modulation of anthracycline-induced myofibrillar...
TRANSCRIPT

Modulation of Anthracycline-Induced Myofibrillar Disarrayin Rat Ventricular Myocytes by Neuregulin-1�
and Anti-erbB2Potential Mechanism for Trastuzumab-Induced Cardiotoxicity
Douglas B. Sawyer, MD, PhD; Christian Zuppinger, PhD; Thomas A. Miller, BA;Hans M. Eppenberger, PhD; Thomas M. Suter, MD
Background—There is an increased incidence of heart failure in patients treated concurrently with anthracyclines and thechemotherapeutic anti-erbB2 agent trastuzumab (Herceptin). On the basis of our previous studies with recombinantneuregulin-1� (NRG-1�), a ligand for the erbB2 receptor tyrosine kinase, we hypothesized that activation of erbB2 byanti-erbB2 versus NRG-1 would cause differential effects on myocyte intracellular signaling as well as anthracycline-induced myofibrillar injury and might potentially account for the clinical toxicity of trastuzumab in the setting ofconcurrent anthracycline therapy.
Methods and Results—We tested this hypothesis using adult rat ventricular myocytes (ARVMs) in culture, assessingmyofibrillar structure by immunostaining for myomesin and filamentous actin. Activation of erbB2, extracellularsignal–regulated kinase 1/2 (Erk1/2), and Akt was assessed by use of antibodies to phosphorylated activated receptoror kinase detected by immunoblot. ARVMs treated with doxorubicin (0.1 to 0.5 �mol/L) showed a concentration-dependent increase in myofilament disarray. NRG-1� (10 ng/mL) activated erbB2, Erk1/2, and Akt in ARVMs andsignificantly reduced anthracycline-induced disarray. In contrast to NRG-1�, anti-erbB2 (1 �g/mL) caused rapidphosphorylation of erbB2 but not Erk1/2 or Akt, with downregulation of erbB2 by 24 hours. Concomitant treatment ofmyocytes with anti-erbB2 and doxorubicin caused a significant increase in myofibrillar disarray versus doxorubicinalone.
Conclusions—NRG-1�/erbB signaling regulates anthracycline-induced myofilament injury. The increased susceptibilityof myofilaments to doxorubicin in the presence of antibody to erbB2 may explain the contractile dysfunction seen inpatients receiving concurrent trastuzumab and anthracyclines. (Circulation. 2002;105:1551-1554.)
Key Words: erbB2 � cardiotoxicity � neuregulins � anthracyclines � myocytes
Treatment of metastatic breast cancer with anthracyclinesand trastuzumab, a novel therapy derived from an anti-
body to the erbB2 receptor tyrosine kinase, results in amarked increase of left ventricular dysfunction and symptom-atic heart failure.1 ErbB2 is a member of the epidermalgrowth factor receptor family, and along with neuregulin andthe erbB4 receptor, it plays an essential role in cardiacdevelopment.2–4 We have shown that recombinant neuregulin1� (NRG-1�) activates both erbB2 and erbB4 receptortyrosine kinase activity and promotes growth, myofilamentorganization, and survival of isolated cardiac myocytes.5,6
The clinical observation of the cardiotoxicity of trastuzumaband anthracyclines suggests that the neuregulin/erbB systemmodulates the response of the myocardium to anthracyclines.Possible mechanisms for this toxicity are alterations in the
structure,7 gene expression,8 and survival9 of cardiac myo-cytes. The main purpose of this study was to test thehypothesis that trastuzumab alters the susceptibility of myo-cytes to anthracycline-induced myofibrillar disarray. Wetherefore characterized the effect of anthracyclines on myo-cyte myofibrillar structure in isolated adult rat cardiac myo-cytes in primary culture and examined the effect of NRG-1�and an antibody to erbB2 with properties similar to trastu-zumab10 on erbB2 signaling and anthracycline-inducedchanges in myofibrillar structure.
MethodsChemicalsRecombinant NRG-1� and NRG-1� were purchased from NeoMar-kers, as were biological-grade antibodies to rodent erbB2 (Clone
Received November 29, 2001; revision received February 5, 2002; accepted February 5, 2002.From the Cardiovascular Medicine Section, Department of Medicine, Boston Medical Center and Myocardial Biology Unit, Boston University School
of Medicine, Boston, Mass (D.B.S., T.A.M.), and the Institute of Cell Biology, Swiss Federal Institute of Technology, Zurich, and Swiss CardiovascularCenter Bern, Inselspital, Bern, Switzerland (C.Z., H.M.E., T.M.S.).
Correspondence to Thomas M. Suter, MD, Cardiology, Inselspital, CH-3010 Bern, Switzerland. E-mail [email protected]© 2002 American Heart Association, Inc.
Circulation is available at http://www.circulationaha.org DOI: 10.1161/01.CIR.0000013839.41224.1C
1551
by guest on May 14, 2018
http://circ.ahajournals.org/D
ownloaded from
B10, Ab-9). All other chemicals, including doxorubicin, werepurchased from Sigma.
Preparation of Cardiac MyocytesAdult rat ventricular myocytes (ARVMs) were isolated from femaleSprague-Dawley rats weighing 150 to 200 g as previously de-scribed.11 Culture medium was based on M-199 (Amimed orGibco-BRL) and contained 20 mmol/L creatine (Sigma), 1% 100-U/mL penicillin/streptomycin (Gibco-BRL), 10% preselected FCS(Seramed), and 10 �mol/L cytosine arabinoside (Sigma-Aldrich).ARVMs were treated on day 7 in culture.
Immunofluorescence MicroscopyCell cultures were fixed and myomesin and filamentous actin werestained as previously described with primary antibody to myome-sin,12 secondary antibodies conjugated to FITC or cyanine-5 fromJackson ImmunoResearch, and rhodamine-phalloidin from Molecu-lar Probes. Stained preparations were analyzed with a Leica confocalscanner TCS NT on the inverted microscope Leica DMIRB-E.Myofibrillar disarray was assessed by an investigator blinded totreatment using a Zeiss Axioplan fluorescence microscope equippedwith a 63[times] oil immersion objective. A total of 200 to 250myocytes were counted for each experimental condition in eachexperiment.
Detection of Erk1/2, Akt/PKB, erbB2, anderbB4 PhosphorylationActivated Erk1/2 and Akt were detected with a PhosphoPlus Akt/PKB (Ser473) antibody kit and p44/42 MAP kinase assay kit (NewEngland BioLabs Inc) as previously described.13 Separate mem-branes were probed with an anti-Akt/PKB and anti-erk1/2 antibodiesto ensure equal loading. Activation of the erbB2 receptor tyrosinekinase was detected as previously described5 by immunoprecipita-tion from volumes of cell lysates in RIPA buffer (500 �g totalprotein) with antibodies to erbB2 and immunodetection using anti-phosphotyrosine antibody (Santa Cruz Biotechnology), horseradishperoxidase–conjugated goat anti-mouse secondary antibody (Sigma-Aldrich), and chemiluminescence detection.
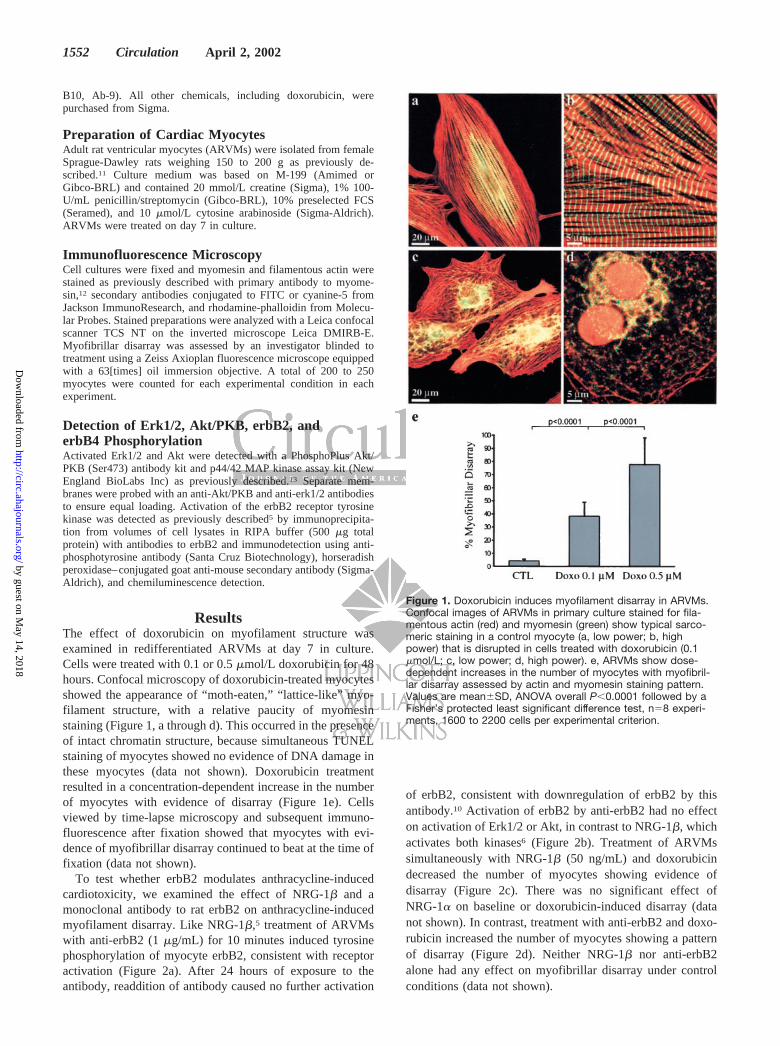
ResultsThe effect of doxorubicin on myofilament structure wasexamined in redifferentiated ARVMs at day 7 in culture.Cells were treated with 0.1 or 0.5 �mol/L doxorubicin for 48hours. Confocal microscopy of doxorubicin-treated myocytesshowed the appearance of “moth-eaten,” “lattice-like” myo-filament structure, with a relative paucity of myomesinstaining (Figure 1, a through d). This occurred in the presenceof intact chromatin structure, because simultaneous TUNELstaining of myocytes showed no evidence of DNA damage inthese myocytes (data not shown). Doxorubicin treatmentresulted in a concentration-dependent increase in the numberof myocytes with evidence of disarray (Figure 1e). Cellsviewed by time-lapse microscopy and subsequent immuno-fluorescence after fixation showed that myocytes with evi-dence of myofibrillar disarray continued to beat at the time offixation (data not shown).
To test whether erbB2 modulates anthracycline-inducedcardiotoxicity, we examined the effect of NRG-1� and amonoclonal antibody to rat erbB2 on anthracycline-inducedmyofilament disarray. Like NRG-1�,5 treatment of ARVMswith anti-erbB2 (1 �g/mL) for 10 minutes induced tyrosinephosphorylation of myocyte erbB2, consistent with receptoractivation (Figure 2a). After 24 hours of exposure to theantibody, readdition of antibody caused no further activation
of erbB2, consistent with downregulation of erbB2 by thisantibody.10 Activation of erbB2 by anti-erbB2 had no effecton activation of Erk1/2 or Akt, in contrast to NRG-1�, whichactivates both kinases6 (Figure 2b). Treatment of ARVMssimultaneously with NRG-1� (50 ng/mL) and doxorubicindecreased the number of myocytes showing evidence ofdisarray (Figure 2c). There was no significant effect ofNRG-1� on baseline or doxorubicin-induced disarray (datanot shown). In contrast, treatment with anti-erbB2 and doxo-rubicin increased the number of myocytes showing a patternof disarray (Figure 2d). Neither NRG-1� nor anti-erbB2alone had any effect on myofibrillar disarray under controlconditions (data not shown).
Figure 1. Doxorubicin induces myofilament disarray in ARVMs.Confocal images of ARVMs in primary culture stained for fila-mentous actin (red) and myomesin (green) show typical sarco-meric staining in a control myocyte (a, low power; b, highpower) that is disrupted in cells treated with doxorubicin (0.1�mol/L; c, low power; d, high power). e, ARVMs show dose-dependent increases in the number of myocytes with myofibril-lar disarray assessed by actin and myomesin staining pattern.Values are mean�SD, ANOVA overall P�0.0001 followed by aFisher’s protected least significant difference test, n�8 experi-ments, 1600 to 2200 cells per experimental criterion.
1552 Circulation April 2, 2002
by guest on May 14, 2018
http://circ.ahajournals.org/D
ownloaded from

DiscussionThe erbB2 receptor tyrosine kinase is expressed in themyocardium by ventricular myocytes and is essential fornormal cardiac development.2 Activation of erbB2 and erbB4by recombinant NRG-1� leads to myocyte growth andinhibition of apoptosis in vitro.5 The present findings supporta role for the NRG/erbB system in modulating myocardialresponse to anthracycline-induced injury. Moreover, theseobservations suggest a potential mechanism for the clinicalobservation of increased cardiotoxicity of anthracyclines inpatients receiving trastuzumab, a monoclonal antibody toerbB2.1
Anthracycline-induced myocardial injury is accompaniedby an increase in myofibrillar disarray that is seen inpatients,14 animals treated with anthracyclines in vivo,15 andin vitro studies of cardiac myocytes.7 The close relationshipbetween myocyte ultrastructural damage and contractile dys-function after anthracycline therapy suggests a mechanisticrelationship.16 We found that even myocytes with evidence ofmyofibrillar degradation continue to beat in culture, althoughpresumably these damaged cells generate less force. Thatthese cells are viable, without evidence of DNA fragmenta-tion, suggests that this damage may be reversible. It is alsointeresting that myofibrillar damage after anthracycline treat-ment both in vivo14 and in vitro (our own data, as well asReference 7) is patchy, with affected myocytes seen adjacentto structurally normal myocytes. This suggests that the effectof anthracyclines on myofibrillar structure is at least in partcontrolled locally.
The NRG/erbB system may be one form of local control.NRG-1� is expressed in microvascular endothelial cells andacts on the erbB2 and erbB4 receptors to alter myofilamentstructure in isolated myocytes via a phosphatidyl inositol (PI)3 kinase–dependent pathway.6 Although both anti-erbB2 andNRG-1� induced similar degrees of phosphorylation of theerbB2 receptor in the short term, they had opposite effects ondoxorubicin-induced changes in myofilament structure. Thedistinct intracellular signaling induced by NRG-1� versus theanti-erbB2 is consistent with a role of Erk1/2 and/or Akt inmediating protection of myocytes against anthracycline-induced myofilament disarray. The deleterious effect ofanti-erbB2, conversely, might occur through downregulationof erbB2 expression, suppression of intracellular signaling, orother mechanisms.
It is important to note that most women receiving trastu-zumab without concurrent anthracyclines did not develop leftventricular dysfunction or overt heart failure.17 This suggestsheterogeneity in the baseline activity of the NRG/erbBsystem among individuals, resulting from either geneticpredisposition or, more likely, environmental stress. Our owndata in vitro with anti-erbB2 support the latter conclusion,because we saw a deleterious effect of anti-erbB2 only in thepresence of anthracyclines, arguably a source of “stress.” Thevariability in the clinical toxicity of trastuzumab may thus bea result of various degrees of superimposed hemodynamic orother stress during trastuzumab therapy. Carefully monitoreduse of trastuzumab in cancer victims with other cardiovascu-
Figure 2. NRG-1� and anti-erbB2 activation of erbB2, intracellular signaling, and effect on anthracycline-induced disarray. a, NRG-1�and anti-erbB2 activate erbB2 phosphorylation to a similar extent (4.8�1.2-fold of control for anti-erbB2, n�5, vs 4.2-fold of control forNRG-1�, n�2). Twenty-four-hour treatment with anti-erbB2 reduced the level of receptor phosphorylation to baseline, and there is nofurther activation of receptor with restimulation. Blot is representative of 4 separate experiments. b, As we have previously reported,6NRG-1� caused �2-fold increase in phospho-Erk in ARVMs. In addition, NRG-1� activated phosphorylation of Akt (2.6�0.8-fold ofcontrol, n�3). There was no activation of either Erk1/2 or Akt by anti-erbB2 (0.9�0.1-fold of control for Erk, 1.2�0.3-fold of control forAkt, n�5). c, d, NRG-1� attenuated doxorubicin-induced myofibrillar disarray, whereas anti-erbB2 increased doxorubicin-induced myo-fibrillar disorganization. Values are mean�SD, 2-way ANOVA, P�0.007 for anti-erbB2, P�0.025 for NRG-1� effect on doxorubicin-induced disarray, n�8 experiments, 1600 to 2200 cells per experimental criterion.
Sawyer et al erbB2 and Myofibrillar Injury 1553
by guest on May 14, 2018
http://circ.ahajournals.org/D
ownloaded from

lar conditions, including aortic stenosis, hypertension, andischemic heart disease, will help to address this hypothesis.
AcknowledgmentsThis work was supported in part by National Institutes of Healthgrant HL-03878 to Dr Sawyer, Swiss National Science Foundation(SNF) grants 3231-054985.98/1 (SCORE A) and 32-55136.98 and agrant from the Swiss Heart Foundation to Dr Suter, SNF SwissPriority Program 5002-046083 to Dr Eppenberger, and a grant fromGenentech to Dr Sawyer. We thank Enzo Buscetti for technicalassistance and Monika Eppenberger-Eberhardt and Jean-ClaudePerriard for helpful discussions.
References1. Slamon DJ, Leyland-Jones B, Shak S, et al. Use of chemotherapy plus a
monoclonal antibody against HER2 for metastatic breast cancer thatoverexpresses HER2. N Engl J Med. 2001;344:783–792.
2. Lee KF, Simon H, Chen H, et al. Requirement for neuregulin receptorerbB2 in neural and cardiac development. Nature. 1995;378:394–398.
3. Meyer D, Birchmeier C. Multiple essential functions of neuregulin indevelopment. Nature. 1995;378:386–390.
4. Gassmann M, Casagranda F, Orioli D, et al. Aberrant neural and cardiacdevelopment in mice lacking the ErbB4 neuregulin receptor. Nature.1995;378:390–394.
5. Zhao YY, Sawyer DR, Baliga RR, et al. Neuregulins promote survivaland growth of cardiac myocytes: persistence of ErbB2 and ErbB4expression in neonatal and adult ventricular myocytes. J Biol Chem.1998;273:10261–10269.
6. Baliga RR, Pimental DR, Zhao YY, et al. NRG-1-induced cardiomyocytehypertrophy: role of PI-3-kinase, p70(S6K), and MEK-MAPK-RSK.Am J Physiol 1999;277(5 pt 2):H2026–H2037.
7. Sussman MA, Hamm-Alvarez SF, et al. Involvement of phosphorylationin doxorubicin-mediated myofibril degeneration: an immunofluorescencemicroscopy analysis. Circ Res. 1997;80:52–61.
8. Ito H, Miller SC, Billingham ME, Akimoto H, et al. Doxorubicin selec-tively inhibits muscle gene expression in cardiac muscle cells in vivo andin vitro. Proc Natl Acad Sci U S A. 1990;87:4275–4279.
9. Sawyer DB, Fukazawa R, Arstall MA, et al. Daunorubicin-induced apo-ptosis in rat cardiac myocytes is inhibited by dexrazoxane. Circ Res.1999;84:257–265.
10. Yarden Y. Agonistic antibodies stimulate the kinase encoded by the neuprotooncogene in living cells but the oncogenic mutant is constitutivelyactive. Proc Natl Acad Sci U S A. 1990;87:2569–2573.
11. Eppenberger-Eberhardt M, Flamme I, Kurer V, et al. Reexpression ofalpha-smooth muscle actin isoform in cultured adult rat cardiomyocytes.Dev Biol. 1990;139:269–278.
12. Grove BK, Kurer V, Lehner C, et al. A new 185,000-dalton skeletal muscleprotein detected by monoclonal antibodies. J Cell Biol. 1984;98:518–524.
13. Xiao L, Pimental DR, Amin JK, et al. MEK1/2-ERK1/2 mediates alpha1-adrenergic receptor-stimulated hypertrophy in adult rat ventricularmyocytes. J Mol Cell Cardiol. 2001;33:779–787.
14. Billingham ME, Mason JW, Bristow MR, et al. Anthracycline cardiomy-opathy monitored by morphologic changes. Cancer Treat Rep. 1978;62:865–872.
15. Lambertenghi-Deliliers G, Zanon PL, Pozzoli EF, et al. Myocardial injuryinduced by a single dose of adriamycin: an electron microscopic study.Tumori. 1976;62:517–528.
16. Bristow MR, Mason JW, Billingham ME, et al. Dose-effect and structure-function relationships in doxorubicin cardiomyopathy. Am Heart J. 1981;102:709–718.
17. Ewer MS, Gibbs HR, Swafford J, et al. Cardiotoxicity in patientsreceiving transtuzumab (Herceptin): primary toxicity, synergistic orsequential stress, or surveillance artifact? Semin Oncol. 1999; 26(4 suppl12):96–101.
1554 Circulation April 2, 2002
by guest on May 14, 2018
http://circ.ahajournals.org/D
ownloaded from

M. SuterDouglas B. Sawyer, Christian Zuppinger, Thomas A. Miller, Hans M. Eppenberger and Thomas
Cardiotoxicity and Anti-erbB2: Potential Mechanism for Trastuzumab-Inducedβby Neuregulin-1
Modulation of Anthracycline-Induced Myofibrillar Disarray in Rat Ventricular Myocytes
Print ISSN: 0009-7322. Online ISSN: 1524-4539 Copyright © 2002 American Heart Association, Inc. All rights reserved.
is published by the American Heart Association, 7272 Greenville Avenue, Dallas, TX 75231Circulation doi: 10.1161/01.CIR.0000013839.41224.1C
2002;105:1551-1554; originally published online March 11, 2002;Circulation.
http://circ.ahajournals.org/content/105/13/1551World Wide Web at:
The online version of this article, along with updated information and services, is located on the
http://circ.ahajournals.org//subscriptions/
is online at: Circulation Information about subscribing to Subscriptions:
http://www.lww.com/reprints Information about reprints can be found online at: Reprints:
document. Permissions and Rights Question and Answer this process is available in the
click Request Permissions in the middle column of the Web page under Services. Further information aboutOffice. Once the online version of the published article for which permission is being requested is located,
can be obtained via RightsLink, a service of the Copyright Clearance Center, not the EditorialCirculationin Requests for permissions to reproduce figures, tables, or portions of articles originally publishedPermissions:
by guest on May 14, 2018
http://circ.ahajournals.org/D
ownloaded from