molecular biology fourth edition chapter 14 messenger rna processing i: splicing lecture powerpoint...
TRANSCRIPT

Molecular BiologyFourth Edition
Chapter 14
Messenger RNA Processing I:
Splicing
Lecture PowerPoint to accompany
Robert F. Weaver
Copyright © The McGraw-Hill Companies, Inc. Permission required for reproduction or display.

14-2
14.1 Genes in Pieces• Consider the sequence of the human -globin
gene as a sentence:This is bhgty the human -globin qwtzptlrbn gene.
• Two italicized regions make no sense– Contain sequences unrelated to the globin coding
sequences surrounding them– Called intervening sequences, IVSs– Usually called introns
• Parts of the gene making sense– Coding regions– Exons
• Some lower eukaryotic genes have no introns

14-3
Evidence for Split Genes• Most higher eukaryotic genes coding for
mRNA and tRNA are interrupted by unrelated regions called introns
• Other parts of the gene, surrounding the introns, are called exons
• Exons contain the sequences that finally appear in the mature RNA product– Genes for mRNAs have been found with
anywhere from 0 to 362 exons– tRNA genes have either 0 or 1 exon

14-4
RNA Splicing• Introns are present in genes but not in mature
RNA• How does the information not find its way into
mature RNA products of the genes?– Introns are never transcribed
• Polymerase somehow jumps from one exon to another
– Introns are transcribed• Primary transcript result, an overlarge gene product is cut
down by removing introns• This is correct process
• Process of cutting introns out of immature RNAs and stitching together the exons to form final product is RNA splicing

14-5
Splicing Outline
• Introns are transcribed along with exons in the primary transcript
• Introns are removed as the exons are spliced together

14-6
Stages of RNA Splicing• Messenger RNA synthesis in eukaryotes occurs
in stages• First stage:
– Synthesis of primary transcript product– This is an mRNA precursor containing introns copied
from the gene if present– Precursor is part of a pool of large nuclear RNAs –
hnRNAs
• Second stage:– mRNA maturation– Removal of introns in a process called splicing

14-7
Splicing Signals• Splicing signals in nuclear mRNA precursors are
remarkably uniform– First 2 bases of introns are GU– Last 2 are AG
• 5’- and 3’-splice sites have consensus sequences extending beyond GU and AG motifs
• Whole consensus sequences are important to proper splicing
• Abnormal splicing can occur when the consensus sequences are mutated

14-8
14.2 Mechanism of Splicing of Nuclear mRNA Precursors
• Intermediate in nuclear mRNA precursor splicing is branched – looks like a lariat
• 2-step model– 2’-OH group of adenosine nucleotide in middle of
intron attacks phosphodiester bond between 1st exon and G beginning of intron
• Forms loop of the lariat• Separates first exon from intron
– 3’-OH left at end of 1st exon attacks phosphodiester bond linking intron to 2nd exon
• Forms the exon-exon phosphodiester bond• Releases intron in lariat form at same time

14-9
Simplified Mechanism of Splicing
• Excised intron has a 3’-OH group
• Phosphorus atom between 2 exons in spliced product comes from 3’-splice site
• Intermediate and spliced intron contain a branched nucleotide
• Branch involves 5’-end of intron binding to a site within the intron

14-10
Signal at the Branch• Along with consensus sequences at 5’- and 3’-ends of
nuclear introns, branchpoint consensus sequences also occur
• Yeast sequence invariant: UACUAAC• Higher eukaryote consensus sequence is more variable• Branched nucleotide is final A in the sequence

14-11
Spliceosomes
• Splicing takes place on a particle called a spliceosome
• Yeast spliceosomes and mammalian spliceosomes have sedimentation coefficients of 40S and 60S
• Spliceosomes contain the pre-mRNA – Along with snRNPs and protein splicing
factors– These recognize key splicing signals and
orchestrate the splicing process

14-12
snRNPs
• Small nuclear RNAs coupled to proteins are abbreviated as snRNPs, small nuclear ribonuclear proteins
• The snRNAs (small nuclear RNAs) can be resolved on a gel:– U1, U2, U4, U5, U6– All 5 snRNAs join the spliceosome to play
crucial roles in splicing

14-13
U1 snRNP• U1 snRNA sequence is
complementary to both 5’- and 3’-splice site consensus sequences– U1 snRNA base-pairs
with these splice sites– Brings the sites together
for splicing is too simple an explanation
• Splicing involves a branch within the intron

14-14
Wild-Type and Mutant U1 snRNA• Genetic experiments completed
• Base pairing between U1 snRNA and 5’-splice site of mRNA precursor – Is necessary – Not sufficient for binding

14-15
U6 snRNP• U6 snRNP associates with the 5’-end of
the intron by base pairing through the U6 RNA
• Occurs first prior to formation of lariat intermediate
• Character may change after first step in splicing
• Association between U6 and splicing substrate is essential for the splicing process
• U6 also associates with U2 during splicing

14-16
U2 snRNP• U2 snRNA base-pairs with the conserved
sequence at the splicing branchpoint• This base pairing is essential for splicing• U2 also forms base pairs with U6
– This region is called helix I– Helps orient snRNPs for splicing
• 5’-end of U2 interacts with 3’-end of U6– This interaction forms a region called helix II– This region is important in splicing in
mammalian cells, not in yeast cells

14-17
Yeast U2 Base Pairing with Yeast Branchpoint Sequence
Source: Adapted from Parker, R., P. G. Sliciano, and C. Guthrie, Recognition of the TACTAAC box during mRNA splicing in yeast involves base pairing to the U2-like snRNA. Cell 49:230, 1987.

14-18
U5 snRNP
• U5 snRNA associates with the last nucleotide in one exon and the first nucleotide of the next exon
• This should result in the two exons lining up for splicing

14-19
U4 snRNP
• U4 base-pairs with U6
• Its role seems to be to bind U6
• When U6 is needed in a splicing reaction U4 is removed
• U6 gene is split by an mRNA-type intron in at least two yeast species

14-20
snRNP Involvement in mRNA Splicing
• Spliceosomal complex contains:– Substrate – U2– U5– U6
• The complex ready for the 2nd step in splicing can be drawn as a group II intron at same stage of splicing
• Spliceosomal snRNPs substitute for elements at center of catalytic activity of group II introns at same stage of splicing

14-21
Spliceosome Catalytic Activity
• Catalytic center of spliceosome appears to include Mg2+ and a base-paired complex of 3 RNAs:– U2 snRNA– U6 snRNA– Branchpoint region of the intron
• Protein-free fragments of these RNAs can catalyze a reaction related to the first step in splicing

14-22
Spliceosome Assembly and Function
• Spliceosome is composed of many components – proteins and RNA
• These components assemble stepwise• The spliceosome cycle:
– Assembly– Function – Disassembly
• By controlling assembly of the spliceosome, a cell can regulate quality and quantity of splicing and so regulate gene expression

14-23
Spliceosome Cycle• Assembly begins with binding of U1 to splicing
substrate forming a commitment complex, a unit committed to to splicing out the intron
• U2 joins the complex next, followed by the others
• U2 binding requires ATP• U6 dissociates from U4 and displaces U1 at the
5’-splice site– This step is ATP-dependent– Activates the spliceosome– Allows U1 and U4 to be released

14-24
snRNP Structure• All snRNP’s have the same set of 7 Sm
proteins– Common targets of antibodies in patients with
systemic autoimmune diseases– Sm protein binds to a common Sm site on the
snRNAs: AAUUUGUGG• U1 snRNP has 3 specific proteins
– 70K has an Mr of 52 kD– A has an Mr of 31 kD– C has an Mr of 17.5 kD
• Sm proteins form a doughnut-shaped structure with a hole through the middle, like a flattened funnel

14-25
Sm Site and RNA
• Five snRNPs participate in splicing
• All contain a common set of 7 Sm proteins and several other proteins that are specific to snRNP
• Structure of U1 snRNP reveals that the Sm proteins form a doughnut-shaped structure to which the other proteins are attached

14-26
A Minor Spliceosome
• A minor class of introns with variant but highly conserved 5’-splice sites and branchpoints can be spliced with the help of a variant class of snRNAs
• Cells can contain minor snRNAs:– U11 performs like U1– U12 acts like U2– U4atac and U6atac perform like U4 and U6
respectively

14-27
Commitment, Splice Site Selection and Alternative Splicing• snRNPs do not have enough specificity
and affinity to bind exclusively and tightly at exon-intron boundaries
• Additional splicing factors are needed to help snRNPs bind
• Some splicing factors are needed to bridge across introns and exons and so define these RNA elements

14-28
3’-Splice Site Selection• Splicing factor Slu7 is required for correct
3’-splicing site selection• Without Slu7, splicing to correct 3’-splice
site AG is suppressed and splicing to aberrant AG’s within 30 nt of the branchpoint is activated
• U2AF is also required for 3’-splice site recognition
• 65-kD U2AF subunit binds to polypyrimidine tract upstream of 3’-splice site and 35-kD subunit binds to the 3’-splice site AG

14-29
Commitment• Commitment to splice at a given site is
determined by an RNA-binding protein• This protein binds to splicing substrate and
recruits other spliceosomal components• The first component to follow is U1• SR proteins SC35 and SF2/ASF commit
splicing on human -globin pre-mRNA and HIV tat pre-mRNA
• Part of the commitment involves attraction of U1 in some cases

14-30
Bridging Proteins and Commitment
• Yeast commitment complex has a branchpoint bridging protein (BBP) binds to:– U1 snRNP protein at the 5’-end of the intron– Mud2p near the 3’-end of the intron– RNA near the 3’-end of the intron
• Bridges the intron and could play a role defining intron prior to splicing
• Mammalian BBP is SF1, may serve same bridging function

14-31
Yeast Two-Hybrid Assay

14-32
Intron-Bridging Protein-Protein Interactions
• Branchpoint bridging protein binds to U1 snRNP protein
• Comparison of yeast to mammalian complexes is seen at right

14-33
Role of the RNA Polymerase II CTD
• C-terminal domain of the Rpb1 subunit of RNA polymerase II stimulates splicing of substrates that use exon definition
• This does not apply to those that use intron definition to prepare for splicing
• CTD binds to splicing factors and could assemble the factors at the end of exons to set them off for splicing

14-34
Alternative Splicing
• Transcripts of many eukaryotic genes are subject to alternative splicing– This splicing can have profound effects on the
protein products of a gene– Can make a difference between:
• Secreted or membrane-bound protein• Activity and inactivity
• Products of 3 genes in sex determination pathway of the fruit fly are subject to alternative splicing

14-35
Sex-Specific Splicing
• Female-specific splicing of tra transcript gives:– An active product that causes female-specific
splicing of dsx pre-mRNA– This produces a female fruit fly
• Male-specific splicing of tra transcript gives:– An inactive product that allows male-specific
splicing of dsx pre-mRNA– This produces a male fruit fly

14-36
Tra and Tra-2
• Tra and its partner Tra-2 act in conjunction with one or more other SR proteins to commit splicing at the female-specific splice site on the dsx pre-mRNA
• Commitment is probably the basis of most, if not all, alternative splicing schemes
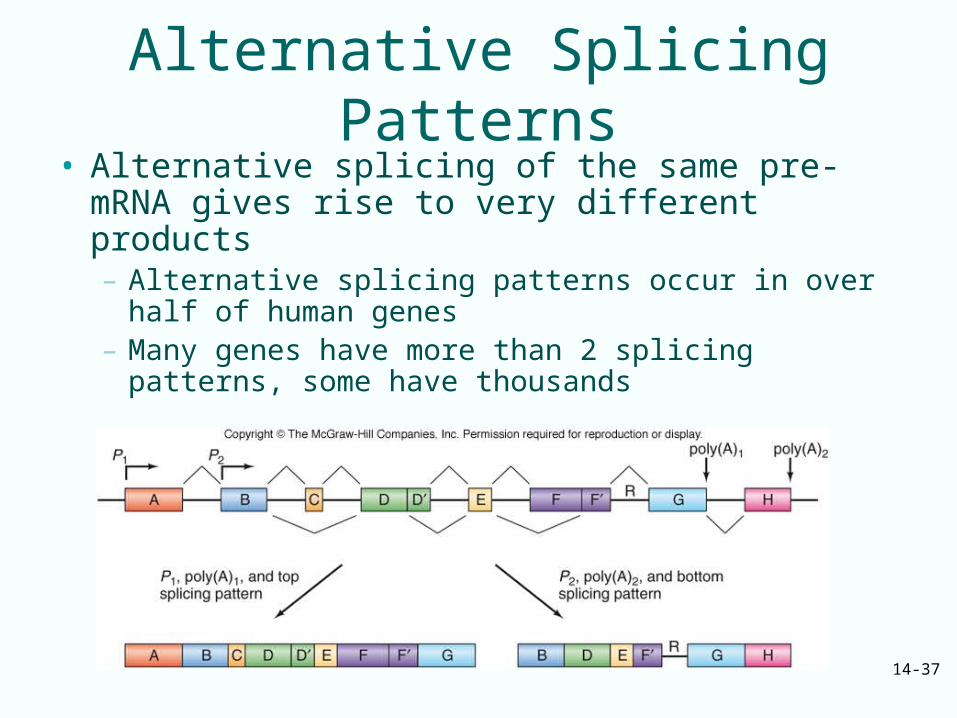
14-37
Alternative Splicing Patterns• Alternative splicing of the same pre-mRNA gives
rise to very different products– Alternative splicing patterns occur in over half of
human genes– Many genes have more than 2 splicing patterns,
some have thousands

14-38
Types of Alternative Splicing• Begin transcripts at alternative promoters• Some exons can simply be ignored resulting in
deletion of the exon• Alternative 5’-splice sites can lead to inclusion or
deletion of part of an exon• Alternative 3’-splice sites can lead to inclusion or
deletion of part of an exon• A retained intron can be retained in the mRNA if
it is not recognized as an intron• Polyadenylation causes cleavage of pre-mRNA
and loss of downstream exons

14-39
Silencing of Splicing• What stimulates
recognition of signals under only some circumstances?
• Exons can contain sequences – – Exonic splicing
enhancers (ESEs) stimulate splicing
– Exonic splicing silencers (ESSs) inhibit splicing

14-40
Reporter Construct Detects ESS Activity

14-41
14.3 Self-Splicing RNAs
• Some RNAs could splice themselves without aid from a spliceosome or any other protein
• Tetrahymena 26S rRNA gene has an intron, splices itself in vitro– Group I introns are a group of self-splicing
RNAs– Another group, Group II introns also have
some self-splicing members

14-42
Group I Introns• Group I introns can be removed in vitro with no
help from protein• Reaction begins with attack by a guanine
nucleotide on the 5’-splice site– Adds G to the 5’-end of the intron– Releases the first exon
• Second step, first exon attacks the 3’-splice site– Ligates 2 exons together– Releases the linear intron
• Intron cyclizes twice, losing nucleotides each time, then linearizes a last time

14-43
Linear Introns

14-44
Group II Introns
• RNAs containing group II introns self-splice by a pathway using an A-branched lariat intermediate, like spliceosome lariats
• Secondary structures of the splicing complexes involving spliceosomal systems and group II introns are very similar