membrane-spanning and periplasmic segments of ccmi have
TRANSCRIPT

JOURNAL OF BACTERIOLOGY, Feb. 2007, p. 789–800 Vol. 189, No. 30021-9193/07/$08.00�0 doi:10.1128/JB.01441-06Copyright © 2007, American Society for Microbiology. All Rights Reserved.
Membrane-Spanning and Periplasmic Segments of CcmI HaveDistinct Functions during Cytochrome c Biogenesis in
Rhodobacter capsulatus�
Carsten Sanders, Clemence Boulay,† and Fevzi Daldal*Department of Biology, Plant Science Institute, University of Pennsylvania, Philadelphia, Pennsylvania 19104
Received 11 September 2006/Accepted 13 November 2006
In gram-negative bacteria, like Rhodobacter capsulatus, about 10 membrane-bound components (CcmABCDEFGHI and CcdA) are required for periplasmic maturation of c-type cytochromes. These components performthe chaperoning and thio-oxidoreduction of the apoproteins as well as the delivery and ligation of the hemecofactors. In the absence of any of these components, including CcmI, proposed to act as an apocytochrome cchaperone, R. capsulatus does not have the ability to produce holocytochromes c or consequently to exhibitphotosynthetic growth and cytochrome cbb3 oxidase activity. Previously, we have demonstrated that nullmutants of CcmI partially overcome cytochrome c deficiency phenotypes upon overproduction of the CcmF-R.capsulatus CcmH (CcmF-CcmHRc) couple in a growth medium-dependent manner and fully bypass thesedefects by additional overproduction of CcmG. Here, we show that overproduction of the CcmF-CcmHRc coupleand overproduction of the N-terminal membrane-spanning segment of CcmI (CcmI-1) have similar suppres-sion effects of cytochrome c maturation defects in CcmI-null mutants. Likewise, additional overproduction ofCcmG, the C-terminal periplasmic segment of CcmI (CcmI-2), or even of apocytochrome c2 also providescomplementation abilities similar to those of these mutants. These results indicate that the two segments ofCcmI have different functions and support our earlier findings that two independent steps are required for fullrecovery of the loss of CcmI function. We therefore propose that CcmI-1 is part of the CcmF-CcmHRc-dependent heme ligation, while CcmI-2 is involved in the CcdA- and CcmG-dependent apoprotein thioreduc-tion steps, which intersect at the level of CcmI during cytochrome c biogenesis.
The c-type cytochromes (cyts) have been found in virtuallyall organisms. In most microorganisms, they function primarilyas electron carriers in energy transduction pathways like res-piration (Res) and photosynthesis (Ps) (41). In some eu-karyotes, they are also involved in other fundamental cellularprocesses, such as programmed cell death (apoptosis) (27).They carry at least one covalently and stereospecifically at-tached heme (iron protoporphyrin IX) moiety via thioetherbonds between the heme vinyl groups and the cysteine thiolsof a highly conserved Cys-Xxx-Yyy-Cys-His signature motifwithin the apoproteins (41). In recent years, this complex post-translation/posttranslocation protein modification process hasbeen under intense study. So far, three evolutionarily distinctmodes of holocytochrome (holocyt) c formation have beenfound in various organisms and are called cyt c maturationsystems I, II, and III (33). The most sophisticated of thesemodes (Ccm system I) is used by �- and �-proteobacteria,deinococci, and mitochondria of plants and protozoa. In gram-negative bacteria, the Ccm system I consists of at least 10genes, designated ccmABCDEFGHI and dsbD or ccdA. All ofthese genes encode membrane-bound components that act onc-type apocytochromes (apocyts) or heme moieties following
their translocation across the cytoplasmic membrane to pro-duce holocyts c (2, 33, 51).
The gram-negative, purple nonsulfur, facultative photosyn-thetic bacterium Rhodobacter capsulatus produces a variety ofmembrane-bound and soluble c-type cyts, including cyts c1, c2,c�, cy, co, and cp, to sustain its versatile growth modes (15, 25,31, 55). Of these proteins, cyt c1 and either cyt c2 or cy arerequired for Ps growth (55), while cyts co and cp are involvedin Res growth as subunits of the cbb3-type cyt c oxidase (31),whose activity can be detected by monitoring the staining ofcolonies following the Nadi (�-naphthol � dimethylphe-nylenediamine 3 indophenol blue � H2O) reaction (30).Studies of R. capsulatus mutants which exhibit Ps�/Nadi� dualphenotypes (32) have identified various components requiredfor cyt c maturation in this species (16, 22, 36, 38). Of thesecomponents, CcmABCD form an ATP-binding cassette(ABC)-containing transporter complex, which is thought tobe required for the translocation of heme groups across thecytoplasmic membrane and for their attachment to the hemechaperone CcmE in the periplasm (24, 42, 52). Recent worksindicate that CcmABCD are also involved in subsequent ATP-dependent release of holo-CcmE providing heme groups forthe putative apocyt c heme lyase component CcmF (21, 24,42, 52).
As disulfide bonds are formed between the cysteines of ex-ported proteins via the DsbA-DsbB pathway for oxidative pro-tein folding (11, 28), similar bonds are also assumed to beformed between the cysteine thiols of the apocyt c heme bind-ing motif (37, 43). Thus, a specific thioreduction pathway (38)is thought to shuttle electrons across the cytoplasmic mem-
* Corresponding author. Mailing address: Department of Biology,Plant Science Institute, University of Pennsylvania, 103B Lynch Build-ing, Philadelphia, PA 19104. Phone: (215) 898-4394. Fax: (215) 898-8780. E-mail: [email protected].
† Present address: Centre d’etudes de Saclay, Direction des Sciencesdu Vivant, Departement de Biologie Joliot-Curie, Service de Bioen-ergetique, 91191 Gif-sur-Yvette, France.
� Published ahead of print on 22 November 2006.
789
Dow
nloa
ded
from
http
s://j
ourn
als.
asm
.org
/jour
nal/j
b on
21
Febr
uary
202
2 by
118
.105
.101
.22.

brane via CcdA (homologous to the central part of Escherichiacoli DsbD) to CcmG and then to CcmH (R. capsulatus CcmH[CcmHRc] or E. coli CcmH [CcmHEc]) in order to reduce thedisulfide bond at the apocyt c heme binding site prior to hemeligation (20, 29, 38). Finally, CcmI is proposed to chaperonethe apocyts c to their heme ligation sites following their trans-location into the periplasm (36).
In R. capsulatus and in some other species, CcmI is a bipar-tite membrane protein with two amino-terminal transmem-brane helices encompassing a leucine zipper-like motif in itscytoplasmic loop (CcmI-1 segment of 112 amino acid residuesin length) and a large periplasmic carboxyl-terminal extensionwith four tetratricopeptide repeat (TPR)-like motifs (CcmI-2segment of 312 amino acid residues in length) (36, 45) (Fig. 1).In R. capsulatus, CcmI mutants lacking the CcmI-2 segment cytc maturation become growth medium dependent. These mu-tants produce several c-type holocyts (e.g., cyts c1, cp, cy, andco) during growth on minimal medium, but they contain onlyholocyt c1 during their growth on enriched medium (36). Thisindicates that CcmI-1 is required for the production of all cytsc, whereas CcmI-2 is to some extent dispensable for cyt c1 (36,45). No homologue of the CcmI-1 segment is found in E. coliwhereas that of CcmI-2 is naturally fused C terminally to thehomologue of CcmHRc, yielding a bifunctional component re-ferred to here as CcmHEc. Remarkably, in E. coli, the lack ofa CcmI-1 segment coincides with the absence of any carboxylterminally membrane-attached c-type cyt, such as cyt c1. Re-cent genetic analyses suggest that R. capsulatus CcmI mightform together with CcmF and CcmHRc a heme ligation corecomplex catalyzing the final steps of heme attachment to apo-cyts c (45, 46).
Interestingly, complete suppression of holocyt c production
defects due to the absence of CcmI requires two distinct andadditive events: overproduction of the CcmF-CcmHRc couplethat overcomes the function of CcmI on minimal but not en-riched medium (17) and additional overproduction of CcmGthat further renders cyt c biogenesis fully proficient in all media(45). The requirement of two genetically independent stepstogether with the bipartite structure of CcmI (Fig. 1) led us tofurther investigate possible functional relationships betweenthe CcmI segments CcmF, CcmHRc, and CcmG. In this work,we demonstrate that overproduction of either the CcmF-CcmHRc couple or CcmI-1, supplemented with the overpro-duction of CcmG, CcmI-2, or even apocyt c2, fully comple-ments CcmI-null mutants for all defects in holocyt c formation.Moreover, we show that the hyperproduction of the CcmF-CcmHRc couple and hyperproduction of CcmI-1 also suppresscyt c deficiency to comparable extents. These findings indicatethat on one hand CcmF, CcmHRc, and CcmI-1 and on theother CcmG and CcmI-2 act together during cyt c maturation.Thus, the membrane-spanning and periplasmic segments ofCcmI interconnect the CcdA- and CcmG-dependent periplas-mic apoprotein thioreduction pathway to the heme ligationpathway involving the CcmF-CcmHRc couple.
MATERIALS AND METHODS
Bacterial strains, plasmids, and growth conditions. The bacterial strains andplasmids used in this work are described in Table 1. R. capsulatus strains weregrown at 35°C on enriched (MPYE) (13) or minimal (Med A) (49) medium,supplemented with appropriate antibiotics (tetracycline, kanamycin, and specti-nomycin at final concentrations of 2.5, 10, and 10 �g/ml, respectively), eitherchemoheterotrophically (Res growth) or photoheterotrophically (Ps growth) inanaerobic jars with H2-CO2-generating gas packs from BBL Microbiology Sys-tems (Cockeysville, MD). The ability of the strains to form visible colonies after3 days of incubation under appropriate growth conditions is denoted by “�”(plus), after 5 days by “slow,” and in other cases by “�” (minus). E. coli strainswere grown on Luria-Bertani (LB) broth supplemented as needed with appro-priate antibiotics (tetracycline, kanamycin, spectinomycin, and ampicillin at finalconcentrations of 12.5, 50, 10, and 100 �g/ml, respectively), as published earlier(13, 26).
Molecular genetic techniques. Standard molecular genetic techniques (44) andconjugal transfer of plasmids from E. coli to R. capsulatus (13) were performedas described elsewhere. The primers (designations and nucleotide sequences)used in this study are listed in Table 2. Plasmid pSVEN was restricted withBamHI and HindIII, and the 1.66-kb fragment containing ccmI and its nativepromoter sequence was cloned into pBluescriptKS� to yield pSVEN-KS(�).The nucleotide sequence of a FLAG epitope (DYKDDDDK) was added in frameto the 3� end of ccmI by PCR with the primers CcmI-FLAGfwd and CcmI-FLAGrevusing a QuikChange XL site-directed mutagenesis kit (Stratagene, La Jolla,CA) according to the manufacturer’s instructions, yielding pNJ1. TheccmI::FLAG-harboring allele was excised from pNJ1 with BamHI and HindIII(1.69 kb) and ligated into the respective sites of pRK415 and pBluescriptIISK�to create pNJ2 and pCS1564, respectively. Plasmid pCS1570 was constructed bydigesting pNJ2 with PstI and then religating the remaining vector fragment todelete the in-frame 126 bp of ccmI (ccmI�Q38-L79::FLAG) corresponding to aCcmI derivative lacking 42 amino acid residues of its cytoplasmic loop. PlasmidpCS1580 harboring ccmI1::FLAG, which produces a CcmI derivative corre-sponding to its N-terminal 121 amino acid residues, fused C terminally with aFLAG tag, was constructed by deleting 912 bp of its 3� end in pCS1564 via PCRamplification using the primers CS1580fwd and CS1580rev and again aQuikChange XL site-directed mutagenesis kit. Plasmid pCS1580 was restrictedwith HindIII and ligated into the same site of pRK415 to obtain pCB1. Similarly,pCS1580 was cloned into pCS1540 carrying an overexpressing ccmG allele (45)to generate pCS1589. Furthermore, a 1,396-bp PCR product containing a 3�fragment of ccmI (ccmI2::FLAG) that corresponds to a gene product lacking theN-terminal 84 amino acid residues of CcmI (i.e., its first transmembrane helixand almost its entire cytoplasmic segment) was amplified from pCS1564 by usingthe primers CS1579fwd, introducing a ribosomal binding site and a start codon,and M13-forward. This PCR product was phosphorylated, restricted with
FIG. 1. Membrane-embedded CcmI-1 and periplasmic CcmI-2 seg-ments of CcmI. The CcmI-1 segment is 112 amino acids long andcomprises the first and second transmembrane helices (black and graycylinders) of CcmI encompassing a cytoplasmic loop with a leucinezipper-like motif (light-gray oval). The CcmI-2 segment refers to the312-amino-acid-long periplasmic portion of CcmI, which contains fourTPR-like motifs (dark-gray rectangles). Q38, S64, L79, A96, and P121are landmark amino acid residues of CcmI that define modified vari-ants of CcmI, such as CcmI�Q38-L79::FLAG, CcmI-1S64::FLAG,CcmI-1A96::FLAG, and CcmI-1::FLAG, produced by the plasmidspCS1570, pCB2, pCS1588, pCB1, pCB6, and pCS1590, respectively.T84M designates the engineered start codon in CcmI to yield aCcmI-2::FLAG derivative produced by pCS1579 or pCS1583, andF175 marks the location of the spectinomycin cassette inserted intoccmI carried by pSEL12 and yielding a CcmI-F175::spe derivative, asdescribed in the text.
790 SANDERS ET AL. J. BACTERIOL.
Dow
nloa
ded
from
http
s://j
ourn
als.
asm
.org
/jour
nal/j
b on
21
Febr
uary
202
2 by
118
.105
.101
.22.
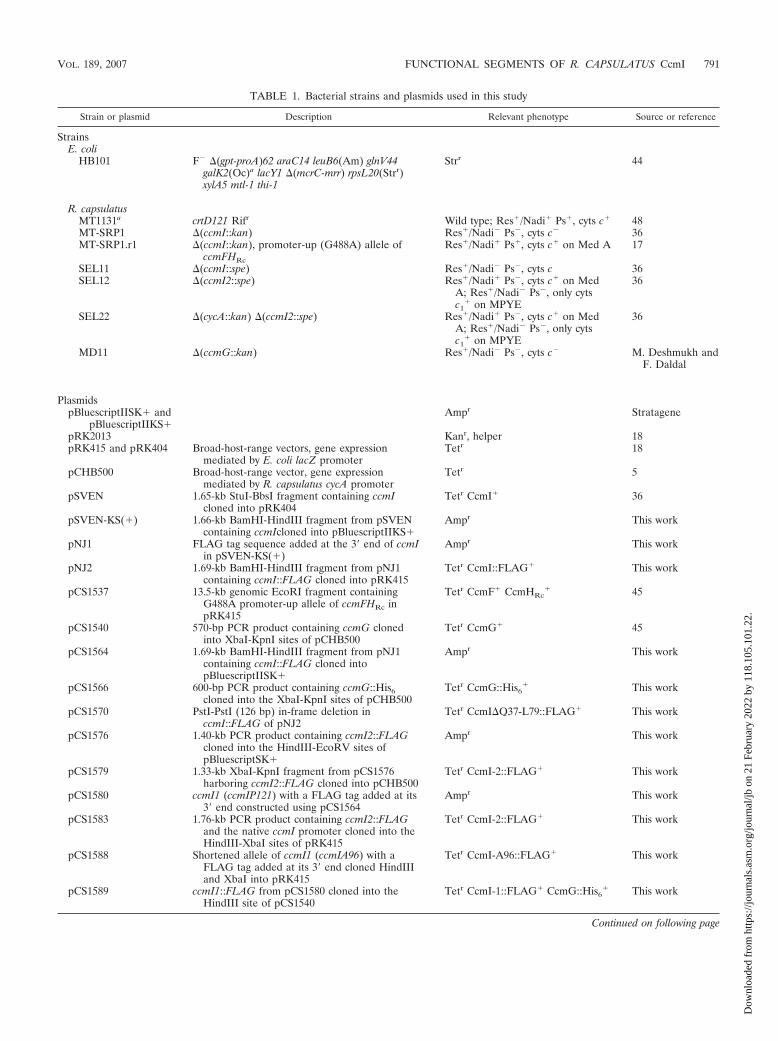
TABLE 1. Bacterial strains and plasmids used in this study
Strain or plasmid Description Relevant phenotype Source or reference
StrainsE. coli
HB101 F� �(gpt-proA)62 araC14 leuB6(Am) glnV44galK2(Oc)a lacY1 �(mcrC-mrr) rpsL20(Strr)xylA5 mtl-1 thi-1
Strr 44
R. capsulatusMT1131a crtD121 Rifr Wild type; Res�/Nadi� Ps�, cyts c� 48MT-SRP1 �(ccmI::kan) Res�/Nadi� Ps�, cyts c� 36MT-SRP1.r1 �(ccmI::kan), promoter-up (G488A) allele of
ccmFHRc
Res�/Nadi� Ps�, cyts c� on Med A 17
SEL11 �(ccmI::spe) Res�/Nadi� Ps�, cyts c 36SEL12 �(ccmI2::spe) Res�/Nadi� Ps�, cyts c� on Med
A; Res�/Nadi� Ps�, only cytsc1
� on MPYE
36
SEL22 �(cycA::kan) �(ccmI2::spe) Res�/Nadi� Ps�, cyts c� on MedA; Res�/Nadi� Ps�, only cytsc1
� on MPYE
36
MD11 �(ccmG::kan) Res�/Nadi� Ps�, cyts c� M. Deshmukh andF. Daldal
PlasmidspBluescriptIISK� and
pBluescriptIIKS�Ampr Stratagene
pRK2013 Kanr, helper 18pRK415 and pRK404 Broad-host-range vectors, gene expression
mediated by E. coli lacZ promoterTetr 18
pCHB500 Broad-host-range vector, gene expressionmediated by R. capsulatus cycA promoter
Tetr 5
pSVEN 1.65-kb StuI-BbsI fragment containing ccmIcloned into pRK404
Tetr CcmI� 36
pSVEN-KS(�) 1.66-kb BamHI-HindIII fragment from pSVENcontaining ccmIcloned into pBluescriptIIKS�
Ampr This work
pNJ1 FLAG tag sequence added at the 3� end of ccmIin pSVEN-KS(�)
Ampr This work
pNJ2 1.69-kb BamHI-HindIII fragment from pNJ1containing ccmI::FLAG cloned into pRK415
Tetr CcmI::FLAG� This work
pCS1537 13.5-kb genomic EcoRI fragment containingG488A promoter-up allele of ccmFHRc inpRK415
Tetr CcmF� CcmHRc� 45
pCS1540 570-bp PCR product containing ccmG clonedinto XbaI-KpnI sites of pCHB500
Tetr CcmG� 45
pCS1564 1.69-kb BamHI-HindIII fragment from pNJ1containing ccmI::FLAG cloned intopBluescriptIISK�
Ampr This work
pCS1566 600-bp PCR product containing ccmG::His6cloned into the XbaI-KpnI sites of pCHB500
Tetr CcmG::His6� This work
pCS1570 PstI-PstI (126 bp) in-frame deletion inccmI::FLAG of pNJ2
Tetr CcmI�Q37-L79::FLAG� This work
pCS1576 1.40-kb PCR product containing ccmI2::FLAGcloned into the HindIII-EcoRV sites ofpBluescriptSK�
Ampr This work
pCS1579 1.33-kb XbaI-KpnI fragment from pCS1576harboring ccmI2::FLAG cloned into pCHB500
Tetr CcmI-2::FLAG� This work
pCS1580 ccmI1 (ccmIP121) with a FLAG tag added at its3� end constructed using pCS1564
Ampr This work
pCS1583 1.76-kb PCR product containing ccmI2::FLAGand the native ccmI promoter cloned into theHindIII-XbaI sites of pRK415
Tetr CcmI-2::FLAG� This work
pCS1588 Shortened allele of ccmI1 (ccmIA96) with aFLAG tag added at its 3� end cloned HindIIIand XbaI into pRK415
Tetr CcmI-A96::FLAG� This work
pCS1589 ccmI1::FLAG from pCS1580 cloned into theHindIII site of pCS1540
Tetr CcmI-1::FLAG� CcmG::His6� This work
Continued on following page
VOL. 189, 2007 FUNCTIONAL SEGMENTS OF R. CAPSULATUS CcmI 791
Dow
nloa
ded
from
http
s://j
ourn
als.
asm
.org
/jour
nal/j
b on
21
Febr
uary
202
2 by
118
.105
.101
.22.

HindIII, and cloned into pBluescriptSK� digested with EcoRV and HindIII,yielding pCS1576. Plasmid pCS1579 was then constructed by restricting pCS1576with XbaI and KpnI and by cloning of the 1,332-bp ccmI2::FLAG-containingfragment into the respective sites of pCHB500. For pCS1566, a 604-bp PCRproduct harboring ccmG was amplified by using the primers helXforward-XbaIand 1566rev, carrying the recognition sites for XbaI and KpnI at their 5� ends,and cloned into the respective sites of pCHB500 containing ccmG::His6 with itsnative ribosomal binding site and expressed from the R. capsulatus cycA pro-moter.
Two-step PCR amplifications were carried out to construct pCS1583,pCB2, pCS1588, pCS1590, and pCB5. As does pCS1579, pCS1583 containsccmI2::FLAG but expresses this CcmI allele from its native ccmI promoter. Thefirst-round PCR used pCS1564 as a template and the primers M13-reverse andCS1583rev to yield a 466-bp fragment and the primers CS1583fwd and M13fwd
to generate a 1,304-bp fragment. These fragments were purified with a QIAquickPCR purification kit (QIAGEN, Valencia, CA) and used as DNA templates withcomplementary sequences of at least 15 bp for a second-round PCR using theprimers M13-reverse and M13-forward. The latter PCR fragment of 1,755 bp wasrestricted with HindIII and XbaI and ligated into the respective site of pRK415to finally create pCS1583. For pCB2, the first round of PCR used pCS1580 as atemplate and the primers M13-reverse and CB2rev or the primers CB2fwd andM13-forward to produce a 658-bp and a 284-bp fragment, respectively. Thefirst-round PCR amplification for pCS1588 was also done using pCS1580 as atemplate and the primers M13-reverse and 1588rev or the primers 1588fwd andM13-forward to generate a 754-bp and a 284-bp fragment, respectively. In bothcases, these fragments were purified, as described for pCS1583 above, and usedas DNA templates for a second round of PCR using the primers M13-reverse andM13-forward. The latter PCR fragments for the construction of pCB2 (927 bp)
TABLE 1—Continued
Strain or plasmid Description Relevant phenotype Source or reference
pCS1590 1.00-kb PCR product containing a gene fusion ofccmI1::FLAG and ccmG::His6 cloned into theXbaI-KpnI sites of pRK415
Tetr CcmI-1::FLAG-CcmG::His6� This work
pCS1591 660-bp PCR product containing a gene fusion ofccmI1::FLAG cloned into the XbaI-KpnI sitesof pCHB500
Tetr CcmI-1::FLAG� This work
pCB1 ccmI1::FLAG from pCS1580 cloned into theHindIII site of pRK415
Tetr CcmI-1::FLAG� This work
pCB2 Shortened allele of ccmI1 (ccmIS64) with aFLAG tag added at its 3� end cloned into theHindIII-XbaI sites of pRK415
Tetr CcmI-S64::FLAG� This work
pCB5 1.00-kb PCR product containing a gene fusion ofccmI1::FLAG and ccmG::His6 cloned into theXbaI-KpnI sites of pCHB500
Tetr CcmI-1::FLAG-CcmG::His6� This work
pCB6 660-bp PCR product containing a gene fusion ofccmI1::FLAG cloned into the XbaI-KpnI sitesof pCHB500
Tetr CcmI-1::FLAG� This work
pHM14 1.25-kb BamHI-HindIII fragment containing R.capsulatus cycA in pRK415
Tetr, cyt c2� 39
pFJ63 1.20-kb MluI-HindIII fragment containing R.capsulatus cycY in pRK404
Tetr, cyt cy� 26
pMT0-404 R. capsulatus petABC gene cluster in pRK404 Tetr, cyt bc1� 4
pOX15 R. capsulatus ccoNOQP gene cluster in pRK404 Tetr, cyt cbb3� 31
a R. capsulatus MT1131 is referred to as a wild-type strain with respect to its cyt c profile and growth properties.
TABLE 2. Designations and nucleotide sequences of primers used in this study
Designation Nucleotide sequence (5� to 3�)
M13-forward.......................................................................................GTA AAA CGA CGG CCA GTM13-reverse........................................................................................GGA AAC AGC TAT GAC CAT GCcmI-FLAGfwd.................................................................................CAC GAT CGG AGC CGC GCC AGA CTA CAA GGA CGA CGA TGA
CAA GTG ATC TGC CCC GGG ATCCcmI-FLAGrev..................................................................................GAT CCC GGG GCA GAT CAC TTG TCA TCG TCG TCC TTG TAG
TCT GGC GCG GCT CCG ATCRccycH-XbaIf ....................................................................................CTC TCT AGA TGC AGG ATA GGA GAC CCC ATGhelXforward-XbaI .............................................................................CGG GTC TAG AAA GCC CAG GAA GAA CGG ACS1566rev...........................................................................................GAG GGT ACC AAG CTT AGT GAT GGT GAT GGT GAT GAG ATC
TAT CCG CCG TCC CGG GCC AGA AGC GGGCS1579fwd..........................................................................................GCG CTG CAG GAG GCG GGC ATG GCG GCG GCC GAG Ga
CS1580fwd..........................................................................................CCG GCC GGT CTG CCC GAC TAC AAG GAC GAC GAT GCS1580rev...........................................................................................GTC GTC CTT GTA GTC GGG CAG ACC GGC CGG GCC GCS1583fwd..........................................................................................GAT AGG AGA CCC CAT GGC GGC GGC CGA GGC GCC GCS1583rev...........................................................................................CGC CTC GGC CGC CGC CAT GGG GTC TCC TAT CCTCS1588fwd..........................................................................................CGC GGC ACG GTG GCG GAC TAC AAG GAC GAC GAC GACCS1588rev...........................................................................................GTC GTC CTT GTA GTC CGC CAC CGT GCC GCG CCG CGGCB2fwd ...............................................................................................GAG GCG GCG CGG TCG GAC TAC AAG GAC GAC GAT GAC AAG TCB2rev ................................................................................................GTC GTC CTT GTA GTC CGA CCG CGC CGC CTC TTC CGG GCT CCB5fwd ...............................................................................................GAC GAC GAT GAC AAG CAG CAA AAT GAC CCG AAT GCC ATG CCB5rev ................................................................................................CGG GTC ATT TTG CTA CTT GTC ATC GTC GTC CTT GTA GTC G
a Underlining indicates the newly introduced ribosome binding site and start codon at the 5� end of ccmI2 in pCS1579 (Table 1).
792 SANDERS ET AL. J. BACTERIOL.
Dow
nloa
ded
from
http
s://j
ourn
als.
asm
.org
/jour
nal/j
b on
21
Febr
uary
202
2 by
118
.105
.101
.22.

FIG. 2. Restriction maps of various plasmids carrying different alleles of ccmI, ccmF-ccmHRc, ccmG, or cycA, and their related phenotypes inselected CcmI-null mutants. Restriction maps of various plasmids and Nadi�, Ps�, or Psslow phenotypes that they confer to the CcmI-null mutantsMT-SRP1 and MT-SRP1.r1 (Table 1) on minimal (Med A) or enriched (MPYE) media are shown on the left and right, respectively. Plasmid pNJ2carries the entire 1,272-bp ccmI gene (ccmI::FLAG) and pCS1570 a 126-bp in-frame deletion (ccmI�Q38-L79::FLAG). Plasmids pCB2 andpCS1588 contain 192-bp and 288-bp 5� fragments of ccmI (ccmI1S64::FLAG and ccmI1A96::FLAG), respectively, and pCB1, pCB6, and pCS1590harbor a 363-bp 5� fragment of ccmI (ccmI1::FLAG) corresponding to the entire CcmI-1 segment. Plasmid pSEL12 carries a spectinomycin cassetteinserted into the XhoI site of ccmI and expresses a truncated allele (ccmIF175::spe). Plasmids pCS1583 and pCS1579 both contain a ccmI fragmentlacking its first 249 bp, with an exogenous ribosomal binding site and a start codon created at its position T84 to ensure the expression of theremaining part of ccmI to yield a derivative corresponding to its CcmI-2 segment. Plasmid pCS1537 carries an up-regulated ccmFHRc (ccmFHRc
up)allele, and pCS1540 and pHM14 harbor ccmG and cycA, respectively. Finally, pCS1589 carries both a ccmG allele as in pCS1540 and a ccmI1 alleleas in pCB1. All ccmI derivatives, except that produced by pSEL12, are fused to a FLAG tag sequence at their 3� ends (indicated by a white rectanglewith a black X). The promoters of ccmI, E. coli lacZ, cycA, and up-regulated ccmFHRc that drive the expression of downstream genes are indicatedas white arrows, black arrows, gray arrows, and striped arrows, respectively. Note that the ccmI derivatives in pNJ2, pCS1570, pCB2, pCS1588,pCB1, pSEL12, pCS1583, and pCS1589 are expressed from their native ccmI promoter, while those in pCB6, pCS1579, and pCB5 are from theR. capsulatus cycA promoter and those in pCS1590 and pCS1591 are from the E. coli lacZ promoter. A, B, E, H, K, P, and X refer to cleavagesites of the restriction enzymes XhoI, BamHI, EcoRI, HindIII, KpnI, PstI, and XbaI, respectively. Numbers 1 and 2 indicate the patterns ofrestriction sites E, K, and B and X, B, P, E, K, and B, respectively, and double dashes refer to truncations used to fit the figure, as appropriate.
VOL. 189, 2007 FUNCTIONAL SEGMENTS OF R. CAPSULATUS CcmI 793
Dow
nloa
ded
from
http
s://j
ourn
als.
asm
.org
/jour
nal/j
b on
21
Febr
uary
202
2 by
118
.105
.101
.22.

and pCS1588 (1,023 bp) were restricted with HindIII and XbaI and then ligatedinto the same sites of pRK415 containing ccmI derivatives (ccmIS64::FLAG orccmIA96::FLAG) that correspond to gene products of the N-terminal 64 and 96amino acid residues of CcmI with C-terminal FLAG tags. The plasmids pCS1590and pCB5 were constructed similarly. In the first round of PCR, a 414-bpfragment was amplified using pCS1580 as a template and the primers RccycH-XbaIf and CB5rev, and a 594-bp fragment was amplified using pCS1566 as atemplate and the primers CB5fwd and M13-forward. This yielded a PCR productcorresponding to a CcmG derivative without its native N-terminal signal se-quence and membrane anchor. These PCR fragments were purified and used asDNA templates for a second round of PCR using the primers RccycH-XbaIf andM13-forward, generating a 1,003-bp fragment, which was then restricted withXbaI and KpnI, ligated into the same sites of pRK415 and pCHB500 harboringa ccmI1::FLAG-ccmG::His6 gene fusion with the ccmI ribosomal binding site,and expressed from different promoters.
Finally, pCS1591 and pCB6 were created by PCR amplification from pCS1580using the primers RccycH-XbaIf and M13-forward, restricting the produced660-bp fragment with XbaI and KpnI and ligating it into the corresponding sitesof the pRK415 and pCHB500. These constructs contained ccmI1::FLAG with itsnative ribosomal binding site but expressed from two different promoters, asdescribed above. All plasmids were verified by DNA sequencing using appropri-ate primers, and DNA sequence analyses, homology searches, and genomesequence comparisons were done using MacVector from Accelrys (San Diego,CA) and BLAST software packages (3). Note that the ccmI alleles in pNJ2,pCS1570, pCB2, pCS1588, pCB1, pSEL12, pCS1583, and pCS1589 are expressedfrom the native ccmI promoter, those in pCB6, pCS1579, and pCB5 from the R.capsulatus cycA promoter of pCB500 derivatives, and those in pCS1590 andpCS1591 from the E. coli lacZ promoter of pRK415 derivatives.
Biochemical techniques. Small-scale crude cell extracts were prepared by usingCelLytic B2X cell lysis reagent (Sigma-Aldrich, St Louis, MO) according to themanufacturer’s suggestions, modified as follows. Ten-ml cultures were centri-fuged for 10 min at 5,000 � g and 4°C, and cell pellets were washed with 2 ml ofTNE buffer (25 mM Tris-HCl, pH 7.5, 150 mM NaCl, 50 mM EDTA, 0.1 mMamino-caproic acid, and 0.1 mM Pefabloc SC [Sigma-Aldrich, St. Louis, MO]).CelLytic B2X cell lysis reagent (200 �l) supplemented with a protease inhibitormixture (final concentration, 50 mM EDTA, 0.1 mM amino-caproic acid, and 0.1mM Pefabloc SC) was added to the pellets, which were resuspended by vortexingfor 2 min before lysozyme (final concentration, 0.2 mg/ml) was added. Afterincubation for 10 min at 25°C, the solution was sonicated (microtip, 60% power,50% pulse) for 30 s using a Sonic Dismembrator 550 (Fisher Scientific, Pitts-burgh, PA) and centrifuged for 10 min at 20,000 � g and 4°C. The supernatantswere used for further protein analyses.
Intracytoplasmic membrane vesicles (chromatophores) were prepared in TNEbuffer by two passages through a French pressure cell at 18,000 lb/in2. Afterelimination of the disrupted cells by centrifugation for 1 h at 20,000 � g and 4°C,supernatants were recentrifuged for 2 h at 150,000 � g and 4°C and membranepellets used for further analyses. Protein concentrations were determined ac-cording to the method of Wiechelman et al. (53) or Bradford (7) by use of abicinchoninic acid kit (Sigma-Aldrich, St. Louis, MO) or bio-safe Coomassiesolution (Bio-Rad, Hercules, CA), respectively. Sodium dodecyl sulfate-polyacryl-amide gel electrophoresis (SDS-PAGE) was performed according to Lae-mmli (34) or Schaegger and von Jagow (47) as appropriate, and the c-type cytswere revealed by determining their endogenous peroxidase activity using3,3�,5,5�-tetramethylbenzidine (TMBZ) and H2O2 (50). Cyt c oxidase activities ofcolonies were detected on agar plates using the Nadi reaction as described above(30). Rabbit antisera against CcmG and CcmHRc used for immunoblot analyseswere described elsewhere (23, 38). Rabbit anti-FLAG immunoglobulin G poly-
clonal antibody (Sigma, St. Louis, MO) was used to detect the FLAG epitopefused to various CcmI derivatives. Monoclonal anti-rabbit immunoglobulin cloneRG-16 alkaline phosphatase conjugate with BCIP (5-bromo-4-chloro-3-in-dolylphosphate)–nitroblue tetrazolium liquid substrate (Sigma-Aldrich, St.Louis, MO) and stabilized goat anti-rabbit immunoglobulin G horseradish per-oxidase conjugate with SuperSignal West Pico chemiluminescent substrate(Pierce, Rockford, IL) were used as secondary antibodies.
Protein purification. Intracytoplasmic membrane vesicles were resuspendedand homogenized in TNE buffer and solubilized for 1 h at 4°C by addition ofdodecylmaltoside (DDM) to the continuously stirring membrane protein solu-tion (1 mg protein/mg DDM; final protein concentration, 1 mg/ml). After cen-trifugation for 30 min at 20,000 � g and 4°C, TNE buffer was exchanged withTNED buffer (25 mM Tris-HCl, pH 7.5, 150 mM NaCl, 2 mM EDTA, and 0.1%[wt/vol] DDM) by using a PD-10 column (Amersham-Pharmacia, Piscataway,NJ). Then, the solution was applied to an anti-FLAG agarose column (Sigma-Aldrich, St. Louis, MO), preequilibrated with 20 ml TNED buffer per ml ofcolumn matrix. Solubilized membrane protein suspensions were passed throughthe column three times, after which the column was washed twice with 5 ml ofTNED buffer. FLAG-tagged CcmI derivatives were subsequently eluted with 5column volumes of TNED buffer supplemented with 0.1 mg per ml of FLAGpeptide (Sigma-Aldrich, St. Louis, MO).
Chemicals. All chemicals were of reagent grade and obtained from commer-cial sources.
RESULTS
The cytoplasmic loop of the membrane-spanning segment ofCcmI is required for efficient cytochrome c biogenesis. In orderto investigate the specific roles of the different CcmI segmentsand motifs (Fig. 1) during cyt c biogenesis, we have constructedvarious ccmI alleles fused to FLAG tag sequences at their 3�ends (Fig. 2) and introduced them into �ccmI mutants MT-SRP1 (CcmI�) and MT-SRP1.r1 (CcmI� CcmFup CcmHRc
up)(where CcmFup and CcmHRc
up denote up-regulated alleles ofCcmF and CcmHRc, respectively) (Table 3), as appropriate.The cytoplasmic loop of CcmI contains a leucine zipper-likemotif, which is implicated in protein-protein interactions (35,40), but its importance for CcmI function is unknown. Twoplasmids, pNJ2 carrying an intact ccmI (ccmI::FLAG) andpCS1570 harboring an allele (ccmI�Q38-L79::FLAG) encod-ing a gene product lacking 42 amino acid residues (Fig. 1, Q38to L79) of the cytoplasmic loop within the CcmI-1 segment(Fig. 2, rows 1 and 2), were constructed, and both plasmidsmediated Ps� and Nadi� phenotypes to a CcmI-null mutant(MT-SRP1) on enriched and minimal media. We noted thatthe latter plasmid exhibited a weaker Ps growth phenotype(Psslow) on enriched medium than the former and that onlypNJ2, not pCS1570, conferred a Ps� phenotype to a doublemutant (SEL22) lacking both the CcmI-2 segment and cyt c2
(36) on all media (Table 3). Thus, the cytoplasmic loop ofCcmI-1, although not absolutely essential for cyt c maturation,
TABLE 3. Phenotypes of various CcmI-null mutants and their derivatives
Strain Relevant characteristic(s)Result witha:
Med A Res/Nadi Ps MPYE Res/Nadi Ps
MT1131 CcmI� CcmF� CcmHRc� �/� � �/� �
MT-SRP1 or SEL11 CcmI� CcmF� CcmHRc� �/� � �/� �
MT-SRP1.r1 CcmI� CcmFup CcmHRcup �/� � �/� �
SEL12 CcmI-2� �/� � �/� �SEL22 or FJM13 CcmI-2� cyt c2
� �/� � �/� �
a Med A and MPYE refer to minimal and enriched growth media, respectively. Res and Ps indicate respiratory and photosynthetic growth conditions, and � and� refer to the ability of the strains to grow under these conditions as defined in Materials and Methods. Nadi � and � indicate the presence and absence of the cytc oxidase activity, respectively.
794 SANDERS ET AL. J. BACTERIOL.
Dow
nloa
ded
from
http
s://j
ourn
als.
asm
.org
/jour
nal/j
b on
21
Febr
uary
202
2 by
118
.105
.101
.22.

becomes important under Ps growth in the absence of cyt c2,which then relies on adequate production of the membrane-bound cyts c1 and cy.
Immunoblot analyses using polyclonal antibodies against theFLAG epitope, CcmHRc, and CcmG were carried out, and thedata indicate that both pNJ2 and pCS1570 produced similarlevels of CcmI in MT-SRP1 (Fig. 3A, lanes 3 and 4). Wenote that, although the theoretical molecular masses forCcmI::FLAG and CcmI-�Q38-L79::FLAG are 45.5 and 40.7kDa, FLAG-specific signals were detected at higher molecularmasses of 55 and 50 kDa, respectively. Interestingly, muchhigher levels of CcmI::FLAG were present in MT-SRP1.r1,which overproduces CcmF and CcmHRc and substitutes par-tially for the function of CcmI during cyt c maturation (Fig. 4A,lanes 3 and 4). Additional experiments suggest that the levels
of CcmHRc and CcmG, known to have different suppressioneffects on holocyt c production when CcmI is absent (17, 45),were comparable to wild type-levels (Fig. 3B and C and 4C,lanes 1, 3, and 4), except for the levels of CcmHRc in thetransconjugant of MT-SRP1.r1, which are much higher, as thisstrain already overexpresses ccmHRc genomically (Fig. 4B,lanes 1, 3, and 4).
Next, the cyt c profiles of strains harboring either pNJ2 orpCS1570 were analyzed using TMBZ and SDS-PAGE. Theamounts of membrane-bound cyts cp, c1, cy, and co were lowerin the CcmI-null mutants expressing ccmI�Q38-L79::FLAGfrom pCS1570 than in those expressing ccmI::FLAG frompNJ2 and the wild type (Fig. 3D and 4D, lanes 1, 3, and 4),while the amounts of soluble cyt c2 were similar in the MT-
FIG. 3. Cytochrome c profiles and immunoblot analyses of variousCcmI mutants grown on minimal medium. Crude cell extracts of R.capsulatus strains grown in minimal medium under Res conditionswere subjected to SDS-PAGE using approximately 100 �g of proteinsper lane. Immunoblot analyses using polyclonal antibodies against theFLAG epitope of CcmI derivatives (A), CcmHRc (B), and CcmG(C) were performed, and holocyts c were detected via their endoge-nous peroxidase activity using TMBZ and H2O2 (D and E) as de-scribed in Materials and Methods. Lanes 1 to 9 correspond to crudecell extracts of the wild type (MT1131/pRK415), �ccmI (MT-SRP1/pRK415), ccmI::FLAG (MT-SRP1/pNJ2), ccmI�Q38-L79::FLAG(MT-SRP1/pCS1570), ccmI1S64::FLAG (MT-SRP1/pCB2),ccmI1A96::FLAG (MT-SRP1/pCS1588), ccmI1::FLAGccmI expressedby its native ccmI promoter from a composite plasmid (MT-SRP1/pCB1), ccmI1::FLAGcycA expressed by the cycA promoter from apRK415 derivative (MT-SRP1/pCB6), and ccmFHRc
up (MT-SRP1/pCS1537), respectively. Molecular mass markers (in kDa) are indi-cated on the left. Note that the two bands around the 30-kDa marker,indicated by an asterisk on the left, were detected due to their perox-idase activities and are not specific to the antibodies used.
FIG. 4. Cytochrome c profiles and immunoblot analyses of variousCcmI mutants overproducing the CcmF-CcmHRc couple grown onenriched medium. Crude cell extracts of R. capsulatus strains grown inenriched medium under Res conditions were subjected to SDS-PAGEusing approximately 100 �g of proteins per lane. Immunoblot analysesusing polyclonal antibodies against the FLAG epitope of CcmI deriv-atives (A), CcmHRc (B), and CcmG (C) were performed, and holocytsc were detected via their endogenous peroxidase activity using TMBZand H2O2 (D and E), as described in Materials and Methods. Lanes 1to 9 correspond to crude cell extracts of the wild type (MT1131/pRK415), �ccmI ccmFHRc
up (MT-SRP1.r1/pRK415), ccmI::FLAG(MT-SRP1.r1/pNJ2), ccmI�Q38-L79::FLAG (MT-SRP1.r1/pCS1570),ccmI::FLAGccmI expressed by its native ccmI promoter from a com-posite plasmid (MT-SRP1.r1/pCB1), ccmI1::FLAGcycA expressed bythe cycA promoter from a pRK415 derivative (MT-SRP1.r1/pCB6),ccmI2::FLAG (MT-SRP1.r1/pCS1579), ccmFHRc
up (MT-SRP1.r1/pCS1537), and ccmG (MT-SRP1.r1/pCS1540), respectively. Molecularmass markers (in kDa) are shown on the left. Note that the two bandsaround the 30-kDa marker, indicated by an asterisk on the left, weredetected due to their peroxidase activities and are not specific to theantibodies used.
VOL. 189, 2007 FUNCTIONAL SEGMENTS OF R. CAPSULATUS CcmI 795
Dow
nloa
ded
from
http
s://j
ourn
als.
asm
.org
/jour
nal/j
b on
21
Febr
uary
202
2 by
118
.105
.101
.22.

SRP1 transconjugants but still lower than amounts in the wildtype (Fig. 3E and 4E, lanes 3 and 4). Therefore, overall dataindicate that the cytoplasmic segment of CcmI with its leucinezipper-like motif is important for efficient holocyt c productionand is more critical for sufficient production of membrane-bound holocyt cy than soluble holocyt c2.
An intact CcmI-1 segment is necessary to suppress porphy-rin excretion and to partially bypass cyt c deficiency pheno-types of a CcmI-null mutant. Various R. capsulatus mutantsdeficient in cyt c maturation excrete heme precursors (uropor-phyrins and protoporphyrins) when grown under Res condi-tions (16, 23). A similar phenotype is also observed with CcmI-null mutants, like MT-SRP1 and SEL11, but not with CcmImutants lacking only its CcmI-2 segment, such as SEL12,SEL22, or FJM13, which produce at least holocyt c1 (36; alsothis work). For Sinorhizobium meliloti, CcmI-null mutants ex-pressing the N-terminal transmembrane helix and the adjacentcytoplasmic loop of the CcmI-1 segment, although still defec-tive for cyt c maturation, alleviate the associated protoporphy-rin IX accumulation phenotype (9). We therefore testedwhether in R. capsulatus the N-terminal part of CcmI is alsoinvolved in porphyrin excretion. Of the three plasmids con-structed, pCB2 and pCS1588 (Fig. 2, rows 3 and 4) producethe N-terminal 64 and 96 amino acid residues of CcmI(CcmI-S64::FLAG and CcmI-A96::FLAG), respectively.These CcmI derivatives contain only the first transmembranehelix of CcmI-1 and portions of the cytoplasmic loop (Fig. 1,S64 and A96). The third plasmid, pCB6 (Fig. 2, row 7), ex-presses via the exogenous cycA promoter the first 121 residuesof CcmI, which include both of the transmembrane helices andthe entire cytoplasmic loop of CcmI-1 (CcmI-1::FLAG) (Fig.1, P121). While pCB2 and pCS1588 could not complement theCcmI-null mutant MT-SRP1 for Ps� or Nadi� phenotypes oneither minimal or enriched medium, pCB6 did so for Nadi� onboth media and for Ps� on minimal medium only. When theporphyrin excretion phenotype was monitored under UV lightat 312 nm by the formation of red fluorescence surroundingcolonies, the transconjugants carrying pCB2 and pCS1588 stillexcreted heme precursors, while those harboring pCB6 did notexhibit this phenotype, like the CcmI-2-null mutants.
Next, immunoblot analyses were carried out using crudeextracts of cells grown by respiration in minimal medium toreassure that various FLAG-tagged CcmI derivatives were pro-duced adequately in appropriate strains tested. Proteins ofapparent molecular masses of 12, 14, and 15 kDa (Fig. 3A and4A, lanes 5, 6, and 8), in all cases higher than the respectivetheoretical molecular masses of 7.9 kDa for CcmI-S64::FLAG,11.2 kDa for CcmI-A96::FLAG, and 13.5 kDa forCcmI-P121::FLAG, respectively, were detected. In addition,similar analyses indicate that, in the strains tested, the levels ofCcmHRc (Fig. 3B, lanes 5, 6, and 8) were similar to those seenin a wild-type control (Fig. 3B, lanes 1 and 2). In contrast, thelevels of CcmG were lower in the wild-type strain and in theMT-SRP1 transconjugant containing pCB6 (Fig. 3C, lanes 1and 8) than in those harboring pCB2 and pCS1588 (Fig. 3C,lanes 5 and 6). This observation indicates that the CcmG levelsare increased in the absence of CcmI and hence of cyt cproduction. Finally, determination of the cyt c profiles of ap-propriate strains confirmed that the membrane-bound cyts cp,c1, cy, and co as well as the soluble cyt c2 could be detected in
cell extracts of MT-SRP1 transconjugants carrying only pCB6and not pCB2 and pCS1588 (Fig. 3D and E, lanes 8, 5, and 6,respectively), in agreement with their Nadi, Ps, and heme pre-cursor excretion phenotypes. Therefore, overall data provideevidence that the presence of an intact CcmI-1 segment com-prised of the first and second transmembrane helices and theleucine zipper-like motif-containing cytoplasmic loop is theminimal topological unit necessary to alleviate porphyrin ex-cretion and to confer cyt c production in R. capsulatus.
Overproduction of CcmI-1, like that of CcmF and CcmHRc,partially overcomes the cyt c maturation defects of CcmI-nullmutants. The fact that the Nadi� and Ps� phenotypes of aCcmI-null mutant overproducing CcmI-1 (MT-SRP1/pCB6)are identical to those of a CcmI-null mutant overproducingCcmF and CcmHRc, like MT-SRP1.r1 (17), suggests that themembrane-spanning segment of CcmI and the CcmF-CcmHRc
couple might play similar roles during cyt c maturation. Thus,three differently replicating plasmids, pCB1, pSEL12, andpCS1591 (Fig. 2, rows 5, 6, and 8), expressing differentamounts of CcmI-1 (due to different promoters and plasmidcopy numbers) were constructed to further probe to what ex-tent these components might substitute for each other, de-pending on their production levels. Plasmid pSEL12 (36) con-tains a spectinomycin cassette insertion in the periplasmicsegment of CcmI and produces a CcmI derivative with itsentire CcmI-1 segment and a short stretch of CcmI-2 but lack-ing the four TPR-like motifs (Fig. 1, F175). Plasmids pCB1 andpCS1591 express a CcmI derivative identical to that producedby pCB6 (Fig. 1, P121), but the latter plasmid lacks the exog-enous cycA promoter, while the former is a composite plasmidassumed to be of higher copy number, due to the cointegrationof ColE1 and RP4 replicons (Fig. 2). All three plasmids con-ferred to a CcmI-null mutant like MT-SRP1 Nadi and Psphenotypes similar to that conferred by pCB6, indicating thatthey all produced CcmI-1. However, pCB1 was also able tomediate to MT-SRP1 a Psslow phenotype on enriched medium.Moreover, all plasmids rendered MT-SRP1.r1 (CcmI� CcmFup
CcmHRcup) Nadi�/Ps� on both media, but only pCB1 comple-
mented SEL22, a double mutant lacking the CcmI-2 segmentand cyt c2 (21), for Ps� growth on minimal medium and Psslow
growth on enriched medium. This suggests that pCB1, whichalso acted differently than pCS1537 carrying an up-regulatedallele of ccmFHRc (55), might overproduce CcmI-1. In addi-tion, comparison of the cyt c profiles of strains harboring pCB1or pCB6 grown by respiration reveals that membrane-boundcyts cp, c1, cy, and co and soluble cyt c2 (Fig. 3D and E, lanes7 and 8, and Fig. 4D and E, lanes 5 and 6) were all produced,although their levels were lower than those detected in a wild-type strain (Fig. 3D and E and 4D and E, lanes 1), as previouslyseen with strains overproducing CcmF and CcmHRc (17).
Immunoblot analyses using polyclonal antibodies against theFLAG epitope indeed showed that the strains harboring pCB1produced a CcmI derivative with an apparent molecular massof 15 kDa at an amount higher than that seen with pCB6 (Fig.3A, lanes 7 and 8, and 4A, lanes 5 and 6), confirming that thisoverproduction was responsible for the additional Psslow phe-notype mediated by pCB1. Additional immunoblot analysesindicated that the levels of CcmHRc in strains carrying pCB1 orpCB6 differed neither among each other (Fig. 3B, lanes 7 and8, and Fig. 4B, lanes 5 and 6) nor compared to their respective
796 SANDERS ET AL. J. BACTERIOL.
Dow
nloa
ded
from
http
s://j
ourn
als.
asm
.org
/jour
nal/j
b on
21
Febr
uary
202
2 by
118
.105
.101
.22.

controls (Fig. 3B and 4B, lane 2). These levels were signifi-cantly higher in strains like MT-SRP1.r1 (Fig. 3B, lane 9, andFig. 4B, lane 8) that contained higher levels of CcmF andCcmHRc than did a wild-type strain (Fig. 4B, lanes 1 and 2),due to the overexpression of the respective chromosomal genecopies (21). On the other hand, the levels of CcmG in theCcmI-null mutants harboring pCB1, pCB6, or pCS1537 re-mained comparable to those seen in a wild-type strain (Fig. 3C,lanes 1, 7, 8, and 9, and 4C, lanes 1, 5, 6, and 8). Clearly, overalldata indicate that overproduction of either CcmI-1 or theCcmF-CcmHRc couple suppresses, either partially or fully (de-pending on the produced amounts), cyt c maturation defectsencountered in CcmI-null mutants, suggesting that the CcmI-1segment and the CcmF-CcmHRc couple function together dur-ing cyt c biogenesis.
The CcmI-2 segment and CcmG play similar roles to bypasscompletely the need for CcmI during cyt c biogenesis. Thefindings that the CcmF-CcmHRc couple and CcmI-1 act simi-larly and that additional overproduction of CcmG overcomesCcmI defects in all growth media led us to inquire whether theCcmI-2 segment and CcmG might also have similar functionsduring cyt c maturation. Plasmids pCS1583 and pCS1579 (Fig.2, rows 9 and 10), producing different amounts of the sameCcmI-2 derivatives via different promoters, were constructed.These derivatives start at position 84 (Fig. 1, T84M) and con-tain the second transmembrane helix and the periplasmic ex-tension of CcmI with its four TPR-like motifs, thought tofacilitate protein-protein interactions (6, 14). Neither pCS1583nor pCS1579 complemented a CcmI-null mutant like MT-SRP1, but they both conferred Nadi�/Ps� phenotypes on en-riched media to MT-SRP1.r1 (CcmI� CcmFup CcmHRc
up),like the overproduction of CcmG (45).
The cyt c profiles of appropriate strains carrying pCS1579grown on enriched medium under Res conditions indicate thatthe amounts of membrane-bound cyts cp, c1, cy, and co as wellas soluble cyt c2 were similar to those carrying pCS1540 (over-producing CcmG) (Fig. 4D and E, lanes 7 and 9). Immunoblotanalyses show that the levels of CcmHRc were not distinguish-able between the strains carrying pCS1579 and pCS1540 (Fig.4B, lanes 7 and 9), while, as expected, those of CcmG weredrastically increased in strains containing pCS1540 (Fig. 4C,lanes 7 and 9) (45). However, the FLAG epitope-taggedCcmI-2 segment of CcmI could not be detected in crude cellextracts of appropriate strains harboring pCS1579 despite theirPs� phenotypes (Fig. 4A, lane 7), possibly due to its rapidproteolytic degradation. This difficulty was circumvented byusing intracytoplasmic membrane vesicles from an appropriatestrain carrying pCS1579 grown under Ps conditions on en-riched medium and affinity-purified FLAG-tagged CcmI-2, asdescribed in Materials and Methods. CcmI-2::FLAG migratedat an apparent molecular mass of 45 kDa (larger than itstheoretical molecular mass of 36.5 kDa) (Fig. 5, lanes 6 to 8).The data establish that the CcmI-2 segment and CcmG playvery similar roles in enhancing cyt c maturation in appropriateCcmI suppressors.
Simultaneous overproduction of CcmI-1 and CcmG also over-comes completely the holocyt c formation defects due to the ab-sence of CcmI. As the CcmI-1 segment and CcmF-CcmHRc cou-ple as well as the CcmI-2 segment and CcmG appearedfunctionally equivalent for cyt c maturation, we surmised that
simultaneous overproduction of CcmI-1 and CcmG might alsobypass completely the need for CcmI to produce adequateamounts of cyts c under all growth conditions. Indeed, plasmidpCS1589 (Fig. 2, row 15), producing a CcmI-1::FLAG derivativeas in pCB1 and an active CcmG as in pCS1540 (45), mediatedNadi�/Ps� phenotypes for appropriate null mutants of CcmI andCcmG (MT-SRP1 or MT-SRP1.r1 and MD11), indicating thatsimultaneous overproduction of the CcmI-1 segment and CcmG,similarly to that of the CcmF-CcmHRc couple and CcmG, sub-stitutes completely for the CcmI function during cyt c maturation.The data therefore suggest that the membrane-spanning segmentof CcmI or CcmF together with CcmHRc on the one hand and theperiplasmic segment of CcmI or CcmG on the other act cooper-atively during cyt c biogenesis.
Notably, additional plasmids, like pCB5 and pCS1590 (Fig.2, rows 13 and 14), carrying a ccmI1 allele fused in frameto a ccmG derivative without its corresponding signal pep-tide and membrane anchor sequence at its 5� end(ccmI1::FLAG-ccmG::His6) and expressing the fusion productfrom different promoters were not able to restore the Nadi�/Ps� phenotypes of a CcmG-null mutant (Table 1, MD11),although they readily complemented a CcmI-null mutant (MT-SRP1) for Nadi� on all media and for Ps� only on minimalmedium. These data point out that the fusion proteins pro-duced by pCB5 or pCS1590 were functional for only theirCcmI-1, but not their CcmG, moieties, suggesting that themembrane anchor of CcmG is important for its function, as hasbeen described similarly for its E. coli homologue (1).
Overexpression of cycA encoding cyt c2 can also substitutefor the function of CcmI-2 during cyt c maturation. Previously,we reported that overexpression of cycA encoding cyt c2 com-plemented a CycA-null mutant (FJM13) carrying an additionalpoint mutation in CcmI-2 for Ps� growth on minimal andenriched media (36). Consistent with this result, plasmidpHM14 harboring cycA also conferred the Ps� phenotype onboth media to a null mutant of CcmI-2 (SEL12) and a nullCcmI-2 and cyt c2 double mutant (SEL22). While pHM14 (Fig.
FIG. 5. Production of CcmI-2 in MT-SRP1.r1 grown under photo-synthetic conditions on enriched medium. Intracytoplasmic membranevesicles prepared from R. capsulatus strain MT-SRP1.r1/pCS1579[�(ccmI::kan) ccmFHRc
up ccmI2::FLAG expressed by the cycA pro-moter] grown on enriched medium under Ps conditions were solubi-lized using DDM, and FLAG-tagged CcmI-2 was purified as describedin Materials and Methods. Samples from different purification steps(CM, intracytoplasmic membrane vesicles; SM, solubilized membraneproteins; FT, flowthrough after three passages through the anti-FLAGagarose column; W, column wash; E1 to E4, elutions 1 to 4 from thecolumn) were subjected to SDS-PAGE (100 �g and 10 �g of proteinsin lanes 2 to 4 and 5 to 9, respectively), and immunoblot analyses werecarried out using polyclonal antibodies against the FLAG epitope, asdescribed in Materials and Methods. Lane 1 corresponds to 100 �g ofintracytoplasmic membrane vesicles from a wild-type strain and thusdoes not contain a FLAG epitope (CM wild type). Molecular massmarkers (in kDa) are shown on the left.
VOL. 189, 2007 FUNCTIONAL SEGMENTS OF R. CAPSULATUS CcmI 797
Dow
nloa
ded
from
http
s://j
ourn
als.
asm
.org
/jour
nal/j
b on
21
Febr
uary
202
2 by
118
.105
.101
.22.

2, row 16) could not complement the CcmI-null mutant MT-SRP1, it could fully complement MT-SRP1.r1 (CcmI� CcmFup
CcmHRcup) on both media. Thus, overproduction of apocyt c2
could substitute for the role of CcmI-2 under appropriate con-ditions, provided that either the CcmI-1 segment is producedor the CcmF-CcmHRc couple is overproduced during cyt cbiogenesis. This effect is seemingly specific to cycA, as cycYencoding cyt cy (pFJ63), petABC encoding the cyt bc1 complex(pMT0-404), and ccoNOQP encoding the cyt cbb3 oxidase(pOX15) (Table 1) could not mediate similar phenotypes toappropriate strains. Furthermore, the cyt c profiles of appro-priate MT-SRP1.r1 transconjugants grown on enriched me-dium under Res conditions indicated that the amounts of sol-uble cyts c2 and of membrane-bound cyts cp, c1, and co weresignificantly increased in the derivatives containing pHM14(Fig. 6, upper and lower panels).
DISCUSSION
In most gram-negative bacteria, including R. capsulatus,CcmI is an essential cyt c maturation component thought to actas a membrane-bound chaperone that directs apocyts c aftertheir translocation across the cytoplasmic membrane to theheme ligation sites (36). CcmI has a bipartite topology with anN-terminal membrane-spanning segment (CcmI-1) formed oftwo transmembrane helices encompassing a cytoplasmic loopcarrying a leucine zipper-like motif and a C-terminal periplas-mic segment (CcmI-2) that contains four TPR-like motifs (36,45). Both of these motifs are considered to facilitate protein-protein interactions between CcmI and its putative partners (6,
14, 35, 40). Furthermore, our earlier work indicated that twodistinct events are required to fully bypass the role of CcmIduring cyt c biogenesis and that CcmI might interact closelywith the CcmF-CcmHRc couple to form a heme ligation com-plex (17, 45). Here, we have dissected these distinct segmentsand their protein-protein interaction motifs to better definethe specific roles of the membrane-spanning and periplasmicsegments of CcmI during cyt c maturation in R. capsulatus.
Our work indicates that production of CcmI derivatives thatcontain (i) only the first transmembrane helix with or withoutany portion of its cytoplasmic loop or (ii) the CcmI-2 segmentpreceded by the second transmembrane helix of CcmI (Fig. 1)is insufficient to support any holocyt c production or to elimi-nate the associated heme precursor excretion phenotype.Moreover, deletion of the cytoplasmic loop of CcmI decreases,but does not abolish completely, cyt c maturation, suggestingthat this portion of CcmI with its leucine zipper-like motif isalso important for its function. Thus, an intact CcmI-1 segmentcomposed of its two transmembrane helices encompassing theleucine zipper-like domain is the minimum topological unitrequired for the formation of at least some c-type holocyts.Interestingly, overproduction of the CcmI-1 segment can fur-ther bypass the need for CcmI during cyt c maturation in agrowth medium-dependent manner, similar to that seen pre-viously with the overproduction of the CcmF-CcmHRc couple(17). Although CcmI-1 and the CcmF-CcmHRc couple behavesimilarly in the presence of cyt c2, strains lacking CcmI-2 andcyt c2, like SEL22, are complemented only with an intactCcmI-1 for adequate production of membrane-anchored cytsc1 and cy required for Ps growth. Together, these findingsprovide evidence that CcmI, CcmF, and CcmHRc act cooper-atively during cyt c maturation (Fig. 7) and further supportearlier proposals by us and others (42, 55) that they might forma core complex catalyzing heme ligation to apocytochromes. Ifsuch a multisubunit protein core complex indeed exists, thenthe membrane-integral segment of CcmI appears to be a crit-ical factor for its stability or activity (Fig. 7).
FIG. 6. Cytochrome c profiles of a CcmI mutant overproducing theCcmF-CcmHRc couple and various c-type cytochromes. Intracytoplasmicmembrane vesicles of appropriate R. capsulatus strains grown in enrichedmedium under Res conditions were subjected to SDS-PAGE using ap-proximately 75 �g of proteins per lane. Membrane-integral (upper panel)and soluble (lower panel) holocyts c were detected via their endogenousperoxidase activities by use of TMBZ and H2O2, as described in Materialsand Methods. Lanes 1 to 5 correspond to intracytoplasmic membranevesicles of the wild type (MT1131/pRK415), �ccmI ccmFHRc
up (MT-SRP1.r1/pRK415), cycA (MT-SRP1.r1/pHM14), cycY (MT-SRP1.r1/pFJ63), and petABC (MT-SRP1.r1/pMT0-404) (Table 1), respectively.Molecular mass markers (in kDa) are shown on the left.
FIG. 7. Roles of the membrane-integral and periplasmic segmentsof CcmI during cytochrome c biogenesis in R. capsulatus. The differentroles of the two CcmI segments during cyt c biogenesis are depictedschematically. We propose that the periplasmic CcmI-2 segment actscooperatively with CcmG to render apocyt c competent for hemeligation (“apoprotein substrate preparation”) and that this step is thencatalyzed by a protein complex (“core ligase”) functionally comprisingthe membrane-integral CcmI-1 segment and the CcmF-CcmHRc cou-ple. CcmE-heme is the holoform of the periplasmic heme chaperone.
798 SANDERS ET AL. J. BACTERIOL.
Dow
nloa
ded
from
http
s://j
ourn
als.
asm
.org
/jour
nal/j
b on
21
Febr
uary
202
2 by
118
.105
.101
.22.

Interestingly, different threshold levels of CcmI-1 and of theCcmF-CcmHRc couple are needed to restore to different ex-tents the Nadi�/Ps� phenotypes of CcmI-null mutants duringgrowth in different media. When these amounts are belowthe required threshold levels, then additional production ofCcmG, CcmI-2, or even the apocyt c2 acts cooperatively to fullybypass the role of CcmI during cyt c maturation. Thus, likeCcmI-1 and the CcmF-CcmHRc couple, CcmI-2 and CcmGalso play similar roles, as indicated by their functional cooper-ation upon their coproduction, albeit the activities currentlyattributed to the two sets of proteins during cyt c maturationare distinct. CcmG is a component of the thio-reductive path-way involved in keeping the thiol groups of the apocyt c hemebinding sites reduced, while the CcmF-CcmHRc pair is neededfor the apocyt c heme ligation process itself (38, 45). Thus, it istempting to conclude that the two segments of CcmI bringtogether two separate steps, namely, the preparation of cata-lytically competent apoprotein substrates and the subsequentheme attachment (Fig. 7), rationalizing why R. capsulatus cellsmight need two additive mutations to fully bypass the absenceof CcmI for efficient production of all c-type cyts under allgrowth conditions.
Cytoplasmic accumulation of heme precursors, as seen withS. meliloti or Rhizobium etli (9, 54), or its extracellular excre-tion, like in R. capsulatus (16, 23), is a notable phenotype oftenassociated with cyt c production defects in bacteria. The mo-lecular basis of this phenotype is unclear, and to what extent itreflects other physiological roles of cyt c maturation compo-nents, like siderophore synthesis or iron acquisition (8), isunknown. S. meliloti or R. capsulatus mutants that lack theentire CcmI, but not those that miss only the CcmI-2 segment,accumulate or excrete porphyrin derivatives. Expression of theN-terminal 96 amino acids of CcmI containing its first trans-membrane helix and the adjacent cytoplasmic loop, but lackingits second transmembrane helix, alleviates porphyrin accumu-lation in CcmI-null mutants of S. meliloti (9), but a similarderivative (CcmI-A96::FLAG) does not suppress porphyrinexcretion in similar R. capsulatus mutants. Physiological con-ditions that affect accumulation or excretion of heme precur-sors in different species need additional studies that are beyondthe scope of this work.
How at the molecular level the functions of CcmG andCcmI-2 are related to each other during cyt c biogenesis and howtheir activities are interconnected to those of CcmF, CcmHRc,and CcmI-1 are unknown, and these issues await biochemicalstudies. Overexpression of cycA, presumably leading to theoverproduction of apocyt c2 and acting like the CcmI-2 seg-ment, suggests that this portion of CcmI might facilitate direc-tion of the catalytically active apocyts c to the heme ligationsites involving CcmI-1, CcmF, and CcmHRc (Fig. 7). If this isthe case, whether the TPR-like motifs in the CcmI-2 segmentfacilitate these functions needs to be tested.
Currently, CcmG is considered to be a component of theperiplasmic apocyt c thioreduction pathway using its thiore-duction function to keep the thiol groups of the apocyt c hemebinding sites reduced (38). However, it has been shown re-cently that the reduced form of ResA, which is the Bacillussubtilis homologue of CcmG in Ccm system II, can recognizeapocyt c and release it in its reduced form to the systemII-specific heme ligation sites formed by ResB and ResC (10,
12). Aside from being a thioredoxin-like protein, if CcmG alsoplays a similar role in Ccm system I at least for some apocytsc (e.g., apocyt c2) prior to heme ligation, then its ability toenhance holocyt c production in the absence of the CcmI-2segment becomes less puzzling. In this respect, it is importantto point out that no mixed disulfide bond formation has beendetected so far between CcmG and CcmHRc in any bacteria todemonstrate electron transfer between these proteins. Fur-thermore, it is now known that the thioreduction function ofCcmG and ResA in Ccm systems I and II, respectively, isrequired for cyt c maturation only in the presence of an activethio-oxidative pathway formed by DsbA and DsbB (19; S.Turkarslan et al., unpublished data). Our ongoing works indi-cate that R. capsulatus CcmG has an additional function(s)distinct from its thioredoxin activity associated with cyt c pro-duction and that CcmG binds apocyt c in vitro (C. Sanders etal., unpublished data). Finally, it is noteworthy that the levelsof CcmG are much higher in CcmI-null mutants (Fig. 3C, D,and E, lanes 2, 5, and 6), as if CcmG is somehow regulated bythe absence of holocyts c.
In summary, the membrane-integral and the periplasmicsegments of CcmI clearly play distinct roles that are related tothose of the CcmF-CcmHRc couple and CcmG, respectively.Our overall findings led us to propose that CcmI coordinateswith its bipartite topology the thio-oxidoreduction and chap-eroning of apocyts c with the covalent and stereospecific hemeattachment steps during cyt c maturation (Fig. 7). Hopefully,future biochemical studies will further define the branchedinteraction networks between the Ccm system I componentsand better elucidate the molecular mechanisms underlying cytc biogenesis processes.
ACKNOWLEDGMENTS
This work was supported by grants from the DOE (91ER20052) andthe NIH (GM38237) to F.D.
We thank Neal Jaffe and Meenal Deshmukh for constructing theplasmids pNJ1 and pNJ2 and Majisha Doleyres and Janice Guntherfor their editorial comments. C.B. was a visiting student from theDepartement Biosciences, Institut Nationale des Sciences Appliqueesde Lyon, and UMR CNRS 5557 Ecologie Microbienne, UniversiteClaude Bernard Lyon I, 69622 Villeurbanne, France.
REFERENCES
1. Ahuja, U., and L. Thony-Meyer. 2006. The membrane anchors of the hemechaperone CcmE and the periplasmic thioredoxin CcmG are functionallyimportant. FEBS Lett. 580:216–222.
2. Allen, J. W., O. Daltrop, J. M. Stevens, and S. J. Ferguson. 2003. C-typecytochromes: diverse structures and biogenesis systems pose evolutionaryproblems. Philos. Trans. R. Soc. Lond. B 358:255–266.
3. Altschul, S. F., W. Gish, W. Miller, E. W. Myers, and D. J. Lipman. 1990.Basic local alignment search tool. J. Mol. Biol. 215:403–410.
4. Atta-Asafo-Adjei, E., and F. Daldal. 1991. Size of the amino acid side chainat position 158 of cytochrome b is critical for an active cytochrome bc1complex and for photosynthetic growth in Rhodobacter capsulatus. Proc.Natl. Acad. Sci. USA 88:492–496.
5. Benning, C., and C. R. Somerville. 1992. Isolation and genetic complemen-tation of a sulfolipid-deficient mutant of Rhodobacter sphaeroides. J. Bacte-riol. 174:2352–2360.
6. Blatch, G. L., and M. Lassle. 1999. The tetratricopeptide repeat: a structuralmotif mediating protein-protein interactions. Bioessays 21:932–939.
7. Bradford, M. M. 1976. A rapid and sensitive method for the quantitation ofmicrogram quantities of protein utilizing the principle of protein-dye bind-ing. Anal. Biochem. 72:248–254.
8. Cianciotto, N. P., P. Cornelis, and C. Baysse. 2005. Impact of the bacterialtype I cytochrome c maturation system on different biological processes.Mol. Microbiol. 56:1408–1415.
9. Cinege, G., A. Kereszt, S. Kertesz, G. Balogh, and I. Dusha. 2004. The rolesof different regions of the CycH protein in c-type cytochrome biogenesis inSinorhizobium meliloti. Mol. Genet. Genomics 271:171–179.
VOL. 189, 2007 FUNCTIONAL SEGMENTS OF R. CAPSULATUS CcmI 799
Dow
nloa
ded
from
http
s://j
ourn
als.
asm
.org
/jour
nal/j
b on
21
Febr
uary
202
2 by
118
.105
.101
.22.

10. Colbert, C. L., Q. Wu, P. J. Erbel, K. H. Gardner, and J. Deisenhofer. 2006.Mechanism of substrate specificity in Bacillus subtilis ResA, a thioredoxin-like protein involved in cytochrome c maturation. Proc. Natl. Acad. Sci. USA103:4410–4415.
11. Collet, J. F., and J. C. Bardwell. 2002. Oxidative protein folding in bacteria.Mol. Microbiol. 44:1–8.
12. Crow, A., N. E. Le Brun, and A. Oubrie. 2005. The role of ResA in type IIcytochrome c maturation. Biochem. Soc. Trans. 33:149–151.
13. Daldal, F., S. Cheng, J. Applebaum, E. Davidson, and R. C. Prince. 1986.Cytochrome c2 is not essential for photosynthetic growth of Rhodopseudo-monas capsulata. Proc. Natl. Acad. Sci. USA 83:2012–2016.
14. D’Andrea, L. D., and L. Regan. 2003. TPR proteins: the versatile helix.Trends Biochem. Sci. 28:655–662.
15. Davidson, E., and F. Daldal. 1987. Primary structure of the bc1 complex ofRhodopseudomonas capsulata. Nucleotide sequence of the pet operon en-coding the Rieske cytochrome b, and cytochrome c1 apoproteins. J. Mol.Biol. 195:13–24.
16. Deshmukh, M., G. Brasseur, and F. Daldal. 2000. Novel Rhodobacter cap-sulatus genes required for the biogenesis of various c-type cytochromes. Mol.Microbiol. 35:123–138.
17. Deshmukh, M., M. May, Y. Zhang, K. K. Gabbert, K. A. Karberg, R. G.Kranz, and F. Daldal. 2002. Overexpression of ccl1-2 can bypass the need forthe putative apocytochrome chaperone CycH during the biogenesis of c-typecytochromes. Mol. Microbiol. 46:1069–1080.
18. Ditta, G., T. Schmidhauser, E. Yakobson, P. Lu, X. W. Liang, D. R. Finlay,D. Guiney, and D. R. Helinski. 1985. Plasmids related to the broad hostrange vector, pRK290, useful for gene cloning and for monitoring geneexpression. Plasmid 13:149–153.
19. Erlendsson, L. S., R. M. Acheson, L. Hederstedt, and N. E. Le Brun. 2003.Bacillus subtilis ResA is a thiol-disulfide oxidoreductase involved in cyto-chrome c synthesis. J. Biol. Chem. 278:17852–17858.
20. Fabianek, R. A., H. Hennecke, and L. Thony-Meyer. 2000. Periplasmic pro-tein thiol:disulfide oxidoreductases of Escherichia coli. FEMS Microbiol.Rev. 24:303–316.
21. Feissner, R. E., C. L. Richard-Fogal, E. R. Frawley, and R. G. Kranz. 2006.ABC transporter-mediated release of a haem chaperone allows cytochromec biogenesis. Mol. Microbiol. 61:219–231.
22. Goldman, B. S., D. L. Beck, E. M. Monika, and R. G. Kranz. 1998. Trans-membrane heme delivery systems. Proc. Natl. Acad. Sci. USA 95:5003–5008.
23. Goldman, B. S., D. L. Beckman, A. Bali, E. M. Monika, K. K. Gabbert, andR. G. Kranz. 1997. Molecular and immunological analysis of an ABC trans-porter complex required for cytochrome c biogenesis. J. Mol. Biol. 268:724–738.
24. Goldman, B. S., and R. G. Kranz. 2001. ABC transporters associated withcytochrome c biogenesis. Res. Microbiol. 152:323–329.
25. Gray, K. A., M. Grooms, H. Myllykallio, C. Moomaw, C. Slaughter, and F.Daldal. 1994. Rhodobacter capsulatus contains a novel cb-type cytochrome coxidase without a CuA center. Biochemistry 33:3120–3127.
26. Jenney, F. E., Jr., and F. Daldal. 1993. A novel membrane-associated c-typecytochrome, cyt cy, can mediate the photosynthetic growth of Rhodobactercapsulatus and Rhodobacter sphaeroides. EMBO J. 12:1283–1292.
27. Jiang, X., and X. Wang. 2004. Cytochrome c-mediated apoptosis. Annu. Rev.Biochem. 73:87–106.
28. Kadokura, H., F. Katzen, and J. Beckwith. 2003. Protein disulfide bondformation in prokaryotes. Annu. Rev. Biochem. 72:111–135.
29. Katzen, F., M. Deshmukh, F. Daldal, and J. Beckwith. 2002. Evolutionarydomain fusion expanded the substrate specificity of the transmembraneelectron transporter DsbD. EMBO J. 21:3960–3969.
30. Keilin, D. 1966. The history of cell respiration and cytochrome. CambridgeUniversity Press, Cambridge, United Kingdom.
31. Koch, H. G., O. Hwang, and F. Daldal. 1998. Isolation and characterizationof Rhodobacter capsulatus mutants affected in cytochrome cbb3 oxidase ac-tivity. J. Bacteriol. 180:969–978.
32. Koch, H. G., H. Myllykallio, and F. Daldal. 1998. Using genetics to explorecytochrome function and structure in Rhodobacter. Methods Enzymol. 297:81–94.
33. Kranz, R., R. Lill, B. Goldman, G. Bonnard, and S. Merchant. 1998. Mo-lecular mechanisms of cytochrome c biogenesis: three distinct systems. Mol.Microbiol. 29:383–396.
34. Laemmli, U. K. 1970. Cleavage of structural proteins during the assembly ofthe head of bacteriophage T4. Nature 227:680–685.
35. Landschulz, W. H., P. F. Johnson, and S. L. McKnight. 1988. The leucinezipper: a hypothetical structure common to a new class of DNA bindingproteins. Science 240:1759–1764.
36. Lang, S. E., F. E. Jenney, Jr., and F. Daldal. 1996. Rhodobacter capsulatusCycH: a bipartite gene product with pleiotropic effects on the biogenesis ofstructurally different c-type cytochromes. J. Bacteriol. 178:5279–5290.
37. Metheringham, R., K. L. Tyson, H. Crooke, D. Missiakas, S. Raina, and J. A.Cole. 1996. Effects of mutations in genes for proteins involved in disulphidebond formation in the periplasm on the activities of anaerobically inducedelectron transfer chains in Escherichia coli K12. Mol. Gen. Genet. 253:95–102.
38. Monika, E. M., B. S. Goldman, D. L. Beckman, and R. G. Kranz. 1997. Athioreduction pathway tethered to the membrane for periplasmic cyto-chromes c biogenesis; in vitro and in vivo studies. J. Mol. Biol. 271:679–692.
39. Myllykallio, H., D. Zannoni, and F. Daldal. 1999. The membrane-attachedelectron carrier cytochrome cy from Rhodobacter sphaeroides is functional inrespiratory but not in photosynthetic electron transfer. Proc. Natl. Acad. Sci.USA 96:4348–4353.
40. O’Shea, E. K., J. D. Klemm, P. S. Kim, and T. Alber. 1991. X-ray structureof the GCN4 leucine zipper, a two-stranded, parallel coiled coil. Science254:539–544.
41. Pettigrew, G. W., and G. R. Moore. 1987. Cytochromes c. Biological aspects.Springer Verlag, New York, NY.
42. Ren, Q., U. Ahuja, and L. Thony-Meyer. 2002. A bacterial cytochrome cheme lyase. CcmF forms a complex with the heme chaperone CcmE andCcmH but not with apocytochrome c. J. Biol. Chem. 277:7657–7663.
43. Sambongi, Y., and S. J. Ferguson. 1996. Mutants of Escherichia coli lackingdisulphide oxidoreductases DsbA and DsbB cannot synthesise an exogenousmonohaem c-type cytochrome except in the presence of disulphide com-pounds. FEBS Lett. 398:265–268.
44. Sambrook, J., and D. W. Russell. 2001. Molecular cloning: a laboratorymanual, 3rd ed. Cold Spring Harbor Laboratory Press, Cold Spring Harbor,NY.
45. Sanders, C., M. Deshmukh, D. Astor, R. G. Kranz, and F. Daldal. 2005.Overproduction of CcmG and CcmFHRc fully suppresses the c-type cyto-chrome biogenesis defect of Rhodobacter capsulatus CcmI-null mutants. J.Bacteriol. 187:4245–4256.
46. Sanders, C., S. Turkarslan, and F. Daldal. 2005. Periplasmic oxidative fold-ing and cytochrome c maturation: a mechanistic view of stereo-selectiveheme attachment, p. 421–424. In A. van der Est and D. Bruce (ed.), Photo-synthesis: fundamental aspects and global perspectives. Allen Press, Law-rence, KS.
47. Schaegger, H., and G. von Jagow. 1987. Tricine-sodium dodecyl sulfate-polyacrylamide gel electrophoresis for the separation of proteins in the rangefrom 1–100 kDa. Anal. Biochem. 166:368–379.
48. Scolnik, P. A., M. A. Walker, and B. L. Marrs. 1980. Biosynthesis of caro-tenoids derived from neurosporene in Rhodopseudomonas capsulata. J. Biol.Chem. 255:2427–2432.
49. Sistrom, W. R. 1960. A requirement for sodium in the growth of Rhodopseu-domonas sphaeroides. J. Gen. Microbiol. 22:778–785.
50. Thomas, P. E., D. Ryan, and W. Levin. 1976. An improved staining proce-dure for the detection of the peroxidase activity of cytochrome P-450 onsodium dodecyl sulfate polyacrylamide gels. Anal. Biochem. 75:168–176.
51. Thony-Meyer, L. 2002. Cytochrome c maturation: a complex pathway for asimple task? Biochem. Soc. Trans. 30:633–638.
52. Thony-Meyer, L. 2003. A heme chaperone for cytochrome c biosynthesis.Biochemistry 42:13099–13105.
53. Wiechelman, K. J., R. D. Braun, and J. D. Fitzpatrick. 1988. Investigation ofthe bicinchoninic acid protein assay: identification of the groups responsiblefor color formation. Anal. Biochem. 175:231–237.
54. Yeoman, K. H., M. J. Delgado, M. Wexler, J. A. Downie, and A. W. Johnston.1997. High affinity iron acquisition in Rhizobium leguminosarum requires thecycHJKL operon and the feuPQ gene products, which belong to the family oftwo-component transcriptional regulators. Microbiology 143:127–134.
55. Zannoni, D., and F. Daldal. 1993. The role of c-type cytochromes in cata-lyzing oxidative and photosynthetic electron transport in the dual functionalplasmamembrane of facultative phototrophs. Arch. Microbiol. 160:413–423.
800 SANDERS ET AL. J. BACTERIOL.
Dow
nloa
ded
from
http
s://j
ourn
als.
asm
.org
/jour
nal/j
b on
21
Febr
uary
202
2 by
118
.105
.101
.22.