galactosidase were present(Table 2). Neither was thiogalactoside transacetylase activ-ity detected in unheated samples of either EC-C and EC-S,although a similar increase in the nonspecific breakdown ofacetyl coenzyme A was observed in both samples. It isunlikely that the failure to detect thiogalactoside transace-tylase was a result of a defective assay procedure since,under similar conditions, the enzyme could be detected ininduced E. coli. Therefore, it was concluded thatthiogalactoside transacetylase activity was absent in bothstrains of E. chrysanthemi.Uptake of 14C-labeled lactose, IPTG, and TMG by EC-C
and EC-S cells grown on different carbon sources. Lactosetransport was constitutive in EC-S and induced by melibioseand raffinose in EC-C (Table 3). Galactose was a poorinducer of lactose transport in EC-C, and neither IPTG,TMG, nor lactose induced a lactose transport system inEC-C.
VOL. 168, 1986
on June 19, 2018 by guesthttp://jb.asm
.org/D
ownloaded from

888 GRAY ET AL.
TABLE 2. Thiogalactoside transacetylase and P-galactosidaseactivity in EC-C, EC-S, and Lac' and Lac- strains of E. coli
Thiogalactoside ,B-Galactosidase
Strain transacetylase (U ml-') (U ml-')Induceda Uninduced Induceda Uninduced
EC-C (Lac-) <0.4 <0.5 610.0 63.1EC-S (Lac') <0.4 <0.5 537.7 40.8E. coli Lac' 33.7 <0.5 2,042.8 13.1E. coli Lac- 1.5 <0.4 1,006.9 56.5
a Overnight cultures of EC-C, EC-S, and both a Lac+ and a Lac- strain ofE. coli K-12 were grown in M63 medium supplemented with 4 g of glycerolliter-' and 0.9 g of yeast extract liter-1 at 30'C on an orbital shaker. Afterdilution (1:25) into fresh medium containing either 1 mM or no IPTG, thecultures were grown to an A6w of about 0.6 and harvested by centrifugation(10,000 x g, 4°C, 10 min). The cells were suspended in 0.1 of the originalculture volume of 0.05 M Tris-0.01 M EDTA (pH 7.9) and sonicated at 4'C.The samples were divided in two. One sample was heated at 70'C for 5 min,centrifuged, and assayed for thiogalactoside transacetylase. The other samplewas not heated before centrifugation and assay for P-galactosidase.
co
a)u
m
L-
CD
cL
ac
4.LeaCL.
co4-
L.
-J
0.4
0.2
0.
0
a v~~~~~~
0~~~~
-V
Neither EC-C nor EC-S was able to transport IPTGirrespective of the nature of the inducer in the growthmedium (data not shown). Measurement of [14C]TMG accu-mulation by EC-C and EC-S produced inconclusive results,because similar levels were found in both formaldehyde-treated and untreated cells (data not shown).The following carbohydrates at a concentration of 2 mM
inhibited lactose uptake by EC-S cells grown on glycerolplus lactose (10-min assay, 30°C; percent inhibition com-pared with the control given in parentheses): TMG (26%),IPTG (87%), thiodigalactoside (91%). Similar values wereobtained in the case of EC-C cells grown on glycerol plusmelibiose. Under the same experimental conditions, 2-deoxy-D-glucose, galactose, and glucose at 1 mM inhibitedlactose transport by EC-S cells grown on glycerol pluslactose by 22, 44, and 56%, respectively. Little or noinhibition of lactose uptake was observed by D-(+)-arabinose, L-(-)-arabinose, L-(-)-fucose, L-rhamnose, man-nitol, or D-mannose (data not shown).
Lactose transport by glycerol-grown EC-S cells did notdeviate greatly from linearity over the first 2 min whenassayed at room temperature (22°C) (Fig. 1). Similar curveswere obtained for melibiose-grown EC-C cells. Therefore,the average rate of transport over this period was used as anapproximation of the initial rate of transport in competitionexperiments, the results of which are presented in Table 4.Raffinose was a potent inhibitor of lactose transport by strain
TABLE 3. Uptake of [14C]lactose by strains EC-C and EC-Sgrown on different carbon sources
Lactose uptake (nmol 10 min'Carbon sourcea 109 cellsl)b
EC-C EC-S
Glycerol 0.17 10.0Glycerol + IPTG 0.43 16.7Glycerol + TMG 0.29 16.3Glycerol + lactose 0.17 10.7Glycerol + melibiose 6.6 6.9Glycerol + raffinose 2.13 8.1Glycerol + galactose 0.96 8.1
a Cultures were grown in M63 medium with various carbon sources (4 gliter-') and assayed for lactose transport as described in Materials andMethods.
b Ten-minute assays performed at 30'C. Each point is the mean of duplicatedeterminations.
50 100
Time (sec)FIG. 1. Inhibition by melibiose and raffinose of lactose transport
by the constitutive lactose transport system of strain EC-S. StrainEC-S was grown to the late exponential phase in M63 mediumsupplemented with 0.9 g of yeast extract liter' and 4 g of glycerolliter-'. The cells were washed and assayed for lactose uptake in theabsence (E) or presence (x) of 2 mM melibiose or raffinose (V).
EC-C grown on raffinose, whereas it was a relatively poorinhibitor of lactose transport by EC-S grown on glycerol.Melibiose, irrespective of the inducer present during growthof the cells, strongly inhibited lactose transport by bothEC-C and EC-S cells.Formaldehyde, 2,4-dinitrophenol, and sodium azide all
inhibited lactose transport by greater than 90%, indicatingthe presence of a carrier involved in active transport.
a-Galactosidase activity in permeabilized EC-C and EC-Scells. The lysis mixture used in the assay of ,B-galactosidasedestroyed a-galactosidase activity in EC-S cells. Conse-quently, the effects of toluene, dithiothreitol, and Mn2+ onot-galactosidase activity were investigated in glycerol-grownEC-S cells. Toluene permeabilization increased a-galactosidase activity by about 30%, with little further in-crease by the addition of dithiothreitol or Mn2+, either singlyor in combination (data not shown). Therefore, cells weremerely tolueneized in all subsequent a-galactosidase assays.High constitutive levels of a-galactosidase were found in
EC-S cells regardless of the carbon source, whereas theenzyme was induced by melibiose and, to a lesser extent, bygalactose and raffinose in strain EC-C (Table 5).
DISCUSSION
The strains of E. chrysanthemi used in this study (EC-Cand EC-S) do not have a lac operon analogous to that foundin E. coli. In E. coli, the lacZ, lacY, and lacA genes areexpressed coordinately (36). In contrast, P-galactosidase inE. chrysanthemi can be induced in EC-C without coordinateinduction of either a lactose permease or a thiogalactosidetransacetylase. Furthermore, in EC-S, a lactose permease isexpressed constitutively, whereas the ,B-galactosidase re-mains inducible. Last, melibiose, an inducer of the lacoperon in E. coli (16), does not induce ,-galactosidase in E.
J. BACTERIOL.
on June 19, 2018 by guesthttp://jb.asm
.org/D
ownloaded from
E. CHRYSANTHEMI LACTOSE AND MELIBIOSE METABOLISM 889
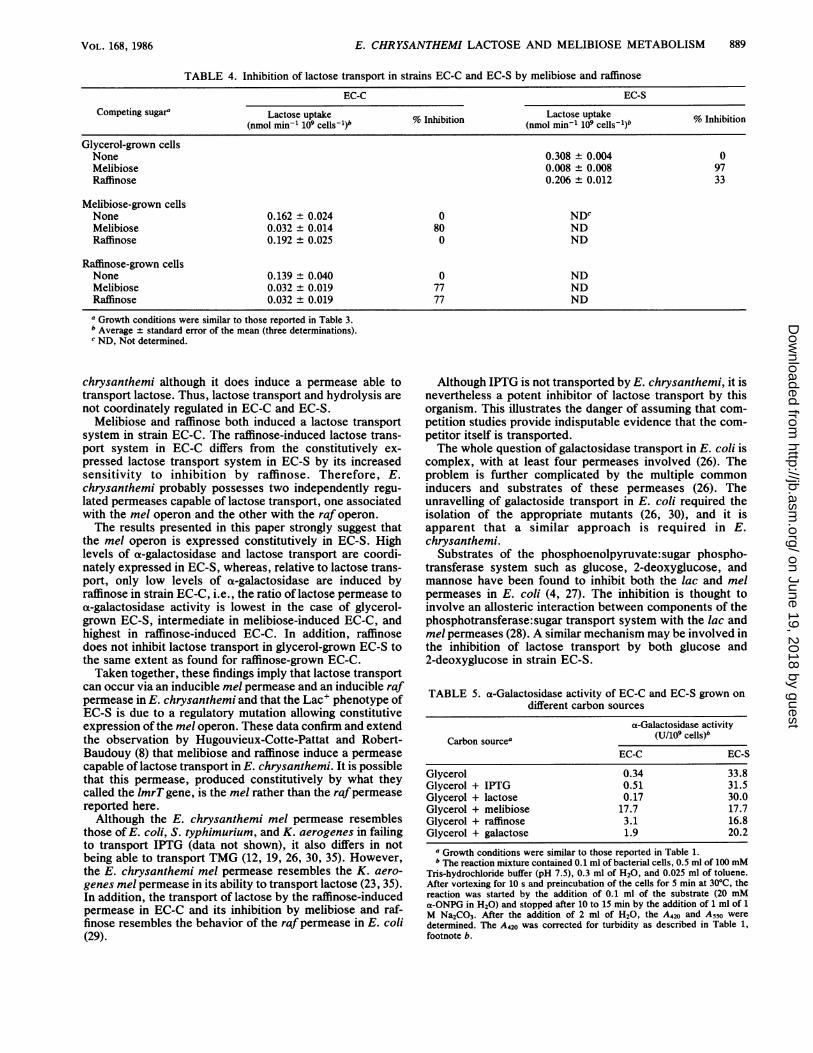
TABLE 4. Inhibition of lactose transport in strains EC-C and EC-S by melibiose and raffinose
EC-C EC-SCompeting sugara Lactose uptake Lcsua%Inhibition (nmol uptake % Inhibition
(nmol min-' 109 cellsl) % niiin(nmol min-' 109 cellsl1)b %IhbtoGlycerol-grown cellsNone 0.308 ± 0.004 0Melibiose 0.008 ± 0.008 97Raffinose 0.206 ± 0.012 33
Melibiose-grown cellsNone 0.162 ± 0.024 0 NDCMelibiose 0.032 ± 0.014 80 NDRaffinose 0.192 ± 0.025 0 ND
Raffinose-grown cellsNone 0.139 ± 0.040 0 NDMelibiose 0.032 ± 0.019 77 NDRaffinose 0.032 ± 0.019 77 ND
a Growth conditions were similar to those reported in Table 3.b Average ± standard error of the mean (three determinations).c ND, Not determined.
chrysanthemi although it does induce a permease able totransport lactose. Thus, lactose transport and hydrolysis are
not coordinately regulated in EC-C and EC-S.Melibiose and raffinose both induced a lactose transport
system in strain EC-C. The raffinose-induced lactose trans-port system in EC-C differs from the constitutively ex-pressed lactose transport system in EC-S by its increasedsensitivity to inhibition by raffinose. Therefore, E.chrysanthemi probably possesses two independently regu-lated permeases capable of lactose transport, one associatedwith the mel operon and the other with the raf operon.The results presented in this paper strongly suggest that
the mel operon is expressed constitutively in EC-S. Highlevels of a-galactosidase and lactose transport are coordi-nately expressed in EC-S, whereas, relative to lactose trans-port, only low levels of a-galactosidase are induced byraffinose in strain EC-C, i.e., the ratio of lactose permease toa-galactosidase activity is lowest in the case of glycerol-grown EC-S, intermediate in melibiose-induced EC-C, andhighest in raffinose-induced EC-C. In addition, raffinosedoes not inhibit lactose transport in glycerol-grown EC-S tothe same extent as found for raffinose-grown EC-C.Taken together, these findings imply that lactose transport
can occur via an inducible mel permease and an inducible rafpermease in E. chrysanthemi and that the Lac' phenotype ofEC-S is due to a regulatory mutation allowing constitutiveexpression of the mel operon. These data confirm and extendthe observation by Hugouvieux-Cotte-Pattat and Robert-Baudouy (8) that melibiose and raffinose induce a permeasecapable of lactose transport in E. chrysanthemi. It is possiblethat this permease, produced constitutively by what theycalled the lmrT gene, is the mel rather than the raf permeasereported here.Although the E. chrysanthemi mel permease resembles
those of E. coli, S. typhimurium, and K. aerogenes in failingto transport IPTG (data not shown), it also differs in notbeing able to transport TMG (12, 19, 26, 30, 35). However,the E. chrysanthemi mel permease resembles the K. aero-
genes mel permease in its ability to transport lactose (23, 35).In addition, the transport of lactose by the raffinose-inducedpermease in EC-C and its inhibition by melibiose and raf-finose resembles the behavior of the rafpermease in E. coli(29).
Although IPTG is not transported by E. chrysanthemi, it isnevertheless a potent inhibitor of lactose transport by thisorganism. This illustrates the danger of assuming that com-petition studies provide indisputable evidence that the com-petitor itself is transported.The whole question of galactosidase transport in E. coli is
complex, with at least four permeases involved (26). Theproblem is further complicated by the multiple commoninducers and substrates of these permeases (26). Theunravelling of galactoside transport in E. coli required theisolation of the appropriate mutants (26, 30), and it isapparent that a similar approach is required in E.chrysanthemi.
Substrates of the phosphoenolpyruvate:sugar phospho-transferase system such as glucose, 2-deoxyglucose, andmannose have been found to inhibit both the lac and melpermeases in E. coli (4, 27). The inhibition is thought toinvolve an allosteric interaction between components of thephosphotransferase:sugar transport system with the lac andmel permeases (28). A similar mechanism may be involved inthe inhibition of lactose transport by both glucose and2-deoxyglucose in strain EC-S.
TABLE 5. a-Galactosidase activity of EC-C and EC-S grown ondifferent carbon sources
a-Galactosidase activityCarbon source"a (U/109 cells)'
EC-C EC-S
Glycerol 0.34 33.8Glycerol + IPTG 0.51 31.5Glycerol + lactose 0.17 30.0Glycerol + melibiose 17.7 17.7Glycerol + raffinose 3.1 16.8Glycerol + galactose 1.9 20.2
a Growth conditions were similar to those reported in Table 1.b The reaction mixture contained 0.1 ml of bacterial cells, 0.5 ml of 100 mM
Tris-hydrochloride buffer (pH 7.5), 0.3 ml of H20, and 0.025 ml of toluene.After vortexing for 10 s and preincubation of the cells for 5 min at 30°C, thereaction was started by the addition of 0.1 ml of the substrate (20 mMa-ONPG in H20) and stopped after 10 to 15 min by the addition of 1 ml of 1M Na2CO3. After the addition of 2 ml of H20, the A420 and A550 weredetermined. The A420 was corrected for turbidity as described in Table 1,footnote b.
VOL. 168, 1986
on June 19, 2018 by guesthttp://jb.asm
.org/D
ownloaded from

890 GRAY ET AL.
We cannot explain why growth on melibiose results insignificantly lower a-galactosidase and permease activity inEC-S (Tables 3 and 5), but catabolite repression might beone cause. Similarly, it is not clear why IPTG and TMG inthe growth medium should enhance lactose transport instrain EC-S (Table 3). One possibility is that the high levelsof P-galactosidase induced by these compounds in E.chrysanthemi play a role by hydrolyzing the intracellularlactose. This would reduce the lactose concentration gradi-ent across the membrane against which the transport systemmust work.The resistance of the a-galactosidase from E.
chrysanthemi to toluene is similar to that of the raf cx-galactosidase from E. coli (29) but is different from the mela-galactosidases of E. coli and S. typhimurium, both ofwhich are sensitive to toluene inactivation (2, 7, 12, 31, 33).Furthermore, neither Mn2+ nor dithiothreitol, both of whichstabilize a-galactosidase activity in cell extracts of E. coli (2,31), activated the Erwinia enzyme. The Erwinia a-galactosidase also differs from the E. coli enzyme in that acofactor such as NAD+ is not required for activity (2).Many members of the Enterobacteriaceae have been
found to acquire new metabolic functions by mutations thatallow the enzymes of different pathways to be used for newfunctions (17). Two types of mutations have been discov-ered. First, regulatory mutations may occur that allowenzymes not inducible by their substrates to be expressedconstitutively, thereby permitting metabolism of the sub-strates (17). A second type of mutation may change thesubstrate specificity of an enzyme and lead to the metabo-lism of a compound that is not normally a substrate of theenzyme (17).The ,B-galactosidases in strains EC-C and EC-S have been
shown to have similar activity on P-ONPG and lactose (6).Therefore, the difference in lactose metabolism betweenEC-C and EC-S cannot be explained by an altered specificityof P-galactosidase for lactose. Our results suggest that EC-Sbecomes phenotypically Lac' owing to a mutation allowingconstitutive expression of the mel operon, i.e., a regulatorymutation.
Lactose metabolism of E. chrysanthemi has not beenstudied in detail, and it is not known whether a lac operonexists in any other strains of this organism. However, thedata presented in this paper may explain why many strains ofE. chrysanthemi are reported to have either a variable or adelayed ability to ferment lactose (3, 5, 11).
ACKNOWLEDGMENTS
This work was supported by grants from the University of FortHare and the Council for Scientific and Industrial Research,Pretoria.
LITERATURE CITED1. Brenchley, J. E., and B. Magasanik. 1972. Klebsiella aerogenes
strain carrying drug-resistance determinants and a lac plasmid.J. Bacteriol. 112:200-205.
2. Burstein, C., and A. Kepes. 1971. The a-galactosidase fromEscherichia coli K12. Biochim. Biophys. Acta 230:463-479.
3. Dickey, R. S. 1979. Erwinia chrysanthemi: a comparative studyof phenotypic properties of strains from several hosts and otherErwinia species. Phytopathology 69:324-329.
4. DilHs, S. S., A. Apperson, M. R. Schmidt, and M. H. Saier, Jr.1980. Carbohydrate transport in bacteria. Microbiol. Rev.44:385-418.
5. Goto, M. 1979. Bacterial foot rot of rice caused by a strain ofErwinia chrysanthemi. Phytopathology 69:213-216.
6. Gray, J. S. S., J. P. Mildenhall, and J. M. Brand. 1984. Lactosemetabolism in Erwinia chrysanthemi. Fort Hare Papers7:367-376.
7. Hogness, D. S., and E. H. Battley. 1957. The a-galactosidase ofAerobacter aerogenes; its characteristics and induced synthe-sis. Fed. Proc. 16:197-198.
8. Hugouvieux-Cotte-Pattat, N., and J. Robert-Baudouy. 1985. Lac-tose metabolism in Erwinia chrysanthemi. J. Bacteriol. 162:248-255.
9. Jones, K. M. 1969. Artificial substrates and biochemical re-agents, p. 448-451. In R. M. C. Dawson, D. C. Elliot, W. H.Elliot, and K. M. Jones (ed.), Data for biochemical research,2nd ed. Clarendon Press, Oxford.
10. Koch, A. L. 1964. The role of permease in transport. Biochim.Biophys. Acta 79:177-200.
11. Lelliot, R. A. 1974. Genus XII. Erwinia, p. 332-339. In R. E.Buchanan and N. E. Gibbons (ed.), Bergey's manual of deter-minative bacteriology, 8th ed. The Williams & Wilkins Co.,Baltimore.
12. Levinthal, M. 1971. Biochemical studies of melibiose metabo-lism in wild type and mel mutant strains of Salmonellatyphimurium. J. Bacteriol. 105:1047-1052.
13. Mildenhail, J. P. 1974. Bacterial stalk rot of maize in the EasternCape Province. Fort Hare Papers 6:29-33.
14. Mildenhall, J. P., B. A. Prior, and L. A. Troilope. 1981. Waterrelations of Erwinia chrysanthemi: growth and extracellularpectic acid lyase production. J. Gen. Microbiol. 127:27-34.
15. Miller, J. H. 1972. Experiments in molecular genetics. ColdSpring Harbor Laboratory, Cold Spring Harbor, N.Y.
16. Miller, J. H. 1980. The Lacd gene, p. 31-88. In J. H. Miller andW. S. Reznikoff (ed.), The operon. Cold Spring Harbor Labo-ratory, Cold Spring Harbor, N.Y.
17. Mortlock, R. P. 1982. Metabolic acquisitions through laboratoryselection. Annu. Rev. Microbiol. 36:259-284.
18. 0rskov, I., and F. 0rskov. 1973. Plasmid determined H2Scharacter in Escherichia coli and its relation to plasmid-carriedraffinose fermentation and tetracycline resistance characters. J.Gen. Microbiol. 77:487-499.
19. Prestidge, L. S., and A. B. Pardee. 1965. A second permease formethyl-thio-o-D-galactoside in Escherichia coli. Biochim. Bio-phys. Acta 100:591-593.
20. Putnam, S. L., and A. L. Koch. 1975. Complications in thesimplest cellular enzyme assay: lysis of Escherichia coli for theassay of I-galactosidase. Anal. Biochem. 63:350-360.
21. Reeve, E. C. R. 1976. The lactose operon of Klebsiella aero-genes V9A. 5. Lac-permease defective mutants of two Kleb-siella Lac plasmids and their apparent reversion to wild type.Genet. Res. 28:61-74.
22. Reeve, E. C. R., and J. A. Braithwaite. 1970. Fk-lac, an episomewith unusual properties found in a wild strain of a Klebsiellaspecies. Nature (London) 228:162-164.
23. Reeve, E. C. R., and J. A. Braithwaite. 1973. The lactose systemin Klebsiella aerogenes V9A. 2. Galactose permeases whichaccumulate lactose or melibiose. Genet. Res. 21:273-285.
24. Reeve, E. C. R., and J. A. Braithwaite. 1973. Lac' plasmids areresponsible for the strong lactose-positive phenotype found inmany strains of Klebsiella species. Genet. Res. 22:329-333.
25. Robbie, J. P., and T. H. Wilson. 1979. Transmembrane effects ofP-galactosides on thiomethyl-,B-galactoside transport in Esche-richia coli. Biochim. Biophys. Acta 173:234-244.
26. Rotman, B., A. K. Ganesan, and R. Guzman. 1968. Transportsystems for galactose and galactosides in Escherichia coli. II.Substrate and inducer specificities. J. Mol. Biol. 36:247-260.
27. Saier, M. H., Jr. 1977. Bacterial phosphoenolpyruvate:sugarphosphotransferase systems: structural, functional, and evolu-tionary interrelationships. Bacteriol. Rev. 41:856-871.
28. Saier, M. H., Jr., M. J. Novotny, D. Comeau-Fuhnnan, T.Osumi, and J. D. Desai. 1983. Cooperative binding of the sugarsubstrates and allosteric regulatory protein (enzyme 111G1c of thephosphotransferase system) to the lactose and melibiosepermeases in Escherichia coli and Salmonella typhimurium. J.Bacteriol. 155:1351-1357.
29. Schmid, K., and R. Schmitt. 1976. Raffinose metabolism in
J. BACTERIOL.
on June 19, 2018 by guesthttp://jb.asm
.org/D
ownloaded from

E. CHRYSANTHEMI LACTOSE AND MELIBIOSE METABOLISM 891
Escherichia coli K12. Purification and properties of a newo-galactosidase specified by a transmissable plasmid. Eur. J.Biochem. 67:95-104.
30. Schmitt4 R. 1968. Analysis of melibiose mutants deficient ina-galactosidase and thiomethylgalactoside permease II in Esch-erichia coli K12. Biochem. Biophys. Res. Commun. 22:473-479.
31. Schmitt, R., and B. Rotman. 1966. a-Galactosidase activity incell-free extracts of Escherichia coli. Biochem. Biophys. Res.Commun. 22:473-479.
32. Schmitt, R., K. Schmid, and J. Roempler. 1973. Plasmid-kontrollierter Raffinose-abbau bei Escherichia coli K12. Hoppe-Seyler's Z. Physiol. Chem. 354:1241.
33. Sheinin, R., and B. F. Crocker. 1961. The induced concurrent
formation of a-galactosidase and ,B-galactosidase in Escherichiacoli B. Can. J. Biochem. Physiol. 39:63-72.
34. Starr, M. P. 1982. The genus Erwinia, p. 1260-1271. In M. P.Starr, H. Stolp, H. G. Truper, A. Balows, and H. G. Schlegel(ed.), The prokaryotes. A textbook on habitats, isolation, andidentification of bacteria, vol. 2. Springer-Verlag, New York.
35. Wilson, T. H., K. Ottina, and D. M. Wilson. 1982. Melibiosetransport in bacteria, p. 33-39. In A. N. Martonosi (ed.),Membranes and transport, vol. 2. Plenum Publishing Corp.,New York.
36. Zabin, I., and A. V. Fowler. 1980. ,B-Galactosidase, the lactosepermease protein, and thiogalactoside transacetylase, p.89-121. In J. H. Miller and W. S. Reznikoff (ed.), The operon.Cold Spring Harbor Laboratory, Cold Spring Harbor, N.Y.
VOL. 168, 1986
on June 19, 2018 by guesthttp://jb.asm
.org/D
ownloaded from