microscopic structure of the retina and vasculature in the...
TRANSCRIPT

Microscopic structure of the retina and vasculature in the human eye
D-Y. Yu1,2, P. K. Yu
1,2, C. Balaratnasingam
1,2, S. J. Cringle
1,2 and E-N. Su
1,2
1 Centre for Ophthalmology and Visual Science, The University of Western Australia, Perth, Australia 2 The ARC Centre of Excellence in Vision Science, The University of Western Australia, Perth, Australia
Retinal diseases with a vascular component account for the majority of blindness in the developed world. The vulnerability
of the retina stems in part from the limitations placed on the retinal vascular system to not obstruct the light path to the
photoreceptors. Some layers of the retina are completely avascular, and in our central region of vision, the fovea, there are
no retinal vessels at all. The retinal vasculature has a highly efficient regulatory system to match blood flow to local
metabolic demands. How this is achieved, and how it is impaired in retinal vascular diseases, is the subject of much
interest in ophthalmic research. In the present study we are employing micro-perfusion fixation and labelling techniques
in human donor eyes. Flat mounted specimens are then studied with multi-wavelength confocal microscopy techniques to
explore the three dimensional relationship between the vascular and neural elements in the human retina. The quality of
label uptake and the efficient fixation of the tissue also allow highly detailed studies of the endothelial cell cyto-
architecture which is providing new insights into the complexity of the control mechanisms in the retinal vasculature.
Such studies will provide the baseline data for future studies using eyes from donors with a known history of retinal
disease.
Keywords Retina; microvasculature; human, macula; optical imaging; cell labelling methods; retinal development;
confocal microscopy
1. Introduction
1.1 General knowledge of retinal structure and function
The human and primate retina is effectively part of the brain that is displaced into the eye during development. The
retina is composed of a thin (~250 µm) sheet in which all retinal neurones are densely packed. In man, the light-
sensitive cells, the photoreceptors, include ~4.6 million cones and 92 million rods [4]. In addition to photoreceptors, the
retina contains four basic classes of neurons organized into three cellular (nuclear) layers, which are separated by two
synaptic (plexiform) layers. There are also two types of glial cells, Muller cells, and astrocytes. Figure 1A shows a
micrograph of the retina and choroid from the parafoveal region of a normal monkey. The layered structure in the
parafoveal region is similar to that in the more peripheral retina, but the thickness of some layers is very different. For
example, the retinal ganglion cell layer (RGC) and outer plexiform layer (OPL) are much thicker. A small region of
retina, the fovea, is morphologically specialized for high-acuity vision. In the fovea the inner layers of the retina
proximal to the photoreceptors are displaced sideways from the fovea so that light can impinge directly on the
photoreceptors. The central rod-free foveal area is located on the visual axis and contains highly specialised cones, the
thinnest and longest in the retina, numbering only 0.1% of the total cone population.
1.2 Significant heterogeneity of metabolic requirements of different retinal layers
The metabolic and functional properties of the retina are highly compartmentalised in terms of their topographic and
geometric distribution. Oxygen is the only molecule serving as the primary biological oxidant [11] and is essential for
the survival of cells. The retina has perhaps the highest oxygen consumption rate of any tissue in the body [1]. Since
oxygen cannot be “stored” in tissue, a constant and adequate supply must be guaranteed in order to preserve function.
Oxygen supply to the retina is arguably more vulnerable to vascular deficiencies than in any other organ. Numerous
studies have demonstrated the marked heterogeneity of oxygen metabolism across the retina, even in different
components of the same cell. The parafoveal region, like most retinal regions except the foveola, is supplied by two
circulations, the retinal and choroidal circulations. The choroid is highly vascularised and lies immediately behind the
retina. The choroid is the dominant oxygen supply for the outer retina. The choroid has a very high blood flow and no
clearly demonstrated regulatory capacity [9], despite the presence of autonomic innervation [2, 8]. However, the
regulatory properties of the choroidal and retinal circulations differ markedly. The retinal circulation is relatively sparse,
presumably to minimize disruption to the light path, and exhibits a well-developed regulatory capacity [10], which is
thought to be purely “local” as there is no apparent autonomic innervation [7]. It seems likely that these opposing
requirements of minimal interference with the light path and provision of sustenance to the highly metabolizing retinal
tissue have led to the development of a system that is delicately, if not precariously, balanced.
The highly layered structure of the retina provides an opportunity for investigating the properties of different sub
cellular components not achievable in the brain. Using oxygen sensitive microelectrode technique, the intraretinal
Microscopy: Science, Technology, Applications and Education A. Méndez-Vilas and J. Díaz (Eds.)
©FORMATEX 2010 867
______________________________________________
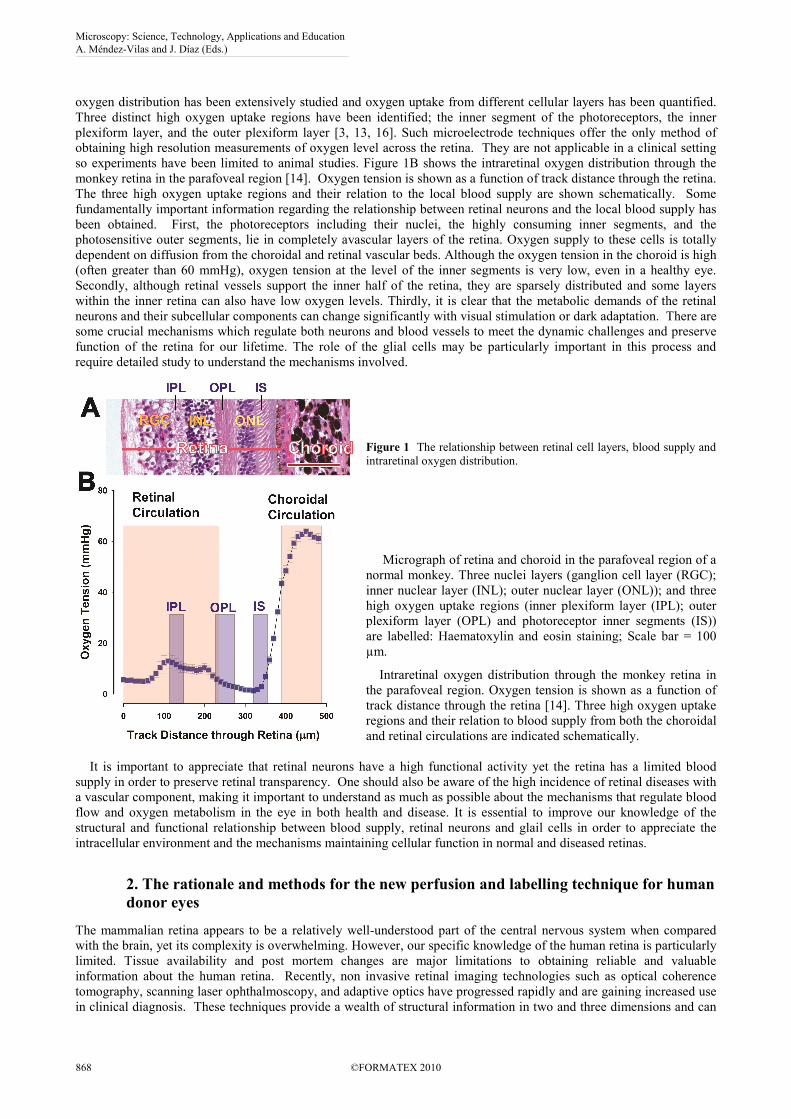
oxygen distribution has been extensively studied and oxygen uptake from different cellular layers has been quantified.
Three distinct high oxygen uptake regions have been identified; the inner segment of the photoreceptors, the inner
plexiform layer, and the outer plexiform layer [3, 13, 16]. Such microelectrode techniques offer the only method of
obtaining high resolution measurements of oxygen level across the retina. They are not applicable in a clinical setting
so experiments have been limited to animal studies. Figure 1B shows the intraretinal oxygen distribution through the
monkey retina in the parafoveal region [14]. Oxygen tension is shown as a function of track distance through the retina.
The three high oxygen uptake regions and their relation to the local blood supply are shown schematically. Some
fundamentally important information regarding the relationship between retinal neurons and the local blood supply has
been obtained. First, the photoreceptors including their nuclei, the highly consuming inner segments, and the
photosensitive outer segments, lie in completely avascular layers of the retina. Oxygen supply to these cells is totally
dependent on diffusion from the choroidal and retinal vascular beds. Although the oxygen tension in the choroid is high
(often greater than 60 mmHg), oxygen tension at the level of the inner segments is very low, even in a healthy eye.
Secondly, although retinal vessels support the inner half of the retina, they are sparsely distributed and some layers
within the inner retina can also have low oxygen levels. Thirdly, it is clear that the metabolic demands of the retinal
neurons and their subcellular components can change significantly with visual stimulation or dark adaptation. There are
some crucial mechanisms which regulate both neurons and blood vessels to meet the dynamic challenges and preserve
function of the retina for our lifetime. The role of the glial cells may be particularly important in this process and
require detailed study to understand the mechanisms involved.
Figure 1 The relationship between retinal cell layers, blood supply and
intraretinal oxygen distribution.
Micrograph of retina and choroid in the parafoveal region of a
normal monkey. Three nuclei layers (ganglion cell layer (RGC);
inner nuclear layer (INL); outer nuclear layer (ONL)); and three
high oxygen uptake regions (inner plexiform layer (IPL); outer
plexiform layer (OPL) and photoreceptor inner segments (IS))
are labelled: Haematoxylin and eosin staining; Scale bar = 100
µm.
Intraretinal oxygen distribution through the monkey retina in
the parafoveal region. Oxygen tension is shown as a function of
track distance through the retina [14]. Three high oxygen uptake
regions and their relation to blood supply from both the choroidal
and retinal circulations are indicated schematically.
It is important to appreciate that retinal neurons have a high functional activity yet the retina has a limited blood
supply in order to preserve retinal transparency. One should also be aware of the high incidence of retinal diseases with
a vascular component, making it important to understand as much as possible about the mechanisms that regulate blood
flow and oxygen metabolism in the eye in both health and disease. It is essential to improve our knowledge of the
structural and functional relationship between blood supply, retinal neurons and glail cells in order to appreciate the
intracellular environment and the mechanisms maintaining cellular function in normal and diseased retinas.
2. The rationale and methods for the new perfusion and labelling technique for human
donor eyes
The mammalian retina appears to be a relatively well-understood part of the central nervous system when compared
with the brain, yet its complexity is overwhelming. However, our specific knowledge of the human retina is particularly
limited. Tissue availability and post mortem changes are major limitations to obtaining reliable and valuable
information about the human retina. Recently, non invasive retinal imaging technologies such as optical coherence
tomography, scanning laser ophthalmoscopy, and adaptive optics have progressed rapidly and are gaining increased use
in clinical diagnosis. These techniques provide a wealth of structural information in two and three dimensions and can
Microscopy: Science, Technology, Applications and Education A. Méndez-Vilas and J. Díaz (Eds.)
868 ©FORMATEX 2010
______________________________________________

significantly improve the level of clinical diagnosis. Many interesting discoveries have been reported from these new
imaging technologies. Our knowledge from conventional microscopic histology, such as retinal sections is insufficient
to interpret the findings from the new generation of imaging technologies. Furthermore, each of the current imaging
methods available has optical and technical limitations. In order to make a correct judgement based on these images in
clinical practice, we need more detailed knowledge at both the gross and cellular levels regarding the structure of the
human retina. We therefore developed a new technique to study post-mortem human eyes to investigate the spatial 2D
and 3D relationship between microvasculature, glial cells and neurons in the retina at both the geographic distribution
and cyto-architecture level. A major question that was considered was how to achieve the most efficient fixation and
staining to minimise post mortem changes. We have used isolated perfused retinal arteriole preparations for many years
to study vascular physiology and pharmacology [15]. We learned from handling post mortem eyes that the vascular
tissue was remarkably robust, both structurally and functionally. We then reasoned that perfusion of the ocular
vasculature offered a means of rapid fixation and staining of the ocular tissues. We chose to perfuse through both the
central retinal artery and a branch of the posterior ciliary artery to flush out the blood and then fix and stain both the
retinal and choroidal circulations. Cannulation of such fine vessels from post mortem tissue does present a technical
challenge, but the majority of post-mortem donor eyes can be cannulated and perfused. Fixative perfusion through the
intact vasculature including all capillaries creates much more effective tissue fixation than immersion fixation.
Perfusion pressure was continuously monitored as a guide to the vascular resistance. This allows the early detection of
blockages or leaks. We were careful not to allow perfusate pressure to exceed the normal physiological range since it is
likely that the vasculature may be weakened in post mortem tissue and we wished to avoid rupture of the vascular wall.
The perfusion technique for fixing and the labelling the vascular endothelium of the retinal and choroidal
microvasculature has been described in detail elsewhere [18]. Briefly, two double suture ties (8-O) were used for
securing each cannula. Syringe pumps (Model 22, Harvard Apparatus, South Natick. MA) were used to deliver an
adjustable flow of perfusate (typically 50 µL/min) and the perfusion pressure was continuously monitored through
conventional transducers (Cobe, Arvada, Colorado), each connected to a bridge amplifier (5B38-02 Analog Devices;
Norwood, MA) and recorded on a chart recorder (LR8100, Yokogawa; Tokyo, Japan). The standard perfusate solution
for washout was oxygenated Ringers solution with 1% Bovine Serum Albumin. At least 15 minutes was allowed to
flush out any residual blood. Then the following solutions were perfused through in order; 4% paraformaldehyde in
0.1M phosphate buffer solution (20 minutes), 0.1% Triton-X-100 in 0.1M phosphate buffer solution (5 minutes), 0.1M
phosphate buffer solution (20 minutes), dye (30U Alexa Fluor 546 or 635 Phalloidins and Hoechst 1.2 µg/ml in 0.1M
phosphate buffer (120 minutes), (Invitrogen A22283 or A34054 & bisBenzimide H 33258 Cat No B2261 from Sigma-
Aldrich), and 0.1M phosphate buffer (30 minutes). Some eyes were perfused with TRITC-Lectins (50 µg/750 µl) in
0.1M phosphate buffer along with the nucleic acid dye bisBenzimide H 33258 (1.2 µg/ml) instead of the Phalloidins.
After dye injection a further period of 0.1M phosphate buffer perfusion (30 minutes) was used to wash away excess
dye. The eyes were then immersed in 4% paraformaldehyde in 0.1M phosphate buffer overnight before the retina and
choroid was flat-mounted for imaging. Orientation and macular location was carefully noted and documented
photographically. Alternatively, we could also use different immunohistochemical labelling such as VE-Cadherin
instead of phalloidin labelling [17].
The posterior globe was dissected at the equator to allow viewing of the posterior retina. The macular region and
fovea was identified by the anatomical orientation of the globe, yellow pigmentation around the fovea, and the
increased choroidal pigmentation in this region. The retina was carefully dissected out without inclusion of the optic
disk region. A few cuts were also made to the retina to enable it to lie flat for mounting.
Retinas that have been perfusion-stained or immersion fixed post-mortem may be immunolabelled as a wholemount
according to the protocol by Xiao and Hendrickson [12] to study the relationship between various retinal cell
components. In brief, the whole retina was washed in 0.1M phosphate buffer on a shaker for 1 to 2 days and floated in
1% sodium borohydride (tetrahydroborate) in phosphate buffer overnight, cryoprotected in 30% sucrose. It was then
stored at -80°C until use. After thawing to room temperature, the retina was floated in a 0.1M phosphate buffer with
0.1% azide and 0.1% Triton-X-100 at 4°C in the following sequence: 10% goat serum (6 to 24 hours), primary
antibodies (3-4 days on a shaker), phosphate buffer wash (1 to 2 days), secondary antibodies and nuclei counterstain
(overnight) and a phosphate buffer wash (2 hours) before being mounted for imaging/viewing
We used a Nikon E800 equipped with Plan Apo lenses ranging from x4, x10, x20, x40 (oil), x60 (oil) and x100 (oil).
In addition to the usual broadband UV, blue and green filter blocks, a special triple filter block was used for
simultaneous imaging of DAPI (or Hoechst)/Texas Red/FITC. ACT-1 software allowed imaging at 9 levels of
resolution, control of exposure time and labelling of image files. Images were captured using a DXM1200 camera
(Nikon, Tokyo, Japan).
The Nikon C1 equipped with three lasers (405, 488 and 532 nm) was used in conjunction with the Nikon E800
microscope. Imaging was done using the EZ-C1 software (v.3.20). Confocal imaging was performed simultaneously for
the different wavelengths with emission signals separated into different channels. Imaging began at low magnification.
Specific regions were zoomed in using high power objective lens (x40, x60 plan apochromatic oil lenses) for detailed
imaging. Z-series were taken through to a depth of 100 µm to obtain 3D information on retinal microvascular
Microscopy: Science, Technology, Applications and Education A. Méndez-Vilas and J. Díaz (Eds.)
©FORMATEX 2010 869
______________________________________________

architecture and to study labelled structures in the vascular endothelium and the retina. The three wavelengths enabled
analysis of the triple labelling of the vascular endothelium and retina.
After imaging, specific regions (e.g. macula, perifovea, surrounding retina) were embedded in OCT for
cryosectioning. Frozen semi-serial sections were mounted on Erie superfrost PLUS gold slides (Biolab, ERIFT-
4981GLPLUS-006E) and allowed to air-dry for an hour prior to staining. Sections from the same region were labelled
with antibodies for identification of different types of retinal neural and glial cells. Intensity of staining with particular
markers can increase or decrease when tissues are exposed to certain physiological or pathologic conditions. Caution is
therefore needed to avoid coming to false conclusions on the basis of single markers.
Procedures for studying the choroidal vasculature required modifying the techniques described above. After
detaching the retina from the posterior globe, as far as possible, the RPE layer was carefully removed using a ferret hair
brush and washing the RPE away from choroid using buffer. A cut was made along the inferior meridian of the intact
choroid/sclera eye cup for purpose of orientation before detachment of the choroid from the sclera. The choroid was
peeled from the underlying sclera with the help of the brush. The area around the macula may require the help of a
metal "scoop" or cutting with fine scissors. The dissected choroid was then flat mounted in glycerol, retina side up, and
multiple radial cuts made to help flatten the specimen before coverslipping and imaging.
3. The major findings from new perfusion and labelling technique for human donor
eye study
3.1 Retinal microvascular distribution and topography
Using confocal microscopy, optic sections were obtained from flat mounted retinas after perfusing and labelling retinal
microvasculature in the retinal tissue. These optic sections can be stacked from selected depths and pseudo-coloured to
identify the depth from 2D images, and also can be constructed as 3D images. Retinal microvasculature is normally
located at half thickness of the retina, only about 150 µm which is a significant advantage for confocal microscopic
imaging. Low magnification images allow the study of vascular distribution and topography.
Figure 2 Confocal images of retinal microvasculature at the macular region obtained from a donor eye after perfusion staining
labelled for filamentous actin. The images show the 2D macular region (A) and 3D images of the immediate region of the fovea as
marked area (B1, B2 and B3). Depth information has been included for this stack of images. The top half is pseudo-coloured red
and the bottom half pseudo-coloured green. The alternating arteriole (a) supply and venous (v) drainage pattern of the macular is
clearly seen. A foveal capillary is clearly visible going around the foveola (*) in the deeper layer of this macula. The 3D images of
the immediate region of the fovea as marked area are showed in B1, B2 and B3 from different angles allowing the detailed spatial
relationship between the microvessels to be seen when played together as a movie. (A) Scale bar: 500 µm. (B1, B2, B3) Each major
division is 50 µm.
Microscopy: Science, Technology, Applications and Education A. Méndez-Vilas and J. Díaz (Eds.)
870 ©FORMATEX 2010
______________________________________________
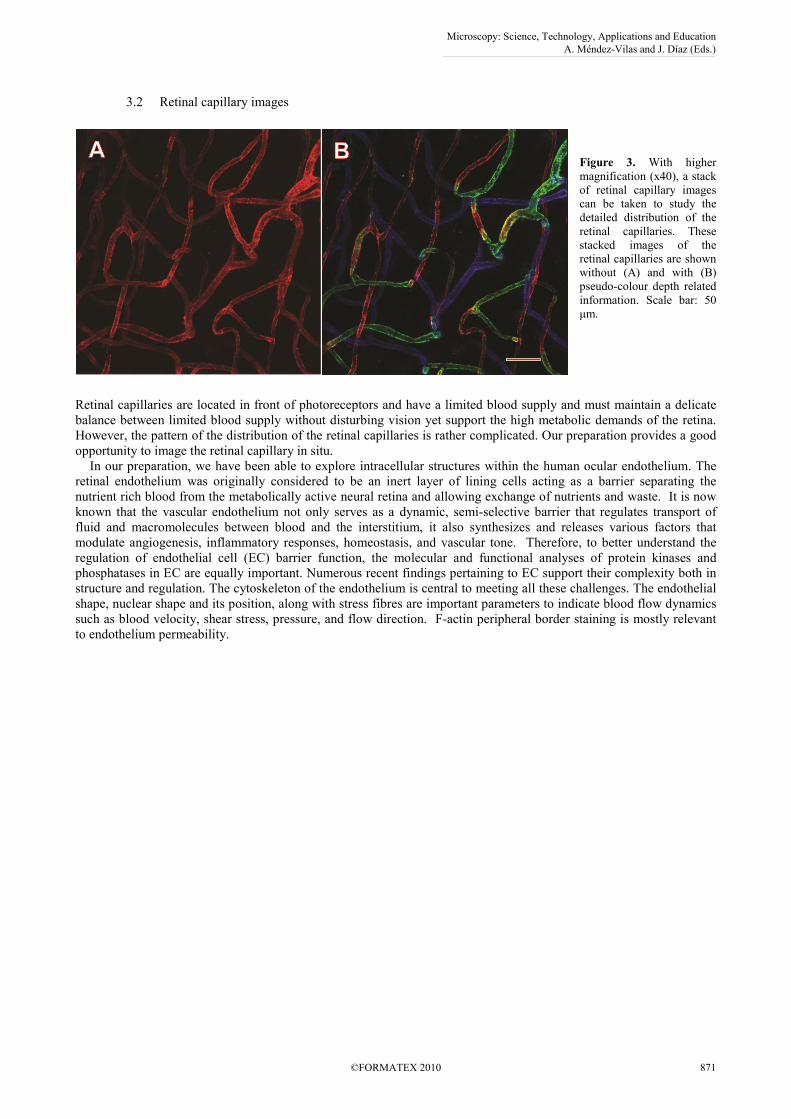
3.2 Retinal capillary images
Figure 3. With higher
magnification (x40), a stack
of retinal capillary images
can be taken to study the
detailed distribution of the
retinal capillaries. These
stacked images of the
retinal capillaries are shown
without (A) and with (B)
pseudo-colour depth related
information. Scale bar: 50
µm.
Retinal capillaries are located in front of photoreceptors and have a limited blood supply and must maintain a delicate
balance between limited blood supply without disturbing vision yet support the high metabolic demands of the retina.
However, the pattern of the distribution of the retinal capillaries is rather complicated. Our preparation provides a good
opportunity to image the retinal capillary in situ.
In our preparation, we have been able to explore intracellular structures within the human ocular endothelium. The
retinal endothelium was originally considered to be an inert layer of lining cells acting as a barrier separating the
nutrient rich blood from the metabolically active neural retina and allowing exchange of nutrients and waste. It is now
known that the vascular endothelium not only serves as a dynamic, semi-selective barrier that regulates transport of
fluid and macromolecules between blood and the interstitium, it also synthesizes and releases various factors that
modulate angiogenesis, inflammatory responses, homeostasis, and vascular tone. Therefore, to better understand the
regulation of endothelial cell (EC) barrier function, the molecular and functional analyses of protein kinases and
phosphatases in EC are equally important. Numerous recent findings pertaining to EC support their complexity both in
structure and regulation. The cytoskeleton of the endothelium is central to meeting all these challenges. The endothelial
shape, nuclear shape and its position, along with stress fibres are important parameters to indicate blood flow dynamics
such as blood velocity, shear stress, pressure, and flow direction. F-actin peripheral border staining is mostly relevant
to endothelium permeability.
Microscopy: Science, Technology, Applications and Education A. Méndez-Vilas and J. Díaz (Eds.)
©FORMATEX 2010 871
______________________________________________

Figure 4 shows confocal images of a major retinal artery. (A) Intracellular structure of endothelial cells (ECs) and smooth muscle
cells (SMCs) was labelled. The white dotted line outlines the area from which the image B was taken [17]. (B): Top: cell border
located F-actin (peripheral bound staining, PBS) outlined the shape of endothelium (yellow arrow heads) and a more structured
filamentous structure (stress fibres, labelled by small white arrowheads) inside the endothelium and nuclei (N) was labelled with
Hoechst. Strong staining of smooth muscle cells made relatively weak appearance of PBS and SF. A schematic drawing was
indicates the PBS and SF location. Scale bar: A: 50 µm, B: 10 µm.
3.3 Retinal vessel, glia and neuron images
Our preparation allows us to detail the spatial distribution of retinal microvasculature and its relationship to neurons and
glial cells at the topographic and cellular level in human donor eyes.
Microscopy: Science, Technology, Applications and Education A. Méndez-Vilas and J. Díaz (Eds.)
872 ©FORMATEX 2010
______________________________________________
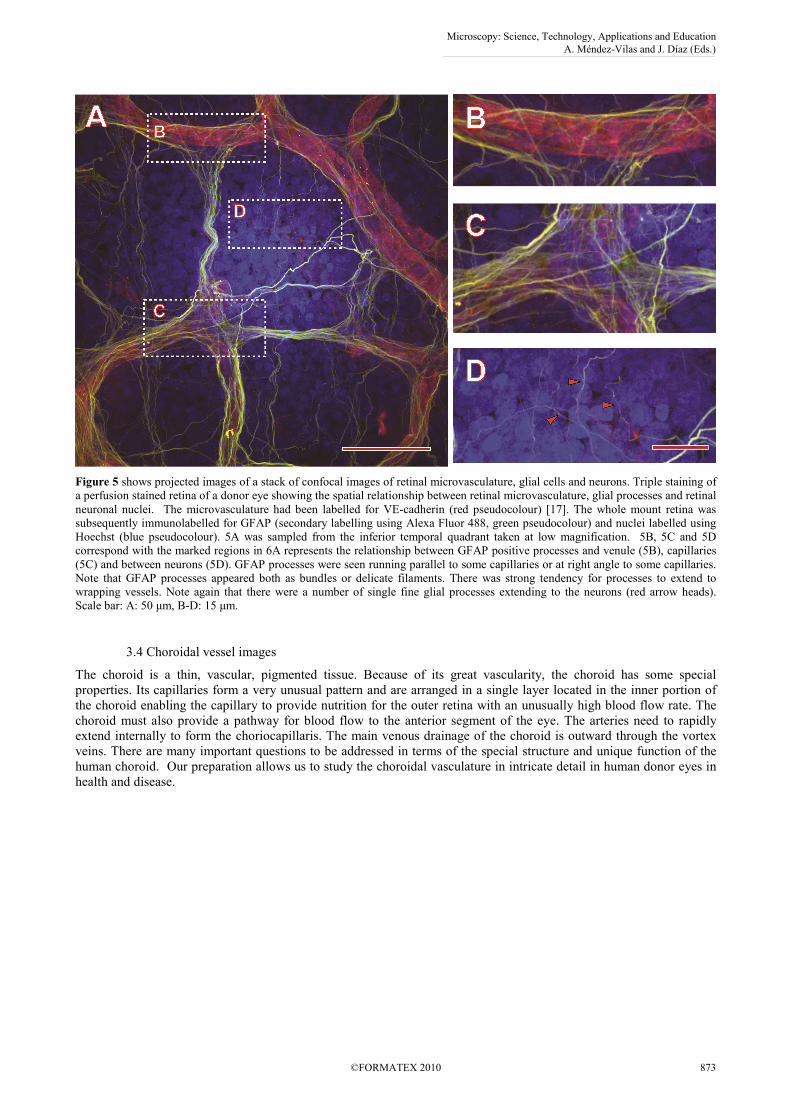
Figure 5 shows projected images of a stack of confocal images of retinal microvasculature, glial cells and neurons. Triple staining of
a perfusion stained retina of a donor eye showing the spatial relationship between retinal microvasculature, glial processes and retinal
neuronal nuclei. The microvasculature had been labelled for VE-cadherin (red pseudocolour) [17]. The whole mount retina was
subsequently immunolabelled for GFAP (secondary labelling using Alexa Fluor 488, green pseudocolour) and nuclei labelled using
Hoechst (blue pseudocolour). 5A was sampled from the inferior temporal quadrant taken at low magnification. 5B, 5C and 5D
correspond with the marked regions in 6A represents the relationship between GFAP positive processes and venule (5B), capillaries
(5C) and between neurons (5D). GFAP processes were seen running parallel to some capillaries or at right angle to some capillaries.
Note that GFAP processes appeared both as bundles or delicate filaments. There was strong tendency for processes to extend to
wrapping vessels. Note again that there were a number of single fine glial processes extending to the neurons (red arrow heads).
Scale bar: A: 50 µm, B-D: 15 µm.
3.4 Choroidal vessel images
The choroid is a thin, vascular, pigmented tissue. Because of its great vascularity, the choroid has some special
properties. Its capillaries form a very unusual pattern and are arranged in a single layer located in the inner portion of
the choroid enabling the capillary to provide nutrition for the outer retina with an unusually high blood flow rate. The
choroid must also provide a pathway for blood flow to the anterior segment of the eye. The arteries need to rapidly
extend internally to form the choriocapillaris. The main venous drainage of the choroid is outward through the vortex
veins. There are many important questions to be addressed in terms of the special structure and unique function of the
human choroid. Our preparation allows us to study the choroidal vasculature in intricate detail in human donor eyes in
health and disease.
Microscopy: Science, Technology, Applications and Education A. Méndez-Vilas and J. Díaz (Eds.)
©FORMATEX 2010 873
______________________________________________
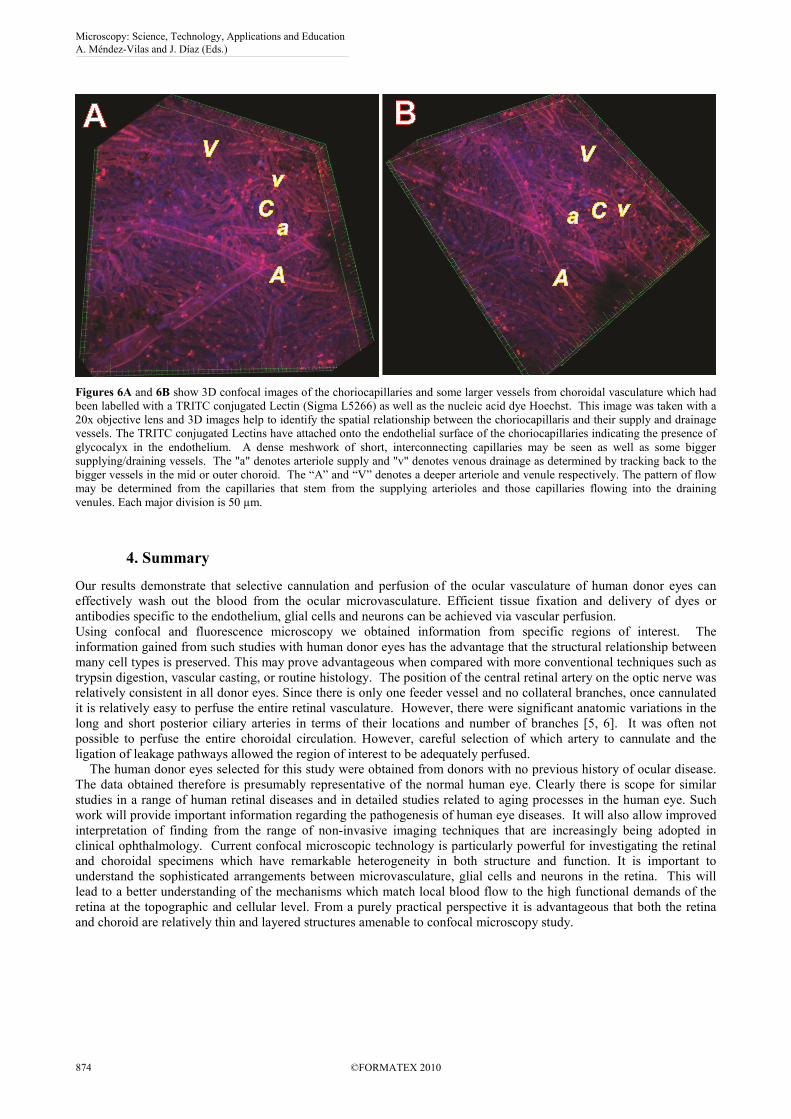
Figures 6A and 6B show 3D confocal images of the choriocapillaries and some larger vessels from choroidal vasculature which had
been labelled with a TRITC conjugated Lectin (Sigma L5266) as well as the nucleic acid dye Hoechst. This image was taken with a
20x objective lens and 3D images help to identify the spatial relationship between the choriocapillaris and their supply and drainage
vessels. The TRITC conjugated Lectins have attached onto the endothelial surface of the choriocapillaries indicating the presence of
glycocalyx in the endothelium. A dense meshwork of short, interconnecting capillaries may be seen as well as some bigger
supplying/draining vessels. The "a" denotes arteriole supply and "v" denotes venous drainage as determined by tracking back to the
bigger vessels in the mid or outer choroid. The “A” and “V” denotes a deeper arteriole and venule respectively. The pattern of flow
may be determined from the capillaries that stem from the supplying arterioles and those capillaries flowing into the draining
venules. Each major division is 50 µm.
4. Summary
Our results demonstrate that selective cannulation and perfusion of the ocular vasculature of human donor eyes can
effectively wash out the blood from the ocular microvasculature. Efficient tissue fixation and delivery of dyes or
antibodies specific to the endothelium, glial cells and neurons can be achieved via vascular perfusion.
Using confocal and fluorescence microscopy we obtained information from specific regions of interest. The
information gained from such studies with human donor eyes has the advantage that the structural relationship between
many cell types is preserved. This may prove advantageous when compared with more conventional techniques such as
trypsin digestion, vascular casting, or routine histology. The position of the central retinal artery on the optic nerve was
relatively consistent in all donor eyes. Since there is only one feeder vessel and no collateral branches, once cannulated
it is relatively easy to perfuse the entire retinal vasculature. However, there were significant anatomic variations in the
long and short posterior ciliary arteries in terms of their locations and number of branches [5, 6]. It was often not
possible to perfuse the entire choroidal circulation. However, careful selection of which artery to cannulate and the
ligation of leakage pathways allowed the region of interest to be adequately perfused.
The human donor eyes selected for this study were obtained from donors with no previous history of ocular disease.
The data obtained therefore is presumably representative of the normal human eye. Clearly there is scope for similar
studies in a range of human retinal diseases and in detailed studies related to aging processes in the human eye. Such
work will provide important information regarding the pathogenesis of human eye diseases. It will also allow improved
interpretation of finding from the range of non-invasive imaging techniques that are increasingly being adopted in
clinical ophthalmology. Current confocal microscopic technology is particularly powerful for investigating the retinal
and choroidal specimens which have remarkable heterogeneity in both structure and function. It is important to
understand the sophisticated arrangements between microvasculature, glial cells and neurons in the retina. This will
lead to a better understanding of the mechanisms which match local blood flow to the high functional demands of the
retina at the topographic and cellular level. From a purely practical perspective it is advantageous that both the retina
and choroid are relatively thin and layered structures amenable to confocal microscopy study.
Microscopy: Science, Technology, Applications and Education A. Méndez-Vilas and J. Díaz (Eds.)
874 ©FORMATEX 2010
______________________________________________

References
[1] Anderson B. Ocular effects of changes in oxygen and carbon dioxide tension. Transactions of the American Ophthalmological
Society. 1968;66:423-474.
[2] Beckers HJM, Klooster J, Vrensen GFJM and Lamers WPMA. Facial parasympathetic innervation of the rat choroid, lacrimal
glands and ciliary ganglion. Ophthalmic Research. 1993;25:319-330.
[3] Cringle SJ and Yu D-Y. A multi-layer model of retinal oxygen supply and consumption helps explain the muted rise in inner
retinal PO2 during systemic hyperoxia. Comparative Biochemistry and Physiology. 2002;132:61-66.
[4] Curcio CA, Sloan KA, Kalina RE and Hendrickson AE. Human photoreceptor topography. The Journal of Comparative
Neurology. 1990;292:497-523.
[5] Erdogmus S and Govsa F. Anatomic characteristics of the ophthalmic and posterior ciliary arteries. Journal of Neuro-
Ophthalmology. 2008;28:320-340.
[6] Hayreh SS. Segmental nature of the choroidal vasculature. British Journal of Ophthalmology. 1975;59:631-648.
[7] Laties AM. Central retinal artery innervation. Archives of Ophthalmology. 1967;77:405-409.
[8] Li H and Grimes P. Adrenergic Innervation of the Choroid and Iris in Diabetic Rats. Current Eye Research. 1993;12:89-94.
[9] Riva CE, Cranstoun SD, Mann RM and Barnes GE. Local choroidal blood flow in the cat by laser Doppler flowmetry.
Investigative Ophthalmology and Visual Science. 1994;35:608-618.
[10] Riva CE, Pournaras CJ and Tsacopoulos M. Regulation of local oxygen tension and blood flow in the inner retina during
hyperoxia. Journal of Applied Physiology. 1986;61:592-598.
[11] Vanderkooi JM, Erecinska M and Silver IA. Oxygen in mammalian tissue: methods of measurement and affinities of various
reactions. American Journal of Physiology. 1991;260:C1131-C1150.
[12] Xiao M and Hendrickson A. Spatial and temporal expression of short, long/medium, or both opsins in human fetal cones.
Journal of Comparative Neurology. 2000;425:545-559.
[13] Yu D-Y and Cringle SJ. Oxygen distribution and consumption within the retina in vascularised and avascular retinas and in
animal models of retinal disease. Progress in Retinal and Eye Research. 2001;20:175-208.
[14] Yu D-Y, Cringle SJ and Su E-N. Intraretinal oxygen distrubution in the monkey retina and the response to systemic hyperoxia.
Investigative Ophthalmology and Visual Science. 2005;4728-4733.
[15] Yu D-Y, Su E-N, Cringle SJ and Yu PK. Isolated preparations of ocular vasculature and their applications in ophthalmic
research. Progress in Retinal and Eye Research. 2003;22:135-169.
[16] Yu D-Y, Cringle SJ, Yu PK and Su E-N. Intraretinal oxygen distribution and consumption during retinal artery occlusion and
hyperoxic ventilation in the rat. Investigative Ophthalmology and Visual Science. 2007;48:2290-2296.
[17] Yu PK, Balaratnasingam C, Morgan WH, Cringle SJ, McAllister IL and Yu D-Y. The structural relationship between the
microvasculature, neurons and glia in the human retina . Investigative Ophthalmology and Visual Science. 2010;51:447-458.
[18] Yu PK, Yu D-Y, Alder VA, Seydel U, Su EN and Cringle SJ. Heterogeneous endothelial cell structure along the porcine retinal
microvasculature. Experimental Eye Research. 1997;65:379-389.
Microscopy: Science, Technology, Applications and Education A. Méndez-Vilas and J. Díaz (Eds.)
©FORMATEX 2010 875
______________________________________________