mitochondrial and peroxisomal oxidation of arachidonic and eicosapentaenoic acid studied in isolated...
TRANSCRIPT

Biochimicu et Biophwicu Actu 879 (1986) 313-321
Elsevier
313
BBA 52428
Mitochondrial and peroxisomal oxidation of arachidonic
and eicosapentaenoic acid studied in isolated liver cells
Erik Christensen, Tor-Arne Hagve and Bjmn 0. Christophersen
Institute of Clinical Biochemistry. Uniuersitv of Oslo, Rikshospitalei, N-0027 Oslo I (Norway)
(Received 7 July 1986)
Key words: Eicosapentaenoic acid: Arachidonic acid; Fatty acid oxidation; Peroxisome; Mitochondrion; Chain elongation; (Rat hepatocyte)
The partitioning between peroxisomal and mitochondrial /3-oxidation of [ l-‘4C]eicosapentaenoic acid (20: 5(n - 3)) and [l-14Cjarachidonic acid (20: 4(n - 6)) was studied. In hepatocytes from fasted rats approximately 70% of the fatty acid substrate was oxidized with oleic, linoleic, eicosapentaenoic and docosahexaenoic (22: 6(n - 3)) acid, even more with adrenic (22: 4(n - 6)) and less with arachidonic acid. When the mitochondriai oxidation was suppressed by fructose refeeding and by ( +)-decanoylcamitine, the fatty acid oxidation in per cent of that in cells from fasted rats was with 18: l(n - 9) 7%, 18: 2(n - 6) 8%, 20: 4(n - 6) 12%, 20: 5(n - 3) 20%, 22: 4(n - 6) 57% and for 22 : 6(n - 3) 29%. The fraction of 14C recovered in palmitate and other newly synthesized fatty acids after fructose refeeding decreased in the order 22 : 4( n - 6) > 22 : 6( n - 3) > 20 : 5( n - 3) > 20 : 4( n - 6) and was very small with 18 : l( n - 9) and 18: 2(n - 6). In cells from both fed and fructose-refed animals 20: 5(n - 3) was efficiently elongated to 22 : 5(n - 3) and 22 : 6( n - 3). 20 : 5(n - 3) and 20 : 4( n - 6) were not elongated after fasting. The phos- pholipid incorporation with [1-‘4C]20: 5(n - 3) decreased during prolonged incubations while it remained stable with [l-14C]arachidonic acid. The results suggest that peroxisomes contribute more to the oxidation of 20 : 5(n - 3) than with 20 : 4(n - 6) although both substrates are probably oxidized mainly in the mitochondria.
Introduction
Since peroxisomal fatty acid /?-oxidation was described [l], the relative roles of mitochondrial and peroxisomal oxidation in the liver and other cells have been discussed. In studies with palmi- tate (16 : 0) and oleate (18 : 1) as substrates differ- ent authors have estimated the peroxisomal oxida- tion to account for less than 10% and up to 30% of the fatty acid oxidation in liver [2-51.
Peroxisomes are believed to play an important role in the oxidation of very long chain fatty acids
Correspondence: Dr. E. Christensen, Institute of Clinical Bio- chemistry, University of Oslo, Rikshospitalet. N-0027 Oslo 1. Norway.
(C,, and longer) [6-lo]. Thus the chain shortening of erucic acid (22:l(n-9)) to 20:1, 18:l and
16 : 1 is mainly a peroxisomal reaction. Very long
chain fatty acids (C&-C,,) are known to accu- mulate in certain inborn errors of metabolism
caused by peroxisomal defects [11,12]. Cz, fatty acids do not accumulate in these diseases.
We have recently studied the oxidation of two polyunsaturated C,, fatty acids, adrenic acid
(22 : 4( n - 6)) and docosahexaenoic acid (22 : 6( n - 3)) in isolated liver cells. The results suggested that 22 : 4(n - 6) was efficiently retroconverted to arachidonic acid (20 : 4(n - 6)) by liver per- oxisomes [13]. The role of peroxisomal P-oxida- tion in liver cells with different fatty acids de- creased in the order 22 : 4( n - 6) > 22 : 6( n - 3) >
00052760/86/$03.50 Q 1986 Elsevier Science Publishers B.V. (Biomedical Division)

314
18 : l(n - 9). This suggests that not only the chain length but also the number and position of double bonds will affect the distribution between per-
oxisomal and mitochondrial aoxidation of each
fatty acid. In assessing the relative roles of mitochondrial
and peroxisomal oxidation it must be kept in mind that both processes are regulated by dietary, hormonal and other factors. Thus fasting and
feeding has strong effects on mitochondrial fatty acid oxidation mediated by malonyl-CoA and other factors [14,15]. The peroxisomal fatty oxida- tion is for example increased by feeding partially hydrogenated fish oil [7,16] and certain drugs such as clofibrate [l].
In the present work the liver metabolism of
arachidonic and eicosapentaenoic acid is com- pared to the metabolism of other unsaturated
fatty acids such as 18: l(n - 9) 18:2(n - 6) 22:4(n-6) and 22:6(n-3).
Materials and Methods
Chemicals [1-‘4C]Arachidonic acid (20 : 4(n - 6)) [1-14C]
eicosapentaenoic acid (20 : $(n - 3)), [1-14C] docosahexaenoic acid (22 : 6(n - 3)), [l-‘4C]oleic acid (18 : l(n - 9)) [l-‘4C]linoleic acid (18 : 2(n - 6)) and [ l-‘4C]linolenic acid (18 : 3( n - 3)) were obtained from New England Nuclear, Boston, MA, U.S.A. [2-‘4C]Docosatetraenoic acid (22 : 4( n - 6))
was from The Radiochemical Centre, Amersham, U.K. ( + )-Decanoylcarnitine was synthesized
according to the method of Biihmer and Bremer [17]. Essentially fatty acid-free bovine serum al- bumin, 4-(2-hydroxyethyl)-l-piperazine-ethane- sulphonic acid (Hepes), collagenase type I and unlabelled arachidonic, eicosapentaenoic, doco- sahexaenoic, oleic, linoleic and linolenic acid were
from Sigma Chemical Co., St. Louis, MO, U.S.A. Docosatetraenoic acid was from Nu Chek Pre. Elysian, MN, U.S.A. 2,6-Di-tert-butyl-p-cresol was from Koch-Light Ltd., Colnbrook, Bucks, U.K. (-)-Carnitine was from Lonza AG, Basle, Switzerland.
Animals Preparation of liver cells. Male weanling rats of
the Wistar strain were from Mollegaard Labora-
tory (Denmark). The animals were fed standard
pellets and had free access to food when not stated otherwise. In some experiments the animals were fasted for 48 h before preparation of liver
cells. Animals which had been fasted for 48 h and then refed 20% (w/v) fructose for 24 h were also
used. Parenchymal liver cells were prepared and
purified according to the method of Seglen [18] except that only 0.03 M Hepes buffer was used in the suspension medium. About (100-300). lo6 cells were obtained from each liver, and 90-95s were viable, as measured by resistance to uptake
of Trypan blue.
Incubations
The concentration of cells in the preparation was approx. 6 . lo6 cells/ml and 1 ml of this
suspension was used (with a total volume of 2 ml of incubation medium in 25-ml Erlenmeyer flasks). Cells were incubated at 37” C in an oxygenated suspension medium [lo] with 1.5% (w/v) bovine
serum albumin with 200 nmol of 14C-labelled fatty acid when not stated otherwise.
The specific activity of all 14C-labelled fatty
acid substrates was 7 mCi/mmol. When indicated the hepatocytes were preincubated with 1 mM ( - )-carnitine or 1 mM ( + )-decanoylcarnitine for
20 min.
Analytical methods The extraction of lipids and the measurement
of radioactive acid-soluble products (as a measure of the rate of P-oxidation) and of radioactive CO, were performed as described by Christiansen [19].
The lipids were extracted by the method of Folch et al. [20] and separated on silicic acid thin-layer plates (Stahl H+) (hexane/ diethyl ether/glacial acetic acid, 70 : 30 : 1, v/v). Aliquots of the total lipid extracts and the free fatty acid, triacylglycerol and phospholipid fractions were transmethylated [21].
In experiments with 14C-labelled fatty acids, the lipid fractions were analyzed by radio-gas chromatography using a Varian 2100 gas chro- matograph connected to an ES1 nuclear radioac- tivity detector with a 1 : 10 outlet splitter. Fatty acid methyl esters were separated at 165 and 200°C using 10% SP-2340 on Supelcoport 100/120

(Supelco Inc. Bellefonte, PA). The peaks were
identified on the basis of the retention time com- pared with the standards.
The solutions used for lipid extraction and thin-layer chromatography contained 2,6-di-tert-
butyl-p-cresol(50 mg/l) as an antioxidant and the lipid extracts were stored under nitrogen gas in
the dark at - 20°C to prevent peroxidation of
unsaturated fatty acids. Cellular protein was determined according to
the method of Lowry et al. [22].
Results
In hepatocytes from fasted rats the oxidation of all the six unsaturated C,,, C,, and C,, fatty acids tested was high (Table I) compared to previ- ous studies with cells from fed animals [13,23]. At low substrate concentrations (0.1 mM) and with
(-)-carnitine approximately 70% of each fatty
acid was oxidized to acid-soluble products and CO, and the rest was esterified under the condi- tions used (Table I). With 22 : 4(n - 6) as the
substrate even more, 80%, was oxidized and with 20 : 4( n - 6) somewhat less, 60%, was oxidized.
( + )-Decanoylcarnitine is known to inhibit mitochondrial oxidation of C,, and C,, fatty acids such as 16: 0 and 18: 1 by competition for the outer long-chain acyl-CoA carnitine acyltransfer-
TABLE I
315
ase. In hepatocytes from fasted rats the degree of
inhibition caused by ( + )-decanoylcarnitine was different with different fatty acid substrates (Ta-
ble I). The formation of acid-soluble oxidation products with ( +)-decanoylcarnitine as a per- centage of that with carnitine was 25-37% with
18 : 1, 18 : 2, 22 : 4 and with 20 : 6 as substrates. In
experiments with 20 : 5( n - 3) and especially with
22 : 4( n - 6) as substrates ( + )-decanoylcarnitine was distinctly less efficient as inhibitor of the fatty
acid oxidation to acid-soluble products (Table I).
Fructose refeeding suppressed the fatty acid oxidation to acid-soluble products and CO, com-
pared to the results with cells from fasted animals (Tables I and II). The degree of suppression caused by fructose refeeding was very different with the different fatty acids used, strongest with the C,, fatty acids and lowest with 22 : 4( n - 6) as sub- strated.
In hepatocytes from rats refed fructose, a con- dition known to stimulate the fatty acid synthesis [24,25], the 14C label from the fatty acid substrates was also recovered in palmitic acid, stearic acid
and oleic acid. These newly synthesized fatty acids are labelled from [‘4C]acetyl-CoA produced by
P-oxidation of the unsaturated fatty acid sub- strates. The total fatty acid P-oxidation can there- fore be calculated from the sum of “C-labelled
acid-soluble products, CO, and newly synthesized
OXIDATION OF 14C-LABELLED FATTY ACIDS IN HEPATOCYTES FROM FASTED RATS IN THE PRESENCE OF
( - )-CARNITINE OR ( + )-DECANOYLCARNITINE
The incubation conditions were as described in Materials and Methods. Labelled fatty acid (0.1 mM) and hepatocytes 16.3-19.2 mg
protein) were incubated for 90 min. The results are expressed as nmol 14C-fattv acid oxidized. The oxidation to acid-soluble products _ in the presence of (+)-decanoylcamitine is also expressed as per cent of the oxidation in the presence of ( - )-camitine given in
parenthesis. Means? SD. of two parallel incubations of hepatocytes form three different livers are given,
Substrate Products and additions
( - )-camitine: ( + )-decanoylcamitine: ( - )-camitine: ( + )-decanoylcamitine: acid-soluble
products
(nmol)
acid-soluble
products
(nmol)
% of result
with
( - )-camitine
CO, CO, (nmol) (nmol)
18:l(n -9) 116.4k 5.7 31.1+ 2.6 27 21(.9*4.3 25.3 * 3.6
18:2(n -6) 117.2* 7.3 39.7 f 1.7 34 32.4kO.9 21.7 k 4.2 20:4(n -6) 104.0* 6.8 38.3 f 3.3 37 16.5 f 3.0 15.2* 1.8
20:5(n -3) 124.05 11.5 70.3 f 6.0 57 23.013.5 19.8 f 3.9
22:4(n -6) 156.7* 7.8 117.0k6.8 75 8.5 * 5.3 9.6 i 2.5
22:6(n-3) 120.35 9.6 41.8i3.6 35 31.0i4.4 18.7i 3.1

316
TABLE II
FORMATION OF 14C-LABELLED 14 : 0, 16 : 0. 16 : 1, 18 : 0 AND 18 : 1 AND OF “-C-LABELLED ACID-SOLUBLE PROD-
UCTS AND t4C0, IN HEPATOCYTES FROM FRUCTOSE-REFED RATS
The incubation conditions were as described in Materials and Methods. Labelled fattv acids (0.1 mM) were incubated with isolated
liver cells (17.5-21.8 mg protein) for 90 min. (-)-Carnitine or (+)-decanoylcarnitine were used as indicated. The results are
expressed as nmol i4C-labelled fatty acid synthesized or oxidized. Means i SD. of two parallel incubations from three different livers
are given.
Substrate Products and additions
18:l(n-9)
18:2(n -6)
20:4(n -6)
20:5(n -3)
22:4(n -6)
22:6(n -3)
acid-soluble products
( - )-camitine ( + )-decanoyl-
camitine
12.1 k 1.8 4.1+ 2.3
21.0 + 2.6 5.2* 1.6
35.6 f 2.3 7.7 f 2.3
45.7 * 5.1 11.3i1.4
92.0 + 6.7 49.1 f 5.5
39.8 * 4.2 21.9+ 3.2
newly synthesized fatty acids
(- )-camitine ( + )-decanoyl-
carnitine
3.OkO.8 3.4 + 0.7
4.6 + 0.5 3.5*0.9
5.6k1.7 5.4 + 0.6
16.1* 1.1 13.2k1.5
29.1 f 5.3 42.4 k 3.6
5.3 f 1.3 19.5 i 1.0
co* (~ )-camitine ( + )-decanoyl-
camitine
5.8kO.S 2.3 i 1.2
9.6 k 3.0 3.1 & 1.7
2.5 + 0.2 0.9 * 0.2
16.2k2.2 2.7il.O
4.2kO.2 2.2kl.l
11.6k1.4 3.3 k 1.7
fatty acids (Table II). Thus calculated from results presented in Tables I and II the total fatty acid oxidation with carnitine in fructose-refed cells as a percentage of the oxidation in fasted cells was for 18 : 1 14%, 18 : 2 238, 20 : 4 34% 20: 5 52%:. 22.4 76% and 22: 6 37%.
Mitochondrial fatty acid oxidation should be very strongly suppressed by the combined used of
two different principles in hepatocytes from fruc- tose-refed animals incubated with (+)-decanoyl-
carnitine. Thus carbohydrate refeeding suppresses mitochondrial fatty acid oxidation probably medi- ated by malonyl-CoA, which inhibits long-chain
acyl-CoA camitine acyltransferase, which is also
the target for ( + )-decanoylcarnitine. When the fatty acid oxidation in hepatocytes
TABLE III
EFFECT OF FASTING AND FRUCTOSE REFEEDING ON FATTY ACID CHAIN ELONGATION IN ISOLATED LIVER
CELLS
The incubation conditions were as described in Materials and Methods, Labelled fatty acids (0.1 mM) ad ( + )-decanoylcamitine were
incubated with hepatocytes (17.5-20.3 mg protein) for 90 min. The results are expressed as nmol fatty acid elongated, desaturated or
remaining as esterified substrate fatty acid. Means + S.D. of two parallel incubations from two different livers are given.
Substrate Fatty acid nmol fatty acid esterified in phospholipid and triacylglycerol
20:5(n -3) 20:5 22:5+22:6
fed
108.Ok 5.8
31.7 * 1.5
20:4(n -6) 20:4 152.0 f 6.8 22:4 7.0 i 2.4
18:3(n -3) 18:3 1140* 9.2 18:4 0
20:4+20:5+22:5 20.9*4.1
18:2(n -6) 18:2 139.05 7.5 18.3 0
20:3+20:4+22:4 4.8k1.7
fasted fructose refed
79.0* 3.5
0
160.0+ 7.4 0
75.1Ifr 3.3 10.8i 3.6
0
129.0* 11.6
7.8+ 2.4
0
137.5* 4.0 35.0* 6.9
172.05 3.5 13.6k 0.7
120.4? 10.1 6.4+ 2.7
60.3i 5.2
167.5f 9.8
0
18.li 3.8

from fructose-refed rats and in the presence of
( + ) - decanoylcarnitine was calculated as a per- centage of the oxidation in cells from fasted rats with (-)-carnitine (Tables I and II) the results were for 18 : 1 7%, 18 : 2 8%, 20 : 4 12%, 20 : 5 20%, 22 : 4 57% and 22: 6 29%.
New 14C-labelled C,,-C,* fatty acids were formed from acetyl-CoA liberated by P-oxidation
of the 14C-labelled fatty acid substrates with
hepatocytes from fructose-refed animals but not in cells from fasted animals.
Table II shows that only small amounts of new
fatty acids were formed with 18: 1, 18: 2, and 20 : 4 as substrates. Distinctly more of the new
fatty acids were formed with 20 : 5 and 22 : 6 as
substrates, while the largest amounts were synthe- sized with 22: 4 as the substrate. With 22: 6 new
fatty acids appeared in larger amounts only in the presence of ( + )-decanoylcarnitine. With 20 : 5 new fatty acids were formed also in the absence of ( + )-decanoylcarnitine.
With all the 14C-labelled substrate fatty acids used palmitic acid accounted for 50-60% of the newly synthesized fatty acids, 14: 0, 16 : 1 and 18 : 1 for approximately 10% each and 14: 1 and 18 : 0 for only trace amounts (not shown).
Table III shows that the feeding state has pro- found effects on the chain elongation of essential
fatty acid substrates. In hepatocytes from rats starved for 48 h no chain elongation of 20: 4 or
20 : 5 was observed. With 18 : 2 and 18 : 3( n - 3)
TABLE IV
317
as substrates A6 desaturation to 18 : 3(n - 6) and
18 : 4(n - 3), respectively, was found. With hepatocytes from fed animals and fructose-refed animals a significant chain elongation of 20 : 4( n
-6)to22:4(n-6)andevenmoreof20:5(n-3)
to 22 : 5(n - 3) and 22 : 6( n - 3) was found. The chain elongation of 20 : 5( n - 3) in the ordinary fed state was nearly as efficient as that in cells from fructose-refed animals. In contrast, the
synthesis of new 14C-labelled C,,-C,, fatty acids from 14C-labelled 20 : 5( n - 3) and 22 : 4( n - 6) is much higher in cells from fructose-refed animals (Table II) than in cells from fed rats where only
trace amounts of new 14C-labelled fatty acid are formed (not shown).
In hepatocytes from fasted rats and with (-)- carnitine very little of all six 14C-labelled fatty
acid substrates was recovered in triacylglycerol
corresponding to rapid fatty acid oxidation under these conditions (Table IV). Table V shows that the fatty acid esterification in triacylglycerol was, as expected, much higher in hepatocytes from animals refed fructose than with cells from fasted animals. ( + )-Decanoylcarnitine increased the incorporation into triacylglycerol with all the fatty acids used in hepatocytes from fasted rats and with cells from fructose-refed rats. The fact that
less fatty acid was oxidized in cells from fructose- refed animals and more substrate was thus availa- ble for esterification than in the fasted state did
not increase the fatty acid incorporation in phos-
ESTERIFICATION IN PHOSPHOLIPIDS AND TRIACYLGLYCEROL OF 14C-LABELLED FATTY ACIDS IN HEPATO-
CYTES FROM FASTED RATS
The incubation conditions were as described in Materials and Methods. Labelled fatty acids (0.1 mM) and hepatocytes (16.3-19.2 mg
protein) were incubated for 90 min with (- )-carnitine or with ( + )-decanoylcamitine. The results are expressed as nmol 14C-fatty
acid esterified. Means & SD. of two parallel incubations from three different livers are given. Data on fatty acid oxidation in the same
experiments are given in Table I.
Substrates
18:l(n-9)
18:2(n -6)
20:4(n -6)
20:5(n -3)
22:4(n -6)
22:6(n-3)
Products and additions
triacylglycerol
( - )-carnitine ( + )-decanoylcarnitine
8.4* 2.9 97.5 f 7.3
6.5 & 1.4 87.7 f 6.7
10.4+ 3.9 61.8 f 2.2
11.4k4.2 66.4+ 5.9 8.6 k 3.5 34.8k4.2
15.4k3.1 45.4+ 5.1
phosphohpids
( - )-camitine
32.8 5 2.2
37.2?4.5
58.3 of- 6.2
28.0+ 2.7
18.0 k 3.2
21.5?4.7
( + )-decanoylcamitine
20.4 * 3.3
35.1+ 5.4
59.3 5 7.6
24.5kO.8
12.5? 3.1 39.6 & 4.0

318
TABLE V
ESTERIFICATION IN PHOSPHOLIPIDS AND TRIACYLGLYCEROL OF 14C-LABELLED FATTY ACIDS IN HEPATO-
CYTES FROM FRUCTOSE-REFED RATS
The incubation conditions were as described in Materials and Methods. Labelled fatty acids (0.1 mM) and hepatocytes (17.3-21.8 mg
protein) were incubated for 90 min with (-)-camitine or (+ )-decanoylcarnitine. The results are expressed as nmol t4C-fatty acid
esterified. Means* SD. of two parallel incubations from three different livers are given. Data on fatty acid oxidation in the same
experiments are given in Table II.
Substrates Products and additions
18:l(n-9)
18:2(n -6)
20:4(n -6)
20:5(n-3)
22:4(n -6)
22:6(n -3)
triacylglycerol
( - )-camitine
128.2* 7.8
128.0& 9.1
92.7+ 5.8
93.2k 7.5
99.65 4.6
84.8 * 10.2
( + )-decanoylcamitine
156.2& 7.5
161.5k 8.9
148.65 4.0
157.3+ 6.2
135.Oi 11.6
137.95 3.3
phospholipids
( - )-camitine
18.7 f 3.1
31.4*4.2
51.6 f 7.8
31.3 f 6.2
26.6 f 3.0
25.7k4.4
(+)-decanoylcarnitine
12.Oi 2.8
19.3~2.1
34.3 * 4.5
19.9 + 2.6
19.6 k 3.7
28.7 k 5.3
pholipids (Tables IV and V). Neither did (+)-de- canoylcarnitine increase the fatty acid esterifica-
tion into phospholipids under the conditions used (Tables IV and V). In some cases (+)-decanoyl- carnitine even seemed to decrease the incorpora-
tion of 14C-labelled substrates into phospholipids at the low substrate concentrations used (0.1 mm). More of the 14C-labelled fatty acid was incorpo-
rated in the phospholipid fraction with 20 : 4( n - 6) as the substrate than with 20 : 5( n - 3) or with any of the other substrates under all conditions used (Tables IV and V).
In most of the present experiments a fixed initial fatty acid substrate concentration (0.1 mM) was used. Fig. 1 shows that the oxidation to acid-soluble products increased linearly with in- creasing initial substrate concentrations both with 20 : 4 and 20 : 5 and with 18 : 1. ( +)-Decanoyl-
carnitine caused the same inhibition (in W) of the
oxidation to acid-soluble products at all substrate concentrations used (0.04-0.4 mM).
The esterification in phospholipids also increased with increasing substrate concentrations with 20 : 4( n - 6) 20 : 5(n - 3) and with 18 : l(n - 9). With oleic acid the incorporation into phos- pholipids was distinctly lower than with 20 : 4(n - 6) and 20 : 5( n - 3) at all substrate concentrations used. ( + )-Decanoylcarnitine had little effect on the incorporation of all three fatty acids at low substrate concentrations. At higher concentrations of 20 : 4 and 20 : 5 ( + )-decanoylcarnitine slightly
increased the phospholipid incorporation of the two 14C-labelled substrates.
these experiments with hepatocytes from fed
[1&c] fatty actd substrate (mM)
Fig. 1. Oxidation to acid-soluble products and esterification in
phospholipids of 14C-labelled fatty acid in hepatocytes from fed rats. The incubation conditions were as described in
Materials and Methods. “C-labelled fatty acid and hepato-
cytes (19.7 mg protein) were incubated for 90 min. 0, incuba-
tions with (- )-camitine, and 0, with ( + )-decanoylcamitine.
Oxidation to acid-soluble products of (a) 20:4(n -6). (b)
20 : 5( n - 3) and (c) 18 : l( n - 9). Esterification of phospholi- pids of (d) 20:4(n -6). (e) 20:5(n -3) and (f) 18:l(n -9).
The results are expressed as nmol labelled fatty acid oxidized
or esterified.
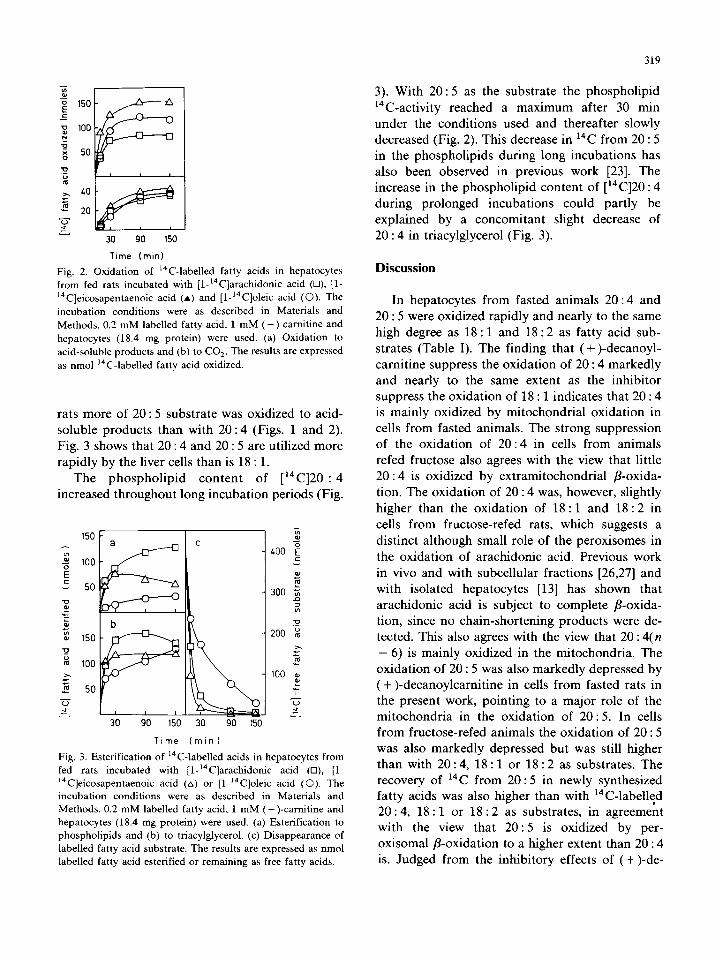
319
30 90 150
Time (min)
Fig. 2. Oxidation of “C-labelled fatty acids in hepatocytes
from fed rats incubated with [l-‘4C]arachidonic acid (0) [l-
“‘Cleicosapentaenoic acid (A) and [1-t4C]oleic acid (0). The
incubation conditions were as described in Materials and
Methods. 0.2 mM labelled fatty acid, 1 mM (- )-carnitine and
hepatocytes (18.4 mg protein) were used. (a) Oxidation to
acid-soluble products and (b) to CO,. The results are expressed
as nmol “C-labelled fatty acid oxidized.
rats more of 20 : 5 substrate was oxidized to acid- soluble products than with 20 : 4 (Figs. 1 and 2).
Fig. 3 shows that 20 : 4 and 20 : 5 are utilized more rapidly by the liver cells than is 18 : 1.
The phospholipid content of [14C]20 : 4 increased throughout long incubation periods (Fig.
Time (mln)
Fig. 3. Esterification of 14C-labelled acids in hepatocytes from fed rats incubated with [l-‘4C]arachidonic acid (o), [l-
t4C]eicosapentaenoic acid (A) or [l-r4C]oleic acid (0). The incubation conditions were as described in Materials and
Methods. 0.2 mM labelled fatty acid, 1 mM (-)-camitine and
hepatocytes (18.4 mg protein) were used. (a) Esterification to
phospholipids and (b) to triacylglycerol. (c) Disappearance of
labelled fatty acid substrate. The results are expressed as nmol labelled fatty acid esterified or remaining as free fatty acids.
3). With 20 : 5 as the substrate the phospholipid
14C-activity reached a maximum after 30 min under the conditions used and thereafter slowly decreased (Fig. 2). This decrease in 14C from 20 : 5 in the phospholipids during long incubations has also been observed in previous work [23]. The increase in the phospholipid content of [14C]20 : 4
during prolonged incubations could partly be explained by a concomitant slight decrease of 20 : 4 in triacylglycerol (Fig. 3).
Discussion
In hepatocytes from fasted animals 20: 4 and 20 : 5 were oxidized rapidly and nearly to the same
high degree as 18 : 1 and 18 : 2 as fatty acid sub- strates (Table I). The finding that ( +)-decanoyl-
carnitine suppress the oxidation of 20 : 4 markedly and nearly to the same extent as the inhibitor
suppress the oxidation of 18 : 1 indicates that 20 : 4 is mainly oxidized by mitochondrial oxidation in cells from fasted animals. The strong suppression of the oxidation of 20: 4 in cells from animals refed fructose also agrees with the view that little 20 : 4 is oxidized by extramitochondrial P-oxida- tion. The oxidation of 20 : 4 was, however, slightly
higher than the oxidation of 18 : 1 and 18 : 2 in
cells from fructose-refed rats, which suggests a distinct although small role of the peroxisomes in
the oxidation of arachidonic acid. Previous work
in vivo and with subcellular fractions [26,27] and with isolated hepatocytes [13] has shown that arachidonic acid is subject to complete P-oxida- tion, since no chain-shortening products were de- tected. This also agrees with the view that 20 : 4( n - 6) is mainly oxidized in the mitochondria. The oxidation of 20 : 5 was also markedly depressed by (+)-decanoylcarnitine in cells from fasted rats in the present work, pointing to a major role of the mitochondria in the oxidation of 20 : 5. In cells from fructose-refed animals the oxidation of 20 : 5
was also markedly depressed but was still higher than with 20 : 4, 18 : 1 or 18 : 2 as substrates. The recovery of t4C from 20 : 5 in newly synthesized fatty acids was also higher than with “C-labelled 20 : 4, 18 : 1 or 18 : 2 as substrates, in agreement with the view that 20: 5 is oxidized by per- oxisomal P-oxidation to a higher extent than 20 : 4 is. Judged from the inhibitory effects of (+)-de-

320
canoylcarnitine and by fructose refeeding the per- oxisomal oxidation of 20: 5 was still distinctly lower than with 22 : 4 as the substrate.
The finding that 20 : 5, 20 : 4 and 18 : 3(n - 3) are not chain-elongated in cells from fasted animals may be due to the low acetyl-CoA carboxylase activity in this state, since malonyl- CoA supplies the C, units used for chain elonga-
tion. Sprecher [28] has shown that in the presence of added malonyl-CoA the chain elongation of
18 : 3(n - 6) in isolated microsomes is reduced by approximately 50% compared to activity in micro-
somes from fed rats. Table III shows that 20 : 5 was nearly as efficiently chain-elongated in the fed
state as in cells from animals refed fructose. New 14C-labelled fatty acids were, however, formed
much more efficiently in cells from fructose-refed rats than in cells from fed animals. Probably the
elongase which forms 22 : 5( n - 3) from 20 : 5( n - 3) and malonyl-CoA is sufficiently active in both the fed and the fructose-refed state, while the fatty acid synthetase is much more active in forming
new 14C-labelled C,, and C,, fatty acids in the fructose-refed state than with cells from fed animals.
At present attempts are made to reduce throm- boxane A, formation in human platelets by giving a dietary supplement of 20 : 5( n - 3). The present
experiments may suggest that 20 : 5(n - 3) is oxidized to a large extent in the fasted state, while it is efficiently elongated to 22 : 5(n - 3) after
feeding.
( +)-Decanoylcarnitine constantly increased the fatty acid incorporation in triacylglycerol, as ex- pected, since it inhibits fatty acid oxidation.
( + )-Decanoylcarnitine decreased the incorpora- tion of “C-labelled fatty acid into the phospholi- pids in some experiments. Thus at low 20 : 4 con- centrations, ( + )-decanoylcarnitine decreased the esterification in the phospholipids. Since ( + )-de- canoylcarnitine will also inhibit the oxidation of endogenous unlabelled fatty acids, such endoge- nous substrates may compete with and thus de- crease the phospholipid incorporation of [14C] 20: 4. In agreement with this view it was found that at high substrate concentrations (+)-de- canoylcarnitine increased the phospholipid incor-
poration of both [14C]20: 4 and [14C]20: 5 (Fig.
1).
20 : 4( n - 6) was esterified in the phospholipids to a greater extent than were 18: 1, 18: 2, 22: 4
and 22 : 6 under all conditions used (Tables IV and V and Fig. 1). The incorporation of 20 : 5( n -
3) into the phospholipids varied with the experi- mental conditions. After short incubation periods distinctly more of 20 : 4(n - 6) and 20 : 5(n - 3)
was recovered in the phospholipids than with 18 : 1. Thereafter during prolonged incubations the phos- pholipid content of 20 : 4( n - 6) increased while
that of 20 : 5(n - 3) decreased. At high substrate concentrations the phospholipid incorporation of
20 : 5(n - 3) was also more efficient. It is possible that 20 : 5( n - 3) is more readily removed from the
phospholipids by the continuous deacylation-re- acylation reactions than 20 : 4( n - 6) is. Thus in feeding experiments Iritani et al. [29] found that dietary 20 : 5( n - 3) is more rapidly removed from
the phospholipids than 20 : 4( n - 6) is after chang- ing to a fat-free diet.
Previous work with erucic acid (22 : l(n - 9)) has shown that this fatty acid is shortened mainly by two cycles of peroxisomal P-oxidation to 18 : l( n - 9) while smaller amounts of 20 : 1( n - 9) and 16 : l(n - 9) are also formed. The present results agree with the view that 22 : 4(n - 6) is shortened probably by the peroxisomes by only one cycle of P-oxidation to arachidonic acid.
The two C,, fatty acid products of peroxisomal /3-oxidation, 20 : l( n - 9) and 20 : 4( n - 6), are thus
treated differently in that 20: l(n - 9) is further shortened while 20 : 4( n - 6) probably leaves the
peroxisomes and is oxidized in the mitochondria or incorporated mainly into the phospholipids.
Acknowledgements
E.C. is Fellow of the Norwegian Cancer Society. The technical assistance of Yngvar Johansen and Siri Tversdal and the secretarial assistance of Tone Omland is greatly appreciated.
References
Lazarow, P.B. and deDuve, C. (1976) Proc. Nat]. Acad. Sci. USA 73,2043-2046
Mannaerts, G.P., Debeer, L.J., Thomas, J. and DeSchepper.
P.J. (1979) J. Biol. Chem. 254, 4585-4595
Foerster, E.-C., FZhrenkemper, T., Rabe, U., Graf, P. and Sies, H. (1981) B&hem. J. 196. 705-712

321
4 Kondrup, J. and Lazarow. P.B. (1985) Biochim. Biophys.
Acta 835, 147-153
5 Veerkamp. J.H. and van Moerkerk. H.T.B. (1986) Biochim.
Biophys. Acta 875. 301-310
6 Christiansen. R.Z.. Christiansen, E.N. and Bremer, J. (1979)
Biochim. Biophys. Acta 673. 417-429
7 Neat, C.E., Thomassen, M.S. and Osmundsen, H. (1981)
Biochem. J. 196. 149-159
8 Osmundsen. H., Neat, C.E. and Norum. K.R. (1979) FEBS
Len. 99, 292-296
9 Norseth, J. and Christophersen, B.O. (1978) FEBS Lett. 88,
353-357
10 Christophersen. B.O.. Norseth. J., Thomassen, M.S.. Chris-
tiansen, E.N., Norum, K.R.. Osmundsen, H. and Bremer, J.
(1982) in Nutritional Evaluation of Long-Chain Fatty Acids
in Fish Oil (Barlow, S.M. and Stansby, M.E., eds.). pp.
89-130, Academic Press, London
11 Kawamura. N., Moser, H.W. and Kishimoto. Y. (1981)
Biochem. Biophys. Res. Commun. 99, 121661225
12 Singh. I., Moser, A.E., Goldfisher, S. and Moser, H.W.
(1984) Proc. Natl. Acad. Sci. USA 81, 4203-4207
13 Hagve. T.-A. and Christophersen, B.O. (1986) Biochim.
Biophys. Acta 875, 165-173
14 McGarry, J.D., Marmaerts. G.P. and Foster, D.W. (1977) J.
Clin. Invest. 60, 265-270
15 Harano. Y., Kashiwagi, A., Kojima, H., Suzuki, M.,
Hashimoto, T. and Shigeta, Y. (1986) FEBS Lett. 188.
267-272
16 Norseth, J. and Thomassen, MS. (1983) Biochim. Biophys.
Acta 751, 312-320
17 Blihmer, T. and Bremer, J. (1968) Biochim. Biophys. Acta
152. 259-267
18 Seglen, P.O. (1973) Exp. Cell. Res. 82. 391-398
19 Christiansen, R.Z. (1977) Biochim. Biophys. Acta 488,
249-262
20 Folch, J., Lees, M. and Sloane-Stanley, G.H. (1957) J. Biol.
Chem. 226, 497-509
21 Metcalf, L.D. and Schmitz. A.A. (1961) Anal. Chem. 33,
363-368
22 Lowry, O.H., Rosebrough, N.J.. Farr, A.L. and Randall,
R.J. (1951) J. Biol. Chem. 193, 265-275
23 Hagve. T.-A. and Christophersen. B.O. (1984) Biochim.
Biophys. Acta 796. 205-217
24 Bruckdorfer, K.R., Kahn, I.H. and Yudkin. J. (1972) Bio-
them. J. 129, 439-446
25 Volpe, J.J. and Vagelos, R.R. (1974) Proc. Natl. Acad. Sci.
USA 71. 889-893
26 Stoffel, W.. Ecker, W., Assad, H. and Sprecher. H. (1970)
Hoppe-Seyler’s Z. Physiol. Chem. 351, 1545-1554
27 Kunau, W.H. and Couzens, B. (1971) Hoppe-Seyler’s Z.
Physiol. Chem. 352,1297-1305
28 Sprecher, H. (1974) Biochim. Biophys. Acta 360. 113-123
29 Iritani. N. and Narita. R. (1984) Biochim. Biophys. Acta
793.441-447